
Temperate grassland Herbicide Conservation Species composition Forage quality

Taxonomic Composition and Temporal Organization of Tropical and Temperate SpeciesAssemblages of Lotic ChironomidaeAuthor(s): William P. Coffman and Carlos L. de la RosaReviewed work(s):Source: Journal of the Kansas Entomological Society, Vol. 71, No. 4, Special PublicationNumber 3: A Festschrift Honoring Mary and Jim Sublette. Part 2: Biology, Ecology andNatural History of Chironomidae (Oct., 1998), pp. 388-406Published by: Allen Press on behalf of Kansas (Central States) Entomological SocietyStable URL: http://www.jstor.org/stable/25085856 .
Accessed: 06/02/2013 11:59
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Kansas (Central States) Entomological Society and Allen Press are collaborating with JSTOR to digitize,preserve and extend access to Journal of the Kansas Entomological Society.
http://www.jstor.org
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY 71(4), 1998, pp. 388-406
Taxonomic Composition and Temporal Organization of Tropical and Temperate Species Assemblages of Lotic Chironomidae
William P. Coffman
Department of Biological Sciences and Pymatuning Laboratory of Ecology, University of Pittsburgh,
Pittsburgh, Pennsylvania 15260
AND
Carlos L. de la Rosa
Riverwoods Field Laboratory, 100 Riverwoods Circle,
Lorida, Florida 33857
abstract: General features of the taxonomic composition and temporal organization of
species assemblages of tropical and temperate lotic Chironomidae are described and com
pared. Seven studies of emergence phenology using pupal exuviae are included in the analy sis: four studies on three streams of northwestern Costa Rica, and three studies on two streams
in western Pennsylvania. Each of the seven studies was conducted for at least one year, but
all phenological analyses were restricted to one-year periods. The collection sites for all seven
studies were 2nd- to 3rd-order sections of the streams. Species richness of the Costa Rican
assemblages ranged from 142-151 for the one-year periods used in the phenological analy ses, and from 82-184 for the Pennsylvanian assemblages. Each assemblage was dominated
by members of the subfamilies Chironominae, Orthocladiinae and Tanypodinae. The aver
age number of species occurring in the tropical stream samples was about 1.8 times the num
ber in the temperate samples (45.9 and 26.1, respectively). The number of species emerging was moderately to greatly influenced by seasonal changes: dry-wet season shifts in the trop ical streams, and summer-winter shifts in the temperate streams. The number of days per year
during which the average species was emerging was determined for all seven assemblages by estimation and direct calculation methods. By both methods, the average Costa Rican species was found to be emerging for many more days (116.2 and 132.9 per year by the two methods) than the average Pennsylvanian species (70.0 per year by both methods). Change in taxo
nomic composition through the year was determined by calculating the change in similarity
(Sorensen Similarity Coefficient) for all combinations of pairs of samples. Pairs of temper ate stream samples were generally less similar the greater the time period separating them.
The mean change between compared samples of the three temperate studies ranged from 0.73
to 0.74. Although taxonomic composition of the tropical stream samples changed throughout the year, the degree of change between compared samples generally did not increase with
time. The mean change between compared samples for the four tropical studies ranged from
0.50 to 0.58. On the whole, the results of these studies are consistent with the hypothesis
(Coffman, 1989) that the life cycles of low-latitude stream species are much less seasonally
regulated than those of temperate latitudes, and that there may be different developmental
stages representing several generations of a species present at the same time.
Latitudinal patterns of taxonomic composition, species richness and phenology of
lotic assemblages of Chironomidae, to the extent that they exist, are not very well
documented. Our knowledge has been limited primarily by three factors: 1) the small
number of detailed taxonomic studies on extra-temperate streams; 2) the obscuration
of such patterns by the fact that other conditions [temperature, productivity, gradi ent, substrate, altitude, stream size (order), etc.] interact in a complicated fashion,
Accepted for publication 15 April 1998.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 389
often resulting in streams of the same latitude having very different taxonomic com
positions and/or levels of species richness; and 3) the very small number of extra
temperate studies which have examined phenology.
Considering many of the factors that may play a role in determining species rich
ness of lotic assemblages of Chironomidae, Coffman (1989) hypothesized that the
richness of low-latitude streams should be less than that of similar streams at mid
latitudes. It was argued that the lack of clear seasonality of temperature and food
types at low latitudes should result in the species assemblage of any one stream con
sisting of the set of chironomid species that was best adapted to these nearly per manent conditions, and that they should become asynchronous in their life histories.
Thus, it was suggested that "all species should be doing all things at all times". This
meant that each species should be represented at all times of the year by most, if not
all, larval instars and that some adult emergence should be taking place at all times.
Each of the instars of each of the species would occupy physical and trophic niche
space that in mid-latitude streams would be occupied by separate, and much more
synchronously developing, species. Therefore, it was predicted that given the finite
supply of niche resources and nearly constant conditions, the species assemblages of low-latitude streams should be smaller and less phenologically distinct than those
of mid-latitude streams. At the time, the only studies on low-latitude species rich
ness and phenology of low order stream assemblages were those of Lehmann (1979,
1981), who found relatively small numbers of species that were, for the most part,
continuously emerging throughout the year in two streams in Zaire. Since then, Fer
rington et al. (1993) have provided similar results for a small stream in Puerto Rico.
However, analyses of pupal exuviae collections from low-latitude streams (Costa Rica and West Africa) have revealed species richness values that are, in general, at
least as great as those of similar mid-latitude streams (Coffman et al., 1992). In this paper we attempt to reconcile the apparently conflicting evidence concern
ing chironomid species richness of low-latitude streams, and test the prediction of
sharply defined differences in emergence phenology between mid-latitude and low
latitude streams. Results of seven studies of species richness and annual phenologi cal patterns (four from Costa Rica and three from western Pennsylvania) will be pre sented to assess the generality of latitude-related phenomena. Since nearly 500
species were encountered in these studies, it is clearly not possible to treat each sep
arately in this paper. Instead, we will limit the comparisons to macrotaxonomic and
macrophenological patterns: taxonomic composition and temporal change in com
position at the subfamily and tribal levels, as well as to estimates of change in sim
ilarity indexes of species collected as a function of time.
Methods
Field procedures
Chironomid pupal exuviae were collected at intervals for at least one year in each of the seven studies, although the year of study, the number of samples, and the in
tervals between samples varied (Table 1). Natural accumulations of floating exuviae
from behind stream flow blocks, e.g., logs, were concentrated using a 0.25 micron sieve and preserved in 70% EtOH. The number of exuviae collected depended on
the size of the accumulated mass of debris, the number of specimens emerging in the 1-2 day period before collection, stream flow and temperature conditions (the
higher the temperature the more rapidly exuviae sink).
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

390 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Table 1. Time periods during which collections were made and the number of samples of exuviae taken;
where different, the one-year time period and number of samples used in phenological analyses.
Geographical area collection site Collection period Number of samples
Costa Rica
Q. Las Yeguitas, Maritza 12 Feb 1989-9 Feb 1990 16
Q. Las Yeguitas, Orosi 12 Feb 1989-9 Feb 1990 13
R. Tempisquito Sur 11 Feb 1989-9 Feb 1990 13
R. Tempisquito, Maritza 10 Feb 1989-9 Feb 1990 23
W. Pennsylvania Powdermill Run, Moul 2 Jan 1983-20 Dec 1983 21
Powder-mill Run, Hdqtrs. 1 Feb 1983-2 Jan 1984 19
Linesville Creek 4 Jan 1991-3 Jan 1992 42
The advantages offered by pupal exuviae are well documented and include: all
species emerge at the water surface, the exuviae remain floating for at least a day,
large numbers of specimens can be rapidly collected, species from all microhabitats
are included, and discrimination of species is usually more readily made than with
either larvae or adults.
Laboratory procedures
The species composition of each sample was determined by the examination of
all specimens with a dissecting microscope. The number of specimens varied from
a few hundred in dry season and winter samples to as many as 70,000 during peak
emergences. Most samples from all seven sites contained more than 1000 specimens.
Examples of each species from each sample were slide mounted in Euparal? to en
sure taxonomic consistency throughout the studies. The specimens are housed in the
Coffman collection at the University of Pittsburgh, Department of Biological Sci
ences. Essentially none of the Costa Rican types of exuviae and less than 50% of
those from western Pennsylvania could be assigned, with certainty, to a described
species. All of the species from western Pennsylvanian sites were referable to a
named genus. Fifty of the 250 species collected from the four Costa Rican sites could
not be placed in a known genus.
Analytical procedures
The length of the average emergence period (in days) for each of the temperate and tropical species assemblages was determined by Estimation and Calculation
Methods. In both cases only those samples taken within the one-year periods were
used. In the Estimation Method, it was assumed that the percent of the year's sam
ples in which a species occurred was proportional to the percent of the year (num ber of days) during which it was actually emerging. This is, obviously, a major as
sumption and its validity is influenced not only by the accuracy of each sample in
reflecting the emerging species, but by the variability of the intervals between sam
ples. This method offers the advantage, however, that it does not require decisions
about how to interpret temporal gaps in the emergence of species. In the Calculation
Method, the average emergence periods for the species of the seven assemblages were measured by direct determination. As long as a species emerged consistently over a series of samples this method was rather straightforward, and began with the
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 391
calculation of the number of days between the extreme samples of the period. Mak
ing the assumption that the actual emergence period of a species would have begun somewhat earlier and ended somewhat later than the days of the extreme samples, the emergence period was increased by half the number of days between the extreme
samples and the flanking samples in which the species did not occur. However, the
interpretation of gaps which occurred in the emergence records for many species (es
pecially in the Costa Rican studies) made the application of this method more prob lematical. Gaps could have resulted from: 1) a failure to detect the presence of the
species because of inadequate samples, or, 2) a real absence reflecting the period be tween generations. In each instance a decision had to be made as to which interpre tation was more likely to be correct. Several factors were considered in determining
whether a gap was long enough for the development of a new generation. Assum
ing, more or less, uniform growth rates, large species were considered to require a
longer gap than small ones. Because they can accumulate sufficient degree days in
approximately the same number of days at anytime of the year, tropical species were
considered to require, on average, a shorter gap than temperate ones. When more
than one generation was detected, the total emergence period for the species was
considered to be the sum of the emergence days for each of the generations. Since frequency data were not available for all of the studies, all analyses of tem
poral change in similarity of samples from a stream were made on the basis of pres ence or absence of a species. Comparisons were made between all combinations of
paired samples using the Sorensen Similarity Coefficient (2C/N, + N2) where C is the number of species in common and N, and N2 are the numbers of species in the two samples (Brown and Gibson, 1983). This coefficient is sensitive to differences
in both the number of shared species and those which occur in only one of the com
pared samples.
All statistical comparisons were made using the Mann-Whitney U-test.
Collection Sites
Costa Rica
The streams of this study [Quebrada (=creek) Las Yegtiitas at two sites, Maritza and Orosi; Rio Tempisquito at Maritza; and Rio Tempisquito Sur at Maritza] are
2nd-3rd order in size, close together (ranging from 10?57/N, 85?29'W to 10?57'N, 85?32'W), and drain the western slopes of Orosi Volcano in the Guanacaste National Park of northwestern Costa Rica. "Maritza" refers to the Maritza Biological Field
Station, owned and operated by the Guanacaste National Park. In general, all of the sites are quite similar in altitude, substrate and temperature, although discharge of
Q. Las Yegtiitas was considerably less than at the other sites. The collection sites on
Q. Las Yegtiitas at Maritza, R. Tempisquito and R. Tempisquito Sur were all at about 600 m. a. s. 1., of moderate to high gradient, and with cobble to boulder substrate. The site on Q. Las Yegtiitas at Orosi is at about 290 m. a. s. 1., has smaller cobble substrate and less gradient than the other sites. The temperature of these three streams varied little during the period of study, remaining close to 20?C throughout the year, but Q. Las Yegtiitas at Orosi was somewhat warmer. The two sites on Q. Las Yegtii tas were intermittent in flow, with only pools remaining during the periods of max imum flow reduction in the dry season (February to May). All sites were located in sections of the streams flowing through Mesoamerican dry forest and were all char
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

392 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Table 2. Subfamily and tribal composition of the four Costa Rican sites. Number of species collected
from all samples and per cent of overall species richness.
Quebrada Quebrada Rio Taxa Las Yeguitas, Las Yeguitas, Tempisquito Rio
at Maritza at Orosi Sur Tempisquito All sites
# % # % # % # % # %~
Chironominae 71 47.0 74 49.3 64 45.1 67 44.4 120 48.0
Chironomini 42 27.8 34 22.7 39 27.5 43 28.5 67 26.8
Tanytarsini 29 19.2 38 25.3 22 15.5 22 14.6 49 19.6
Pseudochironomini 0 0.0 2 1.3 3 2.1 2 1.3 4 1.6
Orthocladiinae 59 39.1 45 30.0 61 42.9 64 42.4 90 36.0
Corynoneurini 26 17.2 24 16.0 25 17.6 25 16.6 37 14.8
Other 33 21.9 21 14.0 36 25.3 39 25.8 53 21.2
Tanypodinae 21 13.9 31 20.7 17 12.0 20 13.2 40 16.0 Pentaneurini 17 11.3 28 18.7 15 10.6 18 11.9 6 14.4
Macropelopiini 4 2.6 3 2.0 2 1.4 2 1.3 4 1.6
# of species 151 150 142 151 250
acterized by well developed riparian forest. Additional data on these streams can be
found in Coffman et al. (1992) and de la Rosa (1992).
Western Pennsylvania
The three study sites differ in a number of respects. Linesville Creek (315 m. a. s. 1.;
41?39/N, 80?26/W) in northwestern Pennsylvania is a headwater stream of the
Shenango-Beaver-Ohio River system. The collection site is located in a 3rd order
woodland section (Wallace Woods) of the stream characterized by a rich glacial de
bris substrate of moderate cobble, gravel and sand, as well as significant quantities of wood and, seasonally, CPOM in the form of leaves and other plant parts. It is of
moderate to low gradient and has a typical annual temperature cycle from about
0.5?C to about 22-25?C. Both sites on Powdermill Run (40?39'N, 80?26/W) are lo
cated in the Powdermill Nature Reserve of the Carnegie Museum, about 60 miles
southeast of Pittsburgh in the Appalachians. Powdermill Run is a headwater stream
in the Loyalhanna-Kiskiminetas-Allegheny-Ohio River System. The Powdermill
Run site at Moul Spring (about 300 m above the confluence of the spring) is a 3rd
order moderate-high gradient section with boulder and cobble substrate intermin
gled with areas of bedrock, gravel and coarse sand. Although it flows through a heav
ily wooded area it has little wood in the stream and the significant autumn leaf fall
is rapidly flushed. At an elevation of 460 m. a. s. 1. it experiences summer tempera
tures somewhat below those of Linesville Creek. The Powdermill Run site at Head
quarters (409 m. a. s. 1.) is a 4th order, moderate-low gradient section of stream with
silty pools and cobble riffles. It typically has greater retention of CPOM but little
wood is retained in the study section. It has an annual temperature cycle similar to
that of Linesville Creek.
Results
Taxonomic composition of the species assemblages
The subfamily and tribal compositions of the four Costa Rican and three western
Pennsylvania sites are given in Tables 2 and 3, respectively.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions
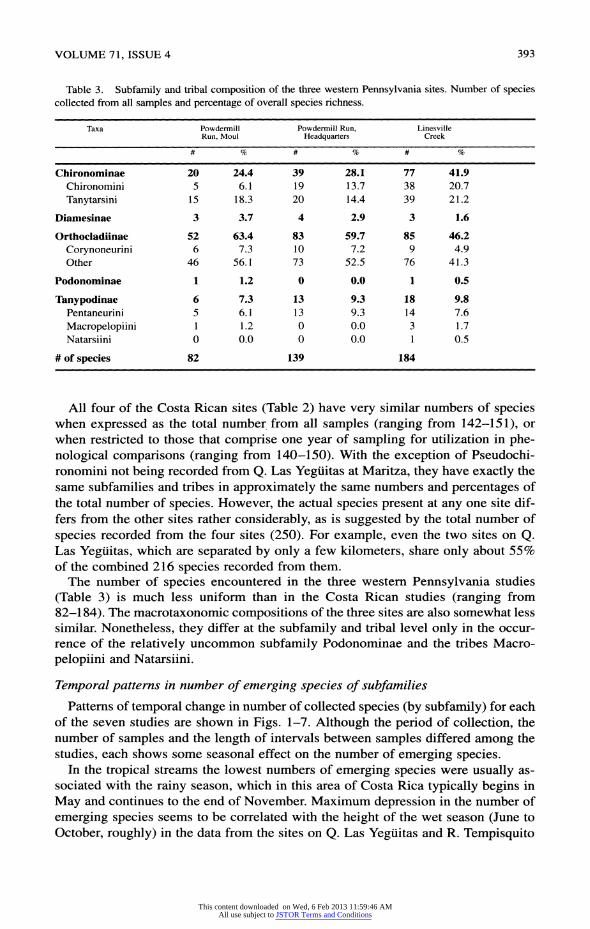
VOLUME 71, ISSUE 4 393
Table 3. Subfamily and tribal composition of the three western Pennsylvania sites. Number of species collected from all samples and percentage of overall species richness.
Taxa Powdermill Powdermill Run, Linesville Run, Moul Headquarters Creek
# % # % # %
Chironominae 20 24.4 39 28.1 77 41.9
Chironomini 5 6.1 19 13.7 38 20.7
Tanytarsini 15 18.3 20 14.4 39 21.2
Diamesinae 3 3.7 4 2.9 3 1.6
Orthocladiinae 52 63.4 83 59.7 85 46.2
Corynoneurini 6 7.3 10 7.2 9 4.9
Other 46 56.1 73 52.5 76 41.3
Podonominae 1 1.2 0 0.0 1 0.5
Tanypodinae 6 7.3 13 9.3 18 9.8
Pentaneurini 5 6.1 13 9.3 14 7.6
Macropelopiini 1 1.2 0 0.0 3 1.7
Natarsiini 0 0.0 0 0.0 1 0.5
# of species 82 139 184
All four of the Costa Rican sites (Table 2) have very similar numbers of species when expressed as the total number from all samples (ranging from 142-151), or
when restricted to those that comprise one year of sampling for utilization in phe
nological comparisons (ranging from 140-150). With the exception of Pseudochi
ronomini not being recorded from Q. Las Yegtiitas at Maritza, they have exactly the
same subfamilies and tribes in approximately the same numbers and percentages of
the total number of species. However, the actual species present at any one site dif
fers from the other sites rather considerably, as is suggested by the total number of
species recorded from the four sites (250). For example, even the two sites on Q. Las Yegtiitas, which are separated by only a few kilometers, share only about 55%
of the combined 216 species recorded from them. The number of species encountered in the three western Pennsylvania studies
(Table 3) is much less uniform than in the Costa Rican studies (ranging from
82-184). The macrotaxonomic compositions of the three sites are also somewhat less
similar. Nonetheless, they differ at the subfamily and tribal level only in the occur
rence of the relatively uncommon subfamily Podonominae and the tribes Macro
pelopiini and Natarsiini.
Temporal patterns in number of emerging species of subfamilies
Patterns of temporal change in number of collected species (by subfamily) for each of the seven studies are shown in Figs. 1-7. Although the period of collection, the
number of samples and the length of intervals between samples differed among the
studies, each shows some seasonal effect on the number of emerging species. In the tropical streams the lowest numbers of emerging species were usually as
sociated with the rainy season, which in this area of Costa Rica typically begins in
May and continues to the end of November. Maximum depression in the number of
emerging species seems to be correlated with the height of the wet season (June to
October, roughly) in the data from the sites on Q. Las Yegtiitas and R. Tempisquito
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions
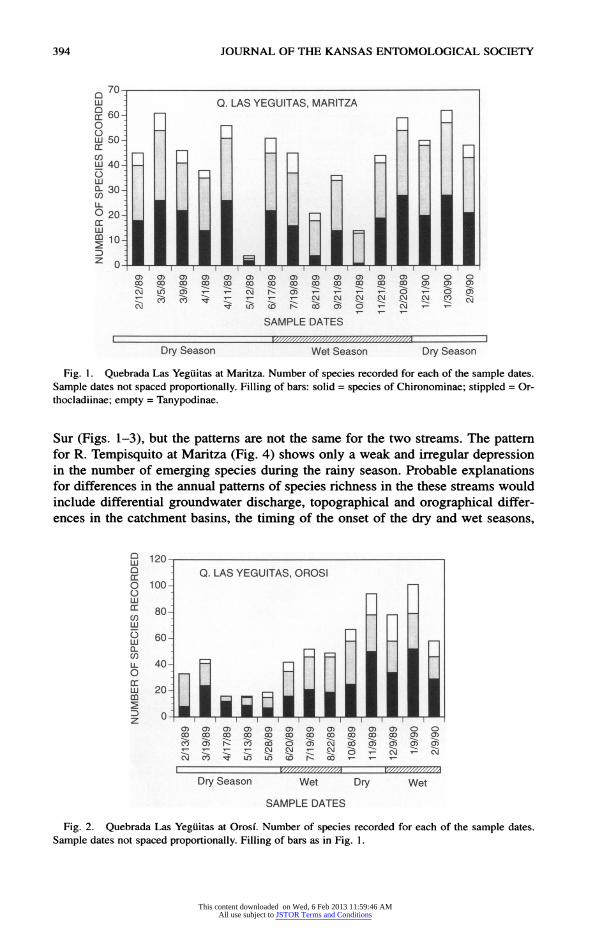
394 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Q701-1 w : Q. LAS YEGUITAS, MARITZA
O)g>g>g>a>a>o>a>o>o>g>o)0>ooo COaO0OCOQO0OaOQO0O0O0O0OCOg>g?gj>
CvIiOO^-t-CVI^OJt-t-^t-O^OP)
SAMPLE DATES
Dry Season Wet Season Dry Season
Fig. 1. Quebrada Las Yeguitas at Maritza. Number of species recorded for each of the sample dates.
Sample dates not spaced proportionally. Filling of bars: solid = species of Chironominae; stippled
= Or
thocladiinae; empty =
Tanypodinae.
Sur (Figs. 1-3), but the patterns are not the same for the two streams. The pattern for R. Tempisquito at Maritza (Fig. 4) shows only a weak and irregular depression in the number of emerging species during the rainy season. Probable explanations for differences in the annual patterns of species richness in the these streams would
include differential groundwater discharge, topographical and orographical differ ences in the catchment basins, the timing of the onset of the dry and wet seasons,
S 120T-1 g Q. LAS YEGUITAS, OROSI
O>0)0)0)0)0>0>0>0)0>0)00 ooooooooooaooooooo oo> oo ot ?>
Dry Season Wet Dry Wet
SAMPLE DATES
Fig. 2. Quebrada Las Yeguitas at Orosi. Number of species recorded for each of the sample dates.
Sample dates not spaced proportionally. Filling of bars as in Fig. 1.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 395
80 t-, Q
g 70 - f~l R|0 TEMPISQUITO SUR CC ; wm
O)O)O)O)O)O>O>O>0>&OOO ooaoaogogojom$ogogog)g)?>
SAMPLE DATES
Dry Season Wet Season Dry Season
Fig. 3. Rio Tempisquito Sur. Number of species recorded for each of the sample dates. Sample dates
not spaced proportionally. Filling of bars as in Fig. 1.
80 t-1
S 70- R- Tempisquito, Maritza |~|
? s
0>G>0)G>0>0)G>0>0>0>ODG>0)a>G>G)Q>C)0>0000
SAMPLE DATES
I- V/MMMMMM/M^^^^^ -1
Dry Season Wet Season Dry Season
Fig. 4. Rio Tempisquito at Maritza. Number of species recorded for each of the sample dates. Sam
ple dates not spaced proportionally. Filling of bars as in Fig. 1.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

396 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
g 35i-"S
g 301 POWDERMILL RUN _ _ _
8 M0UL ny fl
ll I I ml I I i? ill111 mi 11.1 ? 5 lllll.l 1111 111! i
o18,.|l|l|;|j||MI|l||i;,J I s i| i | i i ||11111 Is 1111 SAMPLE DATES
Fig. 5. Powdermill Run at Moul. Number of species recorded for each of the sample dates. Sample dates not spaced proportionally. Filling of bars: diagonal lines = Podonominae; crosshatched = Diamesinae
?otherwise, as in Fig. 1.
the sporadic occurrence of highly localized dry season rains and wet season dry pe riods, and the fact that collections were possible by slamming from standing pools at periods of no flow. More clearly shown by these data is the increase in number of
emerging species during the dry season, which usually commences in December. For
the most part, each of the three subfamilies that occurred in the tropical studies (Chi
ronominae, Orthocladiinae and Tanypodinae) were represented by some species in
all samples. The actual number varied as the total number of species changed, but
the relative proportion of the emerging species belonging to each of the subfamilies
remained remarkably constant.
aOn-, uj POWDERMILL RUN
g 70-i HEADQUARTERS W
^ 5 ? S ? ? S ? '? S ? ? ? S 5 ? ? I: ? S S 8 5 5- ? 5- .5 S 5 5- S ? 2 2 S ^ S? * *
SAMPLE DATES
Fig. 6. Powdermill Run at Headquarters. Number of species recorded for each of the sample dates.
Sample dates not spaced proportionally. Filling of bars: crosshatched = Diamesinae?otherwise, as in
Fig. 1.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions
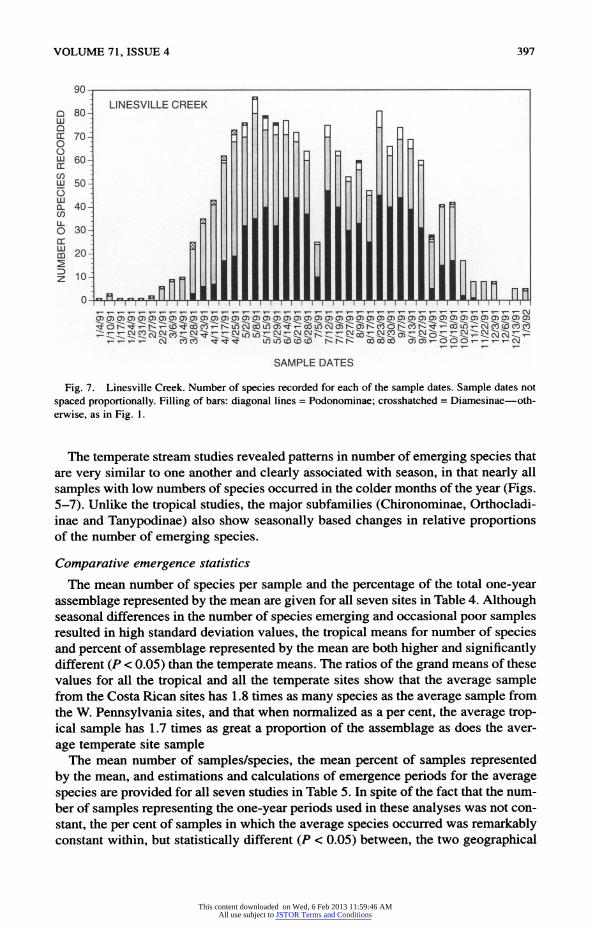
VOLUME 71, ISSUE 4 397
901-1 LINESVILLE CREEK H Q 80- J_ n
o?o bD BBh B LBq o i IM n Ip DIM
8 so- ; 1 o : ''-' i
a. 40-1 |- . .
11
<? : ni I i ' '
! m i : I- I' i ill
| 20- 11 f [
< . ;. , | | | > . ' I
111 | 10- JI | I.
: | I I i
||I|
lllflpifip SAMPLE DATES
Fig. 7. Linesville Creek. Number of species recorded for each of the sample dates. Sample dates not
spaced proportionally. Filling of bars: diagonal lines = Podonominae; crosshatched = Diamesinae?oth
erwise, as in Fig. 1.
The temperate stream studies revealed patterns in number of emerging species that
are very similar to one another and clearly associated with season, in that nearly all
samples with low numbers of species occurred in the colder months of the year (Figs.
5-7). Unlike the tropical studies, the major subfamilies (Chironominae, Orthocladi
inae and Tanypodinae) also show seasonally based changes in relative proportions of the number of emerging species.
Comparative emergence statistics
The mean number of species per sample and the percentage of the total one-year
assemblage represented by the mean are given for all seven sites in Table 4. Although seasonal differences in the number of species emerging and occasional poor samples resulted in high standard deviation values, the tropical means for number of species and percent of assemblage represented by the mean are both higher and significantly different (P < 0.05) than the temperate means. The ratios of the grand means of these
values for all the tropical and all the temperate sites show that the average sample from the Costa Rican sites has 1.8 times as many species as the average sample from
the W. Pennsylvania sites, and that when normalized as a per cent, the average trop ical sample has 1.7 times as great a proportion of the assemblage as does the aver
age temperate site sample The mean number of samples/species, the mean percent of samples represented
by the mean, and estimations and calculations of emergence periods for the average
species are provided for all seven studies in Table 5. In spite of the fact that the num
ber of samples representing the one-year periods used in these analyses was not con
stant, the per cent of samples in which the average species occurred was remarkably constant within, but statistically different (P < 0.05) between, the two geographical
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

398 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Table 4. Mean number of species per sample, and the percentage of the total species assemblage repre sented by the mean for all collections.
Geographical area Mean number of species SD Mean
Costa Rica
Q. Las Yeguitas, Maritza 41.4 ?18.0 27.4
Q. Las Yeguitas, Orosi 51.5 ?27.9 34.3
R. Tempisquito Sur 45.2 ?15.6 31.8
R. Tempisquito, Maritza 45.4 ?15.9 30.7
grand mean = 45.9 grand mean = 31.7
W. Pennsylvania Powdermill Run, Moul 15.9 ?11.3 19.3
Powdermill Run, Hdqtrs. 23.4 ?19.7 16.9
Linesville Creek 39.0 ?30.5 21.2
grand mean = 26.1 grand mean =19.1
ratio of grand ratio of grand means = 1.8 means = 1.7
areas. The ratio of the grand means for the two areas shows that the average species
from the Costa Rican studies was found in 1.7 times the per cent of all samples as
was the average temperate site species. As a result of this difference, the emergence
period in days using the Estimation Method was found to be 1.7 times longer for the
average Costa Rican site species (0.318 of all samples x 365 days =116.2 days) than
that of the Pennsylvania site species (0.192 of all samples x 365 days = 70.0 days).
The determination of the average emergence period using the Calculation Method
shows greater variation within, yet statistically significant (P < 0.05) values between, the two geographical areas. By this method, the average emergence periods of the
tropical and temperate species were found to be 132.9 days and 70.0 days, respec
tively. The ratio of the grand means determined by the Calculation Method suggests that the average emergence period of the Costa Rican species is 1.9 times as long as
its Pennsylvanian counterpart.
Temporal change in similarity of the samples
Change in the taxonomic composition of the collections, as a function of time, was
measured by graphing the value of the Sorensen similarity coefficient versus the
number of days separating the collection of the two samples compared. To do this, each of the samples was defined, in turn, as identity (a value of 1) and compared
with all others. The difference between any pair of samples was calculated as 1 mi
nus the similarity value. A difference of 0 between a pair of samples means that they have an identical taxonomic composition; a difference of 1 means that they have no
species in common. This procedure was repeated for all pairs of samples for each of
the seven studies. When the differences between all paired samples of a study are
plotted as a function of the time interval (in days) between them, it provides a com
prehensive summary of the degree to which the taxonomic composition of the emerg
ing species varies as a function of time (Figs. 8 and 9). All four of the tropical studies (Fig. 8) show a common relationship. The rate of
change is, on average, about the same throughout the year. The degree of change be
tween any two samples seems to be largely independent of the actual number of days
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 399
Table 5. Mean number and percentage of sample dates in which species occurred; estimation and cal
culation of the emergence period in days for the average species.
Mean number Mean percentage Mean emergence Mean emergence Geographical area of sample dates sample dates/species period in days period in days
Costa Rica
Q. Las Yegtiitas, Maritza 4.8 30.0 109.5 116.9
Q. Las Yeguitas, Orosi 4.5 34.6 126.3 145.2
R. Tempisquito Sur 4.2 32.3 117.9 115.8
R. Tempisquito, Maritza 7.0 30.4 111.0 153.8
grand grand grand mean = 31.8 mean = 116.2 mean = 132.9
W. Pennsylvania Powdermill Run, Moul 4.1 19.5 71.2 59.8
Powdermill Run, Hdqtrs. 3.2 16.8 61.3 55.6
Linesville Creek 8.9 21.2 77.4 94.6
grand grand grand mean = 19.2 mean = 70.0 mean = 70.0
ratio of grand ratio of grand ratio of grand means = 1.7 means =1.7 means = 1.9
which separated them. The mean change in the similarity coefficient for all pairs of
samples for these four studies ranges from 0.50-0.58. There is considerable scatter
of the points as the result of two factors: 1) the analysis includes all samples, even
those which were obviously inadequate in size to reliably include all emerging spe cies, and 2) there is a tendency for samples taken very close together in time to be
more similar to one another than are two samples taken at very different times of the
year. There also seems to be a weak tendency for differences to maximize at about
six months, and to diminish as the time interval approaches 365 days. The distributions of the points in the plots of the three temperate studies (Fig. 9)
are similar, but differ greatly from those of the tropical streams. In these plots the differences between samples are, for the most part, a function of the time intervals
between them. The means for the three temperate studies range from 0.73-0.74.
Discussion
Species richness of chironomid assemblages of low-latitude streams
As noted in the introduction, there appears to be conflicting evidence about the
species richness of chironomid assemblages of low-order streams at low latitudes.
Three studies (Lehmann, 1979, 1981; Ferrington et al., 1993) have determined that the number of species in assemblages of small streams of Zaire and Puerto Rico is
relatively low (ranging from 30-46 species) in comparison to temperate streams
which often have more than 100 species (Coffman, 1989). However, results pre sented for Costa Rican streams by Coffman et al. (1992), and more completely in
this paper, demonstrate that species assemblages at low-latitudes may be much richer than have been previously reported: extending well into the range of the richest
known temperate assemblages.
Although it is not possible at the present time to consider every factor that may
play a role in determining the richness of a particular assemblage and how the
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions
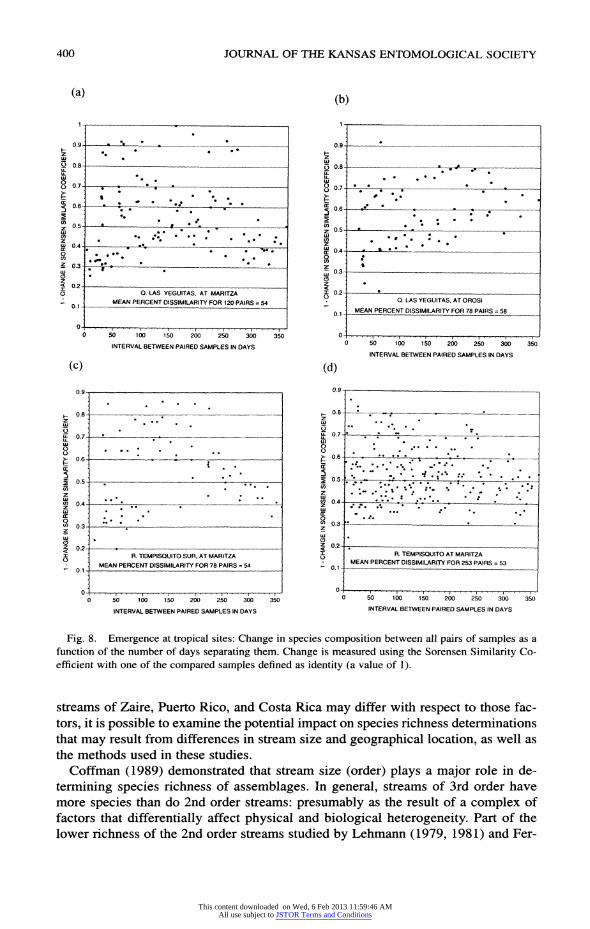
400 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
<a> (b)
1-j-.- H
0.9--?^-!_._-_ 0.9--! ? ? uj ui O 0.8-O 0.8-??r^--? U. J7 _
!? g 0.7-5-.-*?.- -
8 0.7--*. *
.-,-'--5- -
| o,-__-*_-:-*l .' -
| o,-_v^-.-^_^-_ ? * . 5
5 ?:? -.'.'.--' | 0.5?-_!-__-__,_ 1 . ':. .. ;. |
. : . .*
? ?-4-;-^----5-. %.. | o.4--. ? ? * '
2 o.3--^V-*-!_!_
_ 2 ft? _ _ ui * T^
5 0.3 2 ? < 0.2-- < .
O Q. LAS YEGUITAS, AT MARITZA g 0.2-?
J- MEAN PERCENT DISSIMILARITY FOR 120 PAIRS = 54 ^ Q. LAS YEGUITAS, AT OROSI
01"- Q 1 MEAN PERCENT DISSIMILARITY FOR 78 PAIRS =
58_
0) ....,...., - ,.-,., , ,., J 0 50 100 150 200 250 300 350 0 \.i .i . i . i.i.i '
0 50 100 150 200 250 300 350 INTERVAL BETWEEN PAIRED SAMPLES IN DAYS INTERVAL BETWEEN PAIRED SAMPLES IN DAYS
(c) (d)
0.9-j-, ?-9r-1
_ 0.8.?!-1_J-:- 2 o.8?: . ..,-.-s
2 *_ - UI *.* .
8 0.7-??_-_^- It ?-7"-T-*-~-=-r-% u. . Ul . W O .. '.. O . . * . O , .
t ?:-???i1- i
"' ?. ."
' -' .' /. .-^-^?
| ?:-,-=-^^- ?5-:. .: ; ^/ .'^ ,; , /. :. 2 . ui .!"..
* . ui * . co 04-* * * . .* . * * ._;_. <5 0.4-'?i-i-5--, z u*
^yn ^ ^ r r z . ui ... . . UI . cc - . * DC . O
Q .. . co 0.3 - --s_:_ co 0.3-. - z S w . ui . . O O Z 0.2--;
_ : ' R.TEMP.SQU.TO SUR. AT MARITZA 5 R. TEMPISQUITO AT MARITZA
V MEAN PERCENT D.SS.M.LAR.TY FOR 78 PAIRS = 54 ^ 0 , MEA" PERCE"T P'SS>M.LAR,TY FOR 253 PAIRS - 53 - o.1 -\-_____-___
p..,.,.,.,, , I o| ,., ,. 0 50 100 150 200 250 300 350 ? 50 '? 250 300 350
.NTERVAL BETWEEN PAIRED SAMPLES IN DAYS ,NTERVAL BETWEEN PA,RED SAMPLES ,N DAYS
Fig. 8. Emergence at tropical sites: Change in species composition between all pairs of samples as a
function of the number of days separating them. Change is measured using the Sorensen Similarity Co
efficient with one of the compared samples defined as identity (a value of 1).
streams of Zaire, Puerto Rico, and Costa Rica may differ with respect to those fac
tors, it is possible to examine the potential impact on species richness determinations
that may result from differences in stream size and geographical location, as well as
the methods used in these studies.
Coffman (1989) demonstrated that stream size (order) plays a major role in de
termining species richness of assemblages. In general, streams of 3rd order have more species than do 2nd order streams: presumably as the result of a complex of
factors that differentially affect physical and biological heterogeneity. Part of the
lower richness of the 2nd order streams studied by Lehmann (1979, 1981) and Fer
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 401
(a) (b)
1-|- - . .... ? ..?-?.? ..?.- -1 1-T-.---~?M~???-.-1
- " "
v .-**. *
0.9--+??*-^-.- ^ 0.9-- U / /-2
0 08-_M>^?_.. ' %_ O 0.8- *
r. ..-. St
' l
* ? '
! ' " '
in } ui * . n SI S 0.7-as-v? - O 0.7-i?^-^-^^-.
? ;n:-?'* -* I I *..' "
. .
1 V 1 .
s Q.5...y, .-y._j_ s ?-5:k-^?!-1 s . . g .
! z ? z . uj
0.4--^- aj o.4
8 *
8 r '
? 0.3-fc- 2 0.3-j-' g POWDERMILL RUN AT MOUL g I POWDERMILL RUN AT HEADQUARTERS
Z Q 2 MEAN PERCENT DISSIMILARITY FOR 190 PAIRS = 73 i 0 2
MEAN PERCENT DISSIMILARITY FOR 152 PAIRS ? 74
O O '| *"
0.1-- "
0.1-1 -'
0} ,.! !., , J 0 I.,...,, J 0 50 100 150 200 250 300 350 0 50 100 150 200 250 300 350
INTERVAL BETWEEN PAIRED SAMPLES IN DAYS INTERVAL BETWEEN PAIRED SAMPLES IN DAYS
(C)
1-,-. . | ,..nw,t...y."-\M|r
s ::?... .*-. V* ,.-- %
. . O 0.8--... '
?-rr=- -^??? -
ui. . . 8 0.7J .?",:;?':. ?
* . -v.?s?2?
g ;? .-..>'.....?. - '
~
? o.6: v.<-. .- *
.. ;-?
I 0.5-1,,
....*-.-^i . y-:
< 0.2-11 ,...-^-1 5 IJ
LINESVILLE CREEK ,1 I
* MEAN PERCENT DISSIMILARITY FOR 861 PAIRS = 73
0.1--_
0 I'""- ! !., , ,.I 0 50 100 150 200 250 300 350
INTERVAL BETWEEN PAIRED SAMPLES IN DAYS
Fig. 9. Emergence at temperate sites: Description as in Fig. 8.
rington et al. (1993) when compared with the mostly 3rd order Costa Rican streams
may be attributable to this factor.
Species richness of a particular assemblage may also be related to the size and
ecological diversity of the pool of species in the geographical area. It has been noted
(Ashe et al., 1987) that the number of species in the Afrotropical Region is particu
larly small, although Coffman (1992) recorded a large number of species from
streams of West Africa. Insularity may also play a major role in determining the size
of species pools, and therefore, the richness of particular assemblages. Part of the
lower richness of the streams of Zaire and Puerto Rico may be the result of smaller
pools of species from which members of assemblages may be drawn.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

402 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
The use of pupal exuviae in recognizing species and in determining species rich
ness is generally conceded to be more accurate than the use of adults. Morphologi cal differences appear, on average, to be greater between congeneric pupal exuviae
than between their corresponding adults, or, if not greater, more readily discernable.
This may be particularly important in tropical streams where many genera are rep resented by a large number of very similar sympatric species (Coffman et al., 1992).
The greater species richness of the Costa Rican streams when compared to the Cen
tral African streams studied by Lehmann (1979, 1981), who based most identifica
tions on adults, and the Puerto Rican stream studied by Ferrington et al. (1993), who
based all identifications on adults, may be partly responsible for the differences in
the number of species. The ability to capture and process much greater numbers of
pupal exuviae increases the probability that uncommon, and even rare, species will
be included. The use of emergence traps that collect adults from a relatively homo
geneous small area may also limit the number of species in comparison to the sam
pling of large accumulations of pupal exuviae which include species from all mi
crohabitats.
Whatever the reason(s) for the different numbers of species reported from tropi cal streams, it seems that contrary to Coffman's (1989) prediction, low-latitude, 2nd-3rd order, continental, biologically controlled streams do not have consistently lower numbers of species of Chironomidae than do similar temperate streams. Does
it follow, then, that the hypothesis of less synchronization of life history events of
chironomids of low-latitude streams should also be incorrect?
Comparative temporal organization of temperate and tropical lotic assemblages
of Chironomidae
The number of species in temperate assemblages of Chironomidae appears to re
flect saturation, or near saturation, of available niche space, here considered to have
two principle components: an instantaneous component equivalent to the spatial and
trophic heterogeneity available for partitioning at a point in time, and a temporal
component, which reflects change in food resource heterogeneity by allowing for se
quential occurrence of species with different trophic specializations. Supporting ev
idence for this view of the organization of temperate assemblages is both inferential
and empirical. Since the species that comprise an assemblage are drawn from a larger number of available species (the regional pool), each with its particular set of toler
ances and preferences with respect to physical and biotic conditions, it seems likely that, at least, the most abundant and persistent members of an assemblage will be
those that can coexist with a minimal amount of overlap. Empirical evidence comes
from partitioning studies of larvae of assemblages at Powdermill Run at Moul (Coff
man, unpubl.) and Linesville Creek (de la Rosa, 1985). When each species was char
acterized as to trophic role and distribution in time and space (substrate type), it was
found that less than 1% of all species pairs could possibly be in competition. All
other pairs were ecologically isolated from one another by one or more partitioning elements: time, food and substrate.
The interaction of life history patterns and seasonal temperature change also plays a major role in the organization of temperate assemblages. The temporal sequencing of species is not only the result of the availability of preferred substrates and food,
but reflects the fact that the seasonal thermal pattern allows for the acquisition of
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 403
sufficient degree days for life cycle development only at some times of the year. The
interrelationships of the seasonal availability of food resources and thermal change are built into the life histories of chironomid species and determines the number of
generations per year, as well as whether or not a species will diapause and if so, at
what time of the year. To make matters more complicated, it would appear (Coff man, unpubl.) that the larger the species the longer the developmental time (the more
degree days required) and the smaller the number of generations per year. The in
terrelationship of these factors characteristically produces an assemblage in which
the species have major life history events, e.g., rapid growth of 3rd-4th instars and
emergence, restricted to predictable times of the year. Emergence data from
Linesville Creek suggest that, in general, individual species emerge at approximately the same time each year (Coffman, unpubl.). The pronounced seasonal differences
in the number of species emerging from the temperate streams (Figs. 5-7) and the
increasing rate of change in the Sorensen Similarity Coefficient as a function of the number of days between samples (Fig. 9) are to be expected if the view of temper ate assemblages proposed above is correct.
Interpreting the points of the above discussion as postulates concerning the orga nization of chironomid assemblages of small temperate streams, we now consider to
what extent they may hold true for assemblages of small tropical streams. On the ba sis of the results presented by Coffman et al. (1992) and in this paper, there seems
to be no reason to argue that suitable chironomid niche space in tropical streams is
any less species saturated than that of temperate streams, although that may be
achieved in a different manner. The regional pool of species from which members of the assemblages were drawn is at least as large as that available for the temper ate streams. Reiss (1982) speculates that the Central American region may have
1500-2000 chironomid species. In a survey of Costa Rica, Watson and Heyn (1992) found at least 148 species, many belonging to genera not included among the 250
species encountered in the four studies reported here. That the assemblages were
drawn from a larger regional pool is clearly seen in the fact that no more than 151 of the 250 species were found at any one site. Saturation may also be inferred from the observations that all four sites, which were similar in many aspects of hetero
geneity, had essentially the same number of species, ranging from 142 to 151. What is obviously different about these streams is that the instantaneous compo
nent of heterogeneity (spatial and trophic niche space) varies much less as a func tion of the seasonal component of heterogeneity (sequence of food resources) and annual thermal pattern. It was this view of tropical assemblages that led to the hy pothesis that, unlike temperate streams, the conditions suitable for species would be
persistent throughout the year and that consecutive generations would be produced without diapause (Coffman, 1989). Two features of consecutive generations of spe cies that develop without diapause, or any type of temporal constraint, could very
rapidly lead to a chaotic temporal organization of tropical assemblages, contrasting greatly with the strict sequencing of species in temperate assemblages. 1) The fact that perfect synchrony is rarely achievable (for both genetic and environmental in fluence reasons) would mean that each generation would have some individuals
emerging earlier and some later than, perhaps the majority, of individuals. Egg lay ing by these individuals and subsequent hatching of the eggs, larval growth, meta
morphosis and emergence of the adults would then be out of phase with respect to
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

404 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
the "norm". Depending on the original degree of synchrony the number of genera tions required to produce the condition in which "all species would be doing all
things at all times" might be very small. This type of intergenerational time creep is
prevented in temperate streams by the changing conditions which usually prevent immediate development of the next generation. 2) Even if perfect synchrony were
to be maintained generation after generation, unless the period of development were
a factor of 365 days, without diapause and some form of time cue ("zeitgeber"), the
emergence times of succeeding generations would continue to be advanced or re
tarded relative to the calendar. The results of the phenological analyses for the Costa
Rican sites indicate that, although total chaos in temporal organization does not seem
to be the case, the trend relative to the temperate studies is definitely in that direc
tion. Compared to the results from the temperate streams, the higher average num
ber of species per sample, the longer periods of emergence and less change in sim
ilarity of samples as a function of time, suggest that the tropical assemblages are
somewhat less tightly constrained on the time axis.
Departure from the extreme condition of "all species doing all things at all times"
is most likely due to the fact that these streams are not completely aseasonal, as is
shown by the dry and wet season patterns of depression and enhancement of the
number of species emerging. To a great extent, the average emergence period was
kept relatively low in these studies by the large number of species that occurred only once or twice. If their rarity in the collections was simply a matter of inadequate
sampling, it is possible that the actual mean emergence period for these assemblages is much longer than that measured.
General ecological implications
Both theoretical and practical implications can be drawn from the results of these
studies of chironomid assemblages. The results presented in this paper demonstrate
that the number of species emerging from tropical streams at a point in time is at
least as great, and on average greater, than the number emerging from similar tem
perate streams. They also strongly suggest that the temporal organization of chi
ronomid life histories in tropical streams is more chaotic than in temperate streams, and that as a result several, if not all, instars of many, if not most, species are pre
sent at most times. The combination of many species and multiple instars of over
lapping generations of those species produces a large number of distinct "ecological entities" that must be able to pack available niche space. In general, that number
would appear to be higher than that of temperate streams. There are at least five ma
jor possibilities that would allow available niche space to be filled by a larger num
ber of "ecological entities" of Chironomidae. 1) There is more available niche space in tropical streams than in temperate streams. For this to be true physical hetero
geneity and/or trophic heterogeneity would have to be greater in tropical streams.
There seems to be no reason to suggest that the diversity of substrates (microhabi
tats) is any greater since the action of flowing water on geological materials is not
latitude dependent. If anything, the trophic diversity of small tropical streams with
a closed canopy, such as the streams of this report, should have less autochthonous
production and, perhaps, less, certainly less synchronous, allochthonous inputs than
similar temperate streams. 2) Niche space is more finely divided in tropical streams.
The large number of sympatric congeners in the Costa Rican assemblages suggests that their coexistence may be the result of specializations. Until we know more about
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

VOLUME 71, ISSUE 4 405
the distribution and trophic relationships of the larvae, this must be considered a dis tinct possibility. 3) Members of tropical stream assemblages are more tolerant of po tential competitors. This would most easily be facilitated by average low densities of the "entities". There is some evidence in support of this possibility. John Jackson
(Stroud Water Research Center, pers. comm.) reports that chironomid densities in
these streams rarely reach 5000 m2. Densities many times that level are typical of
temperate streams. The question arises, of course, as to why some species don't se
quester a larger share of the limited resources, increase in density, and force less
competitive species out of the assemblage. We simply don't know enough about trop ical streams to address this possibility in definite terms, but such exclusions may be,
perhaps, less likely in a partially chaotic system in which the list of potential com
petitors of a species is changing from generation to generation as the result of gen erational creep. 4) Predation plays a more significant role in influencing species rich ness in the tropical streams. To our knowledge there is no evidence that this is the case. 5) There is less competition from other groups of benthic organisms in the trop ical streams. Until we know more about the overall organization and function of the
other components of the benthos in both tropical and temperate streams, and how
they may interact with the chironomids, it is not possible to usefully speculate on
this possibility. The data presented here suggest that the determination of total species richness of
both temperate and tropical streams requires samples from all periods of the year, but that on average, a single sample from a tropical stream will contain a greater pro
portion of the assemblage. However, because of less rigid seasonal patterns of oc currence and life cycle development, the taxonomic composition of single samples from tropical streams will be less reliable at predicting the composition of samples taken in subsequent years at the same time. The implication is that single year stud ies of larval populations of tropical stream chironomids will provide a less compre hensive view of temporally related ecological interactions of the assemblage than studies of the same length in temperate streams.
Acknowledgments
The collection and preliminary sorting of the Costa Rican samples was carried out with the assistance of the personnel of the Maritza Biological Field Station. We would like to thank the staff of the Guanacaste National Park for logistical and ad ministrative support for the junior author during his stay at the Maritza Biological Station. Thanks are also due to the organizations providing support for scientific proj ects at the Maritza station: the National Science Foundation (grant BSR-9007845), the Stroud Foundation, the Pennswood No. 2 Research Endowment, and the Francis
Boyer Research Endowment. We would also like to thank Dr. Joseph Merritt, Di rector of the Powdermill Nature Reserve of Carnegie Museum for allowing access to the sites on Powdermill Run. Louis Yurasits collected many of the samples from
Linesville Creek and provided advice as to the handling of data. Dr. Frederick Gottlieb
helped solve a number of computer problems and read sections of the manuscript. Dr. John Jackson of the Stroud Water Research Center provided information about chironomid larval densities in the Costa Rican streams. The Department of Biolog ical Sciences of the University of Pittsburgh provided facilities, both on campus and at the Pymatuning Laboratory of Ecology.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions

406 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Literature Cited
Ashe, P., D. A. Murray, and F. Reiss. 1987. The zoogeographical distribution of Chironomidae (Insecta:
Diptera). Annales de Limnologie 23:27-60.
Brown, J. H., and A. C. Gibson. 1983. Biogeography. C. V. Mosby, St. Louis.
Coffman, W. P. 1989. Factors that determine the species richness of lotic communities of Chironomidae.
Acta Biologica Debrecina, Supplementum Oecologica Hungarica 3:95-100.
Coffman, W. P., C. de la Rosa, K. W. Cummins, and M. A. Wilzbach. 1992. Species richness in some
Neotropical (Costa Rica) and Afrotropical (West Africa) lotic communities of Chironomidae
(Diptera). Netherlands Journal of Aquatic Ecology 26:229-237.
de la Rosa, C. 1985. Resource utilization by Chironomidae (Diptera) in a woodland stream ecosystem, Linesville Creek, Crawford County, Pennsylvania. Ph.D. Dissertation. University of Pittsburgh,
Pittsburgh, PA.
de la Rosa, C. 1992. Phoretic associations of Chironomidae (Diptera) on Corydalidae (Megaloptera) in
northwestern Costa Rican streams. Journal of the North American Benthological Society 11:316-323.
Ferrington, Jr., L. C, K. M. Buzby, and E. C. Masteller. 1993. Composition and temporal abundance of
Chironomidae emergence from a tropical rainforest stream at El Verde, Puerto Rico. Journal of the
Kansas Entomological Society 66:167-180.
Lehmann, J. 1979. Chironomidae (Diptera) aus FlieBgewassern Zentralafrikas. Teil I: Kivu-Gebiet,
Ostzaire. Spixiana, Supplement 3:1-144.
Lehmann, J. 1981. Chironomidae (Diptera) aus FlieBgewassern Zentralafrikas. Teil II: Die Region um
Kisangami, Zentralzaire. Spixiana, Supplement 5:1-85.
Reiss, F. 1992. Chironomidae. In: S. H. Hubert and A. Villalobous-Figueroa (eds.), Aquatic Biota of
Mexico, Central America and the West Indies, pp. 433-438. San Diego, California.
Watson, Jr., C. N., and M. W. Heyn. 1992. A preliminary survey of the Chironomidae (Diptera) of Costa
Rica, with emphasis on the lotic fauna. Netherlands Journal of Aquatic Ecology 26:257-262.
This content downloaded on Wed, 6 Feb 2013 11:59:46 AMAll use subject to JSTOR Terms and Conditions
Copyright © 2022 FDOKUMEN




















