Hydroxylated phosphines as ligands for chalcogenide clusters

J. Mol. Biol. (1991) 222, 301-310
Selection of Antibody Ligands from a Large Library of Oligopeptides Expressed on a Multivalent
Exposition Vector
Franc0 Felici
Istituto di Ricerche di Biologia Molecolare (IRBM) via Pontina Km 30.600
00040 Pomexia (Roma), Italy
Luisa Castagnoli, Andrea Musacchiot, Roberto Jappelli and Gianni Cesarenit
Dipartimento di Biologia, II Universitd di Roma Tor Vergata, Via Carnevale, 00173 Roma, Italy
(Received 28 May 1991; accepted 26 July 1991)
Practically any oligopeptide can be exposed on the surface of the bacteriophage capsid by fusion to the major coat protein of filamentous bacteriophages. A phage expressing a particular peptide tag can be selected from a mixture of tens of millions of clones, exposing oligopeptides of random sequence, by affinity purification with a protein ligand. In this respect, pVII1 can be used as an alternative and complement to the exposition vectors based on the product of gene III (~111). We have constructed a phagemid vector that) contains gene VIII under the control of the pLac promoter. This vector can be conveniently used to construct libraries of oligopeptides with a random amino acid sequence. An anti- peptide monoclonal antibody was used to affinity-purify phagemids exposing oligopeptides which can interact with the monoclonal antibody. DNA sequencing of the amino terminus of gene VIII of the recovered clones predicts the synthesis of hybrid proteins whose amino- terminal amino acid sequence is related to that of the oligopeptide used to raise the antibody. In other words, only oligopeptides that bind a very small portion of the immunoglobulin G surface are affinity-purified by this method, implying that the antigen binding site possesses molecular properties that renders it much stickier than the remainder of the molecule.
Keywords: filamentous phages; major capsid protein; ILlb; peptide library
1. Introduction
The possibility of selecting ligands from large libraries of peptides with a random amino acid sequence has recently raised considerable interest. Any peptide domain, free to interact with the solvent, can, in principle, be used as a carrier to present oligopeptides to be probed for binding to specific ligands. If the carrier peptide (exposition vector) is physically linked to its gene, affinity purification of the hybrid protein will result in the selection and cloning of its coding sequence.
This goal was achieved by the development of expression-exposition vectors that exploit the
t Present address: E.M.B.L. Postfach 10.2209, 6900 Heidelberg, Germany.
2 Author to whom all correspondence should be addressed.
-
properties of a phage coat protein, the product of gene III (~111) of the filamentous phage fd (Smith, 1985; Parmley & Smith, 1988). The pII1 protein is present in five copies at one end of the phage filament (for a review on filamentous phages, see Rasched & Oberer, 1986; Model & Russel, 1988). Since pII1 is required for phage adsorption, some peptide insertions that disturb the conformation of the vector protein may result in phage structures that are not infective. However, addition of short peptides to the amino terminus does not affect phage infectivity (Parmley & Smith, 1988). The potential of this approach for the selection of ligands from a large collection of peptides of random sequence has been demonstrated (Cwirla et al., 1990; Devlin et al., 1990; Scott & Smith, 1990).
In principle, any exposition vector can be used for this purpose. For instance, a variety of outer- membrane proteins, or proteins that form the
0022-2836/91/220301-10 $03.00/O 301
0 1991 Academic Press Limited

302 I”. F&i et al
building blocks of bacterial thread-like organelles, are capable of presenting oligopeptides. inserted into specific regions of their structures, to the exterior of the bacterial cell (Charbit et al., 1988; Agterberg et al., 1987; Freudl, 1989; Hedegard & Klemm, 1989; Wu et al.. 1989; Namba et al.. 1989). Ctilizing phages instead of bacteria. however. raises the number of clones that, can be handled in a single experiment by a fact’or of 103, and facilitates the construction and storage of large peptide libraries.
We have explored a different protein vector system t,hat, in principle. should offer some further advantages and opportunities. This is based on the same filamentous bacteriophage utilized by Smith (1986), but exploits a different, protein. the producst of gene VIII (pVIII), which is the major capsid protein of phage fl. The mature protein is 50 amino acid residues long and caan be divided into thrw domains on the basis of amino acid composition. Acidic amino acids predominate at t,hr amino terminus. while the carboxy terminus (which inter- acts with the single-stranded chromosome) is (Aharacterized by four Iysine residues. The cnent,ral part of the protein (residues 21 to 39) is hydra- phobic. pVII1 is initially synthesized as a precursor with an N-terminal extension of 23 amino acid residues. which, after having contributed t’o inserting the protein into the cytoplasmica membrane, is processed by the enzyme leader pepti- dase. (ienet’ic and biochemical st’udies have demorl- &rated that the translocation process is independent from the .sec machinery and requires both t#he amino-terminal extension and the hydro- phobic central region (Kuhn et al.. 1986). PositiveIS charged residues at t,he amino terminus of the leader pept,idr and at the carboxy terminus of the mature protein are also required for translocat’ion (Greenwood bz Perham, 1989; Gallusser 8 Kuhn. 1990).
I)uring the process of phage assembly. approxi- mate& 2700 pVJTT subunits aggregate, /*ia a poorly understood mechanism. to form a helical array around the chromosomal DSA core. Since t,he amino terminus of mature pVII1 is exposed to the solvent. as suggested by structural studies (Marvin. 1990) and demonstrated by the observation that it clonstitutes its antigenic site, we reasoned that an! oligopeptide a,ddition near the N t,erminus of t,hr mature prot,ein, which would not) interfere with translocation across the membrane and/or phage assembly, should result in exposition of the oligo- peptide on the phage capsid.
We describe here how a nonapeptide that forms a p-turn in the interleukin Ifi (ILlfi) protein can be exposed on the surface of filamentous bacterio- phages by inserting its coding sequence after the third codon of the mature pVII1 protein. We have also developed a phagemid vector to facilitate the construction of peptide libraries and shown that we are able t,o select antibody ligands from the library by “biopanning” techniques (Parmley & Smith. 1988).
This expression-exposition system shares most of
t,he desirable caharacteristics of those hased on pl I I and, moreover, should add the further advantages of a simpler and better characterized protein vector that polymerizes at high densit,y.
2. Materials and Methods
Strains used in this work WIW 5 l/IX AIIac-prol th,i
F’[luc1* IncZI~M 15 pro/l I,+] s/rpE (Messing rt ~1.. 1977) and W76 A[/ac-pro] WY, thi F’[Zncl* Inc%AMl. proilH+ 1 .sup*. XL-blue (recd2 ~nd.4 I y?yr.‘LSfi thi h,sdRlT (rK-mK+) supB44 rrl.4 1 inc- F’[~~ro.1/1 Iact* 1ncZ AM15 TnlO] was used for thtb caonstruc+on of the oligopept,id~~ library. Ph. age and reeomhinant, I)XA trcahniques were essentially as described by Sambrook Pf ~1. (1989). Site direct’ed mutagenesis was performed acachording to Kunkrl (1985).
The vrrtor PC%!) t,hat, was used to insert c)ligonu(.lro- tides of random sequence in the 5’-part of the gene VT11 coding sequence was c*onstructed through t,hr following steps. Illitially. the 481 nuclrotide long SnuHl-Hsrrcl I)NA fragment of phagr f’l c*ontaining t,he entire gene VI II and the amino-terminal port,ion of gene HI. was treated with the Klenow subunit of the Dh’A polyrnerase to rrmo\-r the 2 nucleotide 3’ extension of t,he Usml-generat,ed end. purified by low melting agaroae gel electrophoresis and inserted into t,he SmccI site of plQW31,19~t (Dente rt ~1.. 1985). (‘Iones containing the insert, in an orientation that puts gtm~ \‘TTT under pLac control (PC82) form blur, colonies on X-gal? indicator plates since the cr-pept,ide coding sequence is fused in-frame to the nuc~leotide sequence encoding t,he amino-terminal part of gene TTT. In order to construct a plasmid where &galac~tosidase activity would be associat,ed to expression of gene VITI. we removed the DNA sequen(Bp encoding gene ITT and ~VV joined the gene VI11 caoding sequence to the r-peptide coding sequence. This was achieved by &r-directed delr- t’ion utilizing an oligonuc*leotide of sequence
5’ T.-\AAA(‘(:A(‘(:(:(‘(‘i\(:(!TAC:(’TT(:C“I’TT(’(:X 3’
as a primer and PC’82 as single-st,ralrtlrd template. Thus oligonuc~leotidr introduces an am her rodon between the 2 genes. The plasmid cont,aining the desired deletion (P(W) czan therefore be dist,inpuishrd from t’hr parent plasmid since. in sup” hackground. it does not synthesize the cr-pept,ide. However. a .su?~E host cont,aining PC83 firrms blue colonies on X-gal plates. To improve the versatility of the vector’ we removed the polylinker restriction sites left in front of gene VIII hv rutting with Bnm,HI and HindTTl, removal of the 5’ &gle-strsnded termini with mung bean nuclease and recircularization of t)he plasmid in the presence of phage T4 DNA ligase. The resulting plasmid (P(%4). however. s,vnthesized only a pVIII- related protein that could neither be processed nor trans- located across the cytoplasmic membrane. Analysis of t,hr DNA sequence in the proximitv of the deletion suggested that the protein synthesized ‘is probably a t’ranslation product that’ starts at the r-peptide AT(:, and then proceeds t)o translate the in-frame sequence emoding the
t LAbbreviations used: S-gal, .i-brolno-4-c.hloro- 3-indolgl /?-n-galact,oside: TPTG. isopropyl- I-thio- fi-n-galactoside: MAb. monoclonal antibody: Ip. immunoglobulin.

Multivalent Oligopeptide Exposition Vector 303
leader peptide of pVII1 up to the amber codon that separates gene VIII from the a-peptide gene. Xo product corresponding to a translation start at the gene VIII STG could be observed, probably because of a negative inter- ference of the in-frame upstream a-peptide ATG. To over- come this problem we eliminated the a-peptide ATG by site-directed mutagenesis utilizing the oligonucleotide
5’ TGGCGTAATCTCTAGAATAGCTAGATCTTGTGT- GAAATT 3’.
This oligonucleotide introduces unique XbaI and BglII sites in the plasmid (PC84). In order to facilitate the insertion of oligonucleotides in the sequence that codes for the amino terminus of gene VIII: we modified PC84 to introduce an EcoRI and a BarnHI site by site-directed mutagenesis with the oligonucleotide
5 TGCGGGATCCTAGAATTCACCCTCAGC 3’.
Since the 2 new unique restriction sites flank an in-frame amber oodon, the colonies formed by supE strains harbouring this plasmid (PC89) synthesize lower amounts of cc-peptide. as can be recognized by the intensity of the colony colour on indicator plates. This feature offers a ronvenient screening for oligonucleotide insertions that remove the stop codon. Since we have observed that the fl replication origin, present in this family of plasmids, interferes with phage growth and prevents complemen- tation test’s whenever necessary, the fl replication origin was removed by deleting the LVdeI-NaeI DNA fragment. PC89 deleted of the fl origin fragment was named PC108
Fag18 was constructed by hybridizing oligonucleotide
5’ r\ATT(‘eTT(:A(:(:G;IT(:AAGAI1A(:TAAA(~TAACGATAAA 3’
and oligonucleotide
5’ GATCTTTATCGTTACTTTCTTCACCCTGAACG 3’
and ligating them t’o the PC89 plasmid digested with enzymes Ir:coRI and RamHI. The ligation mix was used to transform 71/18 and the colonies formed by cells containing the plasmid with the correct’ insertion were recognized from the intensity of their blue colour on X-gal indicator plates. fag19 was obtained by site-directed mutagenesis of plasmid PC82 utilizing a 56-mer of sequence
.5’ AGTCAAAGGCCGCTTTATCGTTGGATTCTTCAC- (‘CTGAACTGCGGGATCGTCACCC 3’
to introduce t’he ILI~,,,.,,, oligopeptide between residue 7 and 8 of the mature pVII1.
(c) (‘onstruction of an epitope library
The strategy utilized to insert random nucleotide sequences into the region encoding the amino terminus of pVII1 is illustrated in Fig. 1.
In order to synthesize a double-stranded oligonucleo- tide containing a degenerate nucleotide sequence flanked by an EcoRI and a BumHI restriction site, 200 pmol of ohgonucleotide R35 (5’ GCGGGATCCN,,GAATTCACC- CG 3’) were annealed with 200 pmol of oligonucleotide R36 (5’ CGGGTGAATTC 3’) and the complementary strand was synthesized by extension of the 3’ end of R36 with 50 units of the Klenow subunit of DNA polymerase I (Boehringer). The double-stranded 46-mer was digested with EcoRI and BumHI restriction endonucleases and the 32-nucleotide central fragment’ was purified by electro- phoresis on a 2046 (w/v) polyacrylamide gel and sub- sequent’ Sephadex G50 gel filtration.
After EcoRI and RamHI digestion of the plasmid vect’or pC89, the short internal fragment was eliminated
R36 CGGGTGAATTC
Hybrkdization
5' GCGGGATCC-P'GAATTCACCCG 3' :zGmQ3L3333
Incubate with Klenow
5' GCGGGATCC NNNNNNNNNNNNNNNNNNNN GAATTCACCCG 3 3LLW363333
5'GCGG GATCC.P.G AATTCACCCG 3' 3um 3&3333
Insert into vector
r
TTAA GluPhe
GATC
ASpP
r,\ \\\% KS bla
Figure 1. Schematic representation of the strategy utilized to insert random oligonucleotide sequences into the region encoding pVII1. S in the nucleotide sequence indicates that, in t’he corresponding position. an equi- molar mixture of the 4 nucleotide precursors was utilized during oligonucleotide synthesis.
by isopropanol/sodium perchlorate precipitation. Then 1 pg (64 pmol) of vector was ligated with I pmol of double-stranded oligonucleotide at 15°C overnight. The ligation mix was heated at 65°C for 5 min, purified by gel filtration through a 1 ml Sephadex G50 column in water. dried and resuspended in 24 ~1 of water. Transformation was performed by electroporation using a Gene Pulser apparatus (BioRad) at 2.5 kV, 200 Ohm, 25 PF (average time constant’ 48 ms). Portions (2 ~1) of ligation mix were mixed with 80 ~1 of XLl-blue cells (recA 1 endA gyrA96
thi hsdRl7 (rK- mK+) mpE44 relA1 luc- F’(proAB lacIq 1ucZ AM15 TnlO) prepared according tjo Dower et al.
(1988). The transformed cells were immediately resuspended in
a total of 24 ml of SOC medium. were grown for 1 h at 37°C and spread on 48 Petri dishes (15 cm diam.) containing LB agar plus 100 pg ampicillin/ml. The same cell suspension was also titrated on L-agar plates containing 100 pg ampicillin/ml, 16 pg IPTG/ml and 32 pg X-gal/ml to evaluate the total number of independent clones present in the epitope library (approx. 9.4~ 10’ clones) and the percentage of clones forming blur colonies (approx. 40%).
After overnight incubation the colonies were scraped from the plates and resuspended in 48 ml of phage buffer plus 15% (v/v) glycerol. A portion (26 ml) of this bac- terial suspension was stored in samples at - 70 “C, 20 ml were used to prepare plasmid DNA and the remaining 2 ml were added to 2 1 of LB plus 50 pg ampicillin/ml and superinfected with Fl IRl helper phage in order to obtain

304 F. Felici et al
the phage supernatant (approx. 1 x 10” transducing units/ml) (Dent? et al.. 1985). The phages were precipi- tated twice with polyethylene glycol and purified by equilibrium centrifugation in C&l. according to Scot,t & Smith (1990). The resulting phage suspension contained approximately 3 x 10” ampicillin-resistance transducing units/ml. The recipies for the media are from Sambrook et al. (1989).
(d) Library screening
The library was screened by the biopanning technique according to Parmley Br Smith (1988) using a mouse MAb obtained by immunization with an oligopeptide of sequence VQGEESNDK (gift from D. Boraschi) and a secondary biotin conjugated anti-mouse IgC (whole mole- cule) antlbody (Sigma. B-7264). The concentration of thr MAb was 1 pM in the 1st and the 2nd round of biopanning and ti 1 nM in the 3rd round. The 2nd round of biopanninp was preceded by a pre-absorption step of the panned library on the secondary antibody.
The reconstitution experiment to prove that fag18 could expose the IL1/?163.,71 peptide on the phage capsid was performed in the same way, using MAb concentra- tions of 1 PM or 20 nM in different experiments; the yield of M13K07 did not change (average 3 x 10e6 recovered/ panned phages), while the average yield of fag18 was 3 x lo-* using a 1 pM concentration of MAb and 5 x 1O-4 using a 20 nM concentration of MAb in the biopanning procedure.
(e) Electrophoresis and immunostaining
Phage proteins were analysed on a discontinuous SDSj polyacrylamide gel electrophoresis system according to Shggger & von Jagow (1987) using a concentration of 165 y0 T and 3 y0 C as separating gels. This corresponds to 182i, (w/v) acrylamide and 0.5:/, (w/v) bis-acrylamide. Electrophoretically separated proteins were transferred onto a Hybond-ECL membrane (Amersham), using producer instructions. Phage particles were separated by electrophoresis on agarose gels according to Boeke et al. (1980) and transferred onto Hybond-ECL membranes using a 25 m&I-Tris (pH 8.6), 200 mM-glycine, 20% (v/v) methanol solution. A Pharmacia LKB 2117-250 Novablot apparatus was used for the transfer procedure. The transfer conditions were 1 h at 400 mA for protein gels and 1.5 h at 200 mA for phage agarose gels.
The primary mouse MAb (anti-VQGEESNDK peptide) was used at a final concentration of 1 pg/ml. The secondary peroxidase-conjugated anti-mouse TgG (heavy and light chain specific) IgG fraction (USB, 12338 Rabbit) was used at the final dilution of 1:50,000. The rabbit anti-F1 serum (obtained by injection in rabbit of CsCl-purified wild-type fl phage particles) was used at a final dilution of 1:2000 and the secondary peroxidase conjugated anti-rabbit IgG (heavy and light chain specific) fraction (USB, 1251 Goat) was used at a final dilution of 1: 18,000.
The immuno-blotting detection system used was the ECL (Amersham, RPN 2106).
3. Results As a first step in developing a pVII1 expression-
exposition system we wanted to show that the amino acid sequence of the amino terminus of the mature pVII1 does not contain any essential
123456789
wt AlaGluGlyAspAspProAlaLysAla
I I PM28 AlaGluGlyGluAspProAlaLysAla
I \ PM34 AlaGluGlyGluPheGlnAspProAlaLysAla
I I PM34.r AlaGluGlyGluPheSerAspProAlaLysAla
Figure 2. Amino acid sequence of the amino terminub of the wild-type (wt) JJVITI protein and of 3 mutants. PM28 and PM34 were constructed b>r site-directed muta- genesis using 2 oligonucleotides of sequence’
and 5’ T(:(‘(:(:(:,~T(‘(‘T(‘A(:(1(‘T(‘A 3
5’ T(:~I~:(:GATC’(‘TAGAA’I’T(‘A(:(‘(’T(’8~:(’ 3’.
respectively. Amino acid changes are indicated in it&es. The Asp to Glu substitution at position 4 introduces B HamHT site. while the insertion in PM34 corresponds to a BamHI and an EcoRT site flanking an in-frame ITAG stop czodon. Thus. the sequence represent,s the amino witi sequence of the pVTI1 protein synthesized in a .s~pE strain. PM34.r is a spontaneous revertant (TA(: to T(X) of the stop codon in PM34.
information for protein t’ranslocation and phage assembly. RJ site-directed mutagenesis we constructed protein variant’s in which the amino acid sequence of the amino t,erminus is changed either by amino acid substitutions or by insertions of a dipeptide. The codons used to create t#hese mutations were selected with the idea of intro- ducing, at the same time. unique sites recognized by the enzymes SORT and RamHI. These sites will later be used for inserting oligonucleotides. Since viable phages carrying the mutations described in Figure 2 could be constructed. wp concluded that the amino acid sequence at the amino t)erminus of mature pVIII is not, absolutely required either for translocation across the membrane (in accordancr with the observations of Kuhn et nl., 1990) or for phage assembly. This result is consistent wit!h the observation that the amino terminus of the pVTIJ proteins of relat,ed phages is not> conserved (Marvin. 1990).
However, an attempt to use the t’l p\‘llI protein to expose an octapeptide was not successful. 15’e rationalized this failure making the hypothesis that coat proteins carrying oligopeptide insertions cannot form a bact~eriophage capsid. The same insertions, however, might be incorporated at lower density on the phage surface int,erspersed with wild- type coat proteins. The possibility to generate hybrid phages using an extra coat gene cloned in a plasmid was demonstrated by Rowitch et al. (1988).
To test this hypothesis, we constructed a pMB1 derived plasmid (PC108) containing the fl gene VIII under the control of the pI,ac promot,er. In t,he presence of t’he inducer IPTG. this plasmid is able to complement a filamentous phage carrying an amber mutation in gene VIII indicating t’hat “physio- logical” amounts of pVIIJ are synthesized. The

Multivalent Oligopeptide Exposition Vector 305
Figure 3. Genetic map of the phagemid PC89. The figures in parentheses indicate the nucleotide position of the corresponding restriction enzyme sites. The nucleotide sequence of the insertion site and the corresponding amino acid sequence is illustrated above the plasmid map. The wild-type amino acid sequence of the amino terminus of mature pVII1 has been modified to introduce unique EcoRI and BarnHI sites (see PM34 in Pig. 2). The carboxy-terminal amino acid sequence of the leader peptide is in italics. bp, base-pairs.
phagemid PC88 has a genetic structure similar to PC108 with the addition of the fl replication origin. In this plasmid family the gene encoding pVII1 is fused (in frame) with the gene encoding the a-peptide of /%galactosidase via an amber codon. This feature allows us to associate the expression of pVII1, in an amber suppressor strain, to the forma- tion of blue colonies on X-gal indicator plates. In a su- background, however, translation termination occurs after the pVII1 coding sequence and only wild-type pVII1 is synthesized.
Tn order to facilitate the detection of clones containing oligonucleotide insertions in gene VIII, we constructed a derivative of PC88 by inserting into the DNA sequence encoding the amino terminus of the coat protein an EcoRI and a BamHI site flanking an amber codon. su+ strains containing this plasmid (PC89) do not efficiently suppress the two amber codons (the first between the 2 cloning sites and the second immediately preceding the cr-peptide coding sequence) and form white (or pale blue) colonies on /I-galactosidase indicator plates. This offers a convenient way to detect insertions of oligonucleotides that do not contain termination
fag19
wt NH 2 AlaGluGlyAapAspProAlaLyalUalUaPheAsp---
A\ fag18 GluPheValGlnGlyGluGluSerAsnAspLys
Figure 4. The Figure presents the amino acid sequence of the amino terminus of the wild-type (wt) pVTII protein of fl (bold letters). Above and below are the amino acid sequences of the peptide insertions that are synthesized by the phagemids fag19 and fagl8. The insertion points in the wild-type sequence are also indicated. fag18 contains 2 extra residues (GluPhe, encoded by the EcoRI site) at the amino terminus of ILlfl,,,~,,,.
codons since su+ strains containing such plasmids will now form blue colonies. The structure of plasmid PC89 is illustrated in Figure 3.
(a) Exposition of the ILl/?163.171 oligopeptide on the phage capsid
In order to prove that the major capsid protein can tolerate insertions of oligopeptides. we have chosen the nonapeptide of amino acid sequence VQGEESNDK (IL1b163-1,1) as a model system. This sequence corresponds to the oligopeptide span- ning a p-turn in the human interleukin l/l structure from amino acid 163 to 171. TL1fi163.171 was selected because a monoclonal antibody raised against this oligopeptide is available (Boraschi et al., 1989) and because we have shown that its antigeni- city does not depend heavily on the protein context in which it is inserted (our unpublished results). An oligonucleotide encoding the IL1/?,63.171 oligopep- tide was inserted, in the correct reading frame. either between the EcoRI and the BamHI sites (fag18) or approximately ten nucleotides further downstream (fagl9; see Fig. 4).
The corresponding plasmids should direct the synthesis of two modified coat proteins containing an insertion of the oligopeptide ILl/?,,,.,,, either after the third or the seventh amino acid of the mature wild-type protein (see Fig. 4 for the exact protein sequence). Neither of these plasmids, or their derivatives deleted in the fl replication origin, can complement a phage defective in gene VIII, which supports the initial hypothesis that the ILl~,6s.1,,/pVIII hybrid proteins cannot substi- tute the wild-type coat protein. To strengthen this conclusion, we tested eight clones containing inser- tions of nonapeptides of random amino acid sequences for their ability to complement the gene VIII defect. The result was negative in all cases. It remains possible, however, that in bacteria synthe- sizing both wild-type and mutant proteins, hybrid capsids containing a mixture of the two coat pro- teins are formed.
To test this idea, bacteria harbouring the two plasmids, or a control plasmid (PCSS) expressing the
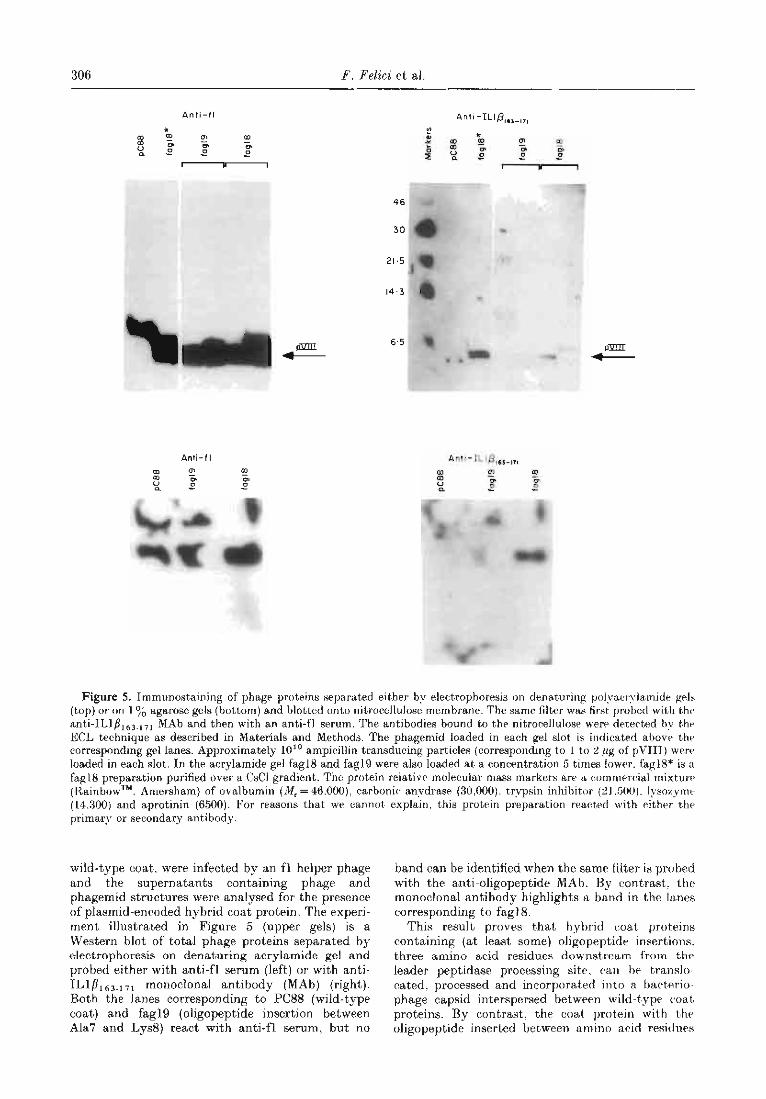
306 F. Felici et al
Anti-f I Anti-ILIP,,,..,,,
Figure 5. Immunostaining of phage proteins separated either by electrophoresis on denaturing polyacrylamide gels (t,op) or on 176 agarose gels (bottom) and blotted onto nitrocellulose membrane. The same filter was first probed with thr anti-IL1p163.17, MAb and then with an anti-f] serum. The antibodies bound to the nitrocellulose were detected by t’hr ECL technique as described in Materials and Methods. The phagemid loaded in each gel slot is indicated abovr the corresponding gel lanes. Approximately 10” ampicillin transducing particles (corresponding to 1 to 2 pg of pVII1) were loaded in each slot.. In the acrylamide gel fag18 and fag19 were also loaded at a concentration 5 times lower. fag18* is a fag18 preparation purified over a C&l gradient,. The protein relative molecular mass markers are a comrnrr~ial mixtuna (Rainbow . , rTM Amersham) of ovalbumin (A!!, = 46.000). carbonic anvdrase (30,000). trypsin inhibitor (21.500). Iysozymr (14.300) and aprotinin (6500). For reasons that we cannot explatn, this protein preparation reart)ed with either the primary or secondary antibody.
wild-type coat. were infected by an fl helper phage and t’he supernatants containing phage and phagemid structures were analysed for the presence of plasmid-encoded hybrid coat protein. The experi- ment illustrated in Figure 5 (upper gels) is a Western blot of total phage proteins separated by electrophoresis on denaturing acrylamide gel and probed either with anti-f] serum (left) or with anti- ILlP163.171 monoclonal antibody (MAb) (right). Both the lanes corresponding to PC88 (wild-type coat) and fag19 (oligopeptide insertion between Ala7 and Lys8) react with anti-f1 serum, but no
band can be identified when the same filter is probed with the anti-oligopeptide MAb. By cont’rast. the monoclonal antibody highlights a band in the lanes corresponding to fag1 8.
This result proves that hybrid coat, proteins containing (at least some) oligopeptide insertions: three amino acid residues downstream from t,he leader peptidase processing site, can he translo- cated. processed and incorporated into a bactrrio- phage capsid interspersed between wild-type coat, proteins. By contrast. the coat protein wit’h t’he oligopeptide inserted between amino acid residues

Multivalent Oligopeptide Exposition Vector 307
Anti-f I An+l-~@163-171 I 2 345678 12345678
Figure 6. Immunostaining of phages spotted on nitrocellulose membrane and probed either with an anti-f1 serum or with the anti-ILlj,,,.,,, MAb. Approximately 10” phage particles were spotted on the filter. Positive clones indicated in Tabk 1 are. respectively: b6 and c2 (p8IL-7); c4 and d6 (p8IL-10): e5 (p8IL-17); b2 (p8IL-4); e4 (p8TL-16): a4. a& ~8. 6% and e7 @ML-1).
Ala7 and Lys8 (fapl9) cannot be identiiied in phape particles. Pulse-chase experiments with 35S-labelled methionine indicate that the hybrid protein is either defective in t)he translocation process or is not a good substrate for the enzyme leader peptidase. since it’ is accumulated in the precursor form (not shown).
The oligopeptide exposed on the phage capsid is in a form available for interaction with the mono- clonal antibody. In fact, after electrophoresis of whole phages on agarose gel and transfer to nitro- cellulose membrane. a phage band can be high- lighted by the IL1/?163.171 antibody in the fag18 lane (Fig. 5. lower gels).
This conclusion is reinforced by an experiment where we demonstrate that phagemids encoding t’he hybrid ILlj3163.17,-p VIII protein can be enriched up to 3000.fold in a biopanning-type experiment over a background of Ml3K07 phages by affinity purification with the anti-oligopeptide MAb. In a control experiment where ,M13K07 was added to a mixture of PGWderived plasmids encoding pVII1 proteins with amino-terminal extensions of nona- peptides of random amino acid sequence. no corre- sponding enrichment was observed.
These experiments show that oligopeptides fused to t’he amino terminus of pVII1 can be incorporated into phagr capsids and can be recognized b.y cognate antibodies. This propert? can be exploited to select phages exposing the ohgopeptide out of a large background of wild-type phage structures with an efficiency comparable to that obtained by exposing oligopeptides on pTIT. The enrichment factor is sufficiently high to allow one to select, by a few rounds of: biopanning, specific ligands present in a library of random oligopeptides at a frequency as low as 10-s or 1W9.
(b) C’onstruction of a peptide library
PC89 is a convenient vector to construct large libraries of random oligopeptides. Its small size and high copy number facilitates preparation of clean vector UNA and permits high yields of transfor- mants/pg of DNA. The presence of two unique restriction sit,es flanking a DNA segment containing
an in-frame amber mutation offers a convenient colour phenotype to evaluate insertion efficiency. To prepare a library of nonapeptides of random amino acid sequences. PC89 DNA linearized by digestion with the enzymes EcoRI and BarnHI was recircular- ized by ligation to a double-stranded oligonucleo- tide having EcoRI and BamHI compatible termini flanking 26 nucleotides of completely degenerate sequence. Transformation yielded 9.4 x IO’ indepen- dent tsansformants. 40°6 of which formed blue colonies, indicating that they contained an oligo- nucleotide insertion without any in-frame nonsense codon.
=\s a test experiment to probe the representative- ness of the library and our ability to select peptide ligands, we performed three rounds of biopanning- type experiments, essentially as described by Scott $ Smith (1990), utilizing the anti-lLlfi,,3.171 MAb for the affinit’y purification. Thirty-eight of the
Table 1 Clones selected with anti-lllfi 163.1 71 from a library
of random oligopeptides inserted into the amino terminus of p VIII
Insertionst Isolat,es$ Spot4 M.P.II
fag18 AEGEF VQGEEDK D pXIL-7 AEGEF RDLHmVV D pXIL-10 AEGEF DKSmSIA D p8IL-17 AEGEF SNDGVWAIP D p8IL-4 AEGEF LFDREmW 1) p8IL-16 AEGEFYTDmRTTD p&311,-2 AEGEF YRDWHQP r) pXILI.5 AEGEF *TFDYTHAS D
+ 1 2 +++ 18 2 +++ 9 I ++ 4 I ++ 4 I + 1 :i + I 1 - < lW3
t Amino acid sequence of the insertions. as deduced from the DNA sequence of the DNA encoding the amino terminus of pVII1. p8IL-15 is a clone containing an ochre codon after F in position .5 and is used as a negative control. The tripeptide that is homologous in the positive clones is underlined.
$ Number of independent isolates in t,he biopanning experiment.
0 Semi-quantitative evaluation of the intensity uf immunostaining of spots of 10” phage particles on a nitrocellulose membrane (see Fig. 6).
jl Figures represent the ratios between the enrichment factor of the corresponding phagemid and that of faglll.

308 F. Felici et al.
clones that survived the third panning were spotted on nitrocellulose membrane and probed with t-he antiCILl/?,,,.,,, MAb. Twelve of these clones reacted with the antibody to varying degrees (Fig. 6). The same clones could be enriched preferen- tially in reconstitution experiments in which the particles containing the hybrid coat proteins were mixed with wild-type particles and then affinity- purified with anti-ILlfi,,3-171 MAb. The pVTT1 genes of the 12 positive clones were sequenced and grouped into six classes of different nucleotide sequences corresponding to six insertions of different nonapeptides (Table 1). The only common feature that could be identified in the amino acid sequence of the six oligopeptides is a tripeptide of sequence SX(D/E), where SND containing oligopep- tides seem to bind more eficiently than the two containing SNE. SND is also present in the original IIJ1P163.171 peptide.
4. Discussion
We have shown that the major capsid protein of filamentous phages, the product of gene VIII, can be exploited to expose oligopeptides on bacterio- phage capsids. pVII1 proteins carrying an amino- terminal extension of nine amino acids, in all the cases tested so far. cannot complement a defect in gene VIII and completely replace the wild-type protein. However, synthesis in the same cell of wild- type and extended pVII1 results in the formation of hybrid phage particles that expose the foreign oligo- peptide in a form available for interaction with an antibody.
We have developed a vector system based on a phagemid that synthesizes pVII1 under the control of the promoter pLac. In this construction, gene \:I11 is fused to the gene encoding the a-peptide of P-galactosidase l/is an amber codon. Furthermore. gene VIII was modified to introduce an EcoRI and a HnmHI site flanking a second in-frame amber codon into t,he sequence encoding the amino terminus of the mature protein. The presence of two amber codons between the translation start and the z-peptide offers a convenient colour screening to identify colonies that contain plasmids with inser- tions of oligonucleotides devoid of any in-frame stop codon.
make the separation of wild-type and hybrid (*oat proteins very difficult in a variet’y of gel systems we tested. Among a collection of random insertions. only a minor fraction of clones (15%) synthesized a hybrid protein that could be easily separated from the wild-type protein. By pulse-chase experiments in the absence of superinfection by wild-type phage, it is possible t,o demonstrate that the remaining clones do synthesize a protein that is eficientl~ translocated and processed to yield a peptide whoscl migration is indistinguishable from that of the mature wild-type coat protein. In the hybrid fag18 phage particles, for instance, no extra band can bc visualized by silver staining of phagr prot,eins separated by electrophoresis on denaturing acryl- amide gels. lmmunostaining by ant,i-Il,l~,h3.171 M,4b reveals that the hybrid protein carrying thr amino-terminal extension of nine amino acid residues co-migrates with the wild-tvpe (*oat pro- t’ein. Furthermore. of the six dikerent) clont~s selected by biopanning from t’he pept,idr library. only three show an ext,ra coat band after elec*tro- phorrtic separation. although they are all able to expose t,he hybrid protein. as demonst,rated by their ability t’o interact with the anti-TLl flih3., 7, MAb. These findings make evaluating the proportion ot random oligopeptCde that clan be ckxposed on the
phage surface a difficult task. However. the characterization of insertions caontaining at least on? C”ys (no (‘ys is present in the pVI1 I sequence). but with an otherwise random sequence. and the idnnti- fication of the ext)ended proteins with ( ‘ys-specifk reagents. has led to the conclusion that, six out of six insertions can br ident,ified on phagfx particles, indi- rating that they art’ eficient,l,v t~ranslocat~c~tl. processed and assembled (A. Luzzago, personal communication). This implies that t’here are ver) few. if any. limit!ations on t,ht> oligopeptidr arnirlo acid sequence that, cxan be exposed by this system, A rough est,imat,e, by Coomassie staining or by im- munostaining. of the ratio between wild-t,ype and hybrid (aoat’ prot)eins incorporated into phage particles suggests t,hat this number is between 30 and 300. with some variation among the different clones.
By modulating the activity of the Lac promoter and by superinfecting with a wild-type phage it should be possible to vary the ratio of wild-type and modified pVTTT, synthesized on the phagemid, and construct chimeric phages with different densities of a specific oligopeptide on their surface. With the bacterium-phagemid system described here, this is not easily done, as we have observed that substan- t,ial amounts of pVIII are synthesized even in the absence of inducer. Addition of IPTG raises pVIIT expression only by a factor of 2. By increasing the lac repressor gene copy number (for instance. by including the gene on the plasmid) it should be possible to obtain a tighter control.
We have shown that the vector FY189 can RF efficiently used to construc*t large oliyopept’ide libraries and to select ligands. Wr have used t,hr biopanning technique described by Parmley &Z Smith (1988) to select phagemids that expose oligo- peptides able to bind a monoclonal antibody raised against’ the oligopeptide VQGEESN DK. After three rounds of biopanning, six classes of clones c>ach exposing a different oligopeptide were eharac%er- ized. Some of t,hem seem t)o havca a mu& higher avidity for the MAb than the peptide exposed on fag1 8 t,hat exactly corresponds to the onp used to raise the antibody. It, is not clear at the moment whether this is due to a higher intrinsic affinit! between the oligopeptide and thr MAb. or to it higher density of the oligopeptide on the, phagt surface.
The unusual electrophoretic properties of pVII1 Surprisingly, all six classes of hybrid olonrs had

Multivalent Oligopeptide Exposition Vector 309
oligopeptide insertions characterized by amino acid sequences related to that of the oligopeptide used to raise the antibody. Namely, they all contained the tripeptide SND (or SNE). This finding has three implications: (1) all the clones that were selected carry oligopeptides that interact with the same binding site in the MAb; (2) this binding site corresponds to the ,MAb antigen binding site; (3) the SND “solution” of making a complementary surface to this site is strongly preferred to any other solution. We find it particularly surprising and interesting that the biopanning technique only selects clones that bind a very small portion of the surface of the TgG target molecule, suggesting that the antigen binding site possesses molecular proper- ties that make it much stickier than any other region of the molecule. Similar results were obtained by Scott & Smith (1990) and by Cwirla et al. (1990) after characterization of a larger number of clones selected for binding to two different MAbs from similar peptide libraries constructed by fusion to the pTTI protein. Whatever its molecular explanation. this phenomenon makes epitope mapping by screening of oligopeptide libraries on filamentous phages a feasible technique, since in all three experi- ments the selected clones were clearly related to the immunizing antigen. Whether the technology can be easily extended to extract ligands specific for molecular surfaces that had not been selected for binding an oligopeptide remains to be seen. The results of Devlin et al. (1990) that reported the selection of peptides able to bind streptavidin is encouraging in this respect.
In conclusion, we have shown that pVTI1 can be used in a manner analogous to the described pIIT vector system to expose oligopeptides on a filamen- tous phage capsid, and can therefore be exploited as an alternative and a complement to pITI. The com- bination of the two could lead to the construction of bivalent phages with one ligand on pTTT and the second on pVII1. While future work, with differ- ently designed libraries. will help in comparing the characteristics and relative merits of the two systems, we can see at least two advantages offered by the combination of a phagemid vector expressing a hybrid pVII1. The smaller dimensions and the higher copy number of the phagemid replicon facili- tat’es and increases t’he efficiency of all the opera- tions required to construct and characterize the oligopept*ide library. Furthermore, a vector based on pVTTT offers the possibility of varying, over a wide range (1 to 2700), the number and, as a conse- quence. the density of the oligopeptide on the phage capsid surface.
The practical applications of this approach are evident. A set of clones that show affinity for a serum against a pathogen could be used for diagnos- tic purposes. Clones that are selected for binding with a hormone receptor should be tested for their ability to simulate or to compete with hormone activity, while a subset of the clones that interact with an enzyme might behave as inhibitors of enzyme activity.
After submission of this paper, our attention was drawn to three recent works that describe related work. Il’ichev et al. (1989) first reported the exposi- tion of an oligopeptide (5 amino acid residues long) on the capsid of bacteriophage Ml3 by fusion to the amino terminus of pVII1. Greenwood et al. (1991) demonstrated that small peptides (up to 6 amino acid residues long) can be exposed by fusion to the pVII1 gene on the phage chromosome, while hybrid coat proteins carrying longer peptides can only be packaged by dispersing them at lower density in an otherwise wild-type capsid. This is in accord with our findings. Finally, Kang et al. (1991) succeeded in using a phagemid system, similar to the one that we have described, to construct hybrid phage capsids exposing an immunoglobulin Fab fragment fused to pVTI1.
F.F. started this work at the Department of Biology. University of Tor Vergata, supported by a grant of the E.C. RAP program to G.C. Phage PM28 was constructed by Helmer Citterich M. This work was partially supported by P.F. Ingegneria Genetica CXR to G.C. and by P.F. Biotecnologie e Biostrumentazione to L.C. We are grate- ful to G. P. Smith for providing detailed biopanning protocols. We thank D. Boraschi for t,he generous gift of the anti-ILlfl,,,.,,, MAb.
References Agterberg, M.. Adriaanse. H. & Tommassen, <J. (1987).
Use of outer membrane protein PhoE as a carrier for the transport of a foreign antigenic determinant to the cell surface of Escherichia coli K-12. (:ene, 58. 145-150.
Boeke, J. D.. Russel, M. & Model, P. (1980). Processing of filamentous phage precoat protein. Effect of sequence variations near the signal peptidase cleavage site. J. ,voZ. Riol. 144. 103-116.
Boraschi. D., Volpini, G., Villa, L., Xencioni, L,., Scapigliati. G.. Nucci, D., Antoni, G.. Matteucci, G., Cioli. F. & Tagliabue. A. (1989). A monoclonal anti- body to the IL-lb peptide 163-171 blocks adjuvancy but not pyrogenicity of IL-la in &Y). J. Immunol. 143. 131-134.
Charbit. *I., Molla. A.. Saurin. A. & Hofnung. MM. (1988). Versatility of a vector for expressing foreign poly- peptides at the surface of Gram-negative bacteria. Gene, 70. 181-189.
Cwirla. S. E.. Peters, E. A., Barret. R. W’. $ Dower, W. ,J. (1990). Peptides on phage: a vast library of peptides for identifving ligands. Proc. Nat. Acad. Sri., C.S.A. 87, 6378&82.
Dente. L.. Sollazzo. M.. Baldari. C.. Cesareni. G. 8: Cortese. R. (1985). The pEMBL family of single- st’randed vectors, In DNA Clon%ng: A Practical Approach (Glover. D. M., ed.). vol. I, pp. 101-107. IRL Press. Oxford.
Devlin. J. J.. Panganiban, L. C. t Devlin, P. (1990). Random peptide libraries: a source of specific protein binding molecules. Science, 249, 404-406.
Dower, W. J.. Miller, J. F. & Ragsdale. C. W. (1988). High efficiency transformation of E. coli by high voltage electroporation. ,Vucl. Acids Res. 16, 61276145.
Freudl, R. (1989). Insertion of peptides into cell-surface- exposed areas of the Escherichia coli GmpA protein does not interfere with export and membrane assembly. Gene, 82. 229-236.

310 I;‘. k’elici et al
Gallusser, A. B Kuhn, A. (1990). Initial steps in protein membrane insertion. Bacteriophage Ml3 procoat pro- tein binds to the membrane surface by electrostatic interaction. EMBO J. 9, 2723-2729.
Greenwood, J. & Perham, R. Ni. (1989). Dual importance of positive charge in the C-terminal region of filamen- tous bacteriophage coat protein for membrane inser- tion and DNA-protein interaction in virus assembly. l’irolagy, 171, 444-452.
Greenwood, ,J.. Willis. A. F:. 8: Perham, R. N. (1991). Multiple display of foreign peptides on a filamentous bacteriophage. Peptides from Plasmodium falciparuwl circumsporozoite protein as antigens. J. Mol. Hiol. 220, 821-827.
Hedegaard, L. & Klemm. P. (1989). Type 1 fimbriar of Escherichia coli as carriers of heterologous antigenic sequences. Gene, 85. I lfi-124.
Il’ichr~. A. A., Minenkova. 0. 0.. Tat’kor. S. 1.. Karpyshev. F. S.. Eroshkin. .4. M., Petrenko. V. A. 8c Sandakhchiev, L. I$. (1989). Production of a viable variant of the Ml3 phage with a foreign peptidr inserted into the basic coat protein. I)&. Akad. Sauk. S.S.S.R. 307, 481-483.
Kang. A. 8.. Barbas. (‘. F.. ,Janda, K. I)., Benkovic. S. J. 8 Lerner. R. A. (1991). Linkage of recognition and replication functions bj- assembling caombinatorial antibody Fab libraries along phage surfaces. Proc. Sat. .4 cad. A”ci.. Iv.R..4 88, 4363-4366.
Kuhn. A.. Kreil, G. & Wirkner. W. (1986). Both hydro- phobic domains of Ml3 procoat are reyuired to initiate membrane insertion. EMBO ./. 5. 3681-36%.
Kuhn. A.. Zhu. H. bt I)albry. R. E. (1990). Eficient translocation of positively charged residues of M I3 procoat protein across the membrane excludes electrophoresis as the primary force for metnbranr insertion. EMBO J. 9. 23852389.
Kunkel. T. A. (1985). Rapid and rficient site-specific mutagenesis without phenotypic selection. I’ux. Sat. rlcad. Sri.. I’.S.A. 82. 488-502.
Marvin. D. A. (1990). Model building studies of Inovirus: genetic variations on a geometric theme. Jnt. J. Hiol. Nacromol. 12. 125-138.
Messing. ,I.. Gronenborn, B.. Miiller-Hill. K. B Hofschneider. P. H. (1977). Filamentous coliphage Ml3 as a cloning vehicle: insertion of a HindIf frag- ment of the lac regulatory region in Ml3 replicativr form in vitro. Proc. Sat. ilcad. Sri.. I’.S.A 74. 3642- 3646.
Model, P. & Russel, M. (1988). Filamentous bacterio- phage. In The Bacteriophage.s ((lalendar. R., ed.). vol. 2, pp. 37.5-456, Plenum, Kew York.
Samba. K.. Yamashita, T. XL Vonderviszt. F. (1989). Structure of the core and cent,ral channel of bacteria1 flagella. Xaturr (London), 342. 648-654.
Parmley. S. F. &, Smibh. (:. I’. (1988). Ant,ibodv-selrct,ablr filamentous fd phage vectors: affinity purification of target genes. Ornw, 73. 305-318.
Rasched. I. & Soberer. E. (1986). Ff cloliphagrs: structural and functional relationships. Microhiol. I~w. 50. 40 l-427.
Rowit’ch. I). H.. Hunter. (:. .J. & Perham. R. X. 11988). Variable eleetrostat’ic intrrartion between D?;A and coat protein in filamentous bacteriophage assembly. .J. Nol. Hiol. 204. 663-674.
Sambrook. .J.. Fritsrh. E. F. & Maniatis. ‘I’. (l!W9). Molecular (‘lon,ing: A Lnhoratoq Manual. (‘old Spring Harbor Laboratory Press. Cold Spring Harbor. ?r’\‘.
Scott. ,J. Ii. & Smit,h, (:. 1’. (1990). Searching for prptidr lipands with an rpitope library. Scirnrv, 249, 386-390.
ShQprr. H. & VOII .Jagow. G. (l!%i). l’ric,iIlr-socliunr dodecyi sulfate-polyacrylamidr gel &c%rophoresis for the separat,ion of proteins in t.he range from I tjo IO0 kl)a. il nal. Biochem. 166. 368-379.
Smith. (:. (1985). .Filammtous fusion J’hagr IloVt'l
expression vectors that display caloned antjigens on
the virion surface. Scl:mer, 228, IYIT, 1317. \Vu, ,J. Y., I\ewton. S.. ,Judd. A.. Stocker. B. B Itobinxol~.
W’. S. (1989). Kxprrssion of immunogenic, epitopes of hepatitis 13 surface antigen with hybrid flagellin prot,rins by a vaccine strain of Stslm~mella. Proc. Sat Acad. Rci., 1Y.8.A. 86. 4726-4730.
Edited by J. Karll
Copyright © 2022 FDOKUMEN