
Tea Chemistry
66
Ii I Critical Reviews ill Plant Sciences, 16(5):41580 (1997) Tea Chemistry Matthew E Harbowy and Douglas A. Balentine Lipton. 800 Sylvan Avenue. Englewood Cliffs. NJ 07660 TABLE OF CONTENTS I. Introduction ....................................................................................................................... 417 II. Chemical Composition, an Overview ...............................................................•....... ; ..... 423 A. Polyphenols .................................................................................................................. 424 B. Caffeine, the Methylxanthines, and Related Compounds ...................................... 424 1. Theobromine .......................................................................................................... 425 2. Other Derivatives of Nucleic Acids ..................................................................... 425 C. Proteins and Amino Acids ......................................................................................... 426 D. Carbohydrates, Pectins, and Fiber ........................................................................... 426 E. Organic Acids and Vitamin C ................................................................................... 426 F. Lipids, Chlorophylls, Carotenoids, and Related Compounds ............................... 427 G. Vitamins and Minerals ............................................................................................... 428 H. Aroma ........................................................................................................................... 428 III. Polyphenols in Tea ........................................................................................................... 429 A. Chemical Classification .............................................................................................. 430 B. Green Tea Polyphenols ................................................................................•.............• 430 1. Catechins and Gallocatechins ............................................................................... 430 2. Flavonols ................................................................................................................. 432 3. Simple Polyphenols ... ............................................................................................. 432 4. Other Polyphenols ................................................................................................. 432 5. Tannins ................................................................................................................... 433 C. Black Tea Polyphenols ................................................................................................ 434 1. Residual Green Tea Polyphenols ......................................................................... 435 a. Catechins .......................................................................................................... 435 b. Flavonols .......................................................................................................... 436 2. Theaflavins and Related Products ............. .......................................................... 437 a. Theaflavins ............................................................................. .......................... 437 b. Theaflavic Acids .............................................................................................. 438 0735·2689/97/$.50 © 1997 by CRC Press LLC 415
-
Upload
bogoragriculturaluniversity -
Category
Documents
-
view
2 -
download
0
Transcript of Tea Chemistry

Ii I
Critical Reviews ill Plant Sciences, 16(5):415-480 (1997)
Tea Chemistry Matthew E. Harbowy and Douglas A. Balentine Lipton. 800 Sylvan Avenue. Englewood Cliffs. NJ 07660
TABLE OF CONTENTS
I. Introduction ....................................................................................................................... 417 II. Chemical Composition, an Overview ...............................................................•....... ; ..... 423
A. Polyphenols .................................................................................................................. 424 B. Caffeine, the Methylxanthines, and Related Compounds ...................................... 424
1. Theobromine .......................................................................................................... 425 2. Other Derivatives of Nucleic Acids ..................................................................... 425
C. Proteins and Amino Acids ......................................................................................... 426 D. Carbohydrates, Pectins, and Fiber ........................................................................... 426 E. Organic Acids and Vitamin C ................................................................................... 426 F. Lipids, Chlorophylls, Carotenoids, and Related Compounds ............................... 427 G. Vitamins and Minerals ............................................................................................... 428 H. Aroma ........................................................................................................................... 428
III. Polyphenols in Tea ........................................................................................................... 429 A. Chemical Classification .............................................................................................. 430 B. Green Tea Polyphenols ................................................................................•.............• 430
1. Catechins and Gallocatechins ............................................................................... 430 2. Flavonols ................................................................................................................. 432 3. Simple Polyphenols ................................................................................................ 432 4. Other Polyphenols ...................................... ........................................................... 432 5. Tannins ........................................................ ........................................................... 433
C. Black Tea Polyphenols ..................................... ........................................................... 434 1. Residual Green Tea Polyphenols ......................................................................... 435
a. Catechins .............................................. ............................................................ 435 b. Flavonols .......................................................................................................... 436
2. Theaflavins and Related Products ....................................................................... 437 a. Theaflavins ............................ ................................................. .......................... 437 b. Theaflavic Acids .................................................. ............................................ 438
0735·2689/97/$.50 © 1997 by CRC Press LLC
415

c. Other Related Structures ............................................. ... ... ............................ 438 3. Further Oxidized Products. The "Thearubigens" ............................................. 439
a. Theafulvins and Theacitrins .......................................................................... 442 b. Gallic Acid Production ................................................................................... 444 c. Bisflavanols and Proanthocyanidins ............................................................ . 444 d. Mixed Oxidation Products of Polyphenols
and Other Compounds ................................................................................... 446 e. Aroma Formation from Polyphenol Oxidation ........................................... 447
4. Oolong Tca Polyphcnols .......... .................................................... ......................... 447 a. Oolongtheanins and Theasinensins ..... ............... .................. ......................... 447
IV. Biochemistry of Tea ...................... ............................................. ........................... ........... 448 A. Caffeine Formation ..................................................................................................... .450 B. Theanine Formation ................................................................................................... 451 C. Biochemistry of Flavonoid Compound Formation ................................................. 452
1. Phenylalanine and the Shikimate Pathway ........................................................ 452 2. Chain Extension, Hydroxylation .......................................................................... 453 3. ChalconelFlavone Tautomerization ..................................................................... 454 4. FlavanolslFlavonols ............................................................................................... 454 5. Esterase ................................................................................................................... 455
D. Latent Enzyme Activity and the Formation of Black Tea ..................................... 456 1. Polyphenol Oxidase ............................................................................................... 456 2. Peroxidase ............................................................................................................... 458
E. Extracellular Enzymatic Activity .............................................................................. 458 V. Chemical Properties of Tea Compounds ....................................................................... 459
A. Formation of Cream and Haze in Black Tea ..... ........................... .......................... 459 B. Complex Formation ........................................................................................ ............ 460 C. Polyphenols as Antioxidants ......................... ......... .................................................... 463
1. Chemical Antioxidant Modcls .............................................................................. 463 2. Biological Antioxidant Models ............................................................................. 465
VI. Trends in Tea Research ................................................................................................... 466 A. Analysis by Chemical Constitution and Technical Innovation ............................. 466 B. Bulk Properties and Correlations to Tea Taster's Profiles .................. ................. 467 C. Tea as an Antioxidant and as a Healthy Beverage ................................................. 467
1. Lipid Oxidation and Cardiovascular Disease .................................................... 468 2. Studies of Tobacco Nitrosamines .......................................... .... ........................... 469 3. Tea and Cancer ...................................................................................................... 470
VII. Conclusion ......................................................................................................................... 470
416

Referee: Dr. Alan P. Davies and Dr. Va Cai, Unilever Research Colworth Lab., Colworth House,
Sharnbrook, Bedford, U.K.
ABSTRACT: The chemistry of tea as a beverage is reviewed in depth, covering both historical and current chemical perspectives. Special attention is given to the polyphenols in tea, although the general composition and properties are also treated. Current trends in tea science. particularly
in the area of polyphenol complexation and antioxidant properties, are also covered. The need for a chemically based understanding, rather than one hypothesized from generalized and indirect observation. is stressed.
KEY WORDS: tea chemistry, polyphenol complexation, antioxidant properties.
I. INTRODUCTION
The real voyage of discovery consists not in seeking flew landscapes but in having new eyes.
·Proust
The universe is 1I0t only queerer than
we suppose, but queerer thall we can
suppose.
-J. B. S. Haldane
It is a picture-perfect image of serenity: relaxing with a good book, sipping a cup of hot tea. For thousands of years, the harvesting, processing, and packaging of the leaf of Camellia sinensis, known worldwide as tea, has developed as an integral part of society and culture. With the advent of the postcolonial scientific era, the perspective of scientific investigation was added to the tradition and mythology of tea cultivation. Today, we can look at a cup of tea and admire the complexities of this beverage and the plant that makes it possible. Tea chemistry has led both consumers and researchers to debate numerous issues and to probe for a deeper understanding of the nature of this beverage. With the growing popularity of tea and increased awareness of the potential health benefits associated with tea consumption, tea chemistry promises to endure as a growing and vibrant field.
After water, tea is the most widely consumed beverage in the world today. Currently in the U.S., per capita consumption of tea is approximately 340 g, which produces
approximately 35 to 40 I of beverage. India has the largest total consumption of tea (540,000 metric tons, 620 g per capita) and Ireland has the largest per capita consumption, at 3220 g (Anon., 1 994). Camellia sinensis is a very i mportant agricultural and commercial product with a unique horticulture and manufacturing process.
Chinese mythology teaches that in the year 2737 B.C. Emperor Shen Nung discovered tea, according to the Chinese medical book, the Pen T'sao, written during the Han Dynasty, circa 25 to 22 1 A.D. The first mention of tea is believed to occur i n the Erh Ya, a Chinese dictionary circa 400 B.C., but the modern character ch'a, signifying tea, was not popularized until the writing of The Classic of Tea, Ch'a Ching, by Lu Yu in 780 A.D. The history of tea is extremely relevant because before the Tang dynasty (61 8 to 906 A.D.) tea was probably only considered as a medicinal, but then became popular as a beverage. As the method of brewing and consuming tea varied as tea moved from culture to culture, the chemical aspects driving acceptability to the consumer varied.
It is likely that tea was consumed as a vegetable in a soup through the Tang dynasty, often mixed with onions, salt, orange peel, and/or ginger. Brick tea, still popular among the modern Mongolians and mountainous people of the Himalayas, was prepared by steaming and compressing the leaf into bricks. During the Song Dynasty (960 to 1279 A.D.), however, this practice fel l from favor and was replaced by a powdered form of the tea, which was whipped into a
417

froth. A bright green color and low astringency (derived from careful shading of the plant) and a delicate aroma accompany powdered green tea. This preparation method of tea is a custom that survives today in Japan as Mattcha. The modem custom of brewed tea leaves arose during the Ming dynasty ( 1 368 to 1 644), coinciding with the West's arrival in China. Although there are many variations on brewing technique, which can impact flavor and chemistry of the brew significantly, it is the basic custom of brewing the dried tea leaves in hot water that has been popularized and spread throughout most of the English-speaking world.
The modern tea industry has its origins in the spread of tea cultivation into India between 1 8 1 8 and 1 834, derived either through import of the tea plant from China or through discovery of native Indian tea var. Assamica, a varietal better adapted to tropical production and having a larger leaf style. Development of tea plantations, and migration of the technology of plantation operation from India to tropical areas in Africa, South America, and Russia (Georgia), has established a variety of localized practices and tea products (Eden, 1 976). Excellent reviews on the culture and production of tea are available (Wilson and Clifford, 1 992). Figure I illustrates the major historical periods in the development of tea cultivation and processing worldwide.
Through cultivation, tea has become an important agricultural product throughout the world, particularly in regions lying close to the equator. Geographical areas that receive annual rainfall of at least 50 inches per year and have a mean average temperature of 30°C are the most favorable for growth and agriculture of tea (Eden, 1 976).
Traditionally, C. sinensis has been propagated, hybridized, and bred through seeds. To maintain clonal purity and to accelerate establishment of new productive stands of tea, vegetative propagation is now a com-
418
mon practice. This involves planting leaf cuttings in nurseries where they develop into seedings within 6 months. The seedings are then transplanted to the fields. The tea plant, once established, will be economically viable for decades barring disease, infestation, or other destructi ve forces.
The tea tree is maintained as a shrub during the growing season through frequent manual harvesting, about every 8 to 12 d during prime growing season. Tea is an evergreen tree, but it is largely dormant through the winter season. Mechanical harvesting methods have been developed, but these are only popular where labor is expensive and where tea is not grown on steep mountain slopes. Plantations also maintain unharvested tea trees for seed production.
Immediately after harvest, the tea leaves (usually the flush, or first two leaves and the bud of the growing tea shoot) are brought to factories si tuated close to the tea gardens for manufacturing. It is the manufacturing process that determines the type of tea produced. There are three general types of manufactured tea: Green (unfermented), Oolong (partially fermented), and Black (fully fermented). The manufacturing processes used to produce each type of tea differ in the degree of enzymatic oxidation or "fermentation". Fermentation refers not to an exogenous, microbial process, as with beer or wine, but the natural browning reaction catalyzed by enzymes endogenous to the plant.
The green tea manufacturing process involves the rapid steaming or pan firing of the freshly harvested leaves to inactivate enzymes, preventing fermentation, producing a dry, stable product. Green teas are typically produced in two categories: "White tea" and "Yellow tea", the latter is withered (wilted), resulting in a small degree of fermentation (Bokuchava and Skobeleva, 1980). There is some variety of terminology between Chinese and Japanese green tea manufacture, and with the increase in popularity

... � '"
\(:!-7.>.m.,,�-> �Wr;'
India 1818-1834 AD
FIGURE 1_ The cultivation of tea worldwide .
A �J:'i)it spreads to Japan 600-800 AD?
Tea'tullivation spreads pacific islands 9Qg"AD
. -.t�';tfu.�t\.
'{) \" ('if _f�J:

of green tea worldwide, pilot production of green teas in other regions, such as Darjeeling in India, from traditionally black tea varieties have led to a wide variety of green tea products on the market.
When oolong and black teas are to be produced, the fresh leaves are allowed to wither until the moisture content of the leaves is reduced to 55 to 72% of the leaf weight. This causes a concentration of polyphenols in the leaves and deterioration of leaf structural integrity. Withering is important for aroma development. The withered leaves are rolled and crushed, initiating fermentation of the tea polyphenols. The fermenting mass formed from rolling and used in black tea manufacture is referred to as dhool. The process used to macerate the leaf plays an important role in the final grade of tea. Two common methods are Orthodox and CTC. Orthodox rolling of the leaf is performed by mechanically applying weight or compression to the leaves. Orthodox processing is typically used for production of large-leaf finished tea products. CTC (crush, tear, curl) processing i s a significant modern improvement of this procedure that minces the leaf in a continuous, high-yielding process and produces smaller-leaf teas.
Oolong teas are prepared by firing the leaves shortly after rolling to terminate the oxidation process and dry the leaves. The rolling process for oolong teas is only designed to slightly damage the leaf and impart 'twist' to the finished product. Black teas are prepared through a separate fermentation process in which cooled air is circulated through the rolled and crushed leaves to moderate the reaction, as the onset of fermentation is accompanied by a rise in temperature from the exothermic fermentation process. This fermentation process results in the oxidation of simple polyphenols to more complex condensed polyphenols that give oolong and black teas their bright red colors and brisk astringent flavor. The degree of
420
fermentation of the dhool largely determines the flavor charateristics of the finished product. Fermented tea leaves are then fired to inactivate the enzymes and dry the leaves. This procedure is accompanied by the final chemical transformations to the product resulting from the high temperatures involved in firing. Figure 2 highlights the differences in manufacture of the most common tea products.
The manufactured teas are then sized, graded, and evaluated for flavor and infusion color by professional tea tasters. The teas are packaged into sacks or wooden chests and are sold at the world tea auctions. Total world production of tea in 1 993 was 2.58 million metric tons, of which 0.59 million metric tons was green tea and 1 .89 million metric tons was black tea. India and China are the major tea-producing countries, manufacturing 53% of tea produced (Anon., 1 994). Figure 3 illustrates available consumption data of tea worldwide.
There are numerous polyphenolic compounds produced by the growing tea plant, and the pathways for in vivo biosynthesis of these phytochemicals have been elucidated generally. The chemistry of green and black tea, therefore, typically centers on the polyphenolic composition of these teas, with polyphenols being the major proportion of extracted solids in black teas. Although this is a Western bias, both i n terms of chemistry of production and beverage as well as the recent interest in the health-promoting aspects of tea, it is important that the overall composition be given careful consideration. The organization of this review attempts to reemphasize these areas, in contrast to previous reviewers.
One of the most comprehensive reviews of tea chemistry available is that of Sanderson (Sanderson, 1972). Complementary to Sanderson's review is that by the Russian tea chemists Bokuchava and Skobeleva (Bokuchava and Skobeleva, 1 980), which

T he Tea Plant
/PIUCking '-I -
T-he-
�-e
-a
-F
-IU-
S-h�1
Withering � degree of 'fermentation'
Withered
Flush
Rolling
'Fermentation'
Fermented Dhool
Firing
Firing
Firing
-------..
"White" tea (green tea)
"Yellow" tea
(green tea)
"Red" tea
(aalang tea)
Firing Black Tea
FIGURE 2. The tea manufacturing process.
421

India 560
China 408
Japan 130
Iran 85 Pakistan 117
USA 91
CIS 132
UK 151
Other 221
Morocco 30 Syr ia 22
Sri lanka 22 Poland 31
Egypt 64
Turkey 124
FIGURE 3. Consumption of tea, 1993 (10' metric tons).
provides a variant perspective and insight into Russian-language literature sources, which are typically not cited in English-language publications. HPLC techniques, now available for rapidly measuring a number of important tea polyphenols, including the catechins, f1avonols, flavonol glycosides, and the theaflavins, have dramatically changed the way tea chemistry is studied, and these methods have been reviewed in detail (Finger et aI., 1 992).
There are a variety of other reviews on the subject of tea, including Roberts' (Rob-
422
erts, 1 942; Roberts, 1 962) and Graham's (Graham, 1983; Graham, 1 984 ) reviews of tea chemistry, a review by Stahl (Stahl, 1962), Wickremasinghe (Wickremasinghe, 1 978), Balentine (Balentine, 1 992), and a review by Robertson (Robertson, 1 992). An excellent comprehensi ve review of the work on tea aroma to the early 1 980s is also available (Bokuchava and Skobeleva, 1 986), as well as a more recent review (Robinson and Owuor, 1 992).
Reviews published subsequent to that of Sanderson appear to attempt an update of the

subject, but none have critically reassessed the field. It is hoped that this review can take some of the historical material, combined with a comprehensive and critical account of recent research, and synthesize a review of tea chemistry as it is currently viewed and applied in our laboratory. An overemphasis on polyphenols, compounds that are significant to color and astringency but less relevant to teas such as shaded green tea production, dominates the l i terature. Low polyphenol content and high chlorophyll content are important for producing an emerald-green and smooth finished green tea product that lacks astringency, and the brothy character of the amino acids, and flowery character of the aroma constituents, play a much more significant role. In addition to the nonpolyphenolic consti tuents, the polyphenols of black tea are a poorly understood but seemingly well-defined group of compounds. The use of "thearubigen" in the literature signifies a wide group of compounds whose chemical identity has not been traced to any identifiable chemical group. Its
use was most significant as a colorimetric indicator for plantation production, but as a chemical identifier its use has seemed to hamper true identification of the mass balance of black tea polyphenols. Work at identifying these compounds in a systematic fasion has begun, and it is hoped that future research will deemphasize the term ' thearubigen' in favor of more chemically accurate descriptors.
II. CHEMICAL COMPOSITION, AN OVERVIEW
In order to divide the subject of tea chemistry into well-ordered issues, it is most convenient to do so on the basis of chemical composition. Frequently, researchers' interest in specific compounds or a class of compounds induces a focusing of attention on a small fraction of the total tea mass. In addi-
tion, there are differences between the composition of the tea leaf and other phytological components, as well as differences between the leaf and the "brew". It is therefore difficult to synthesize an overall picture of the chemistry of tea without resorting to a subdivision of the problem.
With this in mind, we have attempted to focus on the "extract solids", the component of tea leaves that is extracted by boiling water. Table I gives an approximate composition of black and green tea beverage solids by chemical class. Although the length of time for steeping and the amount of water in which the leaves are steeped can vary widely, these factors generally control the amount of solids extracted, and to a lesser extent influence the composition. A typical brew of one tea bag in one cup of water produces a solution of 0.35% wtlwt solids, and from this value the dose expected from consumption of one cup of tea can be calculated. This is typically how tea phytochemicals are consumed. Notes on the composition of the 'flush' or the fresh tea leaves, or other parts
of the tea plant, are added for completeness
TABLE 1 The Composition of a Typical Tea Beverage, %wt/wt Solids
Green tea Black tea
Catechins 30% 9% Theaflavins 4% Simple polyphenols 2% 3% Flavonols 2% 1% Other polyphenols 6% 23% Theanine 3% 3% Aminoacids 3% 3% Peptides/Protein 6% 6% Organic acids 2% 2% Sugars 7% 7% Other carbohydrates 4% 4% lipids 3% 3% Caffeine 3% 3% Other methylxanthines <1% <1% Potassium 5% 5% Other minerals/ash 5% 5% Aroma Trace Trace
423

within the review. We have also subdivided the issue of tea compounds by chemical class. This is done to focus on specific aspects of tea chemistry, such as aroma or polyphenols, and because the methods used for determination of the various chemical classes are quite different.
The strict division of tea chemistry by functional chemical identity has its uses, but begins to lose value i n discussing subjects that cross the boundaries of chemical class or those that have no distinctive chemical identity, such as discussions concerning the 'thearubigens'. Modern tea research has reached the point where simple chemical subdivision has been refined with a fair degree of accuracy, and determination of the synergies and 'boundary violations' present in the chemical composition of tea beverages becomes a necessity.
A. Polyphenols
In terms of human consumption, tea represents a major source of dietary polyphenols. The polyphenolic fraction of tea represents 30 to 40% wt/wt of extract solids and provides astringency, the 'drying' sensation experienced in the mouth after consumption of the tea beverage. A tea drinker typically consumes 1 80 to 240 mg of polyphenols from a strong cup of tea. Recent interest in the health aspects associated with consumption of tea beverages has grown within the scientific community and has generated much excitement about tea polyphenols.
The tea plant produces a diverse number of polyphenolic constituents, presumably as a means of chemical defense against insects, birds, and animals, which would consume the plant as food (Beart et aI., 1 985). The evolution of salivary proline-rich proteins, which bind polyphenols effectively, has ameliorated this defense mechanism, converting it to 'astringency' (Luck et aI., 1 994).
424
B. Caffeine, Methylxanthines, and Related Compounds
Tea has been valued historically for its caffeine content. Caffeine [ 1 ] i s viewed as an important constituent of tea, bestowing mood and cognitive-enhancing properties (Bokuchava and Skobeleva, 1 980). Figure 4 illustrates the methylxanthines of tea. Tea leaves contain between 2 and 5% wt/wt caffeine depending on the variety.
[1]: Caffeine
[2]: Theobromine
NH2 �N�COOH
o
[3]: Theanine
FIGURE 4. Nitrogenous tea phytochemicals.

C. irrawadensis, a member of the Camellia family, lacks caffeine (Roberts et ai., 1 958) but is not processed commercially because it produces a poor finished tea product. The quantity of caffeine that infuses into a tea brew i s determined by infusion time and by leaf style. Longer infusion times lead to greater quantities of caffeine in a tea beverage. Smaller sized tea leaves give a more rapid and stronger infusion, whereas larger leaves and uncut leaves lead to weaker infusions. This results in more or less caffeine extraction, respectively. The caffeiflt, <.:ontent of a typical tea beverage will range from 20 to 70 mg per 1 70 ml of infusion, with a typical infusion being prepared from about 2 to 2.5 g of tea leaves. Coffee brews typically contain from 40 to 1 55 mg caffeine per 170 ml beverage.
There has been little research done on the pharmacology of tea-beverage caffeine. One study suggests a dose of caffeine from tea has a different physiological effect than a pure dose of caffeine (Das et ai., 1 965). This has been attributed to the amino acid theanine, which is unique to tea. However, there are no well-designed clinical studies to support this position. The consensus among scientists today is that caffeine from all beverage sources has a similar physiological effect. The actual content of caffeine depends on many factors, particularly the method of brewing. A brew prepared by the Chinese "gong-fu" style is likely to have a different caffeine impact compared with the Western style of loose tea or to that from a tea bag (Hicks et ai., 1 996). Some reports have suggested that green tea contains significantly less caffeine than black tea. This may be influenced by the clone of leaf used to produce the tea or by the impact of different brewing techniques. No significant differences have been found when brewing green and black teas under similar conditions (Hicks et ai., 1 996), discrediting the theory that withering and fermentation have
a significant impact on caffeine content (Sanderson, 1 972).
Caffeine is one of the most comprehensively studied ingredients in the food supply. Extensive research does not link moderate caffeine intake to any health risks. Studies are needed to better understand the physiological role of tea caffeine and its association with the popularity of tea beverages. Those individuals who are especially sensitive to caffeine can find decaffeinated teas readily available.
1. Theobromine
Theobromine [2] is present in tea in much lower quantities than caffeine. Theobromine is formed as a consequence of the biosynthesis of caffeine (Negishi et ai., 1985a) and is produced in abundance if the methylation path to caffeine i s absent, such as in C. irrawadensis (Roberts et ai., 1 958). Theophylline, a similar di-methylxanthine, has been reported in trace quantities in tea leaves (Michl and Haberler, 1 954; Sanderson, 1972). Recent reports contradict as to the the existence of this compound in tea, some failing to detect these compounds (Hicks et ai., 1 996) and others (Meyer et ai., 1996) reporting small quantities. The xanthine content of teas is clearly an area that requires further, more careful research.
2. Other Derivatives of Nucleic Acids
The RNA and DNA in tea leaves are metabolized naturally as well as digested under the conditions of withering and fermentation by tea nucleases, nucleosidases (Imagawa et ai ., 1 982), and a specific adenine nucleosidase (Imagawa et ai ., 1 979). These catabolic reactions produce purines, which have been detected in very small quan-
425

tities in tea (Michl and Haberler, 1 954; Sanderson, 1 972; Hicks et aI., 1 996).
C. Proteins and Amino Acids
While caffeine is the most well-known ni trogenous component of tea, tea proteins/ pep tides and amino acids contribute significantly to the composition of both the leaf and the tea extract. Figure 4 i l lustrates some of the other important miscellaneous nitrogenous and non nitrogenous components of tea. Recent measurement of the amino acids in two green teas (Liang et aI., 1 990) confirms the presence of 1 8 amino acids, a result that is mirrored for black tea. Amino acids contribute about 6% wt/wt of the extract solids. Tea also contains a significant amount of peptidic material (protein), approximately 6% wt/wt of extract solids. Nitrogenous materials therefore comprise about 15% wt/wt of extract solids. The free amino acid content of tea seems to increase during withering of the fresh tea leaves but decreases duri ng fermentation to black tea (Roberts and Sanderson, 1 966), as it i s likely consumed during aroma biogenesis and through other routes. These pathways have a strong impact on the aroma of the finished product and require more detailed investigation.
In addition to the common amino acids, there is a unique amino acid known only to be present in tea. This amino acid, theanine (y-N-ethyl glutamine, [3]), is believed to be the major amino acid present in tea, comprising about 3% wt/wt of extract solids. Theanine is a significant component of both green tea (Sakato, 1 950) and black tea (Feldheim et aI., 1 986). Theanine has been associated with improved flavor and a modulation of the stimulative effects of caffeine (Kimura and Murata, 1 97 1 ). Recent studies on the antihypertensive effect of theanine found that a large quantity of this compound was required to exhibit an effect in rats (Yokogoshi et aI., 1 995).
426
Optimum production of theanine i n cell culture has been investigated (Furuya et aI., 1 990; Matsuura et ai . , 1 992). Theanine can also be made synthetically on a commercial scale in good yield (Kawagishi and Sugiyama, 1 992).
Tea leaves subjected to anaerobic conditions are found to produce excess amounts of GABA, ory-amino butyric acid (Tsushida, 1 987), derived from glutamic acid due to the action of an endogenous glutamate decarboxylase in tea (Tsushida and Murai, 1 987).
D. Carbohydrates, Pectins, and Fiber
Tea leaves have been shown to contain free sugar residues in addition to pectic subtances, polysaccharides, and fiber (Mizu no et a i . , 1 964; Sanderson and Perera, 1 965). Carbohydrates contribute approximately I 1 % wt/wt of extract solids (Sanderson et aI., 1 976; Graham, 1 984). High molecular weight pectins (polygalacturonic acid) and other polysaccharides have been analyzed on Sephadex G- 100 (Millin et aI., 1 969). As the tea plant matures, increases in the content of lignin and cellulose have been observed (Selvendran et aI., 1 972), which is consistent with their role in providing structural integrity to the growing plant.
A significant portion of the carbohydrate fraction in tea extract has been found to comprise the disaccharide 2-0-(�-L-Arabinopyranosy l)-myo-inositol [4] (Sakata et ai., 1 987), as shown in Figure 5. It was detected by NMR techniques in an aqueous fraction subjected to consecutive extraction with ethyl acetate and butanol (Sakata et aI., 1 989).
E. Organic Acids and Vitamin C
Tea is a significant source of oxalic acid (Sanderson and Selvendran, 1 965) and malic acid (Jayman and Sivasubramanian, 1 975), along with citric, isocitric, and succinic acids (Sanderson and Selvendran, 1 965). Tea

�O�H
OH HO 0
H�O 00H
OH [4]: 2-0-(p-L-Arabinopyranosyl)-myo-inositol
HO OH
HO [5]: Spinasterol
FIGURE 5. Miscellaneous tea phytochemicals.
also contains shikimic and qUIntC acids, which are important to the biosynthesis of the polyphenols (Zaprometov, 1961) . Vitamin C (ascorbic acid) has also been detected in green tea (Liang et aI., 1 990) and black tea (Sanderson, 1 972).
6. Lipids, Chlorophylls, Carotenoids, and Related Compounds
The main pigments in the fresh tea leaf are chlorophylls and carotenoids. Chlorophylls (e.g., [39]) are oxidized during the course of black tea manufacture to the pheophytins and pheophorbides (e.g., [40]), which give the fermented leaf its characteri stic brown-black color. Some of the pheophytins and pheophorbides are extracted into the black tea beverage (Sanderson, 1 972). An efficient HPLC method has been developed for analysis of these pigments
(Taylor and McDowell, 199 1 ). The composition of these components in the green tea leaf has been demonstrated to have a strong impact on the quality of the beverage as perceived by tea tasters (Taylor et aI., 1 992). Tea grown in the shade has been found to have a lower quantity of catechins (resulting in a less astringent beverage) and increased levels of carotenoids and chlorophyll (which may assist in aroma production). This chemical balance is thought to contribute favorably to taste (Mahanta and Baruah, 1 992). The carotenoids play a significant role in the formation of aroma characteristic to black tea (Bokuchava and Skobeleva, 1 986). High chlorophyll levels and low astringency are important to some green tea manufacture, particularly Mattcha, for which a brothy, emerald-colored brew is very important.
Lipids, terpenoids, and saponins make up a large portion of the fresh tea leaf, yet because of their low water solubility, are generally thought to be a minor portion of
427

the water extract solids. However, plant steroids such as spinasterol [5) or lipids such as the plant cuticle wax triacontanol [6) have been shown to comprise an important fraction of tea cream, the precipitate that forms after cooling of concentrated tea extracts (Seshadri and Dhanaraj, 1 988). It is hypothesized that the hydrophobic environment presented by the tea polyphenols and caffeine complex i n tea provide for extra solubility of the lipid components. Lipids are approximately 3 to 4% wtlwt of leaf and has been analyzed in detail (Anan, 1 983; Bhuyan and Mahanta, 1 984). The role of hydrophobic plant materials in the appearance and organoleptic properties of the brew has been typically disregarded in favor of the polyphenols, and this is an important area for which future research is necessary.
G. Viti mans and Minerals
The tea plant has been shown to be rich in potassium (Sanderson ct aI., 1 976) and contains significant quantities of calcium and magnesium, as well as small amounts of manganese, iron, and phosphorus (Kalita and Mahanta, 1 993), copper and nickel (Burke and Albright, 1 970), and sodium, boron, and molybdenum (Hasselo, 1 965). Zinc (Tolhurst, 1 962) and sulfur (Pethiyagoda and Krishnapillai, 1 970) are essential elements for healthy maturation of the tea plant as wel l . Cobalt, lead, and cadmium have been detected in the plant, and concentrations depend principally on soil concentrations of these minerals (Ramakrishna et aI., 1 987).
The tea plant is known to accumulate aluminum (Chenery, 1 955). Aluminum levels can be traced by NMR techniques (Nagata et aI., 1 9 9 1 ) and have been found in complexes with fluoride and catechins in the tea plant (Nagata et aI., 1 993). Tea polyphenols are commonly thought to complex with min-
428
erals, and may be excellent chelating agents. The large number of phenolic hydroxyl groups provides a great number of potential active complexation sites.
Tea beverages are also a significant source of fluoride (Elivin-Lewis et aI., 1980). This is due, in part, to the uptake of aluminum fluoride (Yamada and Hattori, 1977; Yamada and Hattori, 1 980).
H. Aroma
The essential oil or aroma of tea provides much of the pleasing flavor as well as scent of green and black tea beverages, yet comprises only a minor fraction of the total mass of the tea plant or the extracts. Tea aroma contains hundreds of compounds in trace quantities, the composition and mechanism of production of which has been reviewed (Choudhury, 1982; Bokuchava and Skobeleva, 1 986; Robinson and Owuor, 1992).
Withcring has been determined to play an important role in aroma development, as in oolong teas (Takeo, 1 984; Kharebava, 1 986) and black teas (Owuor et aI., 1 987). Many of the aroma components of tea can be found as glycoside derivatives, which are freed during the fermentation process due to the action of glycosidases. Fresh tea enzymes permit the release of additional aroma constituents in the leaf and extract, restoring fresh aroma from stale tea (Guo et aI., 1992). Geranyl, linolyl, terpinyl, and neryl glycosides can be found in fresh tea extracts (Guo et aI., 1 993). �-Glucosidase is the enzyme most likely to be responsible for formation of tea aroma from these glycosides (Morita et aI . , 1994). The exploitation of bound glycosides of aroma components by glycosidase treatement offers the possibility of future improvements i n tea quality.
Part of the aldehyde fraction may be generated from a unique tea leaf amine oxi-

dase (Tsushida and Takeo, 1985). Tea also contains a fatty acid hydroperoxide lyase, which forms volatile aldehydes from the lipid constituents of the tea leaf (Matsui et a!., 199 1 ) . There are also many products that are derived from the oxidizing conditions present during tea fermentation (B okuchava and Skobeleva, 1 986). Figure 6 illustrates one such unique aroma constituent, theaspirone [9], which is produced from oxidation of p-carotene [7].
III. POL YPHENOLS IN TEA
The term 'polyphenoJ' is an inclusive descriptor referring to the millions of natural and synthetic aromatic molecules that are substituted with multiple hydroxyl groups. The polyphenols comprise one of the most distingishing characteristics of the tea plant and have been more thoroughly investigated than any other class of compounds in tea. For this reason, the polyphenols in tea are
[7]: p-Carotene
/ �o [8]: p-Ionone
many other aroma constituents
[9]: Theaspirone
FIGURE 6. Unique tea aroma constituents.
429

treated in greater depth in this section and separate from the overall review of the chemical constituents of tea.
Because of the abundance of polyphenols present in tea leaves and in tea beverages. i t is natural that tea chemistry is often considered to be synonymous with tea polyphenol chemistry. The polyphenols are principally responsible for the color and astringency and partially responsible for the flavor of the tea beverage. The compounds are known antioxidants and are being studied as agents that might reduce risk factors associated with cancer and heart disease. While careful attention needs to be paid to the overall chemistry of the tea plant and beverage. it is no surprise that this review and much of the tea li terature is dominated by the discussion of the tea polyphenols.
A. Chemical Classification
The polyphenols in tea may be subdivided by several chemical backbone structures. Simple tea polyphenols are those thar are synthesized during the early stages of polyphenol biosynthesis. whereas the degree of complexity of the polyphenols increases as one progresses down the biosynthetic pathway. The flavonoids. a subgrouping of polyphenols and the dominant class of green tea polyphenols. are synthesized in part from the simple polyphenols and represent compounds with 1 5 or greater carbon atoms [C" stage] in the basic framework. The polyphenols of black tea represent further chemical transformations of the green tea polyphenols and therefore comprise a third level of complexity. Unique black tea polyphenols are commonly thought to be polymers of the green tea polyphenols and therefore are thought to be comprised of molecules of approximately 30 carbon atoms [C30 stage] or greater. as the simplest polymer would be a dimer such as procyanidin.
430
All too frequently. popular writing on tea compounds confuse the nearly homonymic flavonoid classifications. Flavonoids are the widest subgrouping. i ncluding flavanols. which have a saturated central (C) ring and include the catechins. the major green tea polyphenols. and flavonols. which have an unsaturated central (C) ring and a ketone group. Careful attention must be paid when reading any of these three classifications. These classifications are all inclusive under the broader term polyphenol. which refers to any compound that contains aromatic rings with multiple pendant phenolic OH groups or derivatives thereof.
B. Green Tea Polyphenols
Green tea polyphenols consist of both simple and complex polyphenols. The large majority of polyphenols in green tea are flavonoid monomers called catechins and flavonols.
1. Catechins and Gal/ocatechins
The catechins [ 1 0- 1 3] represent the major polyphenolic constituent of green tea and are i l lustrated in Figure 7. Catechins are members of a more general class of flavonoid. the flavan-3-0Is (also referred to as flavanols). Three subgroupings of the flavanols [afzelechin [ 1 4]. catechin [ 15 ] . gallocatechin [ 1 6]. shown in Figure 8]. representing varying degrees ofB-ring hydroxylation. are the dominant forms. of which the epi-isomers of the catechins and gallocatechins are the principal components found in tea. The tea catechins. a term commonly used to refer to both catechins and gallocatechins. make up as much as 30% wtiwt of dissolved solids.
A large percentage of the catechins present in tea exist as gallic acid esters. While

R1 R2 [10]: Epicatechin EC H H [11]: Epicatechin Gal late ECG Gal late H [12]: Epigailocatechin EGC H OH [13]: Epigailocatechin Gal late EGCG Gal late OH
FIGURE 7. The principal tea catechins.
&OH
HO¢ '� I," � OH
OH
R1 R2 [14] : Afzelechin H H [15]: Catechin C OH H [16] : Gal locatechin GC OH OH
FIGURE 8. The flavan·3·ols.
gallation is found to occur principally at the 3-position, various other gallated species have been isolated, including the epigallocatechin digallates (Nonaka et aI., 1983), and epicatechin digallate (Coxon et aI., 1 972; Hashimoto et aI., 1 987). 3-Methyl gallates of EC and EGC have also been reported (Saijo, 1 982). EGCG was once thought to be unique to the tea plant (Graham, 1983) but
now has been isolated from other sources (Danne et aI., 1 994).
The four most common catechins are epigallocatechin gallate (EGCG, [ 1 3 ] ) , epigallocatechin (EGC, [ 1 2]), epicatechin gallate (ECG, [ 1 1 D, and epicatechin (EC, [ 1 0]). Catechin (C, [ 15]) and gallocatechin (GC, [ 1 6]) are also present i n smaller quantities. While gallocatechin gallate (GCG) and
431

catechin gallate (CG) have also been observed, i t is likely that these are products of racemization and not 'native' to the tea plant (Roberts, 1 962; Robertson, 1 992).
2. Flavonols
The flavonols [kaempferol, quercitin, and myricitin] and their glycosides [ 1 7- 1 9] have only been recognized recently as significant components in tea, although their presence as trace constituents has always been acknowledged. The flavonols are illustrated in Figure 9 Analyses of the flavonol glycosides in general (McDowell et aI., 1 990) and of the flavonol diglycosides (Finger et aI., 1 99 1 a) and triglycosides (Finger et aI., 1 99 1 b) in tea leaf and of flavonol glycosides in tea seed (Sekine et aI., 1 99 1 ; Sekine et al 1 993) have been performed.
Use of hydrolysis to determine flavonol and flavonol glycoside content as their aglycones (Hertog et aI., 1 993) has proven to be useful in determining overall flavonol content (about 0.5 to 2.5% wtiwt extract, as aglycone) of tea infusions.
HO
OH 0
[1 7]: Kaempferol Glycoside [18]: Quercitin Glycoside [19]: Myricitin Glycoside
3. Simple Polyphenols
Gallic acid [20] and its quinic acid ester (or depside, as quinic acid esters are commonly referred), theogallin [2 1 ], have been identified in tea (Cartwright and Roberts, 1 954; Cartwright and Roberts, 1 955) and have been detected by HPLC (Bailey et aI., 1 990; Hashimoto et aI., 1 992). The simple polyphenols and their depsides are shown in Figure 10.
Cinnamic acid derivatives of quinic acid, the coumaryl and carrt:oyl- quinic acids (including chlorogenic acid or 5-caffeoylquinic acid [23]) have also been identified in tea (Cartwright et aI., 1 955). Chi orogenic acid and 4-coumarylquinic acid [22] have been detected by HPLC (Bailey et aI., 1 990).
4. Other Polyphenols
Flavones and their glycosides (Engelhardt et aI., 1 993), such as apigenin [24], have been detected in tea but represent a very small fraction of the polyphenols present. Flavone glycosides can potentially
R, OH
OGlycoside
KaG QuG MyG
H OH OH
H H OH
FIGURE 9. The lIavonol glycosides.
432

OH
Hooc-Q-OH
OH
[20]: Gallic Acid
OH
:�OH
HOOCn OH
OH�OH [21 ] : Theogal l in OH
R
ovd-OH
o HOOCn
OH�OH OH
[22] : Coumarylquinic acid , R=H [23]: Chlorogenic acid , R=OH
FIGURE 10. Gallic acid and the depsides.
be measured quantitatively as aglycones (Hertog et ai., 1 993). Careful resolution of individual flavone glycosides (Engelhardt
et ai., 1 993) has confirmed their presence in tea. Apigenin is illustrated in Figure I I .
Flavan-3,4-diols such as leucocyanidin [27] have also been reported (Roberts et ai., 1 956). These are illustrated in Figure 1 2.
A number of proanthocyanidin species, such as prodelphinidin B 2 [25] gallates, have been isolated and are present in green tea (Non aka et ai., 1 983; Nonaka et ai . , 1 984). The assamicains, such as assamicain A [26] isolated from C. sinensis var. assamica, appear to be ring-opened products of the proanthocyanidins (Hashimoto et ai., 1 989a).
5. Tannins
Although it is commonly stated that there are no tannins (meaning hydrolyzable tannins such as pentagalloylglucose [28]) in tea, this statement is not strictly true. In addition to the gallic acid esters of the catechins and their oxidation products (which can be hydrolyzed to produce gallic acid readily and
433

OH
HO
OH 0
[24] : Flavone (Apigenin)
OH
HO
�OH
" " �OH OH
OH HO
" 'OR1 .J-yOH
o " " lJlOH
" 'OR1
[25] : Proanthocyanidin
(Epigal locatechin-4-a-Epigal locatechin , or Prodelphinidin B2)
FIGURE 1 1 . Miscellaneous polyphenols,
precipitate proteins), there is also a small quantity of hydrolyzable tannin (Nonaka et aI., 1 984; Yoshida et aI., 1 990; Hatano et aI., 1 99 1 ; Han et aI., 1 994). The unique hydrolyzable tannins in tea are typically "hybrid" tannins such as camelliatannin A [29], which is a galloylglucose derivative with pendant catechins, The tannic acid derivatives common to gall-nuts and tree bark are not present in significant quantities in tea infusions. The tea tannins are illustrated in Figure 1 3 ,
434
C. Black Tea Polyphenols
B lack tea polyphenols are produced from the controlled enzymatic reactions involved in the fermentation of green leaf during commercial and model black tea production, The extent and conditions under which fermentation occurs determines the degree to which the polyphenols of green tea are transformed to those unique to black tea. It is reasonable to expect that black tea should contain an amount of polyphenols

OH HO OH
OH
o . . ��6c:: " OGa
OH Ga=Galiate
[26]: Assamicain A
OH
OH
HO
OH OH OH
[27] : Flavan-3,4-diol
(Leucocyanidin)
FIGURE 12. Miscellaneous polyphenols.
similar to green tea. However, the complex nature of these polyphenols, some of which are "polymeric" in nature, has largely resisted chemical identification. The unidentified polyphenolic constituents are often referred to as thearubigens. Despite their complexity, some of the unique black tea polyphenols have been identified and characterized.
1. Residual Green Tea Polyphenols
During the course of fermentation, the polyphenols of green tea are rapidly converted to the polyphenols of black tea. De-
pending on the degree of fermentation, however, some green tea polyphenols remain unconverted. This is particularly true in the case of oolong teas and some Darjeeling teas, which have been known tv resemble green tea both in chemical constitution (Ding et a!., 1 992a) and astringency.
a. Catechins
The catechins represent the major portion of green tea polyphenols and consequently are thought to be the building blocks of black tea polyphenols. Some of the green tea catechins survive the fermentation pro-
435

HO�OH
X HO�O
�O O OH I 0 0 0
HO :::'" 0 0 0 )��-OH OH q �[\4
"'" 0 � OH OH 1 .& � /, HO OH OH OH HO
[28] : Hydrolyzable Tannin (Pentagalloylglucose)
HO
HO HO
HO
HO OH
HO-i >
HO
OH HO
[29]: Cameli iatannin A
FIGURE 1 3. Tannins.
OH
cess and are detected in black tea (Bailey et a!., 1 990). Due to the oxidation reactions and thermal conditions experienced by the tea leaf during black tea production, it i s hypothesized some of the catechins are also epimerized and/or degallated, which explains the appearance of free gallic acid as well as
increased levels of non-epi isomers of the catechins (Coggon et a!., 1 973).
b. Flavonols
It is likely that the majority of flavonols (free as well as glycosides) present in the
436

initial fresh green leaf remain unoxidized and are likewise present in black tea in similar quantities. There is some evidence that some of the flavonols are oxidized during fermentation. It has been suggested that myricitin and myricitin glycosides are the most likely oxidized of the three flavonols (kaempferol, quercitin, myricitin) (McDowell et aI., 1 990).
Despite this, in a recent analysis of flavonols and their glycosides (Hertog et aI., 1 993), no significant difference between the green teas and the black teas was found in terms of total flavonol as aglycone, except for myricitin, which was found to be slightly reduced in the black tea samples compared with green teas. Differences between these samples might have also been derived from different origins for the two analyzed materials, however, owing to the variations in flavonol content of teas grown from different clones and from different regions of the world.
2. Theaflavins and Related Products
One of the key distinctions of black teas compared with green tea is the production of a new type of polyphenol, the theaflavins. The fermentation of green tea leaf also results in the development of characteristic aroma components, a darkening of color of the leaf and extracts, and a decreasing astringency with increased fermentation time.
a. Theaflavins
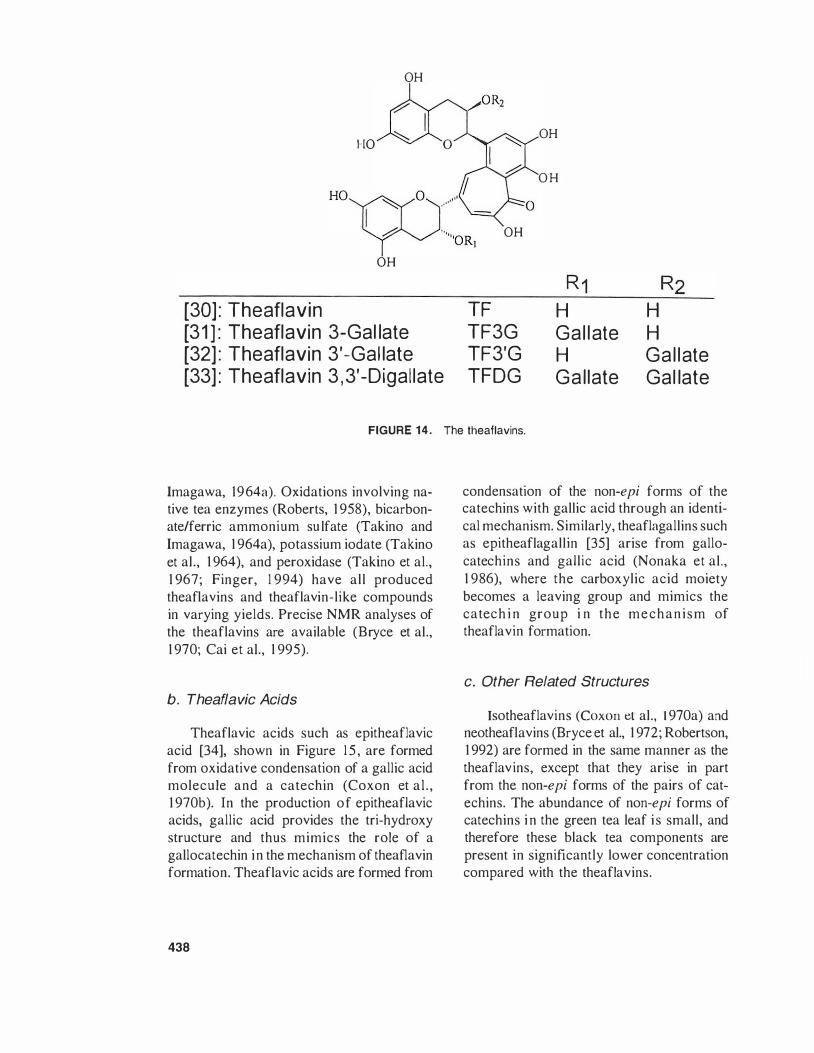
Best known of the fermentation products is the class of compounds known as the theaflavins [30-33), comprising about 3 to 5% wtlwt of the extract solids. They are i l lustrated in Figure 14. Theaflavin provides a bright, red-orange appearance to the tea beverage and has long been positively correlated with market value of tea (Roberts,
1 958). Market value for tea is also influenced by secondary factors such as aroma, as observed with Kenyan teas that normally contain higher theaflavin contents (Owuor et aI., 1 986) and therefore not a distinctive characteristic.
While characterized by a unique benzotropolone ring structure resulting from the dimerization of a catechin and a gallocatechin, there are a series of related compounds, inc luding the i soth eaflav ins , neotheaflavins, and theaflavic acids, which also possess a similar benzotropolone unit. The benzotropolone ring provides the red color and makes the theaflavins easily distinguishable from other components.
Analysis of theaflavins began with extraction of water extracts (Roberts, 1 958; Spiro et aI., 1 987; Robertson and Hall, 1 989) into isobutyl methyl ketone (Roberts and Smith, 1963) or ethyl acetate (Ullah, 1 972), followed by spectrophotometric measurements. Later spectrophotometric methods improved on this technique (Xiao and Li, 1 992). The Flavognost method (Hilton, 1 973) uses diphenyl boric acid ethanolamine to induce a spectrophotometric shift in the benzotropolone ring for better accuracy. Recent improvements in analytical technology include the use of GC (Collier and Mallows, 1 97 1 ), and HPLC (Robertson and Bendall, 1 983; Steinhaus and Engelhardt, 1 989), as well as NIR measurements on the leaf (Hall et aI., 1 988) and absorption onto cartridge columns (Whitehead and Temple, 1 992).
The mechanism of theaflavin formation was fairly well defined in early papers on purpurogallin formation and related experiments (Horner et aI., 1 96 1 ; Critchlow et aI., 1 967; Takino and Imagawa. 1 964b). Papers investigating the fermentation reaction leading to the characteristic benzotropolone ring reported the production of a series of theaflavin-like compounds, most notably erycitrin (Takino and Imagawa. 1 963), categal lin. and pyrogallin (Takino and
437
OH
OH HO
7 Hoyyoi," " _
�""ORI OH
OH
OH °
[30] : Theaflavin [31 ] : Theaflavin 3-Gal late [32] : Theaflavin 3'-Gal late [33] : Theaflavin 3 ,3'-Digal late
TF TF3G TF3'G TFDG
H Gal late H Gallate
H H Gal late Gal late
FIGURE 14. The theallavins,
lmagawa, 1964a), Oxidations involving native tea enzymes (Roberts, 1 958), bicarbonate/ferric ammonium sulfate (Takino and Imagawa, 1 964a), potassium iodate (Takino et aI., 1 964), and peroxidase (Takino et aI., 1 967; Finger, 1 994) have all produced theaflavins and theaflavin-like compounds in varying yields, Precise NMR analyses of the theaflavins are available (Bryce et aI., 1 970; Cai et aI., 1 995),
b. Theaflavic Acids
Theaflavic acids such as epitheaflavic acid [34], shown in Figure 15 , are formed from oxidative condensation of a gallic acid molecule and a catechin (Coxon et aI . , 1 970b). In the production of epitheaflavic acids, gallic acid provides the tri-hydroxy structure and thus mimics the role of a gallocatechin in the mechanism of theaflavin formation. Theaflavic acids are formed from
438
condensation of the non-epi forms of the catechins with gallic acid through an identical mechanism. Similarly, theaflagallins such as epitheaflagallin [35] arise from gallocatechins and gallic acid (Nonaka et aI., 1 986), where the carboxylic acid moiety becomes a leaving group and mimics the catech i n group i n the mechanism of theaflavin formation.
c. Other Related Structures
Isotheaflavins (Coxon et aI., I 970a) and neotheaflavins (Bryce et aI., 1 972; Robertson, 1 992) are formed in the same manner as the theaflavins, except that they arise in part from the non-epi forms of the pairs of catechins. The abundance of non-epi forms of catechins i n the green tea leaf is small, and therefore these black tea components are present in significantly lower concentration compared with the theaflavins.

OH OH
OH
OH Hooe
OH
[34]: Epitheaflavic Acid
H0'Y00 " ' / I -
:::,.. " 'OH OH OH
OH
[35]: Epitheaflagall in
FIGURE 1 5 . The epitheallavic acids and related compounds
3. Further Oxidized Products: The "Thearubigens"
From the early stages of development, simple solvent extractions attempted tu quantify the amount of colored tea compounds present i n the black tea brew (Roberts et aI., 1 957). The theaflavins, a group of bright orange-red compounds, were quickly separated from the remainder. Paper chromatography experiments confirmed that subsequent to solvent extraction, a series of brown-red compounds remained in the aqueous phase, which were not well resolved by 2-D paper
chromatography, but were roughly quantifiable by simple spectrophotometric techniques. These compounds were given the label of thearubigens. However, subsequent research began to compromise the understanding of the thearubigens as a well-defined group of compounds.
Refinements on the procedure for quantifying thearubigens were made beyond the early paper chromatography techniques. Solvent extraction methods, employing ethyl acetate and butanol, or methyl iso-butyl ketone and butanol, used colorimetry differences at -450 nm (for theaflavins) and -350 nm (for thearubigens), with each successive method building on the early assumptions of Roberts. This method was later converted to use C-1 8 cartridge columns (Whitehead and Temple, 1 992) and a series of solvents to elute the appropriate fractions and measure thearubigens and theaflavins by colorimetry.
The significant limitation of use of this method is that there is no clear evidence, other than a correlation, that these measured"thearubigens" and the "thearubigens" identified by Roberts are one and the same. This becomes more evident in later methods. The thearubigens were divided into three subclasses on the basis of paper chromatography: SI, SIIa, and SIIb (Roberts et aI., 1 957). Later HPLC techniques reinterpreted the issue with division of the thearubigens into groups I, n, and III (Bailey et aI., 1 99 1 ; Bailey et aI., 1 994a). In both papers, the presence of an unresolved mass (as illustrated in Figure 16) is taken to represent the same compounds, the "unresolved thearubigens". This is based on loose similarities of HPLC with paper chromatography as well as more recent work with cartridge column fractions (Wellum and Kirby, 1 9 8 1 ).
The arrival of HPLC chromatography led to the belief that for the first time individual thearubigens would be separated and isolated (Hoefler and Coggon, 1 976; Robertson and Bendall, 1 983). However, it
439

�.� .. ' = Area of well resolved spols � = Diffuse, unresolved spots Cinnamic Acids
� ....•.... . -.-... . ..... ... _."::'.:.:::.:.:. r.:.-::::-....... .
Flavonol Glycosides
Catechins
TheafJavins (.
Butanol- etic Acid-Water
225
200
1 75
1 50
. .... 'Thearubigens'
Diffuse spot on bo h paper chromatography a d on H PLC chromatogr m taken to be synonymous
Both identified as 1 26 , 'unresolved thearu igens'
1 00 atechins
75 " 50
25
1 0 20 40
FIGURE 1 6. Paper chromatography vs. HPLC.
became clear early on that chromatography using reversed-phase materials was not an ideal solution, due in part to the observation that some tea components were not eluted from cartridge columns.
Strategies for separation of the thearubigens on normal-phase chromatography
440
(Wedzicha and Donovan, 1 989) and using the technique of counter-current chromatography (Okuda et aI ., 1 988; Wedzicha et aI., 1 990) have shown some promise in separation and identification. The absence of recent reports suggests that use of such "alternative" technologies has fallen into disfavor,

due in part to the extreme simplicity of reversed-phase techniques. Use of reverse phase colums with a step-gradient (Putman and Butler, 1 989) might present a potential useful technique for these compounds as well.
Use of reverse-phase technology has been expanded systematically by a series of recent papers (Opie et aI., 1990; Bai ley et aI., 1990; Bailey et aI., 1 99 1 ; Bailey et aI., 1 994b) and from thesis work (Opie, 1 992). The reverse-phase technique seemed to point to the thearubigens in one of two classes: a series of red (-450 nm) compounds eluting discretely and a diffuse peak appearing as a 'rising baseline' across the same chromatogram.
Investigations into the thearubigen fractions identified by early techniques revealed that part of the thearubigens was the flavonol glycosides (McDowell et aI., 1 990). Being present in green tea, these compounds are clearly not thearubigens. They contribute significantly to the absorbance around -350 nm in both the solvent partitioning and cartridge column techniques and must be excluded from these measurements if the term thearubigen is to retain its original meaning, that is, as a product of oxidation of green tea polyphenols.
Early model fermentation systems successfu l ly identified the paired role of gallocatechins and catechins in theaflavin formation (Sanderson et aI., 1 972). The model fermentation approach was expanded to investigate the role of purified tea PPO (Coggon et aI., 1 973) and peroxidase (Dix et aI., 1 9 8 1 ) on formation of the theaflavins and thearubigens. After the arrival of HPLC, this technique was used in the attempt to justify the individual thearubigin peaks' origin (Robertson and Bendall, 1 983; Robertson, 1 983). Model fermentation of individual catechins was performed (Opie et aI., 1 990), and peaks were identified that are potential thearubigens, but attention was drawn away from individual analysis and focused on a
diffuse rising baseline. A study of a model fermentation of a mixed system oftheaflavins and epicatechin in the presence of polyphenol oxidase showed that epicatechin and the theaflavins, but not the theaflavins alone, resulted in the degradation of the theaflavins (Opie et aI., 1 993). In another study, the presence of a diffuse peak on reverse-phase chromatography (having become a de-facto qualifier for thearubigens) was most significantly formed during model oxidation of epicatechin alone, and such a fermentation brew has been suggested as a strategy for thearubigen isolation (Opie et aI., 1 995). This approach, however, seems limited, as epicatechin is a minor constituent i n tea relative to other catechins, and to suggest that studies of its fermentation products will lead to identification of thearubigens can only result in a very minor portion of the said thearubigens being identified.
Model fermentation systems seem to be the most reasonable approach for determining the origin of individual thearubigens as discrete chemical identities. The use of model fermentation systems eliminates some of the confusion concerning the origin of thearubigen-like compounds that may have been "left over" from the green tea polyp henols. However, until better techniques for analysis are discovered, little information can be gleaned from model fermentation studies. In addition, model fermentation systems will always suffer the question of whether the model is accurately representing thearubigen production.
Use of ultrafi ltration to separate a highmolecular-weight thearubigen fraction confirmed the presence of a high-molecularweight "polymer", but these high-molecularweight polymers represent at most only 2% of the total brew solids (Kuhr et a!., 1 994).
The crude h i storical defi n it ion of thearubigens on the basis of poorly resolved paper chromatograms would seem to represent a stumbling block for systematic
441

chemical identification of tea compounds, because no one unique chemical structure seems to be indicative of a thearubigen to date. It i s therefore the preference of the authors to avoid the use of the broad term "thearubigens" except as a historical artifact and as a semiquantitative analytical number used by tea tasters. One reason is that if the solvent extraction/colorimetry technique (Roberts et ai., 1 957) i s used successfully to measure thearubigens, the thearubigens measured are, in part, composed of flavonol glycosides (McDowell et ai., 1 990). Thus, flavonol glycosides are i n some sense thearubigens. This places the definition of thearubigens as oxidation products of catechins in an awkward position.
Some of the color attributed to the thearubigens may in fact not be flavonoid in nature at ali. Figure 1 7 illustrates some possible alternative explanations for the brown coloration and "acidity" as described in the historical reports. They may be highly rearranged compounds such as catechinic acid [36] (Sears et ai., 1 974). Another explanation is that thearubigens are overoxidation products of theaflavins or direct products of peroxidase, a theory that is well supported by model studies of PPO and peroxidase. In this respect, they may be ring-opened products s i m i lar to m uconic acid [37,38] (Hayaishi and Hashimoto, 1 950; Speier et ai., 1 993) or derivatives thereof (Critchlow et ai., 1 967). The brown color attributed to the thearubigens may also be attributed in part to pheophorb i de (e .g . , [40]) and the pheophytins (Sanderson, 1972), or polysaccharides such as [4] and polymers thereof (Millin et ai. , 1 969). None of these conjectures as shown in Figure 1 7 have been investigated. However, the presence of browncolored products that partition into all of the phases of solvent extraction, including the aqueous phase, butanol phase, and ethyl acetate phase, seems to require that there be a number of possible chemical moieties in-
442
volved and not a simple, single central structure as exhibited by the theaflavins.
a. Theafulvins and Theacitrins
Early reports suggested that the thearubigens, after aci d hydrol ysis, produced anthocyanidins (Brown et ai., 1 969). This would lead one to believe that at least some of the thearubigens are a class of condensed tannin, possessing linkages at the 4-position, which may have benzotropolone units or other chromophores to gi ve the characteristic dark brown appearance of the thearubigens.
Recent work on this approach has isolated a fraction believed to be part of the thearubigens, termed theafulvin (Bailey et ai., 1 992). The buff-tan appearance resembled that of the condensed tannins, and the materials gave simi lar behavior on C- 1 8 chromatography when compared with cider and wine proanthocyanidins. However, acid hydrolysis of the two materials gave widely differing results. Condensed tannins gave reasonable yields of anthocyanidins derived from the hydrolysis products, a behavior that is typical of polymeric flavan-3-0Is. The anthocyanidins are readily identified by their bright color and characteristic absorption spectrum and behavior on PRP-phase chromatography. The products of hydrolysis of the theafulvin fraction, on the other hand, yielded similar colored materials, but which were unretained by PRP chromatography. This led the authors to believe that only the end groups of the polymer were converted to anthocyanidin and that linkage occurred through an alternate location, such as the 3'position (Bailey et ai., I 994a).
In another report, Porter's reagents were used to achieve hydrolysis of the proanthocyanidin fraction, as well as analyze gallate and flavonol content by HPLC (Powell et ai., 1 995). Both the theafulvin fraction, as well as the caffeine-precipitable

X
HOyyO,(X �
" 'OH OH
[1 0] : Epicatechin (colorless)
COOH COOH
c
X OH OH
x
HO [36] : Catechin ic Acid
(brown)
OH I
C H800H - � OH OH
X 0
OH
[37] [30] : Theaflavin [38]
(brown, acidic)
[39]: Chlorophyll B
CHO
(red)
""CH2 (brown, hydrophobic)
H3C 7" r;
HO
o
(Brown, acidic)
CHO
CH3
(green)
CH3 [40]: Pheophorbide
FIGURE 1 7 . Alternative brown pigments.
fraction, were analyzed by this method. In both cases, the proanthocyanidin content of these fractions seemed to be explainable on
the basis of proanthocyanidins observed in green tea polyphenols (Hashimoto et aI., 1 987), and the gallate content of these
443

proanthocyanidins would explain most of the hydrolyzed gallate in the fractions. This result weighs heavily against the possibility that these fractions are thearubigens derived solely from catechin or theaflavin precursors, unless the chemical characteristics of these "thearubigens" had been transformed drastically in chemical nature.
It should be noted that the theafulvin fraction, a buff-tan solid, is isolated in approximately 3% yield from tea extract solids, of which 1 0% is proanthocyanidin in nature and 3% is derived from gallate esters. The low color i mpact this fraction is likely to have on the overall beverage should be considered as small evidence for support of this fraction as a thearubigen. The thearubigens have been suggested to be dark brown and strongly influencing the color of the beverage, i n addition to being -20% of the total extractable solids. Also, gallate esters are the predominant form of catechins in green tea, comprising - 1 5 % wtlwt of extract, of which -35% of the mass of these is gallic acid. Therefore, it is reasonable, to expect -5% of the mass of black tea extract to be gallate, as confirmed by tannase hydrolysis (unpublished results). Therefore, one would expect to find much greater than -0.3% of the extract from which a thearubigen fraction was derived to be present as thearubigen gallate esters. Given this criticism, the theafulvin fraction should not be identified conclusively as a major thearubigen until stronger, direct evidence exists.
Attempts at isolation of the thearubigens ha ve generated at least two other new fractions: the theacitrins (Powell et aI., 1 994) and a caffeine precipitable thearubigen fraction (Powell et aI., 1 993).
b. Gallic Acid Production
One of the products of the fermentation process is the appearance of gallic acid [ 1 2], constituting approxi mately 1 % wtlwt of the
444
extract solids (Graham, 1 984). This simple polyphenol is thought to be a product of degallation of the 3-galloyl substituted catechins and gallocatechins that are abundant in the natural beverage. Although the production of gallic acid is not well understood, either native esterase (tannase) activity or oxidative degallation during the fermentation is a likely pathway to its formation.
A possible method of investigation of thearubigens is to use the presence of gallated polyphenols to establish mass balance of the polyphenolic fraction. Tannase treatment uf both green and black tea produced from the same clone along with measurement of gallic acid content should establish the amount of gallic acid oxidized into polymers. Subtracting the free gallic acid (released in the original black tea extract) and the consumed gallic acid (computed from the gallic acid released by tannase treatment) from the gallic acid released from tannase treatment, one can establish the molar concentration of catechins involved in black tea polyphenols. By then subtracting the molar quantity of known gallated pol yphenols such as catechins and theaflavins, one can establish a molecular basis for the thearubigens and better hypothesize on the true chemical constitution of the thearubigens. Further, the difference in mass balance between recovered gallic acid and original gal lated species in the green leaf would establish the amount of gallic acid polymerized into the "thearubigen mass". Good establishment of cycles of mass balance such as this are notoriously absent from much of the tea chemistry li terature, and this fai lure might explain why true identification of the thearubigens has eluded scientific inquiry for so long.
c. Bisflavanols and Proanthocyanidins
The bisflavanols, such as bisflavanol A [41 l , arise from paired condensation of two gallocatechins (Vuataz and Brandenberger,

196 1). It was initially expected that these would be intermediates in the formation of theaflavins. The extra OH substitution from the use of two gallocatechins, as opposed to a catechin/gallocatechin pair, provides a mechanistic barrier by replacing a hydrogen atom, which is lost easily in tautomerization, with an OH group (see Figure 2 1 ). This does not rule out the possibility of further rearrangement or condensation, possibly to thearubigins.
theasinensins. They were found to be present i n green tea leaf (Nonaka et aI., 1 983) and oolong tea (Hashimoto et aI., 1 988). As well as the gallocatechin-gallocatechin dimer products, the theasinensins include catechingall ocatechin dimer products such as theasinensin F [42]. The theasinensin family is depicted in Figure 1 8.
The bisflavanols were later rediscovered and reclassified under the wider name of
Basic work on the separation, characterization, and chemical identification of black tea constituents has been well advanced in recent l iterature. In a massive compendium of work on tannins and related compounds,
td°H OGallaleOH
9' I I '"
HO �
° .-?
H°'(lr°i., "" · : I
�OGallale OH OH
OH
OH OH
OH
[41] : B i sflavanol A, or Theasinensin A
OH OH
HO OH HOyql ° . . . . . . . 9' � . � I
OGaliale OH OH OH [42] : Theasinensin F
FIGURE 18. The theasinensisns.
445

best summarized in brief by one member of this series (Hashimoto et aI., 1 992), a great number of oxidation products from both oolong and black tea sources have been characterized, as well as novel polyphenols from green tea. This approach is immensely useful, but it is a tedious and expensive route, and i t is anticipated that a good HPLC technique that quantitatively establishes a good mass balance of the unknown tea polyp henols will require many more years of dedicated work unless a working hypothesis for their formation can be established.
The presence of proanthocyanidins and theasinensins in green tea as well as black tea indicates that great care must be taken to exclude the nascent green polyphenols in the quantitation of thearubigens. Furthermore, these compounds do not possess significant absorption in the visible region of the spectrum, which rules them out as contributors to mass of the brown thearubigens.
d. Mixed Oxidation Products of Po/ypheno/s and Other Compounds
While the most widely known coupled oxidation products of green tea fermentation are the theaflavins, resulting exclusively from the reaction of catechins and gallocatechins, there are other possible oxidation products in the fermentation system not arising solely from these materials.
The a-quinone group, once formed, is a highly reactive species and therefore has little selectivity for condensation. It is likely therefore that other nucleophilic species, including active thiols and amine groups, as well as other unoxidized polyphenolic species, can condense with the a-quinone (Van Sumere et aI., 1 975).
Theogallinin [43] and the theaflavonins such as [44] (Hashimoto et aI., 1 992) represent two recently identified species that contribute to the balance of polyphenolic mate-
446
rial present i n black tea. They are the product of the condensation of catechins (which are the l i kely oxidized species) w i th theogallin and myricitrin, respectively. Erycitrin [45] (Takino and Imagawa, 1 963) is a theaflavonin, the discovery of which predates that of the theaflavonins, lending futher confusion to the terminology. These oxidation products are shown in Figure 1 9 .
OH
H
o HOOen
OH�OH OH
OH
OH
OH
OH
[43] : Theogal l in in
OH
OH
HO
OH 0
[44] : Theaflavonin, R=Galloyl [45] : Erycitrin , R=H
FIGURE 19. Mixed oxidation products.

e. Aroma Formation from Po/ypheno/ Oxidation
The aldehyde aroma components arising in black tea aroma have been attributed to condensation of corresponding amino acids with an a-quinone to form an amine-substituted derivative (Bokuchava and Popov, 1 954) during polyphenol oxidation. The citation of this mechanism is interesting as some products (aldehydes, carbon dioxide, and ammonia) have been observed (Popov, 1 956; Skobeleva and Popov, 1 962). Tea leaf amine oxidase produces aldehydes and ammonia as byproducts (Tsushida and Takeo, 1 985). Detection of these byproducts in fermentation studies does not constitute proof that polyphenol oxidation is involved in their production, because these studies do not exclude the possibility of the presence of amine oxidase as a contaminant. Nonetheless, there i s compelling evidence for the role of catechins in aroma generation in black tea. The mechanism also suggests the formation of a nitrogen-substituted catechin [46]. A simi lar nitrogen-substituted catechin is formed by heating epicatechin with alanine in a model system producing 7-C(N-ethy lamino)epicatechin [47] (Anan et a! . , 1 987), which is a brown compound suggestive of the thearubigens. This is consistent with observations of nitrogen incorporation into catechins (Kito et a!., 1 968; Konishi, 1 969). These studies lead one to believe that a small fraction of the thearubigen fraction may possess nitrogen content derived from these compounds. The nitrogen substituted catechins are depicted in Figure 20.
4. Oolong Tea Polyphenols
Typically understood as an intermediate between green and black teas, oolong tea is characterized by a much shorter fermentation time under gentler conditions such that
partial oxidation, rather than total fermentation, occurs. This characterization is also representative of some Darjeeling teas, whose oxidase activity is not permitted to come to full expression. Gentler oxidation apparently creates its own unique set of aroma and polyphenolic compounds.
One residual question concerning the mechanism of theaflavin formation is the concerted oxidation of the catechins. Compounds such as the bisflavonols, also known as theasinensins A-E, represent "dead end" condensation products because they cannot proceed mechanistically to the theaflavins. Figure 2 1 i l lustrates the most plausible mechanism of theasinensin and theaflavin formation. It is likely that the quinone groups are present in low concentrations at any one given time due to their high reactivity. It seems more reasonable that the quinone reacts with an unoxidized polyphenol at low quinone concentrations, and is then oxidized further to theaflavin. Theaflavins may be formed directly from the quinones provided they are present in sufficiently large quantities but might be interrupted at an intermediate stage. Thus, Figure 2 1 also shows how the theasinensins may arise during their formation.
a. O% ngtheanin and Theasinensins
Oolongtheanin and theasinensins F [42] and G (Hashimoto et a!., 1988) may be unique products of the oolong tea fermentation system a n d are shown i n F igure 22. Theasinensins AlB and DIE are atropisomers about the biphenyl link of the same condensation products (the gallocatechins). Oolongtheanin is likely to be a further oxidized form of one of these theasinensins. Intriguing to note, however, is the detection of theasinensins F and G, atropisomers of a mixed condensation of epicatechin gallate and epigallocatechin gallate. Such a mixture
447

�OH
HOyyOi " ,Y � " 'Of-['4HCHRCOOH
OH
[46] : N itrogen Substituted Catechin, R=derived from amino acid N H2CHRCOOH
[47]: 7 -C(N-ethylamino)-epicatech in
FIGURE 20. Nitrogen-substituted catechins.
is expected to produce theaflavin digallate, as epicatechin [ ! O] and epigallocatechin [ 1 2] will produce theaflavin [30]. However, the gentler oxidation present in oolong tea systems may have permitted the detection of this intermediate.
In addition to oolongtheanin [55], two oolonghomobisflavans, and a vitamin C derivative of EGCG, 8-ascorbyl EGCG [56], were also isolated from oolong tea and may be unique oolong tea polyphenols (Hashimoto et aI., 1 989b). It is difficult at this time to place oolong tea polyphenols as either unique polyphenols of the divergent path of oolong tea production or as interme-
448
diates on the path between green and black tea, with green representing the unfermented leaf most closely and black tea representing either complete or near-complete fermentation.
IV. BIOCHEMISTRY OF TEA
Tea is the subject of many biochemical investigations because it produces a number of unique natural products. In contrast to the isolation and quantification of the various components of tea, the mechanisms by which the tea plant forms these compounds, and

OH 7' OH
:::,.. " 'OH OH [1 0]: EC- R=H
[12]: EGC- R=OH
° c
[4�Q) GOH
[1 2]9yOH
X�OH
Low quinone concentration
o
[48ao
xr;.-�) [49� �0
X�O
High quinone concentration
X H Theaflavin
OH
x
..
HO
X
X
[0]
PPO •
OH
X
OH
OH
OH [50]
o
[52]
HO
o
� " 'OH OH [48]: EC- R=H
[49]: EGC- R=OH
0
PPO ..
OH
HO
/
[54]
H
FIGURE 2 1 . Mechanism of theaflavin formation.
449

OH . . ,OH
HO h °
'>-- I " OH H0'Q:)0 ' " � J OH
'OGallale OH
[55]: Oolongtheanin
HO
HO_-J. ...
OH
OH
. . ,QOH " OH
'OGallale
[56]: 8-C-Ascorbyl Epigallocatechin Gallate
FIGURE 22. Oolong lea polyphenols.
the enzymes that activate these mechanisms, provide their own set of challenges to tea chemistry.
A. Caffeine Formation
Caffeine had been thought to he principally synthesized during the withering stage of freshly plucked tea leaves (Roberts and Sanderson, 1 966), although is probably synthesized throughout the life of the plant. Caffeine is most likely synthesized from adenine nucleotides, the dominant free purine forms in tea (Takino et aI., 1 972). Adenosine is a major product of RNA metabolism in tea (Imagawa et aI., 1 976). Adenosine [57] is converted to adenine [58] (Imagawa et aI., 1 979), and through hypoxanthine [59]
450
(or inosine) to xanthine [60] (or xanthosine), from which xanthosine [6 1 ] is the starting branch of caffeine biosynthesis (Negishi et aI., 1 992). Guanosine is also converted to xanthosine, but plays an apparently minor role in caffeine biosynthesis (Negishi et aI., 1 992). Xanthosine is methylated at the 7-position (Negishi et aI., 1 985) to 7-methyl xanthosine [62]. 7-Methyl xanthosine is then hydrolyzed to 7-methyl xanthine [63] (Negishi et aI., 1 988), which is subsequently methylated to theobromine [2] and caffeine [ 1 ] (Suzuki and Takahashi, 1 975; Suzuki and Takahashi, 1 976). The final methylation can be terminated, as found in the variant C. irrawadensis (Roberts et aI., 1 958). Figure 23 illustrates the mechanism of formation of theobromine and caffeine in the tea plant.

NH,
NH, t-1=N
� 0 N�N N N " H �N l:" ,J-) ( Adenine " OH '
� 1 , N 9 " OH [58) N�N O""'N N , l:" ,� � 8 9,··OH
" 'OH "N N / '" Hypoxanthine " �O OH
� � ' N N HO Adenosine I 1 �
[57) O""'N N/xanthOSJne (61)
Xanthine (60)
CH3, 01]
W-(f ,·,OH
�/N;N'H ') o HO 7 -Methyl-xanthosine (62)
�o l CH3 H'N
� I 1 I) O""'N N
I H
7-Methylxanthine (63)
o CH 0 CH3 H'N��
3 CH3-..N��
1 1 ) --- I I ) O""'N N O""'N N
I I C� C� Theobromine (2) Caffeine ( 1 )
FtGURE 23. Biosynthesis of caffeine,
B. Theanine Formation
Theanine [3] (n-ethyl glutamine) is fonned from the action of thea nine synthetase [L-Glutamate:Ethylamine ligase] (Sasaoka, 1 965; Sasaoka et a!., 1 965) on ethylamine derived from alanine (Takeo, 1 974), and
glutamic acid. Theanine degradation products may serve as precursors for synthesis of the catechin A ring, apparently from utilization of the N-ethyl group (Kito et a!., 1 968; Feldheim et a!., 1 986). It is likely that the N-ethyl group enters the general glycolyte pathway, forming the catechin precursor ma-
451

10nyl-CoA. Theanine formation is a lightdependent process requiring the presence of ATP as a cofactor (Sasaoka, 1 965). Theanine synthesis and accumulation is inhibited by production of glutamine (Matsuura and Kakuda, 1 990).
C. Biochemistry of Flavonoid Compound Formation
The pathways for the de novo biosynthesis of f1avonoids i n both soft and woody plants have been generally elucidated and reviewed in detail elsewhere (Jain and Takeo, 1 984; Heller and Forkmann, 1 994). Simi lar pathways for biosynthesis are used by a wide variety of plant species. The regulation and control of these pathways in tea and the nature of the enzymes involved in synthesis in tea have not been studied exhaustively. The following discussion extracts critical information from the pertinent tea literature and otherwise assumes that flavonoid production by Camellia sinensis generally follows the pathways used in other plant species.
1. Phenylalanine and the Shikimate Pathway
Phenylalanine is thought to be the direct precursor of polyphenols in tea plants (Nikolaeva et aI., 1 982); however, tyrosine may also participate in polyphenol production through a l ight-dependent pathway (Zaprometov and Bukhlaeva, 1 97 1 ) . Deamination of phenylalanine [64] forms cinnamic acid [65] by the action of phenylalanine ammonia lyase (Iwasa, 1 976; Shiplova and Zaprometov, 1 977; Zagoskina et aI., 1990). Hydroxylation of cinnamic acid forms coumaric acid [66], which is generated from an enzyme complex referred to as cinnamate
452
4-hydroxylate or cinnamate 4-monooxygenase. Hydroxylation is achieved typically by a cytochrome P-450 enzyme activity associated with the enzyme complex (Heller and Forkmann, 1 988), and a light-independent cinnamate 4-monooxygenase activity has been reported from tea shoots (Saijo, 1 980). The coumaric acid aromatic ring forms the 'B' ring of the f1avonoids. Figure 24 il lustrates the biosynthesis of coumaric acid and the various functions this compound performs in the tea plant.
The gallic and quinic acids origi nate via the shikimate/arogenate pathway. The key enzymes in shikimic acid biosynthesis have been detected in tea (Saijo and Takeo, 1 979). Carbohydrates play an important role, presumably as a precursor to shikimic acid, because radiolabels from both myo-inositol and glucose are incorporated into catechins (Wang and Huang, 1 987). Gallic acid and quinic acid play key roles in forming esters with various polyphenols. Gallic acid i s a key component of tannins and gives the catechins their tannin-like qualities.
Light has been shown to strongly induce biosynthesis of the phenolics by stimulation of the various enzymes along the biosynthetic path. Photosynthesis is associated with production of acetyl-CoA, ATP, and reducing agents such as NADPH, all of which play important roles as various stages of the biosynthesis (Zaprometov, 1 987). Light is very i mportant for catechin production, and shaded tea plants produce finished tea products with reduced astringency due to the lower concentrations of polyphenols (Mahanta and Baruah, 1992).
Coumaric acid, along with other cinnarnic acid derivatives and phenylalanine, is also a precursor to tea plant lignins (Zagoskina and Zaprometov, 1 976; Strekova et aI., 1980). Lignins are ubiquitous throughout the plant kingdom. Tea lignins are methylated (reinforcing low solubility) by the action of

�COOH V NH,
[64] : Phenylalanine
l Phenylalanine ammo nia· lyase
�COOH
o
[65]: Cinnamic Acid
! Cinnamate 4-monooxygenase
O):COOH I NIL HO � '"
,v �COOH/ [67]: Tyrosine � Tyrosine Ammonia-Lyase?
HO)0J
[66]: Coumaric Acid
/ \ to catechins and flavonols to lignins
+ quinic acid
(through navonoid biosynthesis) (through lignin condensation)
o
Hoocv-Z�OH OH"'---{'OH
OH [22]: Coumarylquinic acid
FIGURE 24. Cinnamate biosynthesis.
methyl transferase enzymes (Nikolaeva and Zaprometov, 1 990).
2. Chain Extension, Hydroxylation
After formation of the B ring moiety, the biosynthesis of catechins proceeds with a series of chain extensions to form the C and A ring backbone. 4-Coumaroyl-CoA is the
preferred substrate for chain extension in most plant species, typically formed by a hydroxycinnamate :CoA-ligase (Heller and Forkmann, 1 988).
Malonyl-CoA is produced by carboxylation of the glycolysis product acetyl-CoA via acetyl-CoA carboxylase (Lowenstein, 1981) . A hydroxycinnarnic acid-CoA derivative typically combines with three molecules of malonyl-CoA to form chaIcones via chal-
453

cone synthase (Ebel and Hahlbrock, 1 982). This enzyme apparently requires no cofactors and performs the condensation of the malonyl units and the cyclization to form the phloroglucinol 'A' ring (Heller and Forkmann, 1 994).
Theanine has also been demonstrated to be a significant factor in the synthesis of the phloroglucinol nucleus (Kito et aI., 1 968; Feldheim et aI., 1 986). Incorporation of the ethyl group of theanine into the catechins has been demonstrated by radiolabeling ethylamine, a theanine precursor (Sasaoka et aI ., 1 962; Kito et aI., 1 968). Ethylamine is the best demonstrated substrate of tea amine oxidase (Tsushida and Takeo, 1 985), which is converted to acetaldehyde. Theanine may be a storage mechanism for this amine, with acetaldehyde being the first intermediate on the path to catechin B-ring synthesis, followed by conversion to acetyl-CoA and/or malonyl-CoA.
Camellia sinensis produces flavonoids in which the tri-hydroxy B ring species predominates; however, within the flavonols the di-hydroxy B ring is more common. It is unclear whether 3' and/or 5' hydroxylation is occurring before or after chalcone synthase (Hahlbrock and Grisebach, 1 975). Because 3',4' -hydroxylated forms of all the species can be found throughout, and 4'mono hydroxylated species are found more commonly in the earlier branches of biosynthesis, it is possible that tea contains nonspecific or numerous 3' -hydroxy lases (Heller and Forkmann, 1 988), accounting for the presence of caffeic acids, quercitin, and catechin derivatives; a much more specific 3',5'hydroxylase (Heller and Forkmann, 1988) accounts for the gallocatechins and the smaller quantity of myricetin derivatives.
3. Chalcone/Flavone Tautomerization
Subsequent to formation of the chalcone, the final step in formation of the catechin
454
basic structure is tautomerization of the chalcone structure to a flavanone. The enzyme in this step is chalcone isomerase, which usually bypasses the natural, racemic, anti-addition (Heller and Forkmann, 1 988) to form a characteristic 2S-stereochemistry (Heller et aI., 1 979) of the resulting flavone by syn addition (Ebel and Hahlbrock, 1 982).
Ring closure of chalcones followed by hydroxylation results i n formation of dihydroflavonols. The flavanone is typically hydroxylated by the enzyme flavanone 3-hydroxylase, an oxoglutarate-dependent dioxygenase (Heller and Forkmann, 1 988). The formation of the 2R,3R dihydroflavonol has been shown to form either the f1avonols or the non-epi flavan-3-ols, whereas it is believed that a competing hydroxylase forms the 2R, 3S dihydroflavonol [70], which has been isolated (Nonaka et aI., 1 987).
4. Flavanols/Flavonols
The dihydroflavonols are enzymatically transformed to the f1avonols through action of a 2-oxoglutarate-dependent dioxygenase (Heller and Forkmann, 1 994), forming a double bond through abstraction of vicinal hydrogens on the 2R,3R dihydroflavonols as proposed for flavone synthase (Britsch, 1 990). This activity is referred to as flavonol synthase.
Formation of the 2R, 3S dihydroflavonols is likely to be preferred in the tea plant, as formation of the epi-forms of the f1avan-3-ols is the preferred biosynthesis product. However, while the route to formation of the f1avonols and non-epi-flavan-3-0Is is well defined, the formation of the epi-flavan-3-ols remains conjecture (Stafford, 1 988). Figure 25 illustrates the proposed biosynthetic pathway from cinnamic acids to flavonols and flavan-3-ols.
The formation of proanthocyanidin polymers has also been considered, although i t is unclear if epicatechin [ 1 0] or leucocyanidin [27] is the significant precursor (Nikolaeva

HO
0ySCOA CH,yO Pi OH Jx Acelyl CoA CH, SCoA '"
COAS'y'CH, CoAS I 1661: Coumaric acid
OH
OH °
8 Y (as CoA ester)
/:Chalcone synlhase
OH Chalcone isomerase
-
[68}: chalcone
("""y0H HOyyOj " V
lrY OH ° 1691: 2R-Flavanone
hydroxylase F,av7none
�OH Howo . .. 0 1 .& '. 'OH Dihydronavonol
OH ° � reductase? [70): 2R.3S-dlhydroflavonol �
������� j �OH
�OH Hoyyoil · · ·0 YY " OH
OH ° 1 glycosylalion 172) : navonol
( 1 7) Kaemplerol glycoside
HOyyOj . . ·V
lrY"'OH [7 l}. flavan-3.4-diol
OH 6H ! Flavan-3.4-diol reductase?
r"Y0H
HoYYOj .0
� 'OH OH 1 1 4) novan-J,ol (alzelechin)
FIGURE 25. Biosynthesis of flavonoids.
5_ Esterase et aI., 1 982). Proanthocyanidin polymer formation has been attributed to polyphenol oxidase and the formation of the thearubigens (Brown et aI., 1 969). Further work is necessary to determine the origin of proanthocyanidin polymers and their role in the tea plant and beverage.
Shikimic acid has been shown to be a good precursor for biosynthesis of the gallate group (Zaprometov, 1 962). Gallic acid, found principally in its esterified form in green tea, is thought to be principally syn-
455

thesized through this pathway, although phenylalanine and the cinnamic acids may also be precursors (Saijo, 1 983).
A significant quantity of the small polyphenols exists as quinic acid esters (depsides), particularly the depside of gallic acid, theogallin (Stagg and Swaine, 1 97 1 ). Quinic acid is derived from the shikimic acid pathway via dehydroquinic acid, an intermediate on the pathway to shikimic acid (Sprinson, 1 960; Neish, 1 964). Quinic acid esters of coumaric and caffeic acid (chlorogenic and caffeoylquinic acid) have also been detected i n tea (Sanderson, 1 972).
Gallated f1avan-3-0Is (catechins) are the major flavonoids produced by Camellia sinensis (Sanderson, 1 972). It is of interest that Camellia sinensis forms gallated esters rather than sugar esters, or glycosides, of catechins. Glycosides are the typical plant strategy for rendering the major f1avonoids water soluble (Heller and Forkmann, 1 994), although proanthocyanidin and catechin glycosides, while not rare, are not known to be common (Achmadi et aI., 1 994). It is likely that the catechins are acylated with an activated form of gallic acid, presumably galloylCoA, similar to the aromatic acylations occurring with cinnamic acid acyltransferases in other systems (Kamsteeg et aI., 1 980; Heller and Forkmann, 1 988). Gallic acid esterification is thought to be a slow process compared with its biosynthesis (Zaprometov and Bukhlaeva, 1963) and has been demonstrated through tracer experiments in tea shoots (Saijo, 1 983).
D. Latent Enzyme Activity and the Formation of Black Tea
Conversion of tyrosine and related monophenols to diphenolic compounds is typically accomplished by tyrosinase or monophenol monooxidase. Whether tyrosinase activity or a related enzyme acti vity is
456
responsible for polyphenol oxidase activity (observed in leaf browning and the formation of black tea polyphenols) is unclear.
1. Polyphenol Oxidase
Polyphenol oxidase (EC 1 . 1 4. 1 8 . 1 ; monophenol monooxygenase [tyrosinase] or EC 1 . 1 0.3.2; o-diphenol: O2 oxidoreductase) is one of the more important enzymes involved with the formation of black tea polyphenols. The enzyme is a metallo protein, thought to contain a binuclear copper active site. Polyphenol oxidase (PPO) is an oligomeric particulate protein that is thought to be bound to the plant membranes. The bound form of the enzyme is latent and activation is likely to be dependent on solubilization of the protein (Tolbert, 1 973). PPO is distributed throughout the plant (Durmishidzern and Puridze, 1 980) and has been identified and analyzed in the floral organs (Singh and Ravindranath, 1 994). PPO is localized within plant cells i n the mitochondria (Bokuchava et aI., 1 970), the chloroplasts (Roberts, 1 94 1 ), and the peroxisomes (Kato et aI., 1 976). Using antibody techniques, polyphenol oxidase activity has also been localized in the epidermis palisade cells (Wickremasinghe et aI., 1 967). Recent reviews on the subject of PPO are available (Zawistowski et aI., 1 99 1 ; Whitaker, 1 994; Steffens et aI., 1 994).
The biological role of PPO in plants is thought to be associated with a plant defense mechanism and root development. PPO becomes activated and available when plant tissue is damaged due to injury or infection, catalyzing the formation of insoluble phenolic polymers that aid in wound healing and help prevent the spread of infection (Zawistowski et aI., 1 99 1) . In tea, key reactions occurring during "fermentation" are catalyzed by PPO. These reactions are initiated by crushing and/or tearing withered tea

leaves and lead to the formation of black tea polyphenols and aroma compounds characteristic of black tea.
Polyphenol oxidase catalyzes two general reactions: the hydroxylation of monophenols to o-diphenols, and the oxidation of diphenols via dehydrogenation to o-quinones, both 2-electron transfer reactions. The formation of o-quinones is then followed by condensation to form a wide range of complex products (Mayer and Harel, 1 99 1 ). The copper cofactor associated with the metallo protein is the essential component involved with electron transfer and oxidation of the substrate. The reaction mechanisms associated with PPO of Camellia sinensis have not been determined; however, the reaction mechanisms of PPO from other plants have been characterized (Zawistowski et aI., 1 99 1 ; Whitaker, 1 994; Steffens et aI., 1 994). PPO activity is dependent on oxygen concentration (Robertson, 1 983). The o-quinone species from tea has been looked for in a model system (Korver et aI., 1 973).
Polyphenol oxidase is difficult to purify, due in part to the presence of polyphenols that have a strong affinity for proteins. Work on isolation and characterization of PPO has been reviewed (Mayer, 1 987). The enzyme appears to exist in multiple forms and is comprised of a number of subunit proteins. Reports on the molecular weight of PPO in plants vary widely. The early work on isolation of PPO from green tea leaves reported a molecular weight in the range of 1 30 to 1 60 kDa (Gregory and Bendall, 1 966). Reports on isolation of PPO from other plant species using modern biochemical methods state molecular weights ranging from 40 to 72 kDa, whereas the primary isoform oftypical PPO has a molecular weight of 45 kDa (Steffens et aI., 1 994). The molecular size of PPO isoforms based on cloned genes indicates that PPO has a molecular weight of between 57 and 65 kDa and includes punitive transit peptides of a molecular weight
of I g to 20 kDa. Mushroom tyrosinase has been found to have a quarternary structure comprised of two different subunits of 43 kDa and 1 3.5 kDa, with a total molecular weight of 1 20 kDa and the formula L,H2 (Zawistowski et aI., 1991) . These facts would suggest that the PPO from tea is also comprised of four protein subunits and therefore has a possible molecular weight of - 1 44 kDa. This is consistent with a report of isoforms of 1 17, 56, 4 1 .5 , and 35 kDa (Buzun et aI., 1 974) and another report with isoforms al 1 1 8 and 4 1 kDa (Durmishidzern and Puridze, 1 980). The copper content has been measured at 0.26 wt/wt%, or about 4 to 5 copper atoms for the complex, which is consistent with other literature (Vamos-Vigyazo, 1 98 1 ; Interesse et aI., 1 983; Zawistowski et aI., 1 99 1 ) . This number is probably low because copper diffuses readily away from the enzyme, resulting in a loss of activity (Mayer, 1 987; Zawistowski et aI., 1 99 1 ) . Consistent with observed sigmoidal activity (Pruidze, 1 975), multiple active sites in the enzyme complex are likely to exist.
Polyphenol oxidases act on a broad range of phenolic substrates with both mono-, di-, and trihydroxy substitutions. There is evidence that PPOs from different plant species have preferential substrates (Whitaker, 1 994; Steffens et aI., 1 994). Studies of PPO from tea have mainly been based on model fermentation studies that utilize purified or semipurified substrates (Dix et aI., 1 98 1 ; Robertson and Bendall, 1 983 ; Opie et aI., 1 990; Finger, 1 994). It is clear from these reports that PPO from tea reacts effecti vely with both di- and tri-hydroxylated catechins, reacting with specificity for the o-dihydroxy moiety (Gregory and Bendall, 1 966; Coggon et aI., 1 973). Studies that define the kinetics of PPO from tea in relation to substrate type are needed to better define the enzymology of tea PPO. PPO can be inhibited by a range of chemical constituents that include cyanide, carbon monoxide, EDTA,
457

sulfites, ascorbic acid, erythrobic acid, and thiol-containing compounds (Zawistowski et aI., 1 99 1 ) . In general, compounds that block oxygen, act as antioxidants, and/or bind copper are good inhibitors of PPO. The effect of natural flavonoids and end products of fermentation as inhibitors of tea PPO remains to be defined, although there i s e v i dence of theaflavins and thearubigens as feedback inhibitors (Pruidze and Grigorashvili, 1 975). Most PPO enzymes have a pH optimum in the range of 5.0 to 7.0. Typical solutions of tea polyphenols have a pH of approximately 4.5, and increasing the pH of tea fermentation reactions leads to stronger PPO activity. Tea PPO has an observed optimum pH in the range 4.6 to 5.6 (Takeo, 1 965; Takeo and Uritani, 1 966; Gregory and Bendall, 1966; Perera and Wickremasinghe, 1 972; Coggon et aI., 1 975; Robertson, 1 983).
2. Peroxidase
Peroxidase (EC 1 . 1 1 . 1 .7) is thought to play an integral role in the fermentation process and is found in fresh green leaf (Bokuchava and Popov, 1 948; Gregory and Bendall, 1 966; Bokuchava and Skobeleva, 1 969). Coupled with PPO, which is thought to produce the peroxide, an acti vator of this enzyme system in other systems (Jiang and Miles, 1993), peroxidase is also thought to play a role in the oxidation and formation of the black tea compounds.
Peroxidase is a haem-based enzyme that catalyzes the reductive decomposition of hydrogen peroxide to water and organic peroxide species to the corresponding alcohol. In contrast to superoxide dismutase, whose activity is cycled by coupled oxidation and reduction of peroxide to oxygen and water, respectively, peroxidase cycles its activity through oxidation of a wide variety of substrates.
458
Model system studies using peroxidase and PPO for production of black tea polyphenols suggest that peroxidase activity is responsible not only for decomposition of theaflavins but the formation of thearubigens as well (Dix et aI . , 1 9 8 1 ) . Reactions of epicatechin and PPO have been proposed as a model for thearubigen formation (Opie et aI . , 1 993) and epicatechin oxidation products by this system have been suggested as a good model of thearubigens (Opie et aI., 1 995). This is likely due to the fact that the isolation procedure used for purification of PPO (Opie et aI., 1 990) was found to contain peroxidase activity in an earlier report (Robertson and Bendall, 1 983). Model fermentations using PPO and peroxidase, both separately and in conjunction, show the coupled nature of these enzymes (Finger, 1 994) and point to the need for further investigation of the role of both of these enzymes i n the development of the unique black tea polyphenols and other compounds present in black tea extracts.
E. Extracellular Enzymatic Activity
Many tea products, such as Kombu-cha or Pu-Erh tea, take advantage of enzymatic systems external to the tea plant. In the case ofKombu-cha, sweetened tea brew is seeded with a yeast that produces ethanol and acetic acid. The yeast generates a mushroom-like mass that is typically reused as the seed for future beverage production. Pu-Erh tea is produced by burying green tea leaves in a confined space, thereby allowing microbiological fermentation to blacken the leaves (Shao et aI. , 1995). Pu-Erh is thought to possess many health benefits and is often compressed into bricks and called Tuo-cha tea.
Tea, because of its unique phytochemical composition, offers many opportunities for study of exogenous enzyme systems. One enzyme that has gained a reputation in the

field of tea chemistry is tannase. Tannase enzymes (Gallic acid esterase, Tannin acyl hydrolase, EC 3. I . I .20) such as those isolated from fermentation broths of Aspergil
lus Flavus, A. Niger, or A. Oryzae represent the most significant of these extracellular enzymes. Tannase is significant in terms of its demonstrated effect of hydrolyzing the gallic acid group from tea polyphenols (Deijs and Dijkman, 1 936; Roberts and Wood, 1 95 1 ; Roberts and Myers, 1 959) and its beneficial impact on manufacture of cold water soluble instant teas (Coggun et ai., 1 975). The use of tannase significantly inhibits the ability of tea polyphenols to complex and precipi tate, and reduces the ability of black tea to form tea cream (Nagalakshmi et ai., 1 985). Tannase activity and thermal stability in the presence of tea extracts has been determined (Thomas and Murtagh, 1 985).
V. CHEMICAL PROPERTIES OF TEA COMPOUNDS
Beyond the inherent ecological properties of tea compounds in the tea plant, tea phytochemicals exhibit unusual properties that make them interesting subjects for chemical study.
A. Formation of Cream and Haze in Black Tea
The onset of cooling in a freshly brewed cup of black tea is accompanied by production of a dark red/brown cloud. This cloud is particularly notable in concentrated brews of tea, where the beverage moves from a dark brown to a milky-red color. This apparent 'lightening' in the concentrated solution resembles the milky-red color of tea to which cream has been added and thus was dubbed 'tea cream' (Bradfield and Penney, 1 944; Roberts, 1963; Smith, 1 968).
Tea cream can be separated from the brew most conveniently by centrifugation of a concentrated beverage (Smith, 1 968).
Tea cream was demonstrated by early researchers to contain caffeine, theaflavin, and thearubigens, as well as traces of a number of other substances (Roberts, 1 963; Smith, 1 968). Studies on cream composition (Nagalakshmi and Seshadri, 1 983) and decreaming (Nagalakshmi et ai., 1 985) show that protein and caffeine contribute significantly to cream formation. Additional contributions from lipids in tea such as triacontanol and spi nasterol have been shown (Seshadri and Dhanaraj, 1 988). Simple carbohydrates have been shown to assist in solubilization of cream (Nagalakshmi et ai., 1984) but do not appear to complex significantly with polyphenols in model studies (Williamson et ai., 1 995). High-molecularweight thearubigens have been associated with cream formation (Hazarika et ai., 1 984). HPLC analysis of the phenolic portion of tea cream (Powell et ai., 1 993) revealed 86% 'thcarubigens', 1 2% theafluvins, and 2% flavonol glycosides.
Whi le the polyphenolic constituents' role in cream formation is well understood, the presence of proteins/peptides, carbohydrates, leaf fines (fragments of leaves), and other materials needs to be studied further to ful ly determine the mechanism behind cream formatio n . Analysis of the p H optimum for cream formation (Smith, 1 968), which closely mirrors tannin-BSA pH curves (Hagerman and Klucher, 1 986), combined with knowledge of the significant contribution of protein to the mass of cream (Nagalakshmi and Seshadri, 1 983), provides strong justification for better analysis of the contribution of protein to cream formation.
The study of the physicochemical properties of cream is reviewed and expanded significantly in thesis work (Rutter, 1 97 1 ) and in the literature (Rutter and Stainsby,
459

1 975; Bee et aI., 1 987). Tea cream has been shown to be comprised of ultra fine spherical colloidal droplets that are clearly visualized in a range of concentrations of tea in water, increasing in size with increasing concentration. At very high concentrations, the droplets begin to aggregate further, forming irregular shapes. The colloidal, spherical nature of the tea cream particles distinguishes this material from typical "crystalline" precipitates and is referred to as a coacervate (Bee et aI., 1 987). Tea cream formation has been interpreted in terms of phase separation and is likened to two liquids miscible at high temperatures but immiscible when cooled (Harbron et aI., 1 989).
B. Complex Formation
The onset of cream formation in tea is driven by complexation of the black tea polyphenols. Green tea contains simple polyphenols when compared with the complex polyphenols present in black tea. Green tea exhibits formation of a haze but does not 'cream' to the same extent as black tea. Polyphenols have been observed to have a strong precipitating effect on proteins, denaturing and reducing enzymatic activity (Sekiya et aI., 1 984; Ozawa et aI., 1987). Immobilized polyphenols bind proteins reversibly, with restoration of enzyme activity after elution (Oh et aI., 1 985). Dissolved ash metals such as hard water calcium may influence complexation, as in the case of tea scum formation (Spiro and Jaganyi, 1994).
The mechanism of complex formation is a subject of some dispute in the literature. It has been asserted that polyphenols form a "hydrophobic bond" between the flat aromatic surface of the polyphenol rings and quasi-planar hydrophobic groups such as proline in pep tides and proteins. Evidence for this assertion is deri ved from crystal structures of co-crystallized model polyphenols
460
with caffeine (McManus et aI., 198 I ; Gaffney et aI., 1 986; Martin et aI., 1 986), and of caffeine with I-tryptophan (Nishijo et aI., 1 990). Hydrogen bonding also plays a prominent role (Van Sumere et aI., 1 975; Borazan and AI-Ani, 1 980). Although a full thermodynamic treatment of the issue is beyond the scope of this review, it is hoped that further investigation of the following issues will lead to a better understanding of the mechanism of cream formation.
Polyphenols demonstrate hydrophobic character by partitioning into the cyclodextrin cavity (Spencer et aI., 1 988; Cai et aI., 1 990) and by their octanol-water partition coefficient (Martin et aI ., 1 990). In addition to hydrophobic stacking, there is evidence for a coplanar-type hydrogen bond (Martin et aI., 1986). There is an apparent correlation between hydrophobicity and complexation for hydrolyzable tannins such as pentagalloylglucose and vescalagin (Haslam, 1 993). Hydrophobic association has also been used to explain copigmentation effects observed in solutions of anthocyanins and polyphenols (Mistry et aI., 1 99 I ; Liao et aI., 1992) or xanthy l i u m dyes with caffeine or cyclodextrins (Dangles and Brouillard, 1 993). Polyphenols bind significantly to prolinerich proteins, and this complexation i s thought to drive taste perception and differences among various methods of tea beverage consumption (Luck et aI., 1 994).
Closer inspection of data on condensed tannins (Martin et aI., 1 990) reveals interesting correlations. To illustrate, an increase of one hydroxyl group decreases apparent hydrophobicity yet increases association with caffeine for epicatechin/epigallocatechin, catechin/gallocatechin, and catechin gallate/ epigallocatechin gallate pairs. Despite their hydrophilic nature and twisted structure, procyanidins show simi lar complexation with caffeine. The additional hydroxy group may increase the hydrogen-bonding character, thereby inducing greater association with

caffeine. Those polyphenols should also exhibit less hydrophobicity because the extra
hydroxy group should prefer a hydrophilic environment. Hydrogen bonding has been demonstrated to be a strong driving force for
association, particularly i n systems such as
cyanuric acid with melamine (Mathias et aI.,
1 993) and p-cresol with caffeine (Borazan and AI-An i , 1 980). The hydrogen-bonding character of catechin/polyproline association
has been demonstrated in model systems (Sun
and Mattice, 1996). Simple hydrophobic association would
seem to be insufficient for explanation of
protein complexation, precipitation, and
cream formation. A simpl ified model for the complexation of polyphenols and caffeine (Figure 26) demonstrates that association of
the two hydrophobic surfaces results in a net
decrease of hydrophobicity because the
polyphenols have a hydrophobic core sur
rounded at its edge by hydrophilic groups. Al l (or most) of the hydrophilic surface is
available for solvation, which should result
in a soluble, not precipitated, complex. This
mirrors the formation of micelles, which are commonly understuud tu furm a dispersed '·pseudophase". It also mirrors the observed
association of two "" h;dmphobi, '"""/
key: hydrophobic surface
hydrogen bonding surface
'" results in form ing a
J
complex whose hydrophobic area is reduced, but hydrogen-bonding area is the same (similar to a micelle and caffeine self-aggregation)
However, hydrogen bonding reduces hydrophilic su rface area, which should drive separation of 'cream' phase
01· Caffeine to 01 Caffeine lO @1POIYPhenolt(j 01poIYPhenolto
FIGURE 26. The hydrophobic association model.
461

self-stacking of caffeine aggregates (Thakkar et aI., 1970; Shestopa et aI., 1985), which attempt to compensate for unfavorable hydrophobic association of the plane by aggregating in planar stacks of higher solubility. Cream formation, however, requires the formation of a discrete precipitate analogous to phase separation.
Existence of hydrophobic bonds (Xianqi and Haslam, 1 994) is supported only by circumstantial evidence. Hydrophobicity i s driven by the loss of mutual electrostatic stabilization of water molecules (or similar hydrophilic groups). Hydrophilic groups are forced to form ordered cages around a hydrophobic surface, placing the system at an entropic disadvantage. To overcome this disadvantage, the hydrophobic groups aggregate and separate from the hydrophilic surface in an attempt to minimize the surface area of this ordered cage. The hydrophobic surfaces are held together very loosely by van der Waals' forces. Aromatic surfaces have also been described as hydrophobic, but the mode of hydrophobicity uf aromatic molecules is apparently very different than that of aliphatic hydrophobic surfaces (Makhatadze and Privalov, 1 994). In addition, aromatic and other unsaturated molecules can form hydrogen bonds with the Itelectrons, stabilizing the aromatic group in the presence of water (Rzepa et aI., 1 994; Ernst et aI., 1995). There are also indications that aromatic stacking interactions in aqueous systems have little to do with dispersion forces or hydrophobic interaction (Newcomb and Gellman, 1 994).
A more satisfying model is one in which hydrogen bonding of polyphenols to caffeine (or hydrophobically associated yet soluble complexes to one another) causes a net reduction of hydrophilic surface area with little concomitant reduction of hydrophobic surface area. This places the complex at an electrostatic and entropic disadvantage, causing aggregation and precipitation in the
462
manner of phase separation. Proteins are most easily precipitable at their isoelectric point (Oh and Hoff, 1 987), when the ionic charge on a protein is lowest and the least number of 'water-solubilizing' ionic factors are present. The loose backbone of a protein can hydrogen-bond with the polyphenol, particularly at secondary ami des such as those found in proline residues, because secondary amides have been shown to form stronger hydrogen bonds than primary ami des (Cannon, 1 955; Hagerman and Butler, 1 98 1 ). After binding, there is a reduction in hydrophilic surface area, and the hydrophobic sidechains are reoriented outward into aqueous solution resulting in unfavorable interactions and precipitation. For larger proteins, binding of a polyphenol might occur as the hydrophobic core of the protein associates with the hydrophobic surface of the polyphenol. The strong hydrogen bonding interactions will then cause hydrophilic groups to turn inward, denaturing the protein and causing precipitation. Hydrophobic effects must also play an important role because detergents such as �-octyl-D-glucopyranoside can be used to solubilize the precipitated polyphenol complex (Martin et aI., 1 990). Surfactants are known to form ordered layers at liquid-solid interfaces (Bigelow et aI., 1 946), and the increase in entropy associated with forming ordered mono layers may represent a driving force for reversal of the hydrogen bond complex and aggregate association, resulting in increased water solubility of the polyphenols and their complexes.
Within this model, hydrophobic association and hydrogen bonding interactions are given similar importance, as both are necessary for driving the complex interactions involved in the model. That the proposed model does not currently explain apparent di fferences between the condensed and hydrolyzable tannins i s one l i m i tation. Vescalagin, castagalin, and pentagalloylglucose have simi lar hydrogen bonding sites,

although pentagalloylglucose precipitates proteins much more readily (Haslam, 1 993). This may be related to steric restraints, but different tannins can have different mechanisms of precipitation and thus no single model should be force-fitted to explain all polyphenol interactions, unless the generality can be rigorously proven and the exceptions well defined.
C. Polyphenols as Antioxidants
Research has progressed extensively on the chemical and biological properties and functionality of tea polyphenols. Charcterization of the antioxidant properties of tea flavonoids is a prime example of the newer research inititative.
There are numerous synthetic and natural compounds that block oxidative reactions in vivo and in vitro by quenching free radicals or by preventing free radical formation. These compounds have been termed "antioxidants" (Namiki, 1 990; Pokorny, 1 992). Vitamins A, E, and C and the mineral selenium are common antioxidants occurring in our foods (Pokorny, 1 992), and BHA (butylated hydroxyanisole), BHT (butylated hydroxy toluene), and propyl gallate are synthetic antioxidants approved for food use (Gates, 1 987). A broad range of flavonoid or phenolic compounds have been found to have antioxidant activity (Ho, 1 992). Polyphenols, both synthetic and from numerous plant sources, have also been found to be functional antioxidants in numerous test systems (Ho, 1 992; Lunder, 1 992; asawa, 1 992). Antioxidants are the control agents for regulation of oxidative reactions.
The antioxidant activity of tea extracts and tea polyphenols has been studied in a variety of model systems. It is clear that the polyphenols from green and black tea are effective antioxidants when evaluated in both chemically and biochemically based test
systems. Green tea polyphenols have been shown to be more effective than traditional antioxidants such as BHA, BHT, ascorbic acid, and vitamin E (Tanizawa et aI., 1 983; Tanizawa et aI., 1 984; Namiki and Ozake, 1 986; Zhao et aI., 1 989).
1. Chemical Antioxidant Systems
The effectiveness of purified tea polyphenols and crude tea extracts as antioxidants against the autooxidation of fats has been studied using the Rancimat system. I n one study, purified green tea catechins were evaluated against dl-a-tocopherol and BHA (Ho et aI., 1 992). On a molar basis, the antioxidant activity of the catechins was ranked in the following ascending order: EC <ECG <EGC <EGCG. Each of the catechins was more effective than either dl-a-tocopherol or BHA. A positive, synergistic effect between the catechins and ascorbic acid or dla-tocopherol has also been found (Matsuzake and Hara, 1985). Purified catechins and crude methanol extracts of green, oolong, and black teas have been shown to have significant antioxidant activity. Gallic acid was found to be a more effective antioxidant than any of the tea extracts or purified compounds, suggesting that the gallate moeties of the tea polyphenols are an important part of their expressed antioxidant acitivity (Ho et aI., 1 992). Antioxidant activity of extracts of a variety of green teas and blends of green and black tea has been evaluated. A direct correlation between the antioxidant index of a tea extract and the concentration of EGCG in the extract was shown (Lunder, 1 992).
An alternate method to the rancimat, based on measuring the oxidation of linoleic acid catalyzed by bubbling air (2 h incubation at 60°C with 500 ml/min air flow rate) and determining the peroxide value (POV) or the thiobarbituric acid value (TBA V) has been used to study tea flavonoids (Tanizawa
463

et aI., 1 983; Tanizawa et aI., 1 984). Epicatechin, green tea extract III (n-butanol phase) and green tea extract IV (acetone soluble portion of extract III) were found to have strong antioxidant activity equal to the activity of BHA and were significantly more potent than vitamin E. A similar model system that involves the air oxidation of linoleic acid incorporated into cetyl trimethylammonium bromide (CTAB) micelles has also been used to study the antioxidant activity of flavonoids (Wang and Zheng, 1 992). The micellar system is a better model for food emulsions and biological membrane systems, and micellar studies showed that flavonoids were active as chain-termination antioxidants by reacting with peroxy radicals. The flavonoids had the following descending order of reactivity: a-tocopherol >morin >rutin >quercetin. There is a clear difference between prevention of oxidation of free linoleic acid (the crude extracts of flavonoids were much more active than vitamin E) and linoleic acid in micelles (flavonoids were less active than vitamin E). It is likely that vitamin E is readily incorporated into the micelles, which assures close proximity of the antioxidant to the lipids.
Spin-trapping methods using ESR have been employed to evaluate the ability of green teas and other natural compounds to intercept and measure free radicals. Free radicals were generated by numerous sytems, including stimulated polymorphonuclear leukocytes (Zhao et aI., 1 989); hydroxy radicals produced in reaction mixture containing hydrogen peroxide and Fe++ (from ferrolls ammonium sulfate) (Zhao et aI., 1 989; Shi and Dalal, 1 99 1 ; Uchida et aI., 1 987); a reaction mixture of NADPH, potassium dichromate, and hydrogen peroxide (Shi and Dalal, 1 99 1 ); an irradiated riboflavin system (Zhao et aI., 1 989); and superoxide generated by the hypoxanthine-xanthine oxidase system (Uchida et a I . , 1 987) . DMPO (5 ,6-dimethyl- l pyrroline-N-oxide) was used to trap free radi-
464
cals and monitor their formation. These studies demonstrate tea constitutents as good radical scavengers, suggesting that tea polyphenols can act as antioxidants in termination of both the inititation and chain termination stages of lipid peroxidation.
A classic method of generating hydroxy radicals is the Fenton reaction, a reaction of iron salts and hydrogen peroxide. Studies to determine the effect of polyphenols in scavenging hydroxy radicals generated by the Fenton reaction resulted i n the following ranking of antioxidant potency: curcumin (69% inhibition), vitamin C (56% inhibition), vitamin E (35% inhibition), rosemary extract ( 1 6% inhibition), green tea polyp henol ( 1 2% inhibition), and so-called "green tea fraction 6" ( 1 3 % inhibition) (Zhao et aI., 1 989). Caffeine was also found to be an effective scavenger of hydroxy radicals generated by the Fenton reaction, but its effective concentration was three orders of magnitide higher than of green tea catechins or vitamins (Shi and Dalal, 199 1 ) .
The key active sites of flavonoids for scavenging of free radicals and for antioxidant activity are the o-dihydroxy structure in the B ring, the 2,3-double bond in conjugation with the 4-oxo function in the C ring in flavonols, and the 3- and 5-hydroxy groups with the 4-oxo function in the A and C rings (Nakayama et a I . , 1 99 3 ; Pratt, 1 992; Jovanovic et aI., 1 994). Catechins can react with more than one free radical to form quinones (Zhao et aI., 1992) and have a lower reduction potentials than vitamin E (Jovanovic et al.. 1 995). This should account for the stronger radical-scavenging activity of catechins. Tannins with a greater degree of gallation were more effective in trapping radicals on a molar basis than EGCG, suggesting that theaflavin digallate and gallated thearubigens from black tea could also be effective radical scavengers. Future research is needed to understand the potential healthpromoting properties of antioxidants, and

better characterization and understanding of the polyphenols in both green and black tea extracts is needed.
2. Biological Antioxidant Models
Biologically based model systems for the evaluation of antioxidants have been employed with tea extracts and polyphenols. These model systems mimic the reactions believed to be linked to the pathogenesis of some chronic diseases.
Catechin was found to act as an effective antioxidant in an iron· loaded rat hepatocyte system and was proposed to function through an ion chelation mechanism (Morel et aI., 1993). However, other studies using catechin as an antioxidant have suggested that its antioxidant activity is due to a free radicalscavenging mechanism (Fraga et aI., 1 987; Ratty and Das, 1 988). It is likely that both properties of tea polyphenols contribute to their antioxidant activity.
An erythrocyte membrane ghost system and a rat liver microsome system were used to study dietary antioxidants and oxidation processes (Namiki and Ozake, 1 986). The catechins from tea were found to be very active i n both systems, with EGCG and ECG having the greatest activity in blocking lipid oxidation. These catechins were 1 0 times more effective than vitamin E, but were less active than BHA. EGCG and ECG were effective against peroxidation initiated by either the chemotherapeutic agent adriamycin or the perferyl ion, while EGC was effective only against perferyl-ion induced oxidation (Osawa et aI., 1 992). This suggests that EGCG and ECG were effective as both radical scavengers and chelators, while EGC appears to be a weak radical scavenger but a good chelator. In the stimulated polymorphonuclear leukocyte system, a green tea polyphenol fraction and a green tea extract fraction called 6 (undefined) had much stron-
ger radical scavenging activity than vitamin C, vitamin E, rosemary extract, or curcumin (Zhao et aI., 1 989).
Gallic acid and gallic acid esters have been shown to have both antioxidant and prooxidant activity in vitro (Aruoma et aI., 1993; Cao et aI., 1 996). Noticeable oxidation of deoxyribose was observed with excess concentrations of gallic acid, suggesting that gallic acid may act as a prooxidant and play a role in prooxidant activity in green tea.
The cytotoxic effects of reactive oxygen species (0,· and H,O,) on cells grown in culture was used as a model system for the evaluation of the antioxidant activity of f1avonoids (Nakayama et aI., 1 993; Ruch et aI., 1 989). Chinese hamster lung fibroblast V79 cells in culture were incubated in the presence of f1avonoids for 4 h to allow sufficient time for binding of the polyphenols to the cell surface or cellular absorption after which the cells were washed to remove free f1avonoids. The washed cells were then exposed to reactive oxygen species (either H,O, or superoxide produced enzymatically using a combination of hypoxanthine and xanthine oxidase). In a cytotoxicity test of the f1avonoids, catechin and taxifolin were not toxic at concentrations up to 200 and 1 000 �M, respectively, while quercetin and kaempferol were toxic at concentrations above 1 00 �M. Quercetin and kaempferol were effective in protecting cells from both superoxide and peroxide at concentration of 5 � and 20 �M, while catechin was effective at concentrations of 500 �M and 1000 �M (Nakayama et ai., 1 993). A similar model system used cultured mouse hepatocytes rather than hamster lung fibroblasts (Ruch et aI., 1989). These studies showed that a green tea polyphenol fraction (GTP) was effective in scavenging H,O, and superoxide radicals by demonstrating the ability of GTP to inhibit cell death induced by oxygen radicals and peroxides produced by glucose oxidase or paraquat in
465

a dose-dependent manner. An in vivo experiment was conducted to determine the effect of green tea on radiation-induced lipid peroxidation and lethality (Uchida et aI., 1 992). Epigallocatechin gallate (EGCG) was provided to mice in drinking water for I month at 0.002% or 0.0 I % concentration providing an EGCG intake of 3 or 15 mglkgl d, respectively. The mice were then irradiated with 954 cGy X-ray and killed 3 d later. Oral consumption of EGCG inhibited irradiation-induced formation of hepatic peroxides i n a dose-dependant manner, reducing the formation of peroxides 75% and 90% for the 0.002% and 0.0 I % doses, respectively. Oral consumption of EGCG was also found to significantly extend the lifespan of mice given a lethal dose of radiation. These results support the conclusion that tea catechins are effective scavengers of free radicals in vivo.
Tea extracts, tea polyphenol fractions, and purified catechins have been shown to be effective antioxidants in both chemically and biologically based model systems. In both types of systems, the balance between oxidants and antioxidants is critical. Imbalances between free radicals and antioxidants can occur, caused by an increased production of free radicals andlor decreased effectiveness of the antioxidants within the reaction system. These imbalances can be caused by the radicals overwhelming the antioxidants within the system, or by an excess of antioxidants leading to a prooxidant functionality (Aruoma et aI. , 1 993). It is critical that a proper antioxidant to oxidant balance be maintained in well-controlled biological studies. The balance of different types of antioxidants i s also important, as demonstrated with carotenoids, vitamin E, and vitamin C (Bohm et aI., 1 997).
VI. TRENDS IN TEA RESEARCH
Tea chemistry as a research discipline has been characterized and influenced by the
466
techniques and approaches of each group of researchers. In order to understand much of the language and notation used in the current literature , it becomes necessary to contextualize the development of these terms and theories, and, in effect, deconstruct the science.
The application of HPLC in tea research represents a good example of this: as a new technology, it offered a radical new way of looking analytically and quantitatively at tea components. Yet, the diffuse understanding of the compounds i n tea, combined with qualitative descriptors historically available, has provided a challenging barrier to surmount. A thearubigen i s very clearly assumed to be a complex mixture of polymerized polyphenols that appears as a diffuse spot on paper chromatography. The identity and location of that spot becomes unclear when using a new and different technology. Without a definitive pure standard of a thearubigen, only a qualitative assessment can be made of an already qualitative descriptor. As a new technology breaks a previously homogenous mass of compounds into subdivided fractions, the relati ve differences in concentration of these subdivisions among different samples can negate the idea of homogeneity and cause dramatic reevaluations of theories that depended on the homogeneity of the original mass. Tea research has brought the thearubigens very close to this point, and the homogeneity assumed by this classification is beginning to show weakness.
The advancement of tea chemistry is dependent on comprehension of the li terature in the context of the directions of individual research groups.
A. Analysis by Chemical Constitution and by Technical Innovation
Early determinations of the chemical constitution of tea and tea beverages

(Kursanov, 1 956; Vuataz et aI., 1 959; Millin and Rustige, 1 967) set the stage for a generalized interpretation of the chemistry of tea (Sanderson, 1 972). The subdivisions of each category and the analytical accuracy of new methods have improved the understanding of the mass balance of tea chemistry. The subdivision technique has been propagated in the literature (Graham, 1 984; Balentine, 1 992).
No single technological innovation has propelled the efforts of tea chemistry more in recent years than the introduction of the HPLC (Hoefler and Coggon, 1 976), and the subsequent coupling of the technique with diode-array detection (Bailey et aI., 1 990; Opie et aI., 1 990; Opie, 1992) and mass spectrometry (Lin et aI., 1 993; Bailey et aI., I 994b). Despite its power, HPLC is in a stage of relative infancy compared with the field of tea chemistry, and i t would seem that a large fraction of the task of repeating and reinterpreting the historical results of tea chemistry within the language and context of lhis inslrumenlal meLhod sLil l needs Lo be performed. Progress and innovation in scientific research is frequently the result of dramatic breakthroughs or "revolutions" (Kuhn, 1 970). As each "revolution" propagates itself through a scientific discipline, the paradigms and assumptions that are associated with the discipline must be reevaluated, and the effect of HPLC on tea chemistry needs further investigation in this vein.
B. Bulk Properties and Correlations to Tea Taster's Profiles
Efforts to understand the chemistry of tea can be seen in part as moti vated by the desire of tea plantation owners and manufacturers to understand what factors drive acceptability and market price. Tea imports into the U.S. in 1 993 totaled $ 124 m. (Anon., 1994), a figure that has fluctuated but essentially remained flat for 5 years. Improvements in the quality, marketability, or yield
become strong driving influences for continued business growth and profitability. Improvements in production yield and quality of Indian tea has been attributed to better technological understanding of the underlying factors contro l l ing tea production (Mahanta et aI . , 1993). It is therefore a business necessity as much as an intellectual pursuit to gain a technological advantage, an effort that can only be underpinned by a firm understanding of the chemistry of tea.
Many of the efforts of tea research focus on improving the value of tea by understanding the chemical parameters that determine price. Brisk teas are more astringent and are associated with earlier fermentation times and higher theaflavin content (Owuor and McDowell, 1 994). It fol lows that thearubigens, which are a product of later fermentation times, should be expected to be a negative quality for production of brisk teas. The effect of thearubigens on cream formation (Hazarika et aI., 1 984) can also determine acceptability, especially for iced tea beverages. Principal component analysis has determined that theaflavins are positively correlated (and some flavonol glycosides are negatively correlated) with tea value (McDowell et aI., 1 99 1 ). Theaflavins are very important for quality and price, owing to the bright red color they impart (Hilton and Palmer-Jones, 1975; Ellis and Cloughley, 1 98 1 ), but among similar theaflavin contents, theaflavins are not a good determining factor (Owuor et aI., 1 986). Aroma can also be expected to play an important role, especially i n flowery teas (Robinson and Owuor, 1 992).
C. Tea as an Antioxidant and as a Healthy Beverage
The perception of tea as a healthy beverage is traced to the legendary Shen Nung, who claimed in the Pen Tsao, a Chinese book of herbal medicine, that tea is good for a variety of ailments, including tumors. Tea
467

consumption in ancient times was regarded as healthy, likely due to the fact that the boiling water used to make tea kills many of the water-borne pathogens that caused illnesses common in those times. Vitamin P activity, a reduction of capil lary fragility, has been found in tea and attributed to tea polyphenols (Kursanov et aI., 1 950). Today, research into tea chemistry is a fundamental part of studies on the role of tea as a biological antioxidant and in prevention of chronic diseases.
There is mounting evidence that substances called free radicals play a role in the development of major human diseases such as heart disease and cancer. Research suggests that antioxidants may help protect against these diseases by minimizing the detrimental impact of free radical damage to cells and tissues. Much more research is needed to better identify the processes involved in free radical-mediated diseases and to help develop nutritional strategies to optimize antioxidant status.
The most commonly recognized dietary sources of antioxidants are fruits and vegetables, which contain vitamins C, E, and carotenoids. Preliminary research indicates that the f1avonoids, which include the catechins and f1avonols found i n both black and green tea, also act as antioxidants. However, additional research is required to determine whether tea antioxidants substitute for or complement some of the protective functions identified with the more established antioxidant compounds such as vitamins C, E, and beta-carotene. The role of antioxidants in heart disease is unclear, and their potential role in cancer prevention and the aging process remains speculative.
1. Lipid Oxidation and Cardiovascular Disease
Cardiovascular disease is one of the leading causes of death in the Western World.
468
There is growing evidence that oxidative injury is a major risk factor i n development of cardiovascular disease (Marx, 1 987; Grundy, 1 993). Recent reports that dietary supplementation with vitamin E (at 1 00 IU and less than 400 IU) was associated with a significant reduction in the risk for cardiovascular disease (36 to 54% reduction in risk for women and a 20 to 26% reduction in risk for men) support this hypothesis (Brody, 1 993).
One of the key mechanisms of the hypothesis that oxidative damage is linked to cardiovascular disease is the theory that oxidation of low· density lipoprotein (LDL) leads to damage of the arterial wall, producing sites for fibrous plaque formation. Antioxidant vitamins are believed to inhibit this process, and studies have shown that the antioxidant vitamins ascorbic acid, (X-tocopherol, and �-carotene inhibit LDL oxidation in vitro and reduce the progression of atherosclerosis in animal models.
Studies using a Chinese green tea polyphenol fraction (CGTP) were conducted to determine the effect of tea constituents on oxidation of low-density lipoprotein (LDL) ill vitro in a model system thought to be representative of the effects of antioxidants in modulating risk factors associated with cardiovascular disease (Ding et aI., 1 99 1 ; Ding et aI., 1 992). The antioxidant activity was determined based on lipid peroxide production, thiobarbituric acid reactive substances (TBARS), and the mobility of LDL in an electrophoretic gel. The results of these studies showed that green tea polyphenols block LDL oxidation induced by copper, UV irradiation, or macrophages. The mechanisms for the antioxidant activity of the CGTP have not been determined but are theorized to be metal chelation, free radical scavenging, and an antioxidant "vitamin-sparing" (whereby antioxidant is preferentially consumed, 'sparing' the vitamin) effect. A dose-dependent inhibition of copper-induced LDL oxidation and LDL oxidation induced by transformed

macrophages were also found for catechin (Mangiapane et ai., 1 992).
2. Studies of Tobacco Nitrosamines
Smoking of tobacco products is the main risk factor for development of cardiovascular disease and cancer. The nitrosamines derived from tobacco smoke are good biological oxidants that can damage cellular lipids, LDL lipids, and DNA (Yang, 1 992; Loft et ai., 1 992; Marx, 1 987). One outcome of oxidative damage to DNA is the 8-hydroxylation of guanine bases producing 8-hydroxydeoxyguanosine (80HdG) DNA adducts. Repair of these DNA adducts in vivo leads to the excretion of 80HdG in urine that is a biomarker or index of the current state of oxidative DNA damage and repair (Saul and Ames, 1 986; Loft et ai., 1 992). Tobacco use by men and women has been shown to cause a 50% increase in the excretion of 80HdG, indicating that smoking causes a significant increase in the level of oxidative damage to DNA (Loft et ai., 1 992). Cigarette smoking (> I 0 per day) was found to significantly increase the mutation sensitivity of chromosomes in peripheral lymphocytes (Kim et a!., 1 993). This was determined by treating the cells with mitogens and measuring the frequency of sisterchromatid exchange (SCE). The degree of SCE is a good index of the instability of DNA and of oxidative stress.
A clinical trial was conducted to determine if consumption of green tea or coffee
would alter the sensitivity of DNA to SCE in smokers. Patients were provided three cups of green tea or coffee per day for 6 months. Consumption of three cups of green tea per day inhibited the cigarette smoking-induced increase in frequency of SCE. Coffee consumption did not change the frequency of SCE (Kim et a!., 1 993). This demonstrates that normal tea consumption should be suf-
ficient to block smoke-induced DNA damage due to oxidative events.
A tobacco-specific nitrosamine, 4-(methylnitrosamino)-I (3-pyridyl)-I -butanone, or NNK, is an oxidative carcinogen that induces lung cancer in numerous laboratory animal models (Xu et ai., 1992; Wang et a!., 1 992a). After metabolic activation, NNK reacts with DNA to form oxidized adducts (80HdG). When solutions of green tea, black tea, or the green tea catechin epigallocatechin gallate (EGCG) were provided to mice in place of drinking water the numbers of NNKinduced lung tumors were significantly reduced (Xu et ai., 1 992; Wang et a!., 1 992a). Black and green tea were found to be equally effective in reducing tumor numbers when provided during or after NNK treatment (Wang et a!., I 992a). EGCG was found to be almost as active as green tea infusions (Xu et ai., 1 992). In vivo (Xu et a!., 1 992) studies using mice and in vitro (Shi et ai., 1 993) experiments showed that black and green tea polyphenols inhibit NNK metabolism and DNA adduct formation. EGCG was found to be the most potent inhibitor of the tea polyphenols.
Polyphenols containing 1 ,2- or 1 ,4-diphenol functional groups have been found to be potent inducers of a group of enzymes involved with the metabolism of carcinogens and other xenobiotic compounds, termed phase II enzymes (Sohn et ai., 1 994). Phase II enzymes include NAD(P)H:quinone reductase, glutathione S-transferases, and UDP-glucuronosyl transferases (Talalay, 1992; Prochaska and Talalay, 1 992). Because tea polyphenols fit the criteria for Phase II enzyme inducers, it i s likely that tea polyphenols act as potent inducers of these enzymes, as has been reported (Khan et aI., 1 992). It is likely that one mechanism for the inhibition of NNK-induced lung tumors is blocking of NNK activation. A radical scavenging mechanism might also be involved in the inhibition of NNK-induced DNA adduct
469

formation. The effect of tea on reduction of tumors after NNK treatment indicates that tea can inhibit tumor formation through a second mechanism that does not involve NNK metabolism. Oxidative events have been l inked to the promotion and progression of initiated cells to tumors and cancers. Tea may also act to block these events through an antioxidant mechanism. The promotion of tumor formation in mouse skin by TPA is partially induced by formation of peroxides or free radicals (Wei and Frenkel, 1 993). Tea, GTP (Huang et aI., 1992; Wang et aI., 1 992b), EGCG (Fuj iki et aI., 1 992), and antioxidants (Kozombo et aI., 1983; Smart et aI. , 1 987) were effective in blocking TPA-induced tumor promotion in mouse skin. Mechanistic studies are needed to establish the role of tea in blocking these cellular events.
3. Tea and Cancer
The relationship between consumption of tea and a decreased incidence of cancer has received a significant amount of scientific and media attention during the last few years. The role of dietary constituents (Stavric, 1 994; Wattenberg, 1 992) and tea (Yang and Wang, 1 993) in prevention of cancers has been the subject of numerous scientific papers in recent years. While the emerging research has stimulated a great deal of scientific interest, evidence is still insufficient to allow any clear statements to be made about the relationship between tea consumption and cancer in humans.
VII. CONCLUSION
Understanding the chemistry of tea represents a key technological underpinning after which the systematic study of tea beverages as a healthy addition to the diet can be performed. Achieving mass balance of the
470
polyphenolic constituents of green and black tea is undoubtedly a central focus of tea research. Indirect correlations such as those used to measure thearubigens need to be systematically justified with tangible chemical evidence, whether by isolation of individual compounds as performed by HPLC, or by selective hydrolysis by which the fragments can be measured with quantitative accuracy.
Tea research is moving away from its rudimentary origins as a qualitative outgrowth of tea tasting and the tea industry and moving into a fully quantitative and chemically i nteresting science. The perception of tea not only as a plant and as a beverage but as a healthy part of the human diet and as a rich source of new chemical compounds will propel tea research in new directions.
REFERENCES
Achmadi, S., Syahbirin, G., Choong, E. T., and Hemingway, R. W. 1 994. Phytochemistry 35:
2 1 7-2 1 9 ( 1 994)
Anan, T., 1983. Japan Agric. Res. Quarterly 16:
253-257.
Anan, T., Sakata, K., Yagi, A., and Kato, H. 1987. Agric. Bioi. Chem. 1987: 3395-3397.
Anonymous. 1 994. From The International Tea
Committee, Ltd. , Annual Bulletin of Statistics.
Tea Brokers' Publications, London.
Aruoma, O. I. , Murcia, A., Butler, J" and Halliwell, B. 1993. J. Agric. Food Chem. 41: 1 880-1885.
Bailey, R. G., McDowell, I., and Nurs'en, II. E. 1 990. J. Sci. Food Agric. 52: 509-525.
Bailey, R. G., Nursten, H. E., and McDowell, I. 1 99 1 . J. Chromatog. 542: 1 1 5 .
Bailey, R . G., Nursten, H . E., and McDowell, I . 1992. J. Sci. Food Agric. 59: 365-375.
Bailey, R. G., Nursten, H. E., and McDowell, I. 1994a. J. ehromotog. A. 662: 1 0 1 - 1 1 2 .
Bailey, R . G . , Nursten, H . E . , and McDowell, I . 1994b. J. Sci. Food Agric. 66: 203-208.

Balentine, D. A. 1 992. In: Phenolic Compollnds
in Food and their Effects on Health I, pp.
1 02-107. Ho, c., Lee, C. Y., Huang, M . T.,
Eds., American Chemical Society, Washington, DC.
Beart, J. E., Lilley, T. H., and Haslam, E. 1985.
Phytochem. 24: 33-38.
Bee, R. D., Izzard, M. J., Harbron, R. S., and Stubbs, J. M. 1987. Food Microstructure 6:47-56.
Bhuyan, L. P.and Mahanta, P. K. 1984. l. Sci. Food
Agric. 46: 325-330.
Bigelow, W. c., Pickett, D. L., and Zisman, W. A. 1946. l. Colloid Inteljace Sci. 1: 5 1 3 .
Bohm, F., Edge, R., Land, E. J., McGarvey, D. J., and Truscott, T. G., 1997. lACS 119: 621-622
Bokuchava, M. A. and Popov, V. R. 1948. Dokl,
Akad. Nauk SSSR 60: 6 1 9-622.
Bokuchava, M. A. , Shalamberidze, T. K. , and Sobolcva, G. A. 1970. Dokl. Akad. Nauk SSSR:
192 : 1 374-1375.
Bokuchava, M. A. and Skobeleva, N. I. 1969. Adv.
Food Res. 17: 2 1 5-292.
Bokuchava, M. A .. and Skobeleva, N. I. 1980. CRC Critical Reviews ill Food Science and Nutri
tion. pp. 303-370.
Bokuchava, M. A. and Skobeleva, N. I. 1986. In: Food Flavours: Part B. The Flavour of Bever
ages. pp. 49-84. I. D. Morton and A. J. Macleod, Ed., Elsevier Science Publishers B. V.
Bokuchava, M. A. and Popov, V. R. 1954. Dokl.
Adad. Nauk SSSR 99: 145- 1 5 1 .
Borazan, H . N. and AI-Ani, N. I. 1980. l. Pharm.
Sci. 69: 6 1 3-6 1 5 .
Bradfield, A . E. and Penney, M. J . 1 944. l. Soc. Chen!. Ind. 43: 306--3 1 0.
Britsch, L. 1990. Arch. Biochem. Biophys. 282: 1 52.
Brody, J. E. 1993. The New York Times: May 20.
Brown, A. G., Eyton, W. B. , Holmes, and A., Ollis, W. D. 1969. PhytochemistlY 8: 2333-2340.
Bryce, T., Collier, P. D., Fowlis, I., Thomas, P. E., Frost, D. J., and Wilkins, C. K. 1970. Tet. Lett. 1970: 2789-2792.
Bryce, T., Collier, P. D., Mallows, R., Thomas,
P. E., Frost, D. J., and Wilkins, C. K. Tet. Lett.
1972: 463-466.
Burke, K. E. and Albright, C. H. 1970. l. AOAC 53:
531-533.
Buzun, G. A., Dzemukhadze, K, M., and Mileshko, C. F. 1 974. Subtropical Plants 1 : 48-50.
Cai, Y., Gaffney, S. H., Lilley, T. H .. Magnolato,
D., Martin, R., Spencer, C. M., and Haslam, E. 1 990. l. Chem. Soc. Perkin TrailS. 2 1990: 2 197-2209.
Cannon, C. G. 1955 . Mikrochim. Acta 1955: 555-588.
Cartwright, R. A. and Roberts, E. A. H. 1954. l. Sci.
Food Agric. 5: 593-597.
Cartwright, R. A. and Roberts, E. A. H. 1955. Chent.
Industry 1955: 230-23 1 .
Cartwright, R. A., Roberts, E. A. H., Flood, A. E.,
and Williams, A. H. 1955. Chem. Ind.ustry 1955:
1 062- 1063.
Chenery, E. M. 1955. Plant Soil 6: 1 74-200.
Choudhury, M. N. D. 1982. In: Cultivation and
Utilization of A romatic Plants. pp. 7 1 5-730. Atal, C. K. and Kapur, B. M., Eds., Council of Sci. and Ind. Research, Jammu-Tawi.
Coggon, P., Graham, H. N., and Sanderson, G . W. 1975. British Patent # 1 3 8 0 1 35.
Coggon, P., Moss, G. A., and Sanderson, G. W. 1973. Phytochemistry 12: 1 947-1955.
Collier P. D., and Mallows, R. 1 97 1 . J. Chromatog
raphy 57 : 1 9-27.
Coxon, D. T., Holmes, A., and Ollis, W. D. I 970a. Tet. Lett. 1970: 5241-5246.
Coxon, D. T., Holmes, A. , and Ollis, W. D. 1 970b. Tet. Lett. 1970: 5247-5250.
Coxon, D. T., Holmes, A., Ollis, W. D., Vora, V. C., Grant, M . S., and Tee, J. L. 1972. Tetrahe
dron 28: 2 8 1 9-2826.
Critchlow, A., Haslam, E .. Haworth, R. D., Tinker, P. B., and Waldron, N. M. 1967. Tetrahed. 23: 2829-2847.
Dangles, a., Brouillard, R. 1993. New J. Chen!. 18: 287-296.
471

Danne, A., Petereit, F., and Nahrstedt, A. 1994.
Phytochemistly 37: 533-538.
Das, D. N., Ghosh, J. J., Bhattacharyya, K. c., and
Guha, B. C. 1965. J. Appl. Chem. 28: 1 5-40.
Deijs, W. B., and Dijkman, M. I. 1 936. Arch.
Theecultur. 1936: 1 89.
Ding, Z., Kuhr, S., and Engelhardt, U. H. 1992a. Z Lebellsm. Unlers. Forsch. 195: 108-1 1 I .
Ding, Z.-H .. Chen, Y., Zhou, M., and Fang, Y.-Z. 1 992. Chillese J. Pharm. Toxicol. 6:
263-266.
Ding, Z.-H., Chen, Y., Zhou, M., and Fang, Y.-Z 1 99 1 . Med. Sci. Res. 19: 767-768.
Dix, M. A., Fairley, C. I., Millin, D. I., and Swaine, D. 198 I . J. Sci. Food Agric. 32: 920-932.
Durmishidzern, S . V. and Puridze, G. N. 1980. Soviel Plalll Physiol. 27: 1064-107 1 .
Ebel, I., and Hahlbrock, K. 1982. In: The Flavonoids-Advallces ill Research. pp. 64 1-679. Harborne, I. E. and Mabry, T. I., Eds., Chapman and Hall, London.
Eden, T. 1976. Tea. 3rd ed., Longman Group, London.
Elivin-Lewis, M., Vitali, M., and Kopjas, T. 1980. Prevo Delllistly 6: 273.
Ellis, R. T. and Cloughley, J. B. 198 1 . /111. Tea J. 2:
7-8.
Engelhardt, U. H .. Finger, and A., Kuhr, S. 1993. Z Lebensm. Ulllers. Forsch. 197: 239-244.
Ernst, I. A., Clubb, R. T., Zhou, H. X., Gronenborn, A. M., and Clore, G. M. 1995. Science 267:
1 8 1 3- 1 8 1 7 .
Feldheim, W . , Yongvanit, P., and Cummings, P . H. 1986. J. Sci. Food Agric. 37: 527-534.
Finger, A., Engelhardt, U. H., and Wray, V. 1991a. J. Sci. Food Agric. 55: 3 1 3-32 I .
Finger, A., Engelhard!, U. H., and Wray, V. 1991 b. Phytochem. 30: 2057-2060.
Finger, A. 1994. J. Sci. Food Agric. 66: 293-305.
Finger, A., Kuhr, S., and Engelhardt, U. 1 992. J.
Chromat. 634: 293-3 1 5 .
472
Fraga, C. G., Martino, V. S., Ferraro, G. E., Coussio,
J. D . , and Boveris, A. 1 98 7 . Biochem.
Pharmacal. 36: 7 1 7-720.
Fujiki, H., Suganuma, M., Yoshizawa, S., Yatsunami, I., Nishiwake, S . , Furuya, H., Ok abe, S., Nishiwaki-Matsushima, R., Matsunaga, S., Muto, Y., Okuda, T., and Sugimura, T. 1 992. In: Can
cer Chemoprevellfion. pp. 393-405. Wattenberg,
L., Lipkin, M., Boone, C. W., and Kelloff, G. I., Eds., CRC Press, Boca Raton, FL.
Furuya, T., Orihara, Y., and Tsuda, Y. 1990. Phy
tochemistry 29: 2539-2543.
Gaffney, S. H., Martin, R., Lilley, T. H., Haslam, E., and Magnolato, D. 1986. J. Chem. Soc.
Chem. Comm. 107-109.
Gates, I. C. 1987. Basic Foods, Holt, Rinehart and Winston, Fort Worth, Texas.
Graham, H. 1983. In: The Kirk-Othmer Encyclope
dia of Chemical Technology, 3rd ed. pp. 628-644. Wiley, New York, 22.
Graham, H. 1984. In: The Melhylxalllhille Bever
ages and Foods: Chemistry, Consumption Gnd
Health Effects. pp. 29-74. Alan R. Liss Inc.,
New York.
Gregory, R. P. F., Bendall, D. S. 1966. Bioe/lem. J.
101: 569-58 I .
Grundy, S . M. 1993. Ciin. Cordial. 16 (Suppl. I): 1-3.
Guo, W., Sakata, K., Watanabe, N., Nakajima, R., Yagi, A., Ina, K., and Luo, S. 1 993. Phytochem
istry 33: 1 373-1375.
Guo, W., Sakata, K., Yagi, A., Ina, K., and Luo, S. 1992. Biosci. Biochent. Biotech. 56: 992.
Hagerman, A. E. and Butler, L. G. 198 I. J. Bioi. Cllem. 256: 4494-4497.
IIagerman, A. E.,and Kluchcr. K. M. 1986. In: Progress il/. Clinical and Biological Research (Plant Flavolloids in Biology and Medicine:
Biochemical, Pharmacological, alld Structure
Activity Relationships). pp. 67-76. Alan R. Liss, Inc., New York.
Hahlbrock, K., and Grisebach, H. 1975. In: The Flavonoids. pp. 866-9 1 5 . Harborne, I. B., Mabry, T. I., and Mabry, H., Eds., Chapman
and Hall, London.

Hall, M. N., Robertson, A., and Scotter, C. N. G. 1988. Food Chem. 27: 61-75.
Han, L., Hatano, T., Yoshida, T., and Okuda, T.
1 994. Chem. Pharm. Bull. 42: 1 399-1409.
Harbron , R. S. , Ottewill, R. H., and Bee, R. D. 1989. In: Food Colloids. pp. 283-294. R. D . Bee, Ed.
Hashimoto, F., Nonaka, G., and Nishioka, I. 1987. Chent. Pharm. Bull. 35: 6 1 1-616.
Hashimoto, F., Nonaka, G., and Nishioka, I. 1988. Chem. Phar",. Bull. 36: 1 676-1684.
Hashimoto, F., Nunaka, G., ami Nishiuka, 1. 1989. Chent. Pharm. Bull. 37: 77-85.
Hashimoto, F., Nonaka, G., and Nishioka, I. 1989. Chem. Pilarm. Bull. 37: 3255-3263.
Hashimoto, F., Nonaka, G., and Nishioka, I. 1992. Chem. Pharm. Bull. 40: 1 383-1389.
Haslam, E. 1993. In: Polyphenolic Phenomena. pp. 23-3 1 . Scalbeart, A., Ed., INRA editions, Paris.
Hasselo, H. N. 1965. Tea Quarterly 36: 1 22-1 36.
Hatano, T., Shida, S., Han, L., and Okuda, T. 199 1 .
Chem Pharm. Bull. 39: 876-880.
Hayaishi, 0., and Hashimoto, K. 1950. J. Biochem.
Tokyo 37: 37 1 .
Hazarika, M., Chakravarty, S. K., and Mahanta, P. K. 1 984. J. Sci. Food Agric. 35: 1 208- 1 2 1 8
Heller, W . , Egin-Buhlcr, B . , Gardiner, S . E . , Knobloch, K.-H., Matern, U., Ebel, J., and Hahlbrock, K. 1979. Plant Physiol. 64: 37 1 .
Heller, W., and Forkmann, G. 1988. In: The Fla
vonoids: Advances in Research Since 1980. pp. 399-422. Harborne, 1. B., Ed., Chapman and Hall, London.
Heller, W., and Forkmann, G. 1994. In: The Fla
vonoids: Advances in Research Since 1986. pp. 499-536. Harborne, J. B., Ed., Chapman and Hall, London.
Hertog, M. G. L., Hollman, P. C. H. , and van de Putte, B. 1993. J. Agric. Food Chem. 41: 1 242-1 246.
Hicks, M. B.; Hsieh, Y-H.; Bell, L. N.; 1996. Food.
Res. Int. 29: 325-330.
Hilton, P. J. 1973. In: Encyclopedia of Industrial
Chemical Analysis. 18: 455-5 1 8. John Wiley, New York.
Hilton, P. J. and Palmer-Jones, R. W. 1975. J. Sci.
Food Agric. 26: 1 68 1 - 1 687.
Ho, C. T. 1992. In: Phenolic Compounds in Food
and Their Effects on Health I. pp. 2-7. Ho, C. T., Lee, C. Y., and Huang, M . T., Eds.,
American Chemical Society, Washington, DC.
Ho, C. T., Chen, Q., Shi, H., and Rosen, R. T. 1992. Prevo Med. 2 1 : 520-525.
Hoefler, A. C. and Coggon, P. 1976. J. Chromo 129:
460-463.
Horner, L., Weber, and K. H., Durckheimer, W. 1 96 1 . Chenl. Ber. 94: 2 8 8 1 -2887.
Huang, M . T., Ho, C.T., Wang, Z. Y., Ferraro, T., Finnegan-Olive, T., Lou, Y.-R., Mitchell, J.
M., Laskin, J. D., Newmark, H., Yang, C. S., and Conney, A . H . 1992. Carcinogenesis 13:
947-954.
Imagawa, H., Takino, Y., and Shimizu, M. 1976. Nippon Shokuhin Kogyo Gakkashi 23: 1 38-144.
Imagawa, H., Toryu, H., Ozawa, and T., Takino, Y. 1 982. Agric. Bioi. Chem. 46: 1 2 6 1 - 1 269.
Imagawa, 0., Yamano, H., Inoue, K., and Takino, Y. 1979. Agric. Bioi. Chent. 43: 2337-2342.
Interesse, F. S., Ruggiero, P. , D' Avella, G., and Lamparelli, F. 1983. Phytochemistry 22: 1 885-1 889.
Iwasa, K. 1976. Japan Agric. Res. Quarterly 10:
89-93.
Jain, 1. C. and Takco, T. 1984. Food Bioehem. 8: 243-279.
layman, T. C. Z. and Sivasubramanian, S. 1975. J. Sci. Food Agric. 26: 1 895.
Jiang, Y. and Miles, P. W. 1993. Phytochemistry
33: 29-34.
Jovanovic, S . V " Steenken, S . , Tosie, M . , Marjanovic, B., and Simic, M . G . , 1994. JACS 116: 4846-485 I .
Jovanovic, S . Y., Hara, Y., Steenken, S., and Simic. M. G . , 1995. JA CS 117: 9881-9888.
473

Kalita, J. N. and Mahanta, P. K. 1993. 1. Sci. Food Agric. 62: 1 03-109.
Kamsteeg, J., van Brederode, J., Verschuren, P. M.,
and van Nigtevecht, G. 1 9 8 1 .Z Pjlallzellphysiol.
102: 435.
Kato, C., Uritani, I., Saijo, R., and Takeo, T. 1976. Plallt alld Cell Physio/. 17: 1 045-1052.
Kawagishi, H. and Sugiyama, K. 1992. Biosci.
Biochern. Biotech. 56: 689.
Khan, S. G., Katiyar, L. K. , Agarwal, R., and Mukhtar, H. 1 992. Callcer Res. 52: 4050-4052.
Kharebava, L. G. 1986. Subtropical Plants 4: 162-1 63 .
Kim, Y. H., Shim, J . S . , Kang, M . H., Roh, J . K., Roberts, C., and Lee, I. P. 1 993. Proc. Americall Assoc. Cancer Res. 34: Abstr. #3309.
Kimura, R. and Murata. T. 1 97 1 . Chem. Pharm.
Bull. 19: 1257-126 1 .
Kito, M., Kokura, H., !zalU, J., and Sasoka, K. 1 968. Phytochem 7: 599-603.
Konishi, S. and Takahashi, E. 1 969. Nippoll Do)o
Hiryogaku 20sslii 40: 479-484.
Korver, 0., Wilkins, C. K., and Collier, P. 1 973.
Chem. Illdustry 1973: 89-90.
Kozombo, W. J., Seed, J. L., and Kensler, T. W. 1 983. Callcer Res. 43: 2555-2559.
Kuhn, T. S. 1 970. The Structure of Scientific Revo
lutiollS, 2nd ed. University of Chicago Press,
Chicago.
Kuhr, S., Herzig, B., and Engelhardt, U. H. 1994. Z Lebensm. Ullters. Forsch. 199: 1 3- 1 6.
Kursanov, A. L" Bukin, V. N., Povolotskaya, and K. P., Zaprometov, M. N. 1 950. Biokhim. Cliain. Proizvod. 6: 170.
Kursanov, A. L. 1956. Kulturpf/allze Beilieft 1: 29-48.
Liang, Y. R., Liu, Z. S., Xu, Y. R" and Hu, Y. L. 1990. 1. Sci. Food Agric. 53: 54 1 -548.
Liao, H., Cai, Y., and Haslam, E. 1992. 1. Sci. Food
Agric. 59: 299-305.
Lin, Y. Y., Ng, K. J., and Yang, S. 1993. 1. Chrolllatogr. 629: 389-393.
474
Loft, S . , Vistisen, K., Ewerts, M., Tjonneland, A.,
Overvard, K. , and Poulsen, H. E. 1992. Carcinogenesis 13: 2241 -2247.
Lowenstein, J. M. 198 1 . Methods Ellzymol. 71: 5.
Luck, G., Liao, H., Murray, N. J., Gimmer, H. R., WarminslU, E. E., Williamson, M. P., Lilley, T. H., and Haslam, E. 1994. Phytochemistry 37:
357-37 1 .
Lunder, T. L. 1992. In, Phenolic Compounds in
Food alld their Effects of Health II. pp. 1 14-120. Huang, M. T., Ho, C. T., and Lee, C. Y.,
Eds., American Chemical Society, Washington, DC.
Mahanta, P. K., and Baruah, S. 1 992. 1. Sci. Food
Agric. 59: 2 1 -26.
Mahanta, P. K. Boruah, S. K., Boruah, H. K., and
Kalita, J. N. 1993. 1. Agric. Food Chelll. 41:
272-276.
Makhatadze, G. I. and Privalov, P. L. 1 994. Biophys.
Chern. 50: 285-29 1 .
Mangiapane, H., Thompson, J., Saiter, A., Brown, S., Bell, G. D., and White, D. A. 1992. Biochem.
Pharmaco/. 43: 445-450.
Martin, R., Cai, Y., Spencer, C. M., Lilley, T. H., and Haslam, E. 1990. Bulletin de Liason Groupe
PolyplJenols 15: 304-3 1 8 .
Martin, R., Lilley, T . H . , Bailey, N . A . , Falshaw, C. P., Haslam, E., Magnolato, D., and Begley, M. J. 1 986. 1. Chem. Soc" Chenl. Commun.
1986: 105-106.
Marx, J. L. 1987. Science 235: 529-5 3 1 .
Mathias, J . P., Seto, C . T., and Whitesides, G . M. 1993. Polymer Preprints 34: 92-93.
Matsui, K., Toyota. H.t Kajiwara, T., Kakuno, T., and Hatanaka, A. 199 1 . Phytochemistry 30:
2 1 09-2 1 13.
Matsuura, T. and Kakuda, T. 1 990. Agric. Bioi.
Chem. 54: 2283-2286.
Matsuura, T., Kakuda, T., Kinoshita, T., Takeuchi, N., and Sasaki, K. 1992. Biosci. Biocllelll.
Biotech. 56: 1 1 79- 1 1 8 1 .
Matsuzake, T . and Hara, Y . 1 9 8 5 . Nippon
Nogeikagaku Kaishi 59: 1 29-134.
Mayer, A. M., and Harel, E. 1 99 1 . In: Food Enzy
lIIology. Vol. I . pp. 373-398. Fox, P. F., Ed., Elsevier, London.

Mayer, A. M. 1987. Phytochemistry 26: 1 1 -20.
Meyer, A., Ngiruwonsanga, T., and Henze, G. 1996.
Fresenius J. Ana/. Chem. 356: 284-287
McDowell, I., Bailey, R. G., and Howard, G. 1990.
J. Sci. Food Agric. 53: 4 1 1-414.
McDowell, I., Feakes, J., and Gay, C. 1 99 1 . J. Sci.
Food Agric. 55: 627-64 1.
McManus, J. P., Davis, K. G., Haslam, E., and Lilley, T. H. 198 1 . Chem. Soc. Chem. Camlll.
1981: 309.
Michl, H., and Haberler, F. 1954. Monatsh. Chem. 85: 779-795.
Millin, D. J. and Rustige, D. W. 1967. Process Biocllem. 2: 9- 13.
Millin, D. J. Swaine, D., and Dix, P. L. 1969. J. Sci.
Food Agric. 20: 297-302.
Mistry, T. Y., Cai, Y., Lilley, T. H., and Haslam, E. 1 99 1 . J. Chem. Soc. Perkin Trans. 2, 1991:
1287-1296.
Mizuno, T., Suzuki, Y., Abe, H., and Kampyo, T.
1 964. Nippon Shokuhin Kogyo Gakkeishi 11:
146-152.
Morel, 1., Lescoat, G., Cogrel , P., Sergent, 0., Pasdeloup, N., Brissot, P., Cillard, P., and Cillard, J. 1993. Biochem. Pharmacol 45: 1 3-1 9.
Morita, K . , Wakabayashi, M . , Kubota, K . , Kobayashi, A., and Herath, N . L . 1 994. Biosci.
Biochem. Biotech. 58: 687-690.
Nagalakshmi, S., Jayalaksmi, R., and Seshadri, R. 1 985. J. Food Sci. Tech. 22: 1 9 8-20 I .
Nagalakshmi, S., Ramaswamy, M . S., Nataraj an, C. P., and Seshadri, R. 1984. Food. Chem. 13:
69-77.
Nagalakshrni, S. and Seshadri, R. 1983. J. Fuud Sci. Tech. 20: 243-245.
Nagata, T., Hayatsu, M., and Kosuge, N. 199 1 . Phytochemistry 31: 1 2 1 5 .
Nagata, T . , Hayatsu, M., and Kosuge, N . 1993. Phytochem. 32: 7 7 1 -775.
Nakayama, T., Yamada, M., Osawa, T., and Kawakishi, S. 1993. Biochem. Pahrmacol. 45: 265-267.
Namiki, M. 1990. CRC Crit. Rev. Food Sci. Nutr.
29: 273-300.
Namiki, M. and Ozake, M. 1986. Basic Life Sci. 39:
1 3 1-142.
Negishi, 0., Ozawa, T., and [magawa, H. 1985a. Agric. Bio!. Chem. 49: 25 1 -253.
Negishi, 0., Ozawa, T., and [magawa, H. 1985.
Agric. Bio!. Chenl. 49: 887-890.
Negishi, 0., Ozawa, T., and [magawa, H. 1 988. Agric. Bioi. Chen!. 52: 169-175.
Negishi, 0., Ozawa, T., and [magawa, H. 1992. Biosci. Biotech. Biochem. 56: 499-503.
Neish, A. C. 1 964. In : Biochemistry of Phenolic
Compounds. pp. 295-360. Harborne, J. B., Ed., Academic Press. London.
Newcomb, L. F. and Gellman, S. H. 1994. JA CS
116: 4993-4994.
Nikolaeva, T. N . , Bagrati shvili , D . G., and Zaprometov, M. N. 1982. Soviet Plallt Physiol.
29: 930-933 .
Nikolaeva, T . N. and Zaprometov, M. N . 1990. Soviet Plant Physiol. 37: 286--292.
Nishijo, J., Yonetani, L, Iwamoto, E., Tokura, S., Tagahara, K .. and Sugiura, M. 1990.J. Pharm.
Sci. 79: 14-18.
Nonaka, G., Goto, Y., Kinjo, J. , Nohara, T., and Nishioka, I . 1987. Chem. Pharm. Bull. 35:
1 105.
Nonaka, G., Hashimoto, F., and Nishioka, I. 1986. Chem. Pharm. Bull. 34: 6 1-<i5.
Nonaka, G., Kawahara, 0., and Nishioka, I. 1983. Chenl. Pharm. Bull. 31: 3906-39 14.
Nonaka, G., Sakai, R. , and Nishioka, I. 1984. Phytochemistry 23: 1753-1755.
Oh, H. I., Hoff, J. E., and Haff, L. A., 1985. J. Food
Sci. 50: 1 652-1654.
Oh. I. and Hoff, J. E. 1987. J. Food Sci. 52: 1267-1272.
Okuda, T., Yoshida, T., and Hatano, T. 1988. J. Liq.
Chromo 1 1 : 2447-2454.
Opie, S. c., Clifford, M. N., and Robertson, A. 1 993. J. Sci. Food Agric. 63: 435-438.
475

Opie, S . c., Cl ifford, M. N., and Robertson, A . 1995. J. Sci. Food Agric. 67: 501-505.
Opie, S. C. 1992. PhD. Thesis, School of Biological Sciences, University of Surrey.
Opie, S. C. , Robertson, A. , and Clifford, M. N.
1990. J. Sci. Food Agric. 50: 547-561.
Osawa, T., Ramarathnam, N., Kawakishi, and S., Namiki, M. 1 992. In: Phenolic Compounds ill
Food and Their Effects 011 Health II: Antioxi
dants and Cancer Prevention. pp. 122-149. Huang, M. -T., Ho, C. -T., and Lee, C. Y., Eds., American Chemical Society, Washington, DC.
Owuor, P. O. and McDowell, I . 1 994. Food. Chenl. 51: 25 1 -254.
Owuor, P. O., Reeves, S. F., and Wanyoko, 1. K. 1986. J. Sci. Food Agric. 37: 507-5 1 3.
Owuor, P. o. Tsushida, T., Horita, and H., Murai, T. 1987. Trop. Sci. 27: 159-166.
Ozawa, T., Lilley, T. H., and Haslam, E. 1987. Phytochem. 26: 2937-2942.
Perera, K. P. W. C. and Wickremasinghe, R. 1.. 1 972. Tea Q. 43: 153-163.
Pethiyagoda, U. and Krishnapillai, S. 1 970. Tea
Quarterly 41: 107-120.
Piretti, M. V. and Doghieri, R. 1990. 1. Chromatog
raphy 314: 334-342.
Pokorny, J. 1992. Trends Food Sci. Tech. 2: 223-227.
Popov, V. R. 1956. Biokhimiya 21: 380-384.
Powell, C., Clifford, M. N., Opie, S. c., Ford, M. A., Robertson, A., and Gibson, C. 1.. 1993. J.
Sci. Food Agrie. 63: 77-86.
Powell, C. Clifford, M. N., Opie, S . c., and Gibson, C. L. 1994. From What's in a Cuppa ? Recem Advances ill the Chemistry and Biochemistry
a/Tea. Sci. Conference Papers Series, Londoll,
No. 0027, 1 8 pp.
Pratt, E. E. 1992. In: Phenolic Compounds in Food alld Their Effects Oil Health II: Antioxidants
and Cancer Prevention. pp. 1 1 50-1 1 59. Huang, M.-T., Ho, C. -T., and Lee, C. Y., Eds., American Chemical Society, Washington, DC.
Prochaska, H. 1. and Talalay, P. 1992. In: Phellolic Compounds in Food and their Effects 011 Health
476
II. pp. 1 150-1 1 59. Huang, M.-T., Ho, c. -T.,
and Lee, C. Y., Eds., American Chemical Society, Washington, DC.
Pruidze, G. N. 1975. Fermenty 1975: 25-43.
Pruidze, G. N. and Grigorashvili, G. Z. 1975. Soobshch. Akad. Nauk Curz. SSR 79: 1 85-1 88.
Putman, l.. 1. and Butler, 1.. G. 1989. J. Agric. Food
Chem. 37: 943-946.
Ramakrishna, R. S. , Palmakumbura, S., and Chau, A. 1987. J. Sci. Food. Agric. 28: 33 1-339.
Ratty, A. K. and Das, N. P. 1988. Bioehem. Med. Metab. Bioi. 39: 69-79.
Roberts, E. A. H. 1942. Adv. Ellzymology 1 13-133.
Roberts, E. A. H. 194 I . Bioehem. J. 35: 909-9 19.
Roberts, E. A. H., Cartwright, R. A., and Oldschool, M. 1957. J. Sci. Food Agric. 8: 72-80.
Roberts, E. A. H., Cartwright, R. A., and Wood, D. J. 1 956. J. Sci. Food Agric. 4: 253-257.
Roberts, E. A. H. 1962. In: The Chemistry of Fla
vonoid Substances. pp. 468-5 1 2. Geissmann, T. A., Ed., Pergamon, Oxford.
Roberts, E. A. H. 1958. J. Sci. Food. Agrie. 9: 2 1 2-223.
Roberts, E. A. H. 1 963. J. Sci. Food Agric. 14: 700-
705.
Roberts, E. A. H. and Myers, M. 1959. J. Sci. Food Agric. 10: 172-176.
Roberts, E. A . H. and Smith, R. F. 1963. 1. Sci.
Food Agric. 14: 689-700.
Roberts, E. A. H., Wight, W. and Wood, D. J. 1958. New Phytologist 57: 2 1 1-225.
Roberts, E. A. H. and Wood, D. 1. 195 I . Biochem. 1. 49: 41 4-422.
Roberts, G. R. and Sanderson, G. W. 1966. J. Sci. Food Agrie. 17: 1 82-188.
Robertson, A. and Bendall, D . S . 1983. Phytochem
iSHy 22: 883-887.
Robertson, A. and Hall, M. N. 1989. Food ChenL
34: 57-70.
Robertson, A. 1992. In: Tea: Cultivation to COIl
slllllptioll. pp. 555-601 . Wilson, K . C. and Clifford, M. N., Eds., Chapman Hall, London.

Rubertsun, A. 1983. Phytochemistry 22: 889-896, 897-903.
Robinson, J. M. and Owuor, P. O. 1992. In : Tea:
Cultivatioll and Consumptioll . pp. 603-647.
Wilson, K. C. and Clifford, M. N., Chapman and Hall, London.
Ruch, R. J., Cheng, S . -J . , and Klaunig, J. E. 1989. Carcinogenesis 10: 1003-1008.
Rutter, P. 1 97 1 . Ph. D. Thesis, Procter Department of Food and Lealher Science, University of Leeds, Leeds.
Rutter, P. and Stainsby, G. 1975. Sci. Food Agric. 26: 455-463.
Rzepa, H. S., Smith, M. H., and Webb, M. L. 1994. J. Chen!. Soc. Perkin Trans. 2 1994: 703-707.
Saijo, R. 1982. Agric. BioI. Chem. 46: 1 969-1970.
Saijo, R. 1983. Agric. Bioi. Chenl. 47: 455--460.
Saijo, R. 1980. Plant Cell Physiol. 21: 989-998.
Saijo, R. and Takeo, T. 1979. Agric. BioI. Chern.
43: 1 427-1432.
Sakata, K., Yamauchi, H., Yagi, A., and Ina, K. 1987. Agric. Bioi. Chem. 51: 1 7 3 1-1739.
Sakata, K., Yamauchi, H., Vagi, A. , Ina, K., Parkanyi, L., and Clardy, J. 1989. Agric. BioI.
Chern. 53: 2975-2979.
Sakata, Y. 1950. Agric. Cile",. Soc. Japall 23: 262-267.
Sanderson, G. W., Berkowitz, J. E., Co, H., and
Graham, H. N. 1972. J. Food Sci. 37: 399--404.
Sanderson, G. W. 1972. In: Structllral and Func
tional Aspects of Phytochemistry. pp. 247-3 16. Academic Press, Inc., New York.
Sanderson, G. W. and Perera, B . P. M. 1965. Tea
Quart. 36: 6-13.
Sanderson, G. W., Ranadive, A. S., Eisenberg, L. S., Farrell, F. J., Simons, R., Manley, C. H., and Coggon, P. 1976. In: Sulfur alld Nitrogen
Compoullds in Food Flavors. 26. pp. 14-16. Charalambous, G. and Katz, I., Eds., American Chemical Society, Washington, DC.
Sanderson, G. W. and Selvendran, R. R. 1965. J.
Sci. Food Agric. 16: 25 1-258.
Sasaoka, K. 1965. J. Agric. Chel1l. Soc. Japan 39:
R I-R6.
Sasaoka, K. , Kito, M., Konishi, S. , and Inagaki, H. 1962. Agric. BioI. Chetn. 26: 265.
Sasaoka, K., Kito, M. and Onishi, Y. 1 965. Agric.
Bioi. Chem. 29: 984--988.
Saul, R. L. and Ames, B . N. 1986. Basic Life Sci.
28: 529-535.
Sears, K. D., Casebier, R. L., Hergert, H. L., Stout, G. H., and McCandllish, L. E. 1974. J. Org.
ehe",. 39: 3244-3247.
Sekine, T., Arai, Y., Ikegami, F., Fujii, Y., Shino, S., Yanagasawa, T., Ishida, Y., Okonogi, S., and Murakoshi, I. 1993. Chenl. Phartn. Bull. 41: 1 1 85-1 1 87.
Sekine, T., Alita, J., Yamaguchi, A., Saito, K., Okonogi, S., Morisaki, N., Iwasaki, S" and Murakoshi,
I. 199 1 . Phytochemistry 30: 991-995.
Sekiya, J., Kajiwara, T., Monma, T., and Hatanaka, A. 1984. Agric. BioI Chem. 48: 1963-1967.
Selvendran, R. R., Perera, B. P. M., and Selvendran, S. 1972. J. Sci. Food Agric. 23: 1 1 19-1 1 23 .
Seshadri, R . and Dhanaraj, N . 1988. J. Sci. Food
Agric. 45: 79-86.
Shao, W., Powell, c., and Clifford, M. N. 1995. J. Sci. Food. Agric. 69: 535-540
Shestopa, A. V., Danilov, V. I., and Maleev, V. J.
1 985. Dokl. Akad. Nauk. SSSR 282: IOOG--I 003
Shi, S. T., Wang, Z. Y., Smith, T. J., Cheng, W. F., Ho, C. T., and Yang, C. S. 1993. Proc. Amer.
Assoc. Cancer Res. 34: abstract #935.
Shi, X. and Dalal, N. S. 1 99 1 . Fd. Chenl. Toxico!.
29: 1-6.
Shiplova, S. V. and Zaprometov, M. N. 1977. Soviet Plant Physiol. 26: 657-662.
Singh, H. P. and Ravindranath, S. D. 1 994. J. Sci.
Food Agric. 64: 1 17- 1 20.
Skoboleva, N.!. and Popov, V. R. 1962. Biokhim.
Chaill. Proizvod. 9: 1 85-189.
Smart, R. c., Huang, M.-T., Han, Z. T., Kaplan, M. c., Focella, A., and Canney, A. H. 1987. Callcer Res. 47: 6633-6638.
Smith, R. F. 1968. J. Sci. Food Agr. 19: 53G--534.
Soh", O. S., Surace, A., Fiala, e. S., Richie, J. P. Jr., Colosimo, S., Zans, E., and Weisburger, J. H.
1994. Xenobiotica 24: 1 1 9-127.
477

Speier, G., Tyeklar, Z., Szabo, L., Toth, P., Pierpont, C. G., and Hendrickson, D. N. 1 993. From:
The Activatioll of Dioxygen and Homogeneous
Catalytic Oxidation. Barton, D . H. R., Ed., Plenum Press, NY.
Spencer, C. M., Cai, Y., Marlin, R, Gaffney, S. H., Goulding, P. N., Magnolato, D., Lilley, T. H .. and Haslam, E. 1988. Phytochemistry 27: 2397-2409.
Spiro, M. and Jaganyi, D. 1994. Food Chem. 49: 35 1-357, 359-365.
Spiro, M., Price, W. E., Miller, W. M., and Arami, M. 1 987. Food Chem. 25: 1 1 7-126.
Sprinson, D. B., Advan. 1960. Carbohydrate Chem.
15: 235.
Stafford, H. A. 1988. In: Chemistry and Signifi
cance of Condensed Tannins, Hemingway, R W. and Karchesy, J. 1., Eds., Plenum, New York, 301.
Stagg, G. V. and Swaine, D. 1 97 1 . Phytochemistry 10: 1 67 1 -1673.
Stahl, W. H. 1962. Adv. Food. Res. 1 1 : 201-262.
Stavric, B. 1994. Ciin. Binchem. 27: 3 1 9-332.
Steen ken, S. and Jovanovic, S. V. 1997. lA CS 119:
6 1 7-6 1 8.
Steffens, 1. c., Harel, E., and Hunt, M. D. 1994. Recent Advances in Phytochem. 28: 275-3 1 2 .
Steinhaus, B . , Engelhardt, U . H., and Lebensm, Z. 1989. Ullters. Forseh. 188: 509-5 I I .
Strekova, V . Y., Subbotina, G. A., Zagoskina, N. V., and Zaprometov, M. N. 1980. Soviet Plallt
Physiol. 27: 889-896.
Sun, J.-S. and Mallice, W. L. 1996. Polymer Bull. 37: 69 1-698.
Suzuki, T. and Takahashi, E. 1 975. Biochem. l. 146: 87-96.
Suzuki, T. and Takahashi, E. 1 976. Phytochemisfly
15: 1235- 1 240.
Takeo, T. 1965. Agric. Bioi. Chem. 29: 558-563.
Takeo, T. 1984. Agrie. Bioi. Chenl. 48: 1083-1 085.
478
Takeo, T. 1 974. Phytochemistry 13: 140 I - I 406.
Takeo, T. and Uritani, L. 1966. Agric. BioI. Chen!.
30: 1 55- I 63.
Takino, Y. Ferrelli, A., Flanagan, V., Giantufco, M. A. , and Vogel, M. 1967. Can. l. Chem. 45: 1 949-1956.
Takino, Y. and Imagawa, H. I 964a. Agric. BioI.
Chelll. 28: 1 25- I 30.
Takino, Y. and Imagawa, H. I 964b. Agric. Bioi.
Chem. 28: 255-256.
Takino, Y. and Imagawa, H. 1963. Agric. BioI. Chem. 27: 666--668.
Takino, Y., Imagawa, H., Horikawa, H., and Tanaka, A. 1 964. Agric. BioI. Chem. 28: 64-7 1 .
Takino, Y., Imagawa, H., and Shishido, K . 1972. Nippon Shokuhill Kogyo Gakkaishi 19: 2 I 3-2 1 8.
Talalay, P. 1992. In: Cancer Chemopreventioll, Wallenberg, L" Lipkin, M., Boone, C. W., Kelloff, O. K., Eds., CRC Press, Boca Raton, pp. 469-478.
Tanizawa, H. , Sazuka, Y., and Komatsu, A. 1983. Chenl. Pharo Bull. 31: 4 1 39-4143.
Tanizawa, H" Toda, S., Sazuka, Y., Taniyama, T.,
Hayashi, T., Arichi, S., and Takino, Y. 1984. Chelll. Pharm. Bull. 32: 201 1-20 14.
Taylor, S . J. and McDowell, I. J. 199 I . l. Sci. Food
Agric. 57, 287-29 1 .
Taylor, S" Baker, D., Owuor, P., Orchard, 1., Othieno, c., and Gay, C. 1 992. l. Sci Food.
Agrie. 58: 1 85- 1 9 1 .
Thakkar, A . L., Tensmeyer, I.G., Hermann, R . B., and Wilham, W. L. 1970. l. Chem. Soc. Chelll. Comm. 1970: 524-525.
Thomas, R L. and Murtagh, K. J. 1985. l. Food Sci.
50: 1 1 26-1 1 29.
Tolbert, N. E. 1973. Plant Physiol. 5 1 : 234-244.
Tolhurst, J. A. H. 1962. Tea Quarterly 33: 1 34-1 37.
Tsushida, T. 1987. Kagakuto Seibusto, 25: 699-70 1 .

Tsushida, T. and Murai, T. 1987. Agric. Bioi. Chem.
51: 2865-287 1.
Tsushida, T. and Takeo, T. 1985. Agric. BioI. Chem.
49: 3 1 9-326.
Uchida, S., Edamatsu, R. Hiramatsu, M., Mori, A., Nonaka, G., Nishiokta, I., Niwa, M., and Ozaki, M . 1 987. Med. Sci. Res. 15: 83 1-832.
Uchida, S., Ozake, M., Suzuki, K., and Shikita, M . 1992. Life Sci. 50: 147-152.
Ullah, M. R. 1972. Curl'. Sci. 41: 422-423.
Vamos-Vigyazo, L. 1 9 8 1 . CRC Crit. Rev. Food Sci. Nurr. 15: 49-127.
Van Sumere, C. F., Albrecht, J., Dedonder, A . , De Pooter, H . , and Pe, I. 1975. from The Chem
istry and Biochemistry or Plant Phenolics,
Harborne, J. B. and Van Sumere, C. F., Eds., Academic Press, London, pp. 2 1 1-264.
Vuataz, 1.., Brandenberger, H., and Egli, R. H. 1959.
l. Chromarogr. 2: 1 73-187.
Vuataz, I.. and Brandenberger, H. 1 96 1 . l. Chromatogr. 5: 17-3 1 .
Wang, C . and Huang, Y. 1987. Zhongguo Kexue lishu Doxue Xuebao 17: 469-474.
Wang, P.-F. and Zheng, R.-L. 1992. Chemistry and
Physics of Lipids 63: 37-40.
Wang, Z.-Y., Hong, J.-Y., Huang, M-T., Reuhl, K. R., Conney, A. H., and Yang, C. S. 1992. Can
cer Res. 52: 1943-1947.
Wang, Z.-Y., Huang, M.-T., Ferraro, T., Wong, C.Q., Lou, Y.-R., Reuhl, K., Itropoulos, M., Yang, C. S . and Conney, A. H. 1992. Cancer Res. 52: 1 162-1 1 70.
Wattenberg, I.. W. 1992. In: Cancer Chemoprevention, Wattenberg, L.t Lipkin, M., Boone, C. W., Kelloff, G. J. Eds. CRC Press, Boca Raton, FL, 19: 39.
Wedzicha, B. I.. and Donovan, T. 1. 1 9 89. l. Chron/mogr., 478: 2 1 7-224.
Wedzicha, B. L., La, M. F., and Donovan, T. J. 1990. l. Chromatogr., 505: 357-364.
Wei, H. and Frenkel, K. 1993. Carcinogenesis 14:
Wellum, D. A. and Kirby, W. 198 1 . l. Chromarogr.
206: 400.
Whilaker, J. R. 1994. Food Sci. Tech., 61: 543-556.
Whitehead, D. I.. and Temple, C. M. J. 1 992. Sci.
Food Agric., 58: 1 49- 1 52.
Wickremasinghe, R. I.. 1978. In: Advances ill Food,
Research, Chichester, C. 0., Ed., Academic Press, New York, 24: 229-286.
Wickremasinghe, R. 1.., Roberts, G. R., and Perera, K. P. W. C. 1967. Tea Quarterly 38: 309-3 10.
Williamson, M. P . , Trevitt, C. ami Nohle, J. M. 1995. Carbohyd. Res. 266: 229-235.
Wilson, K. C. and Clifford, M. N. 1992., Eds., Tea:
Cultivation to Consumption, Champ man and Hall, London.
Xianqi, S. B. H. and Haslam, E., 1994. l. Assoc. Leather Chen!. Assoc., 89: 98-104.
Xiao, W. Q. and Li, C. 1992. China Tea. , 1 : 24-26.
Xu, Y., Ho, C.-T., Admin, S. G. Han, c., and Chung, F. I.. 1992. Cancer Res., 52: 3875-3879.
Yamada, H. and Hattori, T. 1977. l. Sci. Soil Mature lpn., 48: 253.
Yamada, H. and Hallori, T. 1980. lap. l. Soil Sci.
Plant Nutr. 51: 3 6 1 .
Yang, C . S . and Wang, Z.-Y. 1993. l. Natl. Cancer IlIst. 85: 1038-1049.
Yang, F.-J., Zhao, B.-I.., and Xin, W.-J. 1992. Res.
Chetll. Intermed. 17: 39-57.
Yokogoshi, H., Kala, Y., Sagesaka, Y. M., Matsuura, T., Kakuda, T., and Takeuchi, N. 1995. Biosci. Biotech. Biochem. 59: 6 1 5-6 18.
Yoshida, T., Chou, T.. Maruyama, Y., and Okuda, T. 1 990. Chent. Pharn/. Bull. 38: 268 1 -
2686.
Zagoskina, N. V., Usik, T. Y., and Zaprometov, M. N. 1990. Soviet Plant Physiol. 37: 388-
393.
Zagoskina, N. V. and Zaprometov, M. N., 1976. In: Abstracts alld Papers Presented at the Third
Ali- Unioll Symposium on Phenolic Compounds,
Metsnierba, Tbilisi, 21.
479

Zaprometov, M. N. 196 1 . Biokhimislry 26: 373-384.
Zaprometov, M. N. and Bukhlaeva, V. Y. 1 97 1 . Plant Physiol. 18: 787-795.
Zaprometov, M. N. and Bukhlaeva, V. Y. 1963. Biokhim. 28: 862-867.
Zaprometov, M. N. 1987. Soviet Plall! Physiol. 34: 5 6 1 -572.
480
Zaprometav, M. N. 1 962. Diokhimistry 27: 366-367. Zawistowski, J., Biliaderis, C. G., and Eskin,
N. A. M. 199 1 . Oxid. Enzymes and Foods,
Robinson, D. S . et aI., Ed., Elsevier, London, pp. 2 1 7-273.
Zhao, B. L., Liu, S. L., Chen, S., and Xin, W.-J. 1 992. Acta Pharmacal. Sinica 13: 9-1 3 .
Zhao, B., Li, X., He, R., Chen, S., and Xin, W.-J. 1989. Gel/ Biop"ys. 14: 1 75-1 85.




















