
Sustainable Highland Rivers Management in Ethiopia - LARIMA
64
Sustainable Highland Rivers Management in Ethiopia DELIVERABLE 1.3 Top‐down operative stream classification system (typology) for Ethiopian highlands
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of Sustainable Highland Rivers Management in Ethiopia - LARIMA

Sustainable Highland Rivers Management
in Ethiopia
DELIVERABLE 1.3
Top‐down operative stream classification system (typology) for Ethiopian highlands

Deliverable No. 1.3
Authors
Aschalew Lakew Haile, EAIR‐NFALRC Otto Moog, BOKU
Suggested citation: Haile, A.L. & O. Moog (2016): LARIMA. Deliverable 1.3: Top‐down operative stream
classification system (typology) for Ethiopian highlands. Appear‐ Austrian Partnership Programme in
Higher Education & Research for Development.
Document title: Top‐down operative stream classification system (typology) for Ethiopian highlands
Work Package No.: 1
Document Type: Intern Deliverable
Date: 23.06.2016
Document Status: Version 2
Acknowledgements
This project has received funding from APPEAR ‐ Austrian Partnership Programme in Higher Education
& Research for Development. APPEAR is a programme of the Austrian Development Cooperation and
is implemented by the OeAD.
Authors would like to thank all of the individuals, whose names are not mentioned as main
contributors, but have contributed by providing comments and photos to this deliverable. Photos used
with permission of the copyright holder.

Deliverable No. 1.3
Content
1 Introduction ..................................................................................................................................... 2
2 Ethiopian eco‐geographical features .............................................................................................. 2
Altitude .................................................................................................................................... 2
Geology .................................................................................................................................... 3
Hydrogeology .......................................................................................................................... 3
Hydrology ................................................................................................................................ 4
Rainfall in Ethiopia ................................................................................................................... 5
Temperatures in Ethiopia ........................................................................................................ 6
Soil types ................................................................................................................................. 7
Land cover ............................................................................................................................... 7
Agro‐ecological zones of Ethiopia ........................................................................................... 8
3 Bio‐geographic classifications applicable for river management .................................................... 9
Freshwater Ecoregions of the World (FEOW) ......................................................................... 9
Detailed description of the FEOW freshwater ecoregion of Ethiopia .................................. 10
3.2.1 Lake Tana (526) ............................................................................................................. 10
3.2.2 Northern Eastern Rift (528) ........................................................................................... 12
3.2.3 Upper Nile (522) ............................................................................................................ 15
3.2.4 Ethiopian Highlands (525) ............................................................................................. 16
3.2.5 Lake Turkana (530) ........................................................................................................ 18
3.2.6 Horn of Africa (529) ....................................................................................................... 21
3.2.7 Lower Nile (523) ............................................................................................................ 23
3.2.8 Shebelle – Juba (531) ..................................................................................................... 26
4 Terrestrial ecoregions .................................................................................................................... 29
5 Other Ethiopian regional typologies ............................................................................................. 44
Ecosystems of Ethiopia .......................................................................................................... 44
6 African subregions by Barber‐James & Gattolliat ......................................................................... 47
7 Proposed river typology based on ecoregions and eco‐geographic features of Ethiopia ............ 48
Examples of stream typology ................................................................................................ 49
8 Preliminary bottom‐up validation of proposed typology ............................................................. 53
9 References ..................................................................................................................................... 57

Deliverable No. 1.3
1 Introduction
Biodiversity is not spread evenly across the Earth but follows complex patterns determined by climate,
hydrology, geomorphology, geology, soil composition, land‐use, vegetation, ecology and the
evolutionary history of the planet. The use of a typology to classify streams has become an accepted
prerequisite of ecological assessment (Hering et al., 2004). A typology generalizes knowledge that can
be applied on a wider scale and improves the comparability of running waters in management,
assessment, and prediction (Hawkes, 1975).
Stream typology is defined as a large area of land that contains an eco‐geographically distinct
assemblage of natural communities that
share a large majority of their species and ecological dynamics
share similar environmental conditions, and
interact ecologically in ways that are critical for their long‐term persistence
Frey (1977) stated that there are recognizable regions within which particular patterns are observed.
These regions generally exhibit similarities in the mosaic of environmental resources, ecosystems, and
effects of humans, and can therefore be termed “ecological regions” or “ecoregions”. Ecological
regions are seen as areas of relative homogeneity in ecological systems and relationships between
organisms and their environment (similar climate, landform, soil, potential natural vegetation,
hydrology, or other ecologically relevant variables) (Omernik, 1987).
In recent years there has been an increasing awareness that these ecoregions exist and effective
management of environmental resources must be undertaken with an ecosystem perspective
(Omernik, 1995; Moog et al., 2004). Consequently it became apparent in the early 1980 in the US, the
early 1990s in Australia and the late 1990s in Europe that administrative regions were no longer a
satisfactory basis for water management planning, monitoring and assessment. The use of ecoregions
or bioregions in water management and conservation is widespread nowadays on many continents,
e.g. the Global 200 ecoregions (WWF), American ecoregions (Omernik, 1987), Australian bioregions
(Thackway & Cresswell, 1995), African ecoregions (Kleynhans & Hill, 1999) and European ecoregions
(Water Framework Directive, 2000).

Deliverable No. 1.3
2
2 Ethiopian eco‐geographical features
Altitude
The topography of Ethiopia is highly diverse, with elevation ranging from 125 m below sea level at the
Denakil Depression to 4620 m a.s.l. at Ras Dejen. More than 45% of the country is dominated by a high
plateau with a chain of mountain ranges divided by the East African Rift Valley. A geographic region
with elevations greater than 1500 m is known as the highlands where 90% of the nation’s population
resides, and a region with elevation less than 1500 m a.s.l. is known as lowland. This large diversity of
terrain has led to wide variations in climate, soils and natural vegetation. Elevation is an important
element in determining the climate of Ethiopia. For every 1000 m, the temperature drops about 6.5
degrees Celcius. Ethiopia has five climatic zones defined by altitude and temperature:
1. Hot, arid zone below 500 m, where average annual rainfall is less than 400 mm and average annual
temperatures range between 28°C and 34°C or higher;
2. The warm to hot, semi‐arid zone includes those areas with an altitude of 500–1,500 m, average
annual rainfall generally of around 600 mm (but as high as 1,600 mm in the western lowlands
of Gambella) and an average annual temperature range of 20–28°C;
3. The warm to cool, semi‐humid zone covers the temperate highlands between 1,500 and 2,500.
Average annual temperatures vary between 16°C and 20°C, and annual rainfall is generally around
1,200 mm, reaching 2,400 mm in the south‐west;
4. The cool to cold humid zone includes the temperate highlands between 2,500 and 3,200 m, where
average temperatures range between 10°C and 16°C, with an annual rainfall of 1,000 mm and up to
2,000 mm in higher areas;
5. The cold, moist temperate zone covers the Afro‐alpine areas on the highest plateaus between 3,200
and above; average temperatures are below 10°C and annual rainfall averages less than 800 mm.
Figure 1: Distribution of Ethiopian highlands across 12 drainage basins
Deliverable No. 1.3
3
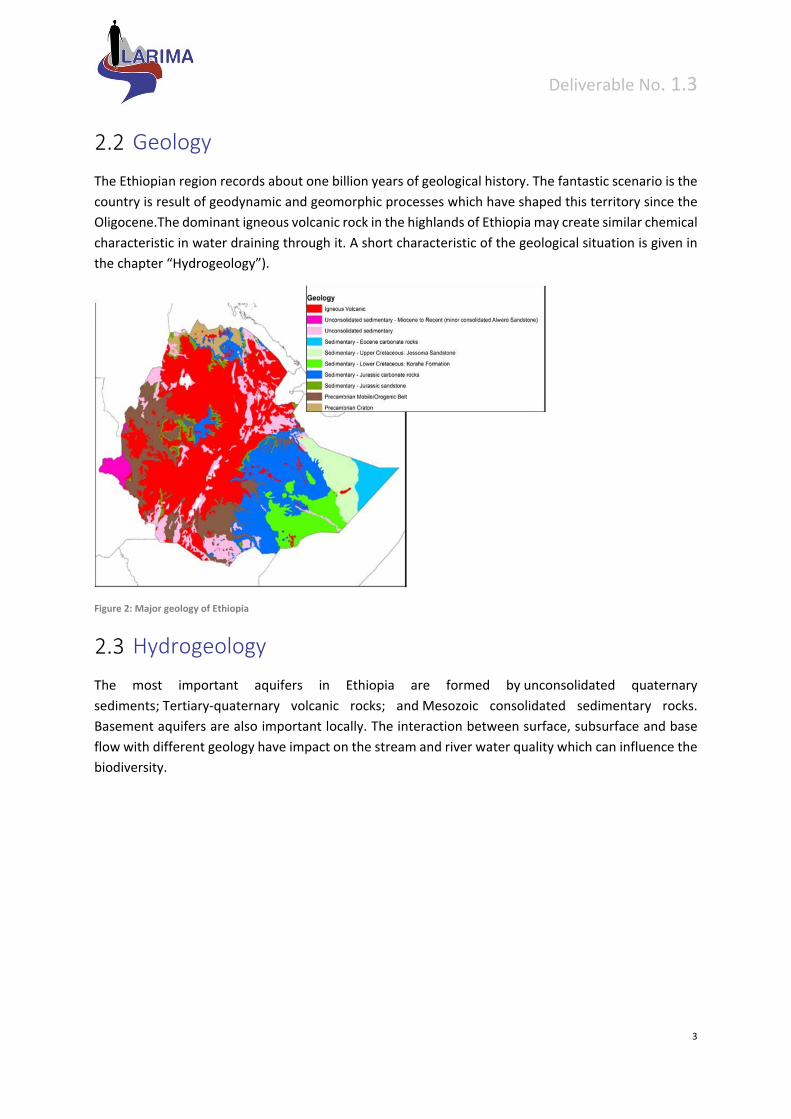
Geology
The Ethiopian region records about one billion years of geological history. The fantastic scenario is the
country is result of geodynamic and geomorphic processes which have shaped this territory since the
Oligocene.The dominant igneous volcanic rock in the highlands of Ethiopia may create similar chemical
characteristic in water draining through it. A short characteristic of the geological situation is given in
the chapter “Hydrogeology”).
Figure 2: Major geology of Ethiopia
Hydrogeology
The most important aquifers in Ethiopia are formed by unconsolidated quaternary
sediments; Tertiary‐quaternary volcanic rocks; and Mesozoic consolidated sedimentary rocks.
Basement aquifers are also important locally. The interaction between surface, subsurface and base
flow with different geology have impact on the stream and river water quality which can influence the
biodiversity.

Deliverable No. 1.3
4
Figure 3: Major hydrogeology of Ethiopia
Hydrology
Most hydrological records in Ethiopia started in the 1960s following the initiation of the Blue Nile Basin
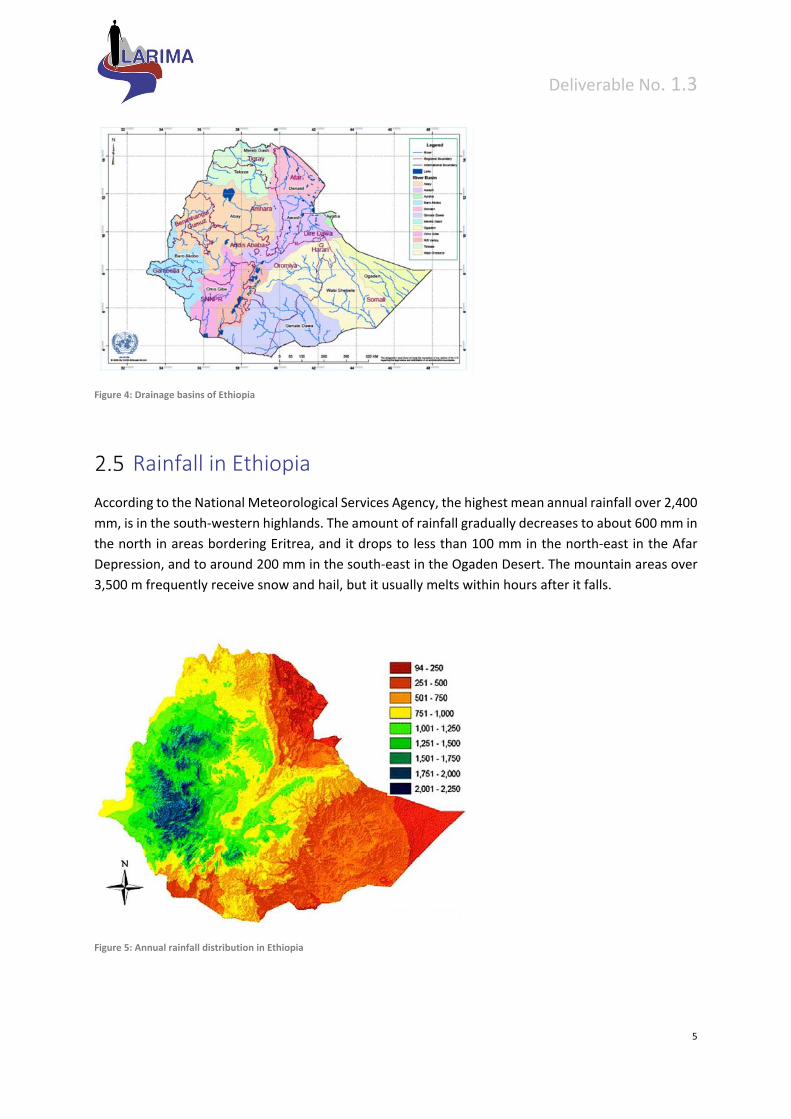
Master Plan study by the USBR (United States Bureau of Reclamation). The country has 12 drainage
basins (8 river basins, 1 lake basin and 3 dry basins) originated from the highlands with the total annual
surface runoff about 122 billion cubic meters and causes about 3 billion tons of top soil erosion per
year. Ethiopia is often referred to as the ‘water tower’ of horn of Africa mainly because of its wide
variety of landforms and climatic conditions, creating an extensive river network system throughout
the country. Despite the huge water resource in the country, very small amount has been developed
for agriculture, hydropower, industry, water supply, fishery and other purposes.
Table 1: Short description of the twelve basins of Ethiopia
River basin Source Catchment (km²) Direction of flow Annual runoff
(BM3) Abay (R) West, South ‐ west highlands 199,812 West to Sudan 52.6 Wabeshebele (R) Bale highlands 202,220 East to Somalia 3.16 Genale (R) Bale highlands 171,042 East to Somalia 5.88 Awash (R) Central highlands 112,696 North east to Djibouti 4.6 Tekeze (R) North Wello highlands 82,350 West to Sudan 8.2 Omo‐Gibe (R) Central, Western highlands 79,000 South to Kenya 17.6 Baro‐Akobo (R) Western Highland 75,912 West to Sudan 23,6 Rift valley (L) Central and Arsi highlands 52,739 Internal 5.6 Mereb (R) Adigirat highlands 5,900 North‐west to Eretria 0,26 Ogaden(D) ‐ 77,120 East ‐ Denakil (D) North Wello highlands 64,380 East 0.86 Aysha (D) ‐ 2,223 East ‐
Source: Modified from Integrated River Basin Master Plan Studies (MoWR, 1998) D= Dry basin R= River basin, L= Lake basin
Deliverable No. 1.3
5
Figure 4: Drainage basins of Ethiopia
Rainfall in Ethiopia
According to the National Meteorological Services Agency, the highest mean annual rainfall over 2,400
mm, is in the south‐western highlands. The amount of rainfall gradually decreases to about 600 mm in
the north in areas bordering Eritrea, and it drops to less than 100 mm in the north‐east in the Afar
Depression, and to around 200 mm in the south‐east in the Ogaden Desert. The mountain areas over
3,500 m frequently receive snow and hail, but it usually melts within hours after it falls.
Figure 5: Annual rainfall distribution in Ethiopia

Deliverable No. 1.3
6
Based on this rainfall distribution pattern, the following four major rainfall regimes can be
distinguished:
1. Central, eastern and northern areas of the country experience a bimodal rainfall pattern,
receiving the majority of their rainfall from the Atlantic, while some derives from the Indian
Ocean. The big rains from June to September come mainly from the Atlantic, while the light
spring rains between February and May come from the Indian Ocean.
2. Western and south‐western parts of the country experience a unimodal rainfall pattern
brought about by wind systems coming from the Indian Oceans and merge with those from
the Atlantic to give continuous rain from March or April to October or November. The amount
of rainfall and length of the rainy season decreases from south to north.
3. Southern and south‐eastern parts of the country experience a bimodal rainfall pattern brought
about by the wind system coming from the Indian Ocean from September to November and
from March to May. The most reliable rainy months are April and May.
4. North‐eastern parts of the country comprise part of the western escarpment of the Pitt Valley
and the adjacent Afar depression. The lowlands have only one rainy season during which only
a little rain falls. However, the escarpment, particularly in the north, can have a third rainy
season brought by moist winds from Asia which have crossed the Arabian Peninsula and cool
as they rise over the Ethiopian escarpment. These can bring mist and rain anytime between
November and February.
Temperatures in Ethiopia
The highest mean maximum temperatures in the country, about 45°C from April to September and
40°C from October to March, are recorded from the Afar Depression in north‐east Ethiopia. The other
hot areas are the north‐western lowlands, which experience a mean maximum temperature of 40°C in
June, and the western and south‐eastern lowlands with mean maximum temperatures of 35°C to 40°C
during April.
The lowest mean temperatures, of 4°C or lower, are recorded at night in highland areas between
November and February (National Meteorological Services Agency, 1989; Ethiopian Mapping
Authority, 1988). Many of those areas, particularly in valley bottoms, have occasional ground frost.
Figure 6; Mean annual temperature in Ethiopia

Deliverable No. 1.3
7
Soil types
The wide ranges of topographic and climatic factors, parent material and land use have resulted in
extreme variability of soils (FAO, 1984). In different parts of the country, different soil forming factors
have taken precedence. According to the Ministry of Agriculture about 19 soil types are identified
throughout the country. The big proportion of the country’s landmass is covered by lithosols, nitosols,
cambisols and regosols in order of their importance. Research showed that Potassium, Nitrogen,
Cation Exchange Capacity (CEC) and organic matter contents of most Ethiopian highland soils are
generally high by international standards (EARO, 1998), whereas their phosphorous content is low to
very low.
Figure 7: Major soil types of Ethiopia
Land cover
Ethiopia's wide ranging altitudes and associated agro‐ecological zones have produced varied landscape
and vegetation types, from tropical moist forests (high forest) in the southwest and the Bale
Mountains, to desert shrubs in the east and northeast and parkland agroforestry on the southern
highlands. Pressures on forestlands from agriculture expansion and wood fuel consumption are likely
to increase in the future as the population in Ethiopia is increasing.

Deliverable No. 1.3
8
Figure 8: Land cover of Ethiopia
Agro‐ecological zones of Ethiopia
The multitude of agro‐ecological zones (AEZs) of Ethiopia is traditionally classified into five categories
with traditional names assigned to each zone, based on altitude and temperature: Bereha, kola,
weinadega, dega and wurch. However, the amount of rainfall and its distribution are also important in
classifying common agro‐ecological zones. It is common to associate the traditional zones with
elevation and temperature and try to recognize agroclimatic and vegetation zones. Many researchers
have classified the vegetation and ecological zones (Zerihun, 1999; MoA, 2000). The current AEZ
classification (MoA, 2000) is based on the basic ecological elements of climate, physiography, soils,
vegetation, farming systems, etc. The intention of better characterization is to suit the country’s
diverse but unique natural and cultural diversity. Eighteen major AEZs are delineated and named by
terms describing the broad moisture and elevation conditions of areas.
Figure 9: Agro‐ecology of Ethiopia

Deliverable No. 1.3
9
3 Bio‐geographic classifications applicable for river management
Several efforts have been made to delineate eco‐geographic regions such as Eco‐Zones, Ecoregions,
Bioregions, and others for the world in general and Ethiopia in particular. The basic grouping into
terrestrial or freshwater regions can be seen as a main unit of discrimination.
Freshwater Ecoregions of the World (FEOW)
FEOW provides a global biogeographic regionalization of the Earth's freshwater biodiversity. The WWF
Conservation Science Program in partnership with The Nature Conservancy and 200 freshwater
scientists from institutions around the world under the guidance of Abell (2008) have developed
FEOW.
The freshwater ecoregion map encompasses 426 units and delineated based on the best available
information. However, data describing freshwater species and ecological processes are characterized
by marked gaps and improved information in the future may warrant map revisions.
Ethiopia shares nine freshwater ecoregions of the FEOW system similar to Thieme regions (Thieme et
al., 2005). Thieme et al. (2005) can be regarded as a quite prominent zonation outcome among the
attempts to delineate freshwater ecoregions. Many authors used freshwater ecoregions in their
analysis because they provide appropriate units for representation of distinct species assemblages,
habitats, and processes at the continental scale. Their ecoregions delineation took into account aquatic
species distributions (with a strong focus on fish, molluscs and herpetofauna) and drainage basins
divides. The 10 African ichthyofaunal provinces of Roberts (1975) served as their basis for their
bioregion delineation.
Table 2: Classification of freshwater ecoregions in Thieme and FEOW systems
Thieme system FEOW system 3 Lake Tana 526 Lake Tana 4 Northern Eastern Rift 528 Northern Eastern Rift
15 Upper Nile 522 Upper Nile 39 Ethiopian Highlands 525 Ethiopian Highlands 56 Lake Turkana 530 Lake Turkana
83 Horn 529 Horn of Africa 86 Lower Nile 523 Lower Nile 89 Red Sea Coast 527 Red Sea Coast 90 Shebella‐Juba‐Catchment 531 Shebelle ‐ Juba

Deliverable No. 1.3
10
Figure 10: The map provides a detail of the FEOW regions that cover the area of Ethiopia
Detailed description of the FEOW freshwater ecoregion of Ethiopia
3.2.1 Lake Tana (526)
Lake Tana is the source of the Blue Nilelocated in the highlands of Ethiopia lies in the north part
of Ethiopia and. The Blue Nile descends from Lake Tana to Tiss‐isat falls isolating the lake’s freshwater
fauna from the rest of the Nile.
Main rivers or other water bodies
Lake Tana was formed by a volcanic blockage that reversed the previously north‐flowing river system
(Beadle, 1981). The total area of the Lake Tana basin is 16,500 km2 and the lake itself covers about
3,150 km2. Numerous seasonal streams and four perennial rivers feed the lake, while only one—the
Blue Nile—leaves it (Nagelkerke, 1997).
The lake is situated in the highlands of Ethiopia at about 1,800 m, and experiences a tropical highland
climate.
Air temperatures range widely, between 7 oC to 31oC, whereas water temperatures stay relatively mild,
normally between 18oC and 26oC (Nagelkerke, 1997). The dry season lasts from October/November to
May/June with maximum monthly rainfall (up to 500 mm/month) in July. Annual rainfall in the vicinity

Deliverable No. 1.3
11
of the lake averages 1315 mm/year, but evaporation is higher at about 1,800 mm/year (Burgis &
Symoens, 1987).
Freshwater habitats
Because evaporation exceeds rainfall, the hydrology of this shallow lake depends largely on the local
climate (Burgis & Symoens, 1987). Lake level varies depending on seasonal rains. The average
difference between the lowest lake level (May‐June) and the highest (September‐October) is 1.5
m (Nagelkerke, 1997). The lake has a mean depth of 8 m, and is well mixed due to relatively strong
winds in the evenings (Nagelkerke, 1997). Cyperus papyrus and other Cyperus spp. line the shores of
the lake (Beadle, 1981).
Fish Fauna
Fish species in the lake are most closely related to those of the Nilo‐Soudanian biogeographic
region. Lake Tana hosts extended cyprinid species flock in Africa. Fifteen species of large barbs have
been described from Lake Tana (Nagelkerke, 1997; Nagelkerke & Sibbing, 1998;. The species flock is
believed to be less advanced in its evolution than Lake Lanao’s cyprinid flock (Mina et al., 1996). Eight
of the large barbs are piscivorous, and Barbus humilis and the newly described small species, Barbus
tanapelagius, are thought to be the major prey species (De Graaf et al., 2000). It is likely that the Lake
Tana barbs evolved from one ancestral species that probably resembled Barbus
intermedius (Nagelkerke, 1997).
About 70% of the fish species in this highland lake are endemic, including eighteen endemic cyprinids.
The tilapia (Oreochromis niloticus) of Lake Tana belongs to a widespread species but is described as an
endemic subspecies, Oreochromis niloticus tana (Seyoum & Kornfield, 1992). The only river loach
(family Balitoridae) known from Africa, Nemacheilus abyssinicus, was described from Lake Tana in
1902 and rediscovered in 1992 in the lake and in the upper Omo River (Dgebuadze et al., 1994).
The large catfish, Clarias gariepinus, widespread throughout Africa, also lives in the lake and forms an
important part of the fishery.
The invertebrate fauna is relatively diverse. Fifteen species of molluscs, dominated by the Planorbidae
family, have been described, including one endemic. An endemic freshwater sponge, Makedia
tanensis, has recently been discovered in the lake. The sponge is small (specimens found were up to
about 2 cm), white and of an encrusting form belonging to a monotypic genus (Manconi et al. 1999).

Deliverable No. 1.3
12
Figure 11: Lake Tana freshwater ecoregion
3.2.2 Northern Eastern Rift (528)
Numerous highly productive lakes lie in the Eastern Rift valley that cleaves the eastern and western
sections of the high altitude Ethiopian dome and extends from the edge of the xeric, Red Sea Coastal
ecoregion in the north to Lake Awassa in the south.
Figure 12: Northern Eastern Rift freshwater ecoregion (red boundary)

Deliverable No. 1.3
13
Main rivers or other water bodies
Two closed basins occur in this section of the Rift Valley: in the south, Lake Awassa basin, consisting of
Lake Awassa and the swampy Lake Shallo; and basin in the north composed of a series of four
interconnected lakes (Abijata, Langano, Shala and Zwai). There is also a number of hot springs adjacent
to the lakes.
This ecoregion also contains a number of crater lakes (Bishoftu, Aranguade, Hora, Kilotes and Pawlo)
located at the northwestern edges of the rift valley around the town of Debrezeit (Mohr 1961), at an
altitude of about 1,900 m. These lakes lie in volcanic explosion craters produced about 7,000 years
ago. The Awash River, which begins in the Ethiopian Plateau, flows north in the Rift Valley and
terminates in Lake Abhe, a closed lake near the border between Ethiopia and Djibouti.
The mean annual air temperature of this ecoregion varies with altitude and ranges between 20oC and
22oC (Tudorancea et al., 1999). There is a four‐month dry season from November to February and an
eight‐month rainy season from March to October. The main rains occur between July and September.
Freshwater habitats
Five major lakes and several rivers lie within this ecoregion. Lake Zwai, the most northerly lake, is
located at an altitude of 1,840m, covers 434 km2 and has average depth of 2.5 meters (Balarin, 1986).
Extensive marshes of Papyrus (Cyperus papyrus) border the lake and produce large numbers of
mosquitoes (Anopheles pharoensis, A. mauritianus and Taeniorhynchus uniformis), (Omer‐Cooper,
1930). The Makki River flows into the lake from the northwest and the River Kattar flows into it from
the northeast. The lake’s waters flow out through the River Bulbula into Lake Abijata.
The three lakes in the chain are Lake Langano, Lake Abijata, and Lake Shala. Lake Langano, located at
an altitude of 1,582 m, covers 241 km2, and has a maximum depth of 47.9 m and a mean depth of 17
m. Salinity is 1.88 g per liter (Wood & Talling, 1988). Lake Langano receives most of its water from small
rivers that drain from the Arsi Mountains, which make up the eastern wall of the Rift valley. The only
outlet from the lake is the Hora Kelo River, which flows into Lake Abijata. Lake Abijata is found at an
altitude of about 1,600 m and is an alkaline lake. The shores slope gradually, and are muddy, with areas
of Juncus vegetation. The depth is about 10 m, and the bottom of the lake is sandy. Three rivers feed
Lake Abijata: Gogessa, Bulbula, and Hora Kelo. Lake Abijata has no outlet, and it loses its water by
evaporation only. Lake Shala is the deepest of the Ethiopian Rift lakes, 266m. It is 28 km long and 15
km wide, and is surrounded primarily by Pleistocene volcanic rocks. The eastern and western shores
are covered by lacustrine deposits and Holocene sands, occasionally blackened by obsidian detritus
(Mohr, 1961). The great depth of Lake Shala may be related to the origin of the basin by intense
faulting. Two permanent creeks and several seasonal streams flow into its closed basin.
Lake Awassa is located at an altitude of 1,680 m, lies south of the other lakes in the ecoregion. It has a
surface area of 88 km2, a maximum depth of 22 m, and an average depth of 11 m. Lake Awassa is a
polymictic lake. The water is murky and alkaline with a pH between 8.75 and 9.05 (Tudorancea et al.,
1988). Like Lakes Abijata and Shala, it is a terminal lake without any visible outlet. Its main tributary is

Deliverable No. 1.3
14
the Tikur Wuha River, which drains swampy Lake Shallo. An extensive belt of submergent and
emergent rooted vegetation, which extends about 150 m offshore, covers the littoral zone.
The flora varies among the different lakes. Three major groups of algae dominate: Chlorophyceae,
Cyanophyceae and Diatomophyceae (Tudorancea et al., 1999). The most common emergent plants
are Scirpus spp., Typha angustifolia, Paspalidium germinatum, and Phragmites sp. Nymphaea
coerulea and Potamogeton spp. are the dominant species of floating and submerged vegetation.
Abijata and Shala Lakes lack aquatic macrophytes.
Terrestrial Habitats
The vegetation to the east and south of Lake Shala is Acacia‐Euphorbia savanna. The most common
trees are the woodland acacias Acacia etbaica, A. tortilis, and Euphorbia abyssinica, and bushes
of Maytenus senegalensis. Beds of bulrushes and sesbania occur where the hot springs and rivers enter
the lake, but most of the shore has steep cliffs.
Fish Fauna
The lakes and streams of the Northern Rift ecoregion support a limited freshwater fauna, including
only eight fish, with few endemic species. The fish fauna of Lake Awassa consists of two species
of Barbus (B. intermedius and B. cf. amphigramma), the North African catfish (Clarias gariepinus),
and Oreochromis niloticus. Oreochromis niloticus is abundant in the lakes and rivers of this ecoregion.
No fish are recorded from Lake Shala. Despite their depauperate number of species, the lakes of this
ecoregion support most of the fish production of Ethiopia.
Three endemic fish, Barbus ethiopicus, B. microterolepis, and Garra makiensis, inhabit Lake Zwai and
its adjacent rivers.Introduced species of Tilapia zilli, Clarias gariepinus, and carps are present in Lake
Zwai.
Only thirteen aquatic frogs, three aquatic reptiles, twelve aquatic mollusks, and four aquatic mammals
live in or adjacent to the freshwater lakes and streams. There are considerable numbers of pelicans,
cormorants, ducks, snipe, stilts, egrets, grebes, ibis, herons, gulls, and darters around Lake Zwai. One
endemic frog, Bufo langanoensis, lives along the shores of Lake Langano and its tributaries. Lakes Zwai
and Langano also harbor hippopotamus (Hippopotamus amphibius).
During the 1970s and 1980s over 400 species of birds were recorded from the Abijata‐Shala National
Park. This park is positioned in one of the narrowest parts of the Great Rift Valley, which is a major
flyway for both Palaearctic and African migrants, particularly raptors, flamingoes, and other water
birds. Many of these birds stop over to rest and feed within the ecoregion. The shallow waters of Lake
Abijatta are remarkably rich in insect life, with large swarms of Corixa in particular, though plankton
diversity is low. The zooplankton fauna in the Ethiopian rift lakes is dominated, in terms of biomass, by
copepods and cladocerans (Tudorancea et al., 1999).

Deliverable No. 1.3
15
Delineation
This ecoregion is defined by the northern lakes of the Ethiopian rift valley and distinguished by lakes
with a distinctive fauna when compared to the more southern rift valley lakes of Chamo and Abaya.
The fish fauna in the northern lakes appears to have been derived from Awash and associated rivers,
while the fish fauna of the latter is Nilo‐Sudanic. These faunal affinities may be explained by the
tumultuous geologic past of the ecoregion, which was exposed to six volcanic events between the
Oligocene and the present (Woldegabriel et al., 1990). Analyses of invertebrate and fish fossils found
in the sediments of the lakes indicate that Lakes Zwai, Abijata, Langano and Shala were once united
into a single freshwater lake draining northward into the Awash River. The present‐day lakes are the
result of subsequent tectonic or volcanic activity (Tudorancea et al., 1999).
3.2.3 Upper Nile (522)
The vast swamps of the Sudd are the primary feature of the Upper Nile ecoregion, which is situated
mainly in Sudan with smaller areas in the Democratic Republic of Congo, Uganda, and Ethiopia. The
ecoregion encompasses the basin of the White Nile River; its major tributaries, the Sobat River and
Bahr el Ghazal; Lake Albert; and Lake Albert’s main influent, the Semliki River. The point at which the
White Nile joins with the Blue Nile marks the northernmost border of the ecoregion (Rzóska 1974;
Dumont 1986).
The southern and highest upstream part of this ecoregion is situated in the Rift Valley. This area is
rugged, with mountain peaks over 4,600 m. Further to the north, the topography changes dramatically
where the Albert Nile flows into the shallow depression of the Sudd. The depression ranges in elevation
between 420 to 380 m asl and stretches for about 600 km from end to end. Most of the depression is
flat with a gradient of 0.01% or less, and it is underlain by clay soils (Food and Agriculture Organization,
1997).
The dynamic Sudd wetland, whose size varies substantially in response to seasonal and inter‐annual
changes in water input, contains a diversity of habitats and supports a rich aquatic and terrestrial fauna
(Rzóska 1974). Water entering the Sudd swamps drains from the hills of the Nile‐Congo watershed
divide, the escarpment of the Uganda Plateau, the Imatong Mountains, and the Ethiopian Plateau.
The swamps and floodplains of the Sudd are among the most important wetlands in Africa and support
a rich biota. Twenty‐two families and 118 species of fish are known to occur within the Upper Nile
ecoregion, including 16 endemics. Cyprinidae is the most diverse family, with Alestiidae, Cichlidae,
Mochokidae, Mormyridae, Poeciliidae, and Schilbeidae also represented by high numbers of species.
Lake Albert has a Nilotic riverine ichthyofauna comprised of 15 families and 46 species.

Deliverable No. 1.3
16
Figure 13: Upper Nile freshwater ecoregion
3.2.4 Ethiopian Highlands (525)
The highlands extend from Eritrea in the north to Kenya in the south. With a long history of isolation,
the Ethiopian Highlands are known to harbour a highly endemic biota.Rivers of the western highlands
generally flow towards Sudan, whereas those of the eastern highlands tend to flow towards the Indian
Ocean.
Water bodies
In the northwestern part of the highlands, the deep, steep‐sided valleys of the major rivers separate
blocks of mountains, and the upper courses of the big rivers such as the Tekezze and Abay (Blue Nile)
plunge through deep gorges. This part of the Ethiopian highlands is also the source of the headwaters
for the Blue Nile. The Blue Nile watershed is the largest basin in Ethiopia. Rivers of this basin drain the
great central plateau and the Blue Nile descends about 1,450 m in a distance of only 350 km from its
source to Khartoum. The westward flowing rivers (the Tekezze, Angereb, Atbara, Abay, Baro and
Akobo) form part of the Nile drainage basin. Three major highland lakes, Lakes Hayq, Ardebo and
Ashengie, lie near the edge of the western escarpment of the rift valley at altitudes between 2,000 and
2,500 m. Lake Hayq, located in northern Ethiopia’s Wollo region, has an area of 5 km2 and a maximum
depth of 23 m, and is noteworthy for its extremely clear water (Kebede et al., 1992). Lake Ardebo is
located about 5‐km southeast of LakeHayq. This lake is smaller in size than Lake Hayq and flows into
Hayq via the Anchercah River (Kebede et al., 1992). Lake Ashengieis located north of Lake Hayq in the
Tigray region, and sits at an altitude of 2,460 m. The lake covers an area of 25 km2 with a maximum
depth of 20 m and a mean depth of 14 m (Wood & Talling, 1988). The lake is fed by a number of small
streams from the surrounding areas and there is no drainage out of the lake. According to
Westphall (1975), uplift of the Ethiopian highlands together with Arabia occurred on an extensive scale
after the regression of the Red Sea towards the southeast in the late Mesozoic to early Tertiary.
The Great Rift Valley bisects the highlands into the eastern and western massifs, which are surrounded

Deliverable No. 1.3
17
by escarpments. This ecoregion contains about 70% of Africa’s highlands. The highest peak, Ras Dejen
(Dashan) at 4,620 meters, is in the northern Ethiopia. The southeastern portion of the Ethiopian
Highlands includes the Sidamo, Bale, Arsi and Harerge Mountains. The highlands in this region are
made up of volcanic rocks, and deep river cuts expose crystalline rocks (Ethiopian Mapping, 1988)
The Ethiopian highlands receive about 950 mm or more of rainfall due to a double passage of the
intertropical convergence zone. The high mountains east of Lake Tana and the southwestern
mountains stand out as places of higher rainfall. They receive 2,000 mm or more of rainfall each
year (Westphal, 1975). A rainfall regime that peaks in March‐May and June‐August is typical for the
Ethiopian Highlands.
Figure 14: Ethiopian highlands freshwater ecoregion
Fish
The fishes of the high mountain torrential streams are largely cyprinids (Harrison, 1995; Getahun &
Stiassny, 1998) adapted to the swiftly flowing floodwaters that occur seasonally. Two genera of fishes
(Barbus and Garra) dominate the fish fauna of these streams. Clarias gariepinus, Varicorhinus
beso and Labeo spp. are also found in high numbers. The Baro‐Akobo basin is apparently particularly
rich in fish diversity (Golubtsov et al., 1995). The fauna is Nilo‐Sudanic and is dominated by Alestes,
Bagrus, Barilius, Citharinus, Hydrocynus, Hyperopisus, Labeo, Malapterurus, and Mormyrus genera.
Endemic fishes
Endemism appears to be high among fish, but the fish fauna is not well known. Endemic fishes of the
genus Garra (e.g. G. dembecha, G. duobarbis and G. ignestii) have recently been described (Getahun,
2000). Lake Hayq is believed to have no indigenous fish species, although the presence of Clarias
gariepinus has been reported (Kebede et al., 1992). Nemacheilus abyssinicus is an endemic species
found in the Baro‐Akobo drainage basin, the Omo‐Gibe drainage basin, and Lake Tana.

Deliverable No. 1.3
18
Other aquatic biota
The invertebrate fauna is less well known than the fishes and it is difficult to estimate endemism among
the aquatic invertebrates. Harrison and Hynes (Harrison & Hynes 1988) indicated
that Dugesia spp., Baetis harrisoni, Pseudocloeon sp., Centroptilum sudafricanus, Afronorus
peringueyi, Neoperla spp., Hydropsyche sp., Simulium spp., nymphs of Aeschna, and chironomid
larvae dominate the benthic communities in the stony runs and torrents of the Ethiopian highlands.
Compared to other highland ecoregions, the Ethiopian Highlands support a rich aquatic mollusc fauna
with over 20 species described.
Delineation
Despite the fact that the Ethiopian highlands are presently separated from both the East African and
the South Arabian mountains, the riverine fauna resembles that of east and southern
Africa (Tudorancea et al. 1999), along with some elements of the Arabian Peninsula. Cyprinids are the
dominant fish in the rivers of this ecoregion. For example, it is known that some smallBarbus species
(e.g., Barbus paludinosus, B. trimaculatus and B. radiatus) have widespread distributions extending
from South Africa to East Africa and into the highlands (Skelton et al., 1995). There are also fish groups
(e.g. Garra) common to the Ethiopian highlands and the Arabian Peninsula. These fish groups are
estimated to have originated in the Lower Tertiary or late Cretaceous, before the separation of India
and the Arabian Peninsula from continental Africa (Briggs 1987); whereas the Red Sea is believed to
have separated the African continent from the Arabian Peninsula in the early Tertiary, between the
Eocene and Oligocene epochs(Getahun, 1998).
Historically, few scientific studies have been made on the fauna of the river systems of Ethiopia;
however, two recent studies have elevated the level of data available for the fish of this
ecoregion (Getahun & Stiassny, 1998; Golubstov et al., 2002). River systems in the Tekezze‐Angereb
basin have not been studied at all due to security problems in the past. Preliminary reports indicate
that the large river bodies of this basin support a rich fish fauna and research is needed to confirm this.
Some information on the benthic fauna of Ethiopian mountain streams and rivers is available in
Harrison and Hynes (1988).
3.2.5 Lake Turkana (530)
The Lake Turkana ecoregion reaches north to include Lakes Abaya and Chamo, as well as the
headwaters of the Omo River southwestern Ethiopia.
Lake Turkana is the largest lake in the eastern portion of the Rift Valley and the fourth largest lake by
volume in Africa (Beadle 1981). Lying in a low closed basin at approximately 365 m asl, the lake is
situated primarily in northwestern Kenya, with its northernmost end inside Ethiopia. Of the twelve
principal rivers that feed Lake Turkana, the River Omo is its only perennial tributary, supplying over
90% of the lake’s inflow (Beadle 1981). The Omo River drains the southwestern portion of the
Ethiopian Massif and flows through the Rift Valley into Lake Turkana. Of the seasonal rivers that flow

Deliverable No. 1.3
19
into the lake, the Turkwell and Kerio Rivers are the largest contributors and enter the lake along its
western edge and in its southern half (Hughes & Hughes, 1992).
Lake Abayaand LakeChamoare located in the northeastern portion of the ecoregion. Five major rivers
feed Lake Abaya, the most important of which is the Bilate. During the rainy season, overspill from
Lake Abaya is carried to Lake Chamo via the Ualo River (Hughes & Hughes, 1992).
The Ethiopian Rift Valley has a mean annual rainfall of 600 mm/year, receiving at least fifty percent of
the precipitation between July and September. The western foothills of the Ethiopian Rift escarpment
receive as much as 800‐1,000 mm of rainfall per year. This heavy rainfall causes the Omo River to flood
(June through September), bringing nutrient rich waters into Lake Turkana (Beadle 1981).
Freshwater habitats
Lake Turkana is 260 km long, with an average width of 30 km, a mean depth of 31 m, and a maximum
depth of 114 m. It has an area of approximately 7,560 km² and a volume of 237 km³ (Coulter et al.,
1986). With no surface outlet, the water budget of the lake is a balance between river and groundwater
inflow and evaporation. Evaporation rates are high, at around 2.3‐2.8 m/yr. An influx of about 19
km3/yr is required to keep lake levels steady, and high inter‐ and intra‐annual fluctuations in water
level occur as a function of the rainfall in distant upland Ethiopia. Generally, the lake level fluctuates
annually with an amplitude of about 1‐1.5 m, but it also undergoes considerable long‐term variations
that exceed those of any other lake of natural origin (Butzer, 1971). The mean retention time of water
in the lake is a short 12.5 years.
The salinity of Lake Turkana is higher than that of any other large African lake. This is due to the fact
that the lake has no outlet, and that it has contracted in volume over the last 7,500 years. Very recent
volcanic activity in the basin has also contributed to the high salinity of the lake (Beadle, 1981). The
most common emergent plants are the grasses Paspalidium geminatum and Sporobolus spicatus, with
extensive beds ofPotamogeton occurring in shallow bays (Hughes & Hughes 1992). The waters of
Abaya and Chamo contain numerous submerged plants, such as Ceratophyllum
demersum, Hydrocotyle sp., and Potamogeton spp., as well as floating plants like Lemna gibba,
Nymphaea spp., and Ottelia ulvifolia (Hughes & Hughes, 1992).
Terrestrial Habitats
The evergreen bush and woodland of the Ethiopian Massif grade into deciduous bush in the Rift
Valley. Extensive seasonal floodplains exist along the Omo River Delta, at the northern tip of Lake
Turkana. Gallery forests of Acacia elatior, Balanites aegyptiaca, and Hyphaena coriacea grow along
Lake Turkana's tributaries (Beadle, 1981; Hughes & Hughes, 1992). Swampy savanna forests of Acacia
and Ficus species line the shores of Lake Abaya and species of Typha and Phragmites are common
along the banks of both Lakes Abaya and Chamo.

Deliverable No. 1.3
20
Figure 15: Lake Turkana freshwater ecoregion
Fish
Lake Turkana is unique among the larger lakes of the eastern Rift Valley in that its aquatic fauna is
dominated by Nilotic riverine species, rather than by species of the cichlid family. Compared to other
large African lakes, Turkana has relatively low fish species richness, providing habitat for about 50
species, 11 of which are endemic. According to Hopson (1982), four fish communities live in the main
lake: a littoral assemblage, an inshore assemblage, an offshore assemblage, and a pelagic assemblage.
Spawning migrations of fish are synchronized with the ecoregion’s seasonal flooding, which occurs
from June through September. During this time, various fish species migrate up the Omo River
(Hydrocynus forskalii, Alestes baremoze, Citharinus citharus,Distichodus niloticus, Barbus bynni) and
other ephemeral affluents (Brycinus nurse, Labeo horie, Clarias gariepinus, Synodontis schall) to breed,
for periods of both long and short duration (Beadle, 1981; Hopson, 1982; Lévêque, 1997).
Endemic fishes
The endemic species nearly all live in the offshore demersal or pelagic zone. Endemic chiclids include
three halpochromine species adapted for deep water: Haplochromis macconneli, H. rudolfianus, and
H. turkanae. Other species endemic to Lake Turkana include Barbus turkanae, Brycinus ferox, B.
minutus, Labeo brunellii, Lates longispinis and Neobola stellae.

Deliverable No. 1.3
21
Other aquatic biota
Lake Turkana is an important site for waterbirds with up to 220,000 congregants having been recorded
at one time and 84 waterbird species, including 34 Palearctic migrants, known from the lake (Bennun
& Njoroge, 1999). Over 100,000 Calidris minutahave been recorded at the lake, in addition to smaller
congregations of other non‐breeding waterbirds (Pelecanus rufescens, Phoenicopterus ruber, Vanellus
spinosus, Charadrius hiaticula, C. asiaticus, C. pecuarius). Bird species present near Lake Abaya
include Anhinga rufa, Bubulcus ibis, Casmerodius albus, Egretta garzetta, Haliaeetus
vocifer, andScotopelia peli (Hughes & Hughes, 1992).
Other aquatic animals in the ecoregion include Hippopotamus amphibius, Crocodylus spp., and an
endemic freshwater turtle, the recently discovered and imperiled Turkana mud turtle (Pelusios
broadleyi) (Hughes & Hughes, 1992; Expert Center for Taxonomic Identification, 2000). Lakes Abaya
and Chamo support notably large populations of Crocodylus niloticus and Hippopotamus
amphibius (Hughes & Hughes, 1992). Three species of frog are endemic to the ecoregion (Bufo
chappuisi, B. turkanae andPhrynobatrachus zavattarii).
Delineation
The Lake Turkana ecoregion is defined by the basins of lakes Turkana, Abaya and Chamo and
the Omo River basin. Fish species in the ecoregion are mainly of Sudanian origin, providing evidence
of a previous connection to the Sobat and the Nile Rivers (Beadle, 1981). For example, the Nilotic
species, Nile tilapia (Oreochromis niloticus), Bagrus domac, and Nile perch (Lates niloticus) are
abundant and common in lakes Turkana, Abaya, and Chamo (Hughes & Hughes, 1992). The Omo River
basin also has several fish species in common with Lakes Turkana, Abaya, and Chamo. The Turkana
basin was formed from tectonic movement during the early Miocene. Evidence suggests thatLake
Turkana was once part of a larger body of water that included present day Lake Baringo (south) and
the Lotikipi Plains (west). In addition, a connection with the Nile and its tributary, the Sobat River, may
have existed more than once during particularly wet periods of the Pleistocene, with the most recent
connection occurring not more than 7,000 years BP (Beadle 1981; Dgebuadze et al., 1994)
3.2.6 Horn of Africa (529)
Habitat Type
The temporary rivers, wadis, and sinkholes of the Horn ecoregion support a depauperated freshwater
fauna adapted to fluctuating environmental conditions. The ecoregion corresponds to the xeric Horn
of Africa, the most easterly portion of the continent covering the northern portion of Somalia and small
portions of Ethiopia and Djibouti. The northern mountains drain primarily to the north toward the Gulf
of Aden and to the southeast where several wadis cut through the plateau and flow to the Indian
Ocean (Hughes & Hughes, 1992).

Deliverable No. 1.3
22
Rivers and other water bodies
From north to south, the major drainages flowing into the Indian Ocean from the interior plateau,
include the ephemeral Jaceyl, Dhuudo, and Nugaal rivers. Among the many small, ephemeral drainages
into the Gulf of Aden are the Durdur and Hodmo rivers.
Topography
A mountain chain runs parallel to the Gulf of Aden, reaching the highest point in Somalia on Shimbiris
Mountain at 2,416 m. The land slopes down from this northern ridge to an interior plateau at about
600 to 1,000 m above sea level in the central portion of the ecoregion. The low‐lying coastal plain
covers a narrow strip along the Gulf of Aden and the Indian Ocean.
The climate of the ecoregion is arid to semi‐arid with an annual rainfall of less than 250 mm. Rainfall
can be particularly sparse at low elevations; for example, Berbera, on the Gulf of Aden, has a mean
annual rainfall of 59 mm. Drought periods are common throughout the ecoregion. Rainfall is bimodal,
with wet seasons from mid‐April to June and then again from October to December (FAO Inland Water
Resources and Aquaculture Service Fishery Resources Division, 1999). The climate is hot and humid on
the coast and hot and arid inland. In the north, temperatures can reach 42o C on the Gulf coast and
can be as low as 0o C in the highlands (Hughes & Hughes, 1992).
Figure 16: Horn of Africa freshwater ecoregion

Deliverable No. 1.3
23
Freshwater habitats
Nearly all water bodies within this arid ecoregion are ephemeral. In the valley of the Nugaal River,
there are several tugs (a temporary watercourse that spreads across flat land) and bullehs (endorheic
depressions) (Hughes & Hughes, 1992). Along the tip of the Horn there are also pans, springs, and pools
in many of the small northern wadis that discharge to the Gulf of Aden and the Indian Ocean.
Sinkholes, salt pans, and subterranean water sources can be found inland.
Terrestrial Habitats
Most of the ecoregion is covered with barren desert or sparsely vegetated mixed scrub and grassland.
The vegetation of the ecoregion is primarily deciduous shrub or bushland inland, with grassy
scrublands on the coast. The most common tree species belong to the deciduous genera Acacia and
Commiphora. The understory consists of shrubby herbs less than one meter high, such as Acalypha,
Barleria, and Aerva. At lower elevations where rainfall is less consistent, vegetation becomes semi‐
desert scrubland. Around sinkholes in the interior limestone country and near tugs and bullehs, Acacia
tortilis grows in association with Commiphora spp. (Hughes & Hughes, 1992). Thickets of dense
vegetation often indicate surface or sub‐surface water. Evergreen and semi‐evergreen scrub grows in
the mountains, and Afro‐montane vegetation including juniper forest grows at the highest
elevations (Hughes & Hughes, 1992).
Fish Fauna
The aquatic fauna of this ecoregion is poorly known, however it is suspected that the ecoregion hosts
a depauperate freshwater fauna able to live in an environment with highly variable water chemistry
and flows. Eight species of freshwater fish are currently known to inhabit the Horn. Many of the fish
species are adapted to life in challenging conditions. The lungfish (Protopterus amphibius) is capable
of aerial breathing and aestivation (burrowing in the mud) for survival during anoxic or dry periods,
respectively. Aphanius dispar is found in oasis pools with hypersaline to fresh water (FishBase, 2001).
There are also several euryhaline fish. For example, the Jarbua terapon (Terapon jarbua) lives in river
mouths, intertidal areas, and also travels up rivers, and Stenogobius gymnopomus and Syngnathus
abaster are primarily estuarine species.
The monotypic Somalian blind barb (Barbopsis devecchii), the only ecoregional endemic, lives in caves
of the ecoregion.
This ecoregion is defined by the xeric Horn of Africa and is characterized by a depauperate and poorly
known aquatic fauna.
3.2.7 Lower Nile (523)
The lower Nile River provides a vital oasis for terrestrial and aquatic wildlife as it runs through the semi‐
arid Sahel and arid Saharan Desert of northern Sudan and Egypt. The boundaries of this ecoregion are
defined by the lower Nile River from Khartoum, where the White and Blue Nile rivers converge,
downstream to the Nile Delta. Notable features of the ecoregion include four major waterfalls over

Deliverable No. 1.3
24
which the Nile flows before emptying into the Aswan High Dam’s massive reservoir, named Lake Nubia
in Sudan and Lake Nasser in Egypt.
Figure 17: Lower Nile freshwater ecoregion
Rivers and other water bodies
Primary water bodies in the ecoregion include the lower Nile River from Khartoum, where the White
and Blue Nile rivers converge, downstream to the Nile Delta ; the areas draining to the lower Nile from
the west; and areas draining from the east, including the Blue Nile and Atbara Rivers up to but
excluding their uppermost reaches. Both the Blue Nile and the Atbara originate in the Ethiopian
Highlands and carry large amounts of sediments, estimated annually at 1.4 billion metric tons
(Waterbury, 2002).
Climate
The arid desert climate in the central portion of the ecoregion becomes increasingly hostile toward the
north. Two seasons occur in the ecoregion. A hot, dry summer exists from April through October, and
a cooler winter exists from November through March. Northern Sudan experiences a high mean daily
temperature of 35oC in summer and 20oC in winter, and receives a mere 20 mm of annual rainfall
(Hughes & Hughes, 1992). At Aswan in Egypt, temperatures range from a mean monthly minimum of
8oC and maximum of 23oC during the coolest month (January), to a minimum of 25oC and maximum of
41oC in the hottest month (August). Little to no precipitation falls in the Egyptian deserts. The western
desert in Egypt experiences periods of years without rainfall, and precipitation in the hills of the
eastern desert is highly variable. Streams can flow violently for several days after a storm drops up to
100 mm of precipitation, and then remain dry for several subsequent years (Hughes & Hughes, 1992).
In the late spring, rain clouds from the South Atlantic reach the Ethiopian highlands and drop their
precipitation, providing the annual Nile flood that reaches Egypt between mid‐May and early July.
Thereafter, the Nile surges with increasing volume for an average of 110 days, reaching its maximum
height and volume in September. At the beginning of the flood in June, a parcel of water in the lower

Deliverable No. 1.3
25
Nile will take twelve days to flow the six hundred miles from the Aswan High Dam to Cairo, but the
same journey requires only six days in full flood in September. The Blue Nile flood is so large that it
effectively blocks the flow of the White Nile at Khartoum, contributing an average 59%, or 52 billion
m³, to the Nile’s total flow. Further north, the discharge from the Atbara contributes another 13% or
11 bilm³ (Collins, 2002). Below the Atbara, the Nile flows through the harsh Nubian and Egyptian
deserts for 2,415 kilometres without receiving any more water before discharging into the
Mediterranean Sea.
A flat, featureless plain covers a majority of the ecoregion, though in the southeast the landscape rises
sharply into the Ethiopian highlands through which flows the Great Abbay (Blue Nile) from Lake Tana.
Fed by powerful tributaries, the Abbay runs through a deep canyon before entering onto the plains of
the Sudan. The Blue Nile gorge has its own unique habitat with a narrow fringe of forest and scrub
lining the river in its upper reaches (Rzóska, 1978). From Khartoum downstream, the Nile valley is a
broad flat plain over 300 km wide at its narrowest point and almost devoid of vegetation throughout
the desert of northern Sudan. However, drought‐tolerant plant species, such as Polygonum spp.
and Potamogeton spp., occur in local stands and create narrow fringes along the mainstem Nile.
Stands of Phragmites proliferate where the Nile flows into Lake Nubia (Dumont 1986; Hughes &
Hughes, 1992).
At the northern edge of the ecoregion, west of the Nile mainstem, Lake Qârûn lies at the bottom of
the Fayum depression, which is 71 km long and 20 km wide. Once fed by the Nile, Lake Qârûn now
receives most of its flow as runoff from surrounding irrigated lands. As a result, its waters are becoming
increasingly saline (Collins, 2002).
Fish Fauna
The lower Nile River provides vital habitat within a desert environment for an array of fish and other
wildlife. Over 70 species of fish live in the ecoregion, many belonging to the families Alestiidae,
Cichlidae, Citharinidae, Claroteidae, Cyprinidae, Mochokidae, and Mormyridae.
Other aquatic biota
In the northeastern corner of the ecoregion, Lake Qârûn supports large number of waterbirds; several
grebes, as well as Aythya fuligula, Fulica atra, and Anas crecca, are abundant. Three species of turtle
live within the lower Nile.
Of the nearly 50 indigenous taxa of molluscs within the entire Nile basin, 9 species are endemic. Within
the Lower Nile ecoregion, 15 gastropods and 9 bivalves occur. Palearctic and Afrotropical species of
gastropods overlap in the Saharan portion of the Nile basin, as well as in parts of the headwaters of
the Blue Nile. For example, Brown (1994) (Brown 1994) observed Armiger cristaliving together
with Ceratophallus natalensis and Segmentorbis angustus in a pool alongside a tributary to the Blue
Nile River.

Deliverable No. 1.3
26
Delineation
The headwaters of the Blue Nile and Atbara Rivers are also separated into the Ethiopian Highlands
ecoregion due to their swift‐flowing, steep nature and different aquatic fauna. The valley of the Nile
River was inundated by the Tethys Sea up through the Cretaceous period (approximately 65 million
years ago) (Dumont 1986). Five geologic phases of the Egyptian Nile can be distinguished, each
separated by a dry period of no flow: the Eonile, Palaeonile, Protonile, Prenile, and the present Neonile
(Rzóska, 1978; Dumont, 1986). The present Nile valley developed at the end of the Miocene. The rise
of the high volcanic plateaus in Ethiopia, probably during the Oligocene, is responsible for the origin
and direction of the Blue Nile and the Atbara River (Rzóska 1978). Tectonic movement and climatic
changes have changed the Nile’s course and flow many times. The Nile Basin has few endemic fish,
due to the frequent cessation of flow that inhibited the evolution and persistence of aquatic
species (Beadle, 1981; Dumont, 1986).
3.2.8 Shebelle – Juba (531)
The xeric systems of this ecoregion include the Wabi Shebelle and Juba basins with the ecoregion
extending from Kenya to Somalia along the coast of the Indian Ocean and inland to the Ethiopian
Highlands. During flooding, the rivers often spill over their banks and inundate adjacent floodplains.
All of the lower Wabi Shebelle and adjacent areas are very dry. The most reliable month for rain is
April, followed by May, whereas no rain falls between October and September. The hottest months
are February and March.
Freshwater habitats
Draining from the southeastern escarpment of the eastern Ethiopian highlands, the Wabi Shebelle and
the Fafan Rivers flow through the Somalian desert, although they do not reach the Indian Ocean. The
Wabi Shebelle is the major river of the central Somali region. Rising between the Arsi and Bale
Mountains, it flows in a southeasterly direction to Somalia. In its lower section, the Wabi Shebelle and
its main seasonal tributary from the east, the Fafan, cut through a series of wide, flat shelves of
sedimentary rocks made of sandstone, limestone, and gypsum. Wabi Shebelle, with a catchment area
of 205,407 km2, winds a length of 1340 km inside Ethiopia, and a further 660 km in Somalia (Ethiopian
Mapping Authority 1988). The Wabi Gestro, the Ghenale River, and the Dawa Parma River drain the
southwestern escarpment of the eastern Ethiopian highlands. These rivers unite and become the Juba
River, which eventually drains into the Indian Ocean (Westphal, 1975). These Juba tributaries arise just
east of Abaya and Chamo Lakes, but are separated from the lake drainages by a high mountainous
divide. According to Roberts (1975), midway between the lower courses of the Wabi Shebelle and the
Juba there is a low‐lying limestone plateau with extensive underground waterways radiating out from
it.

Deliverable No. 1.3
27
Terrestrial Habitats
Floodplains are often covered in a tangled growth of small bushes and herbs, which include wild
relatives of cotton. Large trees are not naturally found on the floodplains, but heat‐tolerant species,
including Hyphaene thebaica, have been planted in settlements.
Figure 18: Shebelle‐ Juba freshwater ecoregion
Fish Fauna
The rivers in this ecoregion are believed to host many Nilo‐Sudanic fishes similar to the southern rift
valley lakes (Lakes Chamo and Abaya). It is believed that these lakes and the Shebelle‐Ghenale River
basins had former connections with the upper White Nile as recently as 7,500 years ago (Roberts 1975;
McClanahan & Young 1996). Some of the rivers in this ecoregion (e.g., Ghenale River) support
abundant populations of fish.
Endemic fish, including Bagrus urostigma, Labeo boulengeri, Labeo bottegi, and Synodontis geledensis,
live in the rivers of this ecoregion. Most of the Nilotic species found in Lake Abaya, with the exception
of Hyperopisus bebe, are also present in the Wabi Shebelle‐Juba drainage (Roberts, 1975). Another
important feature of the area is the presence of subterranean waterways, which are inhabited by the
endemic monotypic fish genera Uegitglanis and Phreatichthys. Both the clariid catfish (U.
zammaranoi) and the cyprinid (P. andruzzii)lack visible eyes and are depigmented and scaleless.
The vegetation alongside the middle section of the Ghenale River also supports populations of the
vulnerable Prince Ruspoli’s turaco (Tauraco ruspolii), white‐winged collared‐dove (Streptopelia
reichenowi), and Jubaland weaver (Ploceus dicrocephalus). Along the coast in Somalia are several areas
of importance for waterbirds, for example the Jasiira lagoon is known to support congregations
of Phoenicopterus ruber and Egretta gularis.
A continuous escarpment, running in a wide curve from the Kenyan border to northern Somalia, forms
the western and northern borders of this ecoregion, while the southwestern part of this escarpment

Deliverable No. 1.3
28
forms the eastern wall of the Rift Valley. The escarpment rises northwards and attains its maximum
elevation of over 3,000m near the Chilalo Massif in Ethi opia. Several endemic fishes live in the streams
and subterranean waters of this xeric ecoregion.
Delineation
A continuous escarpment, running in a wide curve from the Kenyan border to northern Somalia, forms
the western and northern borders of this ecoregion, while the southwestern part of this escarpment
forms the eastern wall of the Rift Valley. The escarpment rises northwards and attains its maximum
elevation of over 3,000m near the Chilalo Massif in Ethiopia. Several endemic fishes live in the streams
and subterranean waters of this xeric ecoregion
The native freshwater flora and fauna of this vast ecoregion has been poorly investigated. Although
the riparian vegetation has been described to some extent, little is known about the upland vegetation.
Without better information, identification of conservation priorities is difficult.

Deliverable No. 1.3
29
4 Terrestrial ecoregions
There are several alternative formal naming schemes for the Earth's terrestrial ecoregions; e.g.
Usdvardy (1975), Encyclopaedia of Earth Ecoregions, Wikipedia Ecoregions, TNC Ecoregion, Global 200
Ecoregions and others.
One of the most widely used, developed by the World Wildlife Foundation, recognizes 867 separate
ecoregions. The ecoregions are categorized within 14 biomes and eight biogeographic realms to
facilitate representation analyses.
The Encyclopaedia of Earth defines the ecoregion approach as following: “An ecoregion is a contiguous
area characterized by well‐defined similarity in flora and fauna as well as geomorphology, climate and
soils. Ecoregions are generally relatively large geographic units on the order of 50,000 square
kilometres or more. Ecoregions may be terrestrial or marine, and do not recognize any political
boundaries or landscape alterations by humans. Generally an ecoregion is depicted by a geographic
descriptor coupled with a biome identity, further articulating one or more specific climatic or dominant
plant community appellations”.
Figure 19: Ecoregions of the world: 14 biome (colour) and 8 biographic realms to facilitate representation analyses (Source: WWF)
With this classification (http://www.globalspecies.org/ecoregions/display/AT0112), Ecoregions of
Ethiopia are classified as follows:

Deliverable No. 1.3
30
Ethiopian montane forests
The Ethiopian Montane Woodland ecoregion is biodiverse, poorly known and highly threatened. The
rugged topography of this ecoregion rings the highlands of Ethiopia and Eritrea, extending to outlying
massifs in Sudan. Formed by volcanic forces 75 million years ago, these highlands were covered with
Eurasian tundra‐like vegetation during the last Ice Ages. Today, remnant patches of natural vegetation
consist mostly of podocarps and juniper forests, with some acacias found at lower elevations. While
soils are rather infertile, this area is densely populated and most land has been converted to
agriculture. Notable endemics found here include the yellow‐throated serin and Prince Ruspoli's
turaco. Many of the endemic species are threatened due to the loss of their habitat.
This ecoregion is highly biodiverse, relatively poorly known and highly threatened. It is mainly found
on the margins of the highlands of Ethiopia and Eritrea. The altitudinal limits of the ecoregion vary
from one locality to another depending upon annual precipitation, but are generally between 1,100
and 1,800 meters (m). From May to October, winds blow from the southwest and bring rainfall to the
Ethiopian portion of the ecoregion. During the rest of the year, onshore winds from the Red Sea bring
moisture to the Eritrean side of the mountains. Rainfall varies from 600 millimetres (mm) in the driest
sites to more than 1,500 mm in wetter areas. Humidity is sometimes higher than would be expected
from these figures, due to cloud precipitation and local interactions between topography and weather.
Unlike the moist equatorial mountains, the effects of cold descend further down on these dry
highlands. Temperatures vary according to the season and elevation, but mean maxima lie between
18°C and 24°C. Mean minima are between 12°C and 15°C.
Ancient Precambrian basement rocks form the substrate of the montane forests in southwestern
Ethiopia and Eritrea. The topography is generally rugged, and soils are rather infertile. The main
Ethiopian and Eritrean dome began to rise 75 million years ago, eventually dividing into two halves,
the northern and southern highlands. A turbulent volcanic period ended four to five million years ago,
followed by climatic fluctuations in the Pliocene and Pleistocene. Glaciers formed on the peaks of the
Ethiopian highlands while surrounding areas, including this ecoregion, were covered with vegetation
similar to Eurasian tundra. Separated by the Great Rift Valley, the northern and southern highlands
were colonized by new species from different directions. The jebels and escarpments along the Red
Sea linked Eritrea and northern Ethiopia with the Palearctic region while southern Ethiopia had a rift‐
wall connection to the Horn of Africa. Both the western and eastern highlands were invaded by tropical
species that could penetrate the Nile floodplains in the west or the Kenyan deserts in the south.
Despite the climatic differences, the surrounding lowlands provided the most consistent source of new
species, so that these highlands show both Afrotropical and Palearctic influences.

Deliverable No. 1.3
31
Figure 20: Ethiopian montane forest ecoregion
This ecoregion is based on the ‘East African evergreen and semi‐evergreen bushland and thicket’ and
‘cultivation and secondary grassland replacing upland and montane forest’ vegetation units. The
cultivation and secondary grassland areas are included in an effort to cover potential vegetation. The
ecoregion lies between 1,100 meters (m) and 1,800 m in elevation.
Ethiopian montane grasslands and woodlands
During the last Ice Age, this entire ecoregion would have been similar to the Eurasian tundras, while
still higher elevations were capped with glaciers. As the climate warmed, these highlands were
recolonized, resulting in a biota with a combination of Palearctic and Afrotropical influences. Ranging
up to 3,000 m, the montane vegetation includes Hagenia, Podocarpus and Juniperus, but intact
vegetation is increasingly fragmented. The region is densely populated because it contains the best
arable land in Ethiopia. A variety of Ethiopian endemics can be found, including the critically
endangered Walia ibex (Capra walie) and endangered mountain nyala (Tragelaphus buxtoni). Plant
endemism in this region peaks in the forest/ woodland/ grassland complex.
This is a biologically rich and severely threatened ecoregion (94,700 square miles) that covers the
majority of two Ethiopian mountain massifs (Eastern and Western), separated by a part of the African
Great Rift Valley. This ecoregion ranges from 1,800 m to 3,000 m in elevation, with montane forest at
lower altitudes and Afroalpine habitat higher up. The climate of these highlands is greatly affected by
their topography, and also by the movement of the Inter Tropical Convergence Zone (ITCZ). As the ITCZ
moves north between May and October, warm moist air is drawn from the Indian Ocean so that rain
falls on the southern slope of the Ethiopian Highlands. During the remainder of the year, the ITCZ lies
south of the highlands and the winds are from the Red Sea to the north and east. These winds typically
contain less moisture, which mainly falls on the northern side of the highland massif. Overall, the
highest annual rainfall (up to 2,500 mm) is on the southwestern scarp faces of the highlands, which
support montane or transitional forests, while over most of the ecoregion the rainfall is around 1,600
mm annually.

Deliverable No. 1.3
32
Figure 21: Ethiopian montane grasslands and woodlands
Geologically the area consists of a Precambrian basement capped in most places by thick Tertiary
basaltic lava flows (the Trap Series). The ancient Precambrian rocks form the substrate of the montane
forests in southwestern Ethiopia and Eritrea. Mesozoic rocks form the surface outcrops of the
southeastern highlands of Ethiopia, while Tertiary basalts form the surface rocks of the remainder of
the Ethiopian and Eritrean massifs. The latter attain a thickness of 3,000 to 3,500 m in the Simien
Mountains. Forest soils in Ethiopia are mainly ferrosols derived from these volcanic substrates.
White (1983) nests this ecoregion within a larger ‘undifferentiated montane vegetation’ unit. The
ecoregion roughly follows the 1,800 m contour for the lower elevation and 3,000 m contour for the
upper elevation. Although the South and Central Highlands are recognized as two areas of bird
endemism (Stattersfield et al., 1998), they contain floral and faunal similarities (WWF, 1998).
Ethiopian montane moorlands
While the Ethiopian Montane Moorlands ecoregion makes up only 2% of the total land area in Ethiopia
(9,700 square miles), it contains 80% of land above 3000 m in the Afrotropical realm. Split into northern
and southern massifs, these highlands were formed by turbulent volcanic forces that ceased only 4 to
5 million years ago. At the end of the last Ice Age, montane species were restricted to higher altitudes
by the warming climate. High levels of endemism are found here and the region’s biota demonstrates
evolutionary links to both the Palaearctic and the Afrotropical realms. The vegetation, known as wurch
to Ethiopians, consists of grassland and moorland with abundant herbs. Most plant species (many of
which are endemic) show adaptations to the extreme conditions found at high altitudes. Stretching
out across Ethiopia, these pockets of high altitude vegetation harbor the last populations of the
critically endangered Ethiopian wolf (Canis simensis) and several small mammal endemics.
Ecoregion Delineation
This ecoregion forms part of the Afroalpine center of plant diversity across East and Northeast Africa
(WWF and IUCN 1994). The lower boundaries are derived from White’s ‘altimontane’ vegetation unit,
roughly following the 3,000 m contour. The Ethiopian Montane Moorlands are poorly developed
compared to other East African moorland areas, lacking typical Afroalpine species like Dendrosenecio.

Deliverable No. 1.3
33
However, they possess considerable endemism and contain affinities to the Palearctic realm, such as
species of Rosa and Primula (Hedberg & Hedberg, 1979, Vuilleumier &Monasterio, 1986).
Figure 22: Ethiopian montane moorlands ecoregion

Deliverable No. 1.3
34
Ethiopian xeric grasslands and shrublands
The Ethiopian xeric grasslands and shrublands is an arid, semi‐desert ecoregion bordering the Red
Sea and the Gulf of Oman. This ecoregion lies mainly between sea level and 800 meters (m) elevation.
There are, however, many hills and massifs, which range up to 1300 m as well as outstanding fault‐
induced depressions, such as the Danakil, lying as low as 155 m below sea level. This region is extremely
active tectonically, experiencing many earthquakes and intermittently active volcanoes. Rainfall is
very low and yearly averages range from 100 to 200 (mm), with less rain falling closer to the coast.
There are many species of interest, including the endemic Archer's lark (Heteromirafra archeri), a
species of dragon tree (Dracaena ombet), and a large suite of desert ungulates, including the last
viable population of African wild ass (Equus africanus somalicus).
Figure 23: Ethiopian xeric grasslands and shrublands ecoregion
This ecoregion extends inland from the Red Sea and the Gulf of Oman, including the Dahlak
Archipelago and other islands, stretching from the Sudanese‐Eritrean border, south
through Ethiopia to Djibouti and eastwards into Somalia, in the Somaliland region of the country.
While it mainly lies between sea level and 800 m, there are many arid hills and massifs up to 1300 m.
Higher massifs such as the Goda and Mabla in Djibouti are considered to be outliers of the Ethiopian
Montane Forest ecoregion. There are also fault‐induced depressions, such as the Danakil Depression
and Lac Assal, lying as much as 160 m below sea level. Elevation generally increases westward towards
the Ethiopian and Eritrean highlands. The region is extremely active tectonically, and it experiences
many earthquakes associated with the continuing enlargement of the Rift Valley. Volcanoes in the
ecoregion are also intermittently active. Basement rocks are composed mainly of Tertiary lava flows,
although there are also Quaternary basinal deposits at the northern end and pre‐Cretaceous basinal
deposits on the northern coast of Somalia. Soils developed over the lava deposits are mainly lithosols,
while regosols are predominant on the Quaternary and pre‐Cretaceous basinal deposits. There are
very few permanent watercourses. The most notable is the Awash River of Ethiopia that terminates in
a series of lakes near the border with Djibouti.

Deliverable No. 1.3
35
The climate is very hot and dry. Mean annual rainfall varies from less than 100 mm close to the coast
to around 200 mm further inland. Mean minimum temperatures range from 21° to 24°C, and mean
maximum temperature is around 30°C. Phytogeographically, White (1983) regarded this ecoregion as
part of the Somali‐Masai regional center of endemism and mapped the vegetation as ‘Somalia‐Masai
semi‐desert grassland and shrubland’. Along the coast, mangroves occur in muddy areas, primarily
around wadi "outwashes" and inlets, while further inland, vegetation changes to grass shrub
steppe. Acacia melliferaand Rhigozum somalense dominate the basaltic lava fields while
scattered Acacia tortilis, A. nubica, andBalanites aegyptiaca can be found in the sandy plains. Stands
of Hyphaene thebaica occur in depressions and along wadis.
The boundaries of this ecoregion follow the ‘Somalia‐Masai semi‐desert grassland and shrubland’
vegetation unit of White (1983), from just north of the Sudan/Eritrea border to the extremely narrow
coastal strip along the northern coast of Somalia (Somaliland). This eastern border follows the range
limit of some species like the Dorcas gazelle (Gazella dorcas), and encompasses the range of others,
such as the African wild ass (Equus africanus). White’s (1983) larger ‘Somalia‐Masai semi‐desert
grassland and shrubland’ unit that extends further east along the Horn of Africa was separated into
distinct ecoregions based on their unique plant and vertebrate compositions.
East Sudanian savanna
The East Sudanian Savanna is a hot, dry, wooded savanna composed mainly of Combretum and
Terminalia shrub and tree species and tall elephant grass (Pennisetum purpureum). The habitat has
been adversely affected by agricultural activities, fire, clearance for wood and charcoal, but large
blocks of relatively intact habitat remain even outside protected areas.
Figure 24: East Sudanian savanna ecoregion
This ecoregion lies south of the Sahel in central and eastern Africa, and is divided into a western block
and an eastern block by the Sudd swamps in the Saharan Flooded Grasslands ecoregion. The western
block stretches from the Nigeria/Cameroon border through Chad and the Central African Republic to

Deliverable No. 1.3
36
western Sudan. The eastern block is found in eastern Sudan, Eritrea, and the low‐lying parts of western
Ethiopia, and also extends south through southern Sudan, into northwestern Uganda, and marginally
into the Democratic Republic of Congo around Lake Albert.
Topographically, the ecoregion is flat, mainly lying between 200 m and 1,000 m in altitude, although
elevation rises slightly in western Ethiopia and around Lake Albert. The climate is tropical and strongly
seasonal. Mean monthly maximum temperatures range from 30° to 33°C and mean minimum
temperatures are between 18°C and 21°C. The annual rainfall is as high as 1,000 mm in the south, but
declines to the north with only 600 mm found on the border with the Sahelian Acacia Savanna. Rainfall
is highly seasonal, and during the rainy season from April to October, large areas of southern Chad and
northern parts of the Central African Republic become totally inundated and inaccessible. During the
dry season most of the trees lose their leaves and the grasses dry up and may burn.
Geologically, the ecoregion overlies a mixture of Precambrian basement rocks, and a number of post‐
Jurassic sedimentary basins. The soils are mainly ultisols and alfisols in the south with entisols in the
north. Some oxisols and vertisols are also found in the east. The ecoregion is sparsely populated, with
typical population densities ranging between 1 to 5 people/km2, although there may be as many as 20
to 30 people/km2 in some places.
White (1983) classified this region phyto‐geographically within the Sudanian regional centre of
endemism, as it supports more than 1,000 endemic species of plants. The vegetation is mapped as
undifferentiated woodland that comprises trees, which are mainly deciduous in the dry season, with
an understory of grasses, shrubs and herbs. Typical trees in the western block of the ecoregion include
Anogeissus leiocarpus, Kigelia aethiopica, Acacia seyal and species of Combretum and Terminalia. In
the eastern block woody vegetation is dominated by Combretum and Terminalia species, as well as
Anogeissus leiocarpus, Boswellia papyrifera, Lannea schimperi and Stereospermum kunthianum.
This ecoregion, along with the West Sudanian Savanna, forms part of the Sudanian regional center of
endemism, and contains two vegetation units, the ‘Sudanian undifferentiated woodland’ and
‘Sudanian woodland with abundant Isoberlina’ (White 1983). Although these two units differ in the
presence of Isoberlina, with the northern unit being slightly drier, they share similar animal
assemblages. The West and East Sudanian Savannas are also similar in terms of their broader species
assemblages, but they were split into two separate ecoregions near the Mandara Plateau because a
number of plant taxa do not cross this boundary (WWF, 1998). Several modifications to White’s
boundaries have been made, including an extension of the Sahelian Acacia Savanna below Lake Chad,
and a northern boundary considerably further south than White’s boundary (WWF, 1998). Edaphic
grassland and communities of Acacia and broadleaved trees identified by White have also been largely
excluded. The eastern portion of this ecoregion forms an extension of undifferentiated woodland,
following White’s ‘Ethiopian undifferentiated woodland’ and ‘Ethiopian transition from
undifferentiated woodland to Acacia deciduous bushland and wooded grassland.’ This area was
extended to include ‘Sudanian undifferentiated woodland’ south towards Lake Albert and Mount
Elgon. Udvardy (1975) delineates a biogeographic boundary between the Western and Eastern Sahel,
which would effectively split the western section of this ecoregion in half.

Deliverable No. 1.3
37
Victoria basin forest‐savanna mosaic
The rolling hills and plateaus of the Victorian Basin Forest‐Savanna Mosaic ecoregion are a unique
landscape where species from West African forest ecosystems converge with those from east African
forest‐savanna mosaics. The region’s scattered lakes, rivers, and marshes add to the great diversity of
habitats supporting a wide variety of species.
Figure 25: ictoria basin forest‐savanna mosaic (http://www.globalspecies.org/ecoregions/display/AT0721)
Sahelian Acacia savanna
Spanning the entire African continent, the Sahelian Acacia Savanna stretches in a continuous band
from the Atlantic Ocean to the Red Sea. Although not particularly rich biologically, these savannas once
supported a large and diverse ungulate community. The first European explorers to visit the region
found vast herds of game, even larger in number than those of eastern and southern Africa. Sadly,
these herds have been reduced to mere remnants due to nearly a century of unregulated over‐hunting
with modern firearms and vehicles, coupled with habitat loss. This ecoregion includes some of the
world’s poorest countries, which have scarce resources for conservation. Existing conservation efforts
are therefore inadequate and the biological values of the ecoregion remain highly threatened.

Deliverable No. 1.3
38
Figure 26: Sahelian Acacia savanna ecoregion
The Sahelian Acacia Savanna stretches across Africa from northern Senegal and Mauritania on the
Atlantic coast to Sudan on the Red Sea, varying in width from several hundred to over a thousand
kilometres. The word "sahel" means "shore" in Arabic and refers to the transition zone between the
wooded savannas of the south and the true Sahara Desert. The ecoregion thus lies south of the
Southern Saharan Steppe and Woodland Ecoregion and north of the West and East Sudanian Savanna
Ecoregions.
The topography is mainly flat and the majority of the ecoregion lies between 200 m to 400 m in
elevation. The few isolated mountain massifs rising from this plateau, some over 3,000 m high, have
been assigned to other ecoregions due to their distinctive fauna and flora. The climate is tropical, hot,
and strongly seasonal. The monthly mean maximum temperatures vary from 33° to 36°C and monthly
mean minimum temperatures are between 18° to 21°C. The annual rainfall is around 600 mm in the
south of the ecoregion, but declines rapidly to the north to around 200 mm. Most rain falls in the
summer months of May to September, followed by a 6 to 8 month dry season, during which time the
woody vegetation loses its leaves and the grasses dry up and may burn. The movements of the
Intertropical Convergence Zone (ITCZ) determine the quantity of rainfall in a particular year ‐ if it
penetrates far to the north there will be a long rainy season and good rains; if it does not move
sufficiently far north, then the rains may fail totally. During the winter, hot dry winds (known in much
of West Africa as the "Harmattan") blow from the north, often bringing dust and sand from the Sahara
with them.
Geologically, the ecoregion mainly overlies a number of post‐Jurassic sedimentary basins, but the
higher land found between the Central African Republic and Sudan is made up of Precambrian
basement materials. There are also recent deposits from huge lakes, which were present during the
pluvial periods of a few thousand years ago, cantered on present day Lake Chad and the Inner Niger
Delta. These lakes have been drying for thousands of years and today cover only fractions of their
former extent, although the region might have been even dryer between 12,000 and 20,000 years BP.
Other smaller lakes would have been present in the past. The soils of the ecoregion are mainly entisols,
with some aridisols found towards the north and alfisols on the highest land. Throughout much of the
region the soils are highly permeable and permanent surface water points or watercourses are rare.

Deliverable No. 1.3
39
The Sahelian Acacia Savanna ecoregion follows two of White’s vegetation units: the ‘Sahel Acacia
wooded grassland and deciduous bushland’ and the ‘Northern Sahel semi‐desert grassland and
shrubland’ (White, 1983). Two small modifications to White’s linework include moving the Adrar de
Iforas from this ecoregion to the West Saharan Montane Xeric Woodland, and extending the eastern
boundary of the ecoregion to the Ethiopian Highlands (WWF, 1998). Udvardy (1975) splits this region
into the Western and Eastern Sahel biogeographical provinces.
Somali Acacia‐Commiphora bushlands and thickets
In the heart of these wide‐sweeping grasslands and associated Acacia‐Commiphora woodlands, the
world’s most spectacular migration of large mammals occurs each year. Wildebeests, plains zebras,
and Thomson’s gazelles traverse the Greater Serengeti Ecosystem, triggered by cyclical wet and dry
seasons. Parts of the world famous Serengeti National Park and the Ngorongoro Conservation Area are
located in this ecoregion; both have been designated as World Heritage Sites and Biosphere Reserves.
Of the three Acacia‐Commiphora Bushland and Thicket ecoregions, this southern unit receives the
most rain and is closely associated with the Serengeti Volcanic Grasslands. The main threats to the
ecoregion are the same as other savanna areas in the region, poaching of large mammals for body
parts and meat, and expansion of pastoralism and agricultural use of the area with associated loss of
tree cover.
Figure 27: Somali Acacia‐Commiphora bushlands and thickets ecoregion (http://www.worldwildlife.org/ecoregions/at0716)
The ecoregion is located in northern and central Tanzania, extending into southwestern Kenya, around
the eastern margins of Lake Victoria. The ecoregion forms the southern border of White’s (1983)
phytogeographical classification for the Somali‐Masai Acacia‐Commiphora deciduous bushland and
thicket. The predominant plants include species of Acacia, Commiphora, and Crotalaria and the grasses

Deliverable No. 1.3
40
Themeda triandra, Setaria incrassata, Panicum coloratum, Aristida adscencionis, Andropogon spp., and
Eragrostis spp. (White, 1983). Within Tanzania, the ecoregion is bisected by two patches of Serengeti
Volcanic Grassland and patches of East African Montane Forest. The volcanic grasslands are an integral
habitat of the greater Serengeti ecosystem. However, the grasslands are considered a separate
ecoregion due to their unique grassland communities, found only on the fine volcanic soil, or "vertisol."
The habitat transitions to miombo woodland towards the south, more Acacia‐Commiphora Bushland
and Thicket towards the north, and Zanzibar‐Inhambane Coastal Forest‐Savanna Mosaic towards the
east. The western portion of the ecoregion is included in the greater Serengeti ecosystem.
Topographically, the ecoregion is situated on the Central African Plateau and slopes upward from east
to west. Elevation ranges from 900 m in the Speke Gulf up to 1,850 m in the Gol Mountains. The
majority of the ecoregion falls between 900 and 1,200 m.
The climate of the region is tropical with seasonal rain that falls in a bimodal pattern. The long rains
occur from March to May and the short rains from November to December. Mean rainfall is 600 to
800 mm annually through most of the region. Extremes include 500 mm in the dry southeastern plains
and 1,200 mm in the northwestern region located in Kenya. Rainfall is variable such that the short rains
may fail in a given year or rain may occur between the two rainy seasons, thereby joining the two.
Temperatures are moderate with mean maximum temperatures as high as 30°C at lower elevations
and as low as 24°C at the highest parts of the ecoregion. Mean minimum temperatures are between
9° and 18°C, and normally between 13° to 16°C.
During the long dry season (August to October), the grasslands can become extremely parched, and
many of the trees and bushes lose their leaves. Fires occur naturally in the ecosystem. Both fire and
elephant browsing play an important role in converting dense thicket and bushland into grassland.
However, a large number of fires are started by pastoralists to promote new vegetative growth for
their livestock.
Geologically, the entire area is underlain by Precambrian basement rocks (up to 2.5 billion years old)
that have been deformed, then eroded over hundreds of millions of years. Ultisols and alfisols lay over
basement rocks, with patches of vertisols close to Lake Victoria. The hills in the ecoregion are formed
of more recent sedimentary rocks, and some are recent volcanics with some active volcanoes. In many
areas late Precambrian outcrops of granitic gneisses and quarzite project from the surface as inselbergs
(locally called kopjes).
This ecoregion forms the southern part of the ‘Somali‐Masai Acacia‐Commiphora bushland and thicket’
vegetation unit of White (1983). It contains higher levels of rainfall than the areas further north, and
floristic and faunistic patterns that are distinct. It is divided from the Northern Acacia‐Commiphora
Bushland and Thickets ecoregion roughly by the chain of mountains along the Kenya‐Tanzania border.
This represents the extent of some species like the gerenuk (Litocranius walleri), which is present in
the north, but is absent in much of the south. The southern boundary of this ecoregion directly follows
White (1983).

Deliverable No. 1.3
41
Northern Acacia‐Commiphora bushlands and thickets
The ecoregion, comprised of semi‐arid mixed woodland, scrub and grassland, is reasonably protected
within a well‐functioning system of national parks and other reserves. However, numbers of humans
and livestock are increasing outside protected areas, and nomadic pastoralism is declining in favor of
settlement, causing environmental degradation through heavy grazing and agricultural expansion.
Water is always in short supply in this region, where one or both rainy seasons commonly fail. Certain
species such as Grevy’s zebra (Equus grevyi) have undergone severe declines because of competition
with livestock for, and often exclusion from, water supplies. Poaching also threatens some of the large
herbivores found here, particularly the black rhinoceros (Diceros bicornis) and, until recently, the
African elephant (Loxodonta africana).
Figure 28: Northern Acacia‐Commiphora bushlands and thickets ecoregion
This ecoregion extends from the southeast corner of Sudan and northeast Uganda, through much of
lowland Kenya, reaching as far as the border with the Northern Zanzibar‐Inhambane Coastal Forest
Mosaic. To the north, it is replaced by drier savanna and semi‐desert vegetation. To the south it grades
into the southern Acacia‐Commiphora bushland and thicket around the Kenya‐Tanzania border.
Climatically, the area falls within the seasonal tropics, with seasonality controlled by movement of the
Inter Tropical Convergence Zone. Mean maximum temperatures are 30°C in the lowlands, falling to
around 24°C in the higher areas. Mean minimum temperatures range from 18 to 21°C. Annual rainfall
ranges from 200 millimetres (mm) in the drier areas near Lake Turkana, to about 600 mm closer to the
Kenyan Coast. It is strongly seasonal, with most precipitation occurring in the long rains, typically from
March to June, and less falling in the short rains of October to December. The timing and amounts of
rainfall vary greatly from year to year, and it is fairly common that one, or both, rainy seasons fail. In
Turkana, Kenya, and southern Sudan, there is typically one rainy period per year. During drier times
the desiccated vegetation becomes highly flammable and large areas of the ecoregion burn every year.
Geologically, the ecoregion is situated on a mixture of basement rocks. These range from Precambrian
basement, through Tertiary volcanic lavas, to Quaternary basin and dune formations. These sediments
outcrop at the surface in many places due to the shallow soils. Apart from these outcrops, the

Deliverable No. 1.3
42
ecoregion is generally level and gently undulating, with lowest elevation in the east, north and
northwest (elevations of 200 meters (m) to 400 m), increasing towards the south and southwest,
where elevations rise up to 1,000 m, but there are a number of notable mountains, such as Moroto in
Uganda, which exceeds 3000 m, and several in Turkana which are above 2000 m. The soils of the
southeast portion of the ecoregion are mainly aridisols, with entisols around Lake Turkana Basin. Along
the western margins of the ecoregion, vertisols can also be found.
The ecoregion is part of White’s Somali‐Masai regional center of phytogeographical endemism and is
predominately Acacia‐Commiphora bushland and thicket. Common plant genera include Acacia,
Commiphora, and Boswellia, and Aristida, Stipa, and Chloris grasses.
This ecoregion forms the central portion of the ‘Somali‐Masai ''Acacia‐Commiphora'' bushland and
thicket’ vegetation unit of White. This larger unit was separated into three ecoregions based on
different bioclimatic and associated floral and faunal patterns. The ‘East African evergreen bushland
and secondary Acacia wooded grassland’ of White was included to better reflect potential vegetation.
The ecoregion is bound by the alluvial plains and rocky plateau of the Masai Xeric Grassland and
Shrubland ecoregion and the Tana River to the northeast, which represent the extent of some species,
such as the common eland (Taurotragus oryx), Bohor reedbuck (Redunca redunca) and Burchell’s zebra
(Equus burchelli). To the south it is divided from the Southern ''Acacia‐Commiphora'' Bushland and
Thicket ecoregion roughly by the chain of mountains along the Kenya‐Tanzania border. This represents
the extent of some species like the gerenuk (Litocranius walleri), which is present in the north, but is
absent in much of the south. Additional studies of herbs, birds and plants may help to better define
this ecoregion.
Masai xeric grasslands and shrublands
This ecoregion lies in northern Kenya, and includes a mix of desert, savanna woodland, wetland, and
bushland. It is known for its cultural history and fossil species, including the early hominids Homo
habilis and Homo erectus, as well as a giant tortoise and giant crocodile. Present day species found in
this ecoregion include three endemic species of amphibians and one endemic turtle associated with
Lake Turkana. In the drier areas several large mammal species occur such as cheetah, lion, elephant,
beisa oryx, Grevy’s zebra and reticulated giraffe. Drought and pastoralism have had a substantial
impact on the habitats of the area, some of which have been reduced to a desert‐like state, and the
populations of many of the large mammals have been greatly reduced. Black rhinoceros (Diceros
bicornis) has been extirpated from the ecoregion.

Deliverable No. 1.3
43
Figure 29: Masai xeric grasslands and shrublands ecoregion
This ecoregion is located in the northern part of Kenya and extreme southwestern Ethiopia. It
encompasses most of Lake Turkana and the Omo River Delta, and grades into the savanna woodlands
of the Northern Acacia‐Commiphora Bushland and Thicket ecoregion to the west and the Somali
Acacia‐Commiphora Bushland and Thicket to the east. Mostly lying between 200 and 700 m elevation
and gently undulating, it includes the Chalbi Desert, a large flat depression between 435 m and 500 m
formed from the bed of an ancient lake.
The climate is hot and dry over most of the year, with mean maximum temperatures of around 30° C
and mean minimum temperatures between 18° C and 21° C. There is a short wet season between
March and June as the Intercontinental Convergence Zone moves north. Mean annual rainfall is
between 200 and 400 mm. Geologically, the ecoregion is located on an outlier of the Tertiary volcanic
materials that make up the Ethiopian massif. The soils of the area are complex, and vary from bare
rocks and lithosols, with solonchaks, yermosols, and regosols indicating the general desiccation of the
area.
This ecoregion forms the southern outlier of White’s ‘Somalia‐Masai semi‐desert grassland and
shrubland,’ including the area around Lake Turkana and the Chalbi Desert. Although, it contains a
similar vegetation structure to the Ethiopian Xeric Grassland and Shrubland ecoregion further north, it
was elevated to ecoregion status due to its disjunct position, arid nature, and elements of savanna
woodland communities.

Deliverable No. 1.3
44
5 Other Ethiopian regional typologies
Ecosystems of Ethiopia
Ethiopia is endowed with diverse ecosystems in which diverse flora and fauna as well as microbial
resources are found. The major ecosystems include: Afroalpine and subafroalpine, montane dry forest
and scrub, montane moist forest, Acacia‐Comiphora woodland, Combretum‐Terminalia woodland,
Lowland humid forest, Aquatic, wetland, Montane grassland, and Desert and semidesert ecosystems
(http://et.chm‐cbd.net/biodiversity/ecosystems‐ethiopia).
Afroalpine and Subafroalpine Ecosystem
The areas which on the average higher than 3200 meters above sea level (m.a.s.l) are generally
referred to as the Afroalpine and Subafroalpine. The lower limit of the afroalpine belt falls at about
3500 m, while the upper limit of vascular plants lies around 5000 m , and subafroalpine areas ranges
between 3200‐ 3500 m. These areas include chains of mountains, mountain slopes and tops of highest
mountains in the country. The highest peak in Ethiopia is Ras Dashen (4533 m a.s.l), where an alpine
climate near 0°C persists all year round, sometimes even with a snow cover lasting a couple of days.
However, dry lowland savannas and deserts surround this moist highland area. Ethiopia has the largest
extent of afroalpine habitats in Africa (Yalden, 1983).
Montane Grassland Ecosystem
The montane grassland ecosystem is distinguished from other types of ecosystems by its physiognomy,
floristic composition and ecology. It consists of herbaceous stratum usually not higher than 30 – 80
cm, very rich in perennial grasses and species of Cyperaceae, but also with sub‐shrubs and perennial
herbs, among which bulbous and rhizomatous plants occur. According to White (1983), the montane
grassland of Ethiopia is a derived vegetation type, although small areas of the grassland may have
existed before human settlement.
Dry Evergreen Montane Forest and Evergreen Scrub Ecosystem
The Ethiopian highlands contribute to more than 50 % of the land area with Afromontane vegetation,
of which dry montane forests form the largest part (Yalden, 1983). The evergreen scrubland vegetation
occurs in the highlands of Ethiopia either as an intact scrub, i.e. in association with the dry evergreen
montane forest or usually as secondary growth after deforestation of the dry evergreen montane
forest. The Dry Evergreen Montane Forest and Evergreen Scrubland vegetations are the characteristic
vegetation types of this ecosystem.
Montane Moist Forest Ecosystem
The montane moist forest ecosystem comprises high forests of the country mainly the southwest
forests, which are the wettest, and also the humid forest on the southeastern plateau known as the

Deliverable No. 1.3
45
Harenna forest.The montane moist forest ecosystem is distinguished by supporting luxuriant growing
epiphytes Canarina, Orchids, Scadoxus and fern plants such as Platycerium and Drynaria. Mosses also
occur in the wettest portion of forests associated to major branches and barks of trees.
Acacia‐Commiphora Woodland Ecosystem
The Acacia‐Commiphora ecosystem is known for its varying soils, topography, and diverse biotic and
ecological elements. These plant species are with either small deciduous leaves or leathery persistent
ones. The density of trees varies from ‘high’, in which they form a closed canopy to scattered
individuals to none at all forming open grasslands. The grasses do not exceed more than one meter,
thus, no true savannah is formed.
Combretum‐Terminalia Ecosystem
This ecosystem is characterized by Cmbretum spp., Terminalia spp., Oxytenanthera abyssinica,
Boswellia papyrifera, Anogeissus lieocarpa, Sterospermem kuntianum, Pterocarpus lucens,
Lonchocarpus laxiflorus, Lannea spp. Albizia malacophylla and Enatada africana. These are small trees
with fairly large deciduous leaves, which often occur with the lowland bamboo‐ Oxytenanthera
abyssinica. The understory is a combination of herbs and grasses. The herbs include Justecia spp.,
Barleria spp., Eulophia, chlorophytum, Hossolunda opposita and Ledeburia spp. The grasses include
Cymbopogon, Hyparrhenia, Echinochla, Sorghum, Pennisetum, etc. Usually the herbs dominate the
ground layer at the beginning of the rainy season while grasses dominate toward the end of the rainy
season.
Lowland Tropical Forest Ecosystem
The characteristic species of this forest are Baphia abyssinica and Tapura fischeri (Friis, 1992). The
common species in the upper canopy include Celtis gomphophylla, Celtis toka, Lecaniodiscus
fraxinifolius, Zanha golungensis, Trichilia prieureana, Alistonia boonei, Antiaris toxicaria, Malacantha
alnifolia, Zanthoxylum lepreurii, Diospyros abyssinica, Milicia excelsa, Baphia abyssinica, Vepris
dainellii and Celtis zenkeri.
Desert and Semi‐desert Ecosystem
It is a very dry zone vulnerable to wind and water erosion even with little or no pressure on the
vegetation from grazing. The vegetation consists of deciduous shrubs, dominated by Acacia sp.
interspersed with less frequent evergreen shrubs and succulents. It has very variable grass vegetation.
The people of the area are pastoral and agro‐pastoral. Large scale irrigated agriculture is gaining
importance in some areas of the ecosystem. This ecosystem is the extreme lowland region of the
country. The flora has developed an advanced xeromorphic adaptation. Shrubs and trees have
developed dwarf growth and have small, sclerenchymatic or pubescent leaves.

Deliverable No. 1.3
46
Wetland Ecosystem
Ethiopia possesses a great diversity of wetland ecosystem (swamps, marshes, flood plains, natural or
artificial ponds, high mountains lake and micro‐dams) as a result of formation of diverse landscape
subjected to various tectonic movements, a continuous process of erosion, and human activities. The
different geological formation and climatic conditions have endowed Ethiopia with a vast water
resources and wetland ecosystem including 12 river basins, 8 major lakes and many swamps,
floodplains, and manmade reservoirs with a total annual surface runoff about 110 billion cubic meter
Aquatic Ecosystem
Aquatic in literal meaning refers to water. As an ecosystem, widely taken, it includes freshwater (rivers,
reservoirs and lakes), marine (oceans and seas) and estuarine (coastal, bays, tidal) ecosystems. The
Ethiopian aquatic ecosystem has high diversity areas such as major rivers and lakes that are of great
national and international importance. The country is well known for its richness in water potential.
There are about 30 major lakes that are located in different ecological zones. These lakes are situated
at altitudes ranging from about 150 m below sea level high up to 4000 m. The surface area of the lakes
vary considerably from less than 1 km² to over 3600 km² and mean depths range from few meters to
over 260 meters. However, the major lakes that are of economic importance are concentrated in the
Rift Valley.

Deliverable No. 1.3
47
6 African subregions by Barber‐James & Gattolliat
Barber‐James and Gattolliat (2012) divided the continent of Africa into five subregions foran
ephemeropterological investigation. These subregions were adaptedfrom the freshwater bioregions
of Thieme et al. (2005) bycombining some of their smaller bioregions to producesubregions of more
or less similar area for comparison.
North Africa, although part of the Palaearctic realm due to its stronger affinities withEuropean fauna
was compared withthe other subregions to provide a complete assessment ofrecorded mayfly
diversity in Africa. The Sahara Desert was included with North Africa in this analysis. According to
Barber‐James and Gattolliat (2012) shares Ethiopia two bioregions: mainly “Central Eastern Africa” und
to a lesser extent in the northwest the “North African & Sahara Desert”.
Ethiopia shares two subregions, the “North Africa & Sahara Desert” and the “Eastern Central Africa”.
Figure 30: Sub‐regions of Ethiopia in Africa

Deliverable No. 1.3
48
7 Proposed river typology based on ecoregions and eco‐geographic features of Ethiopia
There is a long history of organizing biological and ecological systems so that understanding can be
advanced, and management principles and practice can be developed. Because of complex
characteristics within river types which are related to local climate, geology, disturbance,
demographic, economic and political regimes, the regional approach is the realistic one by which
assessment potential can be translated into management prescriptions and application. The regional
classification of streams is an interdisciplinary and applied approach, which bases on a mix of
hydrobiological, geological, geographical and hydrological sciences.
Stream types serve as “units“, for which an assessment system can be applied. A stream type should
always be defined on the basis of natural or near‐natural reference sites, since the comparison with
undisturbed sites of a certain stream type allows defining and classifying different states of
degradation. Biological assessment requires sufficiently stable, integrated stream typologies, which
consider both abiotic and biotic criteria. The most prominent abiotic factors are stream morphology,
geochemistry, altitude, stream size and hydrology.
The approaches to stream typology using single abiotic parameters or both abiotic and biological
elements are nonexistent in Ethiopia. In such condition a “top‐down” approach is chosen on the basis
of existing knowledge and classification schemes such as altitude, freshwater ecoregion, terrestrial
ecoregion, and ecosystem types. Later on a “bottom‐up” approach will be applied for validation of
grouping the streams/rivers. Research on the importance of scale and ecological relevance of
parameters shows that even small‐scale landscape units may only partially explain the distribution of
species and communities. Thus, for practical reasons one should begin with a “top‐down” typological
frame‐work, which then must be verified through “bottom‐up” directed ecological analysisin order to
establish a sound typology.
For a country without any stream/river typology, it is useful to apply top‐down approach for
preliminary classification. This system identifies the stream types based on general landscape
conditions and serves as a first basis for comparisons by providing a common starting point. To begin
with top‐down approach existing broad classifications such as ecoregion, ecosystem, altitude class,
geology, soil type, land use and catchment size can be considered in Ethiopian conditions as discussed
above.
In regions, where stream/rivers are better known additional criteria could be applied for the further
definition and description of the types (e.g. discharge, flow type, channel morphology, water
chemistry, macroinvertebrate community, etc.).
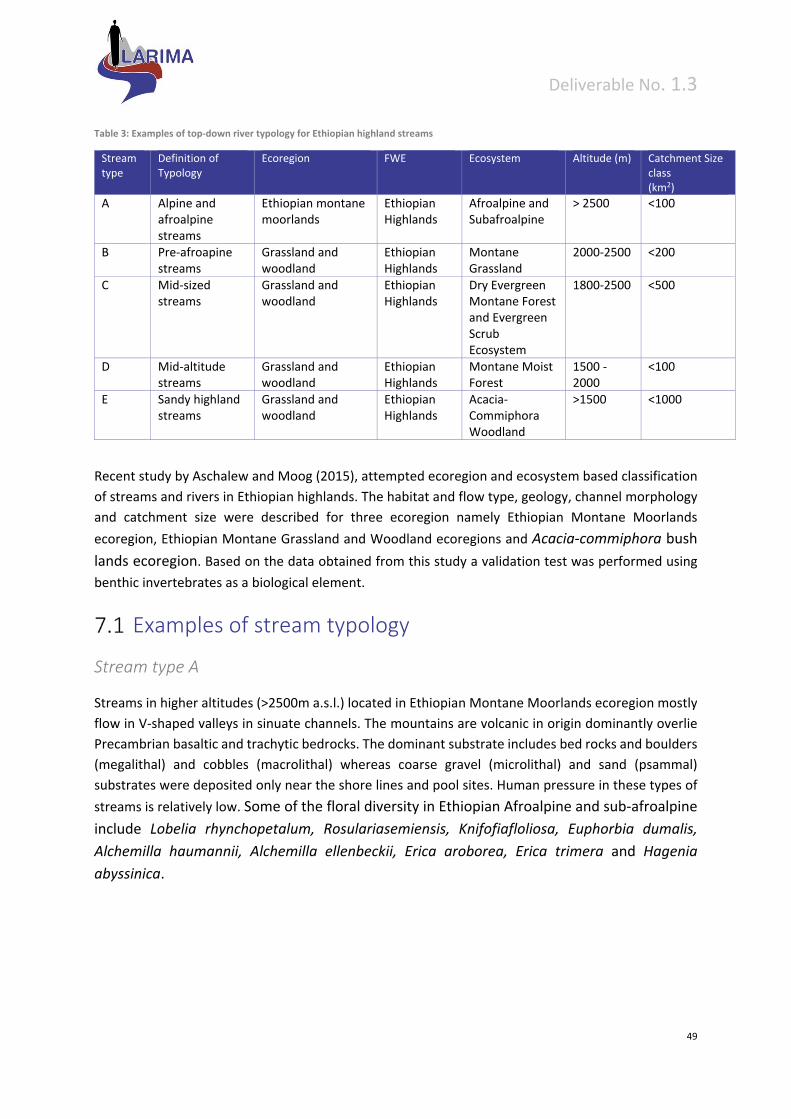
Deliverable No. 1.3
49
Table 3: Examples of top‐down river typology for Ethiopian highland streams
Stream type
Definition of Typology
Ecoregion FWE Ecosystem Altitude (m) Catchment Size class (km2)
A Alpine and afroalpine streams
Ethiopian montane moorlands
Ethiopian Highlands
Afroalpine and Subafroalpine
> 2500 <100
B Pre‐afroapine streams
Grassland and woodland
Ethiopian Highlands
Montane Grassland
2000‐2500 <200
C Mid‐sized streams
Grassland and woodland
Ethiopian Highlands
Dry Evergreen Montane Forest and Evergreen Scrub Ecosystem
1800‐2500 <500
D Mid‐altitude streams
Grassland and woodland
Ethiopian Highlands
Montane Moist Forest
1500 ‐ 2000
<100
E Sandy highland streams
Grassland and woodland
Ethiopian Highlands
Acacia‐Commiphora Woodland
>1500 <1000
Recent study by Aschalew and Moog (2015), attempted ecoregion and ecosystem based classification
of streams and rivers in Ethiopian highlands. The habitat and flow type, geology, channel morphology
and catchment size were described for three ecoregion namely Ethiopian Montane Moorlands
ecoregion, Ethiopian Montane Grassland and Woodland ecoregions and Acacia‐commiphora bush
lands ecoregion. Based on the data obtained from this study a validation test was performed using
benthic invertebrates as a biological element.
Examples of stream typology
Stream type A
Streams in higher altitudes (>2500m a.s.l.) located in Ethiopian Montane Moorlands ecoregion mostly
flow in V‐shaped valleys in sinuate channels. The mountains are volcanic in origin dominantly overlie
Precambrian basaltic and trachytic bedrocks. The dominant substrate includes bed rocks and boulders
(megalithal) and cobbles (macrolithal) whereas coarse gravel (microlithal) and sand (psammal)
substrates were deposited only near the shore lines and pool sites. Human pressure in these types of
streams is relatively low. Some of the floral diversity in Ethiopian Afroalpine and sub‐afroalpine
include Lobelia rhynchopetalum, Rosulariasemiensis, Knifofiafloliosa, Euphorbia dumalis,
Alchemilla haumannii, Alchemilla ellenbeckii, Erica aroborea, Erica trimera and Hagenia
abyssinica.

Deliverable No. 1.3
50
Figure 31: Example showing stream type A in Ethiopian Montane Moorland ecoregion (Bale highlands).
Stream type B
These streams are located in Ethiopian Montane Grassland and Woodland ecoregions, where human
activities are progressively increasing. Geologically, the area consists of a Precambrian basement
covered in most places by thick tertiary basaltic lava flows. The substrate is dominated by megalithal,
macrolithal and mesolithal. Deforestation, agricultural inputs from crop farming and cattle
grazingare stressors that affect the stream system. The width ranges from 2.5 m to 18 m.
Figure 32: Example showing stream type B in Ethiopian Woodland and Grassland ecoregions.

Deliverable No. 1.3
51
Stream type C
These streams are located in Ethiopian Montane Grassland and Woodland ecoregions characterized
by high and intense human activities. Geologically, the area consists of a Precambrian basement
covered in most places by thick tertiary basaltic lava flows. The substrate is dominated by macrolithal
in fast flowing and sloppy section, medium sized stones (mesolithal) and microlithal in slow flowing
water section. Sand and fine silt substrates are dominant on sites exposed to siltation, erosion and
heavy waste disposal.
Figure 33: Example showing stream type C in Ethiopian Woodland and Grassland ecoregions originated from same ecoregion.
Stream type D
These river types are located in Ethiopian woodland and grassland ecoregions but due to its dense
natural forest it is classified as Montane Moist Forest ecosystem. The substrate is dominated by
macrolithal covered with woody debris and mosses. Medium sized and small stones (mesolithal and
microlithal) covered with course particulate organic matter. Sand and fine silt substrates are localized
in adjust sites in between megalithal and macrolithal.
Figure 34: Example showing stream type D in Montane Moist Forest ecosystem

Deliverable No. 1.3
52
Stream type E
These streamtypes are located between 1500 m to 1800 m a.s.l. flow in sinuate channel type and are
located in Acacia‐commiphora bush lands ecoregion and border with Ethiopian Grassland and
Woodland ecoregion. The substrate is characterized by thick sand and silt layer.
Figure 35: Example showing stream type in Acacia‐commiphora bush lands ecoregion

Deliverable No. 1.3
53
8 Preliminary bottom‐up validation of proposed typology
Validation is best accomplished with reference sites that reflect the most natural and representative
condition of the region. Least impacted sites (high ‐good river class according to Aschalew and Moog,
2015) are used as the basis for stream classification. Validation is used to determine whether the
sampled sites should be placed into specific groups that will minimize variance within groups and
maximize variance among groups. Benthic macro invertebrates from reference and good sites
(according to Aschalew and Moog, 2015) were used for bottom up validation. Classification based on
ecoregion, ecosystem, altitude class, catchment class were used as over lay on NMS scatter plot
analysis on benthic invertebrate diversity data. Ecoregion and ecosystem basedstream classification
are identified as good descriptors and from geophysical features, altitude is a major factor for
scattering BMI. Other geophysical features such as soil and temperature contributed limited
information which could be because of limited data used for analysis. Therefore, large biological data
set will be required to test the effectiveness of geophysical features (especially geology) and existing
classification schemes. Moreover temporal and spatial data are recommended in the future research
to establish sound river typology that will be used for effective bioassessment in streams and rivers of
Ethiopia.
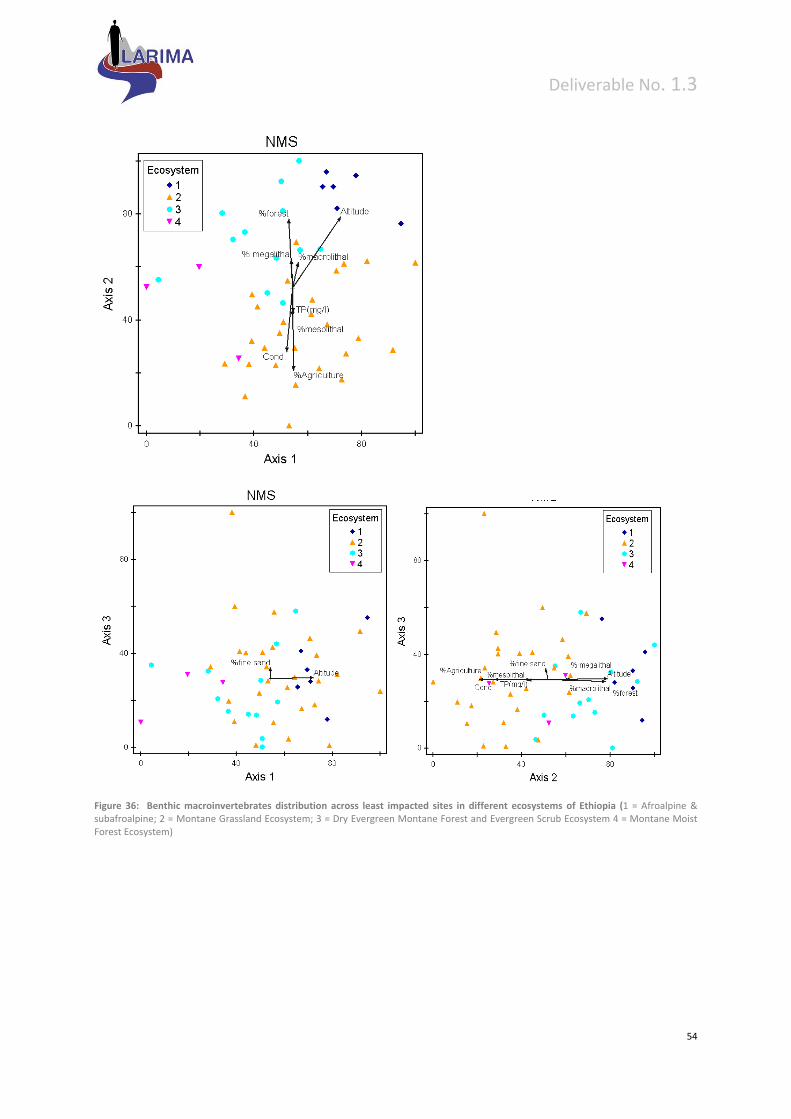
Deliverable No. 1.3
54
Figure 36: Benthic macroinvertebrates distribution across least impacted sites in different ecosystems of Ethiopia (1 = Afroalpine & subafroalpine; 2 = Montane Grassland Ecosystem; 3 = Dry Evergreen Montane Forest and Evergreen Scrub Ecosystem 4 = Montane Moist Forest Ecosystem)
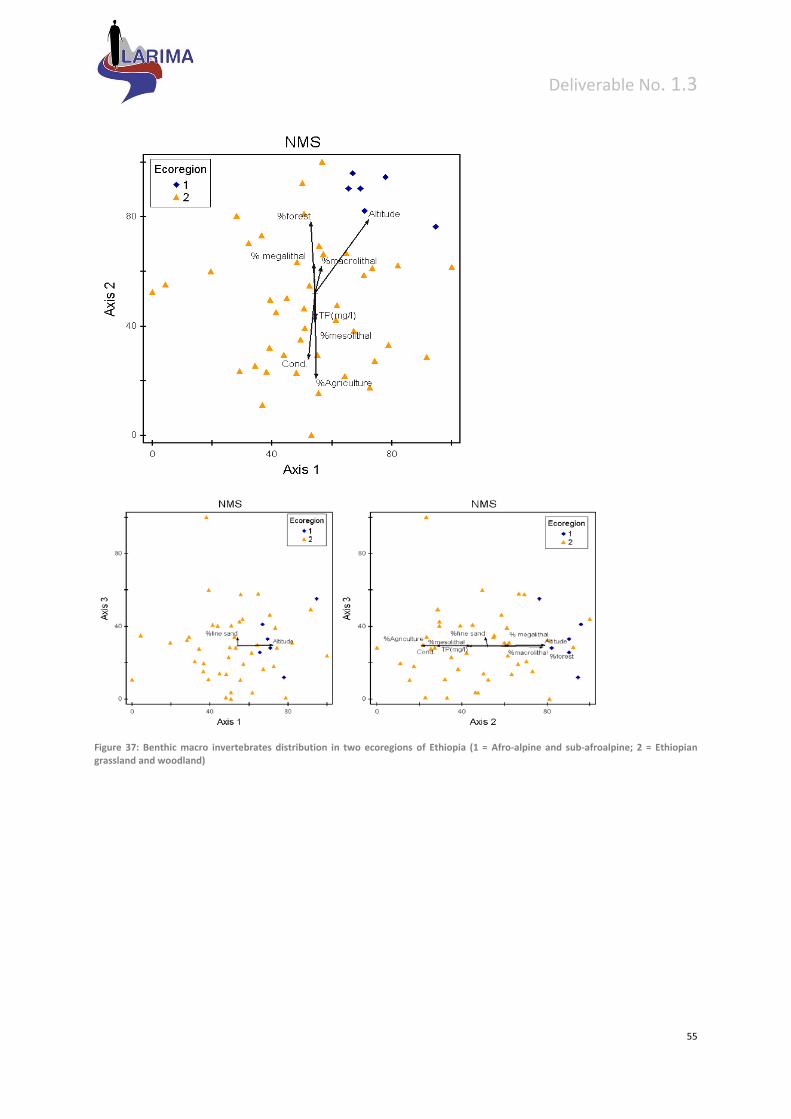
Deliverable No. 1.3
55
Figure 37: Benthic macro invertebrates distribution in two ecoregions of Ethiopia (1 = Afro‐alpine and sub‐afroalpine; 2 = Ethiopian grassland and woodland)
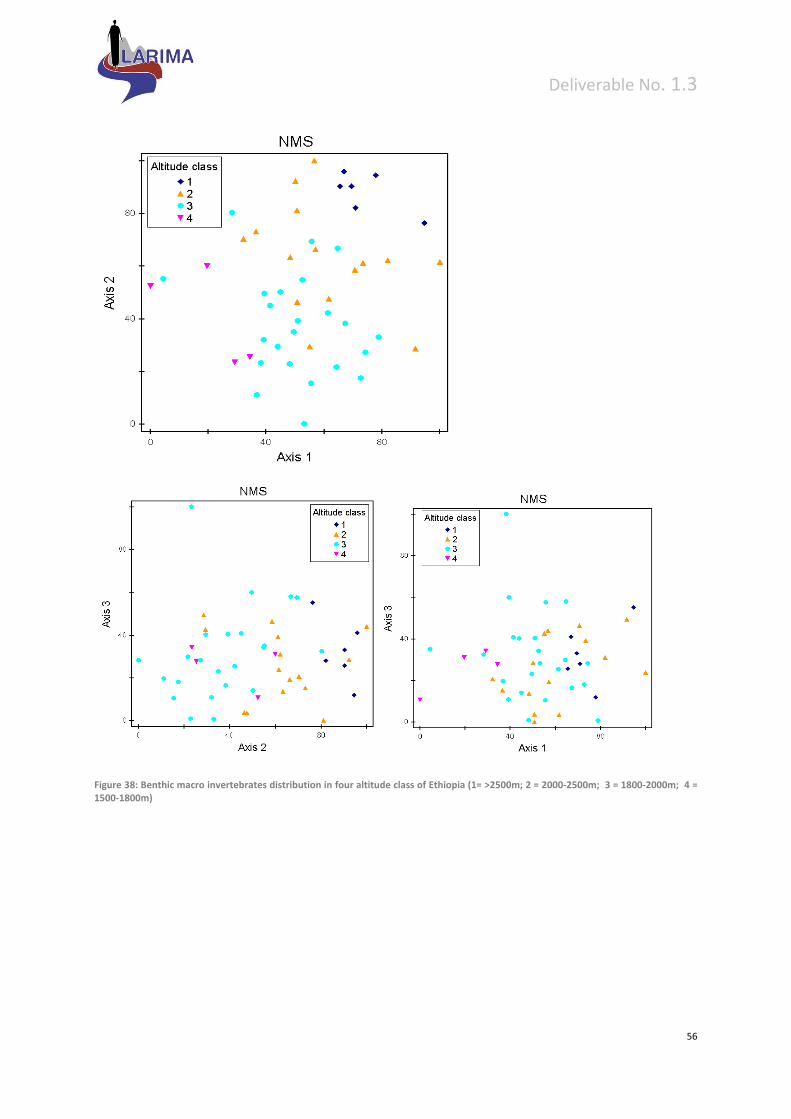
Deliverable No. 1.3
56
Figure 38: Benthic macro invertebrates distribution in four altitude class of Ethiopia (1= >2500m; 2 = 2000‐2500m; 3 = 1800‐2000m; 4 = 1500‐1800m)

Deliverable No. 1.3
57
9 References
Abell, R., M. Thieme, C. Revenga, M. Bryer, M. Kottelat, N. Bogutskaya, B. Coad, N. Mandrak, S.
Contreras‐Balderas, W. Bussing, M. L. J. Stiassny, P. Skelton, G. R. Allen, P. Unmack, A. Naseka,
R. Ng, N. Sindorf, J. Robertson, E. Armijo, J. Higgins, T. J. Heibel, E. Wikramanayake, D. Olson, H.
L. Lopez, R. E. d. Reis, J. G. Lundberg, M. H. Sabaj Perez, &P. Petry, 2008. Freshwater
ecoregions of the world: A new map of biogeographic units for freshwater biodiversity
conservation. BioScience 58:403‐4.
Aschalew, L. & O.Moog, 2015. A multimetric index based on benthic macroinvertebrates for
assessing the ecological status of streams and rivers in central and southeast highlands of
Ethiopia. Hydrobiologia, 751:229–242.
Balarin, J. D. 1986. National reviews for aquaculture development in Africa, Ethiopia. FAO Fisheries
circular.
Barber‐James, H.M. & J.L. Gattolliat, 2012. How well are Afrotropical mayflies known? Status of
current knowledge, practical applications, and future directions. Inland Waters 2: 1‐9.
Beadle, L. C., 1981. The inland waters of tropical Africa. England: Longman Group Limited.
Bennun, L. & P. Njoroge, 1999. Important bird areas in Kenya. Nature Kenya, the East Africa Natural
History Society, Nairobi, Kenya.
Briggs, J. C.,1987. Biogeography and plate tectonics. Amsterdam, The Netherlands: Elsevier.
Burgis, M. J. & J. J. Symoens, 1987. African wetlands and shallow water bodies. Paris, France:
ORSTOM.
Butzer, K. W., 1971. Recent history of an Ethiopian delta: the Omo River and the level of Lake Rudolf,
Department of Geography, Collins, R., 2002. The Nile. Yale University Press, New Haven, CT,
USA.
Coulter, G. W., B. R. Allanson, M. N. Bruton, P. H. Greenwood, R. C. Hart, P. B. N. Jackson, &A. J.
Ribbink, 1986. Unique qualities and special problems of the African Great Lakes. Environmental
Biology of Fishes 17(3):161–183.
De Graaf, M., E. Dejen & F. A. Sibbing, 2000. Barbus tanapelagius, a new species from Lake Tana
(Ethiopia): its morphology and ecology. Environmental Biology of Fishes 59(1) 1‐9.
Dgebuadze, Y. Y., A. S. Golubstov & V. N. Mikheev, 1994. Four fish species new to the Omo‐Turkana
basin, with comments on the distribution of Nemacheilus abyssinicus in Ethiopia.
Hydrobiologia 286 125‐128.
Dumont, H. J. 1986. The Nile River system. In B. R. Davies and K. F. Walker (Ed.). The ecology of river
systems. (pp. 61‐74) Dordrecht, The Netherlands: Dr W. Junk Publishers.

Deliverable No. 1.3
58
Ethiopian Agricultural Research Organization (EARO), 1998. Annual Report of Soil Department. Addis
Ababa, Ethiopia.
European Commission, 2000.Directive 2000/60/EC of the European parliament of the council of 23
October 2000 establishing a framework for Community action in the field of water policy.
Official Journal of the European Communities L327, 1–72.
FAO, 1997. AQUASAT: Morocco (2001).
ww.fao.org/ag/agl/aglw/aquastat/countries/morocco/index.stm).
FAO, 1999. State of the world fisheries and aquaculture 1998. FAO, Rome, Italy.
Friis, I., 1992. Forests and Forest Trees of Northeast Tropical Africa. HMSO, Kew Bulletin Additional
Series XV.
Getahun, A. &M. L. J. Stiassny, 1998. The freshwater biodiversity crisis: The case for conservation.
Ethiopian Journal of Science 21(2) 207‐230.
Getahun, A. 2000. Systematic studies of the African species of the genus Garra (Pisces: Cyprinidae).
Unpublished PhD thesis. New York, USA. The City University of New York.
Getahun, A.,1998. The Red Sea as an extension of the Indian Ocean. In K. Sherman, E. N. Okemwa
and M. J. Ntiba (Ed.). Large marine ecosystems of the Indian Ocean: Assessment, sustainability,
and management. (pp. 277‐281) Cambridge, MA, USA: Blackwell Science.
Golubtsov, A.S., Darkov, A.A., Dgebuadze, Yu.Vu. & M.V. Mina, 1995. An artificial key to fish species
of the Gambela Region (the White Nile Basin in the limits of Ethiopia). Joint Ethio‐Russian
Biological Expedition, Addis Abeba. 84p.
Golubtsov, A.S., Dgebuadze, Yu.Yu. & M.V. Mina, 2002. Fishes of the Ethiopian Rift Valley, pp. 167‐
258. In: C. Tudorancea and W.D. Taylor (eds.) Ethiopian Rift Valley Lakes. Backhuys Publishers,
Leiden, Holland.
Harrison, A. D. & H. B. N. Hynes, 1988. Benthic fauna of Ethiopian mountain streams and rivers.
Archiv für Hydrobiologie/ Supplement, 81 1‐36.
Harrison, A. D., 1995. Northeastern Africa rivers and streams. In C. E. Cushing, K. W. Cummings and
G. W. Minshall (Ed.). River and stream ecosystems. (pp. 507‐517) Netherlands: Elsevier Science
B. V.
Hawkes, H. A., 1975. Origin and development of the Biological Monitoring Working Party
system.Water Research.vol. 32, no. 3, p. 964‐968.
Hedberg, I. & O. Hedberg, 1979. Tropical‐alpine life forms of vascular plants. Oikos 33: 297‐307.
Hering, D., O. Moog, L. Sandin & P. F. M. Verdonschot, 2004. Overview and application of the AQEM
assessment system. Hydrobiologia 516, 1–20.
Hopson, A. J.,1982. Lake Turkana. A report on the findings of the Lake Turkana project 1972‐1975,
Vols. 1‐6. London, UK: Overseas Development Administration.

Deliverable No. 1.3
59
Hughes, R. H. & J. S. Hughes, 1992. A directory of African wetlands. Gland, Switzerland, Nairobi,
Kenya, and Cambridge, UK: IUCN, UNEP, and WCMC.
Kebede, E., G.Teferra&W. D. Taylor, 1992. Eutrophication of Lake Hayq in the Ethiopian highlands.
Journal of Plankton Research 14(10) 1473‐1482.
Kleynhans, C.J. & L., Hill, 1999. Preliminary ecoregional classification system for South Africa.
Department of Water Affairs and Forestry. Resource Directed Measures for Protection of
Water Resources. Main author: H. MacKay. Appendix 1. Volume 3: River Ecosystems Version
1.0, Pretoria.
Lévêque C, ed., 1997. Biodiversity Dynamics and Conservation: The Freshwater Fish of Tropical Africa.
Cambridge (UK): Cambridge University Press.
Manconi, R., T. Cubeddu & R. Pronzato, 1999. African freshwater sponges: Makedia tanensis gen. et
sp. Nov. From Lake Tana, Ethiopia. Memoirs of the Queensland Museum, 44: 361‐367.
McClanahan, T. R. &T. P. Young, 1996. East African ecosystems and their conservation. New York,
USA: Oxford University.
Mina, M. V., Mironovsky, A. N. and Y. Y. Dgebuadze, 1996. Lake Tana large barbs: Phenetics, growth
and diversification. Journal of Fish Biology 48(3) 383‐404.
Mohr, P. A., 1961. The geology, structure, and origin of the Bishoftu explosion craters, Shoa, Ethiopia.
Bulletin of the Geophysical Observatory, University College, Addis Ababa 2 65‐101.
Moog O., A. Schmidt‐Kloiber, T. Ofenböck & J. Gerritsen 2004. Does the ecoregion approach support
the typological demands of the EU `Water Framework Directive'? Hydrobiologia, 516, 21‐33.
Nagelkerke, L. A. J. &F. A. Sibbing, 1998. A revision of the large barbs (Barbus spp., Cyprinidae,
Teleostei) of Lake Tana (Ethiopia), with a description of a new species, Barbus osseensis. In
Nagelkerke, L. A. J. (Ed.). The barbs of Lake Tana, Ethiopia: morphological diversity and its
implications for taxonomy, trophic resource partitioning, and fisheries. (pp. 179‐214)
Wageningen, The Netherlands: Wageningen Agricultural University.
Nagelkerke, L. A. J. & F. A. Sibbing, 1997. A revision of the large barbs (Barbus spp., Cyprinidae,
Teleostei) of Lake Tana (Ethiopia), with a description of a new species, Barbus osseensis. In
Nagelkerke, L. A. J. (Ed.). The barbs of Lake Tana, Ethiopia: morphological diversity and its
implications for taxonomy, trophic resource partitioning, and fisheries. (pp. 179‐214)
Wageningen, The Netherlands: Wageningen Agricultural University.
Omer‐Cooper, J. 1930. Dr. Hugh Scott’s expedition to Abyssinia‐ A preliminary investigation of the
freshwater fauna of Abyssinia. Proc. Zool. Soc. Lond.Part I: 195‐207.
Omernik, J. M., 1987. Ecoregions of the conterminous United States. Ann. Assoc. Am. Geographers
77(1): 118–125.
Omernik, J. M., 1995. Ecoregions: A framework for managing ecosystems. The George Wright Forum
12(1): 35–51.

Deliverable No. 1.3
60
Roberts T.R., 1975. Geographical distribution of African freshwater fishes. Zoological Journal of the
Linnean Society 57: 249–319.
Rzóska, J. 1974. The upper Nile swamps, a tropical wetland study. Freshwater Biology 4 1‐30.
Seyoum, S. &I. Kornfield, 1992. Identification of the subspecies of Oreochromis niloticus (Pisces:
Cichlidae) using restriction endonuclease analysis of mitochondrial DNA. Aquaculture 102(1‐2)
29‐42.
Skelton P.H., J.A. Cambray, A. Lombard &G.A. Benn, 1995. Patterns of distribution and conservation
status of freshwater fishes in South Africa. South African Journal of Zoology 30: 71–81.
Stattersfield, A.J., M.J. Crosby, A.J. Long & D.C. Wege, 1998. Endemic Bird Areas of the World.
Priorities for Biodiversity Conservation. BirdLife Conservation Series No. 7. BirdLife
International, Cambridge, UK. 846 pp.
Thackway, R. & I. D. Cresswell (eds), 1995. An Interim Biogeographic Regionalisation for Australia: A
Framework for Establishing the National System of Reserves, Version 4.0.Australian Nature
Conservation Agency, Canberra.
Thieme M.L, R. Abell, M.L.J. Stiassny & P. Skelton, 2005. FreshwaterEcoregions of Africa and
Madagascar – A Conservation Assessment.Washington (DC): Island Press. 431p.
Tudorancea, C., Fernando, C. H. & Paggi, J. C., 1988. Food and feeding ecology of Oreochromis
niloticus (Linnaeus, 1758) juveniles in Lake Awassa (Ethiopia). Archiv für Hydrobiologie/
Supplement 79 267‐289.
Tudorancea, C., Z. GebreMariam, &E. Dadebo, 1999.Limnology in Ethiopia. In R. G. Wetzel and B.
Gopal (Ed.). Limnology in developing countries, 2. (pp. 63‐118) New Delhi, India: International
Association for Limnology.
Udvardy, M.D.F., 1975. A classification of the biogeographical provinces of the world. IUCN
Occasional Paper No. 18. International Union of Conservation of Nature and Natural
Resources, Morges, Switzerland.
Vuilleumier, F. &M. Monasterio,1986. High Altitude Tropical Biogeography. New York: Oxford
University.
Waterbury, J., 2002. The Nile Basin: National determinants of collective action. New Haven, CT, USA:
Yale University.
Westphal, E., 1975. Agriculture systems in Ethiopia. Joint publication of the College of Agriculture,
Haile Selassie I University, Ethiopia and the Agricultural University, Wageningen, The
Netherlands.
White, F., 1983. The vegetation of Africa, a descriptive memoir to accompany the
UNESCO/AETFAT/UNSO Vegetation Map of Africa UNESCO, Paris.

Deliverable No. 1.3
61
Woldegabriel, G., Aronson, J. L. &R. C. Walter, 1990. Geology, geochronology, and rift basin
development in the central sector of the main Ethiopian rift. Geological Society of America
Bulletin 102 439‐458.
Wood, R. B. & Talling, J. F., 1988. Chemical and algal relationships in a salinity series of Ethiopian
inland waters. Hydrobiologia 158 29‐67.
WWF, 1998. A conservation assessment of terrestrial ecoregions of Africa: World Wildlife Fund,
Washington, DC, USA.
Yalden, D.W., Largen, M.J., Kock, D. & J.C. Hillman, 1996. Catalogue of the mammals of Ethiopia and
Eritrea. 7. Revised checklist, zoogeography and conservation. Tropical Zoology 9: 3‐164.




















