
The commercial production of chemicals using pathway engineering
22
Review The commercial production of chemicals using pathway engineering Gopal Chotani, Tim Dodge, Amy Hsu, Manoj Kumar, Richard LaDuca, Donald Trimbur, Walter Weyler, Karl Sanford * Genencor International, 925 Page Mill Road, Palo Alto, CA 94304, USA Received 21 February 2000; received in revised form 4 September 2000; accepted 28 September 2000 Abstract Integration of metabolic pathway engineering and fermentation production technologies is necessary for the successful commercial production of chemicals. The ‘toolbox’ to do pathway engineering is ever expanding to enable mining of biodiversity, to maximize productivity, enhance carbon efficiency, improve product purity, expand product lines, and broaden markets. Functional genomics, proteomics, fluxomics, and physiomics are complementary to pathway engineering, and their successful applications are bound to multiply product turnover per cell, channel carbon efficiently, shrink the size of factories (i.e., reduce steel in the ground), and minimize product development cycle times to bring products to market. ß 2000 Elsevier Science B.V. All rights reserved. Keywords : Metabolic pathway engineering ; Ascorbic acid ; 1,3-Propanediol ; Isoprenoid ; Biocatalysis ; Aromatic 1. Introduction Metabolic pathway engineering is a rapidly devel- oping technology with great potential to impact dra- matically the development of the biobased economy [1]. Hinman [2] predicted in 1991 a thin harvest for biotechnology in the chemicals area and posited that false expectations, inherent limitations in the technol- ogy, underestimating the di/culty of competing with the power of organic chemistry, and displacing en- trenched manufacturing processes would be di/cult factors to overcome. His forecast has been quite ac- curate to date. However, change and challenge are descriptive of the environment facing the global chemical industry today, resulting in a compelling set of urgent needs that require solutions. The US chemical industry identi¢ed the following key bio- technology needs for the future: (1) biocatalysts that perform at a level not generally available today, (2) application to generation of higher performance and value products, (3) process integration with chemical processing, (4) continuous processing, and (5) better capital e/ciency [3]. Advances in recombinant DNA technology, cell physiology, genetics, functional genomics, and 0167-4838 / 00 / $ ^ see front matter ß 2000 Elsevier Science B.V. All rights reserved. PII:S0167-4838(00)00234-X Abbreviations : DAHP, 3-deoxy-D-arabinoheptulosonate 7- phosphate ; Gpd1p, Gpd2p, glycerol-3-phosphate dehydrogenase ; DHAP, dihydroxyacetone phosphate ; G-3-P, glycerol 3-phos- phate; Gpp1p and Gpp2p, DL-glycerol-3-phosphatase ; pdc, pyru- vate decarboxylase ; adh, alcohol dehydrogenase ; tal, transaldo- lase; tkt, transketolase ; xyl, xylanase; cel, cellulase; 2-KLG, 2- keto-L-gulonic acid ; 5-KDG, 5-keto-D-gluconate ; 2,5-DKG, 2,5- diketo-D-gluconate; IA, idonic acid; 2-KDG, 2-keto-D-gluconate ; GDH, glucose dehydrogenase ; GADH, gluconate dehydroge- nase; 2-KDGH, 2-keto-D-gluconate dehydrogenase ; 2,5-DKGR, 2,5-diketo-D-gluconate reductase ; IADH, idonic acid dehydroge- nase ; PEP, phosphoenolpyruvate ; NTG, NP-methyl-NP-nitro-N- nitrosoguanidine ; PQQ, pyrroloquinoline quinone * Corresponding author. Fax:+1 (650) 8456507; E-mail : [email protected] Biochimica et Biophysica Acta 1543 (2000) 434^455 www.elsevier.com/locate/bba
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The commercial production of chemicals using pathway engineering

Review
The commercial production of chemicals using pathway engineering
Gopal Chotani, Tim Dodge, Amy Hsu, Manoj Kumar, Richard LaDuca,Donald Trimbur, Walter Weyler, Karl Sanford *Genencor International, 925 Page Mill Road, Palo Alto, CA 94304, USA
Received 21 February 2000; received in revised form 4 September 2000; accepted 28 September 2000
Abstract
Integration of metabolic pathway engineering and fermentation production technologies is necessary for the successfulcommercial production of chemicals. The `toolbox' to do pathway engineering is ever expanding to enable mining ofbiodiversity, to maximize productivity, enhance carbon efficiency, improve product purity, expand product lines, andbroaden markets. Functional genomics, proteomics, fluxomics, and physiomics are complementary to pathway engineering,and their successful applications are bound to multiply product turnover per cell, channel carbon efficiently, shrink the size offactories (i.e., reduce steel in the ground), and minimize product development cycle times to bring products tomarket. ß 2000 Elsevier Science B.V. All rights reserved.
Keywords: Metabolic pathway engineering; Ascorbic acid; 1,3-Propanediol; Isoprenoid; Biocatalysis ; Aromatic
1. Introduction
Metabolic pathway engineering is a rapidly devel-oping technology with great potential to impact dra-matically the development of the biobased economy
[1]. Hinman [2] predicted in 1991 a thin harvest forbiotechnology in the chemicals area and posited thatfalse expectations, inherent limitations in the technol-ogy, underestimating the di¤culty of competing withthe power of organic chemistry, and displacing en-trenched manufacturing processes would be di¤cultfactors to overcome. His forecast has been quite ac-curate to date. However, change and challenge aredescriptive of the environment facing the globalchemical industry today, resulting in a compellingset of urgent needs that require solutions. The USchemical industry identi¢ed the following key bio-technology needs for the future: (1) biocatalyststhat perform at a level not generally available today,(2) application to generation of higher performanceand value products, (3) process integration withchemical processing, (4) continuous processing, and(5) better capital e¤ciency [3].
Advances in recombinant DNA technology, cellphysiology, genetics, functional genomics, and
0167-4838 / 00 / $ ^ see front matter ß 2000 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 8 3 8 ( 0 0 ) 0 0 2 3 4 - X
Abbreviations: DAHP, 3-deoxy-D-arabinoheptulosonate 7-phosphate; Gpd1p, Gpd2p, glycerol-3-phosphate dehydrogenase;DHAP, dihydroxyacetone phosphate; G-3-P, glycerol 3-phos-phate; Gpp1p and Gpp2p, DL-glycerol-3-phosphatase; pdc, pyru-vate decarboxylase; adh, alcohol dehydrogenase; tal, transaldo-lase; tkt, transketolase; xyl, xylanase; cel, cellulase; 2-KLG, 2-keto-L-gulonic acid; 5-KDG, 5-keto-D-gluconate; 2,5-DKG, 2,5-diketo-D-gluconate; IA, idonic acid; 2-KDG, 2-keto-D-gluconate;GDH, glucose dehydrogenase; GADH, gluconate dehydroge-nase; 2-KDGH, 2-keto-D-gluconate dehydrogenase; 2,5-DKGR,2,5-diketo-D-gluconate reductase; IADH, idonic acid dehydroge-nase; PEP, phosphoenolpyruvate; NTG, NP-methyl-NP-nitro-N-nitrosoguanidine; PQQ, pyrroloquinoline quinone
* Corresponding author. Fax:+1 (650) 8456507;E-mail : [email protected]
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
Biochimica et Biophysica Acta 1543 (2000) 434^455www.elsevier.com/locate/bba

fermentation process development have driven thedevelopment of commercially viable processes forproduction of products. Establishing metabolic path-ways and production hosts which are capable of de-livering optimal £ow of carbon from substrate to¢nal product at high yields and volumetric produc-tivities requires a focus of attention to the entiremetabolic environment within the production host.To this end, metabolic pathway engineering e¡ortshave expanded to include four key elements: (1)completion and optimization of the primary meta-bolic pathway to the targeted product including re-moval of rate limiting, transcriptional and allostericregulation; (2) genetic blockage of competing path-ways; (3) enhanced carbon commitment to the pri-mary metabolic pathway from central metabolism;and (4) modi¢cation of secondary metabolic path-ways as necessary to enhance energy metabolismand availability of required enzymatic cofactors. Inthis article and from these perspectives we will reviewwork in the aromatic, carbohydrate, organic acid,alcohol, and secondary metabolic pathways, empha-sizing our work over the last decade in certain ofthese areas and providing a view toward the futuredevelopment of this technology.
2. Aromatics
Aromatic compounds provide some of the ¢rstexamples of chemical production using microorgan-isms through the use of pathway engineering such asthe natural end products of the aromatic amino acidpathway, tryptophan [4,5], tyrosine [6], and phenyl-alanine [7]. Extension of the pathways can lead toother important commercial products such as mela-nin and indigo [8]. Intermediates in the aromaticamino acid pathway can also be used as precursorsto other biosyntheses with genes recruited from dif-ferent organisms. Examples include catechol [9],adipic acid [10] and quinic acid [11]. In general, thearomatic pathway illustrates the potential of multipleproduct opportunities from one pathway providing ameans to reduce technical and commercial develop-ment costs.
Initially, work on the aromatic amino acid path-way of Escherichia coli was focused on the construc-
tion of a strain for the overproduction of trypto-phan.
These e¡orts focused on two of the key pathwayengineering elements cited earlier: (i) completion andoptimization of primary aromatic pathway with anemphasis on removal of transcriptional and allostericregulation as well as relief of rate limiting steps; and(ii) enhancement of carbon commitment to the aro-matic pathway through modi¢cation of gene activ-ities within central metabolism.
First, it was found that critical control points lie atthe ¢rst reaction, taking intermediates from centralmetabolism and at branch points to competing path-ways. In the aromatic pathway, these critical controlpoints are the condensation of phosphoenolpyruvateand erythrose 4-phosphate to 3-deoxy-D-arabinohep-tulosonate 7-phosphate, DAHP, by DAHP synthaseand the formation of anthranilic acid from chorismicacid by anthranilate synthase. Second, transcription-al and allosteric regulation of enzyme productionand activity was overcome through the use of alter-native promoters and by the classical technique ofselection for feedback resistant mutants using toxicanalogues of the repressing compounds respectively.
Blocking of competing pathways is generally con-sidered an essential necessity for metabolite overpro-duction. For tryptophan production, the natural reg-ulation controlling production of tyrosine andphenylalanine was su¤cient to keep carbon £owingspeci¢cally to tryptophan. This eliminated the needfor addition of auxotrophic compounds to thegrowth medium.
The same could not be said for the construction ofa strain for the overproduction of phenylalanine.Here the control mechanisms for tyrosine were notsu¤cient to keep signi¢cant carbon from being di-verted. However, instead of using an auxotrophicstrain, technology was developed to keep the genefor chorismate mutase and prephenate dehydroge-nase present during the growth phase of the fermen-tation and then have it excised from the chromosomeduring the production phase [7].
Other important industrial chemicals can be pro-duced o¡ the common portion of the aromatic aminoacids pathway. Two of these are catechol and adipicacid. The key intermediate to these compounds isdehydroshikimic acid. A dehydratase and decarbox-
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 435
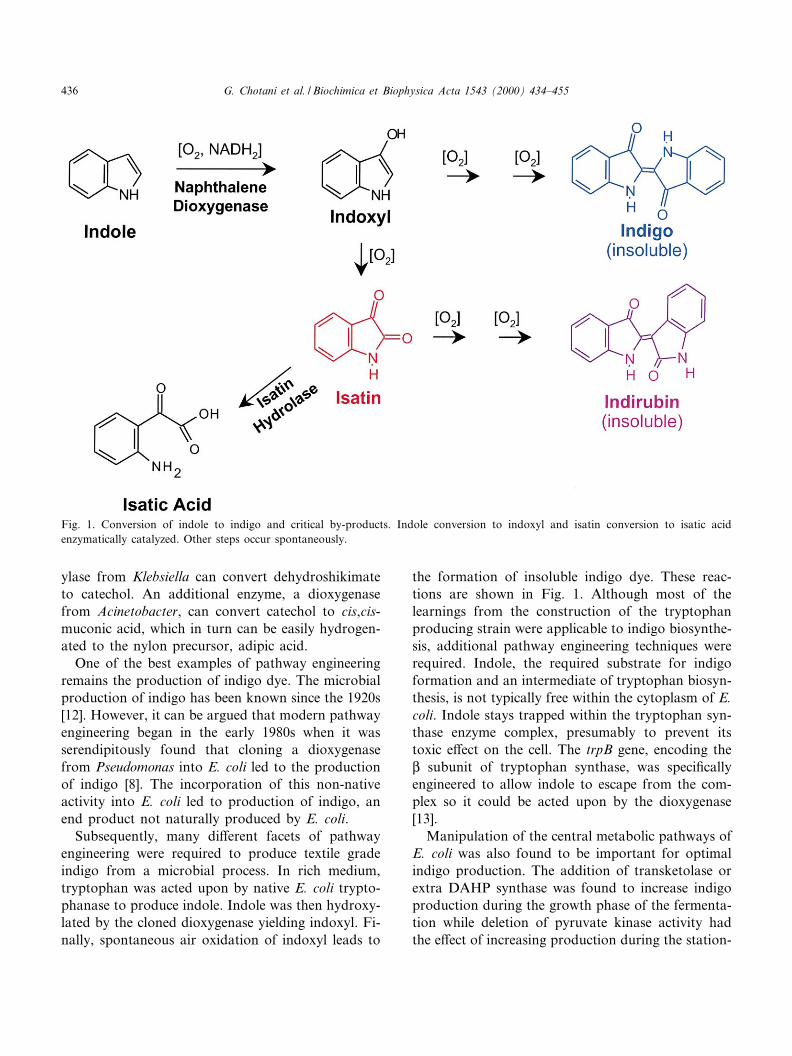
ylase from Klebsiella can convert dehydroshikimateto catechol. An additional enzyme, a dioxygenasefrom Acinetobacter, can convert catechol to cis,cis-muconic acid, which in turn can be easily hydrogen-ated to the nylon precursor, adipic acid.
One of the best examples of pathway engineeringremains the production of indigo dye. The microbialproduction of indigo has been known since the 1920s[12]. However, it can be argued that modern pathwayengineering began in the early 1980s when it wasserendipitously found that cloning a dioxygenasefrom Pseudomonas into E. coli led to the productionof indigo [8]. The incorporation of this non-nativeactivity into E. coli led to production of indigo, anend product not naturally produced by E. coli.
Subsequently, many di¡erent facets of pathwayengineering were required to produce textile gradeindigo from a microbial process. In rich medium,tryptophan was acted upon by native E. coli trypto-phanase to produce indole. Indole was then hydroxy-lated by the cloned dioxygenase yielding indoxyl. Fi-nally, spontaneous air oxidation of indoxyl leads to
the formation of insoluble indigo dye. These reac-tions are shown in Fig. 1. Although most of thelearnings from the construction of the tryptophanproducing strain were applicable to indigo biosynthe-sis, additional pathway engineering techniques wererequired. Indole, the required substrate for indigoformation and an intermediate of tryptophan biosyn-thesis, is not typically free within the cytoplasm of E.coli. Indole stays trapped within the tryptophan syn-thase enzyme complex, presumably to prevent itstoxic e¡ect on the cell. The trpB gene, encoding theL subunit of tryptophan synthase, was speci¢callyengineered to allow indole to escape from the com-plex so it could be acted upon by the dioxygenase[13].
Manipulation of the central metabolic pathways ofE. coli was also found to be important for optimalindigo production. The addition of transketolase orextra DAHP synthase was found to increase indigoproduction during the growth phase of the fermenta-tion while deletion of pyruvate kinase activity hadthe e¡ect of increasing production during the station-
Fig. 1. Conversion of indole to indigo and critical by-products. Indole conversion to indoxyl and isatin conversion to isatic acidenzymatically catalyzed. Other steps occur spontaneously.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455436

ary phase of the culture. From these results, it waspostulated that supply of erythrose 4-phosphate waslimiting during the growth phase while phosphoenol-pyruvate was limiting during the stationary phase[14].
Pathway engineering was not only important foroptimization of yield, but also to improve the qualityof the recovered material. At neutral pH of fermen-tation, the indigo precursor indoxyl yields isatin as asigni¢cant by-product. Dimerization with anothermolecule of indoxyl leads to the formation of indir-ubin, an isomer of indigo with similar dyeing proper-ties, but of deep burgundy color. This relatively largeamount of indirubin caused problems ranging from ared cast to interfering with bleaching during denim¢nishing. In order to make textile grade indigo froma fermentation process, the level of indirubin had tobe reduced.
A potential means to eliminate indirubin was tohydrolyze isatin to isatic acid. An enzyme to carryout this reaction was identi¢ed in a screen of soilsamples predicated on the hypothesis that such anactivity might exist in a natural indole degradationpathway. After cloning and incorporating the newenzyme in the production strain, the indigo productperformed equal to indigo produced chemically [15].
3. Organic acids
3.1. Succinic acid
Succinic acid can be used as an intermediate in thechemical synthesis of 1,4-butanediol, tetrahydrofur-an, and adipic acid, all with large market potential.However, large scale use requires that succinic acidbe produced less expensively than through its currentpetrochemical route. Succinic acid is a common in-termediate in the metabolic pathway of several an-aerobic microorganisms. It is a key intermediate forpropionate producing bacteria and is also producedby anaerobic rumen bacteria. In both cases, however,succinate is produced by mixed acid fermentations inlow yields and concentrations along with several by-products. Anaerobiospirillum succiniproducens, an an-aerobic non-rumen bacterium, and a rumen-faculta-tive anaerobic bacterium Actinobacillus succinogenes130Z have been shown to produce succinic acid as a
major product, in high titer and yield [16^18]. A.succiniproducens can produce 1.2 mol of succinateper mol of glucose [19,20]. A. succiniproducens andA. succinogenes use a phosphoenolpyruvate (PEP)carboxykinase pathway to make succinic acid. Fourkey enzymes include PEP carboxylase, malate dehy-drogenase, fumarase, and fumarate dehydrogenase.Carbon dioxide concentration has been shown toregulate the levels of these enzymes for productionof succinate. Carbon dioxide functions as an electronacceptor and modulates the £ux of PEP. Higher lev-els of CO2 thus favor succinate formation. Electronsgenerated by pyruvate oxidation are used for thereduction of fumarate to succinate. Thus underCO2/H2 atmosphere (hydrogenase), more PEP goesto succinate rather than to pyruvate. A. succinogenesvariants can give titers of 110 g/l and yields of 83^87wt% while maintaining pH with magnesium [21,22].A commercially viable succinic acid fermentationprocess with Actinobacillus sp. or Anaerobiospirillumsp. must be delivered at high yield and speci¢c pro-ductivity. This is being addressed with a genetic en-gineering pathway approach.
E. coli has also been engineered to produce suc-cinic acid [23]. Phosphoenolpyruvate generated inglycolysis is proportionately partitioned to ¢nalproducts that di¡er in their oxidation state. The dis-tribution of fermentation products can be altered byblocking branches of the normal fermentative path-ways or by introducing critical branch point en-zymes. Succinic acid production was increased byoverexpressing phosphoenolpyruvate carboxylase inone E. coli strain with normal glucose fermentationand by overexpressing malic enzyme in anotherblocked in its ability to ferment glucose [24]. Furthermetabolic engineering of the strain led to loss ofglucose fermentation due to inactivation of genesencoding pyruvate: formate lyase and the fermenta-tive lactate dehydrogenase [25]. Furthermore,through spontaneous chromosomal mutation in themembrane bound permease (enzyme IICB) of glu-cose-speci¢c bacterial phosphotransferase system(PTS), succinic acid production in E. coli was en-hanced to 51 g/l and a wt% yield of 95 [26]. Byfurther modifying E. coli, with advances in measur-ing intracellular metabolites and carbon £ux analysis,it should be feasible to develop a commercial bioca-talytic succinic acid process.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 437

3.2. Lactic acid
L-Lactic acid has an ancient history of use as afood preservative and food £avoring compound. Re-cently, lactic acid has received attention since it canbe condensed into a biodegradable polymer. Themarket for lactic acid is rapidly growing, exceedingseveral hundred million dollars annually. Under non-energy limiting batch fermentation conditions, ho-mofermentative bacteria predominantly produce lac-tic acid as their end product. Lactic acid yields arehighest during glycolysis via the homolactic acid fer-mentative pathway [27]. Theoretically, 2 moles oflactate and 2 moles of ATP are formed per mole ofglucose used. Although free lactic acid is preferredfor most industrial processes, anaerobic fermentationfor the production of the organic acid operates opti-mally at pH values where the salt of the organic acidrather than the free acid is formed. To obtain lacticacid in its free form, the fermentation process mustbe carried out at or below its pKa of 3.87. An elegantapproach to this challenge was recently published[28,29]. By insertion of the bovine LDH-A geneinto a Crabtree-negative Kluyveromyces lactis (no py-ruvate decarboxylase gene), pyruvate £ux towardethanol production was fully replaced by lactic acidproduction (1.19 mol lactate per mole of glucose).Transferring the process to a 14 l fermenter gave a
titer of 109 g/l with productivity of 0.8 g/l/h at pH4.5. A doubling of yield as well as titer was achievedin a fermentation carried out with a strain of Sac-charomyces cerevisiae overexpressing the lactate-pro-ton symporter coded for by the gene JEN1 [30].
3.3. L-Ascorbic acid
L-Ascorbic acid (vitamin C) is used in large scaleas an antioxidant in food, animal feed, beverages,pharmaceutical formulations and cosmetic applica-tions. The current world market of ascorbic acid is60 000^70 000 metric tons per year and generates an-nual revenues in excess of US$ 500 million.
The synthesis of L-ascorbic acid [31] developed in1934 by Reichstein and Grussner has remained thecornerstone for the production of vitamin C. It isbelieved that this chemical process currently operatesat a yield of about 50%. Almost all industrial pro-cesses for vitamin C are derived from the Reichsteinand Grussner synthesis, which uses a single biocatal-ysis step within a series of chemically-based unit op-erations. Alternative biological processes exist [32],but almost all involve 2-keto-L-gulonic acid (2-KLG) as the key intermediate (Fig. 2). In all cases2-KLG can be converted to ascorbic acid ¢nal prod-uct using conventional chemical processing technol-ogy.
Fig. 2. Process routes to ascorbic acid.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455438

Four particular e¡orts have gained signi¢cance:D-Sorbitol route. Using genetic engineering meth-
ods, two strains of Gluconobacter oxydans (T-100and G624, 2-KLG producing and sorbose forming,respectively) were modi¢ed. Genes for sorbitol dehy-drogenase and sorbose dehydrogenase were clonedfrom T-100 into G624 to produce 2-KLG from sor-bitol in three steps [33]. Chemical NTG mutagenesiswas used to create a strain leading to diminishedformation of idonic acid, a by-product. Further im-provements of this process can be envisaged by opti-mizing gene expression, fermentation process, andcarbon £ux redirection.
5-Keto-D-gluconate (5-KDG) route. 2-KLG biosyn-thesis using 5-KDG involves oxidation of glucose togluconic acid followed by further oxidation to 5-KDG which can then be reduced to L-idonic acid[34] or L-gulonic acid [35]. Both of these intermedi-ates can be oxidized subsequently to 2-KLG [36].
2,5-Diketo-D-gluconate (2,5-DKG) route in two-step fermentation. 2,5-DKG is formed from glucosein a single step in the genus Erwinia. Two intermedi-ates are D-gluconate and 2-keto-D-gluconate. Fer-mentation using Acetobacter or Erwinia sp. led toover 90% yield of 2,5-DKG from glucose in 20 h[37]. Sonoyama ¢rst described a concept of using
two-step fermentation to produce 2-KLG [38]. Thistechnology resulted from the observation that 2,5-DKG can be reduced to 2-KLG by Brevibacterium[39] or Corynebacterium [40]. It is important to notethat the conversion yield of D-glucose to 2-KLG inthis staged culture with Acetobacter and Corynebac-terium approaches the theoretically maximum ob-tainable yield of the Reichstein process.
One organism route, D-glucose to 2-KLG formation.To simplify the conversion of glucose to 2-KLG,from a two-stage process to a one-step process, arecombinant Erwinia sp. was genetically engineeredat Genentech [41]. The gene of 2,5-DKG reductasefrom Corynebacterium [42] was cloned and expressedin a host strain capable of converting glucose to 2,5-DKG (Fig. 3). This achieved the goal of transform-ing glucose into 2-KLG in a single-step fermentationprocess [43]. This process is fundamentally simplerthan either the current multi-step Reichstein processor the two-stage fermentation process. The single-step fermentation process has been further simpli¢edby Genencor International to produce titers in 14 lfermenters greater than 120 g/l for 2-KLG in lessthan 120 h of fermentation time (Fig. 4) [44,53].
Detailed studies of the metabolism of ketoaldonicacid were carried out to understand the pathway for
Fig. 3. One-step biological production of 2-KLG. Recovered 2-KLG is converted to ascorbic acid through chemical esteri¢cation andlactonization reactions.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 439

Fig. 4. 2-KLG production from glucose in a single production host.
Fig. 5. Carbon £ow to 2-KLG in a Gram-negative production host. Enzymes in metabolic pathway to 2-KLG include: GDH, glucosedehydrogenase; GADH, gluconate dehydrogenase; 2-KDGDH, 2-keto-D-gluconate dehydrogenase, 2,5-DKGR, 2,5-diketo-D-gluconicacid reductase; and IADH, idonic acid dehydrogenase.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455440
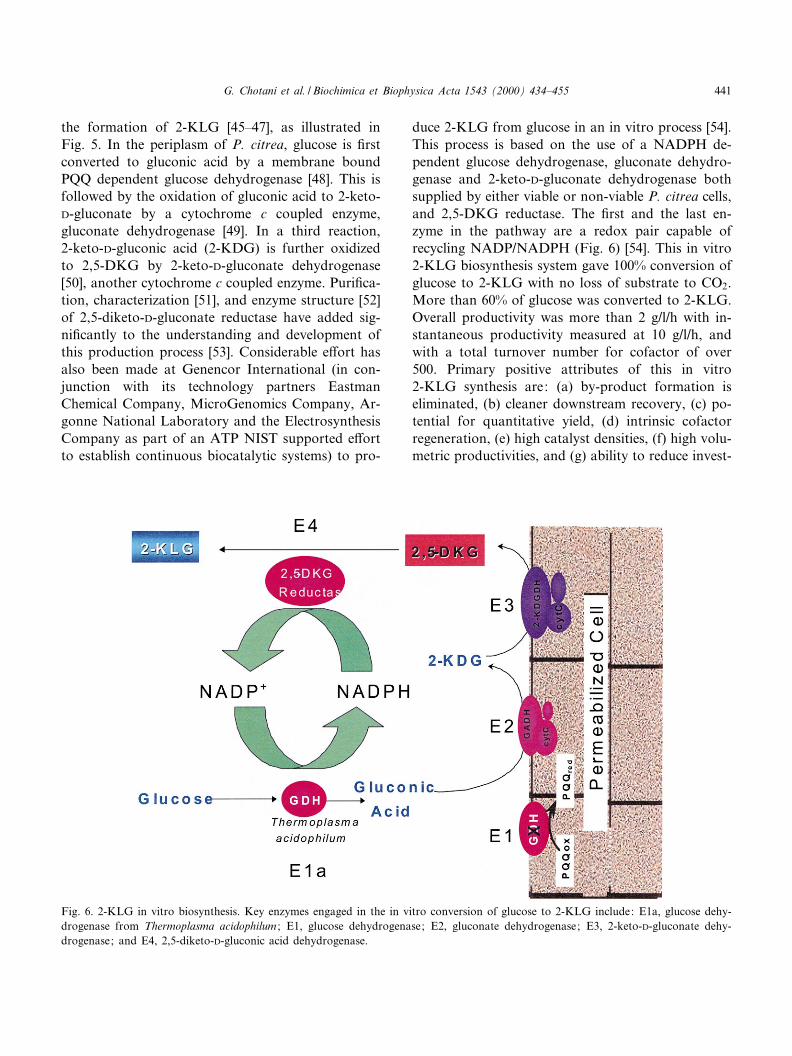
the formation of 2-KLG [45^47], as illustrated inFig. 5. In the periplasm of P. citrea, glucose is ¢rstconverted to gluconic acid by a membrane boundPQQ dependent glucose dehydrogenase [48]. This isfollowed by the oxidation of gluconic acid to 2-keto-D-gluconate by a cytochrome c coupled enzyme,gluconate dehydrogenase [49]. In a third reaction,2-keto-D-gluconic acid (2-KDG) is further oxidizedto 2,5-DKG by 2-keto-D-gluconate dehydrogenase[50], another cytochrome c coupled enzyme. Puri¢ca-tion, characterization [51], and enzyme structure [52]of 2,5-diketo-D-gluconate reductase have added sig-ni¢cantly to the understanding and development ofthis production process [53]. Considerable e¡ort hasalso been made at Genencor International (in con-junction with its technology partners EastmanChemical Company, MicroGenomics Company, Ar-gonne National Laboratory and the ElectrosynthesisCompany as part of an ATP NIST supported e¡ortto establish continuous biocatalytic systems) to pro-
duce 2-KLG from glucose in an in vitro process [54].This process is based on the use of a NADPH de-pendent glucose dehydrogenase, gluconate dehydro-genase and 2-keto-D-gluconate dehydrogenase bothsupplied by either viable or non-viable P. citrea cells,and 2,5-DKG reductase. The ¢rst and the last en-zyme in the pathway are a redox pair capable ofrecycling NADP/NADPH (Fig. 6) [54]. This in vitro2-KLG biosynthesis system gave 100% conversion ofglucose to 2-KLG with no loss of substrate to CO2.More than 60% of glucose was converted to 2-KLG.Overall productivity was more than 2 g/l/h with in-stantaneous productivity measured at 10 g/l/h, andwith a total turnover number for cofactor of over500. Primary positive attributes of this in vitro2-KLG synthesis are: (a) by-product formation iseliminated, (b) cleaner downstream recovery, (c) po-tential for quantitative yield, (d) intrinsic cofactorregeneration, (e) high catalyst densities, (f) high volu-metric productivities, and (g) ability to reduce invest-
Fig. 6. 2-KLG in vitro biosynthesis. Key enzymes engaged in the in vitro conversion of glucose to 2-KLG include: E1a, glucose dehy-drogenase from Thermoplasma acidophilum ; E1, glucose dehydrogenase; E2, gluconate dehydrogenase; E3, 2-keto-D-gluconate dehy-drogenase; and E4, 2,5-diketo-D-gluconic acid dehydrogenase.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 441

ment capital by shrinking the size of the productionreactor. Although redox recycling of cofactor is notan issue, cofactor cost is still high, and enzyme stabil-ities need improvement.
The vision of manufacturing ascorbic acid directlyby fermentation has remained elusive. Nevertheless,e¡orts to this end are ongoing at Genencor Interna-tional [55]. By employing a metabolic selection strat-egy, we have now identi¢ed a 2-KLG to ascorbicacid activity in Candida blankii and Cryptococcusdimmnae yeast sp. [55,56]. Another direct routefrom D-glucose to L-ascorbic acid in microalgae hasalso been developed and reported [32]. Additionalbioengineering is required to advance the direct fer-mentation of glucose to ascorbic acid without theneed to isolate 2-KLG.
4. Alcohols
4.1. Ethanol
Ethanol production by fermentation processes hasreached a turning point in acceptability with the em-phasis of its use as an oxygenated gasoline additive.There will only be su¤cient, low cost ethanol if pro-duction is integrated with e¤cient utilization of re-newable resources. In other words, ethanol has thepotential to become a fully sustainable energy re-source if its production technology is developedalong the 2020 Roadmap http://www.oit.doe.gov/agriculture/. This technology roadmap includesengineered metabolic pathways for conversion of re-newable plant-derived substrates to ethanol. Severalorganisms have already been engineered to overpro-duce ethanol [57,60], as shown in Table 1.
Lignocellulosic hydrolysates, produced via somecombination of treatments (acid, base, solvent, andcocktails of enzymes), contain xylose, arabinose, glu-cose, galactose, mannose, fucose, and rhamnose. Nonaturally occurring organisms rapidly convert all ofthese sugars into ethanol. Natural and rapid ferment-ing organisms such as Saccharomyces and Zymomo-nas mobilis metabolize a limited range of carbon sub-strates. On the other hand, several bacteria such asE. coli, Klebsiella, Erwinia, Lactobacilli, Bacilli, andClostridia utilize a wide variety of carbon sources,but conversion results in limited ethanol yield be-cause of mixed acids and solvents are also producedby their metabolism. Various modes of central me-tabolism [58] of 5- and 6-carbon sugars play majorroles in deciding the ultimate fate of conversion ofpyruvate, the key 3-carbon intermediate, to ethanol,formate, acetate, lactate, propionate, acetoin, ace-tone, isopropanol, butanol, 2,3-butanediol, butyrate,succinate, hydrogen, and carbon dioxide (see http://www.expasy.ch/cgi-bin/show_image?e5).
In general, engineering an e¤cient host is aimed atmaking the ultimate production system faster in rate,better in titer, and cheaper in cost. Because of the`Catch22' situation as explained above, carbon £owto ethanol is compromised by many factors: sugartransport into the cell ; feedback regulation of path-way enzymes by inhibitors including substrates,products, and by-products; multiple pathway branchpoints; turnover of intermediates and cofactors; andethanol e¥ux from the cell. Therefore, classical mu-tagenesis and selection without genetic engineeringcan go only part way towards building the produc-tion system. With advances in metabolic engineeringtools of gene discovery, biodiversity screen, function-al genomics, directed evolution, pathway engineer-
Table 1Microorganisms engineered for ethanol production
Engineered host Engineered gene(s) Donor host
E. coli, Erwinia chrysanthemi, Klebsiella planticola pdc Z. mobilisE. coli adhB Z. mobilisE. coli, Klebsiella oxytoca, E. chrysanthemi, Erwinia carotovora,Enterobacter cloacae, Bacillus subtilis
pdc, adhB Z. mobilis
Z. mobilis tal, tktA, xylA, xylB E. coliE. coli celZ E. chrysanthemiLactococcus lactis, Lactococcus plantarum, Streptococcus mutans pdc, adh Z. mobilisS. cerevisiae pentose pathway genes
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455442

ing, and process design, each of the major metabolicine¤ciencies can now be addressed. Two approacheshave been widely used by pathway engineers: (1)expand pentose utilization capacities of the hosts(Saccharomyces, Zymomonas) which are already e¤-cient in converting hexoses to ethanol, (2) divert car-bon £ow from the native fermentation products toethanol in the hosts (Escherichia, Klebsiella, Erwinia)which are already e¤cient in utilizing mixed sugars.Ingram et al. have demonstrated that it is realistic toborrow enzyme systems of one organism and opti-mally combine them into another host [59]. Suchrecombinant hosts are now being constantly im-proved towards the ultimate goal of sustaining chem-icals production from renewable resources in thenear future.
4.2. Glycerol
Glycerol is a commodity chemical used in cosmet-ics, liquid soaps, food, pharmaceuticals, lubricants,antifreeze solutions, tobacco, and many other appli-cations. It can be synthesized chemically from pro-pylene or biochemically from natural glycerol pro-ducing organisms or hydrolysis of glycerides.Currently the majority of glycerol is producedthrough biochemical routes.
Some species of bacteria, algae, protozoa, andyeast are reported to produce glycerol [61^63]. Forexample, the bacterium Bacillus licheniformis is capa-ble of glycerol overproduction. Most strains of Sac-charomyces produce some glycerol during alcoholicfermentation and this production can be increased bythe application of osmotic stress [63].
Early last century, Carl Neuberg developed twoprocesses that greatly increased the yield of glycerolby strains of S. cerevisiae. These two processes, oftencalled Neuberg's `second' and `third forms' of fer-mentations, require `steering agents' to divert the fer-mentation into di¡erent pathways. In order to divertmore reducing equivalents (NADH) to glycerol pro-duction, steering reagents such as bisul¢tes or alkaliswere added into the fermentation process to inhibitthe conversion of acetaldehyde to ethanol or initiatea Cannizarro disproportionation to yield ethanol andacetic acid from two equivalents of acetaldehyde.Petrovska reported the addition of Na2SO3 to the`second form' of fermentation, which shifts the me-
tabolism of all yeasts toward glycerol formation in amicroaerophilic condition [64].
Both the primary methods for producing glycerolbiochemically today have inherent problems. Isola-tion of glycerol from animal fat and similar sourcesis laborious and ine¤cient. Producing glycerol fromnaturally occurring microorganisms such as Saccha-romyces requires the use of sul¢tes. Pathway engi-neering of microbial organisms can create a betterway. Glycerol is produced from the glycolytic inter-mediate dihydroxyacetone 3-phosphate using twoenzymes: dihydroxyacetone-3-phosphate dehydroge-nase and glycerol-3-phosphatase. A number of thegenes involved in the glycerol biosynthetic pathwayfrom a natural producer have been isolated. For ex-ample, genes encoding a NAD�-dependent glycerol-3-phosphate dehydrogenase (GPD1 and GPD2) havebeen cloned and sequenced from Saccharomyces dia-staticus [65] and S. cerevisiae [66]. Glycerol-3-phos-phate dehydrogenase (EC 1.1.1.8) converts dihy-droxyacetone phosphate (DHAP) to glycerol 3-phos-phate (G-3-P). Exposing Saccharomyces to osmoticstress leads to increased production of glycerol-3-phosphate dehydrogenase due to transcriptional acti-vation of the isogene GPD1 but not GPD2 [67,68].The expression of GPD1 appears to be controlled byexternal osmolarity via an osmosensing mechanismand a mitogen-activated protein (MAP) kinase cas-cade called high osmolarity glycerol response (HOG)pathway [69,70]. Additionally, genes encoding DL-glycerol-3-phosphatase (GPP1 and GPP2) from S.cerevisiae have been identi¢ed [71]. DL-Glycerol-3-phosphatase (Gpp1p and Gpp2p) catalyzes the hy-drolysis of G-3-P to glycerol (EC 3.1.3.21). Underosmotic stress, the intracellular concentration ofGpp2p increases but not that of Gpp1p. BesidesDL-glycerol-3-phosphatase from Saccharomyces spe-cies, glycerophosphatase from other sources hasbeen isolated, including B. licheniformis [72], andPhycomyces blakesleeanus [73].
Other research groups have reported metabolicpathway engineering experiments to produce glycerolin di¡erent microorganisms. For example, in 1997,Michnick et al. reported genetically engineered S. ce-revisiae strains with enhanced levels of glycerol-3-phosphate dehydrogenase (Gpd1p). These strains fer-mented glucose with a strongly modi¢ed [glycerol] :[ethanol] product ratio, which produced glycerol at
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 443
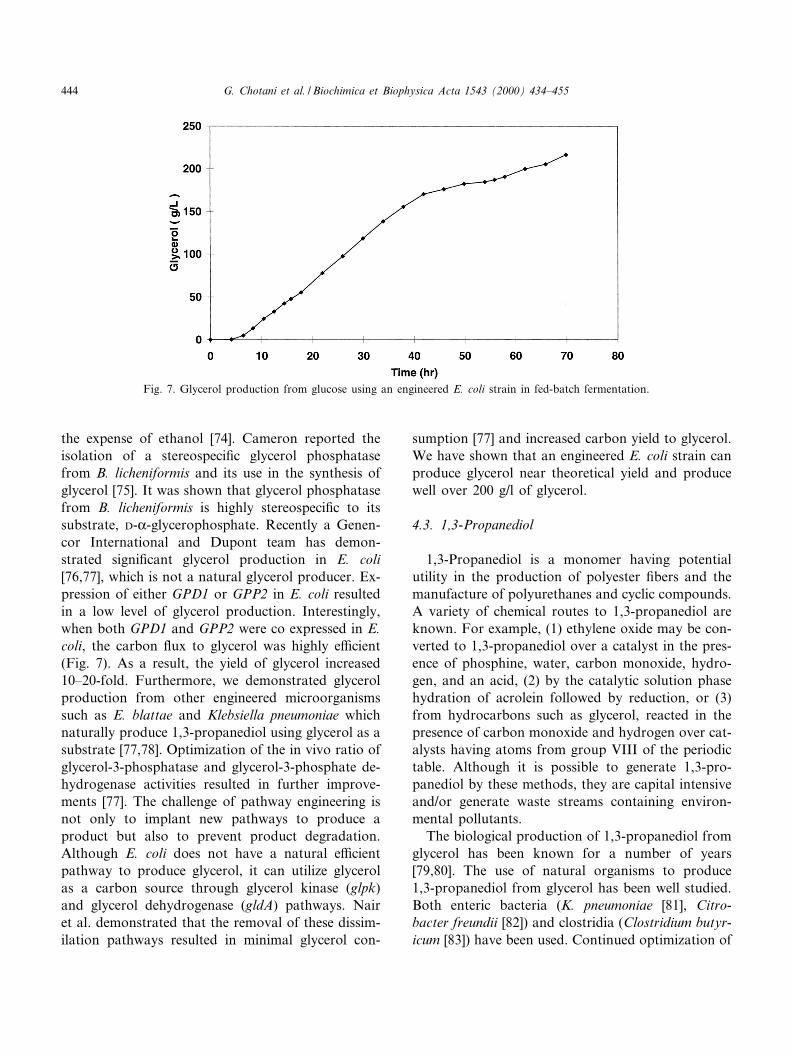
the expense of ethanol [74]. Cameron reported theisolation of a stereospeci¢c glycerol phosphatasefrom B. licheniformis and its use in the synthesis ofglycerol [75]. It was shown that glycerol phosphatasefrom B. licheniformis is highly stereospeci¢c to itssubstrate, D-K-glycerophosphate. Recently a Genen-cor International and Dupont team has demon-strated signi¢cant glycerol production in E. coli[76,77], which is not a natural glycerol producer. Ex-pression of either GPD1 or GPP2 in E. coli resultedin a low level of glycerol production. Interestingly,when both GPD1 and GPP2 were co expressed in E.coli, the carbon £ux to glycerol was highly e¤cient(Fig. 7). As a result, the yield of glycerol increased10^20-fold. Furthermore, we demonstrated glycerolproduction from other engineered microorganismssuch as E. blattae and Klebsiella pneumoniae whichnaturally produce 1,3-propanediol using glycerol as asubstrate [77,78]. Optimization of the in vivo ratio ofglycerol-3-phosphatase and glycerol-3-phosphate de-hydrogenase activities resulted in further improve-ments [77]. The challenge of pathway engineering isnot only to implant new pathways to produce aproduct but also to prevent product degradation.Although E. coli does not have a natural e¤cientpathway to produce glycerol, it can utilize glycerolas a carbon source through glycerol kinase (glpk)and glycerol dehydrogenase (gldA) pathways. Nairet al. demonstrated that the removal of these dissim-ilation pathways resulted in minimal glycerol con-
sumption [77] and increased carbon yield to glycerol.We have shown that an engineered E. coli strain canproduce glycerol near theoretical yield and producewell over 200 g/l of glycerol.
4.3. 1,3-Propanediol
1,3-Propanediol is a monomer having potentialutility in the production of polyester ¢bers and themanufacture of polyurethanes and cyclic compounds.A variety of chemical routes to 1,3-propanediol areknown. For example, (1) ethylene oxide may be con-verted to 1,3-propanediol over a catalyst in the pres-ence of phosphine, water, carbon monoxide, hydro-gen, and an acid, (2) by the catalytic solution phasehydration of acrolein followed by reduction, or (3)from hydrocarbons such as glycerol, reacted in thepresence of carbon monoxide and hydrogen over cat-alysts having atoms from group VIII of the periodictable. Although it is possible to generate 1,3-pro-panediol by these methods, they are capital intensiveand/or generate waste streams containing environ-mental pollutants.
The biological production of 1,3-propanediol fromglycerol has been known for a number of years[79,80]. The use of natural organisms to produce1,3-propanediol from glycerol has been well studied.Both enteric bacteria (K. pneumoniae [81], Citro-bacter freundii [82]) and clostridia (Clostridium butyr-icum [83]) have been used. Continued optimization of
Fig. 7. Glycerol production from glucose using an engineered E. coli strain in fed-batch fermentation.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455444

the basic anaerobic glycerol fermentation process hasproduced promising results. A number of batch orfed-batch fermentations using K. pneumoniae or C.butyricum have produced titers of 50^75 g/l and yieldof 1,3-propanediol 0.44^0.69 mol/mol from glycerol[84,85]. Continuous culture has also been studied for1,3-propanediol production. Typical of continuousprocesses high productivity was achieved, but withlower titers [86,87].
A number of factors have been shown to a¡ect the1,3-propanediol fermentation process: substrate con-centration, co-substrates, H2, and product inhibition.Complete conversion of glycerol to 1,3-propanediolis not possible due to the requirement of an addi-tional reducing equivalent. Therefore the additionof a cheap co-substrate such as glucose should allow100% of the glycerol to be converted to 1,3-propane-diol. Various co-substrates have been tried andshown to increase yields, but problems such as glu-cose inhibition and by-product formation reduce thee¡ectiveness of this strategy [88].
Classical methods have been used to select for 1,3-propanediol resistant Clostridium [89]. The two en-zymes involved in the conversion of glycerol to 1,3-propanediol have been cloned and characterizedfrom several organisms: Klebsiella, Citrobacter, andClostridium. The ¢rst enzyme in the pathway is glyc-erol dehydratase. The dehydratase has been shown tobe a multi-subunit (K2L2Q2) B12-dependent enzyme[90]. The dehydratase has been shown to undergocatalytic inactivation and requires the addition of areactivation complex of two additional proteins [91].The second enzyme in the pathway is NAD-linkeddehydrogenase. Both of these enzymes have beencloned and expressed in E. coli [92,93]. These re-combinant strains of E. coli have been shown toproduce 1,3-propanediol, but generally with low lev-els [94^96]. Improvement in the glycerol to 1,3-pro-panediol fermentation process should be possible bygenetic modi¢cation of natural 1,3-propanediol pro-ducers by increasing expression of the genes in thepathway and limiting by-product formation.
Neither of the chemical nor biological methodsdescribed above for the production of 1,3-propane-diol are well suited for industrial scale productionsince the chemical processes are energy intensiveand the biological processes require the expensivestarting material, glycerol. A method requiring low
energy input and an inexpensive starting material isneeded. A more desirable process would incorporatea microorganism that would have the ability to con-vert basic carbon sources such as carbohydrates orsugars to the desired 1,3-propanediol end product.
Since there are a number of organisms which canproduce glycerol, one can combine a glycerol pro-ducing organism and a 1,3-propanediol producingorganism into a two-step or mixed fermentation.Glucose is known to inhibit 1,3-propanediol produc-tion in enteric bacteria, which would have to beovercome before using a mixed culture. Two-stagefermentation has been demonstrated using S. cerevi-siae and K. pneumoniae [97].
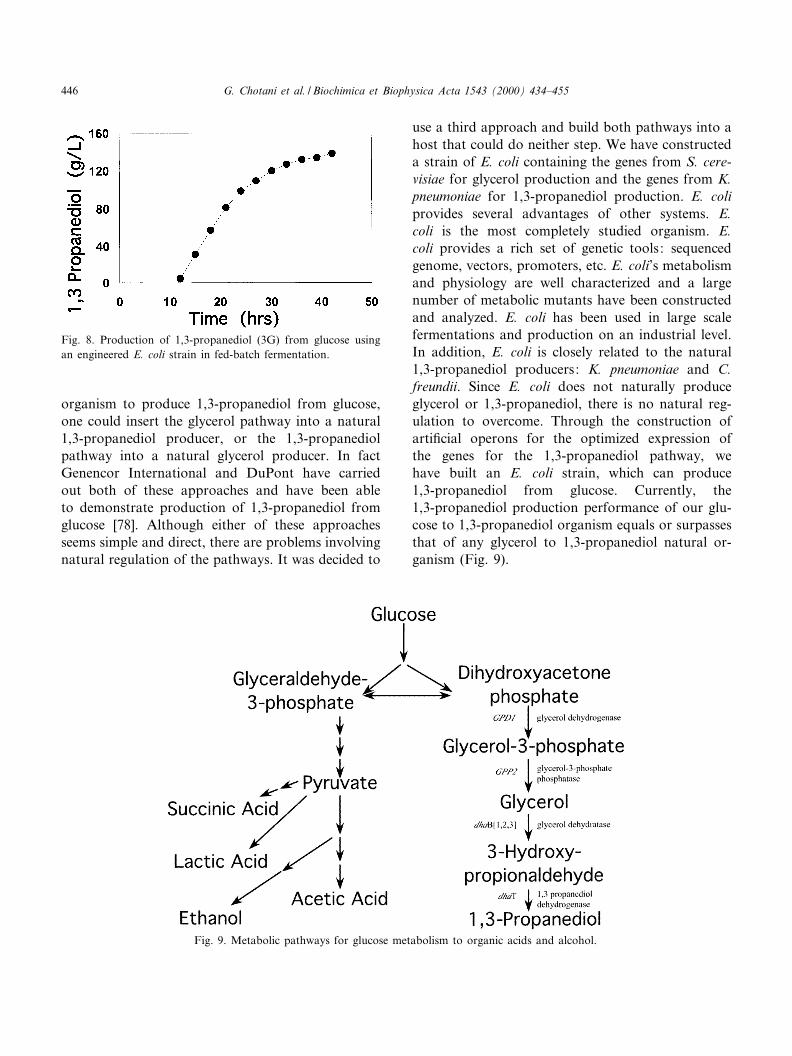
Although biological methods of both glycerol and1,3-propanediol production are known, it has neverbeen demonstrated that a single organism can ac-complish the entire process. The combination ofthe natural biological pathways for the productionof glycerol from glucose and the production of 1,3-propanediol from glycerol into a single host shouldyield a superior process for the production of 1,3-propanediol. The conversion of glucose to 1,3-pro-panediol requires two reducing equivalents per 1,3-propanediol molecule. The classical anaerobic fer-mentation process to produce 1,3-propanediol fromglycerol is limited by the amount of reducing equiv-alents which can be generated from the productionof organic acids such as acetate. The single organismapproach (under development by Genencor Interna-tional and DuPont) overcomes this problem by al-lowing some of the glucose to be converted all theway to CO2 providing more reducing power per glu-cose molecule. The ability to control both carbonand energy £ow in the single organism allows moree¤cient use of the input carbon source.
The conversion of glucose to 1,3-propanediol re-quires the combination of two natural pathways:glucose to glycerol and glycerol to 1,3-propanediol(Fig. 8). The best natural pathways for the produc-tion of glycerol are found in the yeast. S. cerevisiaeproduces glycerol from the glycolytic intermediatedihydroxyacetone 3-phosphate using two enzymes:dihydroxyacetone-3-phosphate dehydrogenase andglycerol-3-phosphate phosphatase. The natural path-way for the production of 1,3-propanediol from glyc-erol requires two enzymes: glycerol dehydratase and1,3-propanediol dehydrogenase. To construct a single
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 445
organism to produce 1,3-propanediol from glucose,one could insert the glycerol pathway into a natural1,3-propanediol producer, or the 1,3-propanediolpathway into a natural glycerol producer. In factGenencor International and DuPont have carriedout both of these approaches and have been ableto demonstrate production of 1,3-propanediol fromglucose [78]. Although either of these approachesseems simple and direct, there are problems involvingnatural regulation of the pathways. It was decided to
use a third approach and build both pathways into ahost that could do neither step. We have constructeda strain of E. coli containing the genes from S. cere-visiae for glycerol production and the genes from K.pneumoniae for 1,3-propanediol production. E. coliprovides several advantages of other systems. E.coli is the most completely studied organism. E.coli provides a rich set of genetic tools: sequencedgenome, vectors, promoters, etc. E. coli's metabolismand physiology are well characterized and a largenumber of metabolic mutants have been constructedand analyzed. E. coli has been used in large scalefermentations and production on an industrial level.In addition, E. coli is closely related to the natural1,3-propanediol producers: K. pneumoniae and C.freundii. Since E. coli does not naturally produceglycerol or 1,3-propanediol, there is no natural reg-ulation to overcome. Through the construction ofarti¢cial operons for the optimized expression ofthe genes for the 1,3-propanediol pathway, wehave built an E. coli strain, which can produce1,3-propanediol from glucose. Currently, the1,3-propanediol production performance of our glu-cose to 1,3-propanediol organism equals or surpassesthat of any glycerol to 1,3-propanediol natural or-ganism (Fig. 9).
Fig. 9. Metabolic pathways for glucose metabolism to organic acids and alcohol.
Fig. 8. Production of 1,3-propanediol (3G) from glucose usingan engineered E. coli strain in fed-batch fermentation.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455446

5. Secondary metabolites
Secondary metabolism in all branches of life pro-duces an astounding array of chemical compounds.The mevalonic acid (terpenes), polyketide synthase,and non-ribosomal polypeptide synthase pathwaysare major contributors to this diversity. Many prod-ucts derived from these pathways ¢nd human use, oras in the case of vitamins are required by us, and assuch are derived from their natural sources, pro-duced by synthetic or semisynthetic means, and aretargets for production by the new biotechnology in-dustry. While sophisticated industrial chemistry hasbeen able to deliver a number of relatively simplenatural products on large scale, chemistry's practicallimits are rapidly approached as functional groupcomplexity and asymmetric centers of the target mol-ecule increase. This complexity should not limit thebiotechnologist. Biotechnology has entered a newrevolution brought about by the advent of genomics,which inherently brings with it the ability for thebiotechnologist to monitor virtually every gene ex-pressed by an organism at a given time, most inter-estingly, in simple organisms such as bacteria or fun-gi under fermentation conditions. With this tool andthe ability to highly control the fermentation envi-ronment with respect to nutrients fed, pH, tem-perature, and oxygenation, it should be possible toengineer optimal organisms dedicated to the pro-duction of a single chemical product. These toolsare also becoming available for plants as well, andwill accelerate the pace of agro-biotechnology [98].
The catalytic complexes providing the vast varietyof molecules are often highly modular in their na-ture, and the speci¢c functionalization of a givenmolecule is determined by the order of arrangementor association of a relatively small number of reac-tion type speci¢c protein domains [99]. This providesthe opportunity for the biotechnologist to carry outcombinatorial biochemistry by rearranging the cata-lytic domains randomly using molecular biologytechniques and creating new biological activities notyet discovered or not yet existing in nature [100].
5.1. Isoprenoids
Isoprenoids constitute a very large group of natu-ral products recruited by humans for a variety of
uses such as imparting £avor or color to foods (zeax-anthines, astaxanthine), fragrance to a variety ofproducts (linalool; US$ 12 000/mt), or as disinfec-tants (camphor; US$ 8700/mt; K-pinene, etc.), med-icines (taxol), or nutrient supplements (e.g., vitaminsA, E, K). In their native role, isoprenoids serve avariety of purposes such as elicitor signals or toxicmolecules in defense against plant pests, pollinatorattractants, antibacterial or antifungal activities, en-zyme cofactors, and other functions. Isoprenoids ofcommodity quantities are usually isolated from plantsources by extraction or are synthesized in part or inwhole by chemical routes [101]. By virtue of beingnatural products, relevant biochemical pathways ex-ist that can be harnessed into fermentation processesby the biotechnology industry. Such processes mighto¡er advantages of production cost, reducing envi-ronmental impact of manufacturing, and providingstereochemically pure products. Recently, the enzymerequired for the biosynthesis of (3R)-linalool wascloned in E. coli [102]. This enzyme was capable ofproducing the desired product in this organism sug-gesting that fermentation may be a route to this spe-cialty chemical.
The cancer therapeutic agent taxol is manufac-tured in a semisynthetic process. Several total syn-theses for taxol have been elaborated; however, theseare not practical for manufacturing due to low over-all enantiomeric yield [103]. For the semisyntheticsynthesis of taxol, the isoprenoid nucleus (baccatinIII) is isolated from the needles of Taxus baccata, ashrub, which grows relatively rapidly. E¡orts, how-ever, are still continuing to produce this compoundin plant cell culture. Another alternative route couldbe to clone the required enzymes into microorgan-isms and produce this valuable drug by fermentation[104,105].
Zeaxanthin and astaxanthin are used in the foodand feed industry to provide desired pigmentation toskins of broiler chickens and the £esh of salmon andtrout in aquaculture, respectively. These food color-ing agents have added value as antioxidants andzeaxanthin provides bene¢ts for the health of theeye [106]. The production of these carotenoids byfermentation is a lucrative challenge because of theirhigh value and appreciable market size. The majorityof these two carotenoids are currently synthesized bychemical methods. It is estimated that fermentation
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 447

could lower cost by 20^35% (market value estimatesfor astaxanthine are at US$ 140 million per year);http://www.mun.ca/seabright/caryeast.html. These es-timates are probably conservative as they assume lowvolumetric yield of the product pigments. Low yieldsobtained in current fermentation processes appear tobe due to inability to channel carbon into the desiredpathway [107,108] and non-optimized fermentationprocesses.
An appropriate alternative to fermentationschemes for vitamin A was recently published. Inorder to help solve a general vitamin A shortageproblem in many parts of the world, Ye et al. [109]genetically modi¢ed rice endosperm (polished rice)by the introduction of several heterologous genes tocomplement existing genes into a complete and func-tioning vitamin A biosynthetic pathway. Regardlessof whether a process is designed as fermentation oras an agro-bioengineering approach, as functionalgenomics tools such as transcriptome and proteomeanalyses become more sophisticated, it will be possi-ble to elucidate the appropriate pathway regulationand circumvent current bottlenecks in plant or mi-croorganism. These compounds are, therefore, cleartargets for the biotechnology industry.
5.2. Polyketides
Investigations of metabolites from the polyketidebiosynthetic pathway have a long and vibrant historywith natural products chemists and biochemists, andthe tradition is continuing with geneticists, molecularbiologists, and biotechnologists. Initial progress to-ward the elucidation of the building blocks of poly-ketide molecules was made through isotopic tracerstudies initiated in the 1950s [110]. The monomersubunits typically consist of acetate, propionate, py-ruvate, and butyrate. Elucidation of the complex en-zymology of these pathways, however, had to awaitthe era of molecular cloning. A comprehensive articlewith key references of the polyketide pathways isprovided by Bentley and Bennett [111].
Early interest in products from this pathway wasdue to the highly colored derivatives of anthraqui-none used as mordant dyes since prehistoric times.The early organic chemicals industry from justaround the turn of the 20th century to the middleof the 20th century almost entirely replaced the im-
portance of these and other naturally occurring dyeswith the ability to manufacture chemically structur-ally similar dyes, initially from coal tar and latercrude oil. As fossil fuels become scarce and environ-mental impact of the chemicals industry needs to beminimized, biotechnology will be ready to bring toprominence the dyes from the polyketide biosyn-thetic pathway once again. Of immediate urgency,however, is the elaboration of new antibiotics, andbioengineering of polyketide pathways appearsto hold signi¢cant promise. This promise has itsbasis in the familiar examples of naturally occur-ring antibiotics from the polyketide biosyntheticpathways such as tetracyclines, erythromycins, nysta-tin and the modular nature of these biosyntheticroutes.
Because of recent large scale DNA sequencing,genes, their organization, and the biochemical orga-nization into clusters of corresponding proteins re-sponsible for the synthesis of numerous antibioticsof the polyketide pathway have been elucidated.From this work the highly modular nature of thissecondary metabolite biochemical pathway has be-come apparent. Polyketides are assembled from sim-ple two to ¢ve carbon atom containing buildingblocks of speci¢c structures. The order in which thesebuilding blocks are assembled by a speci¢c enzymesubunit complex determines the ¢nal structure of theunique compound produced. It has been demon-strated that exchange of the genes coding for theenzyme subunits of varying building block speci¢cityleads to predictable enzyme subunit complexes pro-viding new chemical structures. This observation sug-gests the possibility of producing analog libraries ofantibiotic activities. This approach has been dubbed`combinatorial biochemistry' [112].
Of equal importance and potential is the observa-tion that the biosynthetic pathway for the polyketideanticancer agent epothilone could be transplantedinto a host with better production properties [113].This suggests that these natural products can betransferred to optimal production hosts to increasevolumetric yield and reduce their production cost. Aspolyketide antibiotics in current use are losing e¤-cacy due to the evolution of resistance to them, bio-technology holds the promise of creating new diver-sity and new e¤cacy by recombining existing genesand even creating new genes in the future.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455448

5.3. Non-ribosomal peptides
Another highly modular biosynthetic protein com-plex for the synthesis of a diversity of molecules,including common antibiotics [99,114], starting witha limited set of building blocks, is the non-ribosomalpeptide synthase system. Both D- and L-amino acidsare incorporated into peptides of cyclical or linearstructures containing a relatively small number ofamino acids. These systems are receiving a similarboost in research interest as polyketides are due tothe large expansion of DNA sequences becomingavailable. A functional non-ribosomal peptide syn-thase module has been expressed in E. coli [115].Similar to polyketide research, although there havebeen no reports in the literature as yet, investigatorscan look forward to combinatorial biochemical ap-proaches to generate new diversity. There may alsobe the opportunity for the incorporation of aminoacid analogues into such peptides for even greaterdiversity. Although research in this area lags behindpolyketide research, the rewards for success haveequal potential.
6. Critical success factors
6.1. Titer, yield, and productivity
The last couple of decades have reiterated the mostimportant commercial reality: cost competitivemeans faster, better, and cheaper processes. Manyof the currently commercialized fermentation pro-cesses require de novo synthesis of biomass and themaintenance of other, sometimes unnecessary, cellu-lar functions in addition to the production of thedesired end product. As a consequence, in a numberof processes, greater than 50% of the input carbonsubstrate, typically glucose is expended for cellgrowth and maintenance. In addition, complex car-bon and nitrogen sources are generally necessary asraw materials for cell growth in these processes,thereby complicating and adding costs to down-stream processing of chemical products. In a typicalbatch mode process, each of these biomass buildingfunctions must be repeated with every new fermenta-tion batch, thus wasting valuable raw materials. As aresult, ¢nal titer (measure of product concentration),
or yield (measure of e¤ciency of substrate conver-sion to product), or productivity (measure of volu-metric rate of product formation), and sometimes allthree are limited.
From a kinetic standpoint, carbon £ow into a de-sired biocatalytic pathway is compromised by diver-sion into other metabolic processes. These includesubstrate transport into the cell, feedback regulationdue to inhibition/repression, multiple pathwaybranch points, turnover of intermediates and cofac-tors, and product transport from the cell. The yieldof the end product is thus a complex function ofboth biocatalysis and cell maintenance. Traditionaltools of mutagenesis and selection can only go partway toward optimizing and controlling this process.In addition, appropriate host organisms rarely pos-sess the complete pathway for production of the de-sired intermediate. This dictates that methods mustbe devised for identifying optimal enzymes from oth-er sources to complete the metabolic pathway. Newtechniques must be brought to bear to realize themetabolic goals of developing highly productiveand e¤cient biocatalytic systems.
6.2. Carbon channeling
In the production of chemicals, the ability of bio-logical systems to convert low cost raw materialse¤ciently into chiral products is one of the majoradvantages of biobased over chemical processes.However, one major problem of the biotechnologyapproach, in terms of its cost e¡ectiveness, is theine¤cient use of the raw materials, i.e., the diversionof carbon to metabolites outside of the directed bio-chemical route to the target compound. Pathway en-gineering, which establishes genetic blocks at enzy-matic branch points, has been very successful inlimiting this diversion of resources, but we believe aquantum leap in e¤ciencies could be achieved bymetabolite channeling techniques. By this we meanthe linking of biocatalytic activities (either physicallyor by co-localization) so that pathway intermediatesare transferred from one enzyme to another withoutcomplete equilibration with the surrounding medium[116]. This channeling of pathway intermediates inenzyme networks is not restricted to a speci¢c mo-lecular or physical mechanism. Metabolite channel-ing can also occur as a result of processes resulting in
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 449

the segregation or micro-compartmentation of me-tabolites and enzymes, or it may occur as a resultof a direct transfer of intermediates between catalyticcenters or nodes. This process has been termed `fa-cilitated substrate delivery' since the intermediate isavailable to the next enzyme with a higher probabil-ity than it would be after complete equilibration withthe cellular bulk phase. As a result of this facilitatedtransfer, the catalytic e¤ciency of the overall processis increased without altering the intrinsic catalyticactivities of the components. Numerous examplesof the catalytic advantage a¡orded by multifunction-al biocatalysts have been cited [117]. This structuralorganization of the system may involve multifunc-tional enzymes, multienzyme complexes, dynamic en-zyme associations, or localizations of enzymes onsurfaces. For metabolite channeling, one could useprotein engineering to improve existing methods forco-localizing enzymes or completely new molecularmeans, such as molecular chaperones or sca¡oldingproteins. Channeling may thus represent the initialsteps in the evolution of biotechnology toward cell-free systems. The idea of linking di¡erent enzymeseither via molecular fusion or by docking them toa common supporting element, be it a chaperone-or a cellulosome-type molecule, could lead to fasterand more e¤cient biocatalytic processes. One couldenvision a system in which the conversion of rawmaterial (carbon source) to the desired commercialproduct or to a value-added intermediate could becarried out solely in vitro. This would eliminate thecosts associated with the `wasted' energy required forthe cell mass buildup and maintenance associatedwith traditional fermentation processes.
6.3. Bio-operating systems
Catalytic e¤ciency, sustainable cofactor supply,metabolite channeling, and catalyst life are the coreareas of technology, which enable low cost raw ma-terials conversion to targeted chemical compounds.In other words, we need a bio-operating system(BOS) which will : (1) provide required cofactors toenzyme networks; (2) enhance catalytic e¤cienciesthrough directed channeling of pathway intermedi-ates between sequential enzymatic activities; and(3) provide mechanisms for sustaining and stabilizingbiocatalytic components. The established primary en-
zyme network (i.e., the sequential biocatalytic path-way to the targeted product) and the sustaining en-zyme networks (i.e., those elements of the enzymenetwork which enhance the productive lifespan ofthe process) thus represent the operating system forproduction of chemical products. Application ofBOS for the low cost manufacture of chemicals willalso require innovation and development of ad-vanced bioprocess technologies, which will integratesustainable product synthesis with e¤cient productrecovery.
6.4. Catalytic e¤ciency
A fundamental problem in biotechnology stemsfrom our incomplete knowledge of the enzymes par-ticipating in catalytic pathways as well as the poten-tial variety of pathways that may exist in nature.Since most of our knowledge is based upon enzymesproduced by organisms that are readily cultured inthe laboratory and since we now know that cultur-able organisms represent an in¢nitesimal fraction ofthe naturally occurring species, it is safe to concludethat the natural environment harbors an enormousdiversity of uncharacterized enzymes participating ina myriad of pathways. By using innovative samplingmethods and modifying the use of methods devel-oped for genome sequencing [118] and total proteincharacterizations in whole cells for screening, it willbe possible to penetrate the untapped reservoirs ofenzymes now hidden in unculturable organisms andcryptic pathways. Using modifying procedures suchas directed evolution, gene shu¥ing, molecularbreeding and protein engineering, we can `improve'upon nature's enzymes to optimize them for speci¢cpathway needs. The intent of evolution in biologicalorganisms has been to establish species which aremore ¢t for survival versus ¢t for the synthesis ofchemicals. Over the last 10 years, protein engineeringhas attempted to create this `commercial ¢t' throughrational protein design. We can now apply technol-ogies which will result in the rapid directed evolutionof targeted genes for enzymes having speci¢c indus-trial applications. At the heart of these technologiesare selection procedures which ensure the enrich-ment, from randomly mutagenized populations ofthe targeted gene, of gene activities with `improved'biocatalytic performance. Enriched populations can
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455450

be re-mutagenized, challenged, and enriched againunder selective conditions. In this way, it is possibleto speed up and direct evolution in a microorganismto establish a unique combination of mutations in atargeted gene for an industrially signi¢cant biocata-lyst.
6.5. Sustainable cofactor supply
Enzymatic cofactors are required for many bioca-talytic reactions in the manufacture of chemicals. Re-generation of cofactors can be accomplished bychemical, electrochemical, photochemical, and enzy-matic means [119,120]. Enzymatic processes are themost e¤cient and appear to have the greatest like-lihood for successful application in large scale bio-manufacturing processes. Current whole cell aerobicprocesses which depend on NADH and NADPHcofactors for hydride transfer reactions lose most ofreducing equivalents to oxidation for energy genera-tion. Biological processes, which carry out a series ofreactions requiring movement of intermediates fromthe cytoplasmic to the extracellular compartments,often waste reducing equivalents in the extracellularprocesses through oxidation via molecular oxygen.Pathway engineering o¡ers the opportunity of recov-ering these reducing equivalents through reduction ofNAD(P) to NAD(P)H. By creating reaction pairs,where available, of oxidation and reduction process-es, one can also minimize diversion of metabolic £owdown unproductive pathways. In the case where suchpairing within the required pathway is not possible,the generation of reducing equivalents can be at-tempted with co-substrates whose products will notimpact the ¢nal material [120]. Such methods existfor co-substrates, e.g., formic acid (formate dehydro-genase), whose conversion to CO2 nets one NADH,and methanol, which nets three NADH. Cofactorspeci¢city modi¢cation represents an additional op-portunity for positively in£uencing cofactor supplyrequirements in biocatalytic processes. One can engi-neer cofactor binding pockets utilizing protein engi-neering approaches coupled with sequence informa-tion from related NAD(P)H requiring enzymes.Nishiyama et al. [121] have shown that alterationsin the coenzyme speci¢city of malate dehydrogenasefrom Thermus £avus can be accomplished by site-di-rected mutagenesis with minimal impact on catalytic
e¤ciencies. Alteration of the coenzyme speci¢cityfrom NADPH to NADH in certain cases will makeit possible to nutritionally regenerate cofactors withless expensive substrates, thereby reducing manufac-turing costs.
6.6. High density reactors
Today, nearly all large scale chemical manufactur-ing processes are operated in continuous modeswhereas current industrial biological processes aremostly batch or fed-batch type processes. This dispar-ity in processing maturity and capabilities puts bio-logical processes at a competitive disadvantage. Con-tinuous biological processes will permit the use ofhigh biocatalyst loading and increase the volumetricproductivity of the bioreactors, increase the utiliza-tion e¤ciency of all equipment, and improve conser-vation of energy and water. To establish cost com-petitive routes to chemicals derived from sequentialbiocatalytic systems and to reduce the capital re-quirements for commercialization of these products,processing technologies should focus on: (1) immo-bilized whole cells, or immobilized enzyme reactors[122], and (2) advanced recovery methodologieswhich minimize processing steps as well as wasteby-products [123].
Because of its proven £exibility, the basic stirredtank reactor is still the workhorse of the fermenta-tion industry. Although simple to operate and con-trol, the design su¡ers from oxygen transfer and heatdissipation ine¤ciencies and places real limits on thebioreactor productivity and scale of operation. Weneed bioprocesses amenable to scale up by large fac-tors, such as £uidized or packed column bioreactorsusing immobilized whole cells or enzymes.
6.7. Minimizing capital
Fermentation processes generally require largereactor vessels, a high amount of air/oxygen, andthat in turn requires a high amount of electricity.Metabolism generates a large amount of heat andthus requires cooling systems. They also releasehigh levels of CO2 into the environment and gener-ally produce low aqueous titers, necessitating com-plex recovery schemes. Clearly, integration with pro-cess engineering technologies is required to realize
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 451

the capital targets. The key to overcoming many ofthese barriers is in rational catalyst design, a processpreviously accessible only to conventional chemicalsynthesis. Through advances in biotechnology, majormetabolic ine¤ciencies are becoming addressable.We have the capability to borrow enzyme systemsfrom organisms and combine them into one, opti-mized biocatalytic host. Through control of cellularmetabolic processes, we can not only optimize prod-uct formation but also control waste product gener-ation and other accessory processes, such as simplerrecovery steps, thereby shrinking the size of the fac-tory necessary to complete the synthesis of the tar-geted product (Fig. 10).
6.8. Sustainability impact
Bioprocesses often rely on excessive amounts ofreagents for shifting equilibrium and controllingpH, involve complete isolation of intermediateswith limited use of recycles, and employ short-lived,unrecyclable biocatalysts. Although acceptable forthe production of low volume, high value products,these approaches tend to be expensive, wasteful ofresources, and generally have high environmental im-pact. Identi¢cation of the better alternatives is a keyfactor in successful commercialization of a productunder severe competitive pressures. The ultimate goalis to systematize biochemical processes which havelower environmental impact and which can competesuccessfully with conventional chemical processes in
larger volume, lower value markets. The life cycleinventory (LCI) concept can be used to comparethe environmental performance of bio-manufacturingprocesses. The concept recognizes that all life cyclesteps (raw material acquisition; manufacturing, pro-cessing, and formulation; transportation and distri-bution; use, re-use and maintenance; and recyclingand waste management) have economic, environmen-tal, and energy impacts. However, use of the sustain-ability impact is more robust for evaluation of pro-cess design and comparison since it takes intoaccount social, economic, and environmental aspects.The sustainability impact therefore yields betterunderstanding of the consequences of technologychoices. By addressing the undesirable consequencessuch as carbon ine¤ciencies in the form of carbondioxide emission and energy ine¤ciencies in the formof fossil fuel usage, only renewable feedstock basedtechnologies can move us toward a sustainable soci-ety [124].
7. Concluding remarks
Relative to the title of this review, successful im-plementation of pathway engineering requires morethan a novel research ¢nding or development ofpromising new molecular biology methods for straindevelopment. Multiple disciplines including chemis-try and chemical engineering must be integrated toimplement a commercially viable process. Certainly,
Fig. 10. Shrinking the size of the factory. Use of biocatalysts in the synthesis of chemical products a¡ords opportunities for decreasingcapital investments in large chemical factories.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455452

existing commercial processes have been impactedfavorably by pathway engineering modi¢cations butthe introduction of new biotechnology based process-es awaits further developments [125]. This break-through will occur when the economics of biopro-cesses are compelling and when new biomaterialsare developed which o¡er value to consumers andsociety. The impact of genomics, molecular breedingtechnologies and bioinformatics o¡ers great potentialto accelerate the broad use of pathway engineering inthe chemical and related industries and bring to real-ity the biore¢neries of the future sooner than later[126].
Acknowledgements
Partial funding of `Continuous Biocatalytic Sys-tems for the Production of Chemicals from Renew-able Resources program' at Genencor Internationalby a NIST/ATP grant (70NANB5H1138) is acknowl-edged. We also thank Gregg Whited, Alice Caddow,and Roopa Ghirnikar for helpful suggestions.
References
[1] DOE/GO-10098-385, Plant/Crop-Based Renewable Resour-ces 2020, 1998, pp. 1^24.
[2] R.L. Hinman, BioTechnology 9 (1991) 533.[3] American Chemical Society, Technology Vision 2020, 1996.[4] A. Berry, Trends Biotechnol. 14 (1996) 250^256.[5] M. Ikeda, R. Katsumata, Appl. Environ. Microbiol. 65
(1999) 2497^2502.[6] H. Ito, K. Sato, H. Enei, Y. Hirose, Agric. Biol. Chem. 54
(1990) 823^824.[7] K. Backman, M.J. O'Connor, A. Maruya, E. Rudd, D.
McKay, R. Balakrishnan, M. Radjai, V. DiPasquantonio,D. Shoda, R. Hatch, K. Venkatsubramanian, Ann. NYAcad. Sci. 589 (1990) 16^24.
[8] B.D. Ensley, B.J. Ratzkin, T.D. Osslund, M.J. Simon, L.P.Wackett, D.T. Gibson, Science 222 (1983) 167^169.
[9] K.M. Draths, J.W. Frost, J. Am. Chem. Soc. 113 (1991)9361^9363.
[10] K.M. Draths, J.W. Frost, J. Am. Chem. Soc. 116 (1994)399^400.
[11] K.M. Draths, T.L. Ward, J.W. Frost, J. Am. Chem. Soc.114 (1992) 9725^9726.
[12] P.L. Gray, R. Proc. Proc. B 102 (1927) 263^280.[13] D. Murdock, B.D. Ensley, C. Serdar, M. Thalen, Bio/Tech-
nology 11 (1993) 381^386.
[14] T. Dodge, M. Pepsin, A. Berry, in: Proceedings of the Thir-teenth Forum for Applied Biotechnology, 1999, pp. 281^288.
[15] W. Weyler, T.C. Dodge, J.J. Lau¡, D.J. Wendt, US Patent5866396, 1999.
[16] US Patent 5143833.[17] N.P. Ngheim, B.H. Davison, B.E. Suttle, G.R. Richardson,
Appl. Biochem. Biotechnol. 63-65 (1997) 565^576.[18] J.G. Zeikus, M.K. Jain, P. Elankovan, Appl. Microbiol.
Biotechnol. 51 (1999) 545^552.[19] N. Sumaulov, R. Lamed, S. Lowe, J.G. Zeikus, Appl. Envi-
ron. Microbiol. 57 (1991) 3013^3019.[20] US Patents 5143834, 5168055, 5521075.[21] US Patent 5504004.[22] US Patent 5573931.[23] C.S. Millard, Y.P. Chao, J.C. Liao, M.I. Donnelly, Appl.
Environ. Microbiol. 62 (1996) 1808^1810.[24] L. Stols, M.I. Donnelly, Appl. Environ. Microbiol. 63 (1997)
2695^2701.[25] US Patent 5869301.[26] M.I. Donnelly, R. Chartterjee, C.S. Millard, K.M. Cham-
pion, D.P. Clark, SIM meeting abstract Aug. 1^5, Arlington,VA, 1999.
[27] J.G. Zeikus, Annu. Rev. Microbiol. 34 (1980) 423^464.[28] M.M. Bianchi, M. Tizzani, M. Destruelle, L. Frontalli, M.
Wesoloweskii-louvel, Mol. Microbiol. 19 (1996) 27^36.[29] D. Porro, M.M. Bianchi, L. Brambilla, R. Menghini, D.
Bolzani, V. Carrera, J. Lievense, C.-L. Liu, B.M. Ranzi, L.Frontali, L. Alberghina, Appl. Environ. Microbiol. 65 (1999)4211^4215.
[30] PCT WO 99/14335.[31] T. Reichstein, A. Grussner, Helv. Chim. Acta 17 (1934) 311.[32] WO Patent 9964618A1.[33] Y. Saito, Y. Ishii, H. Hayashi, Y. Imao, T. Akashi, K.
Yoshikawa, Y. Noguchi, S. Soeda, M. Yoshida, M. Niwa,J. Hosoda, K. Shimomura, Appl. Environ. Microbiol. 63(1997) 454^460.
[34] D.J. Steward, Nature 183 (1959) 1133.[35] D.W. Dye, New Zealand J. Sci. 7 (1964) 261.[36] D.A. Kita, US Patent 4555812, 1977.[37] T. Sonoyama, S. Yagi, B. Kageyama, M. Tanimoto, EP
46284, 1982.[38] T. Sonoyama, H. Tani, B. Kageyama, K. Kobayashi, T.
Honjo, S. Yagi, US Patent 3963574, 1976.[39] T. Sonoyama, B. Kageyama, S. Yagi, K. Mitsushima, Agric.
Biol. Chem. 51 (1987) 3039.[40] T. Sonoyama, S. Yagi, B. Kageyama, US Patent 4543331,
1985.[41] S. Anderson, C.B. Marks, R. Lazarus, J. Miller, K. Sta¡ord,
J. Seymour, W. Light, W.H. Rastetter, D.A. Estell, Science230 (1985) 144.
[42] D.A. Estell, D.R. Light, W.H. Rastetter, R.A. Lazarus, J.V.Miller, Eur. Patent 132308, 1985.
[43] US Patents 4757012, 4758514, 5004690, 5008193, 5032514.[44] T. Fowler, S. Causey, WO 98/59054.[45] R.A. Lazarus, R.K. Sta¡ord, J.L. Seymour, M.S. Dennis,
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 453

M.A. Lazarus, E.J.L. Huges, H.I. Miller, C.B. Marks, S.Anderson, GIM90, Strassbourg, 1073, 1990.
[46] R.A. Lazarus, J.L. Seymour, K. Sta¡ord, C.B. Marks, S.Anderson, in: C.L. Hershberger, S.W. Queener, G. Hege-man (Eds.), Genetics and Molecular Biology of IndustrialOrganisms, American Society for Microbiology, Washing-ton, DC, 1989, pp. 187.
[47] S. Trusdell, J.C. Sims, P.A. Boerman, J.L. Seymour, R.A.Lazarus, J. Bacteriol. 173 (1991) 6651.
[48] J.S. Cha, C. Pujol, C.I. Kado, Appl. Environ. Microbiol. 63(1997) 71.
[49] D.Y. Yum, J.G. Pan, K.I. Chang, Y.C. Shin, J. Bacteriol.179 (1997) 6566.
[50] D.Y. Yum, J.G. Pan, K.I. Chang, Y.C. Shin, J. Bacteriol.180 (1998) 5984.
[51] J.V. Miller, D.A. Estell, R.A. Lazarus, J. Biol. Chem. 262(1987) 9016.
[52] S. Khurana, D.P. Powers, S. Anderson, M. Blaber, Proc.Natl. Acad. Sci. USA 95 (1998) 6768.
[53] T. Dodge, G. England, M. Kumar, L. Maheras, BIOT 366,ACS Meeting San Francisco, CA, 2000.
[54] PCT/US99/30918.[55] PCT/US99/28669.[56] PCT/US99/23862.[57] L.O. Ingram, P.F. Gomez, X. Lai, M. Moniruzzaman, B.E.
Wood, L.P. Yomano, S.W. York, Biotechnol. Bioeng. 58(1998) 204^213.
[58] A.T. Bull, H. Dalton (Eds.), Comprehensive Biotechnology,vol. 1, Pergamon Press, London, 1985.
[59] L.O. Ingram, F. Alterthum, K. Ohta, D.S. Beall, in: G.E.Piece (Ed.), Genetic Engineering of Escherichia coli and Oth-er Enterobacteria for Ethanol Production, Developments inIndustrial Microbiology, vol. 31, 1990.
[60] J. Hugenholtz, M. Kleerebezem, FEMS Microbiol. Lett. 179(1999) 492^497.
[61] A. Ben-Amotz, M. Avron, Basic Life Sci. 14 (1979) 91^99.[62] A. Steinbuchel, M. Muller, Mol. Biochem. Parasitol. 20
(1986) 45^55.[63] J. Albertyn, S. Hohmann, J.M. Thevelein, B.A. Prior, Mol.
Cell. Biol. 14 (1994) 4135.[64] B. Petrovska, E. Winkelhausen, Kuzmanova, Can. J. Micro-
biol. 45 (1999) 695-699.[65] H.T. Wang, P. Rahaim, P. Robbins, R.R. Yocum, J. Bacter-
iol. 176 (1994) 7091^7095.[66] K. Larason, R. Ansell, P. Eriksson, L. Adler, Mol. Micro-
biol. 10 (1993) 1101^1111.[67] M. Rep, J. Albertyn, J.M. Thevelein, P. Bernard, S. Hoh-
mann, Microbiology 145 (1999) 715^727.[68] R. Ansell, K. Granath, S. Hohmann, J. Thevelein, L. Adler,
EMBO J. 16 (9) (1997) 2179^2187.[69] T. Maeda, M. Takekawa, H. Saito, Science 269 (1995) 554^
558.[70] J.L. Brewster, T. de Valor, N.D. Dwyer, E. Winter, M.C.
Gustin, Science 259 (1993) 1760^1763.[71] J. Norbeck, A.-K. Pahlman, N. Akhtar, A. Blomberg, L.
Adler, J. Biol. Chem. 271 (1996) 13875^13881.
[72] F.A. Skraly, D. Cameron, Arch. Biochem. Biophys. 349(1998) 27^35.
[73] E. Van Schaftingen, A.J. Van Laere, Eur. J. Biochem. 148(1985) 399^404.
[74] S. Michnick, J.L. Roustan, F. Remize, P. Barre, S. Dequin,Yeast 13 (9) (1997) 783^793.
[75] D. Cameron, F.A. Skraly, WO 9707199, 1997.[76] B.A. Bulthuis, A.A. Gatenby, S.L. Haynie, A.K.-H. Hsu, R.
Lareau, WO 9821340, 1998.[77] V. Nair, M.S. Payne, D.E. Trimbur, F. Valle, WO 9928480,
1999.[78] A.A. Gatenby, S.L. Haynie, Nagarajan, Method for the Pro-
duction of 1,3-Propanediol by Recombinant Organisms, WO9821339, 1998.
[79] A. Freund, Monatsschr. Chem. 2 (1881) 63^641.[80] M.N. Mickelson, C.H. Werkman, Enzymologia 8 (1940)
252^256.[81] K. Menzel, A.-P. Zeng, W.-D. Deckwer, Enzyme Microbiol.
Technol. 20 (1997) 82^86.[82] R. Boenigk, S. Bowien, G. Gottschalk, Appl. Microbiol.
Biotechnol. 38 (1993) 453^457.[83] H. Biebl, Appl. Microbiol. Biotechnol. 35 (1991) 701^705.[84] H. Biebl, S. Marten, H. Hippe, W.-D. Deckwer, Appl. Mi-
crobiol. Biotechnol. 36 (1992) 592^597.[85] A.M. Held, The Fermentation of Glycerol to 1,3-Propane-
diol by Klebsiella pneumoniae, Master's thesis, University ofWisconsin-Madison, Madison, WI, 1996.
[86] K. Ahrens, K. Menzel, A.-P. Zeng, W.-D. Deckwer, Biotech-nol. Bioeng. 59 (1998) 54^552.
[87] R. Boenigk, S. Bowien, G. Gottschalk, Appl. Microbiol.Biotechnol. 38 (1993) 453^457.
[88] H. Biebl, S. Martin, Appl. Microbiol. Biotechnol. 44 (1995)15^19.
[89] S. Abbad-Andaloussi, A.J. Maginot-Durr, E. Petitdemange,H. Petitdemange, Appl. Environ. Microbiol. 61 (1995) 4413^4417.
[90] M. Seyfried, R. Daniel, G. Gottschalk, J. Bacteriol. 178(1996) 5793^5796.
[91] T. Tobimatsu, H. Kajiura, M. Yunoki, M. Azuma, T. Tor-aya, J. Bacteriol. 181 (1999) 4100^4113.
[92] R. Daniel, R. Boenigk, G. Gottschalk, J. Bacteriol. 177(1995) 2151^2156.
[93] I.T. Tong, H.H. Liao, D.C. Cameron, Appl. Environ. Mi-crobiol. 57 (1991) 3541^3546.
[94] I.T. Tong, D.C. Cameron, Appl. Biochem. Biotechnol. 34-35(1992) 149^159.
[95] D.C. Cameron, N.E. Altaras, M.L. Ho¡man, A.J. Shaw,Biotechnol. Prog. 14 (1998) 116^125.
[96] F.A. Skraly, B.L. Lytle, D.C. Cameron, Appl. Environ. Mi-crobiol. 64 (1998) 98^105.
[97] S.L. Haynie, L.W. Wagner, Process for Making 1,3-Pro-panediol from Carbohydrates Mixed Microbial Culture,WO 35799, 1996.
[98] T. Mozo, K. Dewar, P. Dunn, J.R. Ecker, S. Fischer, S.Kloska, H. Lehrach, M. Marra, R. Martienssen, S. Meier-Ewert, T. Altmann, Nat. Genet. 22 (1999) 271^275.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455454

[99] C.L. Bender, F. Alarcon-Chaidez, D.C. Gross, Microbiol.Mol. Biol. Rev. 63 (1999) 266^292.
[100] M. Bycroft, K.J. Weissman, J. Staunton, P.F. Leadlay, Eur.J. Biochem. 267 (2000) 520^526.
[101] P.J. Teisserire, Chemistry of Fragrant Substances, VCHPubl., New York, 1994.
[102] J.W. Jia, J. Crock, S. Lu, R. Croteau, X.Y. Chen, Arch.Biochem. Biophys. 372 (1999) 143^149.
[103] K.C. Nicolaou, Z. Yang, J.J. Liu, H. Ueno, P.G. Nanter-met, R.K. Guy, C.F. Claiborne, J. Renaud, E.A. Coula-douros, K. Paulvannan et al., Nature 367 (1994) 630^634.
[104] S. Jha, D. Sanyal, B. Ghosh, T.B. Jha, Planta Med. 64(1998) 270^272.
[105] K.X. Huang, Q.L. Huang, M.R. Wildung, R. Croteau, A.I.Scott, Protein Expr. Purif. 13 (1998) 90^96.
[106] G.J. Handelman, Z.D. Nightingale, A.H. Lichtenstein, E.J.Schaefer, J.B. Blumberg, Am. J. Clin. Nutr. 70 (1999) 247^251.
[107] C.W. Wang, M.K. Oh, J.C. Liao, Biotechnol. Bioeng. 62(1999) 235^241.
[108] Y. Miura, K. Kondo, T. Saito, H. Shimada, P.D. Fraser,N. Misawa, Appl. Environ. Microbiol. 64 (1998) 1226^1229.
[109] X. Ye, S. Al-Babili, A. Kloti, J. Zhang, P. Lucca, P. Beyer,I. Potrykus, Science 287 (2000) 303^305.
[110] K.B.G. Torssell, Natural Products Chemistry, John Wileyand Sons, New York, 1983.
[111] R. Bentley, J.W. Bennett, Annu. Rev. Microbiol. 53 (1999)411^446.
[112] A.F. Marsden, B. Wilkinson, J. Cortes, N.J. Dunster, J.Staunton, P.F. Leadlay, Science 279 (1998) 199^202.
[113] L. Tang, S. Shah, L. Chung, J. Carney, L. Katz, C. Khosla,B. Julien, Science 287 (2000) 640^642.
[114] R. Kittelberger, M. Pavela-Vrancic, H. von Dohren, FEBSLett. 461 (3) (1999) 145^148.
[115] J. Ku, R.G. Mirmira, L. Liu, D.V. Santi, Chem. Biol. 4(1997) 203^207.
[116] N. Stebbing, in: L. Noyer, F. Lynen, K. Mothes (Eds.),Cell Compartmentation and Metabolic Channeling, Else-vier/North-Holand Biomedical Press, Amsterdam, 1980.
[117] L. Bulow, K. Mosbach, Trends Biotechnol. 9 (1991) 226^231.
[118] A. Milosavljevic, J. Comput. Biol. 2 (1995) 355^370.[119] K.J. Mosbach, Chem. Tech. Biotechnol. 32 (1982) 179^188.[120] H.K. Chenault, G.M. Whitesides, Appl. Biochem. Biotech-
nol. 14 (1987) 147^197.[121] Nishiyama et al., J. Biol. Chem. 268 (1993) 4656^4660.[122] A. Constantinides, G. Chotani, Ann. NY Acad. Sci. 434
(1984).[123] US Patent 4, 435,595.[124] T.U. Gerngross, Nat. Biotechnol. 17 (1999) 541^544.[125] D. Wilke, Appl. Microbiol. 52 (1999) 135^145.[126] L. Lynd, C.E. Wyman, T.E. Gerngross, Biotechnol. Prog.
15 (1999) 777^793.
BBAPRO 36306 21-12-00 Cyaan Magenta Geel Zwart
G. Chotani et al. / Biochimica et Biophysica Acta 1543 (2000) 434^455 455




















