
Sensory and decisional factors in motion-induced blindness
12
Sensory and decisional factors in motion-induced blindness Laboratoire de Psychologie de la Perception (LPP), Paris Descartes University & CNRS, Paris, France Florent Caetta Laboratoire de Psychologie de la Perception (LPP), Paris Descartes University & CNRS, Paris, France Andrei Gorea Department of Neurobiology/Brain Research, Weizmann Institute of Science, Rehovot, Israel Yoram Bonneh The processes underlying motion-induced blindness (MIB) are widely debated. Ultimately, however, they must reduce to a sensitivity drop and/or to an upward shift of the decision criterion. The first possibility was tested by assessing the detection threshold for a contrast (or luminance) increment applied to the MIB target under its visible and suppressed phases. This was performed over a whole range of reference target contrasts (yielding the standard Threshold vs. Contrast [TvC] function) with a 2AFC staircase procedure. The second possibility was tested with a Yes/No procedure, allowing the assessment of both the sensitivities (d V ) and decision criteria (c) yielded by four contrast increments applied to a fixed reference target contrast (the psychometric function). The 2AFC procedure yielded a global upward shift of the TvC function of about 5.3 dB (a factor of 1.84) under the suppressed phase. The Yes/No procedure yielded a commensurate d Vdrop (of about 0.8 A) under the suppressed phase with no change in the slope of the psychometric function and an upward shift of c of about 0.7 A. The presently observed vertical shift (in log–log coordinates) of the TvC function in the suppressed phase is indicative of a divisive inhibition occurring after the transducing stage. The invariance of the psychometric function slope under the visible and suppressed MIB phases supports this conclusion. The present experiments cannot settle the issue of whether the upward shift of the decision criteria is yet another cause of the MIB or a consequence of its underlying inhibitory process. Instead, they make clear that MIB (and perhaps other unstable perceptual phenomena) is associated with both sensory and decisional processes. Keywords: motion-induced blindness, threshold versus contrast function, sensitivity, decision criterion Citation: Caetta, F., Gorea, A., & Bonneh, Y. (2007). Sensory and decisional factors in motion-induced blindness. Journal of Vision, 7(7):4, 1–12, http://journalofvision.org/7/7/4/, doi:10.1167/7.7.4. Introduction From a psychophysicist’s point of view, the Bspontaneous[ disappearance of a physically present and otherwise visible stimulus can result from only two causes: a drop in sensitivity and/or an upward shift of the perceptual decision criterionVwith reference to which an observer will assert that the stimulus is or is not perceived. Each of these two ultimate causes may be instantiated in a number of ways. A sensitivity drop may result from an inhibitory process, an increase of the noise level, and/or a change in the transducing process. Criterion changes can be brought about by a change of the observer’s priors and/or by interference with other decision processes. Any of these internal causes can be brought about and/or modulated by a multitude of external and internal factors such as task switching, multiple tasks and/or stimuli (contextual factors), and changes in vigilance/attentional level. Salient visual stimuli can be blocked from conscious perception under a number of conditions such as binocular rivalry (Levelt, 1965; Wheatstone, 1838), attentional blink (Raymond, Shapiro, & Arnell, 1992), repetition blindness (Kanwisher, 1987), inattentional blindness (Mack & Rock, 1998), change blindness (Rensink, O’Regan, & Clark, 1997), or different forms of masking. The latter, and more specifically metacontrast (Stigler, 1910; see reviews by Breitmeyer, 1984; Breitmeyer & Ogmen, 2000), were used to show that Binvisible[/nonconscious primes may nonetheless affect the perception of their masker (Klotz & Neuman, 1999; McGinnies, 1949), as well as trigger or modulate motor behavior (Goodale & Milner, 1992; Trevarthen, 1968). Equivalent altered perceptual con- sciousness states have been described in stroke patients with extinction (Bender, 1952; Critchley, 1949), neglect Journal of Vision (2007) 7(7):4, 1–12 http://journalofvision.org/7/7/4/ 1 doi: 10.1167/7.7.4 ISSN 1534-7362 * ARVO Received September 29, 2006; published May 23, 2007
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Sensory and decisional factors in motion-induced blindness

Sensory and decisional factors in motion-inducedblindness
Laboratoire de Psychologie de la Perception (LPP)Paris Descartes University amp CNRS
Paris FranceFlorent Caetta
Laboratoire de Psychologie de la Perception (LPP)Paris Descartes University amp CNRS
Paris FranceAndrei Gorea
Department of NeurobiologyBrain ResearchWeizmann Institute of Science
Rehovot IsraelYoram Bonneh
The processes underlying motion-induced blindness (MIB) are widely debated Ultimately however they must reduce to asensitivity drop andor to an upward shift of the decision criterion The first possibility was tested by assessing the detectionthreshold for a contrast (or luminance) increment applied to the MIB target under its visible and suppressed phases Thiswas performed over a whole range of reference target contrasts (yielding the standard Threshold vs Contrast [TvC]function) with a 2AFC staircase procedure The second possibility was tested with a YesNo procedure allowing theassessment of both the sensitivities (dV) and decision criteria (c) yielded by four contrast increments applied to a fixedreference target contrast (the psychometric function) The 2AFC procedure yielded a global upward shift of the TvC functionof about 53 dB (a factor of 184) under the suppressed phase The YesNo procedure yielded a commensurate dVdrop (ofabout 08 A) under the suppressed phase with no change in the slope of the psychometric function and an upward shift of cof about 07 A The presently observed vertical shift (in logndashlog coordinates) of the TvC function in the suppressed phase isindicative of a divisive inhibition occurring after the transducing stage The invariance of the psychometric function slopeunder the visible and suppressed MIB phases supports this conclusion The present experiments cannot settle the issue ofwhether the upward shift of the decision criteria is yet another cause of the MIB or a consequence of its underlying inhibitoryprocess Instead they make clear that MIB (and perhaps other unstable perceptual phenomena) is associated with bothsensory and decisional processes
Keywords motion-induced blindness threshold versus contrast function sensitivity decision criterion
Citation Caetta F Gorea A amp Bonneh Y (2007) Sensory and decisional factors in motion-induced blindness Journal ofVision 7(7)4 1ndash12 httpjournalofvisionorg774 doi101167774
Introduction
From a psychophysicistrsquos point of view theBspontaneous[ disappearance of a physically present andotherwise visible stimulus can result from only twocauses a drop in sensitivity andor an upward shift ofthe perceptual decision criterionVwith reference to whichan observer will assert that the stimulus is or is notperceived Each of these two ultimate causes may beinstantiated in a number of ways A sensitivity drop mayresult from an inhibitory process an increase of the noiselevel andor a change in the transducing processCriterion changes can be brought about by a change ofthe observerrsquos priors andor by interference with otherdecision processes Any of these internal causes can bebrought about andor modulated by a multitude of externaland internal factors such as task switching multiple tasks
andor stimuli (contextual factors) and changes invigilanceattentional levelSalient visual stimuli can be blocked from conscious
perception under a number of conditions such as binocularrivalry (Levelt 1965 Wheatstone 1838) attentional blink(Raymond Shapiro amp Arnell 1992) repetition blindness(Kanwisher 1987) inattentional blindness (Mack amp Rock1998) change blindness (Rensink OrsquoRegan amp Clark1997) or different forms of masking The latter and morespecifically metacontrast (Stigler 1910 see reviews byBreitmeyer 1984 Breitmeyer amp Ogmen 2000) wereused to show that Binvisible[nonconscious primes maynonetheless affect the perception of their masker (Klotz ampNeuman 1999 McGinnies 1949) as well as trigger ormodulate motor behavior (Goodale amp Milner 1992Trevarthen 1968) Equivalent altered perceptual con-sciousness states have been described in stroke patientswith extinction (Bender 1952 Critchley 1949) neglect
Journal of Vision (2007) 7(7)4 1ndash12 httpjournalofvisionorg774 1
doi 10 1167 7 7 4 ISSN 1534-7362 ARVOReceived September 29 2006 published May 23 2007
(Halligan amp Marshall 1993) and blindsight (Weiskrantz1990) Epitomes of the current empirical approach to theproblem of perceptual awareness these phenomena havebeen used as starting blocks for debates on whatconsciousness might be (eg Dennett 1991 Weiskrantz1986) as a means of quantifying awareness-relatedprocesses such as attention (see above) signal processingdisruption (eg Ogmen Breitmeyer amp Melvin 2003) orinterocular and pattern competition (for a review seeTong 2001) and as a substantiation of the dissociationbetween perception and action (Goodale amp Milner 1992)While the interpretations of the above phenomena havebeen scrutinized both at their conceptual level and fromthe standpoint of their neurophysiological substrate theparticularities of the recently discovered motion-inducedblindness (MIB Bonneh Cooperman amp Sagi 2001)phenomenon defy such analysesMIB refers to the spontaneous perceptual vanishing
(followed by reappearance) of highly salient peripheralvisual targets when superimposed on a globally moving(distracter) background Originally MIB was thought of asan instance of the Troxler effect (Troxler 1804) andor oflateral masking (by the moving background) These low-level accounts have been questioned in view of results byBonneh et al (2001) showing that MIB persists for(slowly) moving or flickering targets (hence preventing alocal low-level adaptationVthe main cause of the Troxlereffect Clarke amp Belcher 1962) as well as when thebackground was occluded within relatively large areasaround the targets (hence impeding lateral masking) Thefact that the invisible MIB targets contribute to theformation of negative aftereffects (Hofstoetter Koch ampKiper 2004) and yield an orientation selective adaptation(Montaser-Kouhsari Moradi Zandvakili amp Esteky 2004)was taken as evidence that the neural site of theirsuppression is beyond the primary visual cortex (but seeBlake Tadin Sobel Raissian amp Chong 2006) Thisconclusion is sustained by observations that MIB issensitive to Gestalt-like grouping effects (Bonneh et al2001 Hsu Yeh amp Kramer 2004) and to the manipulationof the depth planes where targets and distracters are pre-sented (Graf Adams amp Lages 2002) The latter obser-vation suggests that target and distracters compete forsaliency consistent with the proposal that MIB is butanother form of bistable perception reflecting the com-petition between neural mechanisms signaling irreconcil-able percepts (Funk amp Pettigrew 2003 see also KastnerDeWeerd Desimone amp Ungerleider 1998) All the findingsabove are compatible with an interpretation according towhich MIB could reveal (at least in part) a disruption ofattentional processing reminiscent of a competition betweenobject representations (Bonneh et al 2001) However thisaccount is in conflict with the most counterintuitiveobservation that MIB correlates positively with the saliencyof the targets (Bonneh et al 2001 Hsu et al 2004)Apart from a very few neglectextinction studies having
suggested that these conditions may also reflect changes in
subjectsrsquo decisional behavior (Azzopardi amp Cowey 1997Campion Latto amp Smith 1983 Gorea amp Sagi 20002002 Klein 1998) the ultimate account of a perceptualawareness impairment has been in terms of sensitivitychanges This was most extensively documented forbinocular rivalry where the sensitivity of the suppressedeye was shown to be significantly deteriorated (Blake1989 Blake amp Camisa 1979 Blake amp Fox 1974 Fox ampCheck 1972 Nguyen Freeman amp Wenderoth 2001Wales amp Fox 1970 Watanabe Paik amp Blake 2004)Watanabe et al (2004) measured the contrast incrementthreshold for the nonsuppressed and suppressed targetsas a function of this targetrsquos contrast Their results showthat the Threshold versus Contrast (TvC) function(Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980) in the suppressed phase isjust an upward shifted version of the TvC in the nonsup-pressed phaseBased on the assumption that the internal response R to
contrast C is a log function of C (R Egrave log(C) FechnerndashWeberrsquos law Fechner 1907) these authors interpretedtheir results as a manifestation of an inhibitory process(during suppression) occurring after the transducing stage(or gain control operator) Indeed as sensitivity ismodeled as the derivative of R with respect to C aposttransducer divisive inhibition m (ie R Egrave mlog(C)with m G 1) yields RC Egrave mC and C Egrave mj1C whichis an upward shift of the TvC function on a logndashlog scaleThe same conclusion would be reached if R were a powerfunction of C (R Egrave C+ Stevensrsquo law 1957) or a NakandashRushton function (R = RmaxC
+(s+ + C+) Naka amp Rushton1966 see Foley 1994) For a pretransducer inhibition thethree putative transducers yield different thresholdchanges no change at all for Fechnerrsquos law a downwardTvC shift for Stevensrsquo law and either an upward or adownward shift for NakandashRushton depending on therelationship between its free parameters (s +) and C1
Hence the processes underlying the suppression of anotherwise highly visible stimulus do not necessarilytranslate into a measurable sensitivity change Althoughthe counterintuitive option of a threshold drop under thesuppressed phase was rejected for binocular rivalry itremains possible for other oscillatory percepts such asMIB In fact given that contrary to other perceptualoscillatory phenomena (see Mueller amp Blake 1989) high-contrast stimuli disappear more frequently than low-contrast stimuli under MIB conditions (Bonneh et al2001) this option is even plausibleThe first purpose of the present study was to compare
the TvC functions (obtained with a 2AFC procedure)under MIB conditions for visible and suppressed targets Itshould be stressed from the start that in no way does thepresent study intend to inform on the validity of one of thethree transducers alluded to above over the remaining two(and even less on its putative location in the visualprocessing stream) In fact the critical literature is stilldebating on this issue (eg Meese 2004 Meese amp
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 2
Holmes 2002 Yu Klein amp Levi 2003) However basedon the logic developed above (see also Footnote 1) anupward TvC shift under the suppressed state will stronglyhint to the reliance on a posttransducer divisive inhibitionregardless of the transducerThe second purpose of the present study was to
determine whether the MIB phenomenon also involves adecision-criterion shift (during the suppressed phase) assuggested (Gorea amp Sagi 2000 2002 Klein 1998) andactually observed (Azzopardi amp Cowey 1997 Campionet al 1983 Ricci amp Chatterjee 2004) for neglect patientswith extinction To answer this question we used in thepresent study a YesNo experimental format and assesseddV and decision criteria for four contrast incrementsapplied to a highly suprathreshold MIB target undervisible and suppressed phases By so doing it allows arough estimation of the psychometric functions for thesetwo perceptual states and thereby an indirect derivation oftheir associated internal noises (related to the slopes ofthese functions) Again the observed constancy of thepsychometric function slopes under suppressed anddominant binocular rivalry phases (Sanders 1980 passimWatanabe et al 2004) is no guarantee of an equivalentresult under MIB conditions
Methods
Observers
Four volunteers who were naBve to the purpose of thisstudy and the first author (all of whom were 24ndash25 yearsold) with normal or corrected-to-normal vision partici-pated in the two experiments
Stimuli
They were presented on a 19-in gamma-correctedscreen (LG Flatron 795FT Plus 1024 750 pixelresolution and 75 Hz refresh rate) 70 cm away from theobservers in a dimly lit room The target stimulus was a03- 16- bar (Figure 1) presented at 64- (in the lefthemifield) from a white fixation cross (057- 057-) andsurrounded by a Bmask[ pattern subtending an 83- 83-area The mask pattern was composed of 49 (7 7)237 cdm2 blue crosses 07- 07- each The whole maskrotated clockwise about its center (coinciding with thecenter of the target) at 523 rads Fixation of a centralcross (at the center of the screen) presumably minimizedeye movements Target (also referred hereafter as pedestal)and mask were displayed on a 107-cdm2 background(L0) The targetpedestal was set at one of five luminancelevels equally spaced on a log scale between 141 and400 cdm2 Luminance increments were randomly applied
for 100 ms to either the upper or lower half of the verticalbar pedestal and observers had to specify which of the twohalves was incremented
Procedure
Luminance (or contrast see below) increment detectionthresholds as a function of pedestal luminance (TvCfunctions Experiment 1) and dVand decision criteria forone pedestal level (L = 312 cdm2) and four incrementlevels (Experiment 2) were measured in independentsessions for each of the two phenomenal states of theobservers that is target visible and suppressed In Btarget-visible[ sessions observers initiated a trial at their leisureby pressing a key while the target was visible in theBtarget-suppressed[ sessions they pressed the key as soonas the target vanished In either case a 100-ms luminanceincrement was applied 50 ms after the key press Thewhole display (mask and target) was erased 850 ms afterthe offset of the incrementIn Experiment 1 (run first) the TvC function was
assessed by means of a 2AFC staircase procedure Anincrement was randomly applied to either the lower or theupper part of the target bar (see Figure 1) and observersindicated its location by pressing one of the two mousebuttons The increment size at the beginning of thestaircase was set on average at 6 dB above the actualthreshold Following three consecutive correct responsesthe magnitude of the increment was decreased by 2 dBand increased by the same amount after each incorrectresponseVa procedure converging to 79 correctresponses (Levitt 1971) which is equivalent to a dVof114 (Macmillan amp Creelman 1991) The staircase wasended after eight reversals and the threshold wascomputed as the mean of the increment amplitudes overthe lasts six reversals Increment thresholds for the fivedifferent pedestal levels and the two phenomenal states
Figure 1 Experimental display (not at scale) used in the twoexperiments The dotted arrow indicates the rigid rotationalmovement of the background crosses The inset shows anenlargement of the target area where L0 LP and LT refer to theluminance of the background of the pedestal and of the testincrement respectively
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 3
were assessed in different sessions and repeated thriceThe order of the sessions (each specified by the pedestalluminance and phenomenal state) was randomized foreach repeat and observerIn Experiment 2 only one pedestal luminance (312 cdm2)
was used throughout and dVvalues together with decisioncriteria were assessed in separate sessions for fourincrement magnitudes by way of a YesNo procedurewith the increment randomly applied to either the lower orthe upper part of the bar in 50 of the trials The fourincrement amplitudes (124 137 150 and 162 dB) werechosen based on preliminary sessions so that the smallestand largest increments yield for all observers dVvaluesnot lower than 04 and not higher than 35 One session(specified by the increment magnitude and by theobserverrsquos phenomenal state) consisted of 100 trials andwas repeated thrice so that dVvalues and decision criteria(defined as c = 05(zH + zFA) where zH and zFA are thez scores of hits and false alarms2) were computed from300 trials per observer The order of the sessions wasrandomized for each observerIt should be noted that both the stimulation setup and
some aspects of the procedure were finalized after a longseries of preliminary experiments meant to maximize theduration and stability of the individual MIB durations andhence the reliability of the thresholddVmeasurementsOriginally these measurements were made with aBstandard[ MIB configuration where the rotating maskwas presented at the center of the screen and where thetarget (a 09- 09- cross) was presented 27- fromfixation in the upper left corner along the main diagonalThis configuration yielded relatively short MIB periods(G15 s) which together with a 100-ms delay between anobserverrsquos key press and the targetrsquos luminance incre-ment produced small and unstable threshold increments(compared to those assessed with visible targets)Postexperimental interviews revealed that in the target-suppressed perceptual states observers frequently expe-rienced premature target reappearance that is before orwithin milliseconds after the increment was appliedPremature target reappearances can be triggered both bythe abrupt luminance increment (Kawabe Yamada ampMiura in press) and by microsaccades set off by anobserverrsquos intention to press the key while focusing hisattention on the target location such premature targetreappearances are most likely favored under shallowsuppression states Hence the preliminary trial-and-errormanipulations were meant to find the stimulation param-eters that strengthen the suppression states In addition tothose preliminary observations indicated that the firsttarget disappearance within each trial tended to last lessand be more easily terminated by the test increment thansubsequent disappearances (presumably because of ashallower adaptation state) Hence observers were encour-aged to refrain from signaling it and await the second oreven the third disappearance in a trial These observations
are of importance as they point at the difficulty ofobtaining reliable threshold or dV assessments duringMIB and at the particular care needed to choose theoptimal stimulating and response recording conditions (fora comprehensive discussion of the topic see Kim ampBlake 2005)
Results
Observations on MIB phenomenology
The time lapse between the beginning of a trial and anobserverrsquos key press to signal a targetrsquos disappearance wasnot measured as observers were encouraged to ignore thefirst or even the first two disappearances (see the Methodssection) This instruction was based on our long-lastingexperience with MIB The duration of a disappearancetends to increase with inspection time and practice(Bonneh Sagi amp Cooperman 2005) suggesting theincremental contribution of an adaptation process Forone observer (out of five) the time lapse before the firstdisappearance was consistently longer (up to 1 min) thanusually reported Observers occasionally signaled partialtarget disappearances (ie of the upper or lower part ofthe pedestal) and were instructed to ignore them Allobservers signaled the reappearance of the suppressedpedestal as if triggered by the test increment when thisincrement was large (at the start of the staircase) but notwhen it was close to threshold Target reappearance wasalso reported on almost all occasions where observerswere aware of their inadvertent eye movements Twoobservers reported that their perception of the rotatingmask alternated between a state where its components(crosses) appeared as individualized items and a statewhere they were grouped to form a unitary rotatingsurface Interestingly they also reported an increasedlikelihood of a targetrsquos disappearance concomitant withthe transition to a latter state A potentially relatedphenomenon was reported by Graf et al (2002)
Experiment 1 TvC functions
The appropriate way of representing luminance incre-ment thresholds as a function of the pedestal luminanceLP has been a matter of debate (eg Kingdom ampMoulden 1991 Whittle 1986) For the present stimulusconfiguration where the reference pedestal was itselfdisplayed on a nonnull background L0 two data displayformats are equally legitimate whereby (a) the liminalluminance increment LE (= ordfLT j LPordf) is plotted as afunction of the pedestal luminance relative to the back-ground luminance that is LPrelative (= ordfLP j L0ordf) or
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 4
(b) the liminal contrast difference CE is plotted againstthe pedestal contrast (CP relative to the backgroundluminance TvC function) that is
CE frac14 jLT j L0jL0
jLP j L0
L0frac14 jLT j LPj
L0eth1aTHORN
and
CP frac14 jLP j L0jL0
eth1bTHORN
As it can be noted the contrast representation is but anormalized (by L0) version of the luminance representa-tion Inasmuch as the incremental threshold is a linearfunction of the pedestal magnitude in logndashlog coordinates(eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980 Watanabe et al 2004) the
two representations yield the same logndashlog slopes butdifferent intercepts For the sake of comparison withWatanabe et alrsquos (2004) equivalent study in binocularrivalry the data below are displayed in the second formatabove (TvC Equations 1a and 1b)Figure 2 displays TvC functions averaged over the five
observers (Figure 2a) and for one of them (Figure 2b)under target-visible (filled symbols) and target-suppressed(open symbols) conditions The mean suppressed-to-visibleCE ratio is 184 (026 log units) which is close to thatobtained in binocular rivalry (Watanabe et al 2004)3
A two-factor (pedestal contrast and phenomenal state)repeated measures ANOVA performed on the log(CE)data shows a significant effect of phenomenal stateF(1 4) = 201 p G 05 and pedestal contrast F(4 16) =1449 p G 001 with no significant interaction betweenthe two F(4 16) = 01 p = 99 This lack of interactionimplies that the visible and suppressed phases yieldequivalent TvC slopes (ie exponents of the fitted powerfunction see below)Whether the target was perceived or not contrast
thresholds (CE) are well fit by power functions of thepedestal contrast CP (CE = kCCP
5 solid and dashedstraight lines in Figure 2 for visible and suppressedphenomenal states respectively) Table 1 displays theexponents 5 (slopes in the logndashlog plots of Figure 2) andthe gain (ie intercept) parameters as estimated from thedata when expressed as contrast kC and as luminance kLthresholds (see above) for each observer together withtheir means The mean 5 is close to unity (Weberrsquos law)which is substantially higher than the exponents obtainedfor spatially modulated gratings (eg Foley amp Legge1981 Gorea amp Sagi 2001 Henning amp Wichmann 2007Legge 1981 Legge amp Foley 1980 Nachmias amp Sansbury1974) but close to the exponents inferred for luminancediscrimination thresholds with spatially localized stimuli(eg Cornsweet amp Teller 1965 Hood amp Finkelstein1986)As suggested by the analysis above the gain parameter
(kC or kL) increases in the suppressed phase (by anaverage factor of 192 and 233 depending on whether itis computed for the contrast or luminance data formatsrespectively) whereas the 5 exponent is equivalent for the
Figure 2 Increment CE thresholds averaged over the fiveobservers (a) and for one of them (b) for visible (filled symbols)and suppressed targets (open symbols) as a function of pedestalcontrast Cpedestal Solid and dashed straight lines are best powerfunction fits for the visible and suppressed conditions respec-tively Vertical bars are T1 SE across observers (a) and over thethree repeats for one observer (b)
Observer
5 (TvL and TvC) kC (TvC) kL (TvL)
Visible Suppressed Visible Suppressed Visible Suppressed
FC 119 124 022 030 014 017CC 115 120 024 064 018 041PC 109 085 032 076 027 110DG 105 114 025 041 023 029MT 141 132 021 028 008 013M 117 T 006 115 T 008 025 T 002 048 T 010 018 T 003 042 T 018
Table 1 Exponents (5) and gains (kC kL) of the power functions fitted to the TvL andor TvC data formats (Figure 2) for visible andsuppressed phenomenal states
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 5
two phenomenal states A linear regression analysisperformed on the log(CE) data of all observers con-firmed the parallelism between the visible and suppressedTvC functions as the difference between their slopes is notsignificant F(1 6) = 0022 whereas their intercepts differsignificantly F(1 7) = 4456 p G 0001 Linear regressionanalyses performed on each observerrsquos data yield equiv-alent effects with the exception of observer MT whoseintercepts for the visible and suppressed states do notdiffer significantlyOverall the present results and statistical analyses point
to a global vertical shift of the TvC function during MIBand are in line with Watanabe et alrsquos (2004) binocularrivalry data As argued in the Introduction section thisshift is compatible with a form of divisive inhibitionoperating during MIB and occurring after the transducingstage whether this transducer is of the form of a logpower or NakandashRushton function (see also Footnote 1)
Experiment 2 Sensitivity (dV) and decisioncriteria (c)
To cite from Green and Swets (1966 p 108) Btheprincipal value of the forced-choice task is that it practi-cally eliminates the need to deal with the observerrsquosdecision criterion[ The main purpose of the present YesNo experiment was precisely not to eliminate thisparameter as our concern was to assess its relation to theobserverrsquos decisional behavior under visible and sup-pressed MIB states To do so we fixed the targetpedestalluminance at 312 cdm2 and assessed dV values anddecision criteria for four increment levels The experi-mental format allows the assessment of the psychometric
functions obtained under the two phenomenal states andmost critically the test of the conjecture that thesuppression phase in MIB reflects at least in part anupward decision-criterion shift with respect to the visiblephaseFigure 3 shows dVvalues averaged over the five subjects
(Figure 3a) and for one of them (Figure 3b) under target-visible (filled symbols) and target-suppressed (opensymbols) conditions as a function of the target incrementmagnitude (in contrast units C) Solid and dotted linesare power function fits (dV= k(C) Pelli 1985 for a dVderivation based on a NakandashRushton transducer type seeYu et al 2003) Observers show an overall sensitivitydrop in the suppressed phase by an average factor of 177(an average of 084 A) A two-way repeated measuresANOVA (with phenomenal state and C as factors)shows a significant effect for the two factors phenomenalstate F(1 4) = 709 p G 05 C F(3 12) = 1939p G 001 with no significant interaction between themF(3 12) = 084 p = 50 This lack of interaction impliesthat the visible and suppressed phases yield psychometricfunctions with similar slopes (see below)The average psychometric function fits yield slightly
larger exponentsslopes () but smaller elevations (k) for thesuppressed ( = 158 k = 3) than for the visible ( = 145k = 49) MIB states A linear regression analysis performedon the mean log(dV) indicates that these slopes are notsignificantly different F(1 4) = 015 p = 72 whereas theelevations are F(1 5) = 20037 p G 001 Equivalentanalyses performed on the slopes and elevations of thepsychometric functions for each observer yield similareffects except for observer PC whose elevations do notdiffer significantly This shift of the psychometric functionalong the ordinate without changes in slope is indicative ofa divisive reduction of signal strength rather than of anincrease in the internal noise The preservation of thepsychometric function slopes under visible and suppressedstates has also been observed for binocular rivalry(Sanders 1980 passim Watanabe et al 2004)The mean CE values derived from the fitted dV
functions (CEExp 2 = (dVk)1) for a dV = 114 (thetheoretical sensitivity at the detection threshold E asassessed with the 2AFC 1-upndash3-down staircase procedureof Experiment 1) are globally lower than those actuallymeasured in Experiment 1 with the 312-cdm2 pedestalfor both the visible (CEExp 1 = 055 CEExp 2 = 037)and the suppressed (CEExp 1 = 113 CEExp 2 = 059)MIB states These differences are substantially larger thanthose previously reported (Green amp Swets 1966 p 112)The only account we can offer is that the lower thresholdsderived from the YesNo data reflect a learning effect asthis experiment was run after the 2AFC one As the 2AFCexperiment was run with five intermingled pedestal levels(as opposed to the one pedestal level used in the YesNoexperiment) it is also possible that the 2AFC thresholdswere relatively increased due to this additional Bnoise[factor
Figure 3 dV averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimuluschange (C) for visible (full symbols) and suppressed targets(open symbols) Standard errors are not shown for the individualdata (as in Panel b) because individual dVvalues were computedfrom the 300 trials run by each observer
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 6
Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

(Halligan amp Marshall 1993) and blindsight (Weiskrantz1990) Epitomes of the current empirical approach to theproblem of perceptual awareness these phenomena havebeen used as starting blocks for debates on whatconsciousness might be (eg Dennett 1991 Weiskrantz1986) as a means of quantifying awareness-relatedprocesses such as attention (see above) signal processingdisruption (eg Ogmen Breitmeyer amp Melvin 2003) orinterocular and pattern competition (for a review seeTong 2001) and as a substantiation of the dissociationbetween perception and action (Goodale amp Milner 1992)While the interpretations of the above phenomena havebeen scrutinized both at their conceptual level and fromthe standpoint of their neurophysiological substrate theparticularities of the recently discovered motion-inducedblindness (MIB Bonneh Cooperman amp Sagi 2001)phenomenon defy such analysesMIB refers to the spontaneous perceptual vanishing
(followed by reappearance) of highly salient peripheralvisual targets when superimposed on a globally moving(distracter) background Originally MIB was thought of asan instance of the Troxler effect (Troxler 1804) andor oflateral masking (by the moving background) These low-level accounts have been questioned in view of results byBonneh et al (2001) showing that MIB persists for(slowly) moving or flickering targets (hence preventing alocal low-level adaptationVthe main cause of the Troxlereffect Clarke amp Belcher 1962) as well as when thebackground was occluded within relatively large areasaround the targets (hence impeding lateral masking) Thefact that the invisible MIB targets contribute to theformation of negative aftereffects (Hofstoetter Koch ampKiper 2004) and yield an orientation selective adaptation(Montaser-Kouhsari Moradi Zandvakili amp Esteky 2004)was taken as evidence that the neural site of theirsuppression is beyond the primary visual cortex (but seeBlake Tadin Sobel Raissian amp Chong 2006) Thisconclusion is sustained by observations that MIB issensitive to Gestalt-like grouping effects (Bonneh et al2001 Hsu Yeh amp Kramer 2004) and to the manipulationof the depth planes where targets and distracters are pre-sented (Graf Adams amp Lages 2002) The latter obser-vation suggests that target and distracters compete forsaliency consistent with the proposal that MIB is butanother form of bistable perception reflecting the com-petition between neural mechanisms signaling irreconcil-able percepts (Funk amp Pettigrew 2003 see also KastnerDeWeerd Desimone amp Ungerleider 1998) All the findingsabove are compatible with an interpretation according towhich MIB could reveal (at least in part) a disruption ofattentional processing reminiscent of a competition betweenobject representations (Bonneh et al 2001) However thisaccount is in conflict with the most counterintuitiveobservation that MIB correlates positively with the saliencyof the targets (Bonneh et al 2001 Hsu et al 2004)Apart from a very few neglectextinction studies having
suggested that these conditions may also reflect changes in
subjectsrsquo decisional behavior (Azzopardi amp Cowey 1997Campion Latto amp Smith 1983 Gorea amp Sagi 20002002 Klein 1998) the ultimate account of a perceptualawareness impairment has been in terms of sensitivitychanges This was most extensively documented forbinocular rivalry where the sensitivity of the suppressedeye was shown to be significantly deteriorated (Blake1989 Blake amp Camisa 1979 Blake amp Fox 1974 Fox ampCheck 1972 Nguyen Freeman amp Wenderoth 2001Wales amp Fox 1970 Watanabe Paik amp Blake 2004)Watanabe et al (2004) measured the contrast incrementthreshold for the nonsuppressed and suppressed targetsas a function of this targetrsquos contrast Their results showthat the Threshold versus Contrast (TvC) function(Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980) in the suppressed phase isjust an upward shifted version of the TvC in the nonsup-pressed phaseBased on the assumption that the internal response R to
contrast C is a log function of C (R Egrave log(C) FechnerndashWeberrsquos law Fechner 1907) these authors interpretedtheir results as a manifestation of an inhibitory process(during suppression) occurring after the transducing stage(or gain control operator) Indeed as sensitivity ismodeled as the derivative of R with respect to C aposttransducer divisive inhibition m (ie R Egrave mlog(C)with m G 1) yields RC Egrave mC and C Egrave mj1C whichis an upward shift of the TvC function on a logndashlog scaleThe same conclusion would be reached if R were a powerfunction of C (R Egrave C+ Stevensrsquo law 1957) or a NakandashRushton function (R = RmaxC
+(s+ + C+) Naka amp Rushton1966 see Foley 1994) For a pretransducer inhibition thethree putative transducers yield different thresholdchanges no change at all for Fechnerrsquos law a downwardTvC shift for Stevensrsquo law and either an upward or adownward shift for NakandashRushton depending on therelationship between its free parameters (s +) and C1
Hence the processes underlying the suppression of anotherwise highly visible stimulus do not necessarilytranslate into a measurable sensitivity change Althoughthe counterintuitive option of a threshold drop under thesuppressed phase was rejected for binocular rivalry itremains possible for other oscillatory percepts such asMIB In fact given that contrary to other perceptualoscillatory phenomena (see Mueller amp Blake 1989) high-contrast stimuli disappear more frequently than low-contrast stimuli under MIB conditions (Bonneh et al2001) this option is even plausibleThe first purpose of the present study was to compare
the TvC functions (obtained with a 2AFC procedure)under MIB conditions for visible and suppressed targets Itshould be stressed from the start that in no way does thepresent study intend to inform on the validity of one of thethree transducers alluded to above over the remaining two(and even less on its putative location in the visualprocessing stream) In fact the critical literature is stilldebating on this issue (eg Meese 2004 Meese amp
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 2
Holmes 2002 Yu Klein amp Levi 2003) However basedon the logic developed above (see also Footnote 1) anupward TvC shift under the suppressed state will stronglyhint to the reliance on a posttransducer divisive inhibitionregardless of the transducerThe second purpose of the present study was to
determine whether the MIB phenomenon also involves adecision-criterion shift (during the suppressed phase) assuggested (Gorea amp Sagi 2000 2002 Klein 1998) andactually observed (Azzopardi amp Cowey 1997 Campionet al 1983 Ricci amp Chatterjee 2004) for neglect patientswith extinction To answer this question we used in thepresent study a YesNo experimental format and assesseddV and decision criteria for four contrast incrementsapplied to a highly suprathreshold MIB target undervisible and suppressed phases By so doing it allows arough estimation of the psychometric functions for thesetwo perceptual states and thereby an indirect derivation oftheir associated internal noises (related to the slopes ofthese functions) Again the observed constancy of thepsychometric function slopes under suppressed anddominant binocular rivalry phases (Sanders 1980 passimWatanabe et al 2004) is no guarantee of an equivalentresult under MIB conditions
Methods
Observers
Four volunteers who were naBve to the purpose of thisstudy and the first author (all of whom were 24ndash25 yearsold) with normal or corrected-to-normal vision partici-pated in the two experiments
Stimuli
They were presented on a 19-in gamma-correctedscreen (LG Flatron 795FT Plus 1024 750 pixelresolution and 75 Hz refresh rate) 70 cm away from theobservers in a dimly lit room The target stimulus was a03- 16- bar (Figure 1) presented at 64- (in the lefthemifield) from a white fixation cross (057- 057-) andsurrounded by a Bmask[ pattern subtending an 83- 83-area The mask pattern was composed of 49 (7 7)237 cdm2 blue crosses 07- 07- each The whole maskrotated clockwise about its center (coinciding with thecenter of the target) at 523 rads Fixation of a centralcross (at the center of the screen) presumably minimizedeye movements Target (also referred hereafter as pedestal)and mask were displayed on a 107-cdm2 background(L0) The targetpedestal was set at one of five luminancelevels equally spaced on a log scale between 141 and400 cdm2 Luminance increments were randomly applied
for 100 ms to either the upper or lower half of the verticalbar pedestal and observers had to specify which of the twohalves was incremented
Procedure
Luminance (or contrast see below) increment detectionthresholds as a function of pedestal luminance (TvCfunctions Experiment 1) and dVand decision criteria forone pedestal level (L = 312 cdm2) and four incrementlevels (Experiment 2) were measured in independentsessions for each of the two phenomenal states of theobservers that is target visible and suppressed In Btarget-visible[ sessions observers initiated a trial at their leisureby pressing a key while the target was visible in theBtarget-suppressed[ sessions they pressed the key as soonas the target vanished In either case a 100-ms luminanceincrement was applied 50 ms after the key press Thewhole display (mask and target) was erased 850 ms afterthe offset of the incrementIn Experiment 1 (run first) the TvC function was
assessed by means of a 2AFC staircase procedure Anincrement was randomly applied to either the lower or theupper part of the target bar (see Figure 1) and observersindicated its location by pressing one of the two mousebuttons The increment size at the beginning of thestaircase was set on average at 6 dB above the actualthreshold Following three consecutive correct responsesthe magnitude of the increment was decreased by 2 dBand increased by the same amount after each incorrectresponseVa procedure converging to 79 correctresponses (Levitt 1971) which is equivalent to a dVof114 (Macmillan amp Creelman 1991) The staircase wasended after eight reversals and the threshold wascomputed as the mean of the increment amplitudes overthe lasts six reversals Increment thresholds for the fivedifferent pedestal levels and the two phenomenal states
Figure 1 Experimental display (not at scale) used in the twoexperiments The dotted arrow indicates the rigid rotationalmovement of the background crosses The inset shows anenlargement of the target area where L0 LP and LT refer to theluminance of the background of the pedestal and of the testincrement respectively
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 3
were assessed in different sessions and repeated thriceThe order of the sessions (each specified by the pedestalluminance and phenomenal state) was randomized foreach repeat and observerIn Experiment 2 only one pedestal luminance (312 cdm2)
was used throughout and dVvalues together with decisioncriteria were assessed in separate sessions for fourincrement magnitudes by way of a YesNo procedurewith the increment randomly applied to either the lower orthe upper part of the bar in 50 of the trials The fourincrement amplitudes (124 137 150 and 162 dB) werechosen based on preliminary sessions so that the smallestand largest increments yield for all observers dVvaluesnot lower than 04 and not higher than 35 One session(specified by the increment magnitude and by theobserverrsquos phenomenal state) consisted of 100 trials andwas repeated thrice so that dVvalues and decision criteria(defined as c = 05(zH + zFA) where zH and zFA are thez scores of hits and false alarms2) were computed from300 trials per observer The order of the sessions wasrandomized for each observerIt should be noted that both the stimulation setup and
some aspects of the procedure were finalized after a longseries of preliminary experiments meant to maximize theduration and stability of the individual MIB durations andhence the reliability of the thresholddVmeasurementsOriginally these measurements were made with aBstandard[ MIB configuration where the rotating maskwas presented at the center of the screen and where thetarget (a 09- 09- cross) was presented 27- fromfixation in the upper left corner along the main diagonalThis configuration yielded relatively short MIB periods(G15 s) which together with a 100-ms delay between anobserverrsquos key press and the targetrsquos luminance incre-ment produced small and unstable threshold increments(compared to those assessed with visible targets)Postexperimental interviews revealed that in the target-suppressed perceptual states observers frequently expe-rienced premature target reappearance that is before orwithin milliseconds after the increment was appliedPremature target reappearances can be triggered both bythe abrupt luminance increment (Kawabe Yamada ampMiura in press) and by microsaccades set off by anobserverrsquos intention to press the key while focusing hisattention on the target location such premature targetreappearances are most likely favored under shallowsuppression states Hence the preliminary trial-and-errormanipulations were meant to find the stimulation param-eters that strengthen the suppression states In addition tothose preliminary observations indicated that the firsttarget disappearance within each trial tended to last lessand be more easily terminated by the test increment thansubsequent disappearances (presumably because of ashallower adaptation state) Hence observers were encour-aged to refrain from signaling it and await the second oreven the third disappearance in a trial These observations
are of importance as they point at the difficulty ofobtaining reliable threshold or dV assessments duringMIB and at the particular care needed to choose theoptimal stimulating and response recording conditions (fora comprehensive discussion of the topic see Kim ampBlake 2005)
Results
Observations on MIB phenomenology
The time lapse between the beginning of a trial and anobserverrsquos key press to signal a targetrsquos disappearance wasnot measured as observers were encouraged to ignore thefirst or even the first two disappearances (see the Methodssection) This instruction was based on our long-lastingexperience with MIB The duration of a disappearancetends to increase with inspection time and practice(Bonneh Sagi amp Cooperman 2005) suggesting theincremental contribution of an adaptation process Forone observer (out of five) the time lapse before the firstdisappearance was consistently longer (up to 1 min) thanusually reported Observers occasionally signaled partialtarget disappearances (ie of the upper or lower part ofthe pedestal) and were instructed to ignore them Allobservers signaled the reappearance of the suppressedpedestal as if triggered by the test increment when thisincrement was large (at the start of the staircase) but notwhen it was close to threshold Target reappearance wasalso reported on almost all occasions where observerswere aware of their inadvertent eye movements Twoobservers reported that their perception of the rotatingmask alternated between a state where its components(crosses) appeared as individualized items and a statewhere they were grouped to form a unitary rotatingsurface Interestingly they also reported an increasedlikelihood of a targetrsquos disappearance concomitant withthe transition to a latter state A potentially relatedphenomenon was reported by Graf et al (2002)
Experiment 1 TvC functions
The appropriate way of representing luminance incre-ment thresholds as a function of the pedestal luminanceLP has been a matter of debate (eg Kingdom ampMoulden 1991 Whittle 1986) For the present stimulusconfiguration where the reference pedestal was itselfdisplayed on a nonnull background L0 two data displayformats are equally legitimate whereby (a) the liminalluminance increment LE (= ordfLT j LPordf) is plotted as afunction of the pedestal luminance relative to the back-ground luminance that is LPrelative (= ordfLP j L0ordf) or
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 4
(b) the liminal contrast difference CE is plotted againstthe pedestal contrast (CP relative to the backgroundluminance TvC function) that is
CE frac14 jLT j L0jL0
jLP j L0
L0frac14 jLT j LPj
L0eth1aTHORN
and
CP frac14 jLP j L0jL0
eth1bTHORN
As it can be noted the contrast representation is but anormalized (by L0) version of the luminance representa-tion Inasmuch as the incremental threshold is a linearfunction of the pedestal magnitude in logndashlog coordinates(eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980 Watanabe et al 2004) the
two representations yield the same logndashlog slopes butdifferent intercepts For the sake of comparison withWatanabe et alrsquos (2004) equivalent study in binocularrivalry the data below are displayed in the second formatabove (TvC Equations 1a and 1b)Figure 2 displays TvC functions averaged over the five
observers (Figure 2a) and for one of them (Figure 2b)under target-visible (filled symbols) and target-suppressed(open symbols) conditions The mean suppressed-to-visibleCE ratio is 184 (026 log units) which is close to thatobtained in binocular rivalry (Watanabe et al 2004)3
A two-factor (pedestal contrast and phenomenal state)repeated measures ANOVA performed on the log(CE)data shows a significant effect of phenomenal stateF(1 4) = 201 p G 05 and pedestal contrast F(4 16) =1449 p G 001 with no significant interaction betweenthe two F(4 16) = 01 p = 99 This lack of interactionimplies that the visible and suppressed phases yieldequivalent TvC slopes (ie exponents of the fitted powerfunction see below)Whether the target was perceived or not contrast
thresholds (CE) are well fit by power functions of thepedestal contrast CP (CE = kCCP
5 solid and dashedstraight lines in Figure 2 for visible and suppressedphenomenal states respectively) Table 1 displays theexponents 5 (slopes in the logndashlog plots of Figure 2) andthe gain (ie intercept) parameters as estimated from thedata when expressed as contrast kC and as luminance kLthresholds (see above) for each observer together withtheir means The mean 5 is close to unity (Weberrsquos law)which is substantially higher than the exponents obtainedfor spatially modulated gratings (eg Foley amp Legge1981 Gorea amp Sagi 2001 Henning amp Wichmann 2007Legge 1981 Legge amp Foley 1980 Nachmias amp Sansbury1974) but close to the exponents inferred for luminancediscrimination thresholds with spatially localized stimuli(eg Cornsweet amp Teller 1965 Hood amp Finkelstein1986)As suggested by the analysis above the gain parameter
(kC or kL) increases in the suppressed phase (by anaverage factor of 192 and 233 depending on whether itis computed for the contrast or luminance data formatsrespectively) whereas the 5 exponent is equivalent for the
Figure 2 Increment CE thresholds averaged over the fiveobservers (a) and for one of them (b) for visible (filled symbols)and suppressed targets (open symbols) as a function of pedestalcontrast Cpedestal Solid and dashed straight lines are best powerfunction fits for the visible and suppressed conditions respec-tively Vertical bars are T1 SE across observers (a) and over thethree repeats for one observer (b)
Observer
5 (TvL and TvC) kC (TvC) kL (TvL)
Visible Suppressed Visible Suppressed Visible Suppressed
FC 119 124 022 030 014 017CC 115 120 024 064 018 041PC 109 085 032 076 027 110DG 105 114 025 041 023 029MT 141 132 021 028 008 013M 117 T 006 115 T 008 025 T 002 048 T 010 018 T 003 042 T 018
Table 1 Exponents (5) and gains (kC kL) of the power functions fitted to the TvL andor TvC data formats (Figure 2) for visible andsuppressed phenomenal states
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 5
two phenomenal states A linear regression analysisperformed on the log(CE) data of all observers con-firmed the parallelism between the visible and suppressedTvC functions as the difference between their slopes is notsignificant F(1 6) = 0022 whereas their intercepts differsignificantly F(1 7) = 4456 p G 0001 Linear regressionanalyses performed on each observerrsquos data yield equiv-alent effects with the exception of observer MT whoseintercepts for the visible and suppressed states do notdiffer significantlyOverall the present results and statistical analyses point
to a global vertical shift of the TvC function during MIBand are in line with Watanabe et alrsquos (2004) binocularrivalry data As argued in the Introduction section thisshift is compatible with a form of divisive inhibitionoperating during MIB and occurring after the transducingstage whether this transducer is of the form of a logpower or NakandashRushton function (see also Footnote 1)
Experiment 2 Sensitivity (dV) and decisioncriteria (c)
To cite from Green and Swets (1966 p 108) Btheprincipal value of the forced-choice task is that it practi-cally eliminates the need to deal with the observerrsquosdecision criterion[ The main purpose of the present YesNo experiment was precisely not to eliminate thisparameter as our concern was to assess its relation to theobserverrsquos decisional behavior under visible and sup-pressed MIB states To do so we fixed the targetpedestalluminance at 312 cdm2 and assessed dV values anddecision criteria for four increment levels The experi-mental format allows the assessment of the psychometric
functions obtained under the two phenomenal states andmost critically the test of the conjecture that thesuppression phase in MIB reflects at least in part anupward decision-criterion shift with respect to the visiblephaseFigure 3 shows dVvalues averaged over the five subjects
(Figure 3a) and for one of them (Figure 3b) under target-visible (filled symbols) and target-suppressed (opensymbols) conditions as a function of the target incrementmagnitude (in contrast units C) Solid and dotted linesare power function fits (dV= k(C) Pelli 1985 for a dVderivation based on a NakandashRushton transducer type seeYu et al 2003) Observers show an overall sensitivitydrop in the suppressed phase by an average factor of 177(an average of 084 A) A two-way repeated measuresANOVA (with phenomenal state and C as factors)shows a significant effect for the two factors phenomenalstate F(1 4) = 709 p G 05 C F(3 12) = 1939p G 001 with no significant interaction between themF(3 12) = 084 p = 50 This lack of interaction impliesthat the visible and suppressed phases yield psychometricfunctions with similar slopes (see below)The average psychometric function fits yield slightly
larger exponentsslopes () but smaller elevations (k) for thesuppressed ( = 158 k = 3) than for the visible ( = 145k = 49) MIB states A linear regression analysis performedon the mean log(dV) indicates that these slopes are notsignificantly different F(1 4) = 015 p = 72 whereas theelevations are F(1 5) = 20037 p G 001 Equivalentanalyses performed on the slopes and elevations of thepsychometric functions for each observer yield similareffects except for observer PC whose elevations do notdiffer significantly This shift of the psychometric functionalong the ordinate without changes in slope is indicative ofa divisive reduction of signal strength rather than of anincrease in the internal noise The preservation of thepsychometric function slopes under visible and suppressedstates has also been observed for binocular rivalry(Sanders 1980 passim Watanabe et al 2004)The mean CE values derived from the fitted dV
functions (CEExp 2 = (dVk)1) for a dV = 114 (thetheoretical sensitivity at the detection threshold E asassessed with the 2AFC 1-upndash3-down staircase procedureof Experiment 1) are globally lower than those actuallymeasured in Experiment 1 with the 312-cdm2 pedestalfor both the visible (CEExp 1 = 055 CEExp 2 = 037)and the suppressed (CEExp 1 = 113 CEExp 2 = 059)MIB states These differences are substantially larger thanthose previously reported (Green amp Swets 1966 p 112)The only account we can offer is that the lower thresholdsderived from the YesNo data reflect a learning effect asthis experiment was run after the 2AFC one As the 2AFCexperiment was run with five intermingled pedestal levels(as opposed to the one pedestal level used in the YesNoexperiment) it is also possible that the 2AFC thresholdswere relatively increased due to this additional Bnoise[factor
Figure 3 dV averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimuluschange (C) for visible (full symbols) and suppressed targets(open symbols) Standard errors are not shown for the individualdata (as in Panel b) because individual dVvalues were computedfrom the 300 trials run by each observer
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 6
Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

Holmes 2002 Yu Klein amp Levi 2003) However basedon the logic developed above (see also Footnote 1) anupward TvC shift under the suppressed state will stronglyhint to the reliance on a posttransducer divisive inhibitionregardless of the transducerThe second purpose of the present study was to
determine whether the MIB phenomenon also involves adecision-criterion shift (during the suppressed phase) assuggested (Gorea amp Sagi 2000 2002 Klein 1998) andactually observed (Azzopardi amp Cowey 1997 Campionet al 1983 Ricci amp Chatterjee 2004) for neglect patientswith extinction To answer this question we used in thepresent study a YesNo experimental format and assesseddV and decision criteria for four contrast incrementsapplied to a highly suprathreshold MIB target undervisible and suppressed phases By so doing it allows arough estimation of the psychometric functions for thesetwo perceptual states and thereby an indirect derivation oftheir associated internal noises (related to the slopes ofthese functions) Again the observed constancy of thepsychometric function slopes under suppressed anddominant binocular rivalry phases (Sanders 1980 passimWatanabe et al 2004) is no guarantee of an equivalentresult under MIB conditions
Methods
Observers
Four volunteers who were naBve to the purpose of thisstudy and the first author (all of whom were 24ndash25 yearsold) with normal or corrected-to-normal vision partici-pated in the two experiments
Stimuli
They were presented on a 19-in gamma-correctedscreen (LG Flatron 795FT Plus 1024 750 pixelresolution and 75 Hz refresh rate) 70 cm away from theobservers in a dimly lit room The target stimulus was a03- 16- bar (Figure 1) presented at 64- (in the lefthemifield) from a white fixation cross (057- 057-) andsurrounded by a Bmask[ pattern subtending an 83- 83-area The mask pattern was composed of 49 (7 7)237 cdm2 blue crosses 07- 07- each The whole maskrotated clockwise about its center (coinciding with thecenter of the target) at 523 rads Fixation of a centralcross (at the center of the screen) presumably minimizedeye movements Target (also referred hereafter as pedestal)and mask were displayed on a 107-cdm2 background(L0) The targetpedestal was set at one of five luminancelevels equally spaced on a log scale between 141 and400 cdm2 Luminance increments were randomly applied
for 100 ms to either the upper or lower half of the verticalbar pedestal and observers had to specify which of the twohalves was incremented
Procedure
Luminance (or contrast see below) increment detectionthresholds as a function of pedestal luminance (TvCfunctions Experiment 1) and dVand decision criteria forone pedestal level (L = 312 cdm2) and four incrementlevels (Experiment 2) were measured in independentsessions for each of the two phenomenal states of theobservers that is target visible and suppressed In Btarget-visible[ sessions observers initiated a trial at their leisureby pressing a key while the target was visible in theBtarget-suppressed[ sessions they pressed the key as soonas the target vanished In either case a 100-ms luminanceincrement was applied 50 ms after the key press Thewhole display (mask and target) was erased 850 ms afterthe offset of the incrementIn Experiment 1 (run first) the TvC function was
assessed by means of a 2AFC staircase procedure Anincrement was randomly applied to either the lower or theupper part of the target bar (see Figure 1) and observersindicated its location by pressing one of the two mousebuttons The increment size at the beginning of thestaircase was set on average at 6 dB above the actualthreshold Following three consecutive correct responsesthe magnitude of the increment was decreased by 2 dBand increased by the same amount after each incorrectresponseVa procedure converging to 79 correctresponses (Levitt 1971) which is equivalent to a dVof114 (Macmillan amp Creelman 1991) The staircase wasended after eight reversals and the threshold wascomputed as the mean of the increment amplitudes overthe lasts six reversals Increment thresholds for the fivedifferent pedestal levels and the two phenomenal states
Figure 1 Experimental display (not at scale) used in the twoexperiments The dotted arrow indicates the rigid rotationalmovement of the background crosses The inset shows anenlargement of the target area where L0 LP and LT refer to theluminance of the background of the pedestal and of the testincrement respectively
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 3
were assessed in different sessions and repeated thriceThe order of the sessions (each specified by the pedestalluminance and phenomenal state) was randomized foreach repeat and observerIn Experiment 2 only one pedestal luminance (312 cdm2)
was used throughout and dVvalues together with decisioncriteria were assessed in separate sessions for fourincrement magnitudes by way of a YesNo procedurewith the increment randomly applied to either the lower orthe upper part of the bar in 50 of the trials The fourincrement amplitudes (124 137 150 and 162 dB) werechosen based on preliminary sessions so that the smallestand largest increments yield for all observers dVvaluesnot lower than 04 and not higher than 35 One session(specified by the increment magnitude and by theobserverrsquos phenomenal state) consisted of 100 trials andwas repeated thrice so that dVvalues and decision criteria(defined as c = 05(zH + zFA) where zH and zFA are thez scores of hits and false alarms2) were computed from300 trials per observer The order of the sessions wasrandomized for each observerIt should be noted that both the stimulation setup and
some aspects of the procedure were finalized after a longseries of preliminary experiments meant to maximize theduration and stability of the individual MIB durations andhence the reliability of the thresholddVmeasurementsOriginally these measurements were made with aBstandard[ MIB configuration where the rotating maskwas presented at the center of the screen and where thetarget (a 09- 09- cross) was presented 27- fromfixation in the upper left corner along the main diagonalThis configuration yielded relatively short MIB periods(G15 s) which together with a 100-ms delay between anobserverrsquos key press and the targetrsquos luminance incre-ment produced small and unstable threshold increments(compared to those assessed with visible targets)Postexperimental interviews revealed that in the target-suppressed perceptual states observers frequently expe-rienced premature target reappearance that is before orwithin milliseconds after the increment was appliedPremature target reappearances can be triggered both bythe abrupt luminance increment (Kawabe Yamada ampMiura in press) and by microsaccades set off by anobserverrsquos intention to press the key while focusing hisattention on the target location such premature targetreappearances are most likely favored under shallowsuppression states Hence the preliminary trial-and-errormanipulations were meant to find the stimulation param-eters that strengthen the suppression states In addition tothose preliminary observations indicated that the firsttarget disappearance within each trial tended to last lessand be more easily terminated by the test increment thansubsequent disappearances (presumably because of ashallower adaptation state) Hence observers were encour-aged to refrain from signaling it and await the second oreven the third disappearance in a trial These observations
are of importance as they point at the difficulty ofobtaining reliable threshold or dV assessments duringMIB and at the particular care needed to choose theoptimal stimulating and response recording conditions (fora comprehensive discussion of the topic see Kim ampBlake 2005)
Results
Observations on MIB phenomenology
The time lapse between the beginning of a trial and anobserverrsquos key press to signal a targetrsquos disappearance wasnot measured as observers were encouraged to ignore thefirst or even the first two disappearances (see the Methodssection) This instruction was based on our long-lastingexperience with MIB The duration of a disappearancetends to increase with inspection time and practice(Bonneh Sagi amp Cooperman 2005) suggesting theincremental contribution of an adaptation process Forone observer (out of five) the time lapse before the firstdisappearance was consistently longer (up to 1 min) thanusually reported Observers occasionally signaled partialtarget disappearances (ie of the upper or lower part ofthe pedestal) and were instructed to ignore them Allobservers signaled the reappearance of the suppressedpedestal as if triggered by the test increment when thisincrement was large (at the start of the staircase) but notwhen it was close to threshold Target reappearance wasalso reported on almost all occasions where observerswere aware of their inadvertent eye movements Twoobservers reported that their perception of the rotatingmask alternated between a state where its components(crosses) appeared as individualized items and a statewhere they were grouped to form a unitary rotatingsurface Interestingly they also reported an increasedlikelihood of a targetrsquos disappearance concomitant withthe transition to a latter state A potentially relatedphenomenon was reported by Graf et al (2002)
Experiment 1 TvC functions
The appropriate way of representing luminance incre-ment thresholds as a function of the pedestal luminanceLP has been a matter of debate (eg Kingdom ampMoulden 1991 Whittle 1986) For the present stimulusconfiguration where the reference pedestal was itselfdisplayed on a nonnull background L0 two data displayformats are equally legitimate whereby (a) the liminalluminance increment LE (= ordfLT j LPordf) is plotted as afunction of the pedestal luminance relative to the back-ground luminance that is LPrelative (= ordfLP j L0ordf) or
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 4
(b) the liminal contrast difference CE is plotted againstthe pedestal contrast (CP relative to the backgroundluminance TvC function) that is
CE frac14 jLT j L0jL0
jLP j L0
L0frac14 jLT j LPj
L0eth1aTHORN
and
CP frac14 jLP j L0jL0
eth1bTHORN
As it can be noted the contrast representation is but anormalized (by L0) version of the luminance representa-tion Inasmuch as the incremental threshold is a linearfunction of the pedestal magnitude in logndashlog coordinates(eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980 Watanabe et al 2004) the
two representations yield the same logndashlog slopes butdifferent intercepts For the sake of comparison withWatanabe et alrsquos (2004) equivalent study in binocularrivalry the data below are displayed in the second formatabove (TvC Equations 1a and 1b)Figure 2 displays TvC functions averaged over the five
observers (Figure 2a) and for one of them (Figure 2b)under target-visible (filled symbols) and target-suppressed(open symbols) conditions The mean suppressed-to-visibleCE ratio is 184 (026 log units) which is close to thatobtained in binocular rivalry (Watanabe et al 2004)3
A two-factor (pedestal contrast and phenomenal state)repeated measures ANOVA performed on the log(CE)data shows a significant effect of phenomenal stateF(1 4) = 201 p G 05 and pedestal contrast F(4 16) =1449 p G 001 with no significant interaction betweenthe two F(4 16) = 01 p = 99 This lack of interactionimplies that the visible and suppressed phases yieldequivalent TvC slopes (ie exponents of the fitted powerfunction see below)Whether the target was perceived or not contrast
thresholds (CE) are well fit by power functions of thepedestal contrast CP (CE = kCCP
5 solid and dashedstraight lines in Figure 2 for visible and suppressedphenomenal states respectively) Table 1 displays theexponents 5 (slopes in the logndashlog plots of Figure 2) andthe gain (ie intercept) parameters as estimated from thedata when expressed as contrast kC and as luminance kLthresholds (see above) for each observer together withtheir means The mean 5 is close to unity (Weberrsquos law)which is substantially higher than the exponents obtainedfor spatially modulated gratings (eg Foley amp Legge1981 Gorea amp Sagi 2001 Henning amp Wichmann 2007Legge 1981 Legge amp Foley 1980 Nachmias amp Sansbury1974) but close to the exponents inferred for luminancediscrimination thresholds with spatially localized stimuli(eg Cornsweet amp Teller 1965 Hood amp Finkelstein1986)As suggested by the analysis above the gain parameter
(kC or kL) increases in the suppressed phase (by anaverage factor of 192 and 233 depending on whether itis computed for the contrast or luminance data formatsrespectively) whereas the 5 exponent is equivalent for the
Figure 2 Increment CE thresholds averaged over the fiveobservers (a) and for one of them (b) for visible (filled symbols)and suppressed targets (open symbols) as a function of pedestalcontrast Cpedestal Solid and dashed straight lines are best powerfunction fits for the visible and suppressed conditions respec-tively Vertical bars are T1 SE across observers (a) and over thethree repeats for one observer (b)
Observer
5 (TvL and TvC) kC (TvC) kL (TvL)
Visible Suppressed Visible Suppressed Visible Suppressed
FC 119 124 022 030 014 017CC 115 120 024 064 018 041PC 109 085 032 076 027 110DG 105 114 025 041 023 029MT 141 132 021 028 008 013M 117 T 006 115 T 008 025 T 002 048 T 010 018 T 003 042 T 018
Table 1 Exponents (5) and gains (kC kL) of the power functions fitted to the TvL andor TvC data formats (Figure 2) for visible andsuppressed phenomenal states
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 5
two phenomenal states A linear regression analysisperformed on the log(CE) data of all observers con-firmed the parallelism between the visible and suppressedTvC functions as the difference between their slopes is notsignificant F(1 6) = 0022 whereas their intercepts differsignificantly F(1 7) = 4456 p G 0001 Linear regressionanalyses performed on each observerrsquos data yield equiv-alent effects with the exception of observer MT whoseintercepts for the visible and suppressed states do notdiffer significantlyOverall the present results and statistical analyses point
to a global vertical shift of the TvC function during MIBand are in line with Watanabe et alrsquos (2004) binocularrivalry data As argued in the Introduction section thisshift is compatible with a form of divisive inhibitionoperating during MIB and occurring after the transducingstage whether this transducer is of the form of a logpower or NakandashRushton function (see also Footnote 1)
Experiment 2 Sensitivity (dV) and decisioncriteria (c)
To cite from Green and Swets (1966 p 108) Btheprincipal value of the forced-choice task is that it practi-cally eliminates the need to deal with the observerrsquosdecision criterion[ The main purpose of the present YesNo experiment was precisely not to eliminate thisparameter as our concern was to assess its relation to theobserverrsquos decisional behavior under visible and sup-pressed MIB states To do so we fixed the targetpedestalluminance at 312 cdm2 and assessed dV values anddecision criteria for four increment levels The experi-mental format allows the assessment of the psychometric
functions obtained under the two phenomenal states andmost critically the test of the conjecture that thesuppression phase in MIB reflects at least in part anupward decision-criterion shift with respect to the visiblephaseFigure 3 shows dVvalues averaged over the five subjects
(Figure 3a) and for one of them (Figure 3b) under target-visible (filled symbols) and target-suppressed (opensymbols) conditions as a function of the target incrementmagnitude (in contrast units C) Solid and dotted linesare power function fits (dV= k(C) Pelli 1985 for a dVderivation based on a NakandashRushton transducer type seeYu et al 2003) Observers show an overall sensitivitydrop in the suppressed phase by an average factor of 177(an average of 084 A) A two-way repeated measuresANOVA (with phenomenal state and C as factors)shows a significant effect for the two factors phenomenalstate F(1 4) = 709 p G 05 C F(3 12) = 1939p G 001 with no significant interaction between themF(3 12) = 084 p = 50 This lack of interaction impliesthat the visible and suppressed phases yield psychometricfunctions with similar slopes (see below)The average psychometric function fits yield slightly
larger exponentsslopes () but smaller elevations (k) for thesuppressed ( = 158 k = 3) than for the visible ( = 145k = 49) MIB states A linear regression analysis performedon the mean log(dV) indicates that these slopes are notsignificantly different F(1 4) = 015 p = 72 whereas theelevations are F(1 5) = 20037 p G 001 Equivalentanalyses performed on the slopes and elevations of thepsychometric functions for each observer yield similareffects except for observer PC whose elevations do notdiffer significantly This shift of the psychometric functionalong the ordinate without changes in slope is indicative ofa divisive reduction of signal strength rather than of anincrease in the internal noise The preservation of thepsychometric function slopes under visible and suppressedstates has also been observed for binocular rivalry(Sanders 1980 passim Watanabe et al 2004)The mean CE values derived from the fitted dV
functions (CEExp 2 = (dVk)1) for a dV = 114 (thetheoretical sensitivity at the detection threshold E asassessed with the 2AFC 1-upndash3-down staircase procedureof Experiment 1) are globally lower than those actuallymeasured in Experiment 1 with the 312-cdm2 pedestalfor both the visible (CEExp 1 = 055 CEExp 2 = 037)and the suppressed (CEExp 1 = 113 CEExp 2 = 059)MIB states These differences are substantially larger thanthose previously reported (Green amp Swets 1966 p 112)The only account we can offer is that the lower thresholdsderived from the YesNo data reflect a learning effect asthis experiment was run after the 2AFC one As the 2AFCexperiment was run with five intermingled pedestal levels(as opposed to the one pedestal level used in the YesNoexperiment) it is also possible that the 2AFC thresholdswere relatively increased due to this additional Bnoise[factor
Figure 3 dV averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimuluschange (C) for visible (full symbols) and suppressed targets(open symbols) Standard errors are not shown for the individualdata (as in Panel b) because individual dVvalues were computedfrom the 300 trials run by each observer
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 6
Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

were assessed in different sessions and repeated thriceThe order of the sessions (each specified by the pedestalluminance and phenomenal state) was randomized foreach repeat and observerIn Experiment 2 only one pedestal luminance (312 cdm2)
was used throughout and dVvalues together with decisioncriteria were assessed in separate sessions for fourincrement magnitudes by way of a YesNo procedurewith the increment randomly applied to either the lower orthe upper part of the bar in 50 of the trials The fourincrement amplitudes (124 137 150 and 162 dB) werechosen based on preliminary sessions so that the smallestand largest increments yield for all observers dVvaluesnot lower than 04 and not higher than 35 One session(specified by the increment magnitude and by theobserverrsquos phenomenal state) consisted of 100 trials andwas repeated thrice so that dVvalues and decision criteria(defined as c = 05(zH + zFA) where zH and zFA are thez scores of hits and false alarms2) were computed from300 trials per observer The order of the sessions wasrandomized for each observerIt should be noted that both the stimulation setup and
some aspects of the procedure were finalized after a longseries of preliminary experiments meant to maximize theduration and stability of the individual MIB durations andhence the reliability of the thresholddVmeasurementsOriginally these measurements were made with aBstandard[ MIB configuration where the rotating maskwas presented at the center of the screen and where thetarget (a 09- 09- cross) was presented 27- fromfixation in the upper left corner along the main diagonalThis configuration yielded relatively short MIB periods(G15 s) which together with a 100-ms delay between anobserverrsquos key press and the targetrsquos luminance incre-ment produced small and unstable threshold increments(compared to those assessed with visible targets)Postexperimental interviews revealed that in the target-suppressed perceptual states observers frequently expe-rienced premature target reappearance that is before orwithin milliseconds after the increment was appliedPremature target reappearances can be triggered both bythe abrupt luminance increment (Kawabe Yamada ampMiura in press) and by microsaccades set off by anobserverrsquos intention to press the key while focusing hisattention on the target location such premature targetreappearances are most likely favored under shallowsuppression states Hence the preliminary trial-and-errormanipulations were meant to find the stimulation param-eters that strengthen the suppression states In addition tothose preliminary observations indicated that the firsttarget disappearance within each trial tended to last lessand be more easily terminated by the test increment thansubsequent disappearances (presumably because of ashallower adaptation state) Hence observers were encour-aged to refrain from signaling it and await the second oreven the third disappearance in a trial These observations
are of importance as they point at the difficulty ofobtaining reliable threshold or dV assessments duringMIB and at the particular care needed to choose theoptimal stimulating and response recording conditions (fora comprehensive discussion of the topic see Kim ampBlake 2005)
Results
Observations on MIB phenomenology
The time lapse between the beginning of a trial and anobserverrsquos key press to signal a targetrsquos disappearance wasnot measured as observers were encouraged to ignore thefirst or even the first two disappearances (see the Methodssection) This instruction was based on our long-lastingexperience with MIB The duration of a disappearancetends to increase with inspection time and practice(Bonneh Sagi amp Cooperman 2005) suggesting theincremental contribution of an adaptation process Forone observer (out of five) the time lapse before the firstdisappearance was consistently longer (up to 1 min) thanusually reported Observers occasionally signaled partialtarget disappearances (ie of the upper or lower part ofthe pedestal) and were instructed to ignore them Allobservers signaled the reappearance of the suppressedpedestal as if triggered by the test increment when thisincrement was large (at the start of the staircase) but notwhen it was close to threshold Target reappearance wasalso reported on almost all occasions where observerswere aware of their inadvertent eye movements Twoobservers reported that their perception of the rotatingmask alternated between a state where its components(crosses) appeared as individualized items and a statewhere they were grouped to form a unitary rotatingsurface Interestingly they also reported an increasedlikelihood of a targetrsquos disappearance concomitant withthe transition to a latter state A potentially relatedphenomenon was reported by Graf et al (2002)
Experiment 1 TvC functions
The appropriate way of representing luminance incre-ment thresholds as a function of the pedestal luminanceLP has been a matter of debate (eg Kingdom ampMoulden 1991 Whittle 1986) For the present stimulusconfiguration where the reference pedestal was itselfdisplayed on a nonnull background L0 two data displayformats are equally legitimate whereby (a) the liminalluminance increment LE (= ordfLT j LPordf) is plotted as afunction of the pedestal luminance relative to the back-ground luminance that is LPrelative (= ordfLP j L0ordf) or
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 4
(b) the liminal contrast difference CE is plotted againstthe pedestal contrast (CP relative to the backgroundluminance TvC function) that is
CE frac14 jLT j L0jL0
jLP j L0
L0frac14 jLT j LPj
L0eth1aTHORN
and
CP frac14 jLP j L0jL0
eth1bTHORN
As it can be noted the contrast representation is but anormalized (by L0) version of the luminance representa-tion Inasmuch as the incremental threshold is a linearfunction of the pedestal magnitude in logndashlog coordinates(eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980 Watanabe et al 2004) the
two representations yield the same logndashlog slopes butdifferent intercepts For the sake of comparison withWatanabe et alrsquos (2004) equivalent study in binocularrivalry the data below are displayed in the second formatabove (TvC Equations 1a and 1b)Figure 2 displays TvC functions averaged over the five
observers (Figure 2a) and for one of them (Figure 2b)under target-visible (filled symbols) and target-suppressed(open symbols) conditions The mean suppressed-to-visibleCE ratio is 184 (026 log units) which is close to thatobtained in binocular rivalry (Watanabe et al 2004)3
A two-factor (pedestal contrast and phenomenal state)repeated measures ANOVA performed on the log(CE)data shows a significant effect of phenomenal stateF(1 4) = 201 p G 05 and pedestal contrast F(4 16) =1449 p G 001 with no significant interaction betweenthe two F(4 16) = 01 p = 99 This lack of interactionimplies that the visible and suppressed phases yieldequivalent TvC slopes (ie exponents of the fitted powerfunction see below)Whether the target was perceived or not contrast
thresholds (CE) are well fit by power functions of thepedestal contrast CP (CE = kCCP
5 solid and dashedstraight lines in Figure 2 for visible and suppressedphenomenal states respectively) Table 1 displays theexponents 5 (slopes in the logndashlog plots of Figure 2) andthe gain (ie intercept) parameters as estimated from thedata when expressed as contrast kC and as luminance kLthresholds (see above) for each observer together withtheir means The mean 5 is close to unity (Weberrsquos law)which is substantially higher than the exponents obtainedfor spatially modulated gratings (eg Foley amp Legge1981 Gorea amp Sagi 2001 Henning amp Wichmann 2007Legge 1981 Legge amp Foley 1980 Nachmias amp Sansbury1974) but close to the exponents inferred for luminancediscrimination thresholds with spatially localized stimuli(eg Cornsweet amp Teller 1965 Hood amp Finkelstein1986)As suggested by the analysis above the gain parameter
(kC or kL) increases in the suppressed phase (by anaverage factor of 192 and 233 depending on whether itis computed for the contrast or luminance data formatsrespectively) whereas the 5 exponent is equivalent for the
Figure 2 Increment CE thresholds averaged over the fiveobservers (a) and for one of them (b) for visible (filled symbols)and suppressed targets (open symbols) as a function of pedestalcontrast Cpedestal Solid and dashed straight lines are best powerfunction fits for the visible and suppressed conditions respec-tively Vertical bars are T1 SE across observers (a) and over thethree repeats for one observer (b)
Observer
5 (TvL and TvC) kC (TvC) kL (TvL)
Visible Suppressed Visible Suppressed Visible Suppressed
FC 119 124 022 030 014 017CC 115 120 024 064 018 041PC 109 085 032 076 027 110DG 105 114 025 041 023 029MT 141 132 021 028 008 013M 117 T 006 115 T 008 025 T 002 048 T 010 018 T 003 042 T 018
Table 1 Exponents (5) and gains (kC kL) of the power functions fitted to the TvL andor TvC data formats (Figure 2) for visible andsuppressed phenomenal states
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 5
two phenomenal states A linear regression analysisperformed on the log(CE) data of all observers con-firmed the parallelism between the visible and suppressedTvC functions as the difference between their slopes is notsignificant F(1 6) = 0022 whereas their intercepts differsignificantly F(1 7) = 4456 p G 0001 Linear regressionanalyses performed on each observerrsquos data yield equiv-alent effects with the exception of observer MT whoseintercepts for the visible and suppressed states do notdiffer significantlyOverall the present results and statistical analyses point
to a global vertical shift of the TvC function during MIBand are in line with Watanabe et alrsquos (2004) binocularrivalry data As argued in the Introduction section thisshift is compatible with a form of divisive inhibitionoperating during MIB and occurring after the transducingstage whether this transducer is of the form of a logpower or NakandashRushton function (see also Footnote 1)
Experiment 2 Sensitivity (dV) and decisioncriteria (c)
To cite from Green and Swets (1966 p 108) Btheprincipal value of the forced-choice task is that it practi-cally eliminates the need to deal with the observerrsquosdecision criterion[ The main purpose of the present YesNo experiment was precisely not to eliminate thisparameter as our concern was to assess its relation to theobserverrsquos decisional behavior under visible and sup-pressed MIB states To do so we fixed the targetpedestalluminance at 312 cdm2 and assessed dV values anddecision criteria for four increment levels The experi-mental format allows the assessment of the psychometric
functions obtained under the two phenomenal states andmost critically the test of the conjecture that thesuppression phase in MIB reflects at least in part anupward decision-criterion shift with respect to the visiblephaseFigure 3 shows dVvalues averaged over the five subjects
(Figure 3a) and for one of them (Figure 3b) under target-visible (filled symbols) and target-suppressed (opensymbols) conditions as a function of the target incrementmagnitude (in contrast units C) Solid and dotted linesare power function fits (dV= k(C) Pelli 1985 for a dVderivation based on a NakandashRushton transducer type seeYu et al 2003) Observers show an overall sensitivitydrop in the suppressed phase by an average factor of 177(an average of 084 A) A two-way repeated measuresANOVA (with phenomenal state and C as factors)shows a significant effect for the two factors phenomenalstate F(1 4) = 709 p G 05 C F(3 12) = 1939p G 001 with no significant interaction between themF(3 12) = 084 p = 50 This lack of interaction impliesthat the visible and suppressed phases yield psychometricfunctions with similar slopes (see below)The average psychometric function fits yield slightly
larger exponentsslopes () but smaller elevations (k) for thesuppressed ( = 158 k = 3) than for the visible ( = 145k = 49) MIB states A linear regression analysis performedon the mean log(dV) indicates that these slopes are notsignificantly different F(1 4) = 015 p = 72 whereas theelevations are F(1 5) = 20037 p G 001 Equivalentanalyses performed on the slopes and elevations of thepsychometric functions for each observer yield similareffects except for observer PC whose elevations do notdiffer significantly This shift of the psychometric functionalong the ordinate without changes in slope is indicative ofa divisive reduction of signal strength rather than of anincrease in the internal noise The preservation of thepsychometric function slopes under visible and suppressedstates has also been observed for binocular rivalry(Sanders 1980 passim Watanabe et al 2004)The mean CE values derived from the fitted dV
functions (CEExp 2 = (dVk)1) for a dV = 114 (thetheoretical sensitivity at the detection threshold E asassessed with the 2AFC 1-upndash3-down staircase procedureof Experiment 1) are globally lower than those actuallymeasured in Experiment 1 with the 312-cdm2 pedestalfor both the visible (CEExp 1 = 055 CEExp 2 = 037)and the suppressed (CEExp 1 = 113 CEExp 2 = 059)MIB states These differences are substantially larger thanthose previously reported (Green amp Swets 1966 p 112)The only account we can offer is that the lower thresholdsderived from the YesNo data reflect a learning effect asthis experiment was run after the 2AFC one As the 2AFCexperiment was run with five intermingled pedestal levels(as opposed to the one pedestal level used in the YesNoexperiment) it is also possible that the 2AFC thresholdswere relatively increased due to this additional Bnoise[factor
Figure 3 dV averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimuluschange (C) for visible (full symbols) and suppressed targets(open symbols) Standard errors are not shown for the individualdata (as in Panel b) because individual dVvalues were computedfrom the 300 trials run by each observer
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 6
Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12
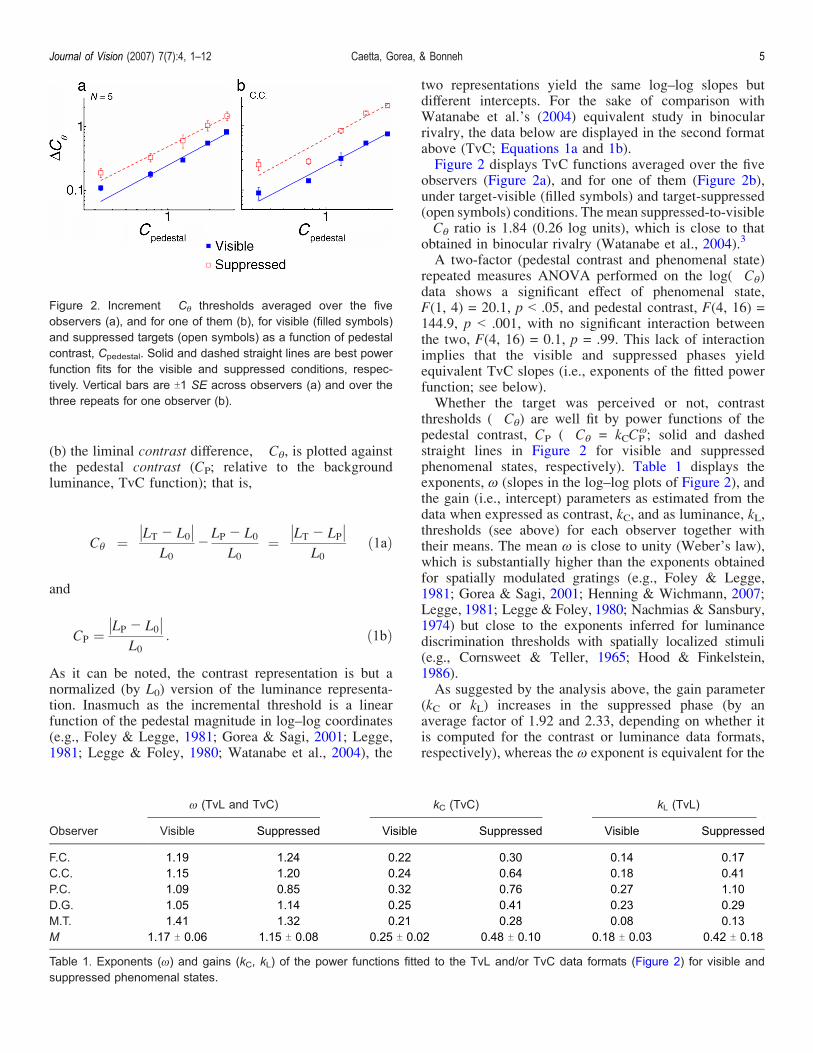
(b) the liminal contrast difference CE is plotted againstthe pedestal contrast (CP relative to the backgroundluminance TvC function) that is
CE frac14 jLT j L0jL0
jLP j L0
L0frac14 jLT j LPj
L0eth1aTHORN
and
CP frac14 jLP j L0jL0
eth1bTHORN
As it can be noted the contrast representation is but anormalized (by L0) version of the luminance representa-tion Inasmuch as the incremental threshold is a linearfunction of the pedestal magnitude in logndashlog coordinates(eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981 Legge amp Foley 1980 Watanabe et al 2004) the
two representations yield the same logndashlog slopes butdifferent intercepts For the sake of comparison withWatanabe et alrsquos (2004) equivalent study in binocularrivalry the data below are displayed in the second formatabove (TvC Equations 1a and 1b)Figure 2 displays TvC functions averaged over the five
observers (Figure 2a) and for one of them (Figure 2b)under target-visible (filled symbols) and target-suppressed(open symbols) conditions The mean suppressed-to-visibleCE ratio is 184 (026 log units) which is close to thatobtained in binocular rivalry (Watanabe et al 2004)3
A two-factor (pedestal contrast and phenomenal state)repeated measures ANOVA performed on the log(CE)data shows a significant effect of phenomenal stateF(1 4) = 201 p G 05 and pedestal contrast F(4 16) =1449 p G 001 with no significant interaction betweenthe two F(4 16) = 01 p = 99 This lack of interactionimplies that the visible and suppressed phases yieldequivalent TvC slopes (ie exponents of the fitted powerfunction see below)Whether the target was perceived or not contrast
thresholds (CE) are well fit by power functions of thepedestal contrast CP (CE = kCCP
5 solid and dashedstraight lines in Figure 2 for visible and suppressedphenomenal states respectively) Table 1 displays theexponents 5 (slopes in the logndashlog plots of Figure 2) andthe gain (ie intercept) parameters as estimated from thedata when expressed as contrast kC and as luminance kLthresholds (see above) for each observer together withtheir means The mean 5 is close to unity (Weberrsquos law)which is substantially higher than the exponents obtainedfor spatially modulated gratings (eg Foley amp Legge1981 Gorea amp Sagi 2001 Henning amp Wichmann 2007Legge 1981 Legge amp Foley 1980 Nachmias amp Sansbury1974) but close to the exponents inferred for luminancediscrimination thresholds with spatially localized stimuli(eg Cornsweet amp Teller 1965 Hood amp Finkelstein1986)As suggested by the analysis above the gain parameter
(kC or kL) increases in the suppressed phase (by anaverage factor of 192 and 233 depending on whether itis computed for the contrast or luminance data formatsrespectively) whereas the 5 exponent is equivalent for the
Figure 2 Increment CE thresholds averaged over the fiveobservers (a) and for one of them (b) for visible (filled symbols)and suppressed targets (open symbols) as a function of pedestalcontrast Cpedestal Solid and dashed straight lines are best powerfunction fits for the visible and suppressed conditions respec-tively Vertical bars are T1 SE across observers (a) and over thethree repeats for one observer (b)
Observer
5 (TvL and TvC) kC (TvC) kL (TvL)
Visible Suppressed Visible Suppressed Visible Suppressed
FC 119 124 022 030 014 017CC 115 120 024 064 018 041PC 109 085 032 076 027 110DG 105 114 025 041 023 029MT 141 132 021 028 008 013M 117 T 006 115 T 008 025 T 002 048 T 010 018 T 003 042 T 018
Table 1 Exponents (5) and gains (kC kL) of the power functions fitted to the TvL andor TvC data formats (Figure 2) for visible andsuppressed phenomenal states
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 5
two phenomenal states A linear regression analysisperformed on the log(CE) data of all observers con-firmed the parallelism between the visible and suppressedTvC functions as the difference between their slopes is notsignificant F(1 6) = 0022 whereas their intercepts differsignificantly F(1 7) = 4456 p G 0001 Linear regressionanalyses performed on each observerrsquos data yield equiv-alent effects with the exception of observer MT whoseintercepts for the visible and suppressed states do notdiffer significantlyOverall the present results and statistical analyses point
to a global vertical shift of the TvC function during MIBand are in line with Watanabe et alrsquos (2004) binocularrivalry data As argued in the Introduction section thisshift is compatible with a form of divisive inhibitionoperating during MIB and occurring after the transducingstage whether this transducer is of the form of a logpower or NakandashRushton function (see also Footnote 1)
Experiment 2 Sensitivity (dV) and decisioncriteria (c)
To cite from Green and Swets (1966 p 108) Btheprincipal value of the forced-choice task is that it practi-cally eliminates the need to deal with the observerrsquosdecision criterion[ The main purpose of the present YesNo experiment was precisely not to eliminate thisparameter as our concern was to assess its relation to theobserverrsquos decisional behavior under visible and sup-pressed MIB states To do so we fixed the targetpedestalluminance at 312 cdm2 and assessed dV values anddecision criteria for four increment levels The experi-mental format allows the assessment of the psychometric
functions obtained under the two phenomenal states andmost critically the test of the conjecture that thesuppression phase in MIB reflects at least in part anupward decision-criterion shift with respect to the visiblephaseFigure 3 shows dVvalues averaged over the five subjects
(Figure 3a) and for one of them (Figure 3b) under target-visible (filled symbols) and target-suppressed (opensymbols) conditions as a function of the target incrementmagnitude (in contrast units C) Solid and dotted linesare power function fits (dV= k(C) Pelli 1985 for a dVderivation based on a NakandashRushton transducer type seeYu et al 2003) Observers show an overall sensitivitydrop in the suppressed phase by an average factor of 177(an average of 084 A) A two-way repeated measuresANOVA (with phenomenal state and C as factors)shows a significant effect for the two factors phenomenalstate F(1 4) = 709 p G 05 C F(3 12) = 1939p G 001 with no significant interaction between themF(3 12) = 084 p = 50 This lack of interaction impliesthat the visible and suppressed phases yield psychometricfunctions with similar slopes (see below)The average psychometric function fits yield slightly
larger exponentsslopes () but smaller elevations (k) for thesuppressed ( = 158 k = 3) than for the visible ( = 145k = 49) MIB states A linear regression analysis performedon the mean log(dV) indicates that these slopes are notsignificantly different F(1 4) = 015 p = 72 whereas theelevations are F(1 5) = 20037 p G 001 Equivalentanalyses performed on the slopes and elevations of thepsychometric functions for each observer yield similareffects except for observer PC whose elevations do notdiffer significantly This shift of the psychometric functionalong the ordinate without changes in slope is indicative ofa divisive reduction of signal strength rather than of anincrease in the internal noise The preservation of thepsychometric function slopes under visible and suppressedstates has also been observed for binocular rivalry(Sanders 1980 passim Watanabe et al 2004)The mean CE values derived from the fitted dV
functions (CEExp 2 = (dVk)1) for a dV = 114 (thetheoretical sensitivity at the detection threshold E asassessed with the 2AFC 1-upndash3-down staircase procedureof Experiment 1) are globally lower than those actuallymeasured in Experiment 1 with the 312-cdm2 pedestalfor both the visible (CEExp 1 = 055 CEExp 2 = 037)and the suppressed (CEExp 1 = 113 CEExp 2 = 059)MIB states These differences are substantially larger thanthose previously reported (Green amp Swets 1966 p 112)The only account we can offer is that the lower thresholdsderived from the YesNo data reflect a learning effect asthis experiment was run after the 2AFC one As the 2AFCexperiment was run with five intermingled pedestal levels(as opposed to the one pedestal level used in the YesNoexperiment) it is also possible that the 2AFC thresholdswere relatively increased due to this additional Bnoise[factor
Figure 3 dV averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimuluschange (C) for visible (full symbols) and suppressed targets(open symbols) Standard errors are not shown for the individualdata (as in Panel b) because individual dVvalues were computedfrom the 300 trials run by each observer
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 6
Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

two phenomenal states A linear regression analysisperformed on the log(CE) data of all observers con-firmed the parallelism between the visible and suppressedTvC functions as the difference between their slopes is notsignificant F(1 6) = 0022 whereas their intercepts differsignificantly F(1 7) = 4456 p G 0001 Linear regressionanalyses performed on each observerrsquos data yield equiv-alent effects with the exception of observer MT whoseintercepts for the visible and suppressed states do notdiffer significantlyOverall the present results and statistical analyses point
to a global vertical shift of the TvC function during MIBand are in line with Watanabe et alrsquos (2004) binocularrivalry data As argued in the Introduction section thisshift is compatible with a form of divisive inhibitionoperating during MIB and occurring after the transducingstage whether this transducer is of the form of a logpower or NakandashRushton function (see also Footnote 1)
Experiment 2 Sensitivity (dV) and decisioncriteria (c)
To cite from Green and Swets (1966 p 108) Btheprincipal value of the forced-choice task is that it practi-cally eliminates the need to deal with the observerrsquosdecision criterion[ The main purpose of the present YesNo experiment was precisely not to eliminate thisparameter as our concern was to assess its relation to theobserverrsquos decisional behavior under visible and sup-pressed MIB states To do so we fixed the targetpedestalluminance at 312 cdm2 and assessed dV values anddecision criteria for four increment levels The experi-mental format allows the assessment of the psychometric
functions obtained under the two phenomenal states andmost critically the test of the conjecture that thesuppression phase in MIB reflects at least in part anupward decision-criterion shift with respect to the visiblephaseFigure 3 shows dVvalues averaged over the five subjects
(Figure 3a) and for one of them (Figure 3b) under target-visible (filled symbols) and target-suppressed (opensymbols) conditions as a function of the target incrementmagnitude (in contrast units C) Solid and dotted linesare power function fits (dV= k(C) Pelli 1985 for a dVderivation based on a NakandashRushton transducer type seeYu et al 2003) Observers show an overall sensitivitydrop in the suppressed phase by an average factor of 177(an average of 084 A) A two-way repeated measuresANOVA (with phenomenal state and C as factors)shows a significant effect for the two factors phenomenalstate F(1 4) = 709 p G 05 C F(3 12) = 1939p G 001 with no significant interaction between themF(3 12) = 084 p = 50 This lack of interaction impliesthat the visible and suppressed phases yield psychometricfunctions with similar slopes (see below)The average psychometric function fits yield slightly
larger exponentsslopes () but smaller elevations (k) for thesuppressed ( = 158 k = 3) than for the visible ( = 145k = 49) MIB states A linear regression analysis performedon the mean log(dV) indicates that these slopes are notsignificantly different F(1 4) = 015 p = 72 whereas theelevations are F(1 5) = 20037 p G 001 Equivalentanalyses performed on the slopes and elevations of thepsychometric functions for each observer yield similareffects except for observer PC whose elevations do notdiffer significantly This shift of the psychometric functionalong the ordinate without changes in slope is indicative ofa divisive reduction of signal strength rather than of anincrease in the internal noise The preservation of thepsychometric function slopes under visible and suppressedstates has also been observed for binocular rivalry(Sanders 1980 passim Watanabe et al 2004)The mean CE values derived from the fitted dV
functions (CEExp 2 = (dVk)1) for a dV = 114 (thetheoretical sensitivity at the detection threshold E asassessed with the 2AFC 1-upndash3-down staircase procedureof Experiment 1) are globally lower than those actuallymeasured in Experiment 1 with the 312-cdm2 pedestalfor both the visible (CEExp 1 = 055 CEExp 2 = 037)and the suppressed (CEExp 1 = 113 CEExp 2 = 059)MIB states These differences are substantially larger thanthose previously reported (Green amp Swets 1966 p 112)The only account we can offer is that the lower thresholdsderived from the YesNo data reflect a learning effect asthis experiment was run after the 2AFC one As the 2AFCexperiment was run with five intermingled pedestal levels(as opposed to the one pedestal level used in the YesNoexperiment) it is also possible that the 2AFC thresholdswere relatively increased due to this additional Bnoise[factor
Figure 3 dV averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimuluschange (C) for visible (full symbols) and suppressed targets(open symbols) Standard errors are not shown for the individualdata (as in Panel b) because individual dVvalues were computedfrom the 300 trials run by each observer
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 6
Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

Figure 4 displays criterion (c) values averaged overthe five observers (Figure 4a) and for one of them(Figure 4b) for visible and suppressed targets (filledand open symbols respectively) as a function of thetarget increment magnitude (C) The horizontal dottedline at c = 0 denotes the optimal behavior Under thevisible state as expected all observers but one (MTc = 49) use a criterion close to optimal (mean c T 1SE = 008 T 004) Under the suppressed state allobservers raise their criterion by an average of 068 A(mean c T 1 SE = 083 T 014) barely below the averagedVdrop (084 A) A two-way repeated measures ANOVAon the c data (with phenomenal state and C as factors)reveals a significant effect of the phenomenal stateF(1 4) = 1557 p G 05 but not of theC factor F(3 12) =049 p = 70 with no significant interaction between thetwo F(3 12) = 100 p = 42 The present data and analysisclearly demonstrate that MIB is associated with a con-servative decision behavior although they cannot specifywhether this behavior is one of its causes or a consequenceof the depleted sensitivity assessed in Experiments 1 and 2
Discussion
The present results indicate that MIB is associated withboth a sensitivity drop and a decisional criterion shift tohigher values In Experiment 1 the TvC functions for thesuppressed phase yielded an average upward shift (relativeto the nonsuppressed phase) of 026 log units (Figure 2)congruent with the results obtained in an equivalentbinocular rivalry study by Watanabe et al (2004) Thissensitivity drop is confirmed by the second experimentshowing that the psychometric functions assessed underthe suppressed phase are shifted downward compared tothe nonsuppressed phase without a change in slope
(Figure 3) This behavior suggests that the sensitivity lossobserved during MIB results from a divisive reduction ofthe signal strength (gain) rather than from an increase inthe internal noiseThe upward shift of the TvC function is compatible
with a posttransducer divisive inhibition (see the Intro-duction section and Footnote 1) As noted a while ago byDealy and Tolhurst (1974) and originally modeled byFoley (1994) and Foley and Chen (1997) such aninhibition may result from prolonged adaptation a processinvoked at least since Kohler (1940) to account forunstable perception phenomena (for a recent review seeBlake Sobel amp Gilroy 2003) At the same time thissame literature also posits a mutual targetndashdistracter short-lived inhibition (ie not due to prolonged inspection)reflecting the antagonism between competitive processes(for a review see Lankheet 2006) Although thiscompetition is readily understood for unstable phenomenasuch as binocular rivalry or Necker cube its substrateremains a matter of debate for the MIB phenomenon Anumber of studies suggest that the relevant competition inMIB occurs at high processing levels between incompat-ible perceptual representations of the static targets and themoving distracters (eg Funk amp Pettigrew 2003 Grafet al 2002 Kanai Paffen Gerbino amp Verstraten 2004)The possibility remains that the MIB target is transientlyinhibited at an earlier processing stage by the movingbackground (eg Breitmeyer Love amp Wepman 1974Gorea amp Fiorentini 1982 Kanai et al 2004 Yantis ampNakama 1998) via lateral interactions (BalochampGrossberg1997 Finkel amp Edelman 1989 Grossberg amp Rudd 1992Tononi Sporns amp Edelman 1992 Zeki amp Shipp 1988)andor feedback loops (Angelucci et al 2002 Felleman ampVan Essen 1991 Zeki amp Shipp 1988) Funk (cited byFunk amp Pettigrew 2003) actually showed that TMS of areaMT (hence disruption of motion processing) decreases theMIB effect Libedinsky and Livingstone (2006) proposedthat both low- and higher level suppression processes(Bdownstream of V1 and upstream of inferotemporalcortex[) are at work in MIB Be it as it may the MIBeffect is most likely contributed to by both long-lived (dueto adaptation as in the Troxler effect eg Hsu Yeh ampKramer 2006 Wilke Logothetis amp Leopold 2003) andshort-lived inhibition (Wilke et al 2003)Experiment 2 shows that in addition to a sensitivity
loss the suppression stage in MIB is also subtended by achange in observerrsquos decisional behavior Responsecriteria during this phase are about 068 A larger thanduring the visible phase (Figure 4) Whether this criterionchange is a (partial) cause of MIB or rather theconsequence of the related sensitivity loss remainsundecided The first alternative could be related toattentional variations triggering the suppressed phases inMIB (via a criterion shift see Gorea amp Sagi 2005) but isshort of a firm theoretical ground and requires furtherinvestigation The second alternative is reminiscent ofGorea and coll (Gorea amp Sagi 2000 2001 2002 2005
Figure 4 Criteria averaged over the five observers (a) with barsindicating T1 SE and for one of them (b) as a function of stimulusincrement (C) for the visible (solid symbols) and suppressedtargets (open symbols)
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 7
Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

Gorea Caetta amp Sagi 2005) findings that lower saliencysensory events are more frequently ignored when pre-sented together (whether simultaneously or in sequence)with more salient ones than without In the presentcontext the two saliency levels tally with the twophenomenal states In any event the decisional behaviorof observers presented with perceptually bistable stimula-tions has not been previously documented and deservesfurther examinationIn summary the present study points to the fact that the
MIB phenomenon is concomitant with a sensitivity drop(most likely due to a posttransducer divisive inhibition)as well as with a shift toward higher values of the decisioncriterion The present experimental paradigm does notallow deciding whether these changes are its (exhaustive)causes or consequences While the sensitivity drop is mostlikely a cause the fact that the criterion shift may well beone of its side effects is consistent with Gorea and Sagirsquos(2000 2001 2002 2005) observation that criterion shiftsresult from a saliency imbalance between simultaneous orintermingled sensory eventsThe question of whether or not the relatively mild
sensitivity changes presently observed (very similar tothose observed during binocular rivalry Watanabe et al2004) may by themselves account for the MIB remainsunsettled That this could be so is consistent with anumber of studies having demonstrated the perceptualsuppression (Kanai amp Kamitani 2003) or the perceptualalternation (Kanai Moradi Shimojo amp Verstraten 2005)of suprathreshold targets subsequent to the transientstimulation with more salient stimuli Kanai and Kamitani(2003) and more recently Simons et al (2006) alsodemonstrated that the brief removal or contrast reductionof a target causes its subsequent fading an indication thatoffset transients per se are sufficient to induce fading Thepoint made here is that in addition to a sustained targetadaptation process the sudden drop in the saliency of theMIB target (due to the sudden change in the transducerrsquosgain) could by itself result in its total suppression (egWilke et al 2003) The criterion shifts presently revealedwould reinforce and prolong the phenomenal disappear-ance of the target during MIB
Acknowledgments
We thank our observers who have patiently spent manyhours out of sheer altruism
Commercial relationships noneCorresponding author Andrei GoreaEmail AndreiGoreauniv-paris5frAddress Laboratoire de Psychologie de la Perception(LPP) Paris Descartes University amp CNRS (FRE 2929)45 rue des Saints POres 75006 Paris France
Footnotes
1
For Stevenrsquos law R Egrave C+ (with + typically below 1eg Foley amp Legge 1981 Gorea amp Sagi 2001 Legge1981) the derivative of R with respect to C is RC Egrave+C+j1 A divisive inhibition m (G1) applied after thetransducer yields R Egrave mC+ with RC Egrave m+C+j1 that isan upward threshold shift (C Egrave mj1+C1j+) The same istrue for an inhibited NakandashRushton transducer (R =mRmaxC
+(s+ + C+) where Rmax and s are the saturationand semisaturation constants respectively and where + isthe maximum steepness of the function but see Meese ampHolmes 2002 Yu et al 2003) as its derivative withrespect to C is m(pRmaxC
+j1s+(s+ + C+)2) so that C isagain proportional to mj1 The case of an inhibition beforethe transducer can be written as R Egrave (mC)+ log(mC) andRmax(mC)
+(s+ + (mC)+) for Stevens Fechner and NakandashRushton respectively The corresponding derivatives withrespect to C are (mC)1j+ 1C and (+Rmaxm
+C+j1s+)(s+ +m+C+)2 Hence a pretransducer inhibition should yield athreshold drop for Stevens (C Egrave (mC)1j+) an unaffectedthreshold for Fechner (C Egrave C) and a threshold that mayeither increase or drop for NakandashRushton depending onthe relationship between C and the free parameters s and +(the proof of the latter is beyond the scope of this studyfor a detailed discussion of these parameters see Meese ampHolmes 2002 and Yu et al 2003)
2
According to this notation (Green amp Swets 1966) anunbiased decision behavior yields c = 0 with negativeand positive values corresponding to Bliberal[ andBconservative[ behaviors respectively
3One of our reviewers was concerned with the fact that
the transition from the suppressed to the visible states mayhave occasionally occurred during the 100-ms presenta-tion of the test increment thereby reducing its effective-ness and hence accounting (at least partly) for the largerthresholds under the suppressed phase To answer thisconcern we have reduced the increment duration to 10 msand measured the suppressed and visible thresholds forone target contrast (128 ie 136 cdm2) For the twotested observers the suppressed thresholds were 193 and143 times larger than the visible ones an increasepractically identical to the one observed with 100-msincrement durations
References
Angelucci A Levitt J B Walton E J Hupe J MBullier J amp Lund J S (2002) Circuits for local andglobal signal integration in primary visual cortex Journalof Neuroscience 22 8633ndash8646 [PubMed] [Article]
Azzopardi P amp Cowey A (1997) Is blindsight likenormal near-threshold vision Proceedings of the
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 8
National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

National Academy of Sciences of the United States ofAmerica 94 14190ndash14194 [PubMed] [Article]
Baloch A A amp Grossberg S (1997) A neural model ofhigh-level motion processing Line motion and for-motion dynamics Vision Research 37 3037ndash3059[PubMed]
Bender M B (1952) Disorders in perception Spring-field IL Thomas
Blake R (1989) A neural theory of binocular rivalryPsychological Review 96 145ndash167 [PubMed]
Blake R amp Camisa J (1979) On the inhibitory natureof binocular rivalry suppression Journal of Experi-mental Psychology Human Perception and Perfor-mance 5 315ndash323 [PubMed]
Blake R amp Fox R (1974) Adaptation to invisiblegratings and the site of binocular rivalry suppressionNature 249 488ndash490 [PubMed]
Blake R Sobel K V amp Gilroy L A (2003) Visualmotion retards alternations between conflicting per-ceptual interpretations Neuron 39 869ndash878[PubMed] [Article]
Blake R Tadin D Sobel K V Raissian T A ampChong S C (2006) Strength of early visual adapta-tion depends on visual awareness Proceedings of theNational Academy of Sciences of the United States ofAmerica 103 4783ndash4788 [PubMed] [Article]
Bonneh Y S Cooperman A amp Sagi D (2001)Motion-induced blindness in normal observersNature 411 798ndash801 [PubMed]
Bonneh Y S Sagi D amp Cooperman A (2005)Learning to ignore Practice can increase disappear-ance in motion induced blindness [Abstract] Journalof Vision 5(8)223 223a httpjournalofvisionorg58223 doi10116758223
Breitmeyer B Love R amp Wepman B (1974) Contoursuppression during stroboscopic motion and meta-contrast Vision Research 14 1451ndash1456 [PubMed]
Breitmeyer B G (1984) Visual masking An integrativeapproach New York Oxford University Press
Breitmeyer B G amp Ogmen H (2000) Recent modelsand findings in visual backward masking A compar-ison review and update Perception amp Psychophy-sics 62 1572ndash1595 [PubMed]
Campion J Latto R amp Smith Y M (1983) Isblindsight an effect of scattered light spared cortexand near-threshold vision Behavioral and BrainSciences 6 423ndash486
Clarke F J J amp Belcher S J (1962) On thelocalization of Troxlerrsquos effect in the visual pathway53ndash68
Cornsweet T N amp Teller D Y (1965) Relation ofincrement thresholds to brightness and luminance
Journal of the Optical Society of America 551303ndash1308 [PubMed]
Critchley M (1949) The phenomenon of tactile inatten-tion with special reference to parietal lesions Brain72 538ndash561
Dealy R S amp Tolhurst D J (1974) Is spatialadaptation an after-effect of prolonged inhibitionJournal of Physiology 241 261ndash270 [PubMed][Article]
Dennett D C (1991) Consciousness explained Bostonamp New York Little Brown
Fechner G T (1907) Elemente der Psychophysik I u IILeipzig Breitkopf amp Hartel
Felleman D J amp Van Essen D C (1991) Distributedhierarchical processing in the primate cerebral cortexCerebral Cortex 1 1ndash47 [PubMed] [Article]
Finkel L H amp Edelman G M (1989) Integration ofdistributed cortical systems by reentry A computersimulation of interactive functionally segregated vis-ual areas Journal of Neuroscience 9 3188ndash3208[PubMed] [Article]
Foley J M (1994) Human luminance pattern-visionmechanisms Masking experiments require a newmodel Journal of the Optical Society of America AOptics image science and vision 11 1710ndash1719[PubMed]
Foley J M amp Chen C C (1997) Analysis of the effectof pattern adaptation on pattern pedestal effects Atwo-process model Vision Research 37 2779ndash2788[PubMed]
Foley J M amp Legge G E (1981) Contrast detectionand near threshold discrimination in human visionVision Research 21 1041ndash1053 [PubMed]
Fox R amp Check R (1972) Independence betweenbinocular rivalry suppression duration and magnitudeof suppression Journal of Experimental Psychology93 283ndash289 [PubMed]
Funk A P amp Pettigrew J D (2003) Does interhemi-spheric competition mediate motion-induced blind-ness A transcranial magnetic stimulation studyPerception 32 1328ndash1338 [PubMed]
Goodale M A amp Milner A D (1992) Separate visualpathways for perception and action Trends in Neuro-sciences 15 20ndash25 [PubMed]
Gorea A Caetta F amp Sagi D (2005) Criteriainteractions across visual attributes Vision Research45 2523ndash2532 [PubMed]
Gorea A amp Fiorentini A (1982) Interactions orienta-tionnelles entre des rOseaux stationnaires et enmouvement Lrsquo AnnOe Psychologique 82 45ndash65
Gorea A amp Sagi D (2000) Failure to handle more thanone internal representation in visual detection tasks
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 9
Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

Proceedings of the National Academy of Sciences ofthe United States of America 97 12380ndash12384[PubMed] [Article]
Gorea A amp Sagi D (2001) Disentangling signal fromnoise in visual contrast discrimination Nature Neuro-science 4 1146ndash1150 [PubMed] [Article]
Gorea A amp Sagi D (2002) Natural extinction Acriterion shift phenomenon Visual Cognition 9913ndash936
Gorea A amp Sagi D (2005) On decision and attentionIn L Itti G Rees amp J K Tsotsos (Eds) Neuro-biology of attention (pp 152ndash159) San Diego CAElsevier
Graf E W Adams W J amp Lages M (2002)Modulating motion-induced blindness with depthordering and surface completion Vision Research42 2731ndash2735 [PubMed]
Green D M amp Swets J A (1966) Signal detectiontheory New York Wiley
Grossberg S amp Rudd M E (1992) Cortical dynamicsof visual motion perception Short-range and long-range apparent motion Psychological Review 9978ndash121 [PubMed]
Halligan P W amp Marshall J C (1993) The history andclinical presentation of neglect In I H Robertson ampJ C Marshall (Eds) Unilateral neglect clinical andexperimental studies (pp 3ndash26) Hove UK LawrenceErlbaum Associates
Henning G B amp Wichmann F A (2007) Someobservations on the pedestal effect Journal of Vision7(1)3 1ndash15 httpjournalofvisionorg713doi101167713 [PubMed] [Article]
Hofstoetter C Koch C amp Kiper D C (2004) Motion-induced blindness does not affect the formation ofnegative afterimages Consciousness and Cognition13 691ndash708 [PubMed]
Hood D C amp Finkelstein M A (1986) Sensitivity tolight In K R Boff L Kaufman amp J P Thomas(Eds) Handbook of perception and human per-formance Vol I Sensory processes and perception(pp 53ndash56) New York Academic Press
Hsu L C Yeh S L amp Kramer P (2004) Linkingmotion-induced blindness to perceptual filling-inVision Research 44 2857ndash2866 [PubMed]
Hsu L C Yeh S L amp Kramer P (2006) A commonmechanism for perceptual filling-in and motion-induced blindness Vision Research 46 1973ndash1981[PubMed]
Kanai R amp Kamitani Y (2003) Time-locked perceptualfading induced by visual transients Journal ofCognitive Neuroscience 15 664ndash672 [PubMed]
Kanai R Moradi F Shimojo S amp Verstraten F A(2005) Perceptual alternation induced by visualtransients Perception 34 803ndash822 [PubMed]
Kanai R Paffen C L Gerbino W amp Verstraten F A(2004) Blindness to inconsistent local signals inmotion transparency from oscillating dots VisionResearch 44 2207ndash2212 [PubMed]
Kanwisher N G (1987) Repetition blindness Typerecognition without token individuation Cognition27 117ndash143 [PubMed]
Kastner S De Weerd P Desimone R amp UngerleiderL G (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functionalMRI Science 282 108ndash111 [PubMed]
Kawabe T Yamada Y amp Miura K (in press) How anabrupt onset cue can release motion-induced blind-ness Consciousness and Cognition [PubMed]
Kim C Y amp Blake R (2005) Psychophysical magicRendering the visible Finvisible_ Trends in CognitiveSciences 9 381ndash388 [PubMed]
Kingdom F amp Moulden B (1991) A model for contrastdiscrimination with incremental and decremental testpatches Vision Research 31 851ndash858 [PubMed]
Klein S A (1998) Double-judgment psychophysicsfor research on consciousness Application toblindsight In S A Hemeroff A W Kaszniak ampA C Scott (Eds) Toward a science of conscious-ness II The second Tucson discussions and debates(pp 361ndash369) Cambridge MA MIT Press
Klotz W amp Neumann O (1999) Motor activationwithout conscious discrimination in metacontrastmasking Journal of Experimental PsychologyHuman Perception and Performance 25 976ndash992
Kohler I W (1940) Dynamics in psychology New YorkLiveright
Lankheet M J (2006) Unraveling adaptation and mutualinhibition in perceptual rivalry Journal of Vision 6(4)1 304ndash310 httpjournalofvisionorg641doi101167641 [PubMed] [Article]
Legge G E (1981) A power law for contrast discrim-ination Vision Research 21 457ndash467 [PubMed]
Legge G E amp Foley J M (1980) Contrast masking inhuman vision Journal of the Optical Society ofAmerica 70 1458ndash1470 [PubMed]
Levelt W J M (1965) On binocular rivalry Assen TheNetherlands Royal VanGorcum
Levitt H (1971) Transformed upndashdown methods inpsychoacoustics Journal of the Acoustical Societyof America 49 467ndash477 [PubMed]
Libedinsky C amp Livingstone M (2006) Multi-levelsuppression during motion-induced blindness
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 10
[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

[Abstract] Journal of Vision 6(6)301 301a httpjournalofvisionorg66301 doi10116766301
Mack A amp Rock I (1998) Inattentional blindnessCambridge MA MIT Press
Macmillan N A amp Creelman C D (1991) Detectiontheory A userrsquos guide New York CambridgeUniversity Press
McGinnies E (1949) Emotionality and perceptualdefense Psychological Review 56 244ndash251
Meese T S (2004) Area summation andmasking Journalof Vision 4(10)8 930ndash943 httpjournalofvisionorg4108 doi1011674108 [PubMed] [Article]
Meese T S amp Holmes D J (2002) Adaptation and gainpool summation Alternative models and maskingdata Vision Research 42 1113ndash1125 [PubMed]
Montaser-Kouhsari L Moradi F Zandvakili A ampEsteky H (2004) Orientation-selective adaptationduring motion-induced blindness Perception 33249ndash254 [PubMed]
Mueller T J amp Blake R (1989) A fresh look at thetemporal dynamics of binocular rivalry BiologicalCybernetics 61 223ndash232 [PubMed]
Nachmias J amp Sansbury R V (1974) Letter Gratingcontrast Discrimination may be better than detectionVision Research 14 1039ndash1042 [PubMed]
Naka K I amp Rushton W A (1966) S-potentials fromluminosity units in the retina of fish (Cyprinidae)Journal of Physiology 185 587ndash599 [PubMed][Article]
Nguyen V A Freeman A W amp Wenderoth P (2001)The depth and selectivity of suppression in binocularrivalry Perception amp Psychophysics 63 348ndash360[PubMed]
Ogmen H Breitmeyer B G amp Melvin R (2003) Thewhat and where in visual masking Vision Research43 1337ndash1350 [PubMed]
Pelli D G (1985) Uncertainty explains many aspects ofvisual contrast detection and discrimination Journalof the Optical Society of America A Optics and imagescience 2 1508ndash1532 [PubMed]
Raymond J E Shapiro K L amp Arnell K M (1992)Temporary suppression of visual processing in anRSVP task An attentional blink Journal of Exper-imental Psychology Human Perception and Perform-ance 18 849ndash860 [PubMed]
Rensink R A OrsquoRegan J K amp Clark J J (1997)To see or not to see The need for attention toperceive changes in scenes Psychological Science8 368ndash373
Ricci R amp Chatterjee A (2004) Sensory and responsecontributions to visual awareness in extinctionExperimental Brain Research 157 85ndash93 [PubMed]
Sanders R K (1980) Binocular suppression duringbinocular rivalry and fusion Unpublished doctoraldissertation University of Rochester
Simons D Lleras A Martinez-Conde S Slichter DCaddigan E amp Nevarez G (2006) Induced visualfading of complex images Journal of Vision 6(10)91093ndash1101 httpjournalofvisionorg6109doi1011676109 [PubMed] [Article]
Stevens S S (1957) On the psychophysical lawPsychological Review 64 153ndash181 [PubMed]
Stigler R (1910) Chronophotische Studien uber denUmgebungskontrast [Effects of exposure duration andluminance on the contrast of the surround] PflugersArchiv fur die Gesamte Physiologie des Menschenund der Tiere 134 365ndash435
Tong F (2001) Competing theories of binocular rivalryA possible resolution Brain and Mind 2 55ndash83
Tononi G Sporns O amp Edelman G M (1992)Reentry and the problem of integrating multiplecortical areas Simulation of dynamic integration inthe visual system Cerebral Cortex 2 310ndash335[PubMed]
Trevarthen C B (1968) Two mechanisms of vision inprimates Psychologische Forschung 31 299ndash348[PubMed]
Troxler D (1804) Uber das Verschwindern gegebenerGegenstande innerhalb unsers Gesichtskrcises InK Himley amp J A Schmidt (Eds)Ophthalmologischesbibliothek (vol II pp 51ndash53) Jena Fromann
Wales R amp Fox R (1970) Increment detection thresh-olds during binocular rivalry suppression Perceptionamp Psychophysics 8 90ndash94
Watanabe K Paik Y amp Blake R (2004) Preservedgain control for luminance contrast during binocularrivalry suppression Vision Research 44 3065ndash3071[PubMed]
Weiskrantz L (1986) Blindsight A case study andimplications Oxford UK Oxford University Press
Weiskrantz L (1990) Outlooks for blindsight Explicitmethods for implicit processes Proceedings of theRoyal Society of London B Bological Sciences 239247ndash278 [PubMed]
Wheatstone C (1838) Contributions to the physiology ofvisionVPart the first On some remarkable andhitherto unobserved phenomena of binocular visionPhilosophical Transactions of the Royal Society ofLondon 128 371ndash394
Whittle P (1986) Increments and decrements Luminancediscrimination Vision Research 26 1677ndash1691[PubMed]
Wilke M Logothetis N K amp Leopold D A (2003)Generalized flash suppression of salient visual targetsNeuron 39 1043ndash1052 [PubMed] [Article]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 11
Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12

Yantis S amp Nakama T (1998) Visual interactions inthe path of apparent motion Nature Neuroscience 1508ndash512 [PubMed]
Yu C Klein S A amp Levi D M (2003) Cross- and iso-oriented surrounds modulate the contrast response
function The effect of surround contrast Journal ofVision 3(8)1 527ndash540 httpjournalofvisionorg381 doi101167381 [PubMed] [Article]
Zeki S amp Shipp S (1988) The functional logic of corticalconnections Nature 335 311ndash317 [PubMed]
Journal of Vision (2007) 7(7)4 1ndash12 Caetta Gorea amp Bonneh 12




















