
THE CALL OF THE SRI LANKAN GOLDEN GECKO CALODACTYLODES ILLINGWORTHORUM, ECOLOGICAL PARALLEL OF THE...
16
Bioacoustics The International Journal of Animal Sound and its Recording, 2008, Vol. 18, pp. 35–49 0952-4622/08 $10 © 2008 AB Academic Publishers THE CALL OF THE SRI LANKAN GOLDEN GECKO CALODACTYLODES ILLINGWORTHORUM, ECOLOGICAL PARALLEL OF THE FAN-TOED GECKOS, GENUS PTYODACTYLUS (REPTILIA: SAURIA: GEKKONIDAE). YEHUDAH L. WERNER 1 , EVGENY CHERNIS 2 AND RONI GRANOT 3 1 Department of Evolution, Ecology and Systematics, The Hebrew University of Jerusalem, 91904 Jerusalem, Israel (y[email protected]) 2 Department of Science Teaching, The Hebrew University of Jerusalem, 91904 Jerusalem, Israel ([email protected]) 3 The Music Cognition Research Laboratory, Department of Musicology, The Hebrew University of Jerusalem, 91905 Jerusalem, Israel ([email protected]) ABSTRACT Based on nine calls of Calodactylodes illingworthorum recorded in the wild in Sri Lanka, the ‘spontaneous’ call of the species is a very rapid series of 4-7 roughly similar chirps. Call parameters are compared to those of Ptyodactylus guttatus of Israel because the species are similar in morphology (size, proportions, digit shape, scutellation, coloration), ecology (rupicolous, nocturnal), and reproductive biology (clutches of two eggs, stuck to rock, in ‘rookeries’). The calls of the two species, despite similarity in basic structure, greatly differ in the number of chirps, chirp repetition rate, total duration, dominant frequency, and relative intensity level. The functional implications of these differences are briefly considered. Keywords: Calodactylodes illingworthorum, gecko, multiple-chirp call, Ptyodactylus, temporal and spectral analysis, vocalization. INTRODUCTION In the animal world, acoustic long-range communication has the advantages of not being dependent on illumination and not being obstructed by intervening objects, while enabling the localization of the caller (Frankenberg & Werner 1992). However, the fidelity of transmission of acoustic signals through natural environments may be affected by various physical and biotic factors (Michelsen & Larsen 1983), though the transmission differences between habitat types may be smaller than often thought (Wiley & Richards 1978). Presumed adaptations of animal vocalizations to their specific acoustic environment have been reported especially for birds (e.g., Morton
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of THE CALL OF THE SRI LANKAN GOLDEN GECKO CALODACTYLODES ILLINGWORTHORUM, ECOLOGICAL PARALLEL OF THE...

BioacousticsThe International Journal of Animal Sound and its Recording, 2008, Vol. 18, pp. 35–490952-4622/08 $10© 2008 AB Academic Publishers
THE CALL OF THE SRI LANKAN GOLDEN GECKO CALODACTYLODES ILLINGWORTHORUM, ECOLOGICAL PARALLEL OF THE FAN-TOED GECKOS, GENUS PTYODACTYLUS (REPTILIA: SAURIA: GEKKONIDAE).
YEHUDAH L. WERNER1, EVGENY CHERNIS2 AND RONI GRANOT3
1Department of Evolution, Ecology and Systematics, The Hebrew University of Jerusalem, 91904 Jerusalem, Israel ([email protected]) 2 Department of Science Teaching, The Hebrew University of Jerusalem, 91904 Jerusalem, Israel ([email protected]) 3 The Music Cognition Research Laboratory, Department of Musicology, The Hebrew University of Jerusalem, 91905 Jerusalem, Israel ([email protected])
ABSTRACT
Based on nine calls of Calodactylodes illingworthorum recorded in the wild in Sri Lanka, the ‘spontaneous’ call of the species is a very rapid series of 4-7 roughly similar chirps. Call parameters are compared to those of Ptyodactylus guttatus of Israel because the species are similar in morphology (size, proportions, digit shape, scutellation, coloration), ecology (rupicolous, nocturnal), and reproductive biology (clutches of two eggs, stuck to rock, in ‘rookeries’). The calls of the two species, despite similarity in basic structure, greatly differ in the number of chirps, chirp repetition rate, total duration, dominant frequency, and relative intensity level. The functional implications of these differences are briefly considered. Keywords: Calodactylodes illingworthorum, gecko, multiple-chirp call, Ptyodactylus, temporal and spectral analysis, vocalization.
INTRODUCTION
In the animal world, acoustic long-range communication has the advantages of not being dependent on illumination and not being obstructed by intervening objects, while enabling the localization of the caller (Frankenberg & Werner 1992). However, the fidelity of transmission of acoustic signals through natural environments may be affected by various physical and biotic factors (Michelsen & Larsen 1983), though the transmission differences between habitat types may be smaller than often thought (Wiley & Richards 1978). Presumed adaptations of animal vocalizations to their specific acoustic environment have been reported especially for birds (e.g., Morton

36
1975) but in some other cases could not be confirmed (e.g., Daniel & Blumstein 1998). The situation in lizards is unknown. Among lizards, the most vociferous family is the Gekkonidae, mostly of which are active nocturnally (Kiester 1977; Tang et al. 2001; Bauer 2002) and many of which boast a vocal repertoire of up to six different vocal expressions (Frankenberg & Werner 1992). Many geckos are also known to possess acute hearing, at least as assessed by certain methods (Werner 1976; Wever 1978). However, the communicatory role of gecko vocalizations has been investigated in few cases (Frankenberg & Werner 1992) and the adaptation of gecko vocalizations to the transmitting medium cannot even be considered before the vocalizations of more species become known in terms of acoustical parameters. Therefore we report here on vocalizations of the Sri Lankan golden gecko.
The Sri Lankan golden gecko Calodactylodes illingworthorum Deraniyagala, 1953a is endemic to Sri Lanka, occurring only or mainly within an area of roughly 20,000 km2 in the island’s south-east (Wickramasinghe & Somaweera 2002). Adults measure about 75mm, maximally about 85mm, RA (Rostrum-Anus, Werner 1971) with a tail of about 110 PERCRA (percents of RA, Werner 1971) and long limbs. The dorsal skin is characterized by small flat scales interspersed with larger flat tubercles. Males have small numbers of femoral and preanal pores (Deraniyagala 1953b; Bauer & Das 2001). The digits of the very similar congener from India, C. aureus Beddome, 1870, are slender, bearing a terminal bi-lobed scansorial expansion, with digits 2-5 (of hand and foot) having an additional subterminal pair of scansors (Russell & Bauer 1989). The dorsal coloration is mottled golden-brown with light grey markings (cross-bands on limbs and tail) and a vertebral series of six darker blotches but the shades vary with location, sex, age, and “mood” (Bauer & Das 2001; Goonewardene et al. 2003).
The geckos of this species inhabit large boulders, rock faces and caves, especially in relatively “dry” locations (i.e. <1,900mm annual rainfall!), within either fire savannah or monsoon forest. They are agile and can leap across 30cm spaces between rock faces. Being nocturnal, in daytime they remain in caves and rock crevices but at night may forage in nearby trees. They usually occur as groups of 3-11 individuals and it has been claimed that one male is dominant and conspicuous through its more intense yellow throat patch. Fights, seemingly territorial, have been observed. Other observations indicate that pair mates may stick together. Their typical calls are rarely heard in daytime; vocal activity climaxes in the late afternoon or early evening and again towards dawn. They oviposit in communal sites in which several females participate during successive years, mostly in caves or dark archways with [reportedly] stable temperature of 28oC (Wickramasinghe & Somaweera 2002; Goonewardene et al. 2003; de Silva et al. 2004b).

37
Thus, as far as known, Calodactylodes illingworthorum seems to closely resemble in both morphology and biology its only congener, C. aureus of India, described in some detail by Bauer & Das (2001). Collectively, the spatially restricted and still relatively little-known Calodactylodes geckos closely resemble in many morphological and biological traits some common and better-known Middle Eastern geckos of the genus Ptyodactylus, as will be detailed later. Therefore the question arises, whether their vocalizations, too, are similar.
MATERIALS AND METHODS
On the occasion of the Fourth World Congress of Herpetology, Bentota, Sri Lanka, 3-9 December 2001, John Rudge (Global Gecko Association) and the first author, guided by Shantha Karunarathne, visited a Calodactylodes habitat near Bibile, Sri Lanka (7o09′N 81o14′E, Altitude 300-305 m) on 2 December 2001. This date would be within the reproductive season because in early January Bauer & Das (2000) found all stages of embryonic development, from shelled oviductal eggs to near-term embryos. Vocalizations were recorded in one boulder cluster a few tens of meters in diameter, where we saw only several geckos of this species, and the typical calls, resembling those of Calodactylodes aureus (Bauer & Das 2000), were recognized by our guide. Recording proceeded between 1000 and 1100h at approx. 27oC (air), on a microcassette recorder with built-in microphone (Panasonic model RN-405, with Panasonic microcassette at the higher tape speed, 2.4 cm/sec). The recording was sampled to digital form with CSL (Computerized Speech Lab, Model 4300B version 2.5.1) hardware and software (sampling rate: 44100Hz) and then analyzed with CSL. The calculations and statistics used MS Excel 2000.
We analyzed nine multiple-chirp calls (following the terminology of Marcellini 1974, 1977) in the temporal and frequency domains. One call yielded only partial information due to pollution by human voice. In the remaining eight calls, the intensity domain could only be analyzed in relative terms as due to the unknown acoustic properties of the recorder, absence of dB meter, and unmeasured level of ambient noise, we cannot estimate the absolute values of emitted voice energy.
Temporal domain
To reveal the temporal properties of each call and its constituent chirps we used the amplitude time envelopes (in CSL “Energy Contour”) as the ratio of energy (dB) to time (msec). The Energy Contour was sampled twice, first with the frame of 1 msec and advance steps of

38
1 msec to accurately derive the chirp durations from the chirp onset-to-onset of the next chirp and to define the relative intensity level of chirps within a call. The smoothed Energy Contour (5msec frame, 1msec steps) was used to derive different temporal parameters, as explained later. Because the ambient noise properties were unknown it was impossible to pinpoint when each chirp ended. Therefore, rather than measuring the onset to offset duration of the chirp we measured the time during which the sound energy of the chirp continuously remained within 20 dB from the chirp energy peak. We assumed that this 10-fold decay would include most of the information significant for communication. This parameter, among others, was derived from the smoothed Energy Contour.
Frequency domain
We first examined the tone structure of vocalizations in spectrograms (sonograms), providing a qualitative visual impression of the temporal and spectral complexity of each call. The power spectrums for each chirp yielded quantitative information of the frequency domain.
Spectrogram PropertiesWe used a 1024 points (63.09 Hz bandwidth) frame size with Hamming filter smoothing. The frequency axis was trimmed to a 0-6000 Hz range, because the burst of energy occurred much below 6000Hz, as seen in power spectrums, and the high-frequency fidelity of the recorder was unknown. In the spectrogram the darkest gray represents maximum energy and each step (shade) of diminishing darkness indicates a 5dB energy decrease.
Power Spectrum PropertiesThe spectrum for each chirp was constructed starting from the point of maximum energy as sampled on Energy Contour, using 1024 points Fast Fourier Transform with Hamming filter window and without smoothing. Each spectrum was then homomorphically smoothed (Hopp et al. 1998) using the Cepstrum function. The Cepstrum allowed us to check whether chirps have a significant fundamental pitch as apparent from some spectrograms. The smoothed spectra served to identify the frequencies with maximum energy and to compare among chirps.
As a result of analyzing the spectrograms we decided to identify two frequencies with maximum energy as derived from the smoothed spectrum. The first peak frequency has absolute maximum energy within the smoothed spectrum. To ensure that the second peak frequency is separate of the first we defined it as a peak separated

39
by an “energy valley” of at least -3 dB from the absolute maximum peak.
Relative chirp intensity
To further explore the structure of the calls we compared the intensity level (dB) of the chirps within the call. In each call we assigned to the energy peak of the most intense chirp the value 0 dB as base line, irrespective of its position in the train of chirps. The other chirps then received negative dB values expressing their difference (delta) from the most intense chirp.
RESULTS
The habitat consisted of groups of large granite boulders in a mixed monsoon forest. Occasionally the geckos were calling, their multiple-chirp calls (we noted no others) like high-pitched rapid chatter, loud enough to be heard at some distance, perhaps about as loud as average human conversational speech (order of magnitude of 70 dB SPL, i.e. relative to 20 μPa). Although sometimes geckos were seen, none were observed calling, they were calling only or mainly from dark shelters in the rock system. Sometimes a call quickly followed another that had emanated from a nearby location (order of magnitude of 1 m apart) so that they appeared to be responding to each other.
Each of nine calls recorded (probably from at least four individuals) comprised a rapid train of chirps, upon analysis seen to lack proper silent intervals. Seven of the calls comprised 4-5 chirps, one 6 and the ninth (for most parameters excluded) 7. Each chirp rose almost promptly to maximal energy, with a rise time of 1-2 msec, then gradually receded but as it waned, the next chirp already burst. Therefore we exclude the rise time from our report of the temporal domain.
Temporal domain
Temporal parameters of the chirp and of the call varied among chirps and among calls (Table 1) but they did not vary in parallel. In six of eight calls, onset-to-onset duration of chirps gradually increased along the call. In four of these six (and also in one of the two other calls), estimated chirp duration (ESD) rose in each successive chirp up to a maximum in the middle of the chirp series, and then receded in successive chirps. In general, all chirp amplitude-time envelopes have a similar shape of abrupt rise to maximum energy and then gradual

40
energy decrease. To further describe the rise-fall characteristics of the chirps we examined the smoothed Energy Contour for the period of time when the energy of sound was maximal (-3 dB interval from the maximum energy peak), reflecting, along with the ESD, two shapes of amplitude-time envelops: “peaked” (relatively even energy decrease) or “plateau” (persisting energy level, then energy decrease). The tendency of peak energy duration of chirps was to roughly parallel ESD. Two calls (No. 7 and 8) may be characterized as strictly “peaked” while other calls have mixed or relatively “peaked” or “plateau” chirp shapes. This typical temporal structure of the call is exemplified in Figure 1. Chirp number per call averaged 4.75 and the overall average chirp rate was 14.7 chirps/s.
Frequency domain
From inspection of the spectrograms (Figure 2A,C) the sound type may be categorized as “Spectrally structured noise” according to Beeman’s classification (Hopp et al. 1998). The low frequency peak that occurs in many chirps seems not to represent the fundamental frequency of the chirp, according to Cepstrum analysis (Snell 1993).
TABLE 1
Summary of the results (temporal domain)
Call Number Chirp Pause Call Chirpnumber of duration1 duration2 duration3 rate4
chirps (msec) (msec) (msec) (chirps/sec)
Mean SD Mean SD
1 4 66.5 25.8 7.3 3.0 288 15.02 5 79 20.3 4 5.0 411 12.73 4 75.5 13.9 21 21.7 365 13.24 6 69 15.4 26 14.1 544 14.65 5 76 17.6 10.2 17.3 421 13.26 4 62.25 1.0 13.5 7.9 341 16.17 5 58.8 6.6 44.3 2.2 471 17.08 5 46.8 11.1 56 4.7 458 21.4
All callsmean 4.75 70.61 13.96 22.79 18.58 412.36 14.73
1Chirp duration is Estimated Chirp Duration (ESD) as explained elsewhere.2Pause Duration equals Onset-to-Onset Chirp Duration (OOD) minus one ESD.3Call duration estimated as the sum of OODs of all chirps in the call except the last one plus the ESD of the last chirp. 4Chirp rate is actually the ESD chirp rate. It was calculated as the reciprocal of mean chirp duration multiplied by 1000.
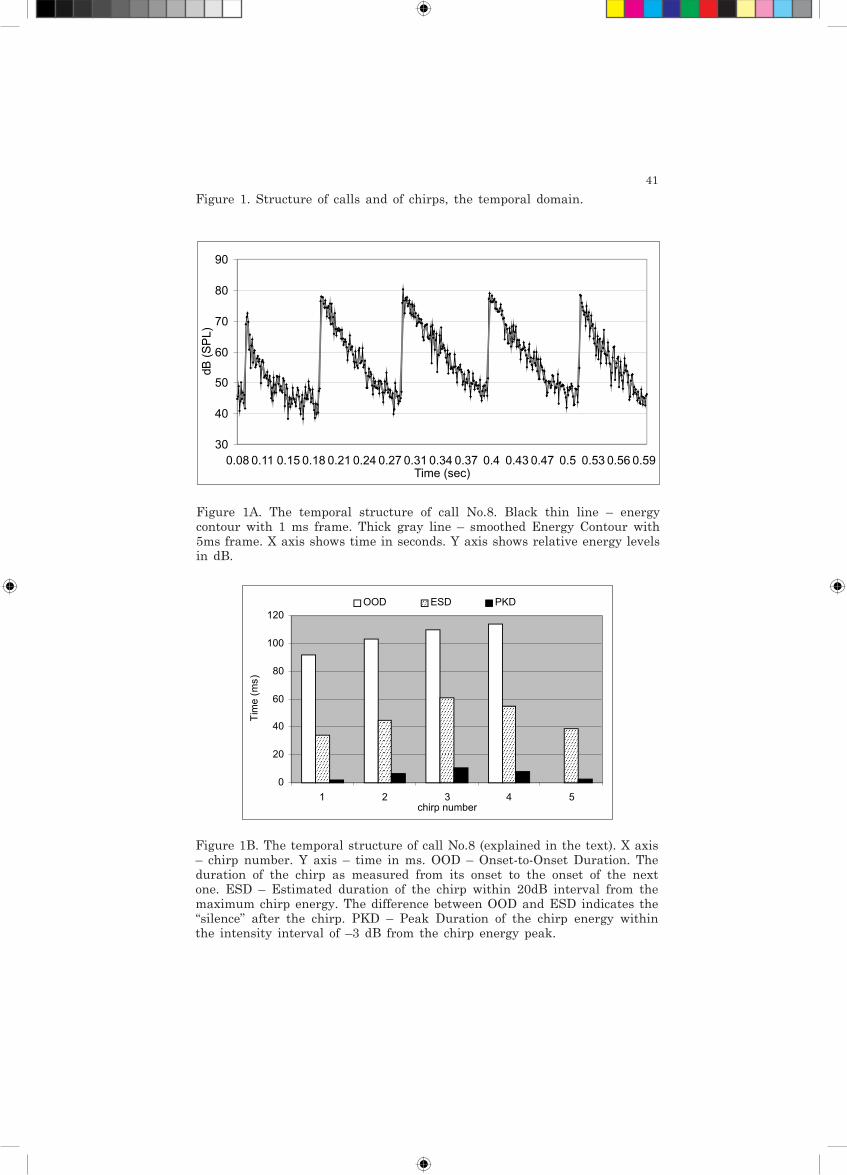
41
Figure 1B. The temporal structure of call No.8 (explained in the text). X axis – chirp number. Y axis – time in ms. OOD – Onset-to-Onset Duration. The duration of the chirp as measured from its onset to the onset of the next one. ESD – Estimated duration of the chirp within 20dB interval from the maximum chirp energy. The difference between OOD and ESD indicates the “silence” after the chirp. PKD – Peak Duration of the chirp energy within the intensity interval of –3 dB from the chirp energy peak.
Figure 1. Structure of calls and of chirps, the temporal domain.
Figure 1A. The temporal structure of call No.8. Black thin line – energy contour with 1 ms frame. Thick gray line – smoothed Energy Contour with 5ms frame. X axis shows time in seconds. Y axis shows relative energy levels in dB.

42
To simplify our primary analysis we decided to work only with the two main frequency peaks in each chirp, i.e. those with maximum energy, as described in Methods (Figure 2B,D). In three calls (of eight) these peaks occurred at uniform frequencies in all chirps of the call (example in Figure 2A,B), in three calls the peak frequencies varied among chirps without clear trends, and in two calls there was a clear trend of the frequencies rising in successive chirps (example in Figure 2 C,D). The overall average frequency of the maximum energy peaks of all calls was 3,418 Hz. The distance of the secondary energy peak from this averaged 1,577 Hz.
Figure 1D. Typical “Plateau” shape of smoothed Energy Contour. Call 6 chirp 1. (Axes as in Fig.1A.)
Figure 1C. Typical “Peaked” shape of smoothed Energy Contour. Call 8 chirp 3. (Axes as in Fig.1A.)

43
Intensity domain
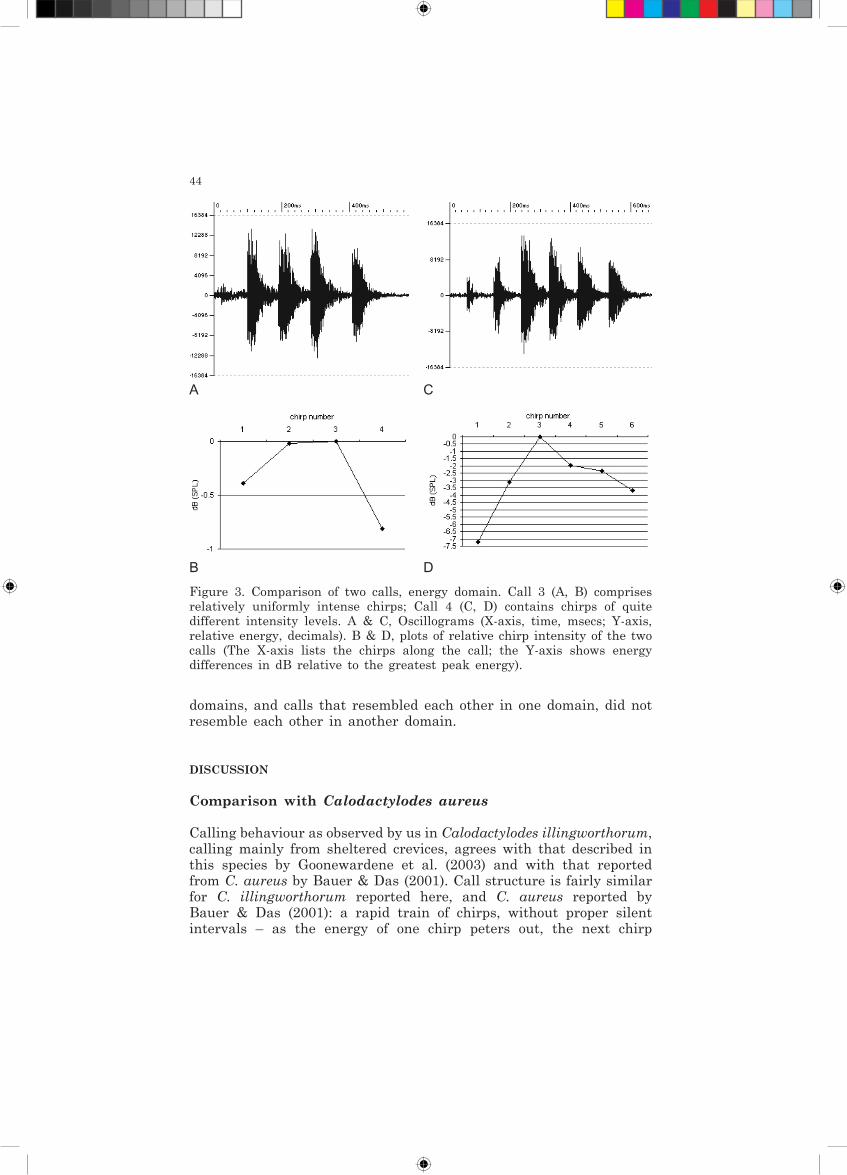
On the oscillograms of complete calls, some calls appear to comprise a chain of equally or similarly intense chirps, and in others a series of equal chirps is preceded by a weak ‘heralding’ chirp. But the analysis revealed a consistent tendency for chirps in the middle of the series to be most intense; the first and last chirps were never the most intense. The difference between the most intense and weakest chirps within a call varied from 1 to 8 dB but commonly was 3-4 dB (Figure 3).
Overall variation
As seen above, in each domain the calls to some extent could be grouped into similar calls. But variation was not correlated among
A C
B D
Figure 2. The structure of two calls, frequency domain. Call 7 (A and B); Call 4 (C and D). A and C, sonograms, the Y-axis comprises 6.5 KHz, B and D show the relative frequencies of the absolute (solid diamonds) and second (open diamonds) energy maxima, as derived from smoothed power spectrum. The X-axis lists the chirps in the call, the Y axis shows the frequency, Hz.
44
domains, and calls that resembled each other in one domain, did not resemble each other in another domain.
DISCUSSION
Comparison with Calodactylodes aureus
Calling behaviour as observed by us in Calodactylodes illingworthorum, calling mainly from sheltered crevices, agrees with that described in this species by Goonewardene et al. (2003) and with that reported from C. aureus by Bauer & Das (2001). Call structure is fairly similar for C. illingworthorum reported here, and C. aureus reported by Bauer & Das (2001): a rapid train of chirps, without proper silent intervals – as the energy of one chirp peters out, the next chirp
Figure 3. Comparison of two calls, energy domain. Call 3 (A, B) comprises relatively uniformly intense chirps; Call 4 (C, D) contains chirps of quite different intensity levels. A & C, Oscillograms (X-axis, time, msecs; Y-axis, relative energy, decimals). B & D, plots of relative chirp intensity of the two calls (The X-axis lists the chirps along the call; the Y-axis shows energy differences in dB relative to the greatest peak energy).
A C
B D

45
already bursts. To the extent that the small samples from both species are representative, the detailed call parameters differ a little. The number of chirps per call is smaller in C. illingworthorum (4-7 chirps, n=9 calls) than in C. aureus (7-11 chirps, n=4); The chirp rate in C. illingworthorum, on the average 14.7 but usually 10-11 chirps/s, may resemble that in C. aureus, 11-12 chirps/s. The resulting call duration is shorter in C. illingworthorum (0.37-0.56 s) than in C. aureus (0.65-0.90 s). The dominant frequency averaged in C. illingworthorum 3,418 Hz and in C. aureus 3,850 Hz but the significance of this apparent difference cannot be evaluated, as in both cases the size of the calling individuals is not known, while in geckos the voice frequency is negatively correlated with body size, both inter- and intraspecifically (Frankenberg 1975). The frequency spectrum of the calls extends to approx. 15 kHz in C. aureus, with a secondary upper energy peak at approximately 11,100 Hz, but we lack comparable information for C. illingworthorum due to instrument limitations.
Comparison with Ptyodactylus
It seems interesting to compare Calodactylodes and Ptyodactylus from the aspects of morphology, biology, and vocal behaviour. The two species of Calodactylodes closely resemble each other. The genus Ptyodactylus comprises several species and subspecies in the northern half of Africa and in south-western Asia (Heimes 1987). In the best studied area, Israel, there occur three species. One of these, P. puiseuxi Boutan, 1893, is diurnal, with relatively smaller eyes and somewhat stocky proportions; but two, P. guttatus Heyden, 1827, and P. hasselquistii Donndorff, 1798, resemble Calodactylodes. With Ptyodactylus represented by these two species, its morphological similarity to Calodactylodes encompasses body size (around 80 mm RA), proportions of limbs (long) and tail (equalling or a little exceeding RA), digits that are distally expanded with a pairs of terminal scansors, with a claw between them (plus a subterminal scansor pair on most Calodactylodes digits), and dorsal scutellation (small scales plus dispersed small or flat tubercles). Its coloration is similarly mottled (not uniform, striped or sharply cross-banded), and presumably cryptic; especially that of P. guttatus, being yellowish, beige or grey with both lighter and darker spots, resembling that of Calodactylodes (Werner & Frankenberg 1982; Werner & Sivan 1993, 1994).
Biologically, Ptyodactylus likewise inhabits rocks in both open and forest situations, although precipitation is much less for it than for Calodactylodes. Both genera are basically nocturnal, mitigated to varying extent by some diurnal activity, including calling (Werner 1965; Werner & Sivan 1994). Most conspicuously, Ptyodactylus also

46
oviposits clutches of two subspherical eggs on sheltered rock faces, in communal rookery sites used by several females year after year. In this case it is also known that oviposition recurs there a number of times in the same year, by the same females (Werner 1986).
Comparative bioacoustics
The calls of Calodactylodes concerned here, a series of similar simple chirps more or less evenly spaced, resemble in this basic structure the multiple-chirp calls of many geckos of diverse affinities and habitats (Frankenberg & Werner 1992; Rickert et al. 2001), including Ptyodactylus guttatus (Frankenberg 1974) and the African P. hasselquistii ragazzi (Werner, Katsir and Rickert, unpublished). The analogous calls of some other geckos have a more complicated structure; these include Gekko gecko (Tang et al. 2001) but also P. h. hasselquistii and P. puiseuxi (Frankenberg 1974; Frankenberg et al. 1978). In some species, even among those with simple multiple-chirp calls, differences have been reported among male-to-male calls, male-to-female calls, and female calls, besides the occurrence of assorted ‘distress’ calls (Frankenberg & Werner 1984, 1992; Rickert et al. 2001). The calls of Calodactylodes illingworthorum cannot be interpreted in these terms. As their three-dimensional variation precluded classification and they were emitted by unidentified individuals in unobserved situations, their variation may be random or individual, and they probably represent the common male-to-male (territorial) call, the male-to-female(courting) call, or possibly both (if we have overlooked the distinction).
Beyond their basic similarity, the call of Calodactylodes illingworthorum greatly differs from that of Ptyodactylus guttatus in chirp rate, which averaged 14.7 chirps/s in the former, compared with means of 0.98 – 1.28 in the latter, depending on social situation and geographical location. The mean number of chirps per call, 4.75 in the former, was below those in the latter, 7-14, depending on social situation and geographical location. As a result, mean call duration, <1s in the former, profoundly differs from the latter, 5.5-12.25s, depending on social situation and geographical location. The (upper) dominant frequency of C. illingworthorum, 3418 Hz is lower than that of P. guttatus, 3000-5600 Hz (Frankenberg 1974). The relative intensity level of chirps within a call has not yet been studied in P. guttatus but overall its call is much louder than that of C. illingworthorum (Werner, from general experience in the field).
Conceivably these differences relate to a possible functional difference, that the calls of P. guttatus serve to communicate between inhabitants of distant rocks while those of C. illingworthorum mainly serve within the colony. The intensity difference obviously accords

47
with this but perhaps also the lower frequency of C. illingworthorum, whose voice, at close range, must “go around corners”. Even the slow rhythm of the P. guttatus might be more efficient in building up attention at a distance.
In the matter of repetition rate, at least, some other geckos are intermediate between Calodactylodes and Ptyodactylus, for example Hemidactylus frenatus (Marcellini, 1974) and H. turcicus (Frankenberg 1982). However, too few geckos have had their vocalizations analyzed, to warrant any generalizations, beyond the negative correlation of dominant frequency with body size (Frankenberg 1975), about ecological or phylogenetic relations of call parameters. The latter is particularly intriguing because species vary in the nature of the difference between vocalizations differing in social role (Frankenberg & Werner 1992). In this matter it is unfortunate that we do not know the social context of the calls reported here.
The fact that Calodactylodes illingworthorum has been defined as threatened or endangered (Wickramasinghe & Somaweera 2002) has the ambiguous potential to both encourage research into its biology and hamper such research. But because the gecko is and has long been popular with local people (De Silva et al. 2004a, 2004c) surely facilitates further research that may help safeguard the species.
ACKNOWLEDGEMENTS
The first author thanks Anslem de Silva for arranging his visit to the habitat and especially Shantha Karunarathne for his efficient guidance in the field. We thank Aaron Bauer, John Rudge and especially Ruchira Somaweera for help with literature and information.
REFERENCES
Bauer, A.M. (2002). Lizards. In Encyclopedia of amphibians and reptiles (Ed. by T. Halliday & K. Adler), pp. 138–175. Abingdon: Andromeda Oxford Ltd.
Bauer, A.M. & Das, I. (2001). Review of the gekkonid genus Calodactylodes (Reptilia: Squamata) from India and Sri Lanka. J.S. Asian Nat. Hist. 5, 25-35.
De Silva, A., Bauer, A.M., Austin, C.C., Perera, B.J.K, Jayaratne, R.L., Goonasekera, M.M., Goonawardene, S., Hawke, Z., Vanneck, V. & Drion, A. (2004a). Some cultural traits of the inhabitants of the Nilgala fire savannah, Sri Lanka, towards animals: with special reference to the herpetofauna. Lyriocephalus, 5,183-191.
De Silva, A., Bauer, A.M., Austin, C.C., Goonawardene, S., Hawke, Z., Vanneck, V., Drion, A., de Silva, P., Perera, B.J.K, Jayaratne, R.L., & Goonasekera, M.M. (2004b). Distribution and natural history of Calodactylodes illingworthorum (Reptilia: Gekkonidae) in Sri Lanka: preliminary findings. Lyriocephalus, 5, 192-198, pl. 5.
De Silva, A., Das, I., Bauer, A.M. & Goonawardene, S. (2004c). Vedda rock art in Sri Lanka depicting reptiles, with special reference to golden gecko Calodactylodes illingworthorum (Reptilia: Gekkonidae). Lyriocephalus, 5, 213-219, pl. 8.

48
Frankenberg, E. (1975). Distress calls of gekkonid lizards from Israel and Sinai. Israel J. Zool., 24, 43-53.
Frankenberg, E. (1982). Vocal behavior of the Mediterranean house gecko, Hemidactylus turcicus. Copeia, 1982, 770-775.
Frankenberg, E. & Werner, Y.L. (1984). The defensive vocal “distress” repertoire of gekkonid lizards: intra- and inter-specific variation. Amphibia-Reptilia, 5, 109-124.
Frankenberg, E. & Werner, Y.L. (1992). Vocal communication in the Reptilia – facts and questions. Proceedings of the First Argentinian and First South American Congress of Herpetology, Tucomán, 1987. Acta Zool. Lilloana, 41, 45-62.
Frankenberg, E., Werner, Y.L. & Adar, O. (1978). Further observations on the distinctive vocal repertoire of Ptyodactylus hasselquistii cf. hasselquisti (Reptilia: Gekkoninae). Israel J. Zool., 27,176-188.
Goonewardene, S., Hawke, Z., Vanneck, V., Drion, A., de Silva, A., Jayaratne, R. & Perera, J. (2004). Report of Project Hoona - 2003. Diversity of Nilgala Fire Savannah, Sri Lanka: with special reference to its herpetofauna. The University of Edinburgh, U.K. 223 pp.
Hopp, S., Owen, M. & Evans, C. (Eds.). (1998). Animal Acoustic Communication. Sound Analysis and Research Methods. Berlin and Heidelberg: Springer-Verlag.
Kiester, A.R. (1977). Communication in amphibians and reptiles. In How animals communicate (Ed. by T.A. Sebeok), pp. 519-544. Bloomington: Indiana University Press.
Marcellini, D.L. (1974). Acoustic behavior of the gekkonid lizard, Hemidactylus frenatus. Herpetologica, 30, 44-52.
Marcellini, D. (1977). Acoustic and visual display behavior of gekkonid lizards. Amer. Zool., 17, 251-260.
Michelsen A. & Larsen, O. (1983). Strategies for acoustic communication in complex environments. In Neuroethology and behavioral physiology (Eds. F. Huber and H. Marx), pp. 321-331. Berlin: Springer.
Morton, E. (1975). Ecological sources of selection on avian sounds. Am. Nat., 109, 17-34.
Snell, R. 1993. CSL Cepstrum Tutorial. Victoria, British Columbia, Canada: STR – Speech Technology Research Ltd.
Tang, Y.Z., Zhuang, L.Z. & Wang, Z.W. (2001). Advertisement calls and their relation to reproductive cycles in Gekko gecko (Reptilia, Lacertiliea). Copeia, 2001, 248-253.
Werner, Y.L. (1965). Über die israelischen Geckos der Gattung Ptyodactylus und ihre Biologie. Salamandra, 1, 15-25 (in German).
Werner, Y.L. (1976). Optimal temperatures for inner-ear performance in gekkonoid lizards. J. Exp. Zool., 195, 319-352.
Werner, Y.L. (1986). Ecology of eggs and laying sites of Ptyodactylus geckos. In Studies in herpetology (Proc. Europ. Herp. Meeting, Prague, 1985), (Ed by Z. Roček), pp. 441-444. Prague: Charles University.
Werner, Y.L. & Frankenberg, E. (1982). Multidimensional investigation of a group of animals, exemplified by Ptyodactylus (Reptilia: Gekkonidae). in Neurobiology and strategies of adaptation (Ed. By F.G. Barth), pp. 60-63. Frankfurt: Goethe Universitaet.
Werner, Y.L. & Sivan, N. (1993). Systematics and zoogeography of Ptyodactylus (Reptilia: Sauria: Gekkonidae) in the Levant: 1, Biometry of three species in Israel. Rev. Española de Herpetol., 7, 47-64.
Werner, Y.L., Rickert, M. & Schröder, E. (2001). Polysyllabic calls of diplodactyline geckos, genera Oedura and Rhacodactylus. Herpetol. Nat. Hist. 8, 37-48.
Wever, E.G. (1978). The reptile ear. Princeton: Princeton University Press.Wickramasinghe, M. & Somaweera, R. (2002). Distribution and current status of the
endemic geckos of Sri Lanka. Gekko, 3 (1), 2-13.

49
Wiley, R.H. & Richards, D.G. (1978). Physical constraints on acoustic communication in the atmosphere: implications for the evolution of animal vocalizations. Behav. Ecol. Sociobiol. 3, 69-94.
Received 11 October 2007, revised 8 November 2007 and accepted 8 November 2007





















