
Seasonal and Spatial Patterns of Distribution of Subtidal Benthic Invertebrate Communities in the...
16
Seasonal and spatial patterns of distribution of subtidal benthic invertebrate communities in the Mondego River, Portugal – a poikilohaline estuary P. Chainho 1, *, J.L. Costa 1 , M.L. Chaves 1 , M.F. Lane 2 , D.M. Dauer 2 & M.J. Costa 1,3 1 Instituto de Oceanografia, Faculdade de Cie ˆncias da Universidade de Lisboa, Campo Grande, 1749-016, Lisboa, Portugal 2 Department of Biological Sciences, Old Dominion University, Norfolk, Virginia, 23529, USA 3 Departamento de Biologia Animal, Faculdade de Cie ˆncias da Universidade de Lisboa, Campo Grande, 1749-016, Lisboa, Portugal (*Author for correspondence: E-mail: [email protected]) Key words: benthic condition, spatial patterns, seasonal variations, environmental gradients, Mondego River estuary, Venice salinity system Abstract The use of benthic assemblages to assess ecological quality of estuarine environments is a major tool for the implementation of the Water Framework Directive (2000/60/CE) for European aquatic eco- systems. Benthic communities show spatially heterogeneous distributions and experience seasonal variations due to both natural and anthropogenic stresses. The major goal of this study was to quantify the relationships between environmental gradients and the spatial and temporal patterns of the benthic communities along a Portuguese estuary. Seasonal and spatial variations relating macro- benthic communities and measures of water and sediment quality along the northern branch of the Mondego River estuary were examined at seven sampling stations from July 2000 to June 2001. Cluster analysis of biological data indicated three major groups of communities based on spatial distribution patterns: (1) a lower sector with stronger marine influence and dominated by Streblospio shrubsolii and Cerastoderma glaucum; (2) a middle sector with dominance of S. shrubsolii and Coro- phium multisetosum; (3) and an upper sector where C. multisetosum dominates a community charac- terized by a lower number of species. Canonical correspondence analyses of biological and environmental data determined a major salinity gradient influencing benthic communities. Seasonal changes of benthic communities were mainly determined by freshwater input and salinity changes that imposed a strong decrease in densities and number of species during winter, followed by a recovery during spring. Benthic ecological structure and contaminant levels indicated that the Mondego northern branch is moderately disturbed, although opportunistic species dominated the benthic com- munity, suggesting that natural and anthopogenic sources of stress may be acting together. The Mondego River estuary, a poikilohaline-type estuary, characterized by strong seasonal changes in water flow and salinity, cannot be consistently stratified into salinity regions based upon the Venice classi- fication system. Biotic communities, exemplified here by the benthic communities, are seasonally dis- placed, compared to a homiohaline-type estuary where the Venice system can be applied without modification. Future identification of reference conditions and design of monitoring programs cannot be accomplished without understanding how interactions between biotic and physico-chemical dynamics differ between homiohaline and poikilohaline estuaries. Results obtained in this study could be used to assist future assessments in other Portuguese estuaries. Hydrobiologia (2006) 555:59–74 ȑ Springer 2006 H. Queiroga, M.R. Cunha, A. Cunha, M.H. Moreira, V. Quintino, A.M. Rodrigues, J. Seroˆ dio & R.M. Warwick (eds), Marine Biodiversity: Patterns and Processes, Assessment, Threats, Management and Conservation DOI 10.1007/s10750-005-1132-2
Transcript of Seasonal and Spatial Patterns of Distribution of Subtidal Benthic Invertebrate Communities in the...

Seasonal and spatial patterns of distribution of subtidal benthic invertebrate
communities in the Mondego River, Portugal – a poikilohaline estuary
P. Chainho1,*, J.L. Costa1, M.L. Chaves1, M.F. Lane2, D.M. Dauer2 & M.J. Costa1,31Instituto de Oceanografia, Faculdade de Ciencias da Universidade de Lisboa, Campo Grande, 1749-016, Lisboa, Portugal2Department of Biological Sciences, Old Dominion University, Norfolk, Virginia, 23529, USA3Departamento de Biologia Animal, Faculdade de Ciencias da Universidade de Lisboa, Campo Grande, 1749-016, Lisboa,Portugal
(*Author for correspondence: E-mail: [email protected])
Key words: benthic condition, spatial patterns, seasonal variations, environmental gradients, Mondego River estuary,Venice salinity system
Abstract
The use of benthic assemblages to assess ecological quality of estuarine environments is a major toolfor the implementation of the Water Framework Directive (2000/60/CE) for European aquatic eco-systems. Benthic communities show spatially heterogeneous distributions and experience seasonalvariations due to both natural and anthropogenic stresses. The major goal of this study was toquantify the relationships between environmental gradients and the spatial and temporal patterns ofthe benthic communities along a Portuguese estuary. Seasonal and spatial variations relating macro-benthic communities and measures of water and sediment quality along the northern branch of theMondego River estuary were examined at seven sampling stations from July 2000 to June 2001.Cluster analysis of biological data indicated three major groups of communities based on spatialdistribution patterns: (1) a lower sector with stronger marine influence and dominated by Streblospioshrubsolii and Cerastoderma glaucum; (2) a middle sector with dominance of S. shrubsolii and Coro-phium multisetosum; (3) and an upper sector where C. multisetosum dominates a community charac-terized by a lower number of species. Canonical correspondence analyses of biological andenvironmental data determined a major salinity gradient influencing benthic communities. Seasonalchanges of benthic communities were mainly determined by freshwater input and salinity changes thatimposed a strong decrease in densities and number of species during winter, followed by a recoveryduring spring. Benthic ecological structure and contaminant levels indicated that the Mondegonorthern branch is moderately disturbed, although opportunistic species dominated the benthic com-munity, suggesting that natural and anthopogenic sources of stress may be acting together. TheMondego River estuary, a poikilohaline-type estuary, characterized by strong seasonal changes in waterflow and salinity, cannot be consistently stratified into salinity regions based upon the Venice classi-fication system. Biotic communities, exemplified here by the benthic communities, are seasonally dis-placed, compared to a homiohaline-type estuary where the Venice system can be applied withoutmodification. Future identification of reference conditions and design of monitoring programs cannotbe accomplished without understanding how interactions between biotic and physico-chemicaldynamics differ between homiohaline and poikilohaline estuaries. Results obtained in this study couldbe used to assist future assessments in other Portuguese estuaries.
Hydrobiologia (2006) 555:59–74 � Springer 2006H. Queiroga, M.R. Cunha, A. Cunha, M.H. Moreira, V. Quintino, A.M. Rodrigues, J. Serodio & R.M. Warwick (eds),Marine Biodiversity: Patterns and Processes, Assessment, Threats, Management and ConservationDOI 10.1007/s10750-005-1132-2

Introduction
Benthic invertebrate communities have beenwidely used as indicators of the ecological status ofcoastal and estuarine ecosystems (e.g. Pearson &Rosenberg, 1978; Hily et al., 1986; Dauer, 1993;Weisberg et al., 1997; Borja et al., 2003) and wereincluded in the biological elements indicated by theWater Framework Directive (WFD) (2000/60/EC)for use in environmental monitoring. Benthicinfaunal species live in the sediments and showrelatively low mobility, being exposed to stress dueto contaminants, low dissolved oxygen, limitingnutrient levels and physical disturbances (Daueret al., 1992; Weisberg et al., 1997; Cowie et al.,2000). Benthic communities include species withdifferent life cycles and specific tolerances to stressevents, which make them suitable to be classifiedinto different functional groups that reflect themagnitude of disturbances (e.g. Bilyard, 1987).They also play an important role in the chemicalfluxes of the water/sediment interface (Aller &Aller, 1998; Aller et al., 2001) and are one of themain compartments of aquatic food webs, beingeffective indicators of impacts at higher levels ofbiological organization (Bilyard, 1987; Aldenet al., 1997).
In spite of all the advantages mentioned abovesome biological characteristics of benthic com-munities have to be taken into account wheninterpreting results of benthic condition assess-ment. Benthic communities show high spatialheterogeneity in estuaries that are related to theinfluence of natural gradients of different factorscontributing to the overall distribution of species.Many benthic species occur along a wide spectrumof an estuarine environment while some others areconfined to a narrower habitat, according to theirtolerance to environmental variables such assalinity, sediment type, depth, etc. In addition tospatial patterns, temperate estuarine invertebratecommunities also show important temporal vari-ations related to seasonal and interannual changes.Seasonal fluctuations in abundance and composi-tion can be due to recruitment pulses that occurduring spring and autumn for most species, butalso to the occurrence of extreme environmentalconditions such as low temperatures, floods anddroughts (Alden et al., 1997; Attrill & Power,2000; Salen-Picard & Arlhac, 2002). Freshwater
flow variability is one of the main factors influ-encing the high temporal and spatial changes inphysical, chemical and biological conditions inestuaries, particularly in rivers that show strongseasonal changes (Kimmerer, 2002). Thesehydrodynamic fluctuations have an importanteffect on the erosion and depositional cycles,influencing the sediment composition and there-fore the colonization by particular benthic com-munities. In addition, widely varying salinitypatterns in an estuary will alter local benthiccommunity composition due to seasonal flowpatterns (Boesch, 1977b) or extreme episodicstorm events (Boesch et al., 1976a, b).
The Mondego River estuary, located in thewestern Atlantic Portuguese coast is divided intotwo branches that diverge 7.5 km upstream fromthe river mouth and have different hydrographiccharacteristics (Fig. 1). In the southern branch thewater circulation is mainly driven by tidal excur-sion and the only freshwater input comes from thePranto River, a small tributary. The northernbranch receives most of the freshwater input and isstrongly influenced by seasonal water flow fluctu-ations (Flindt et al., 1997). These two brancheswere originally in contact through a small channelin the upstream area but the southern branch be-came gradually silted up and the connection oc-curs only during strong spring tides (Cunha &Dinis, 2002). The southern branch subsystem hasbeen widely studied concerning physical andchemical variables that influence ecological pro-cesses. These studies identified a eutrophicationgradient as one of the main factors influencingbenthic communities (Flindt et al., 1997; Marqueset al., 1997; Lillebø et al., 1999; Martins et al.,
Figure 1. Location of sampling stations selected in the northern
branch of the Mondego River estuary.
60

2001; Cardoso et al., 2002). In contrast few studieshave been carried out in subtidal communities ofthe northern branch (Marques et al., 1993; Pardalet al., 1993) and they did not cover the regionupstream of the link between the southern andnorthern branches.
Understanding the spatial and temporal varia-tions of the benthic communities is a basic tool fordiscriminating between pollution induced changesand natural variations (Boesch, 1973; Hollandet al., 1987) and the first step towards the devel-opment of environmental indicator tools for Por-tuguese estuaries. Assessment of the benthiccondition of the northern branch of the MondegoRiver estuary can give a reasonable representationof both natural and man-made impacts in the entireriver basin since it receives most of the freshwatergenerated upstream. This paper gives an overviewof the spatial distribution of benthic communitiesin the northern branch of the Mondego Riverestuary, identifying the main environmental gra-dient generating the distribution of those commu-nities. Seasonal changes were also analyzed byexamining variations in the dominant species.These findings emphasize the ecological impor-tance of the poorly understood biotic differencesbetween homiohaline and poikilohaline estuaries.
Methods
Study area
The Mondego River estuary is located on thePortuguese Atlantic coast (40�08¢ N; 8�50¢ W), a
temperate-warm region influenced both by Atlanticand Mediterranean climates (Fig. 1). This region ischaracterized by a rainfall period that extends fromNovember to May and a drought period of verylow water flow between June and October(Loureiro et al., 1986). River flow data for theperiod between 1987 and 1997 measured at theCoimbra dam indicated an annual mean flow of812 m3 s)1. Maximum flows were measured fromDecember to March and minimum flows occurredbetween June and October (Fig. 2). Averagemonthly flows varied between a maximum value of167 m3 s)1 in January and aminimum average flowof 15 m3 s)1 in September. The Mondego Riverflow regime is very irregular and important dailychanges can occur due to the action of dams con-trolling the discharges. This estuary is affected by amesotidal semi-diurnal regime and is normally to-tally mixed, except for periods of extreme floods ordroughts when it can be only partially mixed(Cunha & Dinis, 2002). During the last decades theMondego River estuary was severely altered by theconstruction of dams located upstream, the drain-age of some mudflat areas, embankment of theriver margins, dredging activities to maintain anavigation channel and intensive agriculture use.Sediment grain size has a very heterogeneousspatial distribution along the estuary, much coarserin the upstream areas where it consists mainly ofvery coarse sand and some marginal locationscovered with fine sand. The lower section has adominant composition of medium to fine sandalthough there are small areas near the river bankswere sandy mud sedimentation occurs (Cunha &Dinis, 2002).
( g )
Figure 2. Average monthly flows measured at Coimbra dam between 1987 and 1997 (source: Portuguese Water Institute).
61

Sampling design
A total of seven sampling stations were selectedalong the longitudinal extension of the northernbranch of the estuary (Fig. 1) in an attempt to coverthe salinity range between the vicinity of the rivermouth and the upper tidal reaches. Three benthicinvertebrate samples were taken at each stationusing a modified van Veen grab (0.05 m2) and theircontents were fixed and preserved with 4% bufferedformalin. Grab contents were sieved in the labora-tory using a 500 lm mesh and preserved in 70%alcohol. All samples were sorted using amicroscopeand invertebrates were identified to the highestpossible taxonomic separation. Bottom dissolvedoxygen (DO) (mg l)1), water temperature (�C) andsalinity were measured in situ using a Data SondeSurveyor 4 and a Sechii disc was used to measuretransparency (m). Additional water samples werecollected and frozen for subsequent analysis ofnitrates (mg l)1), nitrites (mg l)1), phosphates(mg l)1) and ammonia (mg l)1). The analyses weremade by the laboratory of the Portuguese Envi-ronmental Institute using methods certified by thePortuguese Quality Institute. At each station,another grab sample was taken and frozen forsediment grain size, total organic content (TOC)and heavy metals analysis. Sediment grain sizecomposition was determined using an AFNORtype sieve battery (0.063 mm; 0.25 mm; 0.5 mm;2 mm; 9.25 mm) after drying the sediment (60 �C)for a period of 48 h. Samples were classified usingthe Roux (1964) scale. TOC was obtained as thedifference between dry weight, measured after dry-ing the sample at 60 �C during 24 h and ash weight,obtained after ignition at 480 �C for a period of12 h. Heavy metal concentrations (mg kg)1) in thesediment (arsenic, chromium, lead, cooper andzinc) were also determined by the PortugueseEnvironmental Institute, using certified methods.
Seasonal variations were assessed by repeatingthe surveys every 3 months, namely in July (sum-mer) and October 2000 (autumn) and in February/March (winter) and June (spring) 2001. All sam-ples were collected on low tide, during spring tideperiods. The winter cruise took place immediatelyafter a flood event and Stations 1 and 7 could notbe sampled due to the strong currents impedingthe use of the collecting devices.
Data analysis
Spatial patternsStations were classified into spatial groups usingthe ‘‘mean variance per comparison’’ techniquedescribed by Williams & Stephenson (1973). Byapplying this technique it was possible to groupstations into spatial groups independent of theeffects associated with collections conducted dur-ing different seasons. The technique estimates thevariance between stations and between samplingevents by calculating the Euclidean distance be-tween stations and sampling events (over all spe-cies) after stations and sampling events werecentred to their respective means. The varianceestimates were used as a measure of dissimilaritybetween stations for cluster analyses in order toassign the spatial groups. A flexible sorting strat-egy was used for the cluster analysis with anintensity coefficient or value of )0.25 (Boesch,1977a). Dissimilarity coefficients were calculatedusing a program written in the SAS/IML� matrixprogramming language while the dendrogramsfor this analysis were produced using PROCCLUSTER of the SAS /Stat� software package.All species counts were standardized to have anoverall mean value of zero and a standard devia-tion of one prior to conducting this analysis. Theanalysis was conducted using data collected duringonly three seasons (summer, autumn and spring)due to the missing observations in winter. Anoverall test for a significant difference in speciescomposition between site groups was accom-plished using a MANOVA (Wilk’s Lambda) whilepairwise comparisons between individual sitegroups were made using an F-test on pairwisesquared Mahalanobis distances. Descriptive mea-sures determined for the benthic communities ofeach group identified by the cluster analysis werethe average density, the Shannon-Wiener diversityindex (H¢ log e), the number of species (consider-ing each taxon as a species), the Simpson’s domi-nance index (k¢) and the top 10 dominant species.Similarity percentage breakdown procedure(SIMPER) (Clarke & Warwick, 1994), includedin the PRIMER software package was used todetermine the contribution of individual taxatowards the dissimilarity between and similaritywithin the groups identified by Cluster analysis.
62

Seasonal variationMultivariate ordination techniques were applied toanalyze spatio-temporal variation in the speciesdensity reduced data set and identify the relationsbetween environmental and biological data, usingCANOCO 4.5 software. Relations between envi-ronmental and biological data were analyzed byperforming a Canonical Correspondence Analysis(CCA). CCA constrains the axes to be linearcombinations of the environmental variables (terBraak & Smilauer, 2002).
The species data matrix was reduced by elimi-nating taxa that occurred in less than five samplesand accounted for less than 0.1% of the totalabundance, to obtain an interpretable ordinationdiagram.Amatrix of explanatory variableswas alsoconstructed to determine the variation in the speciesdata that was related to environmental factors. Thereduced environmental matrix excluded variablesthat showed collinearity (silt and clay) and variableswhich registered values under the detection limit(nitrates, nitrites, phosphates and ammonia). Met-als were also excluded because they never exceededLong et al.’s (1995) effects range-median (ERM) orCrommentuijn et al.’s (2000) maximum permissibleconcentrations (MPC) and any differences foundbetween stations would not have a biologicallymeaningful interpretation. All other variablesmeasured were included, namely dissolved oxygen,water temperature, salinity, transparency, depth,sediment type (stones, coarse sand, medium sand,fine sand) and total organic content.
Selection of variables was based on stepwisemultiple regressions and the statistical significanceof the variables added to the analysis was testedusing a Monte Carlo permutation test (499 unre-stricted permutations) (ter Braak & Verdonschot,1995). Decision about the ordination model to usein the constrained analysis (CCA) was based onlength of gradient calculated by a previous Detr-ended Canonical Correspondence Analysis(DCCA). Since the longest gradient was 4.05 theCCA was based on a unimodal model. DCCA wasalso used to obtain the ordination of species andsamples, since an ‘‘arch effect’’ was apparent in theinitial canonical correspondence analysis (CCA)(Gauch, 1982).
Seasonal variations were analyzed by plottingmajor taxonomic groups and ecological groups foreach station group-season combination based on
the classification ofBorja et al. (2000) (Appendix 1).These authors assigned taxa to one of five ecologicalgroups, according to their sensitivity to an increas-ing stress gradient (I to V). Contributions fromindividual taxa to dissimilarities between seasonswere estimated using the similarity percentagebreakdown procedure (SIMPER). Densities of thedominant species were plotted for each season.
Results
Benthic macrofauna general characterization
A total of 38 394 invertebrate specimens werecollected and 84 taxa identified (Appendix 1). Thehighest densities were registered during spring atStation 9 (56,613 ind m)2) while the lowest valueswere observed during winter at Station 4 (160 indm)2). The 10 dominant species accounted for 93%of the total average density with the amphipodCorophium multisetosum Stock, 1952 (47%) andthe polychaete Streblospio shrubsolii (Buchanan,1890) (21%) being the two dominant species. Thedominance of C. multisetosum was due to the highnumbers collected during spring at the threeupstream stations. S. shrubsolii was the mostabundant species during all the other seasons.Dominant species found during summer and au-tumn were typical estuarine surface and subsurfacedeposit feeders (e.g. S. shrubsolii, Spio martinensisMesnil, 1896, Caulleriella caputesocis (Saint-Joseph, 1894)) but some freshwater invertebratessuch as insect larvae (Ephemeroptera and Diptera)were collected during winter and spring, mainly inthe upstream stations.
Spatial patterns in benthic communities
Cluster analysis indicated three main groups ofstations in the northern branch of the MondegoRiver estuary (Fig. 3). There was a significantdifference in species composition across all sitegroups (Wilk’s Lambda = 0.063; DF = 28:33;p<0.0001) and between each of the individual sitegroups (Table 1). Spatial Group A consisted ofStations 1 and 2, which are located in the lowersector of the estuary. These stations are charac-terized by medium sand sediments, a stronger tidalinfluence and a salinity decrease associated with
p p y ( )
63
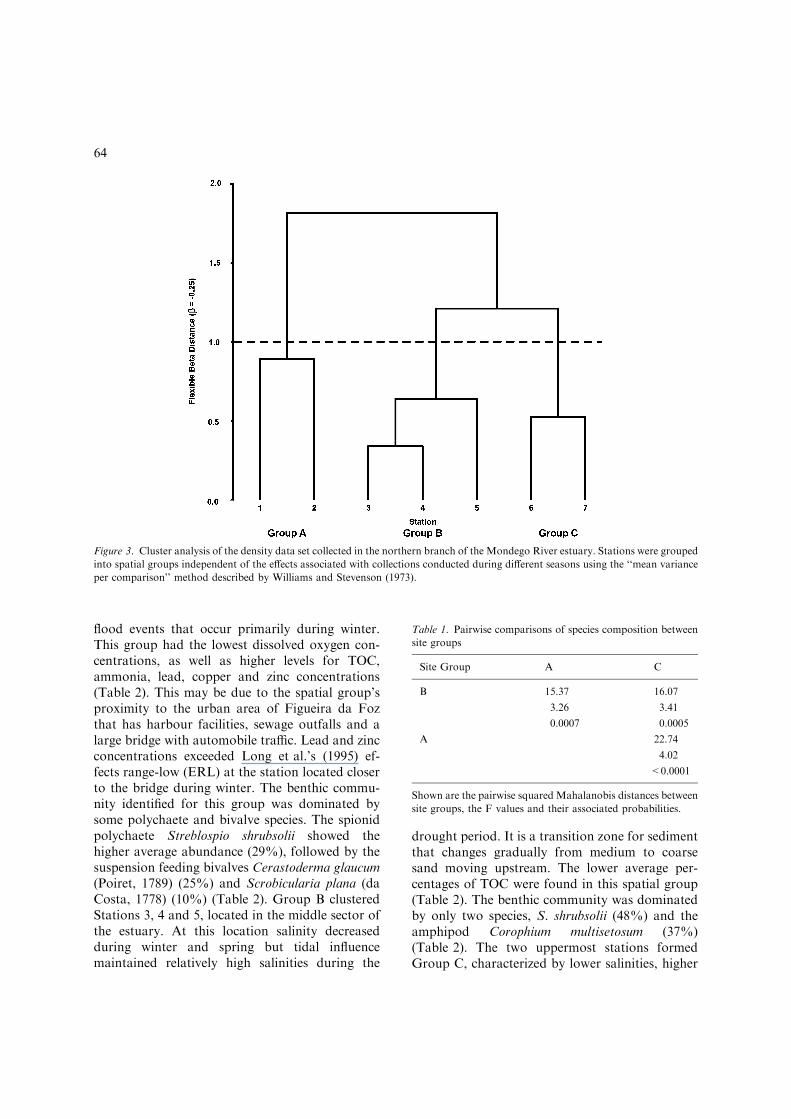
flood events that occur primarily during winter.This group had the lowest dissolved oxygen con-centrations, as well as higher levels for TOC,ammonia, lead, copper and zinc concentrations(Table 2). This may be due to the spatial group’sproximity to the urban area of Figueira da Fozthat has harbour facilities, sewage outfalls and alarge bridge with automobile traffic. Lead and zincconcentrations exceeded Long et al.’s (1995) ef-fects range-low (ERL) at the station located closerto the bridge during winter. The benthic commu-nity identified for this group was dominated bysome polychaete and bivalve species. The spionidpolychaete Streblospio shrubsolii showed thehigher average abundance (29%), followed by thesuspension feeding bivalves Cerastoderma glaucum(Poiret, 1789) (25%) and Scrobicularia plana (daCosta, 1778) (10%) (Table 2). Group B clusteredStations 3, 4 and 5, located in the middle sector ofthe estuary. At this location salinity decreasedduring winter and spring but tidal influencemaintained relatively high salinities during the
drought period. It is a transition zone for sedimentthat changes gradually from medium to coarsesand moving upstream. The lower average per-centages of TOC were found in this spatial group(Table 2). The benthic community was dominatedby only two species, S. shrubsolii (48%) and theamphipod Corophium multisetosum (37%)(Table 2). The two uppermost stations formedGroup C, characterized by lower salinities, higher
Figure 3. Cluster analysis of the density data set collected in the northern branch of the Mondego River estuary. Stations were grouped
into spatial groups independent of the effects associated with collections conducted during different seasons using the ‘‘mean variance
per comparison’’ method described by Williams and Stevenson (1973).
Table 1. Pairwise comparisons of species composition between
site groups
Site Group A C
B 15.37 16.07
3.26 3.41
0.0007 0.0005
A 22.74
4.02
<0.0001
Shown are the pairwise squared Mahalanobis distances between
site groups, the F values and their associated probabilities.
64
oxygen levels and coarser sand. Chromium andzinc measured at these sites exceeded Crom-mentuijn et al.’s (2000) Negligible Concentration(NC) values and Long et al.’s (1995) ERL valuesand higher nitrates and TOC levels were alsomeasured, probably due to the intensive agricul-ture in this area. The benthic community wasstrongly dominated by C. multisetosum (77%),mainly because of high numbers collected duringspring. Oligochaetes (10%) and an introducedbivalve species Corbicula fulminea (Muller, 1774)
(8%) were also abundant. Although in low num-bers, insects larvae (Ephoron virgo (Olivier, 1791)and Chironomidae) occurred in this area associ-ated with higher winter and spring freshwater in-put (Table 2).
SIMPER analysis showed higher dissimilaritiesbetween Groups A and C (91%) and highercloseness of stations of Group B to A than to C.Streblospio shrubsolii gave the highest contributionto the dissimilarities between the lower and themiddle sector of the estuary (16%), except for the
g g g p ( j , ) y g p g y
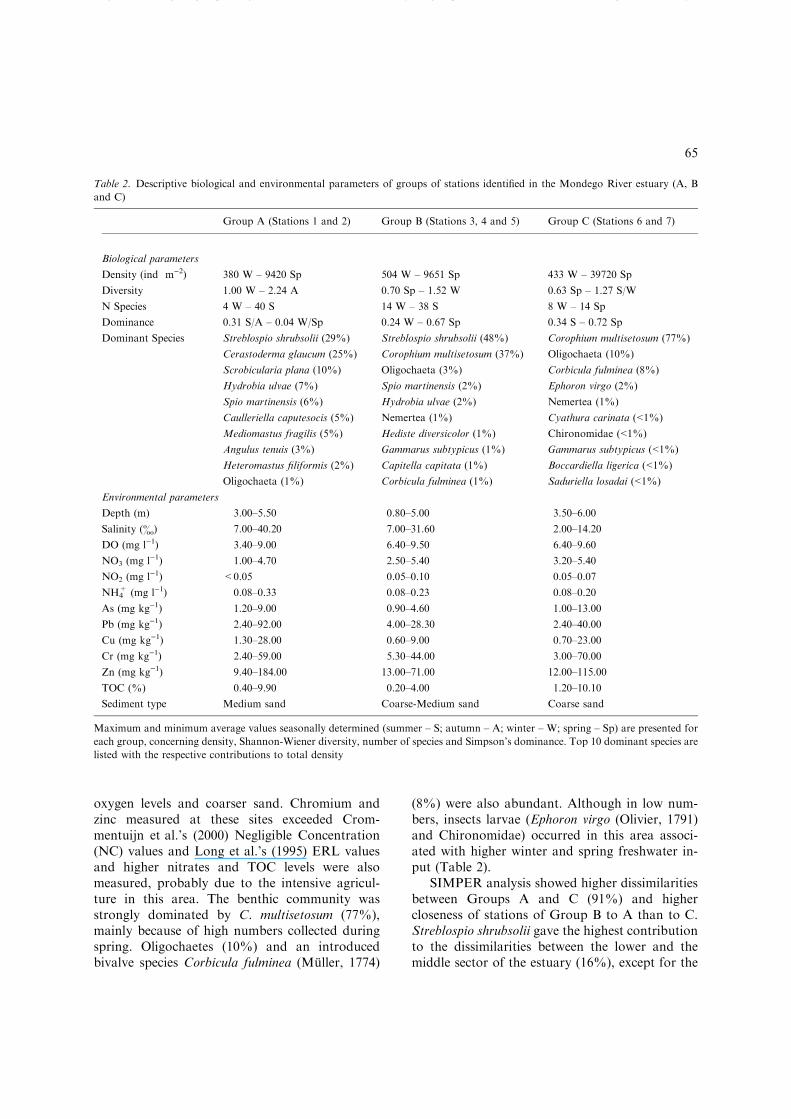
Table 2. Descriptive biological and environmental parameters of groups of stations identified in the Mondego River estuary (A, B
and C)
Group A (Stations 1 and 2) Group B (Stations 3, 4 and 5) Group C (Stations 6 and 7)
Biological parameters
Density (ind m)2) 380 W – 9420 Sp 504 W – 9651 Sp 433 W – 39720 Sp
Diversity 1.00 W – 2.24 A 0.70 Sp – 1.52 W 0.63 Sp – 1.27 S/W
N Species 4 W – 40 S 14 W – 38 S 8 W – 14 Sp
Dominance 0.31 S/A – 0.04 W/Sp 0.24 W – 0.67 Sp 0.34 S – 0.72 Sp
Dominant Species Streblospio shrubsolii (29%) Streblospio shrubsolii (48%) Corophium multisetosum (77%)
Cerastoderma glaucum (25%) Corophium multisetosum (37%) Oligochaeta (10%)
Scrobicularia plana (10%) Oligochaeta (3%) Corbicula fulminea (8%)
Hydrobia ulvae (7%) Spio martinensis (2%) Ephoron virgo (2%)
Spio martinensis (6%) Hydrobia ulvae (2%) Nemertea (1%)
Caulleriella caputesocis (5%) Nemertea (1%) Cyathura carinata (<1%)
Mediomastus fragilis (5%) Hediste diversicolor (1%) Chironomidae (<1%)
Angulus tenuis (3%) Gammarus subtypicus (1%) Gammarus subtypicus (<1%)
Heteromastus filiformis (2%) Capitella capitata (1%) Boccardiella ligerica (<1%)
Oligochaeta (1%) Corbicula fulminea (1%) Saduriella losadai (<1%)
Environmental parameters
Depth (m) 3.00–5.50 0.80–5.00 3.50–6.00
Salinity (&) 7.00–40.20 7.00–31.60 2.00–14.20
DO (mg l)1) 3.40–9.00 6.40–9.50 6.40–9.60
NO3 (mg l)1) 1.00–4.70 2.50–5.40 3.20–5.40
NO2 (mg l)1) <0.05 0.05–0.10 0.05–0.07
NH4+ (mg l)1) 0.08–0.33 0.08–0.23 0.08–0.20
As (mg kg)1) 1.20–9.00 0.90–4.60 1.00–13.00
Pb (mg kg)1) 2.40–92.00 4.00–28.30 2.40–40.00
Cu (mg kg)1) 1.30–28.00 0.60–9.00 0.70–23.00
Cr (mg kg)1) 2.40–59.00 5.30–44.00 3.00–70.00
Zn (mg kg)1) 9.40–184.00 13.00–71.00 12.00–115.00
TOC (%) 0.40–9.90 0.20–4.00 1.20–10.10
Sediment type Medium sand Coarse-Medium sand Coarse sand
Maximum and minimum average values seasonally determined (summer – S; autumn – A; winter – W; spring – Sp) are presented for
each group, concerning density, Shannon-Wiener diversity, number of species and Simpson’s dominance. Top 10 dominant species are
listed with the respective contributions to total density
65

spring assemblage. Although dominant in bothassemblages this spionid occurs with lower densityat Group A stations. On the other hand, Ceras-toderma glaucum, the polychaete Spio martinensisand the gastropod Hydrobia ulvae (Pennant, 1777)were found with much higher densities in Group Aand explained 14%, 7% and 6% of the dissimi-larity between Groups A and B, respectively. Thehigh density of Corophium multisetosum in theupstream stations accounted for 27% of the dis-similarity between Groups B and C. Oligochates,S. shrubsolii and Corbicula fulminea were alsoimportant in separating these two groups and ex-plained 15%, 15% and 12% of the dissimilaritiesbetween Groups B and C, respectively.
Ecological groups obtained by classifying taxaaccording to their sensitivity to pollution (Borjaet al., 2000) were used as a measure of theecological structure of the assemblages showing anaverage density domination of 18 taxa belongingto Group III, which includes species tolerant toorganic enrichment. Over 92% of the organismscollected during the study were classified in thisgroup, while only two taxa, Oligochaeta andCapitella capitata (O. Fabricius, 1780), or about5% of the total number of organisms were classi-fied into Group V (Fig. 4), considered as first-or-der opportunistic species.
Seasonal variation in benthic communities
Major variations were found in winter assemblagesthat showed a reduced number of species and
extremely low abundances (Table 2). In contrast,higher numbers and stronger dominance were reg-istered during spring in all station groups withincreased densities in Group C. As previouslymentioned the ecological structure of the MondegoRiver estuarywas numerically dominated byGroupIII species (Fig. 4). However, some variations of thespecific composition occurred between seasons.Summer and autumn registered a higher heteroge-neity of the ecological groups represented in GroupA, mainly because of the occurrence of rare speciesof marine influence. Spatial Group C registered thehighest numerical representationofGroupVduringsummer and winter, due to the density of Oligo-chaetes (Fig. 4). Spring showedhigher homogeneitydue to the strong numerical abundance ofGroup IIIspecies that lowered the relative contribution ofother groups.
SIMPER analysis showed which species con-tributed the most to the seasonal variations withineach station group:
Group A – higher dissimilarities were foundbetween winter assemblages and all other seasons.Only four taxa were collected with very lownumbers in this period, namely Streblospio shrub-solii, Hesionidae, the isopod Cyathura carinata(Kroyer, 1847) and Turbellaria. Cerastodermaglaucum, Scrobicularia plana, Spio martinensis,Mediomastus fragilis (Rasmussen, 1973) andCaulleriella caputesocis were the species which bestdiscriminated between summer and all other sea-sons, showing higher densities during summer,decreasing towards winter (Fig. 5). S. shruboslii
Figure 4. Ecological groups (Borja et al., 2000) identified seasonally for groups of stations defined in the Mondego River estuary.
66
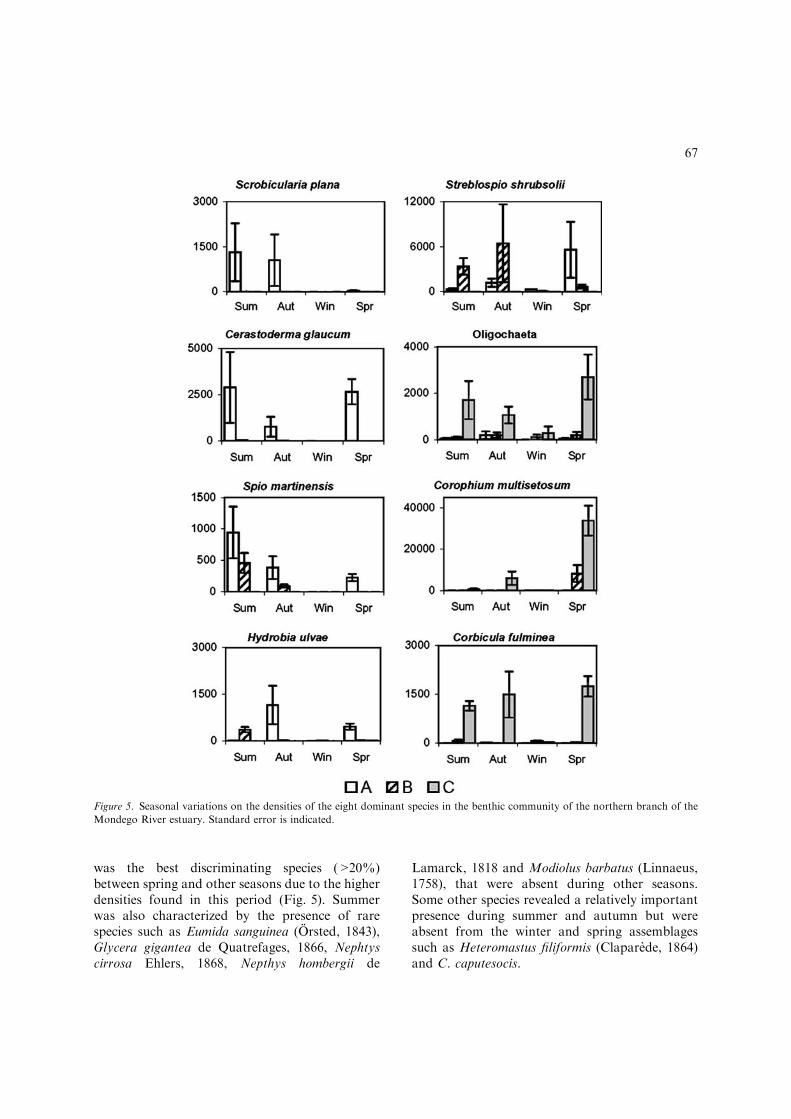
was the best discriminating species (>20%)between spring and other seasons due to the higherdensities found in this period (Fig. 5). Summerwas also characterized by the presence of rarespecies such as Eumida sanguinea (Orsted, 1843),Glycera gigantea de Quatrefages, 1866, Nephtyscirrosa Ehlers, 1868, Nepthys hombergii de
Lamarck, 1818 and Modiolus barbatus (Linnaeus,1758), that were absent during other seasons.Some other species revealed a relatively importantpresence during summer and autumn but wereabsent from the winter and spring assemblagessuch as Heteromastus filiformis (Claparede, 1864)and C. caputesocis.
Figure 5. Seasonal variations on the densities of the eight dominant species in the benthic community of the northern branch of the
Mondego River estuary. Standard error is indicated.
67

Group B – high dissimilarities between seasonswere found, with the lowest dissimilarities regis-tered between summer and autumn assemblages(57%). Streblospio shrubsolii gave the highestcontribution to dissimilarities between all seasons,except for spring, due to density changes. A highernumber of species was collected during autumnand summer (Table 2). Corophium multisetosumoccurred only during winter and spring (Fig. 5)and contributed most to the dissimilarities betweenthese seasonal assemblages (30%). Spio martinen-sis, Caulleriella caputesocis, Glycera gigantea andthe bivalve Angulus tenuis (da Costa, 1778)occurred only during summer and autumn periods,showing the same pattern as Group A, althoughwith lower densities. Chironomidae occurred onlyduring the winter period.
Group C – upper sector assemblages showed ahigher homogeneity between seasons, except forwinter and spring (80% dissimilar) due to thecontribution of Corophium multisetosum (60%)that registered a strong numerical increase duringspring (Fig. 5). This species accounted for thehighest contribution to the dissimilarities betweenspring and other seasons (>50%). Oligochaetesand Corbicula fulminea were also abundant inGroup C, except for the winter period (Fig. 5).Chironomidae occurred exclusively during winterand spring and Ephoron virgo occurred only duringspring.
Relationships between environmental andbiological variables
After data reduction 28 taxa were retained, thataccounted for more than 99% of the total abun-dance.
The constrained ordination (CCA) of speciesdensity with stepwise forward selection of envi-ronmental variables retained only three environ-mental variables (p<0.05): salinity, medium sandand TOC. The ordination plot showed an appar-ent arch effect on species and samples, suggestingthe need for detrending. DCCA of species abun-dance data produced an ordination in which thefirst four axes were statistically significant(p<0.01), with respective eigenvalues of 0.66,0.14, 0.06 and 0.03. The first two axes explained34.9% of the total variance in species data and56.8% of the total variance on the species–envi-
ronment relation. Salinity presented a negativecorrelation with the first axis ()0.90) and it was thevariable with the highest explanatory power re-lated to this axis (Fig. 6). Medium sand contentalso explained some of the variation on the firstaxis, with a correlation of )0.67. In spite ofexplaining some of the variance in species distri-bution, TOC showed low correlations with the firsttwo axes (<0.30) (Fig. 6).
Salinity and sediment grain size established themain gradient separating species that apparentlyhave an optimal distribution in lower salinities andcoarser sands, such as Corophium multisetosum,Corbicula fulminea, Ephoron virgo, Chironomidae,Amage adspersa (Grube, 1863) and Oligochaeta(Fig. 5). These species were mainly identified in theGroup C assemblage. The ordination indicates amedium location of Streblospio shrubsolii,Cyathura carinata, Hediste diversicolor (O.F.Muller, 1776), Saduriella losadai Holthuis, 1964,Nemertea, Lekanesphaera hookerii (Leach, 1814)and Websterinereis glauca (Claparede, 1870) alongthe saline gradient. Some of these species occurredwith higher densities in Group B stations but all ofthem demonstrated tolerance to changes occurringalong the estuary since they showed a wide dis-tribution along the three different regions identi-
Figure 6. DCCA of benthic invertebrate density for the stations
sampled in the Mondego River estuary during summer (S),
autumn (A), winter (W) and spring (Sp). Taxa abundance data
were square root transformed. Environmental variables –
Salinity (Sal), Medium sand (Ms) and Total Organic Content
(TOC) – were plotted on the ordination as arrows. See
Appendix 1 for taxa abbreviations.
68

fied. In the DCCA plot the remaining species werelocated closer to the positive end of the salinityand sediment vectors, associated with spatialGroup A stations. A small part of the speciesvariation was still explained by the second axis(6.2%), separating species characteristic of GroupsA and C assemblages from Group B. Although thecorrelation of TOC to this axis was not very high,this variable apparently explains part of the dif-ferences mentioned, since the Group B stationsregistered lower values. DCCA ordination of spe-cies and samples also revealed temporal differencesin similarity within groups, separating the plots ofthe same stations in different seasons (Fig. 6).
Discussion
The identification of the spatial and seasonal pat-terns of change in the benthic communities of theMondego River estuary may be an importantcontribution to the development of biological cri-teria to assess the environmental condition ofPortuguese estuaries. The study design led to theidentification of three spatial groups with distinctenvironmental and biological characteristics in thenorthern branch of this estuary, instead of the fiveclasses established by the Venice system. Thatsystem, based on the identification of salinityclasses, is widely accepted and indicated by theWFD to identify different water types. Salinitydecreases moving upstream from spatial Group Anear the mouth of the River to the spatial Group Cupstream and is coupled with a transition frommedium to coarse sands upstream. Several sourcesof anthropogenic disturbance were identified inGroup A, such as a well developed urban area withharbour facilities that require periodic dredging ofthe access channel, dumping and a bridge with hightraffic levels. This sector showed organic enrich-ment and higher levels of heavy metals were mea-sured in the bridge vicinity. Lower organicenrichment was found in spatial Group B while theGroup C stations showed higher levels of organicmatter and nutrient concentrations, probablyrelated to nutrient loads from intensive agriculture.
Benthic communities presented similar patternsof longitudinal change along the estuary. The lowersector assemblages were dominated by the surfacedeposit feeder Streblospio shrubsolii and some
suspension feeding (Cerastoderma glaucum) anddeposit-feeding (Scrobicularia plana) bivalve spe-cies. Group B showed a stronger dominance ofS. shrubsolii populations, shared with the amphi-pod Corophium multisetosum. The last speciesmentioned accounted for almost 80% of the uppersector assemblages due to high densities foundduring spring. DCCA identified a saline gradientinfluencing species distribution. Seasonal differ-ences found in community composition andstructure, were apparently also related to salinitychanges caused by high freshwater input fromupstream. Although salinity values were obtainedwith a single measurement for each season, dailyand monthly flows measured upstream stronglyindicate that they were representative of seasonalchanges (Fig. 2). Colonization of Group A stationsby marine species was observed during the droughtseason while Group C stations were colonized byfreshwater species during higher flow periods.Species with higher tolerances to salinity changessuch as S. shrubsolii,Hediste diversicolor, Cyathuracarinata and Saduriella losadai showed a widerdistribution and persistence between seasons.These results are consistent with previous studies inthe Mondego River estuary (Marques et al., 1993;Pardal et al., 1993), although the number of speciesand densities collected during this study were muchhigher. The higher densities in the present study areprobably due to the use of a 1 mm mesh sieveduring previous studies, compared to the use of a0.5 mm mesh sieve in the present study.
Salinity effects may be acting together withphysical disturbances caused by the strong currentsoccurring during the rainfall period and conse-quent alterations of the erosion–deposition cycles.Winter communities were very impoverished bothnumerically and in terms of the number of speciesand stronger changes occurred in the lower sectorof the estuary, where most stenohaline species werefound. In the middle sector lower salinities per-sisted for a longer period and the benthic com-munity was apparently adapted to this salineregime showing lower changes between seasons.Streblospio shrubsolii, a typical estuarine speciesdominated the community over all seasons, exceptfor spring when Corophium multisetosum increasedits density. This polychaete is classified as anopportunistic species that colonises organicallyenriched sediments (Pearson & Rosenberg, 1978;
69

Sarda & Martin, 1993). This change in the domi-nance may be due to competition as both speciesoccupy similar spatial niches (surface feedersdwelling in the upper 1 cm of the sediment) andoccur in high densities. Previous studies carried onPortuguese estuaries showed the prevalence of thisamphipod in salinities ranging from 2.5 to 10 andcoarser sediment (Queiroga, 1990). Cunha et al.(2000) concluded that C. multisetosum abundancemay be associated with increased freshwater inflowfollowing rainy periods and subsequent decreasingabundance with the higher summer temperatures,since it is a cold-temperature species, with itssouthern limit in Portugal. During spring, C. mul-tisetosum seems to replace the dominant spionidS. shrubsolii in Group B, since increasing densitiesoccur in the lower sector for this species in the sameperiod. The benthic community of the upper sectorseems to be more stable with lower fluctuations inspecific composition and densities, except for thespring boom of C. multisetosum.
The AZTI Marine Biotic Index (AMBI) ap-proach of Borja et al. (2000) produces an overallevaluation of benthic community condition, rela-tive to anthropogenic stress, by scoring benthicspecies based upon relative tolerance to pollution.Using this approach the Mondego River estuarywould be considered moderately disturbed sinceGroup III species are dominant. These species aretolerant of excess organic matter and occur innormal conditions but increase their numbers whenstimulated by organic enrichment. Based on AMBIvalues, winter communities in the upper sectorwould be considered severely degraded due to thedominance of oligochaetes and summer and au-tumn communities in Group A would be less dis-turbed since a higher number of pollution sensitivespecies occur (e.g. A. tenuis, Nephtys spp., Glyceraspp., Owenia fusiformis delle Chiaje, 1842, Eteonepicta de Quatrefages, 1866, Eumida sanguinea, Di-opatra neapolitana delle Chiaje, 1841). Pollutionsensitive species did occur in all sectors but gener-ally in low densities and with great variation be-tween seasons. These results suggest caution inapplying and interpreting the AMBI approachwhen strong seasonal patterns occur. Nevertheless,benthic communities are still under seasonalphysical and physiological stress that favours thesettlement of species with opportunistic life histo-ries and/or wide tolerance to changes in environ-
mental conditions, such as Streblospio shrubsolii(Sarda & Martin, 1993; Rossi & Lardicci, 2002).The present data demonstrate the ability of thisspecies to colonize and produce high-density localpopulations in the lower sector of the MondegoRiver between winter and spring. Lardicci et al.(1997) indicated peaks of fecundity occurring every2 months for this species when food availability isnot a limiting factor.
Estuaries may be classified in many waysincluding geomorphology (e.g. drowned river val-leys, fjords, etc.), degree of saline stratification (e.g.highly stratified, partially stratified, etc.), and tidalmixing (e.g., macrotidal, microtidal, etc.) (Lauff,1967;Elliott&McLusky, 2002).Major variations inamount of freshwater flow may be due to strongseasonal differences in rainfall and such differencescan be amplified by dam placement. Boesch (1977b)states that poikilohaline estuaries, where strongseasonal changes in salinity and water flow occur atspecific locations, cannot be assessed using theVenice system.He also states that euryhalinemarinespecies are displaced as an effect of poikilohalinity,allowing the domination of estuarine endemic spe-cies. The dominance of Streblospio shrubsolii inGroup B during summer and autumn and in GroupA during winter and spring indicates an agreementof the Mondego River estuary results with poiki-lohalinity effects theory. Apparently, strongerfreshwater inputs displace euryhaline species foundin the lower sector of the estuary during lower flowperiods, allowing the dominance of the spionid S.shrubsolii. At the same time, this estuarine endemicspecies is displaced from the middle sector byfreshwater species such as the amphipodCorophiummultisetosum.
The Mondego River estuary, a poikilohaline-type estuary, characterized by strong seasonalchanges in water flow and salinity, cannot beconsistently stratified into salinity regions basedupon the Venice classification system as indicatedby the WFD. Biotic communities, exemplified hereby the benthic communities, are seasonally dis-placed, compared to a homiohaline-type estuarywhere the Venice system can be applied withoutmodification. Future identification of referenceconditions and design of monitoring programscannot be accomplished without understandinghow interactions between biotic and physico-chemical dynamics differ between homiohaline
70

and poikilohaline estuaries (Elliott & McLusky,2002). Our results indicate that seasonal and spa-tial stratification may be necessary to be able toseparate natural and anthropogenic stresses. Inaddition, to the biotic effects further research isnecessary in understanding the relative roles ofriver flow versus groundwater as sources of pol-lutants. For example, during low river flow sea-sons groundwater flow may become relativelymuch more important than during times of highriver flow. Understanding ecological patterns,particularly where spatial and/or temporal varia-tion, is great, also requires a comprehensiveappreciation of the interactions of geomorphol-ogy, hydrology, and climatology.
Acknowledgments
This study was financially supported by two Ph.D.fellowships (SFRH/BD/5144/2001 and SFRH/BD/6365/2001) granted by FCT (Science andTechnology Foundation) and ESF in the aim ofthe III European Community Support Frame-work. Financial support was also given to a pro-ject sponsored by Instituto de Ambiente. The SAS/IML program used here was written by Dr. Ray-mond W. Alden III of the University of Nevada,Las Vegas. We would also like to thank ProfessorsJean-Claude Dauvin and Joao Castro for the helpwith the identification of some invertebrate spe-cies. Finally, we thank Gilda Silva and NunoPrista for their valuable comments and contribu-tions to the final draft of this manuscript.
References
Alden, R. W., S. B. Weisberg, J. A. Ranasinghe & D. M.
Dauer, 1997. Optimizing temporal sampling strategies for
benthic environmental monitoring programs. Marine Pollu-
tion Bulletin 34: 913–922.
Aller, R. C. & J. Y. Aller, 1998. The effect of biogenic irrigation
intensity and solute exchange on diagenetic reaction rates in
marine sediments. Journal of Marine Research 56: 905–936.
Aller, R. C., J. Y. Aller & P. F. Kemp, 2001. Effects of particle
and solute transport on rates and extent of remineralization
in bioturbated sediment. In Aller, J. Y. S. A. Woodin, & R.
C. Aller (eds) Organism–Ssediment Interactions. University
of South Carolina Press, Columbia, USA: 402 pp.
Attrill, M. J. & M. Power, 2000. Effects on invertebrate pop-
ulations of drought-induced changes in estuarine water
quality. Marine Ecology Progress Series 203: 133–143.
Bilyard,G., 1987.The valueof benthic infauna inmarine pollution
monitoring studies. Marine Pollution Bulletin 18: 581–585.
Boesch, D. F., 1973. Classification and community structure of
macrobenthos in the Hampton Roads area, Virginia. Marine
Biology 21: 226–244.
Boesch, D. F., 1977a. Application of Numerical Classification
in Ecological Investigations of Water Pollution. EPA Report
600/3–77–033 U.S. EPA. Corvallis, oregan, USA: 115 pp.
Boesch, D. F., 1977b. A new look at the zonation of the ben-
thos along the estuarine gradient. In Coull, B. (ed.) Ecology
of Marine Benthos. University of South Carolina Press,
Columbia, USA: 467 pp.
Boesch, D. F., R. J. Diaz & R. W. Virnstein, 1976a. Effects of
tropical storm Agnes on soft-bottom communities of the
James and York estuaries and the lower Chesapeake Bay.
Chesapeake Science 17: 246–259.
Boesch, D. F., M. L. Wass & R. W. Virnstein, 1976b. The
dynamics of estuarine benthic communities. In Wiley, M.
(ed.) Estuarine Processes, Vol. I. Academic Press, New
York: 541 pp.
Borja, A., J. Franco & V. Perez, 2000. A Marine Biotic Index to
establish the ecological quality of soft bottom benthos within
European estuarine and coastal environments. Marine Pol-
lution Bulletin 40: 1100–1114.
Borja, A., I. Muxika & J. Franco, 2003. The application of a
marine biotic index to different impact sources affecting soft-
bottom benthic communities along European coasts. Marine
Pollution Bulletin 46: 835–845.
ter Braak, C. J. F. & P. Smilauer, 2002. CANOCO Reference
Manual and CanoDraw for Windows Users Guide: Software
for Canonical Community Ordination (version 4.5). Micro-
computer Power, Ithaca, New York, USA: 500 pp.
ter Braak, C. J. F. & P. F. M. Verdonschot, 1995. Canonical
correspondence analysis and related multivariate methods in
aquatic ecology. Aquatic Sciences 57: 255–289.
Cardoso, P. G., A. I. Lillebø, M. A. Pardal, S. M. Ferreira &
J. C. Marques, 2002. The effect of different primary pro-
ducers on Hydrobia ulvae population dynamics: a case study
in a temperate intertidal estuary. Journal of Experimental
Marine Biology and Ecology 277: 173–195.
Clarke, K. R. & R. M. Warwick, 1994. Change in Marine
Communities: an Approach to Statistical Analysis and
Interpretation. Natural Environmental Research Council,
Plymouth Marine Laboratory, United Kingdom: 144 pp.
Cowie, P. R., S. Widdicombe & M. C. Austen, 2000. Effects of
physical disturbance on an estuarine intertidal community:
field and mesocosm results compared. Marine Biology 136:
485–495.
Crommentuijn, T., D. Sijm, J. de Bruijn, M. Van den Hoop
K. van Leeuwen & E. van de Plassche, 2000. Maximum
permissible and negligible concentrations for metals and
metalloids in the Netherlands, taking into account back-
ground concentrations. Journal of Environmental Manage-
ment 60: 121–143.
Cunha, M. R., M. H. Moreira & J. C. Sorbe, 2000. The
amphipod Corophium multisetosum (Corophiidae) in Ria de
71

Aveiro (NW Portugal). II. Abundance, biomass and pro-
duction. Marine Biology 137: 651–660.
Cunha, P. P. & J. Dinis, 2002. Sedimentary dynamics of the
Mondego estuary. In Pardal, M. A. J. C. Marques, & M. A.
S. Graca (eds) Aquatic Ecology of the Mondego River
Basin: Global Importance of Local Experience. Imprensa da
Universidade de Coimbra, Portugal: 43–62.
Dauer, D. M., 1993. Biological criteria, environmental health
and estuarine macrobenthic community structure. Marine
Pollution Bulletin 26: 249–257.
Dauer, D. M., J. A. Ranasinghe & A. J. Rodi, 1992. Effects of
low dissolved oxygen events on the macrobenthos of the
lower Chesapeake Bay. Estuaries 15: 384–391.
Elliott, M. & McLusky, 2002. The need for definitions in
understanding estuaries. Estuarine, Coastal and Shelf Sci-
ence 55: 815–827.
Flindt, M. R., L. Kamp-Nielsen, J. C. Marques, M. A. Pardal,
M.Bocci,G.Bendoricchio,J.Salomonsen,S.N.Nielsen&S.E.
Jørgensen, 1997. Description of three shallow estuaries:
Mondego river (Portugal), RoskildeFjord (Denmark) and the
lagoon of Venice (Italy). EcologicalModelling 102: 17–31.
Gauch, H. G., 1982. Multivariate Analysis in Community
Ecology. Cambridge University Press, Cambridge, United
Kingdom: 314 pp.
Hily, C., H. le Bris & M. Glemarec, 1986. Impacts biologiques
des emissaires urbains sur les ecosystemes benthiques. Oce-
anis 12: 419–426.
Holland, A. F., A. T. Shaughnessy &M. H. Hiegel, 1987. Long-
term variation in mesohaline Chesapeake Bay macroben-
thos: spatial and temporal patterns. Estuaries 10: 227–245.
Kimmerer, W. J., 2002. Effects of freshwater flow on abundance
of estuarine organisms: physical effects or trophic linkages?.
Marine Ecology Progress Series 243: 39–55.
Lardicci, C., G. Ceccherelli & F. Rossi, 1997. Streblospio
shrubsolii (Polychaeta: Spionidae): temporal fluctuations in
size and reproductive activity. Cahiers de Biologie Marine
38: 207–214.
Lillebø, A. I., M. A. Pardal & J. C. Marques, 1999. Population
structure, dynamics and production of Hydrobia ulvae
(Pennant) (Mollusca: Prosobranchia) along an eutrophica-
tion gradient in the Mondego estuary (Portugal). Acta
Oecologica 28: 289–304.
Lauff, G. H., 1967. Estuaries. American Association for the
Advancement of Science, Washington, D.C. 757 pp.
Long, E. R., D. D. McDonald, S. L. Smith & F. D. Calder,
1995. Incidence of adverse biological effects within ranges of
chemical concentrations in marine and estuarine sediments.
Environmental Management 19: 81–95.
Loureiro, J. J., M. C. Almeida, M. L. Machado & E. Teixeira,
1986. Bacia hidrografica do rio Mondego. In D.G.R.A.H.,
(ed.) Monografias Hidrologicas dos principais cursos de
agua de Portugal continental. Lisboa, Portugal: 240–278.
Marques, J. C., P. Maranhao & M. A. Pardal, 1993. Human
impact assessment on the subtidal macrobenthic community
structure in the Mondego estuary (western Portugal). Estu-
arine, Coastal and Shelf Science 37: 403–419.
Marques, J. C., M. A. Pardal, S. N. Nielsen & S. E. Jorgensen,
1997. Analysis of the properties of exergy and biodiversity
along an estuarine gradient of eutrophication. Ecological
Modelling 102: 155–167.
Martins, I., M. A. Pardal, A. I. Lillebø, M. R. Flindt & J. C.
Marques, 2001. Hydrodynamics as a major factor controlling
the occurrence of green macroalgal blooms in a eutrophic
estuary: a case study on the influence of precipitation and
river management. Estuarine, Coastal and Shelf Science 52:
165–177.
Pardal, M. A., J. C. Marques & G. Bellan, 1993. Spatial dis-
tribution and seasonal variation of subtidal polychaete
populations in the Mondego estuary (western Portugal).
Cahiers de Biologie Marine 34: 497–512.
Pearson, T. H. & R. Rosenberg, 1978. Macrobenthic succession
in relation to organic enrichment and pollution of the marine
environment. Oceanography and Marine Biology. Annual
Review 16: 229–311.
Queiroga, H., 1990. Corophium multisetosum (Amphipoda:
Corophiidae) in Canal de Mira, Portugal: some factors that
affect its distribution. Marine Biology 104: 397–402.
Rossi, F. & C. Lardicci, 2002. Role of the nutritive value of
sediment in regulating population dynamics of the deposit-
feeding polychaete Streblospio shrubsolii. Marine Biology
140: 1129–1138.
Roux, R. M., 1964. Les sediments de l’etange de Reine.
Recueil des Travaux de la Station Maritime d’Endoume
35: 257–285.
Salen-Picard, C. & D. Arlhac, 2002. Long-term changes in a
Mediterranean benthic community: relationships between
the Polychaete assemblages and hydrological variations of
the Rhone river. Estuaries 25: 1121–1130.
Sarda, R. & D. Martin, 1993. Populations of Streblospio
Webster (Polychaeta: Spionidae) in temperate zones:
demography and production. Journal of the Marine Bio-
logical Association of the United Kingdom 73: 769–784.
Weisberg, S. B., J. A. Ranasingue, C. Schaffner, R. J. Diaz, D.
M. Dauer, L. Frithsen & J.B. , 1997. An estuarine benthic
index of biotic integrity (B-IBI) for Chesapeake Bay. Estu-
aries 20: 149–158.
Williams, W. T. & W. Stephenson, 1973. The analysis of three-
dimensional data (sites · times· species) in marine ecology.
Journal of Experimental Marine Biology and Ecology 11:
207–227.
72

Appendix
List of taxa identified in the northern branch of the Mondego River estuary and respective ecological groups, assigned according to
Borja et al. (2000) criteria. Abbreviations of taxa used in the DCCA are also shown.
Phyllum Plathyelmintha Family Phyllodocidae Family Gammaridae
ClassTurbellaria Eteone picta (II) Gammarus subtypicus (I)
Turbellaria n.i. (NA) Eumida sanguinea (II) Esa Family Haustoriidae
Phyllum Nemertea Ophiodromus flexuosus (II) Haustorius arenarius (I)
Nemertea n.i. (III) Nem Family Pilargiidae Family Hyalidae
Phyllum Annelida Sigambra tentaculata (NA) Hyale pontica (I)
Class Oligochaeta Family Pisionidae Family Melitidae
Oligochaeta n.i. (V) Oli Pisione remota (II) Melita palmata (III) Mpa
Class Hirudinea Family Polygordiidae Order Decapoda
Hirudinea n.i. (NA) Polygordiidae n.i. (NA) Family Crangonidae
Class Polychaeta Family Saccocirridae Crangon crangon (I)
Family Ampharetidae Saccocirrus papillocercus (NA) Family Portunidae
Amage adspersa (NA) Aad Family Sabellariidae Carcinus maenas (III)
Amage gallasii (NA) Aga Sabellariidae n.i. (I) Family Processidae
Family Capitellidae Family Spionidae Processa sp. (I)
Capitella capitata (V) Cca Boccardiella ligerica (IV) Class Insecta
Heteromastus filiformis (III) Hfi Polydora ciliata (IV) Order Colembola
Mediomastus fragilis (III) Mfr Prionospio fallax (IV) Colembola n.i. (NA)
Family Cirratulidae Prionospio cirrifera (IV) Order Ephemeroptera
Caulleriella caputesocis (III) Ccapu Pygospio elegans (III) Sma Ephemeroptera n.i. (NA)
Family Glyceridae Spio martinensis (III) Ssh Family Polymitarcidae
Glycera tridactyla (II) Streblospio shrubsolii (III) Ephoron virgo (II)
Glycera gigantea (II) Ggi Phyllum Arthropoda Family Caenidae
Goniada emerita (II) Class Crustacea Caenis sp. (NA)
Goniada norvegica (II) Order Ostracoda Order Plecoptera
Family Hesionidae Ostracoda n.i. (NA) Family Leuctridae
Hesionidae n.i. (NA) Hes Order Tanaidacea Leuctridae n.i. (NA)
Family Nereididae Family Paratanaidae Order Diptera
Hediste diversicolor (III) Hdi Leptochelia dubia (NA) Family Chironomidae
Websterinereis glauca (III) Wgl Heterotanais sp. (NA) Chironomidae n.i. (IV)
Family Nephtyidae Order Mysidacea Phyllum Mollusca
Nephtys caeca (II) Family Mysidae Class Gastropoda
Nephtys cirrosa (II) Acanthomysis longicornis (NA) Family Hydrobiidae
Nephtys hystricis (II) Gastrosaccus spinifer (NA) Hydrobia ulvae (III) Hul
Nephtys hombergii (II) Nho Leptomysis gracilis (NA) Class Bivalvia
Nephtys longesetosa (II) Order Isopoda Family Corbicularidae
Nephtys pulchra (II) Npu Family Anthuridae Ccar Corbicula fulminea (III) Cfu
Family Opheliidae Cyathura carinata (III) Family Cardiidae
Ophelia radiata (I) Family Chaetiliidae Slo Cerastoderma glaucum (III) Cgl
Family Onuphidae Saduriella losadai (NA) Family Mytilidae
Diopatra neapolitana (I) Family Gnathiidae Modiolus barbatus (I) Mba
Family Orbiniidae Paragnathia formica (III) Family Solenidae
Scoloplos armiger (I) Sar Family Sphaeromatidae Lho Solen marginatus (I)
Family Oweniidae Lekanesphaera hookeri (II) Family Scrobiculariidae
Continued on p. 74
73

Appendix (Continued)
Owenia fusiformis (I) Order Amphipoda Scrobicularia plana (III) Spl
Family Paraonidae Amphipoda n.i. (NA) Family Tellinidae
Paraonidae n.i. (NA) Family Ampeliscidae Angulus tenuis (I)
Family Pectinariidae Ampelisca lusitanica (I) Class Opisthobranchia
Pectinaria koreni (I) Family Amphithoidae Opisthobranchia n.i. (NA)
Family Pholoidae Amphithoe sp. (NA) Phyllum Echinodermata
Pholoe minuta (II) Family Corophiidae Class Ophiuroidea
Corophium acherusicum (III) Cmu Ophiuroidea n.i. (NA)
Corophium multisetosum (III)
74




















