
Soil organic carbon sequestration potential of cropland in China
Upload
independentCategory
view
7download
0
• .' JOllrtla! ofChemical Ec% gy . vor 2/. No, /2. /995
SEQUESTRATION OF LICHEN COMPOUNDS BY LICHEN-FEEDING MEMBERS OF THE ARCTIIDAE
(LEPIDOPTERA)
SONJA HESBACHER, I INGRID GIEZ, I GERNOT EMBACHER,2 KONRAD FIEDLER,) WALDEMAR MAX,4 ALOIS TRAWÖGER,5
ROMAN TÜRK,6 OTTO L. LANGE, land PETER PROKSCH 1• *
'Julius-von-Sachs-lnstitUl für BiowisseTlschaften Universität Würzburg
MillLerer DaUenbergweg 64 D-97082 Würzburg, Germany
2Anton Brucknerstr, 3 A-5020 Salzburg , Austria
J LehrstuhL für Zoologie JI Theodor-Boveri-lnstitut für Biowisserlschaften
Am Hubland D-97074 Würzburg, Germany
'Am Stadtpark 18 D-38642 Gaslar, Germany
s Lindenstr. 9 A-6020 Innsbruck, Austria
61nstitut für PflanzenphysioLogie Universität Salzburg Hel/brunnerstr. 34
A-5020 Salzburg, Austria
(Received January 31 , 1995; accepted August 9, 1995)
Abstract-A survey for the presence of sequestered lichen compounds in 103 wild-caughl imagines represenling eight different genera and 16 different speeies of the Arctiidae was conducted. Known lichen compounds were detected for the first time in 24 of the analyzed specimens (representing live different
genera and ll different species) based on their HPLC retention times and on their UV -absorplion spectra, The anthraquinone parietin, the depside atranorin, as weil as a hydrolytie c1eavage product of the latter were among the lichen compounds most frequently detected in wild-caught imagines, The observed variation of sequestered lichen compounds in wild-caught imagines
*To whom correspondence should be addressed.
2079
-/1
009R-0331 /95/1200-2079$07 .50iO © 1995 Plenum Publlshing Corporation
•

• 2080 HESBACHER ET AL
with unknown feeding history may be due to several reasons . Lack of lichen compounds in imagines may have been caused. for example. by larvae feeding on lichens with no or only minute amounts of phenolic products. The age of the specimens analyzed mayaiso inlluence the results obtained. Avoidance of lichen compounds by selective feeding on those parts of lichen thalli that have no or little lichen products may be another reason for the lack of lichen compounds in imagines. Preliminary feeding experiments conducted with lar
vae of Ei/ema comp/ana . for example, indicated that the larvae fed exclusively on the algal layer and cortex of the lichen Cladonia pyxida(a, whereas the medulla. which is rich in fumarprotocctraric acid . was avoided. As expected. imagines hatching from the larvae were free 01' this lichen compound . Any ecological role of the sequestered lichen compounds for the herbivores is unknown. It is possible. however, that sequestered lichen compounds may be utilized for the chemical defense of arctiid moths or against microbial path
ogens.
Key Words-Arctiidae , Lepidoptera, lichens, lichen compounds, HPLC, sequestration, chemical defense.
INTRODUCTION
Lichens comprise some 20,000 different species and show a worldwide distribution, commonly growing on rocks and poorly developed soils such as those of arid lands and boreal-arctic regions, or as epiphytes on trees and shrubs (Crittenden and Porter, 1991). Especially in habitats that are poor in nutrients and characterized by extreme environmental conditions (such as hot or cold deserts), lichens may fonn the dominant floral elements and may thus provide important potential resources for herbivores. The large concentrations of mostly phenolic secondary compounds-the so-called lichen compounds-that are accumulated in copious amounts (sometimes exceeding 20% of the dry weight) by many lichens have long been suspected to protect these symbiotic organisms from general ist herbivores (Zukai, 1895; Stahl, 1904). Recent experiments with polyphagous larvae of the insect Spodoptera littoralis (Noctuidae) demonstrated pronounced acute toxicity and feeding deterrency for frequently occurring lichen compounds such as (+)- and (- )-usnic acid or vulpinic acid at concentrations comparable to or even below those found in many lichens (Emmerich et al., 1993). Other lichen compounds, such as oxyphysodic acid or fumarprotocetraric acid, had no effect on larval survival, but prolonged the larval period of S. littoralis and caused malfonnations of imagines when added to the larval diet (Giez et al., 1994).
In spite of the deterrent or even toxic effects of many lichen products toward generalist herbivores, there are examples of specialized lichen feeders, including oribatid mites (Seyd and Seaward, 1984; Reutimann and Scheidegger, 1987), terrestrial gastropods (Baur et al., 1992; Hesbacher et al. , 1995) and Lepidop-

• SEQUESTRATION OF LI CHEN COMPOUNDS 2081
tera, forexample , from the family Arctiidae (Rambold, 1985; Embacher. 1994; Trawöger, 1991). We are interested in the mechanisms that aIlow this kind of specialization and especially in the face of lichen compounds in these specialized lichen-insect associations. Recently , we have demonstrated for the first time that the lichen-feeding snails, Ba/ea perversa and Chondrina clienta, sequester lichen compounds in their soft bodies (Hesbacher et al., 1995). In the present study we report for the first time on the occurrence of lichen compounds in wild-caught imagines of the Arctiidae that are known to feed on lichens in nature (Rambold, 1985; Embacher, 1994; Trawöger, 1991), as weIl as in insects that were raised in the laboratory on a known lichen diet.
METHODS AND MATERIALS
Wild-Caught lnsects. Dates and localities for c01lection of wild-caught imagines in which lichen compounds could be identified are given in Table 2 below. FoIlowing air or freeze drying, a1l coIlected specimens were individually ground in a mortar and exhaustively extracted with acetone . The resulting extracts were subsequently reduced to a known volume and stored in a freezer until HPLC analysis.
Feeding E.xperiments with Eilemu complana. Larvae of Ei/ema complalla collected near Stallhof (Niederösterreich) were reared for the last five days of their larval period in the laboratory on the lichen Cladonia pyxidata (L.) Hoffrn., containing fumarprotocetraric acid (7, Figure 1) as the major lichen compound. During these five days, larval feces were collected daily for HPLC analysis. The imagines emerged from the pupae after 12-14 days in darkness at 25 °C. Feces, the lichen, and freeze-dried imagines were extracted in a known amount ofacetone at room temperature (20-22°C) . The extracts were stored in a freezer until HPLC analysis.
Loca/ization 0/ Fumarprotocetraric Acid (7) in the Lichen Cladonia pyxidata. Thallus pieces of the lichen Cladonia pyxidata (L) Hoffm. including basal squamules of the primary thallus as weIl as segments of podetia were soaked in H20 followed by a treatment (10 min) with 1.8% HCI to dissolve oxalic acid present in the thallus. Air bubbles were removed from the thallus under vacuum. Subsequently, transversal sections (25 /Lm) were cut with a freezing microtome (Kryomat, Leitz; -25°C). The sections obtained were analyzed microscopically using polarized light.
HPLC Analysis. The HPLC system (Pharmacia) was equipped with a photodiode array detector (Waters). SampIes were injected on a Nova-Pak C I8
column (Waters) (150 x 3.9 mm, 4-/Lm pore size) . Separation of the compounds took place by using a linear gradient from 100% A (10% MeOH , 90% H20 adjusted to pH 2 with o-phosphoric acid) to 100% B (MeOH) in 40 min. Iden-
-
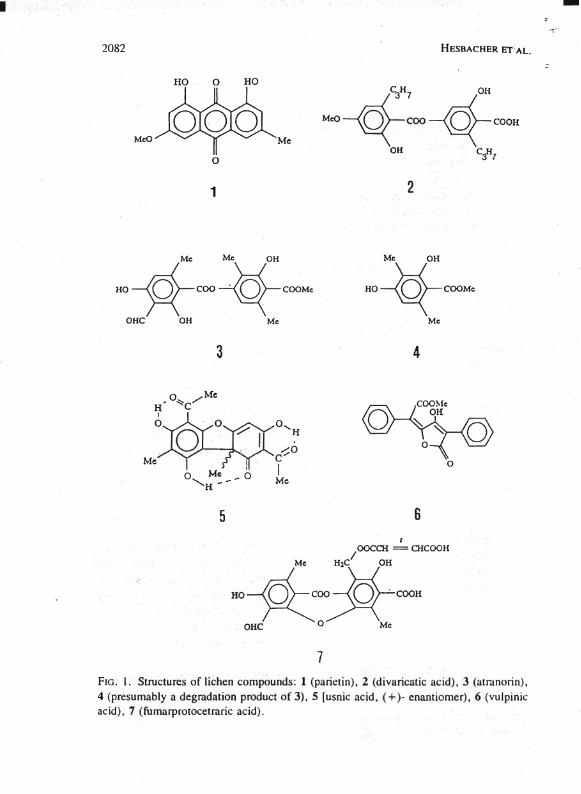
• 2082
HO o
McO
o
1
3
5
HO
Me
0, H
~O c I Me
HESBACHER ET 'AL.
2
4
o
6 I
/OOCCH = CHCOOH
Ho~)fCOOH OHC 0 Me
7 FIO . I. Structures of lichen compounds: 1 (panetin), 2 (divancatic acid), 3 (atranonn), 4 (presumably adegradation product of 3), 5 [usnic acid, (+)- enantiomer), 6 (vulpinic acid), 7 (fumarprotocetranc acid).
-

• SEQUESTRATION OF LICHEN COMPOUNDS 2083
titieation of liehen eompounds was by eomparison of retention times and by eomparison of the online-reeorded UV absorption speetra with those of eommereially available liehen substanees (Roth) or with liehen eompounds isolated and identified previously in our laboratory . Liehen eompounds were quantified by the external standard method using eommereially available liehen produets (Roth).
RESULTS
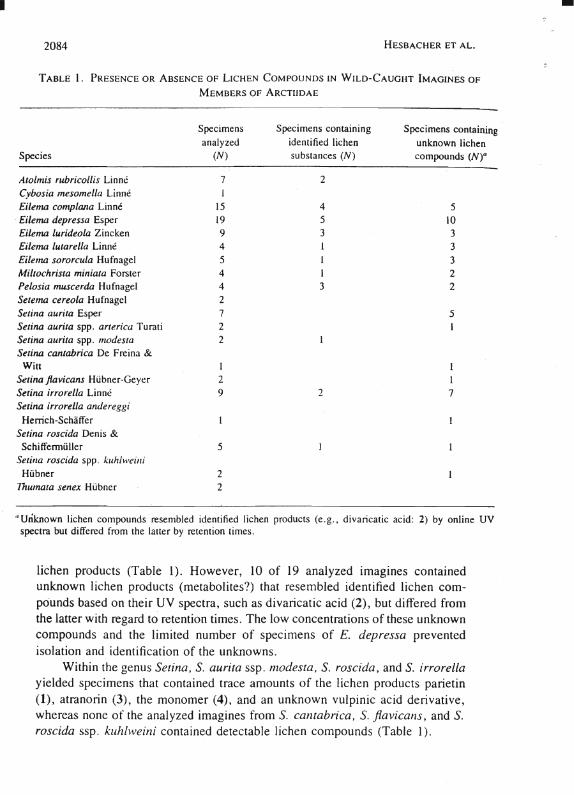
Table 1 summarizes the data obtained in our survey for liehen eompounds in 103 wild-eaught imagines representing eight different genera and 16 different speeies of the Aretiidae. Liehen eompounds 1-6 (Figure I) were readily identified by HPLC based on their retention times and online UV absorption speetra as weil as by comparison with authentie standards. The anthraquinone parietin (1), the depside atranorin (3), as weil as the monomer (4), whieh presumably originates from atranorin by hydrolytie cleavage, were among the major lichen eompounds deteeted in the inseets (Table 2). Other lichen eompounds that were sporadieally found included usnic aeid (5) [differentiation between (+)- or (-)usnie aeid was not possible due to the small amountsl as weil as divarieatic acid (2). In addition to compounds 1-6, several eonstituents were deteeted in the imagines that resembled liehen produets (e.g., usnie or vulpinie aeid) based on their eharaeteristie UV absorption spectra but differed with regard to their retention times.
Considerable inter- as weil as intraspeeific variation with regard to the nature of the sequestered liehen eompounds and to their amounts (mierograms per imago) was observed. Relatively large amounts of parietin (1) were found in one imago of Atolmis rubricollis (5.64 jLg/imago), along with atranorin (3) (0.23 jLg/imago), and the monomer (4) (0.09/jLg/imago), whereas a seeond speeimen of A. rubricollis yielded only parietin (1) (1.85 JLg/imago). Five additional imagines of the same speeies were found to be devoid of any detectable liehen eompounds (Table 1).
Analysis of 52 speeimens from the genus Eilema showed also a high variability with regard to sequestered lichen eompounds. Even though each of the six analyzed speeies of this genus was represented by one to several speeimens in whieh liehen eompounds could be unequivoeally identified, a comparatively large number was devoid of deteetable lichen products. For example, of 19 wild-caught imagines of E. depressa (syn. E. deplana), only five specimens were found to contain the liehen compounds parietin (1) (trace-5.43 JLg/imago), divaricatic acid (2) (trace) as weil as the monomer (4) and usnic aeid (5) (both also in traee amounts), whereas the remaining 14 spccimcns were free of tho~e
-
• 2084 HESBACHER ET AL.
T ABLE 1. PRESENCE OR ABSENCE OF LICHEN COMPOUNDS IN WILD-CAUGHT IMAGINES OF
MEMBERS OF ARCTllDAE
Species
AlOlmis rubricollis Linn': Cybosia mesomella Linn': Eilema complana Linn': EilerruJ depressa Esper EiierruJ lurideola Zincken EiierruJ lurarella Linne EiierruJ sororcula Hufnagel MillOchrisra miniara Forster Pelosia muscerda Hufnagel Serema cereola Hufnagel Serina aurira Esper Serina aurira spp. arrerica Turati Serina aurira spp. modesta Serina canrabrica Oe Freina &
Win Serina ftavicans Hübner-Geyer Serina irrarella Linne Serina irrarella andereggi
Herrich-Schätfer Serina roscida Oenis & Schitfermüller
Serina roscida spp. kuh/weini Hübner
7humara senex Hübner
Specimens analyzed
(N)
7 I
15 19 9 4 5 4 4 2 7 2 2
I 2 9
5
2 2
Specimens containing
identified lichen substances (N)
2
4 5 3
3
2
Specimens containing
unknown lichen compounds (N)"
5 10 3 3 3 2 2
5
7
" uriknown lichen compounds resembled identified lichen products (e.g .• divaricatic acid: 2) by online UV spectra but ditfered from the latter by retention times.
lichen products (Table I). However, 10 of 19 analyzed imagines contained unknown lichen products (metabolites?) that resembled identified lichen compoünds based on their UV spectra, such as divaricatic acid (2), but differed from the latter with regard to retention limes . The low concentrations of these unknown compounds and the Iimited number of specimens of E. depressa prevented isolation and identification of the unknowns.
Within the genus Setina. S. aurita ssp. modesta. S. roscida, and S. irrorella yielded specimens that contained trace amounts of the lichen products parietin (1), atranorin (3), the monomer (4), and an unknown vulpinic acid derivative, whereas none of the analyzed imagines from S. cantabrica. S. jiavicans, and S. roscida ssp. kuhlweini contained detectable lichen compounds (Table I).
•

•
TABLE 2. DISTRIBUTION AND AMOUNTS OF LICHEN COMPOUNDS DETECTED IN WILD-CAUGHT SPECIES 01' ARCTIIDAE"
'" m Lichen eompounds identified ÜLg/imago)'" I:)
c Dry wt. m
'" Speeies Speeimen (mg) Year and loeality of eollection" 2 3 4 5 6 -I
'" > I (E)"
-I Alo/mis rubricollis 15.7 1983 Salzburger Land (Austria) 5.64 0.23 0.09 Ö
2 (M) 15.4 1993 Harz (FRG) 1.85 z 0
Ei/ema comp/alJa I (E) 8.3 ? Salzburger Land (Austria) + "Tl
r 2 (F) 10.6 1984 Spessart (Germany) + i) 3 (M) 9.4 1993 Harz (Gemlany) + :t
m 4 (M) 8.9 1993 Harz (Germany) + z
n Ei/ema depressli I (E) 11.1 ? Salzburger Land (Austria) + 0
::: 2 (E) 9.8 1993 Salzburger Land (Austria) 5.43 + "0
0 3 (F) 12.6 1984 Spessan (Gennany) 0.99 c
z 4 (F) 9.1 1984 Spcssan (Gcrmany) + Cl
'" 5 (M) 11.0 1967 Harz (FRG) + Ei/ema /urideo/a I (E) 9.1 1993 Salzburger Land (Austria) 1.18 +
2 (M) 8.6 1993 Harz (Gennany) + 3 (M) 6.6 1959 Harz (Germany) 0.093
Ei/ema !tllarellll I (M) 4.6 1977 Harz (Germany) + Ei/ema .wrorcu/a I (E) IO.M ? Salzburger Land (Austria) + Mi/rvchrisla milliara I (E) 7.6 1984 Salzburger Land (Auslria) + Pe/osia muscerda I (E) 7.7 1993 Salzburger Land (Austria) +
2 (E) 8.2 1993 Salzburger Land (Austria) + + 3 (E) 7.4 1993 Salzburger Land (Austria) +
Serina aurilll ssp. I (S) 13.6 1973 Tessin (Switzerland) + II/odesw
.)clillil irrorcll" I (E) 7.H 1')1)1 Sall.hllrgcr I.alld (Allstria) ... 2 (10) 6.2 ? Salzburger L:lI1d (Austria) +
Selill<l roscida I (E) 4.6 1993 Salzburger Land (Austria) +
nol dClcctcd , +: prcscnt (trac.: amounts). N 0 00
I, Details "I' the lucalitics ur collcctiun ean be obtaincd from the authors. Ul
, Numbers of compounds as in Figure I. "Sourees: E = G. Embaeher, F = K. Fiedler, M = W. Max, S = M. Sleincit.
-

• 2086 HESBACHER ET AL.
Controlled feeding experiments were conducted with larvae of Eilema com-plana. Due to the small sampIe size (only two larvae were available), however, the experiments can only be regarded as preliminary in nature . Larvae of E. complana (second and third instars) were observed feeding on the lichen Cladonia pyxidata. When transfered from the natural habitat to the laboratory , the larvae were found to feed exclusively on the cortical and algal layers of the lichen thalli but avoided the medulla, as evidenced by the whitish appearence of the remaining parts of the thallus after the cortex and green algal layer had been completely removed by the larvae. Light microscopic analysis of transverse sections of the lichen thallus indicated that the lichen compound fumarprotocetraric acid (7) was confined exclusively to the medulla but appeared to be either lacking or negligible in the algal layer or in the cortex. Larval feces that were collected daily as weil as the imaginesthat developed from the larvae were devoid of fumarprotocetraric acid (7) (Figure 2) . Hence, the larvae avoided uptake of the lichen compound by selective feeding on those parts of the thalli that were essentially free of fumarprotocetraric acid (7).
DISCUSSION
Sequestration of secondary plant constituents such as alkaloids (Ehmke et al., 1990), ftavonoids (Wiesen et al., 1994), or cardiac glycosides (Malcolm, 1990) by herbivorous insects has frequently been reported for interactions of herbivores with higher plants. Species of the Arctiidae, for example , are known to sequester pyrrolizidine alkaloids from plants of the Asteraceae, Fabaceae, or Boraginacae and to utilize these compounds either for their own chemical defense (Hartmann, 1991) or as precursors for pheromones (Boppre and Schneider, 1985). The present srudy provides, for the first time, evidence for sequestration of lichen compounds by Iichen-feeding members of the Arctiidae. Of 103 specimens, 64 specimens were found to contain identified lichen compounds or metabolites resembling identified lichen products by their online UV spectra (Table I). However, when comparing the number of species with sequestered lichen compounds to the number of species lacking detectable lichen compounds, the different sam pIe sizes (number of analyzed specimens per species) of both groups have to be taken into account. Species of the Arctiidae that lacked identified lichen compounds, such as Setina cantabrica or Setema cereola, were represented by only one or two specimens compared, for example, to the 19 specimens of E. depressa and the 15 specimens for E. complana (including five and four specimens, respectively, that contained lichen products). Considering the pronounced intraspecific variation with regard to presence vs. absence of lichen compounds observed in this study, sequestration of lichen compounds may therefore be more widespread within lichen-feeding members of the Arctiidae than suggested by the results of Table I .
•

• SEQUESTRATION OF LICHEN COMPOUNDS
A
B
c
o !
10 !
20 !
7
30 !
2087
FIG. 2. (A) Detection of fumarprotöc·etraric acid, 7, in medulla portion of thalli of Cladonia pyxidata not eaten by Eilema camplana larvae; a lack of fumarprotocetraric acid in (8) larval feces and (C) adults of Eilema complana after feeding on cortical and algal layers of thalli of Cladonia pyxidata .
•

• 2088 HESBACHER ET AL: ,
The observed variation of sequestered lichen cornpounds in wild-caught imagines may be due to several reasons. The data presented in Tables land 2 are from insects with an unknown feeding history . Lack of lichen compounds in some imagines may, for example, have been caused by feeding on lichens with no or with only minute amounts of phenolic lichen compounds . Selective feeding on parts of thalli that are free of lichen compounds may be another reason for the observed variability . Preliminary feeding experiments with E. complana and the lichen Cladonia pyxidata, for example, indicate that the larvae feed only on the algal and corticallayers of the thallus, which are virtually free of phenolic compounds, but avoid the medulla, wh ich contains the bulk of fumarprotocetraric acid (7) . It is possible that similar feeding behavior that results in avoidance of lichen compounds deposited in the medulla regions of lichens may exist also in other species ofthe Arctiidae. The age ofthe specimens analyzed mayaiso influence the results obtained . Phenolic lichen compounds are usually stable and are not expected to be chemically altered during storage. However, if sequestered lichen compounds are primarily accumulated in the scales (which is unknown so far) , repeated handling of specimens, in addition to the natural wear and tear with age, could influence the results obtained. Nevertheless, this study demonstrates for the first time that sequestration of lichen compounds is variable, but widespread, in wild-caught arctiid moths.
It is not known yet if the lichen compounds sequestered by species of the Arctiidae or by lichen-feeding snails, which recently have been shown to sequester lichen compounds from their diet (Hesbacher et al. , 1995), are of ecological importance to the herbivores. However, it has been demonstrated that anthraquinone derivatives, such as chrysazin or chrysophanol, which are structurally closely related to parietin, are deterrent toward ants like Mynnica sabuleti (Hilker et al. , 1992). Anthraquinones are also known for their antimicrobial activities (Cudlin el al., 1976), and may be involved in chemical defense against microbial pathogens. It is therefore possible that storage of lichen compounds may result in chemical protection of insects or of snails from predators or from pathogens. Further studies on the roles of sequestered lichen compounds in invertebrate herbivores are underway.
Acknowledgments-Financial support from the Deutsche Forschungsgemeinschaft as a project of the "Sonderforschungsbereich 251 der Universität Würzburg" is gratefully acknowledged.
REFERENCES
BAUR, A., BAUR, B., and FRÖBERG, L. 1992. The effect of lichen diet on growth rate in the rockdwelling land snails Chondrina clienta (Westerlund) and Balea perversa (Linnaeus). J. Moll. Stud. 58:345-347.

I
SEQUESTRATION OF LICHEN COMPOUNDS 2089
BOPPRE , M., and SCHNEIDER , D. 1985 . Pyrrolizidine alkaloids quantitatively regulate both scent organ morphogenesis and pheromone biosynthesis in male Creatonotos moth (Lepidoptera: Arctiidae). 1. Comp. Physiol . 157(5):569-577.
CRITTENDEN, P.D., and PORTER , N. 1991. Lichen fonning fungi : Potential sources of novel metabolites. Tib. Tech. 9:409-414 .
CUDLlN , J ., BLUMAUEROVA, M., STEINEROVA, N. , MATEJU , J., and ZALABAK, V. 1976. Biological activity of hydroxyanthraquinones and their glucosides toward microorganisms. Folia Microbiol. 21 :54-57.
EHMKE, A., WITTE, L., BILLER , A. , and HARTMANN, T. 1990. Sequestration, N-oxidation and transfonnation of plant pyrrolizidine alkaloids by the arctiid moth Tyriajacobaeae L. Z. Naturforsch . 45c : 1185-1192.
EMBACHER , G. 1994. An Flechten lebende Grossschmetterlinge Salzburgs (Lepidoptera: Geometridae, Arctiidae, Noctuidae). Entomofauna 15 :209-224.
EMMERICH, R. , GIEZ, 1. , LANGE, O.L., and PROKSCH, P. 1993 . TOl(icity and antifeedant activity of lichen compounds against the polyphagous herbivorous insect Spodoptera littoralis. Phytochemütry 33: 1389-1394.
GIEZ, 1., LANGE, O.L. , and PROKSCH , P. 1994 . Growth retarding activity of lichen substances against the polyphagous herbivorous insect Spodoptera littoralis. Biochem. Syst. Ecol. 22 : 113-120.
HARTMANN. T. 1991. Alkaloids . pp. 79-121 , in G.A. Rosenthai and M.R. Berenbaum (eds.). Herbivores. Their Interactions with Secondary Plant Metabolites. Academic Press. New York.
HES BACHER. S .. BAUR, B., BAUR , A .. and PROKSCH. P. 1995. Sequestration of lichen compounds by three species of terrestrial snails. J . Chem. Ecol. 2:233-246 .
HILKER , M., ESCHBACH, U. , and DETT:-IER . K. 1992 . Occurrence of anthraquinones in eggs and larvae of several Galerucinae (Coleoptera: Chrysomelidae) . NaturwissenschafteIl 79:271-274.
MALCOLM . S.B. 1990. Chemical defense in chewing and sucking insect herbivores: plant derived cardenolides in the monarch butterfly and oleander aphid . Chemoecology I: 12-21.
RAMBOLD, G. 1985. Fütterungsel(perimente mit den an Flechten lebenden Raupen von Setina aurita Esp. Nachrichtenbl. bayer. Entomologen. 34 :82-90.
REUTIMANN . P .• and SCHEIDEGGER, C. 1987. Importance oflichen secondary products in food choice of two oribatid mites (Acari) in an alpine meadow ecosystem. J. Chem. Ecol. 13:363-370.
SEYD. E.L., and SEAWARD, M.R.D. 1984. The association of oribatid mites with lichens . Zool. J. Unn . Soc. 80:369-420.
STAHL, E. 1904. Die Schutzmittel der Flechten gegen Tierfraß, pp . 357-375, in Festschrift zum 70. Geburtstag von Ernst Haeckel. G. Fischer Verlag, Jena .
TRA WÖGER . A. 1991. Die evolutive Bedeutung introgressiver Hybridisation zwischen zwei Arten der Gattung Setina Schrank, 1802. Ber. Nat. -Med. Verein Innsbruck 78: 177-201 .
WIESEN. B. , KRUG. E. , FIEDLER, K., WRAY, V., and PROKSCH , P. 1994. Sequestration of hostplant derivcd flavonoids by the Lycaenid butterfly Polyommatus icarus. 1. Chem. Ecol. 20 :2523-
2538. ZUKAL, H. 1895. Morphologische und biologische Untersuchungen über die Flechten. Sber. K.
Böhm. Ces. Wiss. MatJz. -Nat. Kl . 104: 1303-1395 .
-
Copyright © 2022 FDOKUMEN




















