
This book has been specifically designed to accomplish two things... #1

Shifting visual attention away from fixation is specificallyassociated with alpha band activity over ipsilateralparietal regions
DIEGO COSMELLI,a,b VLADIMIR LOPEZ,a JEAN-PHILIPPE LACHAUX,c,d
JAVIER LOPEZ-CALDERON,e,f BERNARD RENAULT,g,h,i JACQUES MARTINERIE,g,h,i andFRANCISCO ABOITIZb,f
aEscuela de Psicologıa, Pontificia Universidad Catolica de Chile, Santiago, ChilebCentro Interdisciplinario de Neurociencia, Pontifica Universidad Catolica de Chile, Santiago, ChilecUnite 280, INSERM 151, Lyon, FrancedUniversite Lyon 1, Lyon, FranceeCenter for Mind and Brain, University of California, Davis, Davis, California, USAfLaboratorio de Neurociencia Cognitiva, Departamento de Psiquiatrıa, Escuela deMedicina, Pontificia Universidad Catolica de Chile, Santiago, ChilegUPMC Universite Paris 06, UMR 7225, UMR-S 975, Centre de Recherche de l’Institut Cerveau-Moelle (CRICM), Paris, FrancehCNRS, UMR 7225, CRICM, Paris, FranceiINSERM, UMR-S 975, CRICM, Paris, France
Abstract
We studied brain activity during the displacement of attention in a modified visuo-spatial orienting paradigm. Using abehaviorally relevant no-shift condition as a control, we asked whether ipsi- or contralateral parietal alpha bandactivity is specifically related to covert shifts of attention. Cue-related event-related potentials revealed an attentiondirecting anterior negativity (ADAN) contralateral to the shift of attention and P3 and contingent negative variationwaveforms that were enhanced in both shift conditions as compared to the no-shift task. When attention was shiftedaway from fixation, alpha band activity over parietal regions ipsilateral to the attended hemifieldwas enhanced relativeto the control condition, albeit with different dynamics in the upper and lower alpha subbands. Contralateral-to-attended parietal alpha band activity was indistinguishable from the no-shift task.
Descriptors: EEG, Alpha band, Parietal cortex, No-shift, Central cue
Waking life is typically characterized by ongoing shifts of atten-tion among a vast number of possible objects. Behaviorally, at-tending shortens reaction times, inhibits distracters, andfacilitates recognition, discrimination, and recall (Kastner &Ungerleider, 2000). Neurophysiological studies have shown at-tention-related modulations at different levels, including en-hanced baseline firing patterns (Nakamura & Colby, 2000),changes in temporal relations among neuronal populations(Fries, Reynolds, Rorie, & Desimone, 2001; Jensen, Kaiser, &Lachaux, 2007), and selective modulation of several componentsof the event-related potential (ERP; Hillyard & Anllo-Vento,1998; Luck, Woodman, & Vogel, 2000). Neuroimaging and neu-ropsychological approaches have consistently shown that a dis-tributed brain network is implicated in the control of attention,
including both cortical and subcortical structures (Raz, 2004;Weiskrantz, Warrington, Sanders, & Marshall, 1974). At thecortical level, fronto-parietal networks are thought to play acritical role in attentional selection, and the right hemisphere isusually considered dominant (Corbetta, Miezin, Shulman, &Petersen, 1993; Corbetta & Shulman, 2002; Green &McDonald,2008; Hopfinger, Camblin, & Parks, 2010).
Traditionally, electrophysiological studies of attention ori-enting in humans have focused on the modulation of brain ac-tivity evoked by a stimulus that has or has not been attended(Hillyard & Kutas, 1983; Hillyard, Luck, & Mangun, 1994).When one is studying covert visuo-spatial attention shifts, themost common experimental manipulation is to instruct the sub-ject to orient his or her attention to a cued locationFwithoutmoving the eyesFand then detect a relevant stimulus that mayormay not appear at the cued location (Posner, 1980). However,more recently, the question about the brain mechanisms under-lying the process of orienting or shifting attention, in contradis-tinction to its effects, has begun to be addressed inelectroencephalogram (EEG) work. To do this, studies have fo-cused on analyzing brain responses during the cue–stimulus in-terval in addition to that evoked by the subsequent stimulus.
The authors thank Steven Hillyard for insightful comments on aprevious version of this work. This work was supported by FONDECYTprojects 3060094, 1090612, and PBCT-CONICYT PSD72.
Address correspondence to: Diego Cosmelli, Escuela de Psicologıa,Pontificia Universidad Catolica de Chile, Av. Vicuna Mackenna 4860,Macul, Santiago de Chile, Chile. E-mail: [email protected]
Psychophysiology, 48 (2011), 312–322. Wiley Periodicals, Inc. Printed in the USA.Copyright r 2010 Society for Psychophysiological ResearchDOI: 10.1111/j.1469-8986.2010.01066.x
312

Among these, studies of oscillatory activity during the orien-tation of visuo-spatial attention in the cue–stimulus interval haveshown consistent changes in the alpha (8–12 Hz) band (Foxe,Simpson, & Ahlfors, 1998; Freunberger et al., 2008; Fu et al.,2001; Kelly, Lalor, Reilly, & Foxe, 2006; Rihs, Michel, & Thut,2007, 2009; Sauseng et al., 2005; Thut, Nietzel, Brandt, & Pas-cual-Leone, 2006; Worden, Foxe, Wang, & Simpson, 2000;Yamagishi, Callan, Anderson, & Kawato, 2008). Because an in-crease in alpha rhythm has been considered indicative of reducedcortical activity (Pfurtscheller, 2001; but see Nunez, Wingeier, &Silberstein, 2001; Palva & Palva, 2007), occipito-parietal con-tralateral-to-cued decrease (Sauseng et al., 2005; Thut et al.,2006; Yamagishi et al., 2008) and/or ipsilateral-to-cued increase(Doesburg, Roggeveen, Kitajo, & Ward, 2008; Fu et al., 2001;Kelly et al., 2006; Rihs et al., 2007; Worden et al., 2000) in alphapower has been interpreted as reflecting facilitation of the cued(attended), or active suppression of the noncued (ignored) loca-tion, respectively. Whether an ipsilateral increase or a contralat-eral decrease in posterior alpha band power is specific to covertvisuo-spatial shifts of attention remains an open question, al-though recent studies suggest that both can be elicited dependingon task requirements such as the cue–stimulus interval duration(Rihs et al., 2009).
Because of the way the cue–stimulus paradigm is constructed,being able to identify brain activity that is specific to the processof orienting attention and not merely related to sensory or in-terpretative aspects of the cue is necessary (Jongen, Smulders, &Van Der Heiden, 2007; Nobre, Sebestyen, &Miniussi, 2000; vanVelzen & Eimer, 2003). Different approaches have been devel-oped to address this question, such as using bilaterally symmetriccues (Nobre et al., 2000; van Velzen & Eimer, 2003), nonlater-alized cues (Jongen et al., 2007), neutral noninformative cues(Jongen, Smulders, & van Breukelen, 2006; Luck et al., 1994;McDonald & Green, 2008; Posner, 1980; Slagter, Kok, Mol,Talsma, & Kenemans, 2005; Talsma, Slagter, Nieuwenhuis,Hage, & Kok, 2005), or including trials in which the subject hasto attend to or interpret the cue but not perform any further task(Grent-’t-Jong & Woldorff, 2007).
However, as used, neutral noninformative, nonlateralized, orinterpret-cue controls do not target the main distinction that hasto be made in order to specify brain activity engaged by displac-ing the focus of attention away from fixation. For nonlateralized,noninformative, or neutral cues, this is because the subsequenttask is always required at the (otherwise cued) peripheral loca-tions. As a number of studies show (de Gonzaga Gawryszewski,Riggio, Rizzolatti, & Umilta, 1987; Jongen et al., 2006; Luck etal., 1994; Yantis & Jonides, 1990), after a neutral cue, subjectswill either expand their attentional focus or divide their expec-tations between possible target locations. As the attentional focushas been shown capable of adopting rather complicated spatialdistribution patterns (Malinowski, Fuchs, & Muller, 2007;Muller, Malinowski, Gruber, & Hillyard, 2003), it seems un-likely that no attentional displacement (i.e., spatial modulation)will occur after noninformative cues with peripheral, behavior-ally relevant stimuli. On the other hand, for the interpret-cuecondition, a different but equally problematic situation arises:Because subjects know that no target will appear after the cue,there is no relevant task at fixation and theFcovertFdistribu-tion of attention is left uncontrolled.
In this study we took a different approach to this problem byusing an informative no-shift cue to instruct the subject that abehaviorally relevant task would have to be done at fixation.
This way, we used a natural control as a contrast for covert shifts,namely, attending to something where one is looking.We appliedthis approach to ask whether alpha band activity over parietalregions ipsilateral or contralateral to the attended hemifield wasspecifically related to the covert displacement of visuo-spatialattention.
Methods
ParticipantsTwenty-five college student volunteers initially participated inthis study. Because of excessive blinking or failure to comply withfixation during the task, 7 subjects were rejected for furtheranalysis (see Data Preprocessing below for details). Thus, theresults from 18 subjects (7 women) are reported here (meanage5 24.7, range5 20–31). All participants were right-handed,had normal or corrected-to-normal vision, and reported nocolor-vision deficiency. Participants gave informed consent andhad no history of drug abuse or neurological or psychiatric con-ditions. The Ethics Committee of Pontificia UniversidadCatolica de Chile approved the study, and all experiments wereperformed at the Cognitive Neuroscience Laboratory of the De-partment of Psychiatry of the University.
Stimuli and ProcedureAmodified version of the spatial-cueing paradigm (Posner, 1980)was used in this study (see Figure 1). All stimuli were presentedon a black background on a computer screen situated 57 cmaway from the subject using Stim 1.0 software (CompumedicsNeuroscan, Texas, USA). The fixation symbol was composed oftwo white small concentric circles flanked by white left- andrightward pointing arrowheads extending in total 1.51 horizon-tally and 0.41 vertically of visual angle. Centered at 81 to the leftand right of the fixation symbol, two thin white vertical rectan-gles of 1 ! 21were presented. These lateral rectangleswere addedto indicate the peripheral locations and avoid visual search in thetime period between cue and stimulus. Both the central fixationsymbol and the two lateral rectangular outlines were presentthroughout the entire task without interruption except betweenblocks (see below). Subjects maintained fixation on the centralsymbol throughout the entire block. Each trial began at t5 0 (seeFigure 1), and, after an intertrial interval of 600, 900, or 1200ms,the central fixation symbol changed according to one of threepossibilities: either the left- or the right-pointing arrowhead orthe central concentric rings briefly changed from white to blue(150 ms), cueing the subject as to where to orient his attentioncovertly: left, right, or maintain center (no-shift), respectively.Each cueing direction occurred in 33% of trials in randomizedorder. Target or distracter stimuli (150 ms duration) were pre-sented 1100 ms after cue onset in one of the three possible lo-cations (center, left peripheral, or right peripheral). All stimuli(target and distracters) consisted of two small colored squares(0.61/side) presented in vertical arrangement separated by 0.61.Red (RGB: 255, 0, 0), magenta (RGB: 255, 0, 255), yellow(RGB: 235, 255, 0), and green (RGB: 155, 255, 0) colors wereused, and all combinations excluding same-color arrangementwere presented. In the central location, stimuli appeared aboveand below the fixation symbol with the same size and spacingbetween colored squares as the lateral stimuli. The trial ended1100 ms after the presentation of the target/distracter stimulus,
Attention shifts and parietal alpha band activity 313

although the central fixation symbol and the lateral rectanglesremained present continuously so that the subject did not knowwhen the next trial would commence.
A total of 1,062 trials were presented to each subject in sixblocks of 10.5 min each, with 5-min pauses between blocks and alonger pause halfway through the experiment. Target stimuli(11.8% of all trials) were 100% valid with respect to the cuedlocation, and subjects were informed of this fact prior to theexperiment, stressing that they should always comply with thecue direction in order to detect the corresponding target. Addi-tionally, distracters (88.2% of all trials) were presented 50%valid and 50% invalid with respect to the cued location. Whenlateral shifts were cued, invalid distracters were presented atthe opposite (peripheral) location and never at fixation. Whenno-shift was cued, invalid distracters appeared with equal prob-ability at either left or right positions. Therefore, in order to keepa comparable number of trials across conditions, when attentionwas cued to stay at fixation, ERP waveforms to lateral stimuliwere obtained, collapsing across all (left and right) invalid dis-tracters. Subjects were instructed to ignore invalid distracters andmaintain attention on the central fixation. Target stimuli wereused only to control for task performance, and all further anal-ysis of electrophysiological data was done on distracters.
At the beginning of each block, the subject was informedabout which of the four colors would be considered as the targetand instructed to indicate its presence among the two coloredsquares by pressing a button as fast as possible. Subjects wereasked to withhold response if the target color was not present atthe cued position. Response hand was alternated halfwaythrough the experiment, and the order was counterbalancedacross subjects. Color combinations prescribed two possibletypes of blocks (i.e., difficulty levels) based on differences be-tween the target color and the distracters: red and magenta wereexpected to be easier whereas yellow and green were expected tobe more difficult to discriminate. Separate analysis of perfor-mance during the two types of blocks was used to rule out un-specific effects such as effort or alertness (see Discussion). Theabsence of differences in alpha band power between difficultylevels despite significant behavioral differences suggested thatchanges in oscillatory activity were not related to unspecificeffects (see Supplementary Material), so we chose to collapseacross levels in all further analysis.
Behavioral MeasurementsTask performance was studied through reaction times (RT), hitrate (HR), and false alarm (FA) rate for all target trials. Anybutton press occurring prior to 200 ms or after 1100 ms post-target was excluded from the analysis. The hit rate was defined asthe percentage of correctly identified targets. The false alarm ratewas defined as the percentage of distracters identified as targets(Stanislaw & Todorov, 1999).
Electrophysiological RecordingsContinuous EEG recording was performed with an 80-electrodeNuAmps EEG System (Compumedics Neuroscan). Subjectsused a chin rest throughout the entire experiment except duringpauses. All impedances were kept below 10 kO, or else the elec-trode was rejected. Electrode impedance was retested duringpauses to assure stable values throughout the experiment. Be-cause of hardware constraints of the setup, all electrodes werereferenced to CPz during acquisition but off-line re-referenced toaveraged mastoids. The signal was bandpassed filtered between0.1 and 100 Hz prior to being digitized at 1 kHz. Diagonalelectro-oculogram (EOG) was obtained with two electrodes po-sitioned in the outer canthi of the eyes, displaced 1 cm upward onthe left derivation and 1 cm downward on the right derivation(Jervis, Ifeachor, & Allen, 1988). All recordings were acquiredusing Scan 4.3 software (Compumedics Neuroscan) and storedfor off-line treatment.
Data PreprocessingBecause of the importance of ensuring adequate fixation for co-vert visuo-spatial orienting, detection of eye movements wasperformed in two steps. First, an automated procedure imple-mented in the acquisition software was used, and any segment ofthe data presenting voltage deflections exceeding " 60 mV wasrejected. Full trials (# 1700 ms to 1100 ms around target/dis-tracter presentation) were obtained from the remaining data, andonly data from subjects with 60 or more trials per condition afterthis step were kept. This resulted in the rejection of data from 5of the 25 initial subjects duemainly to excessive blinking. Second,the resulting set of trials were linearly detrended, using a # 1700to # 1050-ms (# 600 to # 50 ms relative to cue presentation)baseline segment as reference for the linear trend and then vi-sually inspected using EEGLAB v6.03b Toolbox (Delorme &Makeig, 2004). Any trials presenting further artifacts or evi-dences of horizontal or vertical eye movements were rejected andonly data from subjects with 30 trials or more per condition afterthis second step were retained for further analyses, resulting inthe rejection of data from 2 other subjects. After this procedure,the average number of clean trials was 97.3 for shift left(SD5 25.4, range 46–138), 94.6 for shift right (SD5 27.5, range39–134), and 97.1 for no shift (SD5 27.7, range 41–139). Sta-tistical analysis showed that there were no significant differencesin the number of kept trials between the three attentional con-ditions, F(2,34)5 0.95, p5 .4, as well as no significant pairwisedifferences (po.05, Bonferroni corrected comparisons, withinMS5 44.365, df5 34). To further control for eyemovements, weobtained average EOGs for all three conditions following thesame processing for ERPs (see below). The average EOG wave-forms for the remaining trials for all three conditions is presentedin Figure 3D, below, as left–right difference and confirms thatno systematic eye movements were associated with any of thethree conditions.
314 D. Cosmelli et al.
Figure 1. Schematic depiction of experimental paradigm. Blackrectangles represent screens. Horizontal arrangements representsuccessive views. Vertically arranged screens represent alternative views.Fixation and lateral outlines were present throughout the entire block.The fixation figure has been augmented in the first two verticalarrangements for presentation purposes.
Data AnalysisAll subsequent data analysis was done with in-house imple-mented Matlab 7.1 (The Mathworks Inc.) routines. Two setsof analysis were performed on the data: ERPs to cues and dis-tracters and oscillatory activity in the alpha band during thecue–stimulus interval.
ERPs to the cues and distracters were obtained by averagingall trials for each condition, electrode, and subject separately, andthen filtering the resulting data to reject frequencies above 30 Hz.Average amplitude values for the different ERP peaks were ob-tained in prespecified windows for ERPs to the cue and aroundindividual peak values for early potentials evoked by the dis-tracters.
A pilot study with 8 subjects and a passive task was run toassess the dependency of early (170–220 ms) ERPs to the cue’sphysical properties. We presented the same cues as a distracter ina go/no-go task at fixation. Although the short intertrial intervalwe used to ensure a high number of trials precluded analysis ofoscillatory activity, we observed that, despite the absence of in-structions to orient attention, a contralateral occipital negativityto the lateral cues and a bilateral occipital negativity to the centralcues was apparent (data not shown). This reproduces the earlynegative deflections seen in the present paradigm and is in linewith the results of van Velzen and Eimer (2003). We therefore donot include further analysis of this time window here.
Following Kiss, van Velzen, and Eimer (2008), we measuredthe average amplitude in a 350–650-ms postcue window to study
the attention-directing anterior negativity (ADAN) component,in bilateral fronto-central electrodes (see inset in Figure 3, below,for the corresponding electrode locations). Additionally, wemeasured a midline parieto-central broad P3-type positivity(250–400 ms postcue) and a midline fronto-central contingentnegative variation (CNV; 400–50 ms prestimulus). During thepoststimulus interval, we focused on the early P1 and N1 wavesevoked by valid and invalid distracters. We first determined thetwo posterior electrodes (contralateral to attended stimulus) thatshowed the strongest voltage deflection for both P1 and N1components in the average across subjects. Electrodes 22 (leftoccipital lateral) and 65 (right occipital lateral) were chosen (seeinset in Figure 2), and a 20-ms window average was obtainedfrom the voltage traces of these electrodes, centered on each peakand for each subject individually. Average peak latencies acrosssubjects were 98.1 ms for P1 (SD5 15.8 ms) and 168.1 ms for N1(SD5 30.2 ms).
To study oscillatory activity, data were subsampled at 250 Hzprior to applying a seven-cycleMorlet wavelet transform (Tallon-Baudry & Bertrand, 1999) in 35 frequency steps from 4 to 80 Hzon single trials. Raw time-frequency charts for each trial wereaveraged across trials in order to obtain the induced power foreach condition. Values were log-transformed and baseline-cor-rected with respect to a # 400 to # 50-ms window prior to cuepresentation; all power values are therefore presented here asrelative change with respect to this precue period. Because wewere interested in discriminating posterior ipsilateral versus con-
Attention shifts and parietal alpha band activity 315
B
A
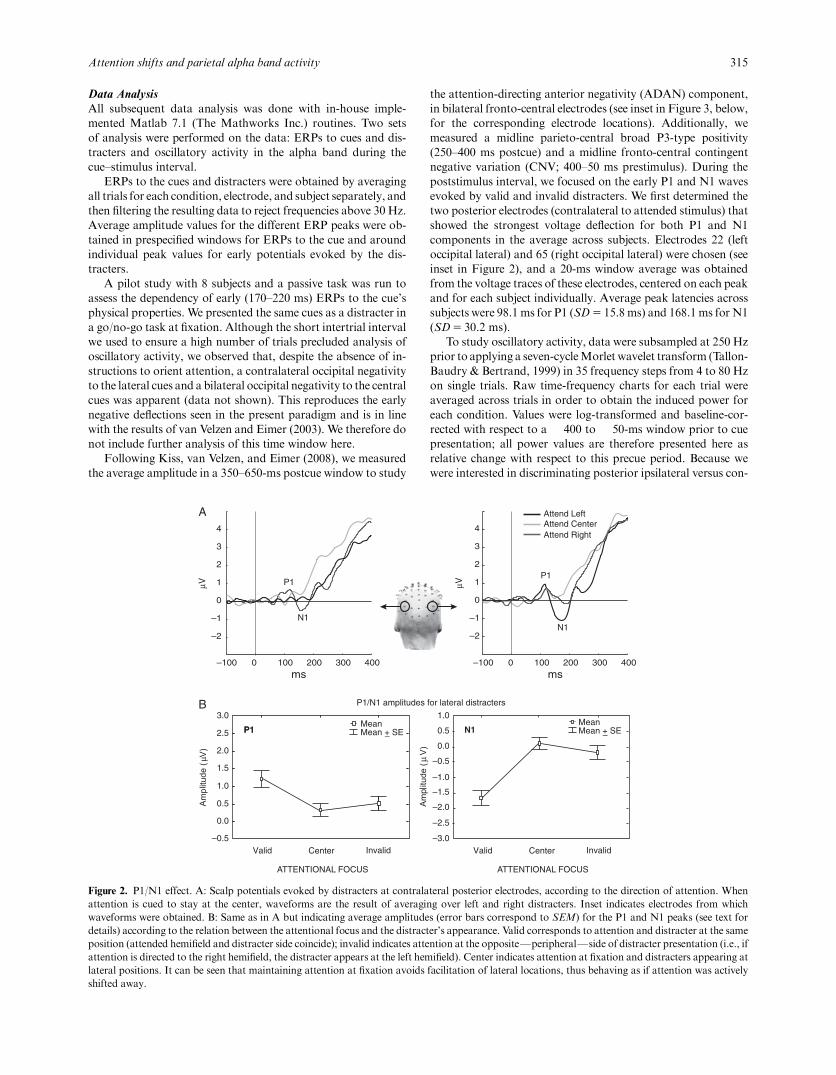
Figure 2. P1/N1 effect. A: Scalp potentials evoked by distracters at contralateral posterior electrodes, according to the direction of attention. Whenattention is cued to stay at the center, waveforms are the result of averaging over left and right distracters. Inset indicates electrodes from whichwaveforms were obtained. B: Same as in A but indicating average amplitudes (error bars correspond to SEM) for the P1 and N1 peaks (see text fordetails) according to the relation between the attentional focus and the distracter’s appearance. Valid corresponds to attention and distracter at the sameposition (attended hemifield and distracter side coincide); invalid indicates attention at the oppositeFperipheralFside of distracter presentation (i.e., ifattention is directed to the right hemifield, the distracter appears at the left hemifield). Center indicates attention at fixation and distracters appearing atlateral positions. It can be seen that maintaining attention at fixation avoids facilitation of lateral locations, thus behaving as if attention was activelyshifted away.
tralateral variations in alpha power, all statistical analysis oftime-frequency data was performed on the regional average oftwo parietal electrodes for each hemisphere (see inset in Figure 4,below).We found important differences in the frequency locationof the alpha peak for the different subjects’ spectra, sowe decidedto adjust for individual alpha frequency (see SupplementaryFigure S1). Moreover, because upper and lower alpha subbandscan react in different ways according to the task (Freunberger etal., 2008; Klimesch, 1999), we separated for each subject a lower(four frequency bins below and including the peak bin) and anupper (three frequency bins above the peak bin) alpha band. Allfurther analyses were performed on the corresponding individ-ually adjusted subbands separately.
Statistical Designt test statistics for dependent samples were used to compare thebehavioral performance between the different conditions. Allstatistical analyses of electrophysiological measures were per-formed using the individual values obtained from the electrodes/latencies described above in a repeated measures analysis ofvariance (ANOVA) design. For the P1 and N1 components thefactors Visual Field (two levels: left and right) and AttentionalCondition (three levels: lateral valid, central invalid, and lateralinvalid) were considered. For the cue-related ERP, the repeatedmeasures ANOVA included the factors Hemisphere (two levels:right hemisphere and left hemisphere) and Cue relative direction(two levels: ipsilateral and contralateral). Oscillatory alpha-bandactivity during the cue–stimulus interval was statistically testedusing repeated measures ANOVA for each frequency band.Within-subject factors were Cue (three levels: shift left, no shiftand shift right), Hemisphere (two levels: left parietal and rightparietal), and Cue-Stimulus Interval Latency (three levels: L1through L3). In all cases Greenhouse–Geisser corrections wereapplied to compensate for violations of assuming sphericity, and
planned univariate comparisons were carried out when necessary(see Results section for further details). Scalp interpolation on astandard head (obtained from the EEGLAB toolbox;Delorme&Makeig, 2004) was used only for presentation purposes.
Results
Behavioral PerformanceMean RTs to left, right, and central targets were 517 ms(SD5 81.7 ms), 515.1 ms (SD5 86.8 ms), and 468 ms(SD5 74.6 ms), respectively. Separate two-tailed t tests fordependent samples revealed no significant differences in RT toleft versus right visual field targets (t5 0.33, p5 .74) and sig-nificant differences between both left (t5 7.57, po.05) and right(t5 6.63, po.05) versus central targets. Mean HR to left, right,and central targets were 80.7% (SD5 16.6%), 81.8%(SD5 16.2%), and 94.3% (SD5 8%), respectively. A similarpattern was obtained for HR, so that no significant differencebetween left and right visual fields was observed (t5 # 0.87,p5 .39), whereas HR to targets in both left (t5 # 5.93, po.05)and right (t5 # 5.42, po.05) visual fields were significantlylower than HR to central targets. Mean FA rates to left, right,and central distracters were 7.7% (SD5 9.9%), 9.7%(SD5 10.8%), and 1.2% (SD5 2.8%), respectively. When sub-ject to statistical testing, FA rates followed a slightly differentpattern. Although differences between lateral versus central tar-gets behaved as in RTs and HRs (t5 4.41 and t5 5.16 for leftand right vs. center, respectively, both po.05), we found signifi-cantly higher FAs for distracters on the right visual field whencompared to the left visual field (t5 # 2.57, po.05).
Electrophysiology
ERPs to distracters. In order for the central no-shift condi-tion to be an adequate control, it is necessary to verify that at-
316 D. Cosmelli et al.
A B
DC
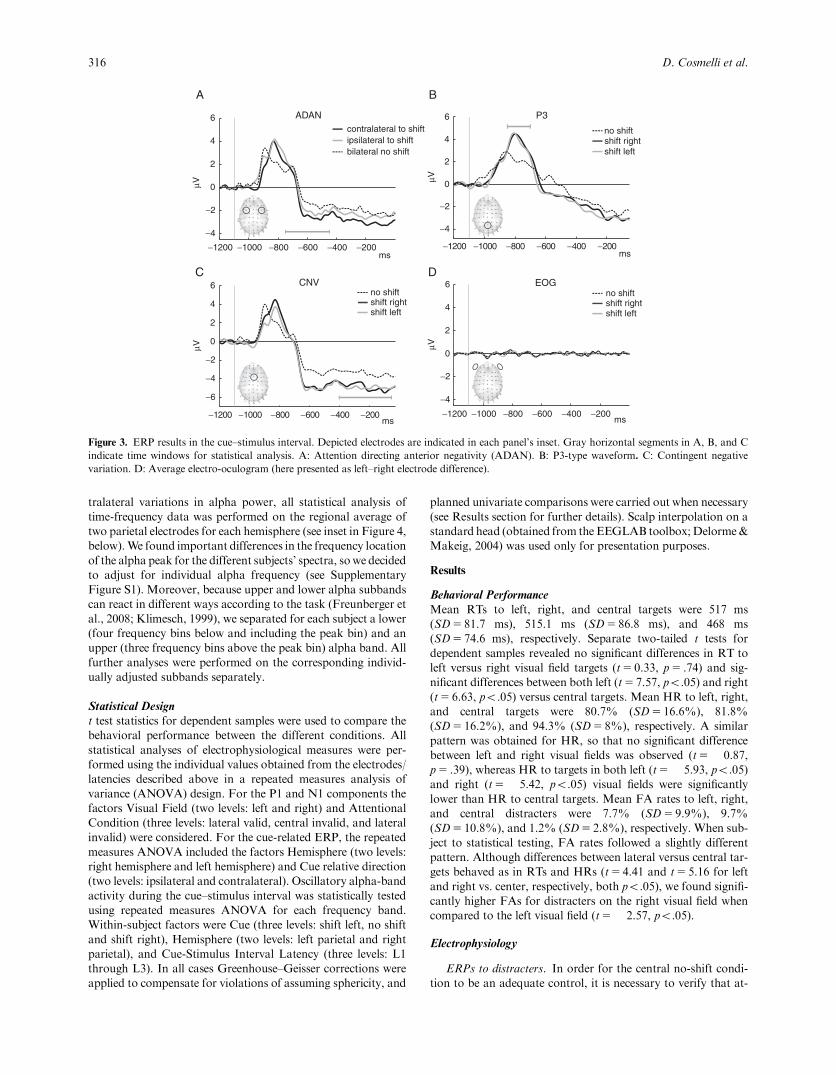
Figure 3. ERP results in the cue–stimulus interval. Depicted electrodes are indicated in each panel’s inset. Gray horizontal segments in A, B, and Cindicate time windows for statistical analysis. A: Attention directing anterior negativity (ADAN). B: P3-type waveform. C: Contingent negativevariation. D: Average electro-oculogram (here presented as left–right electrode difference).
tention is not distributed to lateral positions when the subject isinstructed to perform the task at fixation.We hypothesized that ifattention is maintained at fixation as a consequence of the no-shift informative cue, invalid distracters will evoke early (P1/N1)brain responses comparable to when attention is directed to lat-eral positions and the stimulus appears at the opposite location.This is in contrast to neutral noninformative cues, where a diffuseor divided attention state is induced and therefore an interme-diate response between valid and invalid conditions is observed(Hillyard et al., 1994; Jongen et al., 2006; McDonald & Green,2008; Wright, Richard, & McDonald, 1995).
Figure 2 presents the results obtained for the P1/N1 analysison the distracters. It can be seen that when compared to valid
distracters, invalid distracters evoke a reduced P1/N1 responseafter lateral shifts of attention. When attention is cued to stay atfixation, peripheral distracters evoke a smaller P1/N1 responsethat is comparable to when attention has been shifted away. Toquantify these apparent differences, the data obtained in the P1and N1 range were subject to a separate repeated measuresANOVA with the factors Visual Field (two levels: left and right)and Attentional Condition (three levels: lateral valid, central in-valid, and lateral invalid). For the P1 peak, statistical analysisrevealed no main effect of Visual Field, F(1,17)5 1.71, p5 .21,indicating no overall differences regarding where the stimuluswas presented. A main effect of Attentional Condition wasfound, F(2,34)5 5.42, po.05 (see Figure 2B, left: P1 amplitude).
Attention shifts and parietal alpha band activity 317
A
B
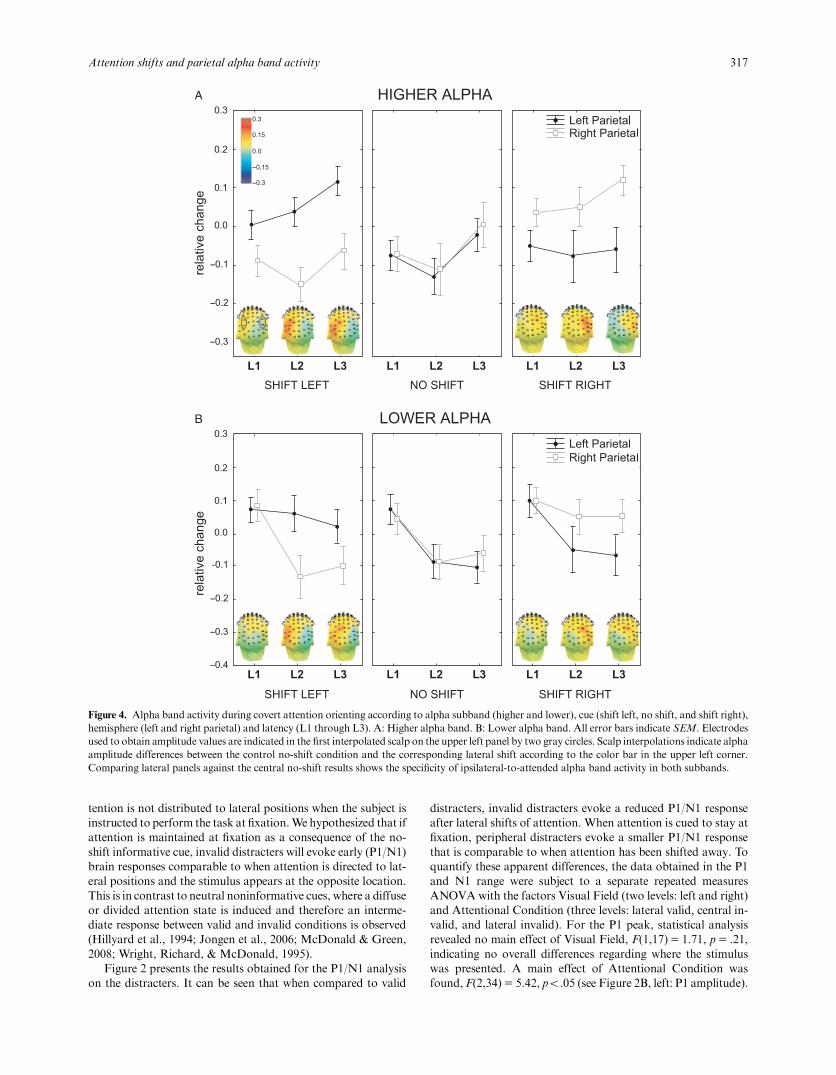
Figure 4. Alpha band activity during covert attention orienting according to alpha subband (higher and lower), cue (shift left, no shift, and shift right),hemisphere (left and right parietal) and latency (L1 through L3). A: Higher alpha band. B: Lower alpha band. All error bars indicate SEM. Electrodesused to obtain amplitude values are indicated in the first interpolated scalp on the upper left panel by two gray circles. Scalp interpolations indicate alphaamplitude differences between the control no-shift condition and the corresponding lateral shift according to the color bar in the upper left corner.Comparing lateral panels against the central no-shift results shows the specificity of ipsilateral-to-attended alpha band activity in both subbands.

Planned comparisons between lateral valid and lateral invalidconditions revealed a significantly higher P1 amplitude for thevalid condition, F(1,17)5 8.02, po.05, and no differences be-tween central invalid and lateral invalid conditions,F(1,17)5 1.15, p5 .3. The two-way Visual Field ! AttentionalCondition interaction was not significant, F(2,34)5 0.72,p5 .49. For the N1 peak, a similar pattern emerged from thestatistical analysis (see Figure 2B, right: N1 amplitude): Wefound no main effect of Visual Field, F(1,17)5 1.72, p5 .21 ortwo-way Visual Field ! Attentional Condition interaction,F(2,34)5 2.88, p5 .07. As for the P1 peak, a main effect ofAttentional Condition was also detected, F(2,34)5 12.67,po.05. Subsequent planned comparisons revealed that N1 am-plitude was more negative for lateral valid than lateral invalid,F(1,17)5 26.96, po.05, and showed no difference between cen-tral invalid and lateral invalid conditions, F(1,17)5 1.49,p5 .24.
These results reproduce previous reports (Hillyard & Anllo-Vento, 1998; Luck, Heinze, Mangun, & Hillyard, 1990) regard-ing the effects of lateral shifts of attention, confirming that thesubjects correctly oriented their attention in our experiment.More importantly, they show that when attention is cued to stayat fixation, peripheral stimuli evoke responses that are indistin-guishable from those evoked when attention has been activelyshifted away. The no-shift informative cue is, therefore, an ad-equate control condition because, when presented, it does notproduce facilitation of peripheral positions.
Cue-related ERPs. The results for the evoked responses in thecue–stimulus interval are summarized in Figure 3. As shown inpanel 3A, we observed a clear frontal negativity contralateral tothe direction of attentional orienting in the 350–650-ms postcuewindow, consistent with a traditional ADAN (Hopf &Mangun,2000; Nobre et al., 2000; van Velzen & Eimer, 2003). We ana-lyzed these apparent differences first by comparing contralateralversus ipsilateral ERP amplitudes using a repeated measuresANOVA with factors Hemisphere (two levels: right hemisphereand left hemisphere) and Cue Relative Direction (two levels:ipsilateral and contralateral). We observed a main hemisphereeffect whereby the left frontal regions showed a more negativedeflection than the right frontal regions, F(1,17)5 9.88, po.05.The classical ADAN effect was observed as themain effect of cuerelative directionality, F(1,17)5 6.11, po.05, indicating that co-vert orienting of attention was associated with a more negativepotential over contralateral frontal electrodes (corresponding toFC5/6; see inset in Figure 3A). No two-way interaction wasobserved, F(1,17)5 0.03, p5 .86. Interestingly, when we ana-lyzed the threeFabsoluteFcueing directions (shift left, shiftright, and no shift) in a separate ANOVA, we observed a maineffect of cue direction, F(2,34)5 3.63, po.05), indicating thatbesides being specifically contralateral to the shift direction, bothleft and right shifts produce a more negative deflection than theno-shift instruction. We found no evidence of a late directingattention positivity (LDAP) in our data, observing only a slighttendency to a positive deflection over the right posterior elec-trodes upon left shifts that did not reach significance,F(1,17)5 1.25, p5 .28, Univariate Test of Significance.
Besides the more traditional cue-related potentials, we ob-served two stereotyped deflections in the cue–stimulus interval.As shown in Figure 3B, a clear P3-type positivity peaking around300 ms after the presentation of the cue is evident. Analysis ofvariance of the amplitude in the 250–400-ms postcue window
confirmed a clear cue direction effect, F(2,34)5 13.82, po.05,due to a more positive amplitude during lateral shifts. A post hocBonferroni test (within MS5 1.1808, df5 34) showed that therewas no difference between lateral shifts in the amplitude of thispotential but confirmed the apparent difference between no shiftand both left and right shifts (both po.01). A similar patternemerged for the CNVanalysis (see Figure 3C), which showed thatlateral shifts were associated with a stronger negative deflection inthe 400–50-ms prestimulus interval, F(2,34)5 6.90, po.01. Apost hoc Bonferroni test again showed no differences betweenlateral shifts and amore negative amplitude for both left and rightshifts when compared to the no-shift condition (both po.01).
Oscillatory alpha-band activity during the cue–stimulusinterval. We then turned to study the specificity of alpha bandresponses to attentional displacement by contrasting lateral shiftswith the no-shift control condition. To quantify dynamicalchanges in alpha band power, we divided the 1100-ms cue–stim-ulus interval into three equal time windows (latencies, L) of 365ms each (L1:# 1100 to # 735 ms; L2: # 734 to # 370 ms; L3:# 369 to # 4 ms) and compared the average alpha band am-plitude over parietal regions in these windows for all three cues.
Average values of the corresponding time–frequency regions,defined by latency and frequency limits, were subject to three-wayrepeated measures ANOVA designs for both upper and lower al-pha subbands separately. Within-subjects factors were Cue (threelevels: shift left, no shift, and shift right), Hemisphere (two levels:left parietal and right parietal), and Latency (three levels: L1through L3). No main effects of any factor were observed in eitheralpha band except for a latency effect in the lower alpha subband,F(2,34)5 5.56, po.01, due to an overall drop in power during L2andL3of the cue–stimulus interval.A two-wayCue ! Hemisphereinteractionwas evident in both higher, F(2,34)512.43, po.01, andlower, F(2,34)55.68, po.01, alpha subbands. This was due tooverall higher amplitude over parietal regions ipsilateral to the di-rection of attention when covert orienting was cued. No Cue !Latency (higher alpha: F[4,68]50.80, p5 .53; lower alpha:F[4,68]50.35, p5 .84) or Hemisphere ! Latency (higher alpha:F[2,34]50.61, p5 .55; lower alpha: F[2,34]5 2.10, p50.14) in-teractions were detected. However, a clear three-way Cue ! Hemi-sphere ! Latency interaction was observed for both higher,F(4,68)5 2.83, po.05, and lower, F(4,68)58.04, po.01, alpha.Figure 4 is the graphical representation of this three-way interactionand presents the results obtained for both alpha subbands, accord-ing to hemisphere, cueing condition, and cue–stimulus interval la-tency. Scalp interpolations in lateral panels present the differencebetween lateral shifts and the no-shift condition (i.e., the differencebetween lateral and central panels).
A clear pattern can be observed when the three conditions arecontrasted (post hoc Bonferroni-corrected comparisons, onlypo.05 are informed; lower alpha, within MS5 0.00808; higheralpha, within MS5 0.00769; both df5 68). When comparingonly left and right shifts (i.e., ignoring central panels in Figure4A,B) it can be observed that ipsilateral (contralateral) parietalactivity was always higher (lower) than contralateral (ipsilateral)activity in mid- and late latencies for both alpha bands. Yet,taken in isolation, these results cannot distinguish whether ipsi-lateral increase or contralateral drop is specific for the process ofcovert orienting; comparing them against the no-shift conditionis necessary to resolve this ambiguity. Indeed, we observed thatwhen attention was cued to stay at fixation, left and right parietalregions exhibited the samebehavior for bothhigher and lower alpha
318 D. Cosmelli et al.

bands across the entire cue–stimulus interval (Figure 4A,B respec-tively, central panel): Amplitude in the lower alpha band droppedfrom L1 to L2 and then stayed at the same level until the appear-ance of the stimulus; in the higher alpha the pattern was somewhatdifferent, showing an initial drop and a subsequent tendency toincrease back to baseline levels. However, when attention wasshifted away from fixation toward lateral positions, only the ipsi-lateral parietal regions showed activity that was different from theno-shift condition. This was observed in both subbands, albeit withdifferent dynamics (see lateral panels in Figure 4, A vs. B):Whereasin the lower alpha band ipsilateral parietal regions maintained alevel near to baseline activity, showing only a slight drop, amplitudein the higher alpha band showed a consistent increase throughoutthe cue–stimulus interval. Alpha band amplitude over the contra-lateral hemisphere, on the other hand, was indistinguishable fromthe no-shift condition, as can be seen by comparing in Figures 4Aand 4B lateral versus central panels.
These results show that alpha band activity over ipsilateral-to-attended parietal regions is specifically engaged during thedisplacement of the focus of attention away from fixation. Theylikewise show that alpha band activity over contralateral parietalregions behaves the same way as if no shift was required.
Discussion
The main results of this study can be summarized as follows: (i)Behavioral data confirmed that subjects were able to perform thetask consistently, but, as expected, performance was always muchbetter at fixation than at the periphery. (ii) The use of a centralinformative no-shift cue was effective in keeping attention at fix-ation as revealed by smaller P1/N1 amplitudes evoked by invalid(lateral, peripheral) stimuli. (iii) The cue-relatedADANcomponentwas consistently more negative over frontal regions contralateral tothe shift of attention and more pronounced than the no-shift con-dition. Additionally, P3-type and CNV waveforms in the cue–stimulus interval were enhanced in both shift conditions as com-pared to the no-shift task. (iv) When attention was covertly dis-placed from fixation, alpha band activity over the parietal regionsipsilateral to the attended hemifield (contralateral to the ignoredside) was enhanced relative to the no-shift condition. Alpha bandactivity over parietal regions contralateral to attended hemifield, onthe other hand, were comparable to the no-shift task. In the fol-lowing we discuss a number of points raised by these results.
The informative no-shift cue/central task conditionTraditional studies in attentional shifting follow Posner’s (1980)now classical cue–target paradigm whereby the subject is in-structed to direct attention to one of two possible peripherallocations upon endogenous or exogenous cues. In some cases,neutral (noninformative regarding the most probable location ofstimulus appearance) cues are used. Usually, brain activity iscontrasted between conditions, and differences between shiftingto one side or another are revealed. However, such comparisonscan only show what is going on in the brain that differentiatesshifting to the right from shifting to the left (or any other alter-native locations used) but donot contrast shifting attention to theperiphery against not doing so (Nobre et al., 2000). By incor-porating a behaviorally relevant no-shift condition at fixation,we sought to transcend the limitations of using neutral or inter-pret cue conditions as contrasts that do not ensure the absence ofattentional covert modulation (see de Gonzaga Gawryszewski etal., 1987; Jonides, 1980).
Although some studies have dealt with the question of differ-ences between central and peripheral attention (de Gonzaga Ga-wryszewski et al., 1987;Miniussi, Rao, &Nobre, 2002; Neville &Lawson, 1987; Tassinari, Aglioti, Chelazzi, Marzi, & Berlucchi,1987), these have been mainly behavioral or have focused ontargets rather than on cues, therefore being unable to specifybrain activity related to the orientation phase. Nevertheless, theydo show that attention can be controlled to stay put at fixationthrough central cues (Yantis & Jonides, 1990), and that this hasbehaviorally analogous consequences as attending to a periph-erally cued location in the sense that nonattended locations sufferperceptual detriment. Our results confirm and extend these find-ings: Keeping attention at fixation by central informativeno-shift cues avoids any lateral facilitation, as revealed by the re-duced P1/N1 to subsequent (invalid) peripheral stimuli (Figure 2).We believe that the incorporation of this condition is therefore avaluable contrast for the study of brain activity specifically re-lated to the covert orienting of visuo-spatial attention.Moreover,we propose to consider distinguishing between noninformative(neutral) cues, which do not necessarily ensure a no-shift con-dition (McDonald & Green, 2008), and the more explicit infor-mative no-shift cue as used here, which is tailored to keep theattentional focus at fixation.
Behavioral results did show, however, that the task at fixationwas always easier than the lateral (peripheral) tasks. This resulthas important consequences for the validity of the no-shift con-trol as used in our study. Indeed, because alpha band activity isknown to be highly dependent on changes in alertness or mentaleffort (Dockree, Kelly, Foxe, Reilly, & Robertson, 2007; Keil,Mussweller, & Epstude, 2006; Ray & Cole, 1985), it poses thequestion as to what extent unspecific effects such as these mightbe at play when the subject is confronted with the more de-manding task of shifting attention away from fixation. One pos-sibility would be to match the central and lateral tasks fordifficulty. However, this has the drawback ofmaking the no-shiftcondition a less ecologically valid control: Displacing the focus ofattention from where one is fixating is inherently effortful andenergy demanding, whereas fixating where one expects some-thing to happen is comparatively easy. The validity of the centralcontrol, therefore, cannot depend on it demanding equivalenteffort, because voluntary covert displacements of attention arenever effortless (James, 1950). Trying to match these conditionsin terms of effort would mean distancing oneself from the wayvoluntary covert shifts of attention are undertaken naturally.
This said, and as pointed out by an anonymous reviewer, theexistence of two difficulty levels in our task offers a way of es-timating if effort has a substantial impact on our results. If effortor alertness were responsible for changes in alpha band dynamicsduring covert shifts, the two difficulty levels would differ in thisvariable. As the analysis presented in the supplementary materialshows, however, no significant changes in alpha band activitywere detected between difficulty levels during lateral shifts despiteclear behavioral differences. This suggests that the modulationswe see cannot be explained by unspecific changes in effort oralertness but seem more likely to reflect a topographical redis-tribution of cortical activity in the alpha band related to brainmechanisms underlying covert attentional shifts.
Cue-Related ERPsAlthough we did not find evidence for a clear LDAP in our data,we observed a significant ADAN waveform in the 350–650-mspostcue window (Figure 3). In addition to showing the tradi-
Attention shifts and parietal alpha band activity 319

tional contralateral preference, we observed that both shifts wereassociated with more negative values as compared to the no-shiftcondition. This suggests that, although some of the brain pro-cesses underlying the ADAN might overlap with activity relatedto the control of nonspatial attention, an important part likelyreflects activity that is specific for the covert orienting of attention(Jongen et al., 2007; Kiss et al., 2008; van Velzen & Eimer, 2003).Furthermore, our results appear consistent with the study byGrent-’t-Jong and Woldorff (2007) that showed that a frontalnegative deflection beginning around 400 ms postcue remainedwhen an interpret-cue conditionwas subtracted from lateral shifts.
The P3-type component we observed was much greater inamplitude in the shift versus the no-shift condition, especiallyover superior parietal regions, but with no evident lateralization.This could indicate the presence of an underlying location-inde-pendent mechanism for displacing the focus of attention. Indeed,in a recent study (Slagter, Kok, Mol, Talsma, et al., 2005) asimilar pattern of parieto-central P3-type positivity around 260–520 ms was observed when spatial versus nonspatial attentionalcontrol was compared: P3-type activation was stronger for trialswhere the cued feature had changed regardless of whether thefeature was spatial or not. In view of these results, the authorssuggested that this P3 component could be indicative of processesunderlying the generic switching (updating) of the ongoing at-tentional set (see also Slagter, Kok, Mol, & Kenemans, 2005).This interpretation would be compatible with our results, as bothshift-left and shift-right conditions imply an important cognitivechange with respect to the previous moment, whereas the centralcueing ismore of an ongoingmaintenance and focalization of thecurrent state. Finally, an influential work by Corbetta et al.(1993) used a central-cueing control in a positron emission to-mography study. The authors showed that, when compared tolateral shifts, central cues produced lower overall superior frontaland superior parietal activity, which is in line with the P3-typeresults presented here (see Figure 3B).
The last cue-related component we studied was the CNV(Brunia & van Boxtel, 2001; Walter, Cooper, Aldridge,McCallum, & Winter, 1964). The results presented here are inoverall agreementwith previous reports regarding the presence ofthe CNV component, as stimulus appearance was completelypredictable in our setting. Interestingly, more negative valueswere observed for both attentional shifts, although there was nodifference between left versus right shifts. The neural origin of theCNV is not fully understood, but it is proposed to have frontal,fronto-central, medial frontal, and superior and medial parietalgenerators depending both on the task and whether it is an earlyor late CNV (Gomez et al., 2001; Sieroka, Dosch, Specht, &Rupp, 2003). An enhanced CNV would be compatible with anaturally more effortful gesture during covert shifts of attentionif, indeed, these were of premotor origin (Rizzolatti, Riggio,Dascola, & Umilta, 1987). One would expect a stronger involve-ment of supplementary motor cortex and related frontal andfronto-medial areas in such cases (see also Small et al., 2003).This interpretation is supported by a study on covert orienting(Wright, Geffen, &Geffen, 1995), where directional cues inducedstronger CNVwhen compared to nondirectional, neutral cues.
Alpha Band Oscillatory ActivityOscillatory activity that presented clear attention-shift-relatedmodulations was confined to a broad alpha band (6–14Hz). Thisis in linewith numerous noninvasive studies in human beings thathave shown that this electroencephalographic spectral compo-
nent is highly reactive to attentional manipulations (Babiloni etal., 2003; Bastiaansen, Bocker, Cluitmans, & Brunia, 1999;Foxe et al., 1998; Fu et al., 2001; Kelly et al., 2006; Klimesch,1999; Rihs et al., 2007, 2009; Sauseng et al., 2005; Thut et al.,2006; Worden et al., 2000). However, a number of studies havereported modulations in posterior alpha band activity whensubjects are expecting forthcoming events, even if orienting at-tention to a different location in space is not required (Babiloniet al., 2003; Bastiaansen & Brunia, 2001; Filipovic, Jahanshahi,& Rothwell, 2001). On the other hand, differential reactivity ofintra-alpha bands has been observed depending on whether thesubject has to memorize, make semantic judgments, or simplyattend to upcoming stimuli (Fink, Grabner, Neuper, & Neu-bauer, 2005; Klimesch, 1999), thus suggesting a rather complexfunctional role of this widespread cortical oscillation (Palva &Palva, 2007). In the specific case of visual attention, this raisesthe question of to what extent changes in cortical activity, inparticular occipito-parietal alpha and subbands within, are spe-cifically related to the spatial displacement of the focus of at-tention or rather reflect a complex mixture of preparation,expectation, and so forth and the actual spatial orientation pro-cesses. More specifically, an important issue that remains con-troversial in the literature pertains the relative corticaldistribution of changes in alpha power (Kelly et al., 2006; Rihset al., 2007, 2009; Sauseng et al., 2005; Worden et al., 2000):whether lateral covert shifts of attention are related to contra-lateral alpha decrease (reflecting putative facilitation of the at-tended visual field) or ipsilateral alpha increase (reflectingputative inhibition of the ignored visual field). Making this dis-tinction in light of the informative no-shift cue control has beenthe main objective of this study.
Our results are in line with the notion that the overall processof selective attention depends on a complex mechanism involv-ing both facilitation of potentially relevant features and inhibi-tion of irrelevant aspects (Hillyard et al., 1994; Rihs et al., 2007,2009; Suffczynski, Kalitzin, Pfurtscheller, & Lopes da Silva,2001). A recent study by Rihs et al. (2009) revealed both con-tralateral-to-attended decrease and contralateral-to-unattendedincrease depending onwhether the early or late phase of the cue–stimulus interval was considered, respectively. The results pre-sented here are in partial agreement with these findings and ex-tend them by suggesting potential mechanisms of suppression/facilitation in the differential dynamics present within subbandsin the alpha range (see Figure 4). Indeed, we observed a sus-tained increased activity in the higher alpha range over parietalsites ipsilateral to the attended location (contralateral to theunattended site) that became more evident in the later phase ofthe cue–stimulus interval. This increase in alpha oscillatory ac-tivity was specific for the attentional orientation process as con-trasting it to the informative no-shift cue demonstrates:Contralateral-to-attended parietal sites were indistinguishablefrom the no-shift control. These results are consistent with re-cent work on the relation between P1 and alpha band oscilla-tions showing that upper alpha band activity could bespecifically related to inhibition of task-irrelevant brain regions(Freunberger et al., 2008). In the lower alpha band, on the otherhand, parietal activity contralateral to the attended hemifieldshowed a rapid decrease that was maintained throughout thecue–stimulus interval. This result appears initially in agreementwith previous reports showing that attentional orienting is as-sociated with a specific drop in posterior alpha band power overregions that process the attended hemifield (Sauseng et al., 2005;
320 D. Cosmelli et al.

Thut et al., 2006). Yet only contralateral-to-ignored (ipsilateral-to-attended) regions showed specific differences when comparedto the no-shift control, whereas contralateral-to-attendedparietal cortices maintained the same level of activity as whenno attentional orienting was undertaken. It seems possibleto hypothesize, therefore, that facilitatory processes during selec-tive attention, if indexed by reduced posterior alpha activity, mightbe more related to expectation or general preparatory activitynecessary for the subsequent target discrimination than to theactual covert displacement of attention (see also Capotosto,Babiloni, Romani, & Corbetta, 2009; Yamagishi et al., 2008).The latter seems to be associated with selective, dynamicallycomplex, ipsilateral-to-attended augmented parietal alphaoscillations.
ConclusionsTo better understand the dynamics of EEG oscillatory activityduring visuo-spatial attention orienting, we contrasted lateral co-vert shifts with a behaviorally relevant no-shift control at fixation.In this way, we were able to disentangle which aspects of alphaband dynamics are specifically reactive to covert orienting: In theupper alpha band, a gradual increase over parietal regions ipsi-lateral to the attended hemifield was observed during lateral shifts.In the lower alpha band, a drop in power was evident duringcovert orienting over both hemispheres. However, only ipsilateral-to-attended parietal regions were different from the control,maintaining a comparatively higher level of activity. In both alphasubbands, the activity of parietal regions contralateral to the at-tended hemifield did not differ from the no-shift task.
REFERENCES
Babiloni, C., Brancucci, A., Babiloni, F., Capotosto, O., Carducci, F.,Cincotti, F., et al. (2003). Anticipatory cortical responses during theexpectancy of a predictable painful stimulation. A high-resolutionelectroencephalography study. European Journal of Neuroscience, 18,1692–1700.
Bastiaansen, M. C., Bocker, K. B., Cluitmans, P. J., & Brunia, C. H.(1999). Event-related desynchronization related to the anticipation ofa stimulus providing knowledge of results. Clinical Neurophysiology,110, 250–260.
Bastiaansen, M. C., & Brunia, C. H. (2001). Anticipatory attention: Anevent-related desynchronization approach. International Journal ofPsychophysiology, 43, 91–107.
Brunia, C. H., & van Boxtel, G. J. (2001). Wait and see. InternationalJournal of Psychophysiology, 43, 59–75.
Capotosto, P., Babiloni, C., Romani, G. L., & Corbetta, M. (2009).Frontoparietal cortex controls spatial attention through modulation ofanticipatory alpha rhythms. Journal of Neuroscience, 29, 5863–5872.
Corbetta,M.,Miezin, F.M., Shulman, G. L., & Petersen, S. E. (1993). APET study of visuospatial attention. Journal of Neuroscience, 13,1202–1226.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed andstimulus-driven attention in the brain. Nature Reviews Neuroscience,3, 201–215.
de Gonzaga Gawryszewski, L., Riggio, L., Rizzolatti, G., & Umilta, C.(1987). Movements of attention in the three spatial dimensions andthe meaning of ‘‘neutral’’ cues. Neuropsychologia, 25, 19–29.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source toolboxfor analysis of single-trial EEG dynamics including independentcomponent analysis. Journal of Neuroscience Methods, 134, 9–21.
Dockree, P.M., Kelly, S. P., Foxe, J. J., Reilly, R. B., &Robertson, I. H.(2007). Optimal sustained attention is linked to the spectral content ofbackground EEG activity: Greater ongoing tonic alpha (approxi-mately 10 Hz) power supports successful phasic goal activation. Eu-ropean Journal of Neuroscience, 25, 900–907.
Doesburg, S. M., Roggeveen, A. B., Kitajo, K., & Ward, L. M. (2008).Large-scale gamma-band phase synchronization and selective atten-tion. Cerebral Cortex, 18, 486–499.
Filipovic, S. R., Jahanshahi, M., & Rothwell, J. C. (2001). Uncouplingof contingent negative variation and alpha band event-relateddesynchronization in a go/no-go task. Clinical Neurophysiology,112, 1307–1315.
Fink, A., Grabner, R. H., Neuper, C., & Neubauer, A. C. (2005). EEGalpha band dissociation with increasing task demands. Brain Re-search. Cognitive Brain Research, 24, 252–259.
Foxe, J. J., Simpson, G. V., & Ahlfors, S. P. (1998). Parieto-occipitalapproximately 10 Hz activity reflects anticipatory state of visual at-tention mechanisms. NeuroReport, 9, 3929–3933.
Freunberger, R., Holler, Y., Griesmayr, B., Gruber, W., Sauseng, P., &Klimesch, W. (2008). Functional similarities between the P1 compo-nent and alpha oscillations. European Journal of Neuroscience, 27,2330–2340.
Fries, O., Reynolds, J. H., Rorie, A. E., & Desimone, R. (2001). Mod-ulation of oscillatory neuronal synchronization by selective visual at-tention. Science, 291, 1560–1563.
Fu, K. M., Foxe, J. J., Murray, M. M., Higgins, B. A., Javitt, D. C., &Schroeder, C. E. (2001). Attention-dependent suppression of dis-tracter visual input can be cross-modally cued as indexed by antic-ipatory parieto-occipital alpha-band oscillations. Brain Research, 12,145–152.
Gomez, C. M., Delinte, A., Vaquero, E., Cardoso, M. J., Vazquez, M.,Crommelinck, M., et al. (2001). Current source density analysisof CNV during temporal gap paradigm. Brain Topography, 13, 149–159.
Green, J., &McDonald, J. (2008). Electrical neuroimaging reveals timingof attentional control activity in human brain. Plos Biology, 6, e81.
Grent-’t-Jong, T., & Woldorff, M. G. (2007). Timing and sequence ofbrain activity in top-down control of visual-spatial attention. PlosBiology, 5, e12.
Hillyard, S., Luck, S. J., & Mangun, G. R. (1994). The cueing of at-tention to visual field locations: Analysis with ERP recordings. In H.J. Heinze, T. F. Munte, & G. R. Mangun (Eds.), Cognitive electro-physiology (pp. 1–25). Boston: Birkhauser.
Hillyard, S. A., & Anllo-Vento, L. (1998). Event-related brain potentialsin the study of visual selective attention. Proceedings of the NationalAcademy of Sciences, USA, 95, 781–787.
Hillyard, S. A., & Kutas, M. (1983). Electrophysiology of cognitiveprocessing. Annual Review of Psychology, 34, 33–61.
Hopf, J. M., &Mangun, G. R. (2000). Shifting visual attention in space:An electrophysiological analysis using high spatial resolution map-ping. Clinical Neurophysiology, 111, 1241–1257.
Hopfinger, J. B., Camblin, C. C., & Parks, E. L. (2010). Isolating theinternal in endogenous attention. Psychophysiology (in press).
James, W. (1950). Principles of psychology (vol. 1). New York: DoverPublications. (Originally published in 1890).
Jensen, O., Kaiser, J., & Lachaux, J.-P. (2007). Human gamma-frequency oscillations associated with attention and memory. Trendsin Neuroscience, 30, 317–324.
Jervis, B. W., Ifeachor, E. C., & Allen, E. M. (1988). The removal ofocular artifacts from the electroencephalogram: A review.Medical &Biological Engineering & Computing, 26, 2–12.
Jongen, E., Smulders, F., & Van Der Heiden, J. (2007). Lateralized ERPcomponents related to spatial orienting: Discriminating the directionof attention from processing sensory aspects of the cue. Psychophys-iology, 44, 968–986.
Jongen, E. M., Smulders, F. T., & van Breukelen, G. J. (2006). Varietiesof attention in neutral trials: Linking RT to ERPs and EEG frequen-cies. Psychophysiology, 43, 113–125.
Jonides, J. (1980). Towards a model of the mind’s eye’s movement. Ca-nadian Journal of Psychology, 34, 103–112.
Kastner, S., & Ungerleider, L. (2000). Mechanisms of visual attention inthe human cortex. Annual Review of Neuroscience, 23, 315–341.
Keil, A., Mussweller, T., & Epstude, K. (2006). Alpha-band activityreflects reduction of mental effort in a comparison task: A sourcespace analysis. Brain Research, 1121, 117–127.
Kelly, S. O., Lalor, E. C., Reilly, R. B., & Foxe, J. J. (2006). Increases inalpha oscillatory power reflect an active retinotopic mechanism fordistracter suppression during sustained visuospatial attention. Journalof Neurophysiology, 95, 3844–3851.
Attention shifts and parietal alpha band activity 321

Kiss, M., van Velzen, J., & Eimer, M. (2008). The N2pc component andits links to attention shifts and spatially selective visual processing.Psychophysiology, 45, 240–249.
Klimesch,W. (1999). EEG alpha and theta oscillations reflect cognitive andmemory performance: A review and analysis.Brain Research, 29, 169–195.
Luck, S. J., Heinze, H. J., Mangun, G. R., & Hillyard, S. A. (1990).Visual event-related potentials index focused attention within bilat-eral stimulus arrays. II. Functional dissociation of P1 and N1 com-ponents. Electroencephalography and Clinical Neurophysiology, 75,528–542.
Luck, S. J., Hillyard, S. A.,Mouloua,M.,Woldorff,M.G., Clark, V. O.,& Hawkins, H. L. (1994). Effects of spatial cuing on luminance de-tectability: Psychophysical and electrophysiological evidence for earlyselection. Journal of Experimental Psychology, 20, 887–904.
Luck, S. J., Woodman, G. F., & Vogel, E. K. (2000). Event-relatedpotential studies of attention. Trends in Cognitive Science, 4, 432–440.
Malinowski, O., Fuchs, S., &Muller,M.M. (2007). Sustained division ofspatial attention to multiple locations within one hemifield. Neu-roscience Letters, 414, 65–70.
McDonald, J., & Green, J. (2008). Isolating event-related potentialcomponents associated with voluntary control of visuo-spatial atten-tion. Brain Research, 1227, 96–109.
Miniussi, C., Rao, A., & Nobre, A. C. (2002). Watching where you look:Modulation of visual processing of foveal stimuli by spatial attention.Neuropsychologia, 40, 2448–2460.
Muller, M. M., Malinowski, O., Gruber, T., & Hillyard, S. A.(2003). Sustained division of the attentional spotlight. Nature, 424,309–312.
Nakamura, K., & Colby, C. L. (2000). Visual, saccade-related, and cog-nitive activation of single neurons in monkey extrastriate area V3A.Journal of Neurophysiology, 84, 677–692.
Neville, H. J., & Lawson, D. (1987). Attention to central and peripheralvisual space in a movement detection task: An event-related potentialand behavioral study. I. Normal hearing adults. Brain Research, 405,253–267.
Nobre, A. C., Sebestyen, G. N., & Miniussi, C. (2000). The dynamics ofshifting visuospatial attention revealed by event-related potentials.Neuropsychologia, 38, 964–974.
Nunez, P. L., Wingeier, B. M., & Silberstein, R. B. (2001). Spatial-temporal structures of human alpha rhythms: Theory, microcurrentsources, multiscale measurements, and global binding of local net-works. Human Brain Mapping, 13, 125–164.
Palva, S., & Palva, J. M. (2007). New vistas for alpha-frequency bandoscillations. Trends in Neuroscience, 30, 150–158.
Pfurtscheller, G. (2001). Functional brain imaging based on ERD/ERS.Vision Research, 41, 1257–1260.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Ex-perimental Psychology, 32, 3–25.
Ray,W. J., & Cole, H.W. (1985). EEG alpha activity reflects attentionaldemands, and beta activity reflects emotional and cognitive processes.Science, 228, 750–752.
Raz, A. (2004). Anatomy of attentional networks. Anatomical Record,281, 21–36.
Rihs, T., Michel, C., & Thut, G. (2007). Mechanisms of selective inhi-bition in visual spatial attention are indexed by alpha-band EEGsynchronization. European Journal of Neuroscience, 25, 603–610.
Rihs, T., Michel, C., & Thut, G. (2009). A bias for posterior a-bandpower suppression versus enhancement during shifting versus main-tenance of spatial attention. NeuroImage, 44, 190–199.
Rizzolatti, G., Riggio, L., Dascola, I., & Umilta, C. (1987). Reorientingattention across the horizontal and vertical meridians: Evidence infavor of a premotor theory of attention.Neuropsychologia, 25, 31–40.
Sauseng, O., Klimesch, W., Stadler, W., Schabus, M., Doppelmayr, M.,Hanslmayr, S., et al. (2005). A shift of visual spatial attention isselectively associated with human EEG alpha activity. EuropeanJournal of Neuroscience, 22, 2917–2926.
Sieroka, N., Dosch, H. G., Specht, H. J., & Rupp, A. (2003). Additionalneuromagnetic source activity outside the auditory cortex in durationdiscrimination correlates with behavioural ability. NeuroImage, 20,1697–1703.
Slagter, H. A., Kok, A., Mol, N., & Kenemans, J. L. (2005). Spatio-temporal dynamics of top-down control: Directing attention to lo-cation and/or color as revealed by ERPs and source modeling. BrainResearch, 22, 333–348.
Slagter, H. A., Kok, A., Mol, N., Talsma, D., & Kenemans, J. L. (2005).Generating spatial and nonspatial attentional control: An ERP study.Psychophysiology, 42, 428–439.
Small, D. M., Gitelman, D. R., Gregory, M. D., Nobre, A. C., Parrish,T. B., &Mesulam, M. M. (2003). The posterior cingulate and medialprefrontal cortex mediate the anticipatory allocation of spatial atten-tion. NeuroImage, 18, 633–641.
Stanislaw, H., & Todorov, N. (1999). Calculation of signal detectiontheory measures. Behavior Research Methods, Instruments, & Com-puters, 31, 137–149.
Suffczynski, P., Kalitzin, S., Pfurtscheller, G., & Lopes da Silva, F.(2001). Computational model of thalamo-cortical networks: Dynam-ical control of alpha rhythms in relation to focal attention. Interna-tional Journal of Psychophysiology, 43, 25–40.
Tallon-Baudry, C., & Bertrand, O. (1999). Oscillatory gamma activity inhumans and its role in object representation. Trends in Cognitive Sci-ence, 3, 151–162.
Talsma, D., Slagter, H. A., Nieuwenhuis, S., Hage, J., & Kok, A. (2005).The orienting of visuospatial attention: An event-related brain po-tential study. Brain Research, 25, 117–129.
Tassinari, G., Aglioti, S., Chelazzi, L., Marzi, C. A., & Berlucchi, G.(1987). Distribution in the visual field of the costs of voluntarily al-located attention and of the inhibitory after-effects of covert orient-ing. Neuropsychologia, 25, 55–71.
Thut, G., Nietzel, A., Brandt, S. A., & Pascual-Leone, A. (2006). Alpha-band electroencephalographic activity over occipital cortex indexesvisuospatial attention bias and predicts visual target detection. Jour-nal of Neuroscience, 26, 9494–9502.
van Velzen, J., & Eimer, M. (2003). Early posterior ERP components donot reflect the control of attentional shifts toward expected peripheralevents. Psychophysiology, 40, 827–831.
Walter, W. G., Cooper, R., Aldridge, V. J., McCallum,W. C., &Winter,A. L. (1964). Contingent negative variation: An electric sign of sen-sorimotor association and expectancy in the human brain. Nature,203, 380–384.
Weiskrantz, L., Warrington, E. K., Sanders, M. D., & Marshall, J.(1974). Visual capacity in the hemianopic field following a restrictedoccipital ablation. Brain, 97, 709–728.
Worden, M. S., Foxe, J. J., Wang, N., & Simpson, G. V. (2000). An-ticipatory biasing of visuospatial attention indexed by retinotopicallyspecific alpha-band electroencephalography increases over occipitalcortex. Journal of Neuroscience, 20, RC63.
Wright, M. J., Geffen, G. M., & Geffen, L. B. (1995). Event relatedpotentials during covert orientation of visual attention: Effects of cuevalidity and directionality. Biological Psychology, 41, 183–202.
Wright, R. D., Richard, C. M., & McDonald, J. J. (1995). Neutral lo-cation cues and cost/benefit analysis of visual attention shifts. Cana-dian Journal of Experimental Psychology, 49, 540–548.
Yamagishi, N., Callan, D. E., Anderson, S. J., & Kawato, M. (2008).Attentional changes in pre-stimulus oscillatory activity within earlyvisual cortex are predictive of human visual performance. BrainResearch, 1197, 115–122.
Yantis, S., & Jonides, J. (1990). Abrupt visual onsets and selective at-tention: Voluntary versus automatic allocation. Journal of Experi-mental Psychology, 16, 121–134.
SUPPORTING INFORMATIONAdditional Supporting Information may be found in the onlineversion of this article:
Table S1: Behavioral measures (Reaction Times, Hit Ratesand False Alarms) according to Attentional Condition andDifficulty level.
Table S2: Alpha Band Power, Easy vs Difficult comparisons,Ipsilateral-to-shift hemisphere parietal electrodes, Latencies 2–3.
Figure S1: Illustration of individual alpha peak differences.
Please note: Wiley-Blackwell are not responsible for the con-tent or functionality of any supporting materials supplied by theauthors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
(Received October 30, 2009; Accepted March 31, 2010)
322 D. Cosmelli et al.
Attention shifts and parietal alpha band activity
1
Supplementary Material
Separate analysis of difficult versus easy blocks was used to rule out modulations of alpha band
activity due to non-specific changes (such as changes in alertness or effort) between central no-
shift conditions and the more demanding lateral shifts. We first present the behavioral results for
the Difficult and Easy blocks separately and then the corresponding analysis of the alpha band
power.
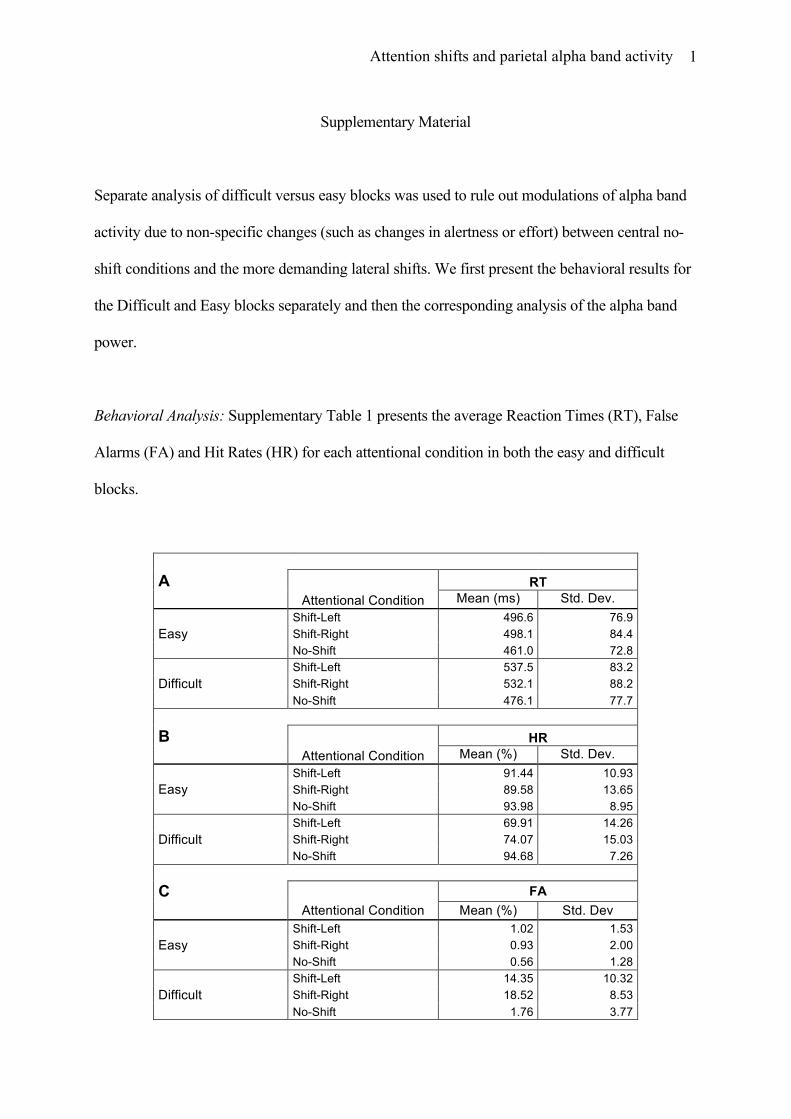
Behavioral Analysis: Supplementary Table 1 presents the average Reaction Times (RT), False
Alarms (FA) and Hit Rates (HR) for each attentional condition in both the easy and difficult
blocks.
A RT Attentional Condition Mean (ms) Std. Dev.
Easy Shift-Left 496.6 76.9 Shift-Right 498.1 84.4 No-Shift 461.0 72.8
Difficult Shift-Left 537.5 83.2 Shift-Right 532.1 88.2 No-Shift 476.1 77.7
B HR Attentional Condition Mean (%) Std. Dev.
Easy Shift-Left 91.44 10.93 Shift-Right 89.58 13.65 No-Shift 93.98 8.95
Difficult Shift-Left 69.91 14.26 Shift-Right 74.07 15.03 No-Shift 94.68 7.26
C FA Attentional Condition Mean (%) Std. Dev
Easy Shift-Left 1.02 1.53 Shift-Right 0.93 2.00 No-Shift 0.56 1.28
Difficult Shift-Left 14.35 10.32 Shift-Right 18.52 8.53 No-Shift 1.76 3.77

Attention shifts and parietal alpha band activity
2
Supplementary Table 1: Behavioral measures (Reaction Times, Hit Rates and False Alarms) according to Attentional Condition and Difficulty level.
Statistical analysis of RT, HR and FA was performed using a repeated measure ANOVA design
with factors Difficulty (2 levels: Easy/Difficult) and Cue Orientation (3 levels: Shift-Left, No-
Shift and Shift-Right).
As expected, we observed a main effect of difficulty for RT, F(1, 17)=17.54, p<0.01.
Main effect of cue orientation was also significant, F(2, 34)=41.73, p<0.01, due to systematic
shorter RT for the No-shift condition. Interaction between these two factors did not reach
significance, F(2, 34)=2.02, p=0.15. Post-hoc Bonferroni-corrected comparisons showed no
differences in RT between Shift-Left and Shift-Right within difficulty levels. However, as
expected, Shift-Left (p < 0.05, df: 34) and Shift-Right (p < 0.05, df: 34) RTs where shorter in the
Easy compared to the Difficult condition. In contrast, no significant differences were observed in
RT during the No-Shift between difficulty levels. Finally, RTs for Shift-Left and Shift-Right
were significantly longer than in the No-Shift condition, when compared within difficulty levels
(p<0.05; all df: 34).
As with the RT measure, a main effect of difficulty was significant for HR, F(1,
17)=18.48, p<0.01. Main effect of cue orientation was also significant, F(2, 34)=44.85, p<0.01,
here again due to much higher HR in the central position regardless of the difficulty level. The
Difficulty x Cue Orientation interaction was significant for this measure, F(2, 34)=33.09, p<0.01,
due to the fact that HR to central targets did not exhibit significant changes between difficulty
levels, while HR to lateral targets did. Post-hoc Bonferroni-corrected comparisons revealed no
differences in HR between Shift-Left and Shift-Right within Easy or Difficult levels, although
HR where significantly lower for the Difficult condition compared to the Easy condition in both

Attention shifts and parietal alpha band activity
3
lateral Shifts (p<0.05, df: 34). As for the RT measure, no significant differences where found in
HR between difficulty levels for the No-Shift condition.
Main effect of difficulty for FA was similar to the other behavioral measures, F(1,
17)=18.48, p<0.01, as well as for the main effect of cue orientation, F(2, 34)=48.27, p<0.01.
There was a significant Difficulty x Cue Orientation interaction, F(2, 34)=41.69, p<0.01: here
again the No-Shift task did not show significant changes between difficulty levels in contrast to
lateral shifts. Interestingly, Bonferroni-corrected comparisons revealed a left/right asymmetry for
this measure: FA after Shift-Left cues where significantly lower when compared to Shift-Right,
yet this effect was only evident in the difficult level (p<0.05, df: 34). No differences were found
in the Easy condition regardless of the attended visual field.
To summarize, easy and difficult blocks where distinguishable in terms of all three
behavioral measures, yet due mainly to lateral shifts showing significant differences between
easy and difficult tasks. When attention was maintained at fixation, the more difficult task
produced only a slight increase in RT relative to the easy task (see supplementary Table 1) that
did not reach significance.
Alpha Band Analysis: Because lateral shifts showed sensitivity to changes in task difficulty,
differences in alpha band during lateral shifts across difficulty levels, if present, should be
indicative of the degree of influence effort has on our results regarding specifically the covert
orienting process. We therefore studied the specific behavior of alpha power over
contralateral and ipsilateral parietal sites during the cue-distracter interval for the three cuing
conditions and the two difficulty levels.
Alpha band power over the parietal electrodes during the cue-distracter interval was
analyzed with a similar repeated measures ANOVA design to the one used in the main text,
but considering Difficulty as an additional factor (2 levels: Easy/Difficult). We found no main

Attention shifts and parietal alpha band activity
4
effects of difficulty (Alpha Lower: F(1, 17) = 0.0147, p = 0.905; Alpha Upper: F(1, 17) =
0.0085, p = 0.927). Neither the two-way Difficulty x Cue interaction (Alpha Lower: F(2, 34)
= 0.6412, p = 0.533; Alpha Upper: F(2, 34) = 1.844, p = 0.174) or the two-way Difficulty x
Hemisphere interaction (Alpha Lower: F(1, 17) = 0.001, p = 0.975; Alpha Upper: F(1, 17) =
0.1385, p = 0.714) were apparent. There was no significant three-way Difficulty x Cue x
Hemiphere interaction either (Alpha Lower: F(2, 34) = 0.0607, p = 0.941; Alpha Upper: F(2,
34) = 0.1132, p = 0.893).
In order to further scrutinize the data we performed a series of univariate planned
comparisons between difficulty levels for left and right shifts. To maximize sensitivity we
used the ipsilateral-to-shift electrode over latencies 2 and 3 in the cue-distracter interval
(where the main changes related to attentional orienting where apparent). No significant
differences where found for any attentional condition between difficulty levels for either
Alpha sub-bands as can be seen in Table 2 of the Supplementary Material:
Sub-band F p Alpha Lower 1.09374 0.3103 0.44687 0.5128 Alpha Upper 1.14245 0.3001 3.40786 0.0824
Supplementary Table 2: Alpha Band Power, Easy vs Difficult comparisons, Ipsilateral-to-shift hemisphere parietal electrodes, Latencies 2-3
These analyses show that despite behavioral differences between difficulty levels during
covert shifts, these do not appear related to significant changes in alpha band activity. This
strongly suggests that unspecific effects such as effort or alertness cannot account for the
observed results in alpha band dynamics. In view of this and in order to maintain more
adequate number of trials, we have chosen to collapse across difficulty levels for all analyses
presented in the main text.

Attention shifts and parietal alpha band activity
5
Supplementary Figure Captions
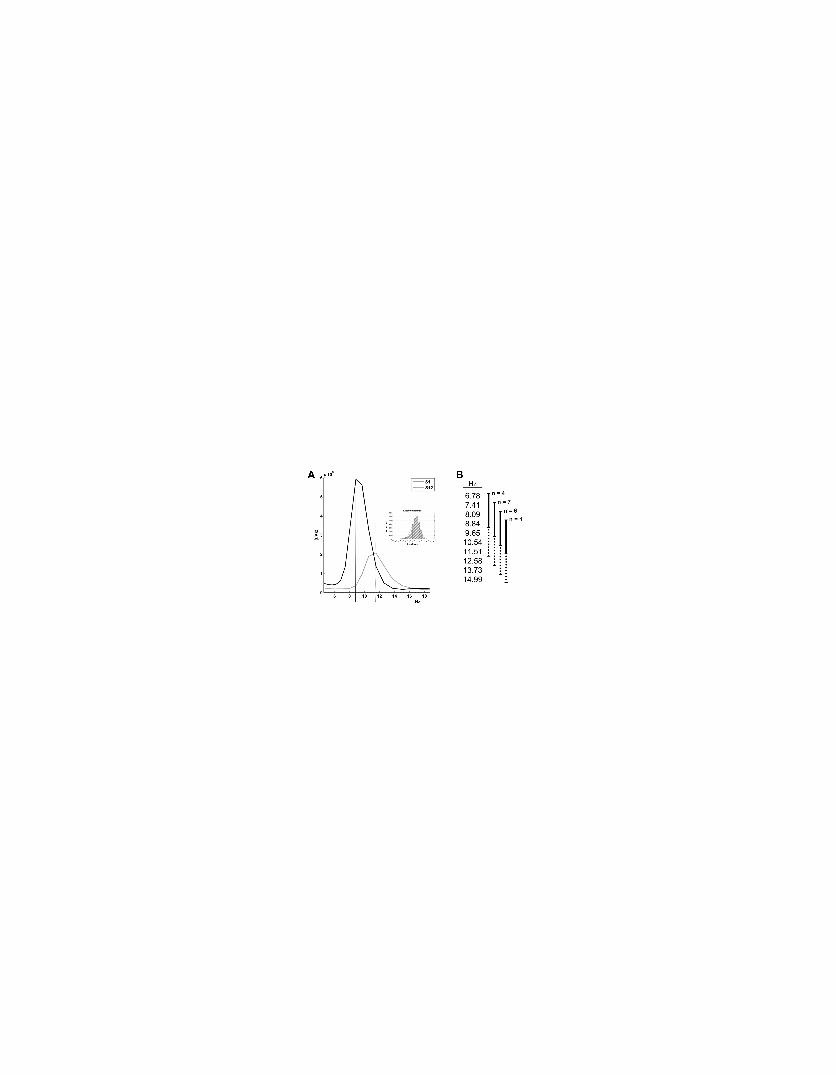
Supplementary Figure 1: Illustration of individual alpha peak differences. A. The main figure
presents a raw average time-frequency chart collapsed across time for a parieto-central electrode
(CPz) in the 4-18Hz range, for two subjects. Continuous and dotted vertical lines indicate peak
values for two subjects respectively (Subject 1 and Subject 12). Inset shows distribution of
overall alpha power log-transformed and baseline corrected and normal distribution overlay in
red continuous line justifying the use of a General Linear Model for the statistical analysis of this
dataset (Kiebel, Tallon-Baudry & Friston., 2005). B. The diagram indicates the four analysis
groups adjusted according to individual alpha peaks with the corresponding frequency bins.
Continuous/dotted lines indicate lower/upper-alpha ranges.
Copyright © 2022 FDOKUMEN




















