
Mutation at the phenylalanine hydroxylase gene (PAH) and its ...
Upload
independentCategory
view
0download
0
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 290, No. 1, October, pp. 160-166, 1991
Steroid Hormone Hydroxylase Specificities of Eleven cDNA-Expressed Human Cytochrome P45Os’
David J. Waxman,*,’ David P. Lapenson,* Toshifumi Aoyama,? Harry V. Gelboin,? Frank J. Gonzalez,? and Ken Korzekwat *Department of Biological Chemistry and Molecular Pharmacology and Dana-Farber Cancer Institute, Harvard Medical School, Boston, Massachusetts 02115; and TLaboratory of Molecular Carcinogenesis, National Cancer Institute, Bethesda, Maryland 20892
Received March 29, 1991, and in revised form May 2, 1991
Steroid hydroxylation specificities were determined for 11 forms of human cytochrome P450, representing four gene families and eight subfamilies, that were synthe- sized in human hepatoma Hep G2 cells by means of cDNA- directed expression using vaccinia virus. Microsomes isolated from the P450-expressing Hep G2 cells were isolated and then assayed for their regioselectivity of hydroxylation toward testosterone, androstenedione, and progesterone. Four of the eleven P45Os exhibited high steroid hydroxylase activity (150-900 pmol hydroxy- steroid/min/mg Hep G2 microsomal protein), one was moderately active (30-50 pmol/min/mg) and six were inactive. In contrast, 10 of the P45Os effectively cata- lyzed 0-deethylation of 7-ethoxycoumarin, a model drug substrate, while only one (P450 2A6) catalyzed signifi- cant coumarin 7-hydroxylation. Human P450 4B1, which is expressed in lung but not liver, catalyzed the 6@-hydroxylation of all three steroids at similar rates and with only minor formation of other hydroxylated products. Three members of human P450 family 3A, which are expressed in liver and other tissues, also cat- alyzed steroid G&hydroxylation as their major activity but, additionally, formed several minor products that in- clude 2&hydroxy and 15j3-hydroxy derivatives in the case of testosterone. These patterns are similar to those exhibited by rat family 3A P45Os. Although several ro- dent P45Os belonging to subfamilies 2A, 2B, 2C, 2D are active steroid hydroxylases, four of five human P45Os belonging to these subfamilies exhibited very low activity or were inactive, as were the human 1A and 2E P45Os examined in the present study. These studies demonstrate that individual human cytochrome P450 enzymes can
i Supported in part by Grant DK33765 the National Institutes of Health (D.J.W.).
‘To whom correspondence should be addressed at: Dana-Farber Cancer Institute, JF-525,44 Binney Street, Boston, MA 02115.
160
hydroxylate endogenous steroid hormones with a high degree of stereospecificity and regioselectivity, and that some, but not all of the human cytochromes exhibit me- tabolite profiles similar to their rodent counterparts. 0 1991 Academic Press, Inc.
Eukaryotic cytochrome P450 (P450)3 enzymes catalyze the oxidative metabolism of structually diverse lipophilic chemicals, including many drugs, other foreign com- pounds, and endogenous substances such as steroid hor- mones and fatty acids. P450-catalyzed steroid hydroxyl- ation reactions are key to several important metabolic pathways, including vitamin D3 activation, catabolism of cholesterol to bile acids, and the biosynthesis of all major classes of steroid hormones (1). In rat liver, steroid hor- mones are subject to regiospecific hydroxylation reactions catalyzed by many, but not all, of the more than 20 distinct P450 enzymes that can be expressed in this tissue (2). Hepatic P45Os purified from rats and mice have been shown to exhibit unique patterns of steroid hormone hy- droxylation (e.g., Refs. (3-5)). These patterns can serve as useful fingerprints that are diagnostic of the identity and purity of isolated P450 preparations. Hydroxysteroid metabolite patterns can also be useful in monitoring the relative concentrations of individual P450 forms present in liver and other tissues that simultaneously express multiple cytochromes P450 (6). Finally, steroid hydrox- ylation reactions can be used to probe the active sites of these cytochromes, with dramatic changes in regiospe- cificity and/or hydroxylase activity made possible by as few as one to three amino acid changes (7-9).
In contrast to rodent P45Os, human cytochrome P450 enzymes (10) have been much less extensively studied
3 Abbreviations used: P450, cytochrome P450; TLC, thin-layer chro- matography.
0003-9861/91 $3.00
Copyright 0 1991 hy Academic Press, Inc. All rights of reproduction in any form reserved.

STEROID HYDROXYLATION BY HUMAN CYTOCHROME P45Os 161
with respect to their specificities for hydroxylation of en- dogenous steroid hormones. Indirect approaches, includ- ing antibody and chemical inhibition experiments and correlative analyses, have been used to show that P450 enzymes belonging to the CYP3A gene family4 are major contributors to steroid 6@-hydroxylation reactions cata- lyzed by human liver microsomes (12,13). This conclusion is supported by direct analysis using purified human P450 3A proteins (see footnote 4), which can be shown to cat- alyze 6/Shydroxylation of testosterone, as well as 2-hy- droxylation of 17P-estradiol and 16&-hydroxylation of dehydroepiandrosterone and its 3-sulfate, albeit at low catalytic rates (14-17). Low hydroxylase activities have also been observed with purified rat 3A P45Os (e.g., Refs. (18, 19)), and this is apparently due to the requirement of specialized conditions for reconstitution of P45Os en- coded by this gene family (20, 21).
The catalytic specificities of P45Os that are difficult to isolate or otherwise study in the purified state can be studied using suitable cDNA expression systems. cDNA- directed P450 expression in hepatoma Hep G2 cells in- fected with recombinant vaccinia virus (22) is particularly useful for studies of human P45Os, insofar as the host is a liver-derived human cell line that has a low endogenous P450 background. These cells also have good levels of endogenous human NADPH P450 reductase, which is re- quired for all microsomal P450 hydroxylation reactions, and synthesize the vaccinia-encoded P45Os in enzymat- ically active form and at levels comparable to expression levels observed for individual P450 enzymes in human liver. Using this expression system our laboratories have recently shown that two human 3A P45Os actively catalyze steroid 6@-hydroxylation reactions at rates of up to 15 mini’ P450-’ for one of the cytochromes (23). In the present study we apply the vaccinia expression system to the characterization of the hydroxylation specificities of a panel of 11 human cytochrome P45Os toward three ste- roid hormone substrates. Included are several P45Os that are present at very low levels in human tissues and have never been purified. These P45Os are shown to hydrox- ylate testosterone, androstenedione, and progesterone to give unique patterns of hydroxylated metabolites.
MATERIALS AND METHODS
Hep G2 microsomal P45Os. Recombinant vaccinia viruses containing individual human P450 cDNAs were constructed and then used to infect human Hep G2 hepatoma cells at a multiplicity of infection of 10 as described elsewhere (24). The specific P450 cDNAs included in these experiments are listed in Table I. cDNA-expression of rat P450 2Bl was carried out using similar methods (7). Wild type vaccinia strain WR was used as a control for non-P450-expressing cells. Cells were
* See Ref. (11) for an updated listing of the systematic designations given to the human P450 genes (designated CUP) and P450 gene products included in the present study. For instance, gene CYP3A4 encodes P450 3A4, which belongs to gene family 3, subfamily A, and has been desig- nated gene product 4.
TABLE I
0-dealkylation and Hydroxylation Reactions Catalyzed by Expressed Human P450 cDNAs in Vaccinia Virus-Infected
Hep G2 Microsomes Using Model Drug Substrates”
7-benzyloxy 7-ethoxycoumarin resorufin Coumarin
0-deethylase 0-debenzylase ‘I-hydroxylase Human P450’ activity activity activity
(pmol/min/mg microsomal protein)
lA2 57 <5 <3 2A6 75 <5 418 2B6 46 9 21 2C8 18 20 <3 2c9 22 <5 <3 2D6 <3 10 <3 2El 177 <5 <3 3A3 21 18 <3 3A4 36 25 13 3A5 15 -6 13 4Bl 22 8 <3 Rat 2Bl 197 440 9
“Microsomes were isolated then assayed for 0-dealkylation of 7- ethoxycoumarin or 7-benzyloxy resorufin and for 7-hydroxylation of coumarin as described under Materials and Methods. Activities shown were corrected for background activities catalyzed by Hep G2 microsomes prepared from wild type vaccinia-infected cells, typically ~2-3 pmol/ min/mg for all three substrates.
* See Ref. (24) and references therein for further characterization of these human P450 cDNAs. P450 2A6 was previously designated 2A3 and P450 2B6 was previously designated either IIB7 or hIIB1 (11).
harvested 24 h after infection, and then washed twice with 50 mM NaP, buffer, pH 7.4,0.14 M NaCl. Cells were stored at -8O’C until disrupted by sonication followed by microsome preparation by differential cen- trifugation as specified elsewhere (25). Typical yields were 1 mg cell lysate protein/l.5 X lo7 cells.
Steroid hydroxylation catalyzed by expressed P45Os. Steroid hydrox- ylation assays (with the exception of the KJV,, studies; see below) were carried using TLC methods detailed elsewhere (6, 12). In brief, Hep G2 microsomes (60 pg protein) were incubated with 10 nmol of %-labeled testosterone, androstenedione, or progesterone (58-60 mCi/ mmol; Amersham) in 0.1 M Hepes buffer, pH 7.4, containing 1 mM
NADPH for 20 min at 37°C (0.2 ml, final volume and 50 pM, final steroid concentration). Reaction mixtures were extracted with ethyl ac- etate and then chromatographed on TLC plates developed sequentially in solvent systems AB (testosterone metabolites), PQ (androstenedione metabolites), or N (progesterone metabolites). (See Table 1 of Ref. 26 for a listing of the compositions of these TLC solvent systems). TLC plates were exposed to X-ray film overnight and the individual metab- olites quantitated by liquid scintillation counting (6).
K,,,/V,, studies. For the KJV, studies, Hep G2 cells were lysed with a probe sonicator, and centrifuged at 450,OOOg for 10 min. The pellet was resuspended in 50 mM KP,, pH 7.4. Incubation mixtures (1 ml, 50 mM KP,, pH 7.4) contained -20 pmol P450. Substrate was in- troduced in 5 ~1 methanol and the reactions were initiated by the addition of 0.83 mg NADPH. The reactions were terminated after 10 min by the addition of 2.5 ml dichloromethane. For the testosterone and andro- stenedione assays, llcu-hydroxyprogesterone was then added as an in- ternal standard and 1 wmol progesterone was added to prevent nonspe- cific binding. For the progesterone studies, ll@-hydroxytestosterone was used as an internal standard. The incubations were extracted and cen-
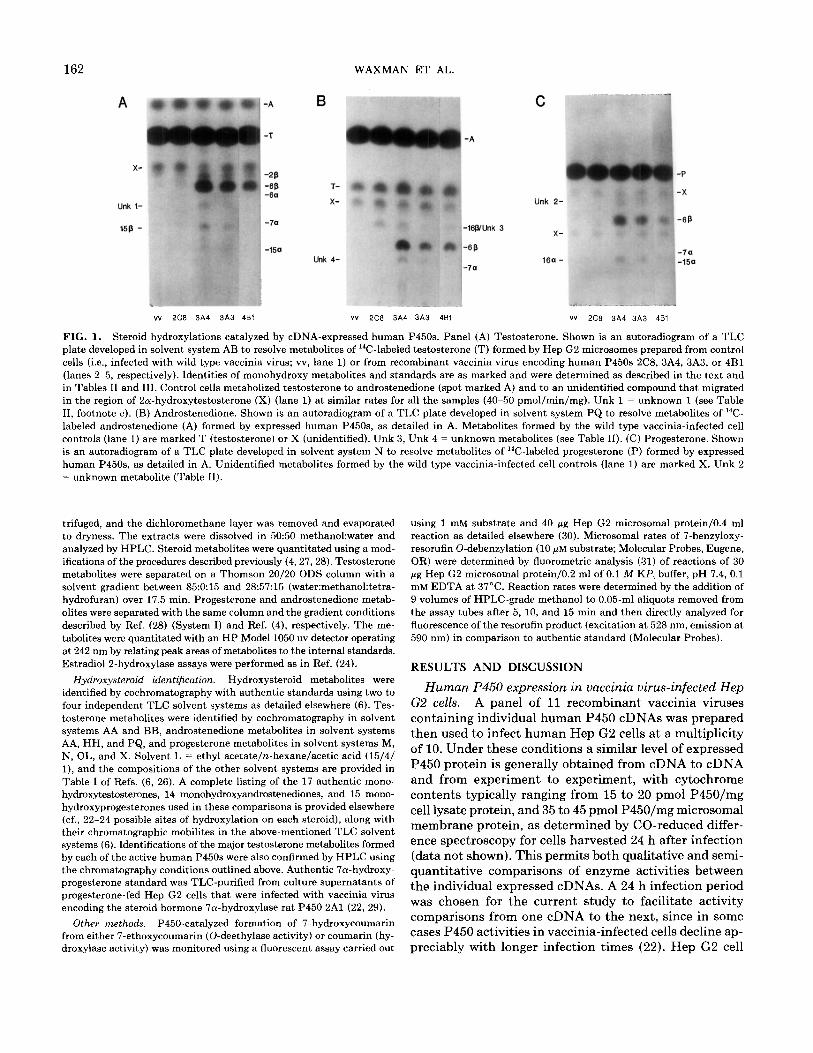
162
Unk I-
WAXMAN ET AL.
-T
-2P -6P -6Cl
T-
X-
-,a
-15a Unk 4-
-A
-16pl
-6P
-7a
‘Unk 3
Unk 2-
X-
ia -
VY 2C6 3A4 3A3 481 “V 2C6 3A4 3A3 481 Y” 2C6 3A4 3A3 461
FIG. 1. Steroid hydroxylations catalyzed by cDNA-expressed human P45Os. Panel (A) Testosterone. Shown is an autoradiogram of a TLC plate developed in solvent system AB to resolve metabolites of ‘%-labeled testosterone (T) formed by Hep G2 microsomes prepared from control cells (i.e., infected with wild type vaccinia virus; vv, lane 1) or from recombinant vaccinia virus encoding human P45Os 2C8, 3A4, 3A3, or 4Bl (lanes 2-5, respectively). Identities of monohydroxy metabolites and standards are as marked and were determined as described in the text and in Tables II and III. Control cells metabolized testosterone to androstenedione (spot marked A) and to an unidentified compound that migrated in the region of 2a-hydroxytestosterone (X) (lane 1) at similar rates for all the samples (40-50 pmol/min/mg). Unk 1 = unknown 1 (see Table II, footnote c). (B) Androstenedione. Shown is an autoradiogram of a TLC plate developed in solvent system PQ to resolve metabolites of “C- labeled androstenedione (A) formed by expressed human P45Os, as detailed in A. Metabolites formed by the wild type vaccinia-infected cell controls (lane 1) are marked T (testosterone) or X (unidentified). Unk 3, Unk 4 = unknown metabolites (see Table II). (C!) Progesterone. Shown is an autoradiogram of a TLC plate developed in solvent system N to resolve metabolites of “C-labeled progesterone (P) formed by expressed human P45Os, as detailed in A. Unidentified metabolites formed by the wild type vaccinia-infected cell controls (lane 1) are marked X. Unk 2 = unknown metabolite (Table II).
trifuged, and the dichloromethane layer was removed and evaporated to dryness. The extracts were dissolved in 5050 methanol:water and analyzed by HPLC. Steroid metabolites were quantitated using a mod- ifications of the procedures described previously (4,27,28). Testosterone metabolites were separated on a Thomson 20/20 ODS column with a solvent gradient between 85015 and 28:57:15 (water:methanol:tetra- hydrofuran) over 17.5 min. Progesterone and androstenedione metab- olites were separated with the same column and the gradient conditions described by Ref. (28) (System I) and Ref. (4), respectively. The me- tabolites were quantitated with an HP Model 1050 uv detector operating at 242 nm by relating peak areas of metabolites to the internal standards. Estradiol 2-hydroxylase assays were performed as in Ref. (24).
Hydroxysteroid identification. Hydroxysteroid metabolites were identified by cochromatography with authentic standards using two to four independent TLC solvent systems as detailed elsewhere (6). Tes- tosterone metabolites were identified by cochromatography in solvent systems AA and BB, androstenedione metabolites in solvent systems AA, HH, and PQ, and progesterone metabolites in solvent systems M, N, OL, and X. Solvent L = ethyl acetate/n-hexanelacetic acid (15/4/ l), and the compositions of the other solvent systems are provided in Table I of Refs. (6, 26). A complete listing of the 17 authentic mono- hydroxytestosterones, 14 monohydroxyandrostenediones, and 15 mono- hydroxyprogesterones used in these comparisons is provided elsewhere (cf., 22-24 possible sites of hydroxylation on each steroid), along with their chromatographic mobilites in the above-mentioned TLC solvent systems (6). Identifications of the major testosterone metabolites formed by each of the active human P45Os were also confirmed by HPLC using the chromatography conditions outlined above. Authentic 7a-hydroxy- progesterone standard was TLC-purified from culture supernatants of progesterone-fed Hep G2 cells that were infected with vaccinia virus encoding the steroid hormone 7a-hydroxylase rat P450 2Al (22, 29).
Other methods. P450catalyzed formation of 7-hydroxycoumarin from either 7-ethoxycoumarin (0-deethylase activity) or coumarin (hy- droxylase activity) was monitored using a Auorescent assay carried out
using 1 mM substrate and 40 fig Hep G2 microsomal protein/O.4 ml reaction as detailed elsewhere (30). Microsomal rates of 7-benzyloxy- resorufin 0-debenzylation (10 pM substrate; Molecular Probes, Eugene, OR) were determined by Auorometric analysis (31) of reactions of 30 wg Hep G2 microsomal protein/O.2 ml of 0.1 M KP; buffer, pH 7.4, 0.1 mM EDTA at 37°C. Reaction rates were determined by the addition of 9 volumes of HPLC-grade methanol to 0.05-ml aliquots removed from the assay tubes after 5, 10, and 15 min and then directly analyzed for fluorescence of the resorufin product (excitation at 528 nm, emission at 590 nm) in comparison to authentic standard (Molecular Probes).
RESULTS AND DISCUSSION
Human P450 expression in vaccinia virus-infected Hep G2 cells. A panel of 11 recombinant vaccinia viruses containing individual human P450 cDNAs was prepared then used to infect human Hep G2 cells at a multiplicity of 10. Under these conditions a similar level of expressed P450 protein is generally obtained from cDNA to cDNA and from experiment to experiment, with cytochrome contents typically ranging from 15 to 20 pm01 P450/mg cell lysate protein, and 35 to 45 pmol P45O/mg microsomal membrane protein, as determined by CO-reduced differ- ence spectroscopy for cells harvested 24 h after infection (data not shown). This permits both qualitative and semi- quantitative comparisons of enzyme activities between the individual expressed cDNAs. A 24 h infection period was chosen for the current study to facilitate activity comparisons from one cDNA to the next, since in some cases P450 activities in vaccinia-infected cells decline ap- preciably with longer infection times (22). Hep G2 cell

STEROID HYDROXYLATION BY HUMAN CYTOCHROME P45Os 163
TABLE II
Steroid Hydroxylations Catalyzed by Human P45Os 3A3,3A4 and 3A5”
3A3 3A4 3A5
pmol/min/mg Hep G2 cell microsomes*
Testosterone Go-hydroxy 2/%hydroxy 156-hydroxy Unknown 1’ Total activity
Progesterone Go-hydroxy 16a-hydroxy Unknown 2d Total activity
Androstenedione Go-hydroxy Unknown 3’ Unknown 4’ Total activity
330 550 115 26 40 8 14 23 <3 8 16 13
378 -
629 123
410 730 135 50 85 <3
36 70 4 496 885 139
240 535 130 19 40 14 17 40 7
276 615 151
a Metabolites were identified as described in the text. Activities were measured in Hep G2 microsomes isolated from cells infected with re- combinant vaccinia virus encoding the indicated human P450 cDNAs, as described under Materials and Methods. Values shown were de- termined in parallel incubations, and are representative of at least three separate experiments.
* Net activities, after subtraction of background activities catalyzed by microsomes isolated from wild type vaccinia-infected Hep G2 cells (typically corresponding to 62-3 pmol/min/mg). An activity of 10 pmol/ min/mg corresponds to -1200 cpm hydroxysteroid product formed during a 20 min assay with 60 pg Hep G2 microsomal protein.
’ Unknown 1 comigrated with llp-hydroxytestosterone and 19-hy- droxytestosterone in TLC solvent system AA but was distinguished from those compounds in system BB. Its migration was similar to that of l/3- hydroxytestosterone (but not 16 other standards) in both solvents (6), but the lp standard was not available for direct comparison.
’ Comigrated with 2rY-hydroxyprogesterone in TLC solvent system M, but migrated more slowly than the 2a derivative in systems OL and N. It was also distinguishable from 15 other available monohydroxy- progesterone standards (6).
’ Migration similar to [‘“Cl 16/J-hydroxyandrostenedione formed by purified rat P450 2Bl. TLC purification and rechromatography in TLC solvent systems AA and PQ evidenced heterogeneity and/or decompo- sition, as is characteristic of the acid-labile 16/3-hydroxyandrostenedione (31.
’ Migrates between 7o- and 7/3-hydroxyandrostenedione in TLC sol- vent systems AA and PQ and slightly ahead of Ga-hydroxyandroste- nedione in system HH.
microsomes were isolated for use in the enzymatic anal- yses, since this fraction contains both the expressed P45Os and sufficient endogenous NADPH P450 reductase to support P450-dependent hydroxylation reactions at maximal rates (e.g., Ref. (22)). Cytochrome b5, which plays a facilitatory role in some microsomal P450 hy- droxylation reactions, is also present in the Hep G2 mi- crosomes (data not shown). Moreover, preliminary studies
using crude lysates revealed the presence of interfering steroid-metabolizing activities in the cytosol (probably steroid oxidoreductases) when using progesterone as sub- strate.
In order to insure that the expressed P45Os are enzy- matically competent, the isolated Hep G2 microsomes were assayed for 7-ethoxycoumarin 0-deethylation activ- ity, an 0-dealkylation reaction that is known to be cat- alyzed by P45Os belonging to several different gene fam- ilies. Five of the eleven expressed human P45Os actively catalyzed this reaction (36-177 pmol product/min/mg microsomal protein), with human P450 2El exhibiting the highest activity. Five other P45Os metabolized 7- ethoxycoumarin at rates that were lower (15-22 pmol/ min/mg) but still significant compared to microsomes prepared from mock P450-infected Hep G2 cell controls (<2-3 pmol/min/mg) (Table I). These values can be compared to an activity of 197 pmol/min/mg when vac- cinia-expressed rat P450 2Bl was analyzed in parallel (Table I). One of the human P45Os was judged inactive with 7-ethoxycoumarin as substrate (P450 2D6); however, in other studies this P450 has been shown to actively catalyze debrisoquine 4-hydroxylation. In other experi- ments six of the human P45Os were shown to catalyze the 0-debenzylation of 7-benzyloxyresorufin at rates that were low but measureable (Table I). With coumarin as substrate, human P450 2A6 exhibited a high activity (418 pmol/min/mg; Table I), in agreement with earlier studies
TABLE III
Steroid hydroxylations catalyzed by human P45Os 4Bl and X8”
Testosterone Androstenedione Progesterone
pmol/min/mg
P450 4Bl Go-hydroxy Unknown
P450 2C8 Unknown
190 205 195 <lo” 21’
27d 30’ 49’
’ Activities shown are from a single representative experiment and were determined as described in Table II, footnotes a and b.
* Migration similar to 2&hydroxytestosterone, but characterization was hampered by the overlaps with metabolite formed by wild type vaccinia-infected cell microsomes that migrated just below spot X (Fig. 1A). Tentative identification as 2P-hydroxytestosterone is supported by HPLC analysis.
’ Migration similar to 160.hydroxyandrostenedione. These uniden- tified androstenedione metabolites of P45Os 4Bl and 2C8 were chro- matographically indistinguishable from the metabolite designated “un- known 3” in Table II (also see footnote e to Table II) and probably correspond to 16@-hydroxyandrostenedione.
d Comigrates with G&hydroxytestosterone in TLC solvent system BB, but migrates faster than the S/3 metabolite in system AB.
e Migrates similar to 6@-hydroxyprogesterone in TLC solvent system OL, similar to 21-hydroxyprogesterone in system X, and between 1201. and 6o-hydroxyprogesterone in system M.

164 WAXMAN ET AL.
TABLE IV
Kinetic Analysis of Testosterone and Estradiol Metabolism Catalyzed by Human P45Os”
P450 form
A. Testosterone Go-hydroxylation 3A3 3A4 3A5 4Bl
B. E&radio1 2-hydroxylation
55 14 47 20
430 19 152 24
3A3 109 1.9 3A4 156 3.3 3A5 (Nonlinear kinetics) lA2 32 3.3
“Steady-state kinetic parameters were monitored by HPLC using isolated Hep G2 microsomal fractions, as described under Methods. Shown are values derived from a single experiment representative of at least two independent determinations. Lineweaver-Burke double recip- rocal plots are shown in Fig. 2. V,, values are in units of nmol hy- droxysteroid/nmol P450/min. Data are given for the primary metabolite only. In cases where the minor metabolites could be quantitated reliably over the range of substrate concentrations employed in these experi- ments, the product ratios did not vary appreciably (110%) with substrate concentration, suggesting that the Km values are the same as for the major metabolite.
from one of our laboratories (32; cytochrome designated IIA3 in that study). Human P450 2B6 and rat P450 2Bl exhibited much lower activities (21 and 9 pmol/min/mg, respectively), while the other 10 P45Os were inactive.
Steroid hydroxylation catalyzed by cDNA-expressed hu- man P45Os. Each of the 11 expressed human P45Os was then assayed using testosterone, androstenedione and progesterone as substrates. Four of the P45Os exhibited high hydroxylase activity with all three steroids (3A3,3A4, 3A5,4Bl), one was moderately active (2C8) and six were inactive. Representative autoradiographs of TLC plates resolving the hydroxysteroid metabolites formed by 4 of the P45Os are shown in Fig. 1. These P45Os were each characterized by a unique metabolic profile, with the ex- ception of P45Os 3A3 and 3A4, which yielded indistin- guishable metabolite patterns.
Individual steroid metabolites were eluted from the TLC plates and then rechromatographed on TLC plates devel- oped in solvent systems chosen to resolve closely migrating monohydroxysteroids (6). Product identifications were based on the cochromatography of 14C-labeled steroid me- tabolites with authentic, unlabeled standards in multiple TLC solvent systems. This was accomplished using meth- ods that we have previously shown to be highly sensitive and reliable, particularly when a large collection of au- thentic standards is available (e.g., Refs. (6, 12, 26)). Tes- tosterone metabolite identifications were also confirmed by HPLC (see Methods). The product identifications de- termined in this manner are marked on Fig. 1 and are listed in Tables II and III, along with the hydroxylation
rates measured for each of the products. Several of the steroid metabolites did not corn&rate with any of the available standards, and thus could not be identified.
6P-Hydroxylation by human family 3A and 4B P45Os. Steroid 6P-hydroxylation was the most prominent reac- tion catalyzed by four of the human P45Os, three belonging to gene family CYP3A and one to gene family CYP4B. Family 3A P45Os have previously been implicated as major catalysts of steroid 6P-hydroxylation in human liver mi- crosomes by antibody inhibition and other indirect meth- ods, including correlative analyses (12, 13), as well as expression studies (23). The present study extends those previous findings to include P450 3A3, which was found to have a hydroxylation specificity indistinguishable from that of P450 3A4, and also includes the resolution and characterization of several minor metabolites that were not studied previously (Table II). P450 3A3 and P450 3A4, which have 98% amino acid sequence identity (23), exhibited indistinguishable hydroxylation profiles with all three steroids. P450 3A5, which is only 82-84% similar to the other two P45Os at the amino acid level (23), was distinguished by its higher regioselectivity for 6P-hydrox- ylation, particularly in the case of testosterone and pro- gesterone (93-97% of total product formation). P450 3A5 also exhibited a distinctly lower steroid hydroxylase ac- tivity than 3A4 (Table II), despite the fact that the two cytochromes metabolize nifedipine at similar rates (23). Kinetic analysis (Table IVA) revealed that this lower ac- tivity (determined at 50 PM steroid substrate) reflects an eight- to ninefold higher K,,, exhibited by 3A5 as compared to 3A3 and 3A4. Expressed P450 3A3 was somewhat less active than P450 3A4 under standard assay conditions, but this effect is primarily due to differences in V,,, (Table
10, 1 I
08 t 0 P450 3A3 . P450 3A4 v P450 3A5 . P450 481
1
FIG. 2. Lineweaver-Burke double-reciprocal plots analyzing testos- terone Go-hydroxylation catalyzed by cDNA-expressed human P45Os. Data shown were obtained in a single representative experiment as de- scribed under Methods. Kinetic constants are summarized in Table IV.

STEROID HYDROXYLATION BY HUMAN CYTOCHROME P45Os 165
IV and Fig. 2). In contrast, both K,,, and V,,, differences characterize the estradiol2-hydroxylase activities of 3A3 and 3A4 (Table IVB). In the case of 3A5, which also cat- alyzes estradiol 2-hydroxylation (24), nonlinear kinetics were observed (data not shown).
The pattern of testosterone metabolism catalyzed by human P45Os 3A3 and 3A4, i.e., formation of 6p, 2& and 15P-hydroxytestosterone, is highly similar to that exhib- ited by human liver microsomes, an observation that is consistent with the identification of the family 3A P45Os as major contributors to those reactions in human liver (12). Rat 3A P45Os exhibit a similar metabolite pattern (20, al), suggesting the conservation between species of important active site determinants of P450 3A substrate specificity.
study exhibited steroid metabolite profiles similar to their rodent counterparts. Thus, both human and rat P45Os belonging to family 3A hydroxylate testosterone at the 6p > 2p J=- 15p positions, all P-face positions that are not shielded by the Cl8 or Cl9 methyl groups. In contrast, human P450 2B6 was inactive with the three steroids examined in this study, even though rat 2B1, which shares 76% similarity to human 2B6 (34), actively metabolizes all three substrates (2). Human lung P450 4Bl was iden- tified as a steroid Go-hydroxylase. The activities of the corresponding rabbit and rat lung 4B P45Os (35) are not known.
G/3-Hydroxylation of all three steroids was also cata- lyzed by human P450 4Bl (Table III). This P450 exhibited a threefold higher K, for testosterone as compared to 3A3 and 3A4 (Table IVA, Fig. 2), and this is reflected in the somewhat lower testosterone G/3-hydroxylase activity of 4Bl as compared to 3A3 and 3A4 in assays carried out at 50 PM steroid substrate (Table III versus Table II). Minor metabolites formed by P450 4Bl were tentatively identified as 2&hydroxytestosterone and 16P-hydroxy- androstenedione (Table III, footnotes b and c, respec- tively). Northern blot analyses indicate that the mRNA encoding this P450 is expressed in human lung, but not liver, albeit at levels that vary greatly from one individual to the next (33).
Hydroxylation specificities of human family 2 and family 1 P45Os. Of the six human family 2 P45Os examined in the present study, only one was moderately active (2C8; 27-49 pmol/min/mg) (Table III). The others, human P45Os 2A6, 2B6, 2C9, 2D6, and 2E1, were essentially in- active, as was human P450 lA2. In contrast, several rodent P450 members of the 2A, 2B, 2C, and 2D subfamilies (each represented in our panel of 12 human P45Os) ac- tively metabolize these same steroid hormones at one or more sites (e.g., Refs (2, 5)). One P450 2C8 metabolite was detected for each of the three steroid substrates, but these metabolites could not be identified (Table III).
The physiological significance of many of these hy- droxylation reactions is unclear, but may include androgen deactivation, or alternatively, generation of hydroxyste- roids that have important endocrine functions that remain to be discovered (2). Some of these hydroxylation activ- ities, particularly those characterized by high K,,, values (c.f., Table IV), may derive from the unusually broad sub- strate specificities of these P45Os, many of which will also oxygenate a large variety of drugs and other foreign com- pounds. Androgen 7a-hydroxylation, such as catalyzed by rodent 2A P45Os, but not by the human P45Os char- acterized in this study, may lead to the formation of com- pounds that play an important role in the production and metabolism of testosterone in the testis (36, 37). The rat steroid 7a-hydroxylase P450 2Al is not only expressed in liver (a), but is expressed in testis, where it is the major catalyst of testosterone 7a-hydroxylation (38). Testos- terone 7a-hydroxylase activity is also detectable in human testes (39), but the corresponding P450 has yet to be identified.
REFERENCES
1.
2.
3.
4.
5.
CONCLUSION 6.
The studies described in this report provide a quali- tative characterization of the steroid hydroxylase speci- ficities, as well as a semi-quantitative comparison of the activities of 11 human cytochrome P45Os representing a total of four gene families and eight subfamilies. Several of these cytochromes are present at low levels in human tissues and have never been purified. The present findings complement recent studies of 17&estradiol metabolism catalyzed by these same human P45Os (24) and provide further insight into the range of steroid hydroxyiation reactions that can be catalyzed by human P450 enzymes. Some, but not all of the human P45Os examined in this
7.
8.
9.
10.
11.
Waterman, M. R., John, M. E., and Simpson, E. R. (1986) (Ortiz de Montellano, P. R., Ed.), pp. 345-386, Pergamon, New York.
Waxman, D. J. (1988) Biochem. Pharmacol. 37, 71-84.
Waxman, D. J., Ko, A., and Walsh, C. (1983) J. Biol. Chem. 258, 11,937-11,947.
Wood, A. W., Ryan, D. E., Thomas, P. E., and Levin, W. (1983) J. Bid. Chem. 258,8839-8847.
Devore, K., Harada, N., and Negishi, M. (1985) Biochemistry 24, 5632-5637.
Waxman, D. J. (1991) in Methods in Enzymology, (Waterman, M. R., and Johnson, E. F., Eds.), Vol. 206, pp. 462-476, Academic Press, San Diego.
Aoyama, T., Korzekwa, K., Nagata, K., Adesnik, M., Reiss, A., La- penson, D. P., Gillette, J., Gelboin, H. V., Waxman, D. J., and Gon- zalez, F. J. (1989) J. Biol. Chem. 264, 21,327-21,333.
Lindberg, R. L. P., and Negishi, M. (1989) Nature 339, 632-634.
Kronbach, T., Larabee, T. M., and Johnson, E. F. (1989) Proc. Natl. Acad. Sci. USA 86,8262-8265.
Guengerich, F. P. (1989) Annu. Rev. Pharmacol. Toxicol. 29, 241- 264.
Nebert, D. W., Nelson, D. R., Coon, M. J., Estabrook, R. W., Fey- ereisen, R., Fujii-Kuriyama, Y., Gonzalez, F. J., Guengerich, F. P., Gunsalus, I. C., Johnson, E. F., Loper, J. C., Sato, R., Waterman, M. R., and Waxman, D. J. (1991) DNA Cell. Biol. 10, 1-14.

166 WAXMAN ET AL.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
Waxman, D. J., Attisano, C., Guengerich, F. P., and Lapenson, D. P. (1988) Arch. Biochem. Biophys. 263, 424-436.
Wrighton, S. A., Ring, B. J., Watkins, P. B., and Vandenbranden, M. (1989) Mol. Pharmacol. 36, 97-105.
Wrighton, S. A., Brian, W. R., Sari, M. A., Iwasaki, M., Guengerich, F. P., Raucy, J. L., Molowa, D. T., and Vandenbranden, M. (1990) Mol. Pharmacol. 38,207-213.
Guengerich, F. P., Martin, M. V., Beaune, P. H., Kremers, P., Wolff, T., and Waxman, D. J. (1986) J. Biol. Chem. 261, 5051-5060.
Kawano, S., Kamataki, T., Yasumori, T., Yamazoe, Y., and Kato, R. (1987) J. Biochem. 102,493-501.
Kitada, M., Kamataki, T., Itahashi, K., Rikihisa, T., and Kanakubo, Y. (1987) J. Biol. Chem. 262, 13,534-13,537.
Elshourbagy, N. A., and Guzelian, P. S. (1980) J. Biol. Chem. 255, 1279-1285.
Waxman, D. J., Dannan, G. A., and Guengerich, F. P. (1985) Bio- chemistry 24,4409-4417.
Imaoka, S., Terano, Y., and Funae, Y. (1988) J. Biochem. (Tokyo) 104,481-487.
Halvorson, M., Greenway, D., Eberhart, D., Fitzgerald, K., and Par- kinson, A. (1990) Arch. Biochem. Biophys. 277, 166-180.
Aoyama, T., Korzekwa, K., Nagata, K., Gillette, J., Gelboin, H. V., and Gonzalez, F. J. (1989) Eur. J. Biochem. 181, 331-336.
Aoyama, T., Yamano, S., Waxman, D. J., Lapenson, D. P., Meyer, U. A., Fischer, V., Tyndale, R., Inaba, T., Kalow, W., Gelboin, H. V., and Gonzalez, F. J. (1989) J. Biol. Chem. 264, 10,388-10,395.
Aoyama, T., Korzekwa, K., Nagata, K., Gillette, J., Gelboin, H. V., and Gonzalez, F. J. (1990) Endocrinology 126, 3101-3106.
25
26
27.
28.
29. 30.
31.
32.
33.
34.
35.
36.
37. 38.
39.
Waxman, D. J., Lapenson, D. P., Morrissey, J. J., Park, S. S., Gel- boin, H. V., Doehmer, J., and Oesch, F. (1989) Biochem. J. 260, 81-85.
Waxman, D. J., Lapenson, D. P., Nagata, K., and Conlon, H. D. (1990) Biochem. J. 265, 187-194. Korzekwa, K. R., Trager, W. F., Nagata, K., Parkinson, A., and Gillette, J. R. (1990) Drug Metab. Dispos. 18, 974-979. Swinney, D. C., Ryan, D. E., Thomas, P. E., and Levin, W. (1987) Biochemistry 26, 7073-7083. Waxman, D. J. (1984) J. Biol. Chem. 259,15,481-15,490. Waxman, D. J., and Walsh, C. (1982) J. Biol. Chem. 257, 10,446- 10,457. Burke, M. D., Thompson, S., Elcombe, C. R., Halpert, J., Haapa- ranta, T., and Mayer, R. T. (1985) Biochem. Pharmacol. 34,3337- 3345. Yamano, S., Tatsuno, J., and Gonzalez, J. F. (1990) Biochemistry 29,1322-1329. Nhamburo, P. T., Gonzalez, F. J., McBride, 0. W., Gelboin, H. V., and Kimura, S. (1989) Biochemistry 28, 8060-8066. Yamano, S., Nhamburo, P. T., Aoyama, T., Meyer, U. A., Inaba, T., Kalow, W., Gelboin, H. V., McBride, 0. W., and Gonzalez, F. J. (1989) Biochemistry 28, 7340-7348. Gasser, R., and Philpot, R. M. (1989) Mokc. Pharmucol. 35, 617- 625. Inano, H., Suzuki, K., Wakabayashi, K., and Tamaoki, B.-I. (1973) Endocrinology 92, 22-30. Mittler, J. C. (1985) Steroids 45, 135-142. Sonderfan, A. J., Arlotto, M. P., and Parkinson, A. (1989) Endo- crinology 125,857-866. Sunde, A., Tveter, K., and Eik-Nes, K. B. (1980) Acta Endocrinolog. 93,243-249.
Copyright © 2022 FDOKUMEN




















