
Craniometrical variability in the cave bears (Carnivora, Ursidae): Multivariate comparative analysis
Upload
cadic-conicetCategory
view
3download
0
Taxonomic status of southern South AmericanConepatus (Carnivora: Mephitidae)
MAURO I. SCHIAFFINI1,2*, MAGALÍ GABRIELLI1,3, FRANCISCO J. PREVOSTI1,4,YAMILA P. CARDOSO1,3, DIEGO CASTILLO5, ROBERTO BO6, EMMA CASANAVE5 andMARTA LIZARRALDE1,3
1Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina2Laboratorio de Investigaciones en Evolución y Biodiversidad, Facultad de Ciencias Naturales,Universidad Nacional de la Patagonia San Juan Bosco, sede Esquel, Sarmiento 849, CP 9200Esquel, Argentina3Centro Regional de Estudios Genómicos, Universidad Nacional de La Plata, La Plata, Argentina4División Mastozoología, Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’, Av. AngelGallardo 470, C1405DJR Buenos Aires, Argentina5GECM-Cátedra Fisiología Animal, Departamento de Biología, Bioquímica y Farmacia, UniversidadNacional del Sur, Bahía Blanca, Argentina6Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
Received 8 July 2012; revised 21 November 2012; accepted for publication 30 November 2012
Despite recent taxonomic evaluations of Mephitidae and North American hog-nosed skunks, southern SouthAmerican species of Conepatus have not been thoroughly examined in a systematic context. Conepatus chingaand Conepatus humboldtii were described more than 150 years ago, based on external characters such as haircoloration and size. Although historically recognized as valid species, to date no detailed systematic analysis hasbeen performed for either of these taxa. Herein, we evaluated the taxonomic status of C. chinga and C. humboldtiiwithin the southern part of South America using geometric morphometrics of the skull and mandible, mitochon-drial DNA analysis using the cytochrome b and cytochrome oxidase c subunit I genes, and also control region andpelage pattern variation. We failed to find morphological (skull shape and pelage coloration patterns) or moleculardifferences between these two species; thus, we considered that the specimens assigned to C. chinga andC. humboldtii belong to the same species. Our results indicate that environmental variation seems to beresponsible for shape and size variation in Conepatus skulls from southern South America.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344.doi: 10.1111/zoj.12006
ADDITIONAL KEYWORDS: geographical variation – geometric morphometrics – mitochondrial DNAanalysis – taxonomy.
INTRODUCTION
In the last two decades there has been an increasingnumber of studies dealing with the taxonomic statusof the skunks, long recognized as part of the Musteli-dae family, which have introduced several relevant
modifications of their systematics (Dragoo et al., 1993;Dragoo & Honeycutt, 1997; Flynn et al., 2005). Infact, molecular data now indicate that skunks belongto their own family, Mephitidae (Dragoo et al., 1993;Dragoo & Honeycutt, 1997; Flynn et al., 2005; Satoet al., 2006, 2012; Eizirik et al., 2010).
The South American skunks are placed in the genusConepatus, the ‘hog-nosed skunks’, a Nearctic clade re-presented by fossils in South America since the middle*Corresponding author. E-mail: [email protected]
bs_bs_banner
Zoological Journal of the Linnean Society, 2013, 167, 327–344. With 9 figures
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344 327

Pleistocene (1.8–0.78 Mya), with a dubious record fromthe late Pliocene (ª 3 Mya) (Wang & Carranza-Castañeda, 2008; Prevosti & Pardiñas, in press). Threeliving species of Conepatus are recognized in SouthAmerica, Conepatus semistriatus (Boddaert, 1785)(Brazil, Belize, Guatemala, Honduras, Nicaragua,Mexico, and Peru), Conepatus chinga (Molina, 1782)(Argentina, southern Brazil, Bolivia, Chile, Paraguay,Peru, and Uruguay), and Conepatus humboldtii Gray1837 (southern Argentina and southern Chile)(Redford & Eisenberg, 1992; Wozencraft, 2005; Díaz &Lucherini, 2006; Wilson & Mittermeier, 2009).
The taxonomic history of South American Conepa-tus remains complex with several species describedin the 19th and 20th centuries that later went on to besynonymized (Cabrera, 1958). Using skin colour pat-terns and size, Cabrera & Yepes (1940) described fivespecies for Argentina and Chile: Conepatus suffocans,Conepatus castaneus, C. humboldtii, C. chinga, andConepatus rex. Cabrera (1958) recognized only fourspecies, believing C. suffocans to be synonymouswith C. chinga. This author also included Conepatusbudini and Conepatus gibsoni in C. chinga, and com-mented that C. castaneus might represent a northernpopulation of C. humboldtii although it differed inpelage patterns and undescribed cranial features(Cabrera, 1958). Kipp (1965) was the first to reviewthe systematics of South American Conepatus usingquantitative analysis applied to a large sample ofskins. She included C. rex in C. chinga and C.castaneus in C. humboldtii, and separated thesespecies based on the shape of the white bands in thedorsal pelage, and the presence of bicoloured hairs inC. humboldtii but not in C. chinga. Kipp’s work reit-erated patterns identified 70 years earlier by Milne-Edwards, Vaillant & Gervais (1891) who realized thatthe slight differences between C. castaneus andC. humboldtii did not support two different and rec-ognizable species. Later, Van Gelder (1968) studied apopulation of skunks from Uruguay, identified asC. chinga, corroborating the presence of wide varia-tion in skin patterns, even from a single locality.Van Gelder (1968) and later authors (e.g. Honacki,Kinman & Koeppl, 1982; Wozencraft, 2005; Wilson &Mittermeier, 2009), did not follow the systematicschema of Kipp (1965) in some regards (i.e. they didnot include C. suffocans, C. gibsoni, and Conepatusmendosus in C. humboldtii), but did in others (i.e. theinclusion of C. castaneus in C. humboldtii). This sys-tematic arrangement is what is currently recognizedby most authors (e.g. Redford & Eisenberg, 1992;Díaz & Lucherini, 2006; Emmons & Helgen, 2008; seealso Osgood, 1943), although Parera (2002) includedC. castaneus in C. chinga.
The two Recent southern South American specieswere described on the basis of external characters
such as size and colour patterns of the fur, as indicatedin the original description of C. humboldtii writtenby Gray (1837): ‘. . . black, with some rather shininghairs, with a broad white band along each side of theback . . .’. However, several authors described exten-sive variation amongst these characters (Van Gelder,1968; see also Yepes, 1936; Cabrera & Yepes, 1940;Cabrera, 1958; Kipp, 1965; Dragoo, Honeycutt &Schmidly, 2003; Dragoo & Sheffield, 2009). In addition,other authors have pointed to the difficulty in separat-ing these species based on skull anatomy, at least withqualitative and traditional morphometric approaches(i.e. using linear measurements; see Kipp, 1965).
The absence of clear diagnostic features generates aconfusing situation in the delimitation of the distribu-tion of these species (compare Redford & Eisenberg,1992 against Wozencraft, 2005), and a circularitybecause the specimens were identified to species levelbased solely on their geographical location. Further-more, there are several authors that clearly saw thatthe group needed a systematic revision of these skunks(e.g. Galliari, Pardiñas & Goin, 1996; Forasiepi, 2003;Wang & Carranza-Castañeda, 2008). Recent revisionsof other Conepatus species have shown that the NorthAmerican skunks Conepatus leuconotus and Conepa-tus mesoleucus represent a single species, and onlyC. mesoleucus figginsi probably represents a differentlineage relative to the larger clade containing theremaining taxa from North America (Dragoo et al.,2003). It is probable that a similar situation couldbe taking place between C. chinga and C. humboldtiiin South America. In areas where both species arepresent (e.g. northern Patagonia), the difficulty inseparating these species introduces a serious concernwhen it comes to the conservation of these skunks,as C. humboldtii is listed in the appendix II of CITES,but not C. chinga (Emmons & Helgen, 2008). Also,molecular data such as mentioned above are neededto identify potentially unique genetic stocks and toevaluate better the taxonomic status of populationsof Conepatus if management strategies are to beemployed to conserve these skunks.
In this paper, we evaluated the taxonomic statusof the southern hog-nosed skunk species (C. chingaand C. humboldtii) in Argentina and Uruguay, study-ing skull anatomy using geometric morphometrics,skin coloration patterns, and genetic divergence usingmitochondrial DNA (cytochrome b, cytochrome coxidase subunit I, and control region).
MATERIAL AND METHODSSAMPLE
We examined 350 specimens, of which 288 skulls andmandibles from museum collections were used for
328 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

geometric morphometrics, 186 skins for colorationpatterns (Table S1), and 25 tissue samples for DNAanalysis (Table S2). The specimens cover a wide areaof Argentina, Uruguay, and some areas of Chile(Fig. 1). We included holotypes of C. rex Thomas,1898, Conepatus proteus Thomas, 1902, C. gibsoniThomas, 1910, C. budini Thomas, 1919, and Conepa-tus calurus Thomas, 1919, in the geometric morpho-metrics analyses. Holotype skulls of C. humboldtiiGray, 1837 and C. castaneus D’Orbigny & Gervais,1847, were also analysed in two complementarydata sets because both skulls were broken and didnot allow the application of the same landmarks(see Figures S1–S5)
We explored the existence of differences betweenC. chinga and C. humboldtii as geographically delim-ited by Osgood (1943), Redford & Eisenberg (1992),Parera (2002), Díaz & Lucherini (2006), and Emmons
& Helgen (2008). Following this criterion, C. chinga ispresent from northern Río Negro province to thenorth of Argentina, adjacent areas in Chile, Uruguay,south-east Brazil, Paraguay, and Bolivia, whereasC. humboldtii is in Santa Cruz, Chubut, southern andcentral parts of Río Negro provinces, and adjacentareas in Chile. We also evaluated the systematicschema of Kipp (1965) and the difference betweenskunks from different ecoregions of Argentina andUruguay in the same way, but we got similar results,which are presented in Figures S1–S5.
GEOMETRIC MORPHOMETRIC ANALYSIS OF THE SKULL
We determined three age categories following VanGelder (1968): juvenile, with dentition not fullyerupted; subadult, with permanent dentition eruptedbut basisphenoid-basioccipital suture visible; and
Figure 1. Locality records for Conepatus chinga (grey shading) and Conepatus humboldtii (black shading).
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 329
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

adult, with permanent dentition erupted and nosutures visible. Only subadults and adults wereanalysed.
We took digital photographs of the skull in ventralview and mandible in lateral view from a total of 288specimens. Skulls were orientated with the palateand sagittal of the horizontal ramus of the mandibleparallel to the photographic plane. The semiland-marks were positioned using MakeFan 6 (Sheets,2002), which draws fans on photographs, enablingdigitization of semilandmarks that result from theintersection of the fan lines and the outlines of theanatomical structures. We used the program tpsUtil1.40 (Rohlf, 2008a) to compile image files and bothlandmarks and semilandmarks were digitized ontothe images using tpsDig 2.12 (Rohlf, 2008b; Fig. 2).Landmark configurations were superimposed throughgeneralized Procrustes analysis (GPA Goodall, 1991;Rohlf, 1999), minimizing the sum of squared dis-tances between homologous landmarks by translat-ing, rotating, reflecting, and scaling them to best fit,using tpsRelw 1.35 (Rohlf, 2003). For sliding semilan-dmarks we used the minimized bending energy of thethin-plate spline (Bookstein, 1997). The matrix withthe aligned coordinates of both landmarks and semi-landmarks were included in a principal componentanalysis with MorphoJ 1.03b software (Klinberger,2011).
We tested for the existence of shape and sizedifferences between subadults and adults usingone-way multivariate analysis of variance (MANOVA)and t-tests, respectively. Owing to the presenceof significant differences in shape (MANOVA,F = 4.87, P < 0.0001) but not in size (t-test, T = 0.49,P = 0.6259), we chose to only present the analysis of
adult specimens, although a more inclusive analysiswith the two age groups combined yielded similarresults (data not shown). The presence of sexualdimorphism in shape and size was tested using thesame tests, but because of the absence of significantdifferences in shape (MANOVA, F = 0.93, P = 0.5553)and size (t-test, T = 0.98, P = 0.3373), the samplewas pooled. All the MANOVAs and t-tests were per-formed with InfoStat v.2010 software (Di Rienzoet al., 2010).
We explored the existence of differences betweenC. chinga and C. humboldtii using principal com-ponent analysis and discriminant analysis withcross-validation in order to calculate percentage ofreclassification of each group and 10 000 permutationsto obtain the significance of Procrustes distances inMorphoJ 1.03b software (Klinberger, 2011).
The relationship between size (centroid size) andshape were tested with multivariate regression inMorphoJ 1.03b software (Klinberger, 2011). The sig-nificance of the regression was tested with a permu-tation test and 10 000 resamples. We explored therelationship between skull shape (aligned Procrustescoordinates) and latitude/longitude with canonicalcorrelation analysis (CCA), to test for the presence ofvariation across the geographical ranges, which are inturn linked to environmental changes. We performeda Mann-Whitney U-test to evaluate the differencesbetween centroid size of C. chinga and C. humboldtiibecause of the lack of normality in this variable.A possible correlation between centroid size andlatitude/longitude was evaluated with simple regres-sion in MorphoJ software (Klinberger, 2011) and mul-tiple regression in R version 2.12.0 (R DevelopmentCore Team, 2005).
Figure 2. Distribution of the landmarks (white circles) and semilandmarks (white squares) used in morphometricanalysis. A, skull; B, mandible.
330 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

Using new methodologies that allow one to uselandmark configurations to infer phylogenetic trees(Catalano, Goloboff & Giannini, 2010; Goloboff &Catalano, 2010), we obtained a phylogeographicaltree based on the shape of the skull from a ventralview and the mandible from a lateral view undermaximum parsimony with the software TNT 1.1(Goloboff, Farris & Nixon, 2008). We included only 70specimens that had both mandibles and skullsand the tree was rooted with C. semistriatus. Thesearch was performed with 30 random sequence addi-tions plus tree bisection-reconnection (TBR), usingthe superimpositions (alignments) obtained from theGPAs. The three shorter trees were used to performnew searches using TBR and change the alignmenttrying to find shorter trees, until two successive TBRrounds found the same tree (Catalano et al., 2010; S.Catalano, pers. comm., 2011).
ANALYSIS OF SKIN COLORATION PATTERNS
Photographs of 145 museum skins, 17 live captures,and 23 camera trap photos were used to observevariation in colour patterns. These samples included164 specimens of C. chinga and 21 of C. humboldtii.
Each specimen was classified into one of fourdorsal stripe patterns (Fig. 3A). Specimens were alsogrouped into categories based on the presence andshape of white coloration on the head (Fig. 3B) andthe region of the body where the stripes merged. Werecorded the presence of white and black monochro-matic hair in the tail. Finally, we looked for bicolouredhair (white base, black tip) in the tail because thischaracter had been previously used to separate thetwo species (Kipp, 1965; Wozencraft, 2005).
MITOCHONDRIAL DNA ANALYSIS
Tissue samples of specimens were collected by per-sonnel of the Wildlife National Office (DirecciónNacional de Fauna Silvestre, Secretaría de Ambientey Desarrollo Sustentable). Skulls and mandibles ana-lysed were deposited in the Mammal Collections ofMuseo Argentino de Ciencias Naturales ‘BernardinoRivadavia’ (MACN) and Museo de Ciencias Naturalesde La Plata (MLP), Argentina.
We used the cytochrome b (Cyt b) and cytochromeoxidase c subunit I (COI) genes, and also the controlregion (D-loop), to examine genetic diversity amongst25 individuals from 12 localities in Argentina (Fig. 1,
Figure 3. A, four categories of dorsal stripe patterns in hog-nosed skunks (Conepatus chinga and Conepatus humboldtii)in Argentina and Uruguay. The categories are pattern 1, stripes absent; pattern 2, stripes extending from the crown tothe mid-dorsal region; pattern 3, stripes extending from the crown to the hips; pattern 4, stripes extending the full lengthof the body and onto the base of the tail. B, three head white colour patterns in hog-nosed skunks (C. chinga andC. humboldtii) in Argentina and Uruguay.
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 331
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

Table S2). Tissue samples were collected in the fieldand preserved in 96% ethanol. DNA was extractedfrom fixed or dried tissues (liver, spleen, muscle, andskin) using the salt-extraction protocol (Aljanabi &Martinez, 1997).
The Cyt b gene was amplified using primersdescribed in Smith & Patton (1993). The COI genewas amplified and sequenced at the Canadian Centrefor DNA Barcoding, Biodiversity Institute of Ontario(University of Guelph, Guelph, Ontario) using thecocktail primers described in Ivanova et al. (2007) aspart of collaborations between CONICET and iBOL(International Barcode of Life). Finally, the D-loopwas amplified using the primers described in Poljaket al. (2010).
Cyt b and D-loop amplifications of the double-stranded product were performed in a total volume of50 mL, containing 5 mL of 10¥ reaction buffer, 1 mLdeoxyribonucleoside triphosphate mix at 10 mMeach, 1 mL of each primer at 10 mM, 0.2 mL Taq DNApolymerase equivalent to 1 unit of polymerase pertube, and 1 mL DNA. The thermal profiles consisted ofan initial denaturation at 94 °C for 3 min, followed by35–40 cycles at 94 °C for 30 s, 55–57 °C for 30 s, andat 74 °C for 1 min, with a final extension step at 74 °Cfor 5 min. Double-stranded PCR products were puri-fied with a Qiaquik PCR Purification Kit (Qiagen),examined on 1% agarose gels, and directly sequencedin both directions using the same primers as thoseused for amplifications. Sequencing was conducted atMAGROGEN Sequencing Service (Korea).
A molecular phylogenetic approach was used toinvestigate the taxonomic status of Conepatus speciesfrom Argentina. In the three molecular analyses, weused Mephitis, Spilogale, and Taxidea as outgroupsequences.
For the Cyt b marker, we sequenced 14 specimensof Conepatus from Argentina and used one availablesequence of C. chinga from Bolivia and two of C. leu-conotus. For the COI marker, we sequenced 19 speci-mens of Conepatus from Argentina and used twoavailable sequences of C. leuconotus (see Table S2)and one of Conepatu semistratus. Twelve smaller(188 bp) sequences of Conepatus from Brazil were alsoused. Chaves et al. (2011) identified these specimensas C. chinga and C. semistratus although they did notform reciprocally monophyletic groups. For the D-loopmarker, we sequenced 14 specimens of Conepatusfrom Argentina and used one available sequence ofC. chinga from Bolivia and two of C. leuconotus.
Sequences were aligned manually using BioEdit(Hall, 1999). First, separate analyses were conductedon the Cyt b, COI, and D-loop markers. For the threemolecular markers, appropriate substitution modelswere estimated with the Akaike information criterionas implemented in MrAIC (Nylander, 2004). Three
phylogenetic reconstruction methods were analysed.First, maximum likelihood (ML) phylogenetic recon-structions were performed using TreeFinder (Jobb,von Haeseler & Strimmer, 2004). Confidence valuesfor the edges of the ML tree were computed viabootstrapping (Felsenstein, 1985), based on 1000replicates. Second, Bayesian inference (BI) analysiswas conducted in MrBayes 3.1.2 (Huelsenbeck &Ronquist, 2001; Ronquist & Huelsenbeck, 2003). Fourchains were run simultaneously (three heated, onecold) for 20 000 000 generations, with tree spacesampled every 100th generation. After a graphicalanalysis of the evolution of the likelihood scores,the first 300 000 generations were discarded as burn-in. The remaining trees were used to calculate theconsensus tree. Third, a maximum parsimony (MP)analysis was performed with the software TNT 1.1(Goloboff et al., 2008). We performed a traditionalsearch using TBR with 1000 random sequence addi-tions, and jackknife support values based on 1000replicates. Finally, a concatenated sequence analysiswas then performed with the three markers: Cytb, COI, and D-loop. Each of the three estimationmethods, ML, BI, and MP, were performed on theconcatenated data set as described above. We used allavailable data, and a DNA supermatrix with 41 speci-mens of Conepatus and 1742 bp was obtained. Thismatrix had 50% missing data. For Conepatus species,we computed a distance matrix of the Kimura ‘two-parameter’ model (Kimura, 1980) for each marker. Inthe case of COI, the specimens from Brazil were notanalysed for genetic distances because the speciesdetermination was not solved in the original work(Chaves et al., 2011).
RESULTSSKULL MORPHOMETRICS AND SIZE: DIFFERENCES
BETWEEN C. CHINGA AND C. HUMBOLDTII
PCA did not show any separation between C. chingaand C. humboldtii in the shape of the skull in ventralview or the mandible in lateral view in any compo-nent axis. The first and second axes of the skull in theventral view analysis (Fig. 4A) explained 22.39 and9.49% of the variance, respectively. Along the firstprincipal component (PC), toward its positiveextreme, are specimens with narrow skulls, smallertemporal fossa, more anteriorly placed upper carnas-sial (P4) and first upper molar (M1), smaller bulla,larger magnum foramen, and more posteriorly placedmastoid and paracondylar processes (the opposite ten-dency is observed towards the negative extreme of thefirst PC; Fig. 4A). Along the second PC, towards itspositive extreme, are specimens with longer skulls,smaller temporal fossa and glenoid cavity, bulla and
332 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

Fig
ure
4.Sc
atte
r-pl
ots
ofpr
inci
pal
com
pone
ntan
alys
isof
skul
l(A
)an
dm
andi
bles
(B).
Bla
ckdo
tsre
pres
ent
Con
epat
usch
inga
,gre
ydo
tsC
onep
atus
hum
bold
tii,
and
cros
ses
holo
type
s.F
requ
ency
ofcr
oss-
valid
atio
ndi
scri
min
ant
scor
esan
dde
form
atio
nw
iref
ram
egr
aphs
ofsh
ape
diffe
renc
esof
skul
ls(C
)an
dm
andi
bles
(D).
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 333
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

mastoid process and posterior border of the palate,and P4-M1 larger and more posteriorly placed (theopposite tendency is observed towards the negativeextreme of the second PC; Fig. 4A). Regarding thelateral mandible analysis (Fig. 4B) the values were32.06% for the first axis and 12.94% for the second.Those individuals with positive values for the firstPC displayed longer and more robust mandibles, withmore vertical coronoid process, whereas the oneswith negative values presented shorter and moreslender mandibles, with a posteriorly inclined coro-noid process. On the second PC, individuals withpositive values displayed a lower coronoid process andstronger mandibular body, whereas the specimenswith negative values demonstrated the opposite ten-dencies (Fig. 4B).
Discriminant function analysis (DFA) showed sig-nificant differences (P < 0.0001) in the ventral viewof the skull (Fig. 4C) with Procrustes (0.0173) andMahalanobis (3.493) distances between means of eachgroup. The reclassification percentage reached 83.33%.Analysis based on the lateral mandible view (Fig. 3D)displayed significant differences (P < 0.0003) in Pro-crustes (0.019) and Mahalanobis (3.387) distances andthe reclassification percentage was 71.62%.
The Mann-Whitney U-test for ventral skull meas-urements showed no significant differences (P = 0.111)between centroid size of C. chinga (mean = 1031.975)and C. humboldtii (mean = 1017.536). The same testfor lateral mandible dimensions showed no signifi-cant differences (P < 0.8632) between centroid sizeof C. chinga (mean = 639.4267) and C. humboldtii(mean = 642.7335).
The most parsimonious phylogenetic tree (21.03steps) obtained from the matrix of skull and mandibleshape showed that C. chinga and C. humboldtii arecompletely mixed and did not recover any mono-phyletic groups (Fig. 5). In addition, specimens fromgeographically disparate localities were often clus-tered together (e.g. Buenos Aires and Salta provinces).
SKULL MORPHOMETRICS AND SIZE: ALLOMETRY AND
GEOGRAPHICAL VARIATION
The shape of skulls was positively correlated withsize (Fig. 6A), as expressed by the regression analy-
sis with P < 0.0001. The percentage of the varianceexplained by size was 55.7%. Larger specimens havesmaller P4-M1, smaller magnum foramen and bulla,larger temporal fossa, narrower palate at or posterior
Figure 5. Phylogeographical tree of skulls and mandiblesunder maximum parsimony. Csem, Conepatus semistria-tus; Cch and , Conepatus chinga; Chum and , Conepa-tus humboldtii; Ecu, Ecuador; Uru, Uruguay; Salt, Salta;BsAs, Buenos Aires; ER, Entre Ríos; LaPamp, La Pampa;Mend, Mendoza; Chu, Chubut; Cord, Córdoba; Jjy, Jujuy;Tuc, Tucumán; Corr, Corrientes; Rngr, Río Negro; Nqn,Neuquén; SE, Santiago del Estero; m, male; f, female. Foracronyms see Table S1.
!
334 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

Fig
ure
6.Sc
atte
r-pl
ots
ofre
gres
sion
scor
esof
shap
ean
dce
ntro
idsi
zefo
rsk
ulls
(A)
and
man
dibl
es(B
).D
efor
mat
ion
grid
sar
eal
sosh
own.
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 335
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344
to the M1, and the palate more posteriorly expanded.Small specimens showed the opposite pattern (e.g.large bulla, small temporal fossa). The shape of man-dibles (Fig. 6B) was also positively correlated withsize (P < 0.05) with 50.5% of the variance explainedby size. Large specimens have a higher mandibularbody (especially below premolars and canine), and acoronoid process that is more posteriorly inclined,whereas in small specimens the mandibular body isslender and the coronoid process more vertical.
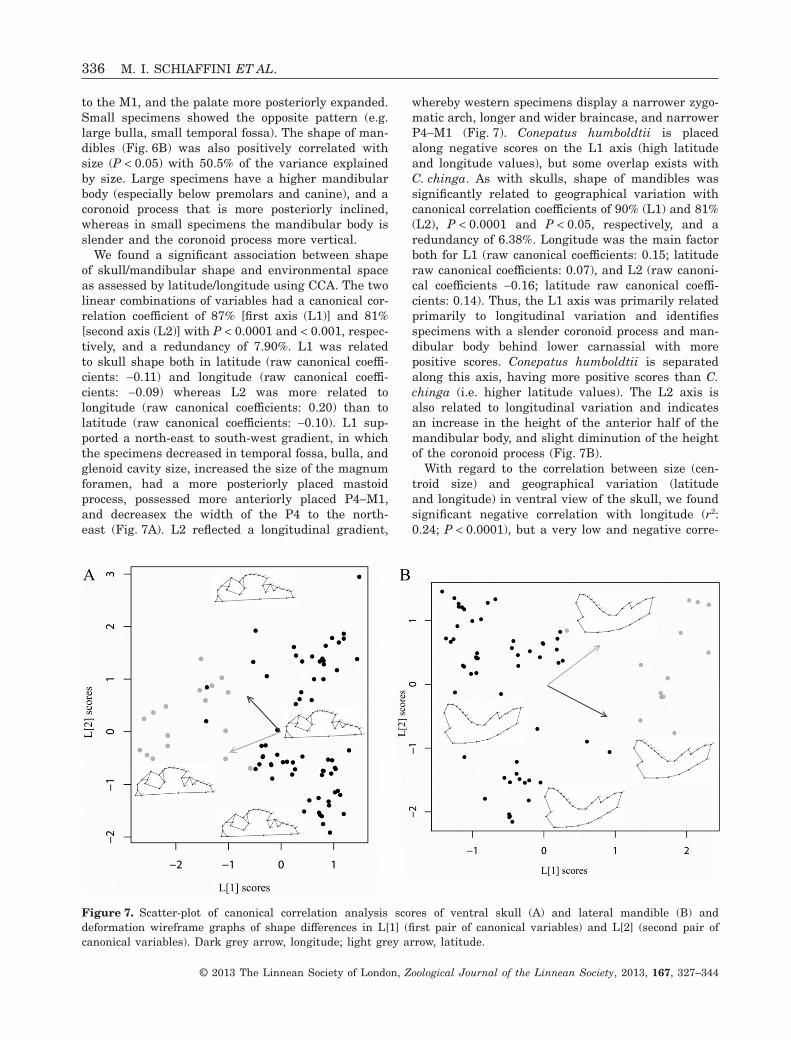
We found a significant association between shapeof skull/mandibular shape and environmental spaceas assessed by latitude/longitude using CCA. The twolinear combinations of variables had a canonical cor-relation coefficient of 87% [first axis (L1)] and 81%[second axis (L2)] with P < 0.0001 and < 0.001, respec-tively, and a redundancy of 7.90%. L1 was relatedto skull shape both in latitude (raw canonical coeffi-cients: -0.11) and longitude (raw canonical coeffi-cients: -0.09) whereas L2 was more related tolongitude (raw canonical coefficients: 0.20) than tolatitude (raw canonical coefficients: -0.10). L1 sup-ported a north-east to south-west gradient, in whichthe specimens decreased in temporal fossa, bulla, andglenoid cavity size, increased the size of the magnumforamen, had a more posteriorly placed mastoidprocess, possessed more anteriorly placed P4-M1,and decreasex the width of the P4 to the north-east (Fig. 7A). L2 reflected a longitudinal gradient,
whereby western specimens display a narrower zygo-matic arch, longer and wider braincase, and narrowerP4-M1 (Fig. 7). Conepatus humboldtii is placedalong negative scores on the L1 axis (high latitudeand longitude values), but some overlap exists withC. chinga. As with skulls, shape of mandibles wassignificantly related to geographical variation withcanonical correlation coefficients of 90% (L1) and 81%(L2), P < 0.0001 and P < 0.05, respectively, and aredundancy of 6.38%. Longitude was the main factorboth for L1 (raw canonical coefficients: 0.15; latituderaw canonical coefficients: 0.07), and L2 (raw canoni-cal coefficients -0.16; latitude raw canonical coeffi-cients: 0.14). Thus, the L1 axis was primarily relatedprimarily to longitudinal variation and identifiesspecimens with a slender coronoid process and man-dibular body behind lower carnassial with morepositive scores. Conepatus humboldtii is separatedalong this axis, having more positive scores than C.chinga (i.e. higher latitude values). The L2 axis isalso related to longitudinal variation and indicatesan increase in the height of the anterior half of themandibular body, and slight diminution of the heightof the coronoid process (Fig. 7B).
With regard to the correlation between size (cen-troid size) and geographical variation (latitudeand longitude) in ventral view of the skull, we foundsignificant negative correlation with longitude (r2:0.24; P < 0.0001), but a very low and negative corre-
Figure 7. Scatter-plot of canonical correlation analysis scores of ventral skull (A) and lateral mandible (B) anddeformation wireframe graphs of shape differences in L[1] (first pair of canonical variables) and L[2] (second pair ofcanonical variables). Dark grey arrow, longitude; light grey arrow, latitude.
336 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

lation with latitude (r2: 0.08, P < 0.01). The size ofmandibles in the lateral view showed the samepattern, that is it was uncorrelated with latitude(r2: 0.0002, P < 0.8846), but was correlated with lon-gitude (r2: 0.15, P < 0.00001). Multiple regressionsalso showed that longitude was the only significantfactor correlated with size (figures not shown).
PELAGE VARIATION
All of the examined specimens of Conepatus wereblackish or brownish black with a varying amountof white hair. Of the total specimens attributed toC. chinga, 30 were brownish and 134 blackish. Analy-sis of the four categories of dorsal stripe patternsrevealed that C. chinga was represented by all ofthem, with 6.7% of the specimens in category 1, 16%in category 2, 28.8% in category 3, and 41.5% incategory 4 (Fig. 8A). The white coloration on the headregion was absent in 22.4% of the specimens, spottedin 6.1%, and striped in 72% (Fig. 8B). The stripesmerged on the head region in 69 individuals, on theneck region in 30, and behind the neck in 18. A totalof 81 individuals specimens of C. chinga displayedstripes that did not merge (Fig. 8C).
Of the 26 specimens assigned to C. humboldtii, tenwere brownish and 16 blackish. Conepatus humbold-tii had individuals represented in three of the fourcategories of dorsal stripe pattern, with 80.8% incategory 4, 15.4% in category 3, and 3.8% in category2 (Fig. 8A). In 15.4% of the individuals white colora-tion on the head region was absent, whereas in 84.6%white hair constituted two well-defined stripes. Wedid not find individuals with white spots on the head(Fig. 8B). Thirteen individuals had two separateddorsal stripes from the head to the tail, in 12 skunksthe stripes merged on the head region, and only inone individual did the stripes merge on the neckregion (Fig. 8D). Finally, 12 individuals of C. hum-boldtii possessed stripes that did not merge.
The amount and distribution of white hair on thetail in both species was highly variable and did notallow the identification of different patterns. Acrossthe 129 individuals classified as C. chinga, 126 (98%)had black hair on the tail, whereas 70% of specimenshad white. In C. humboldtii (N = 20), 95% of individu-als had black hair on the tail and 85% had white.
We examined 149 specimens (C. chinga = 129,C. humboldtii = 20) for the presence of bicoloured hairon the tail. For C. humboldtii, bicoloured hairs werefound in 80% of the analysed specimens whereas forC. chinga 79.1% of the skunks had only unicolouredhair on the tail.
Albeit the existence of a difference in the percent-age of representation of each state in C. chinga andC. humboldtii, there is no clear difference between
these species, because some specimens of C. chingahave states not present in C. humboldtii (see aboveand Fig. 8).
MITOCHONDRIAL DNA ANALYSIS
Sequence alignment of the Cyt b marker comprised826 bp positions, from which 88 bp were variablewithin the ingroup. The model of sequence evolutionthat fitted our sequence data best was HKY (Hase-gawa Kishino Yano, Hasegawa et al., 1985) + gamma,according to MrAIC (Nylander, 2004). The ML, BI,and MP phylogenetic trees obtained had similar
Figure 8. Observed frequency for hog-nosed skunks(Conepatus chinga and Conepatus humboldtii) inArgentina/Uruguay and adjacent states for A, dorsal stripepattern; B, white coloration on the head, and C, region ofthe body at which the white dorsal stripes merge.
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 337
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

topologies. The BI tree with the support values of thethree analyses is shown in Figure 9A. Conepatusleuconotus appears as the sister group of Conepatussequences from Argentina and Bolivia. The speci-mens identified as C. chinga formed a paraphyleticgroup, whereas C. humboldtii formed a monophyleticgroup.
For the COI marker the sequence alignmentcomprised 658 bp positions, from which 106 werevariable within the ingroup. The model of sequenceevolution that best fitted our sequence data set wasHKY + gamma, according to MrAIC (Nylander, 2004).The ML, BI, and MP phylogenetic trees obtained havesimilar topologies. The BI tree is shown in Figure 9B.In this analysis, the sister-species of Conepatusspecies from Argentina and Brazil is C. semistratus.Then, these species cluster with C. leuconotus. Thespecimens identified as C. chinga again formed aparaphyletic group with four specimens from Brazil,Specimens identified as C. humboldtii clustered withfive specimens of Conepatus sp. from Brazil, to form amonophyletic group.
The sequence alignment of the D-loop markercomprised 255 bp positions, from which 22 bp werevariable within the ingroup. The model of sequenceevolution that best fitted our sequence data set wasgeneral time reversible + gamma, according to MrAIC(Nylander, 2004). The ML, BI, and MP phylogenetictrees obtained have similar topologies. The BI tree isshown in Figure 9C. In this analysis, the specimensidentified as C. chinga and C. humboldtii were recip-rocally monophyletic groups but branch support waslow for ML and MP.
In the concatenate analysis, the ML, BI, and MPphylogenetic trees obtained have similar topologies.The BI tree is shown in Figure 9D. Well-supportednodes in this analysis indicate that the sister-speciesof Conepatus species from Argentina and Brazilis C. semistratus. Then, these species cluster withC. leuconotus. The specimens identified as C. chingaagain formed a paraphyletic group with specimensfrom Brazil, but C. humboldtii and the specimensfrom Brazil are a monophyletic group.
Per cent sequence divergence amongst taxa wasestimated for each marker separately (Fig. 9). We alsocalculated the intraspecies sequence divergence to
determine the species limits of Conepatus. All possiblepairwise comparisons of Conepatus species indicatedthat the smallest per cent sequence divergenceamongst species occurred between the specimens iden-tified as C. chinga and C. humboldtii (3.4% for Cyt b;2.5% for D-loop, and 1.5% for COI). These values are3.7 times smaller for Cyt b and 1.44 times smallerfor D-loop than the divergence between C. chinga vs.C. leuconotus or C. humboldtii vs. C. leuconotus. Forthe COI gene the divergence between C. chinga andC. humboldtii was five times smaller than the diver-gence between C. chinga vs. C. semistratus or C. hum-boldtii vs. C. semistratus (see Fig. 9).
DISCUSSION
During the 19th century and the first half of the 20th
century, many Conepatus species were described basedon external characters, such as size and pelage/colourpatterns. For South America in particular, Mr O.Thomas described several species of the genus includ-ing C. rex (1898), C. arequipae (1900), C. chorensis(1902), C. gibsoni (1910), C. ajax (1913), and two newspecies from a single locality: C. budini and C. calurus(1919) from Otro Cerro, La Rioja. Surprisingly,Thomas himself in 1902 (referenced in Thomas, 1920)pointed to ‘the difficulty of a proper determination ofthe members of Conepatus, and the probability thatmost of the forms grade into each other’ and that ‘. . .every character varies individually . . .’. Despite thesewarnings, Cabrera & Yepes (1940), Cabrera (1958),and Kipp (1965) synonymized several of the aforemen-tioned species relying exclusively on external charac-ters. The absence of clear characters to discriminatethese species often resulted in specimens beingassigned specific status using published distributionmaps instead of diagnostic morphological characters.This introduced ambiguity in previous taxonomicassignation and complicated the evaluation of theirvalidity. Thus, this is the first rigorous work in whichreliable systematic characters are employed for SouthAmerican Conepatus, including novel techniques suchas geometric morphometrics and mitochondrial DNAanalysis.
Both the PCA and MP analysis of the morphometricdata show a wide overlap between C. chinga and
Figure 9. Bayesian inference trees for the Conepatus genus. A, tree derived from cytochrome b; B, tree derived fromcytochrome oxidase c subunit I; C, tree derived from D-loop marker and D, tree derived from the analysis concatenatingthese markers. Numbers next to branches are Bayesian posterior probabilities, bootstrap values followed by jackknifesupport values, respectively. The tables show the percentage of sequence divergence (Kimura, 1980) of Conepatus speciesfor each marker. ER, Entre Ríos; BsAs, Buenos Aires; LaPamp, La Pampa; Chu, Chubut; Rngr, Río Negro; Nqn, Neuquén;SaFe, Santa Fe; Scrz, Santa Cruz; BrGo, Brasil-Goias; BrPi, Brasil-Piaui; BrMG, Brasil-Minas Gerais; BrRGS, Brasil RioGrande do Sul; Bol, Bolivia; , Conepatus chinga; , Conepatus humboldtii; , Conepatus sp. For GenBank accessionnumbers or iBOL accession numbers see Table S2.
!
338 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 339
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

C. humboldtii, allowing no separation between thesetwo groups (Figs 4, 5). The PCA also demonstratesthat specimens from different regions of Argentinaoverlap widely in their character states (data notshown, see also Schiaffini et al., 2010). Contrary tothe PCA results, DFA did support significant differ-ences in Mahalanobis and Procrustes distances withsome overlap, but several factors and bias presentin the DFA preclude us from accepting this result(see Kovarovic et al., 2011). First, the specimens weredetermined a priori as C. chinga and C. humboldtiiaccording to their geographical location, without anyclear morphological or molecular justification (a non-ambiguous prior classification of specimens is neededin DFA; see Davis, 1986; Legendre & Legendre, 1998).This could be a problem given the geographical patternof variation in shape and size found in our analysis (seeabove), because if the sample is separated in two usinga geographical distribution (e.g. Patagonia-northernareas) it is easy to find significant differences. Weconfirmed this, because extracting the skull variationexplained by latitude and longitude (or size), thedistances are not significant and the reclassificationpercentages fall to 31–36% (worse than the randomexpectation of 50%), indicating that the differences aregiven by a geographical pattern of variation. It is alsoimportant to point out that Mahalanobis and Pro-crustes distances in DFA under Kipp’s (1965) system-atic scheme (see Figures S1–S5), or even alternativedivision (Pampean Region against other areas; datanot shown), were also significant if we did not take inaccount the pattern of geographical variation in theskull and mandible. This demonstrates that evenarbitrary divisions could lead to significant differen-tiation when using DFA.
Results of the pelage colour pattern analysis indi-cated that frequency differences in the distribution ofstates were present. However, such differences wereinconsistent enough that we failed to find a patternthat could help separate or recognize the two species.This agrees with previous studies that found broadvariation in pelage coloration patterns (e.g. Kipp,1965; Van Gelder, 1968; Dragoo et al., 2003). In arecent popular book, Canevari & Vaccaro (2007) usedthe presence of long and soft hairs in C. humboldtiibut long and rough hairs in C. chinga as a characterto separate these species. Although we did not quan-tify it, the variations observed in the studied skinsdid not allow us to use this as a good diagnosticcharacter to separate these taxa. In a similar fashion,in another popular book, written by Chebez (2009),the author used pelage characteristics of study skinsto recognize four valid species of Conepatus in SouthAmerica (C. chinga, C. castaneus, C. rex, and C. hum-boldtii). The author further stated that C. semistria-tus is present in the Chaco Region of Argentina.
However, as discussed in this paper, most of thecharacters used by this author are highly variableand do not allow complete and reliable separation ofthese skunks. In addition, there is no evidence tosupport the presence of C. semistriatus in Argentina.Most likely, this variation in pelage patterns isrelated to different environments occupied by popula-tions of Conepatus that inhabit southern SouthAmerica, and not to specific differences (e.g. Cabrera,1971; Cabrera & Willink, 1980; García, 1990; Burkartet al., 1999; Olson et al., 2001).
The results obtained from the mitochondrial genesagree with the absence of separation between thesespecies in morphological features. Trees constructedusing Cyt b and COI sequences failed to find recipro-cal monophyly of C. humboldtii and C. chinga,although the first formed a natural group with Cyt b(see Fig. 9A). Only the D-loop tree recovered a recip-rocal monophyletic relationship for these taxa, butthe branch support was not very high. Additionallythe observed molecular distances between these taxaare low and smaller than the observed distancesbetween them and C. leuconotus and C. semistriatus.Dragoo et al. (1993, 2003) found similar results usingthe same methodology applied to North AmericanConepatus, whereby they synonymized C. leuconotusand C. mesoleucus. Moreover, for Cyt b, the geneticdistances found here between the specimens deter-mined as C. humboldtii and C. chinga are comparableto the intraspecific genetic distances in other speciesof carnivores such as Nasua narica (4.4–4.9%; Helgenet al., 2009).
For COI, the mean intraspecific variability foundfor 152 species of mammals was 1.1 ± 1.5% (Meier,Zhang & Ali, 2008) and the range of this distance forsome species of carnivore was 0.065–2.3% (Tobe,Kitchener & Linacre, 2010). In this work we foundgenetic distances that are in the same range of theseprevious results.
It is interesting that Patagonian skunks (i.e. des-ignated as C. humboldtii) are clustered with someBrazilian ones that could belong to C. chinga orC. semistriatus (see Chaves et al., 2011) following theCOI and combined analysis (Fig. 9B, D). However, itmust be stated that a sequence of C. semistriatus(from Mexico, its type ‘locality’) falls outside the cladeof C. chinga-C. humboldtii (and has 7.618–7.319%divergence from them; Fig. 9B), suggesting thatspecimens from Brazil do not belong to C. semistria-tus, but to C. chinga-C. humboldtii. We note thatthe specimens from Brazil that cluster with C. hum-boldtii are from Rio Grande do Sul, the southernregion of this country. Moreover, the individualsthat cluster with the C. chinga are from Goias, Piaui,and Minas Gerais, regions in the north of Brazil,and these clusters clearly did not agree with geo-
340 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

graphical distances. A broader analysis with moresamples from Brazil, northern South America, andCentral-Northern America is needed to corroboratethis finding.
The geographical pattern observed in the skull mor-phology of Conepatus could be linked to environmen-tal variation. The region that we studied has verydissimilar environments: high and low altitudes (i.e.Puna and Patagonian Steppe), wet and dry regions(i.e. Humid Pampas and Dry Chaco), and hot andtemperate areas (i.e. High Monte and PatagonianSteppe) (Burkart et al., 1999; Olson et al., 2001). Thesample area comprises a wide range of latitudes,including a conspicuous temperature gradient and aprecipitation gradient that is present from north-eastArgentina to the south-west and Patagonia, and othermore localized gradients might be present along alti-tudinal differences in the Andes (see García, 1990).These changes impose different abiotic conditions, butalso different vegetation and resource availability(e.g. McNab, 2010). The clinal pattern observed in theskull is possibly related to these environmental pat-terns, especially as demonstrated by the correlationwith longitude, which could be generated by thestrong north-east to south-west diminution of precipi-tation. The decrease in skull size to the west could beseen as an effect of the increase of aridity and sea-sonality, but shape is also affected by latitude andlongitude. Some of the characters observed in thisclinal variation (development of the zygomatic arch,size and position of P4-M1) could also be related todiet differences (see Van Valkenburgh, 1989, 2007;Biknevicius & Van Valkenburgh, 1996). Specimensfrom the south-west tend to have larger temporalfossa, have more anteriorly placed P4-M1, and widerP4, and there is also a pattern along longitudewhereby eastern specimens have wider M1-P4(Fig. 7A). Other changes associated with latitude andlongitude include the development of the coronoidprocess and the mandible body (Fig. 7B). Severalstudies have shown that changes in these charactersare related to different diet types (e.g. Ewer, 1973;Turnbull, 1970; Van Valkenburgh, 1989, 2007;Biknevicius & Van Valkenburgh, 1996; Popowics,2003); thus, it is possible that they could be explainedby differences in prey availability along the studiedarea. In this regard, Baryshnikov, Puzachenko &Abramov (2002) found a great amount of size varia-tion of cheek teeth in Eurasian badgers related togeographical variation, which they interpreted asadaptations to different diets. It is interesting tonote that other carnivores, like the South Americangrey fox (Lycalopex gymnocercus-Lycalopex griseus)present a similar pattern (body size increases to thenorth-east, along the same area: see Zunino et al.,1995; Prevosti et al., 2011) This hypothesis must
be tested with a deeper biomechanical and morpho-metric analysis and complemented with ecologicalinformation.
In summary, skull morphometry, variation of pelagecoloration patterns, and mitochondrial DNA do notsupport the recognition of two distinct Conepatusspecies in Argentina. We do not include the typeof C. chinga or DNA and skins from its type locality,but do for C. humboldtii and junior synonyms ofthese species (e.g. C. castaneus, C. ajax; see above).As we included skulls and skins from the typelocality (Magallanes Strait, Chile; see Cabrera, 1958)of C. humboldtii, and sequences from areas nearit (Esperanza, Santa Cruz province, Argentina),C. humboldtii could be the valid name of the skunksof Argentina, Uruguay, and the Chilean Patagonia.However, we also failed to find differences in skullsbetween specimens from the type locality of C. chinga(around Valparaiso, Chile; see Osgood, 1943; Cabrera,1958) and from other localities. With the availableinformation we prefer to consider the skunks of south-ern South America (Uruguay, central and southernChile, Argentina) as belonging to one species, andC. chinga has page priority and is the valid name forthis taxon. The phylogenetic position of the sequencesfrom Brazil (Chaves et al., 2011) indicates that theyalso should be included in C. chinga. More morpho-logical samples from Chile, DNA samples from thiscountry and from north-west Argentina, and othergenes will help to test this hypothesis and the con-clusions presented in this paper.
ACKNOWLEDGEMENTS
We thank the following people for allowing access tospecimens under their care: Sergio Bogan (FundaciónFélix de Azara), Rubén Barquez and Mónica Díaz(Instituto Miguel Lillo), Jaime Cárcamo (Instituto dela Patagonia, Zoología), David Flores (Museo Argen-tino de Ciencias Naturales ‘Bernardino Rivadavia’),Enrique González (Museo de Historia Natural deMontevideo), Diego Verzi and Itatí Olivares (Museode La Plata), Damián Romero (Museo Municipal deCiencias Naturales ‘Lorenzo Scaglia’), GeraldineVerón (Muséum National d’Histoire Naturelle),Barbara Herzig (Naturhistorischen Museum Wien –Mammalogy), Doris Moerike (Staatliches Museum fürNaturkunde Sttutgart – Mammalogy). We also thankMaría J. Corriale (UBA) and G. Porini (DFS, SAyDS)for field collaboration. María J. Corriale also preparedthe skulls collected from Río Negro, La Pampa, andBuenos Aires provinces. Paula Jenkins and RobertoPortela Miguez kindly sent us photographs of holo-types specimens from the Natural History Museum.We would especially like to acknowledge CécileCallou for some pictures from the Muséum National
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 341
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

d’Histoire Naturelle. We are especially grateful to S.Catalano for helping us with the maximum parsi-mony tree using the cluster of the Instituto MiguelLillo (Tucumán, Argentina) and to Adam W. Fergusonfor a critical reading of the manuscript. Thanks tothe Dirección Nacional de Fauna Silvestre (SAyDS),Argentina for their collaboration. We acknowledgesupport from Consejo Nacional de InvestigacionesCientíficas y Técnicas (CONICET), Argentina andInternational Barcode of Life Project (Canada).Thanks to CONICET (PIP 112-201101-00164) andAgencia Nacional de Promoción (PICT 2011-0309) forfinancial support.
REFERENCES
Aljanabi SM, Martinez I. 1997. Universal and rapid salt-extraction of high quality genomic DNA for PCR-basedtechniques. Nucleic Acids Research 25: 4692–4693.
Baryshnikov GF, Puzachenko AY, Abramov AV. 2002.New analysis of variability of cheek teeth in EurasianBadgers (Carnivora, Mustelidae, Meles). Russian Journal ofTheriology 1: 133–149.
Biknevicius AR, Van Valkenburgh B. 1996. Design forkilling: craniodental adaptations of predators. In: GittlemanJL, ed. Carnivore behavior, ecology, and evolution, Vol. II.Ithaca, NY: Cornell University Press, 393–428.
Bookstein FL. 1997. Morphometric tools for landmarkdata: geometry and biology. Cambridge: Cambridge Univer-sity Press.
Burkart R, Bárbaro NO, Sánchez RO, Gómez DA. 1999.Eco-regiones de la Argentina. Administración de ParquesNacionales, PRODIA, 1–43.
Cabrera AL. 1958. Catálogo de los Mamíferos de América delSur. Revista del Museo Argentino de Ciencias Naturales‘Bernardino Rivadavia’. Buenos Aires: Instituto Nacional deInvestigación de las Ciencias Naturales.
Cabrera AL. 1971. Fitogeografía de la República Argentina.Boletín de la Sociedad Argentina de Botánica 16: 1–42.
Cabrera AL, Willink A. 1980. Biogeografía de AméricaLatina. Secretaría General de la Organización de los EstadosAmericanos. Programa Regional de Desarrollo Científico yTecnológico. Serie de Biología. Monografía N°13.
Cabrera AL, Yepes J. 1940. Historia Natural Ediar:Mamíferos Sud-Americanos: vida, costumbres y descripción.Buenos Aires: Cia. Argentina de Editores.
Canevari M, Vaccaro OB. 2007. Guía de mamíferos del surde América del Sur. Buenos Aires: Ed LOLA.
Catalano SA, Goloboff PA, Giannini NP. 2010. Phyloge-netic morphometrics (I): the use of landmark data in aphylogenetic framework. Cladistics 26: 11–20.
Chaves PB, Graeff VG, Lion ML, Oliveira LR, Eizirik E.2011. DNA barcoding meets molecular scatology: shortmtDNA sequences for standardized species assignment ofcarnivore noninvasive samples. Molecular Ecology Resources12: 18–35.
Chebez JC. 2009. Otros que se van. Especies argentinas enpeligro. Buenos Aires: Ed. Albatros.
Davis JC. 1986. Statistics and data analysis in geology, 2ndedn. New York: John Wiley and Sons.
Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L,Tablada M, Robledo CW. 2010. InfoStat versión 2010.Grupo InfoStat, FCA, Universidad Nacional de Córdoba,Argentina.
Díaz MM, Lucherini M. 2006. Orden Carnivora. In: BarquezRM, Díaz MM, Ojeda RA, eds. Mamíferos de Argentina.Sistemática y Distribución. Tucumán: Sociedad Argentinapara el Estudio de los Mamíferos, 89–107.
Dragoo JW, Bradley RD, Honeycutt RL, Templeton JW.1993. Phylogenetic relationships among the skunks: amolecular perspective. Journal of Mammalian Evolution 1:255–267.
Dragoo JW, Honeycutt RL. 1997. Systematics of mustelid-like carnivores. Journal of Mammalogy 78: 426–443.
Dragoo JW, Honeycutt RL, Schmidly DJ. 2003. Taxo-nomic status of white-backed hog-nosed skunks, genusConepatus (Carnivora: Mephitidae). Journal of Mammalogy84: 159–176.
Dragoo JW, Sheffield SR. 2009. Conepatus leuconotus.Mammalian Species 827: 1–8.
Eizirik E, Murphy WJ, Koepfli KP, Johnson WE, DragooJW, Wayne RK, O’Brien SJ. 2010. Pattern and timing ofdiversification of the mammalian order Carnivora inferredfrom multiple nuclear gene sequences. Molecular Phyloge-netics and Evolution 56: 49–63.
Emmons L, Helgen K. 2008. Conepatus chinga. In: IUCN,2011. IUCN Red List of Threatened Species. Version 2011.1.Available at: http://www.iucnredlist.org
Ewer RF. 1973. The carnivores. Ithaca, NY: Cornell Univer-sity Press.
Felsenstein J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39: 783–791.
Flynn JJ, Finarelli JA, Zehr S, Hsu J, Nedbal MA. 2005.Molecular phylogeny of the Carnivora (Mammalia): assess-ing the impact of increased sampling on resolving enigmaticrelationships. Systematic Biology 54: 317–337.
Forasiepi AM. 2003. Nuevo registro de Conepatus primaveus(Mammalia, Carnivora, Mustelidae) del Pleistoceno de laProvincia de Buenos Aires, Argentina. Revista del MuseoArgentino de Ciencias Naturales 5: 21–29.
Galliari CA, Pardiñas UFJ, Goin FJ. 1996. Lista comen-tada de los mamíferos argentinos. Mastozoología Neotropi-cal 3: 39–62.
García NO. 1990. Síntesis climatográfica de la RepúblicaArgentina. Publicación Facultad de Ingeniería y CienciasHidricas-Universidad Nacional del Litoral 36: 1–33.
Goloboff PA, Catalano SA. 2010. Phylogenetic morphomet-rics (II): algorithms for landmark optimization. Cladistics26: 1–10.
Goloboff PA, Farris JS, Nixon KC. 2008. TNT, a freeprogram for phylogenetic analysis. Cladistics 24: 774–786.
Goodall C. 1991. Procrustes methods in the statistical analy-sis of shape. Journal of the Royal Statistical Society 52:285–339.
342 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

Gray JE. 1837. Conepatus humboldtii. In: Charlesworth E,ed. The magazine of natural history, and journal of zoology,botany, mineralogy, geology and meteorology, Vol. I. London,581.
Hall T. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symposium Series 41: 95–98.
Hasegawa M, Kishino H, Yano T. 1985. Dating of thehuman-ape splitting by a molecular clock of mitochondrialDNA. Journal of Molecular Evolution 22: 160–174.
Helgen KM, Kays R, Helgen LE, Tsuchiya-Jerep MTN,Pinto CM, Koepfli KP, Eizikirk E, Maldonado JE.2009. Taxonomic boundaries and geographic distributionsrevealed by an integrative systematic overview of the moun-tain coatis, Nassuela (Carnivora: Procyonidae). Small Car-nivore Conservation 41: 65–74.
Honacki JH, Kinman KE, Koeppl JW. 1982. Mammalspecies of the world. Lawrence, KS: Allen Press andAssociates.
Huelsenbeck JP, Ronquist F. 2001. MrBayes: Bayesianinference of phylogenetic trees. Bioinformatics 17: 754–755.
Ivanova NV, Zemlak TS, Hanner RH, Hebert PDN. 2007.Universal primer cocktails for fish DNA barcoding. Molecu-lar Ecology Notes 7: 544–548.
Jobb G, von Haeseler A, Strimmer K. 2004. TreeFinder: apowerful graphical analysis environment for molecular phy-logenetics. BMC Evolutionary Biology 4: 18.
Kimura M. 1980. A simple method for estimating evolution-ary rates of base substitutions through comparative studiesof nucleotide sequences. Journal of Molecular Evolution 6:111–120.
Kipp VH. 1965. Beitrag zur Kenntnis der Gattung ConepatusMolina, 1782. Zeitschrift für Säugetierkunde 30: 193–232.
Klinberger CP. 2011. MorphoJ: an integrated softwarepackage for geometric morphometrics. Molecular EcologyResources 11: 353–357.
Kovarovic K, Aiello LC, Cardini A, Lockwood CA. 2011.Discriminant function analyses in archaeology: are classifi-cation rates too good to be true? Journal of ArchaeologicalScience 38: 3006–3018.
Legendre P, Legendre L. 1998. Numerical ecology, 2ndEnglish edn. Amsterdam: Elsevier.
McNab BK. 2010. Geographic and temporal correlations ofmammalian size reconsidered: a resource rule. Oecología164: 13–23.
Meier R, Zhang G, Ali F. 2008. The use of mean instead ofsmallest interspecific distances exaggerates the size of the‘Barcoding Gap’ and leads to misidentification. SystematicBiology 57: 809–813.
Milne-Edwards A, Vaillant L, Gervais P. 1891.Mission scientifique du Cap Horn 1882–1883. TomeVI. Zoologie. Première partie. Gauthier-Villars et. Fils.Imprimeurs- Libraires De L’Ecole Polytechnique, Du BureauDes Longitudes.
Nylander JAA. 2004. MrAIC.pl. Program distributed by theauthor. Evolutionary Biology Centre, Uppsala University.
Olson DM, Dinerstein E, Wikramanayake ED, BurgessND, Powell GVN, Underwood EC, D’Amico JA, Itoua I,
Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Rick-etts TH, Kura Y, Lamoreux JF, Wettengel WW, HedaoP, Kassem KR. 2001. Terrestrial ecoregions of the world: anew map of life on earth. Bioscience 51: 933–938.
Osgood WH. 1943. The mammals of Chile, Vol. 30, Zoologicalseries, field museum of natural history. Chicago, IL: FieldMuseum Press.
Parera A. 2002. Los mamíferos de Argentina y la regiónaustral de Sudamerica. Buenos Aires: Ateneo.
Poljak S, Confalonieri V, Fasanella M, Gabrielli M, Liz-arralde MS. 2010. Phylogeography of the armadillo Cha-etophractus villosus (Dasypodidae Xenarthra): post-glacialrange expansion from Pampas to Patagonia (Argentina).Molecular Phylogenetics and Evolution 55: 38–46.
Popowics TE. 2003. Postcanine dental form in the Musteli-dae and Viverridae (Carnivora: Mammalia). Journal of Mor-phology 256: 322–341.
Prevosti FJ, Pardiñas UFJ. in press. The heralds: carni-vores (Carnivora) and sigmodontine rodents (Cricetidae) inthe Great American Biotic Interchange. In: Rosenberger AL,Tejedor MF, eds. The Origins and Evolution of CenozoicSouth American Mammals. Vertebrate Paleobiology andPaleoanthropology Book Series. New York: AmericanMuseum of Natural History.
Prevosti FJ, Segura V, Martin GM, Schiaffini MI. 2011.Sobre el estado sistemático de Lycalopex griseus y Lycalopexgymnocercus: análisis de morfometría geométrica y modelosde distribución potencial. La Plata: XXIV Jornadas Argen-tinas de Mastozoología.
R Development Core Team. 2005. R: a language and envi-ronment for statistical computing. Vienna: R Foundation forStatistical Computing.
Redford KH, Eisenberg JF. 1992. Mammals of theNeotropics. The southern cone, Vol. 2, Chile, Argentina,Uruguay, Paraguay. Chicago, IL: The University of ChicagoPress.
Rohlf FJ. 1999. Shape statistics: procrustes super-impositions and tangent spaces. Journal of Classification16: 197–223.
Rohlf JF. 2003. TPSRelw, version 1.35. Ecology and Evolu-tion, SUNY at Stony Brook.
Rohlf JF. 2008a. TPSUtil, version 1.40. Ecology andEvolution, SUNY at Stony Brook.
Rohlf JF. 2008b. TPSDig2, version 2.12. Ecology and Evolu-tion, SUNY at Stony Brook.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19: 1572–1574.
Sato JJ, Wolsan M, Prevosti FJ, D’Elía G, Begg C, BeggK, Hosoda T, Campbell KL, Suzuki H. 2012. Evolution-ary and biogeographic history of weasel-like carnivorans(Musteloidea). Molecular Phylogenetics and Evolution 63:745–757.
Sato JJ, Wolsan M, Suzuki H, Hosoda T, Yamaguchi Y,Hiyama K, Kobayasi M, Minami S. 2006. Evidence fromnuclear DNA sequences sheds light on the phylogeneticrelationships of Pinnipedia: single origin with affinity toMusteloidea. Zoological Science 23: 125–146.
TAXONOMY OF SOUTHERN SOUTH AMERICAN SKUNKS 343
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344

Schiaffini MI, Prevosti JF, Bo R, Corriale MJ, Porini G,Lizarralde M. 2010. Morfometría craneana de Conepatus(Carnivora: Mephitidae) de Argentina: implicancias tax-onómicas. Bahía Blanca: XXIII Jornadas Argentinas deMastozoología.
Sheets HD. 2002. IMP-integrated morphometricspackage. Buffalo, NY: Department of Physics, CanisiusCollege.
Smith MF, Patton JL. 1993. The diversification of SouthAmerican murid rodents: evidence from mitochondrial DNAsequence data for the akodontine tribe. Biological Journalof the Linnean Society 50: 149–177.
Thomas O. 1920. Report on the Mammalia collected byMr. Edmund Heller during the Peruvian expedition of 1915under the auspices of Yale University and the NationalGeographic Society. Proceedings of the United StatesNational Museum 58: 217–249.
Tobe SS, Kitchener AC, Linacre AMT. 2010. Reconstruct-ing mammalian phylogenies: a detailed comparison of theCytochrome b and Cytochrome Oxidase Subunit I mitochon-drial genes. PLOS ONE 5: e14156.
Turnbull WD. 1970. Mammalian masticatory apparatus.Fieldiana, Geology 18: 147–356.
Van Gelder RG. 1968. The genus Conepatus (Mammalia,Mustelidae): variation within a population. AmericanMuseum Novitates 2322: 1–37.
Van Valkenburgh B. 1989. Carnivore dental adaptationsand diet: a study of trophic diversity within guilds. In:Gittleman JL, ed. Carnivore behavior, ecology, and evolu-tion. Ithaca, NY: Cornell University Press, 410–435.
Van Valkenburgh B. 2007. Déjà vu: the evolution of feedingmorphologies in the Carnivora. Integrative and ComparativeBiology 47: 147–163.
Wang X, Carranza-Castañeda O. 2008. Earliest hog-nosedskunk, Conepatus (Mephitidae, Carnivora), from the earlyPliocene of Guajanato, Mexico and origin of South Americanskunks. Zoological Journal of the Linnean Society 154:386–407.
Wilson DE, Mittermeier RA, eds. 2009. Handbook of themammals of the world, Vol. 1, Carnivores. Barcelona: Lynxeditions in association with Conservation International andIUCN.
Wozencraft WC. 2005. Order Carnivora. In: Wilson DE,Reeder DM, eds. Mammal species of the world. A taxonomicand geographic reference, Vol. 1, 3rd edn. Baltimore: TheJohns Hopkins University Press, 601–624.
Yepes J. 1936. Mamíferos coleccionados en la parte central yoccidental de la provincia de La Rioja. Physis 12: 31–42.
Zunino GE, Vaccaro OB, Canevari M, Gardner AL. 1995.Taxonomy of the genus Lycalopex (Carnivora: Canidae) inArgentina. Proceedings of the Biological Society of Washing-ton 108: 729–747.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article:
Table S1. Examined specimens from museum collections.Table S2. GenBank accession numbers or iBOL accession numbers (*), for nucleotide sequences used in thisstudy.Figures S1–S5. Alternative principal component analysis and discriminant function analysis using differentsystematic schemes and reduced landmark data sets.
344 M. I. SCHIAFFINI ET AL.
© 2013 The Linnean Society of London, Zoological Journal of the Linnean Society, 2013, 167, 327–344
Copyright © 2022 FDOKUMEN




















