
Success with Intravenous Alpha Lipoic Acid (thioctic acid) for Amanita hepatotoxic poisoning.

at SciVerse ScienceDirect
Plant Physiology and Biochemistry 70 (2013) 43e51
Contents lists available
Plant Physiology and Biochemistry
journal homepage: www.elsevier .com/locate/plaphy
Research article
Salicylic acid-mediated establishment of the compatibility betweenAlternaria brassicicola and Brassica juncea is mitigated by abscisic acidin Sinapis alba
Mrinmoy Mazumder a, Srirupa Das a, Upala Saha a, Madhuvanti Chatterjee a,Kaushik Bannerjee b, Debabrata Basu a,*
aDivision of Plant Biology, Bose Institute, P1/12 C. I. T. Scheme VIIM, Kolkata 700054, West Bengal, IndiabNational Research Center for Grapes, P.B. No-3, P.O. Manjri Farm, Solapur Road, Pune 412307, Maharashtra, India
a r t i c l e i n f o
Article history:Received 14 December 2012Accepted 30 April 2013Available online 21 May 2013
Keywords:Abscisic acidAlternaria brassicicolaBrassica junceaJasmonic acidNecrotrophic pathogenSalicylic acidSinapis alba
* Corresponding author. Tel.: þ91 (0) 33 25693312E-mail addresses: mrinmoy_mazumder82@yah
[email protected] (S. Das), Upala.saha@[email protected] (M. Chatter(K. Bannerjee), [email protected],(D. Basu).
0981-9428/$ e see front matter � 2013 Elsevier Mashttp://dx.doi.org/10.1016/j.plaphy.2013.04.025
a b s t r a c t
This work addresses the changes in the phytohormonal signature in the recognition of the necrotrophicfungal pathogen Alternaria brassicicola by susceptible Brassica juncea and resistant Sinapis alba. AlthoughB. juncea, S. alba and Arabidopsis all belong to the same family, Brassicaceae, the phytohormonal responseof susceptible B. juncea towards this pathogen is unique because the latter two species express non-hostresistance. The differential expression of the PR1 gene and the increased level of salicylic acid (SA)indicated that an SA-mediated biotrophic mode of defence response was triggered in B. juncea uponchallenge with the pathogen. Compared to B. juncea, resistant S. alba initiated enhanced abscisic acid(ABA) and jasmonic acid (JA) responses following challenge with this pathogen, as revealed by moni-toring the expression of ABA-related genes along with the concentration of ABA and JA. Furthermore,these results were verified by the exogenous application of ABA on B. juncea leaves prior to challengewith A. brassicicola, which resulted in a delayed disease progression, followed by the inhibition of thepathogen-mediated increase in SA response and enhanced JA levels. Therefore, it seems thatA. brassicicola is steering the defence response towards a biotrophic mode by mounting an SA response insusceptible B. juncea, whereas the enhanced ABA response of S. alba not only counteracts the SA responsebut also restores the necrotrophic mode of resistance by enhancing JA biosynthesis.
� 2013 Elsevier Masson SAS. All rights reserved.
1. Introduction
Plants are under constant threat from thousands of microbes.However, only a handful of these microbes are able to colonise andspread disease. To combat pathogen attack, plants evoke bothconstitutive (pre-formed physical and chemical barriers) as well asinduced immune responses. It is a well-known fact that phyto-hormones and their cross-talk play a crucial role during the innatedefence response in determining the susceptibility/resistivity to aparticular pathogen [1e3]. Plants activate distinctive defencepathways depending on the lifestyle of the attacker, i.e., whether itis a necrotroph or a biotroph. The lifestyle of the attacker
; fax: þ91 (0) 33 2355 3886.oo.co.in (M. Mazumder),gmail.com (U. Saha),
jee), [email protected][email protected]
son SAS. All rights reserved.
determines which set of distinct complex network of phytohor-mone signalling pathways is to be initiated to combat the challenge[4,5]. Therefore, early recognition of pathogens is one of the pre-requisite step for any plants. In resistant plants, the first level ofresistance response results from recognition of pathogen associ-ated molecular pattern (PAMP) and known as PTI (PAMP TriggeredImmunity) but pathogen specific effectors interfere with PTI andcauses ETS (Effectors Triggered Susceptibility). The second level ofresistance response starts on recognition of pathogen specific ef-fectors by plant specific factors (R-gene) and develops ETI (EffectorsTriggered Immunity). The susceptible plant fails to block/recognizethe effectors and thus results ETS (Effectors Triggered Susceptibil-ity) [6].
Different phytohormones have distinctly different mechanisticfunctions that provide protection to plants against diverse types ofpathogens and pathogen-induced stress conditions. Hormones,such as salicylic acid (SA), jasmonic acid (JA), and ethylene (ET)along with reactive oxygen species (ROS) were previously consid-ered as to have primary roles in the initiation of defence response
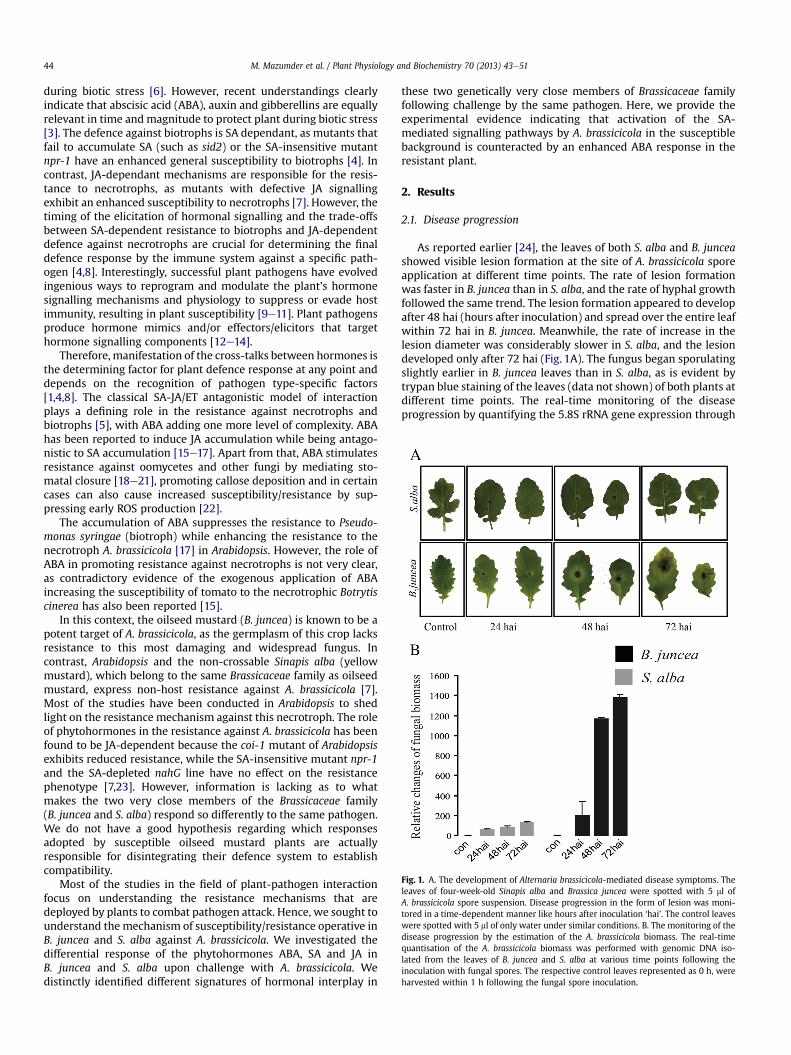
Fig. 1. A. The development of Alternaria brassicicola-mediated disease symptoms. Theleaves of four-week-old Sinapis alba and Brassica juncea were spotted with 5 ml ofA. brassicicola spore suspension. Disease progression in the form of lesion was moni-tored in a time-dependent manner like hours after inoculation ‘hai’. The control leaveswere spotted with 5 ml of only water under similar conditions. B. The monitoring of thedisease progression by the estimation of the A. brassicicola biomass. The real-timequantisation of the A. brassicicola biomass was performed with genomic DNA iso-lated from the leaves of B. juncea and S. alba at various time points following theinoculation with fungal spores. The respective control leaves represented as 0 h, wereharvested within 1 h following the fungal spore inoculation.
M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e5144
during biotic stress [6]. However, recent understandings clearlyindicate that abscisic acid (ABA), auxin and gibberellins are equallyrelevant in time and magnitude to protect plant during biotic stress[3]. The defence against biotrophs is SA dependant, as mutants thatfail to accumulate SA (such as sid2) or the SA-insensitive mutantnpr-1 have an enhanced general susceptibility to biotrophs [4]. Incontrast, JA-dependant mechanisms are responsible for the resis-tance to necrotrophs, as mutants with defective JA signallingexhibit an enhanced susceptibility to necrotrophs [7]. However, thetiming of the elicitation of hormonal signalling and the trade-offsbetween SA-dependent resistance to biotrophs and JA-dependentdefence against necrotrophs are crucial for determining the finaldefence response by the immune system against a specific path-ogen [4,8]. Interestingly, successful plant pathogens have evolvedingenious ways to reprogram and modulate the plant’s hormonesignalling mechanisms and physiology to suppress or evade hostimmunity, resulting in plant susceptibility [9e11]. Plant pathogensproduce hormone mimics and/or effectors/elicitors that targethormone signalling components [12e14].
Therefore,manifestation of the cross-talks between hormones isthe determining factor for plant defence response at any point anddepends on the recognition of pathogen type-specific factors[1,4,8]. The classical SA-JA/ET antagonistic model of interactionplays a defining role in the resistance against necrotrophs andbiotrophs [5], with ABA adding one more level of complexity. ABAhas been reported to induce JA accumulation while being antago-nistic to SA accumulation [15e17]. Apart from that, ABA stimulatesresistance against oomycetes and other fungi by mediating sto-matal closure [18e21], promoting callose deposition and in certaincases can also cause increased susceptibility/resistance by sup-pressing early ROS production [22].
The accumulation of ABA suppresses the resistance to Pseudo-monas syringae (biotroph) while enhancing the resistance to thenecrotroph A. brassicicola [17] in Arabidopsis. However, the role ofABA in promoting resistance against necrotrophs is not very clear,as contradictory evidence of the exogenous application of ABAincreasing the susceptibility of tomato to the necrotrophic Botrytiscinerea has also been reported [15].
In this context, the oilseed mustard (B. juncea) is known to be apotent target of A. brassicicola, as the germplasm of this crop lacksresistance to this most damaging and widespread fungus. Incontrast, Arabidopsis and the non-crossable Sinapis alba (yellowmustard), which belong to the same Brassicaceae family as oilseedmustard, express non-host resistance against A. brassicicola [7].Most of the studies have been conducted in Arabidopsis to shedlight on the resistance mechanism against this necrotroph. The roleof phytohormones in the resistance against A. brassicicola has beenfound to be JA-dependent because the coi-1 mutant of Arabidopsisexhibits reduced resistance, while the SA-insensitive mutant npr-1and the SA-depleted nahG line have no effect on the resistancephenotype [7,23]. However, information is lacking as to whatmakes the two very close members of the Brassicaceae family(B. juncea and S. alba) respond so differently to the same pathogen.We do not have a good hypothesis regarding which responsesadopted by susceptible oilseed mustard plants are actuallyresponsible for disintegrating their defence system to establishcompatibility.
Most of the studies in the field of plant-pathogen interactionfocus on understanding the resistance mechanisms that aredeployed by plants to combat pathogen attack. Hence, we sought tounderstand the mechanism of susceptibility/resistance operative inB. juncea and S. alba against A. brassicicola. We investigated thedifferential response of the phytohormones ABA, SA and JA inB. juncea and S. alba upon challenge with A. brassicicola. Wedistinctly identified different signatures of hormonal interplay in
these two genetically very close members of Brassicaceae familyfollowing challenge by the same pathogen. Here, we provide theexperimental evidence indicating that activation of the SA-mediated signalling pathways by A. brassicicola in the susceptiblebackground is counteracted by an enhanced ABA response in theresistant plant.
2. Results
2.1. Disease progression
As reported earlier [24], the leaves of both S. alba and B. junceashowed visible lesion formation at the site of A. brassicicola sporeapplication at different time points. The rate of lesion formationwas faster in B. juncea than in S. alba, and the rate of hyphal growthfollowed the same trend. The lesion formation appeared to developafter 48 hai (hours after inoculation) and spread over the entire leafwithin 72 hai in B. juncea. Meanwhile, the rate of increase in thelesion diameter was considerably slower in S. alba, and the lesiondeveloped only after 72 hai (Fig. 1A). The fungus began sporulatingslightly earlier in B. juncea leaves than in S. alba, as is evident bytrypan blue staining of the leaves (data not shown) of both plants atdifferent time points. The real-time monitoring of the diseaseprogression by quantifying the 5.8S rRNA gene expression through
M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e51 45
genomic amplification in B. juncea and S. alba leaves at differenttime points on challenge with A. brassicicola indicated, anextremely high rate of hyphal growth in B. juncea compared toS. alba (Fig. 1B). In this connection, it is noteworthy to mention thatour earlier result indicated that when allowing infected plants togrow under sunlight (both infected/control plants of S. alba andB. juncea), the S. alba plants survived and grew well for more than30 days, whereas infected B. juncea plants died within a week [24].
2.2. Differential expression of the hormone-responsive genes inB. juncea and S. alba plants during interaction with A. brassicicola
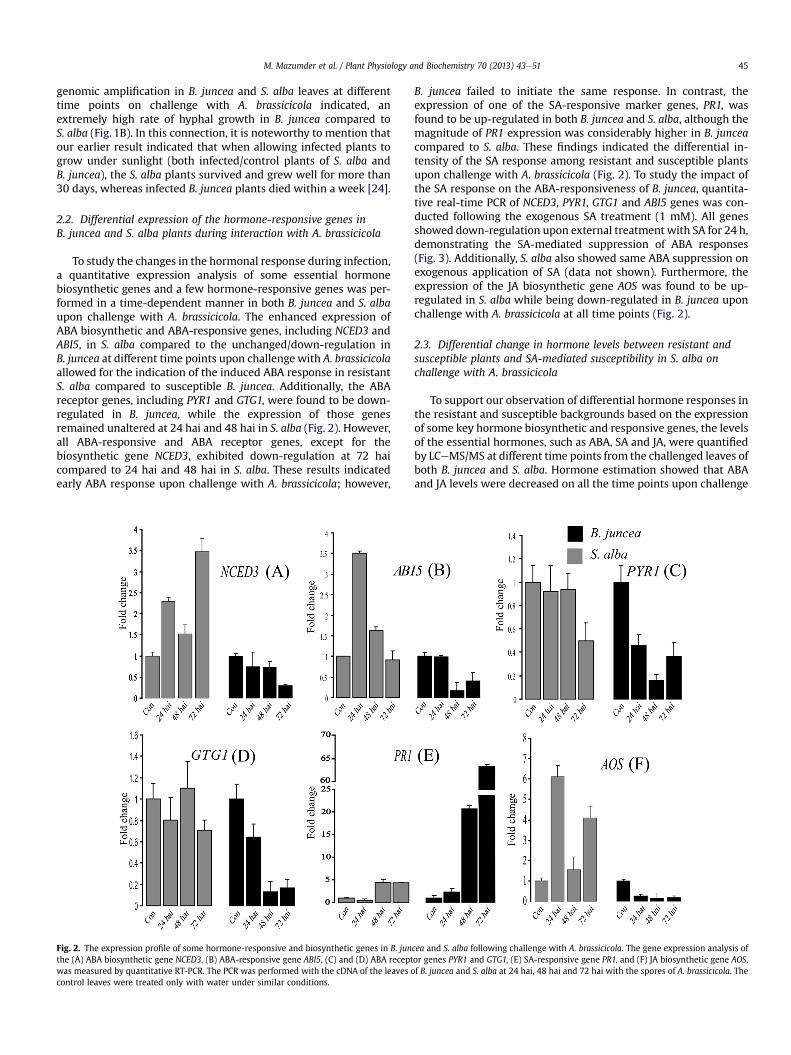
To study the changes in the hormonal response during infection,a quantitative expression analysis of some essential hormonebiosynthetic genes and a few hormone-responsive genes was per-formed in a time-dependent manner in both B. juncea and S. albaupon challenge with A. brassicicola. The enhanced expression ofABA biosynthetic and ABA-responsive genes, including NCED3 andABI5, in S. alba compared to the unchanged/down-regulation inB. juncea at different time points upon challenge with A. brassicicolaallowed for the indication of the induced ABA response in resistantS. alba compared to susceptible B. juncea. Additionally, the ABAreceptor genes, including PYR1 and GTG1, were found to be down-regulated in B. juncea, while the expression of those genesremained unaltered at 24 hai and 48 hai in S. alba (Fig. 2). However,all ABA-responsive and ABA receptor genes, except for thebiosynthetic gene NCED3, exhibited down-regulation at 72 haicompared to 24 hai and 48 hai in S. alba. These results indicatedearly ABA response upon challenge with A. brassicicola; however,
Fig. 2. The expression profile of some hormone-responsive and biosynthetic genes in B. junthe (A) ABA biosynthetic gene NCED3, (B) ABA-responsive gene ABI5, (C) and (D) ABA receptwas measured by quantitative RT-PCR. The PCR was performed with the cDNA of the leaves ocontrol leaves were treated only with water under similar conditions.
B. juncea failed to initiate the same response. In contrast, theexpression of one of the SA-responsive marker genes, PR1, wasfound to be up-regulated in both B. juncea and S. alba, although themagnitude of PR1 expression was considerably higher in B. junceacompared to S. alba. These findings indicated the differential in-tensity of the SA response among resistant and susceptible plantsupon challenge with A. brassicicola (Fig. 2). To study the impact ofthe SA response on the ABA-responsiveness of B. juncea, quantita-tive real-time PCR of NCED3, PYR1, GTG1 and ABI5 genes was con-ducted following the exogenous SA treatment (1 mM). All genesshowed down-regulation upon external treatment with SA for 24 h,demonstrating the SA-mediated suppression of ABA responses(Fig. 3). Additionally, S. alba also showed same ABA suppression onexogenous application of SA (data not shown). Furthermore, theexpression of the JA biosynthetic gene AOS was found to be up-regulated in S. alba while being down-regulated in B. juncea uponchallenge with A. brassicicola at all time points (Fig. 2).
2.3. Differential change in hormone levels between resistant andsusceptible plants and SA-mediated susceptibility in S. alba onchallenge with A. brassicicola
To support our observation of differential hormone responses inthe resistant and susceptible backgrounds based on the expressionof some key hormone biosynthetic and responsive genes, the levelsof the essential hormones, such as ABA, SA and JA, were quantifiedby LCeMS/MS at different time points from the challenged leaves ofboth B. juncea and S. alba. Hormone estimation showed that ABAand JA levels were decreased on all the time points upon challenge
cea and S. alba following challenge with A. brassicicola. The gene expression analysis ofor genes PYR1 and GTG1, (E) SA-responsive gene PR1, and (F) JA biosynthetic gene AOS,f B. juncea and S. alba at 24 hai, 48 hai and 72 hai with the spores of A. brassicicola. The
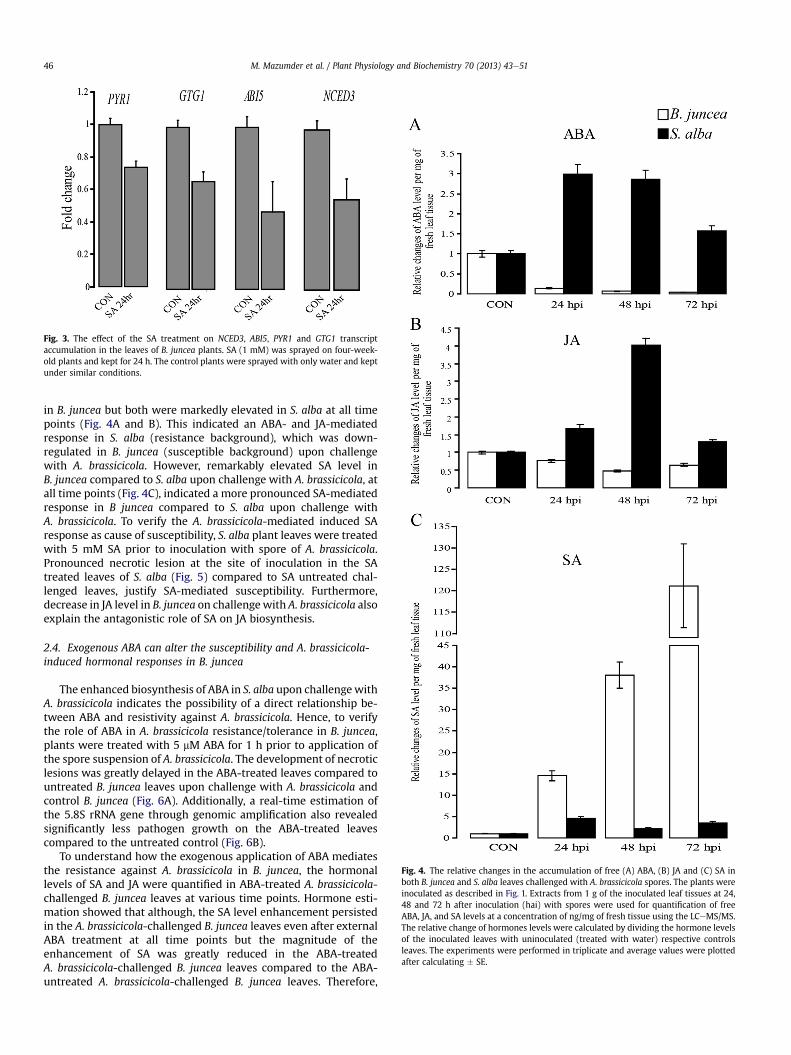
Fig. 4. The relative changes in the accumulation of free (A) ABA, (B) JA and (C) SA inboth B. juncea and S. alba leaves challenged with A. brassicicola spores. The plants wereinoculated as described in Fig. 1. Extracts from 1 g of the inoculated leaf tissues at 24,48 and 72 h after inoculation (hai) with spores were used for quantification of freeABA, JA, and SA levels at a concentration of ng/mg of fresh tissue using the LCeMS/MS.The relative change of hormones levels were calculated by dividing the hormone levelsof the inoculated leaves with uninoculated (treated with water) respective controlsleaves. The experiments were performed in triplicate and average values were plottedafter calculating � SE.
Fig. 3. The effect of the SA treatment on NCED3, ABI5, PYR1 and GTG1 transcriptaccumulation in the leaves of B. juncea plants. SA (1 mM) was sprayed on four-week-old plants and kept for 24 h. The control plants were sprayed with only water and keptunder similar conditions.
M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e5146
in B. juncea but both were markedly elevated in S. alba at all timepoints (Fig. 4A and B). This indicated an ABA- and JA-mediatedresponse in S. alba (resistance background), which was down-regulated in B. juncea (susceptible background) upon challengewith A. brassicicola. However, remarkably elevated SA level inB. juncea compared to S. alba upon challenge with A. brassicicola, atall time points (Fig. 4C), indicated a more pronounced SA-mediatedresponse in B juncea compared to S. alba upon challenge withA. brassicicola. To verify the A. brassicicola-mediated induced SAresponse as cause of susceptibility, S. alba plant leaves were treatedwith 5 mM SA prior to inoculation with spore of A. brassicicola.Pronounced necrotic lesion at the site of inoculation in the SAtreated leaves of S. alba (Fig. 5) compared to SA untreated chal-lenged leaves, justify SA-mediated susceptibility. Furthermore,decrease in JA level in B. juncea on challengewith A. brassicicola alsoexplain the antagonistic role of SA on JA biosynthesis.
2.4. Exogenous ABA can alter the susceptibility and A. brassicicola-induced hormonal responses in B. juncea
The enhanced biosynthesis of ABA in S. alba upon challengewithA. brassicicola indicates the possibility of a direct relationship be-tween ABA and resistivity against A. brassicicola. Hence, to verifythe role of ABA in A. brassicicola resistance/tolerance in B. juncea,plants were treated with 5 mM ABA for 1 h prior to application ofthe spore suspension of A. brassicicola. The development of necroticlesions was greatly delayed in the ABA-treated leaves compared tountreated B. juncea leaves upon challenge with A. brassicicola andcontrol B. juncea (Fig. 6A). Additionally, a real-time estimation ofthe 5.8S rRNA gene through genomic amplification also revealedsignificantly less pathogen growth on the ABA-treated leavescompared to the untreated control (Fig. 6B).
To understand how the exogenous application of ABA mediatesthe resistance against A. brassicicola in B. juncea, the hormonallevels of SA and JA were quantified in ABA-treated A. brassicicola-challenged B. juncea leaves at various time points. Hormone esti-mation showed that although, the SA level enhancement persistedin the A. brassicicola-challenged B. juncea leaves even after externalABA treatment at all time points but the magnitude of theenhancement of SA was greatly reduced in the ABA-treatedA. brassicicola-challenged B. juncea leaves compared to the ABA-untreated A. brassicicola-challenged B. juncea leaves. Therefore,
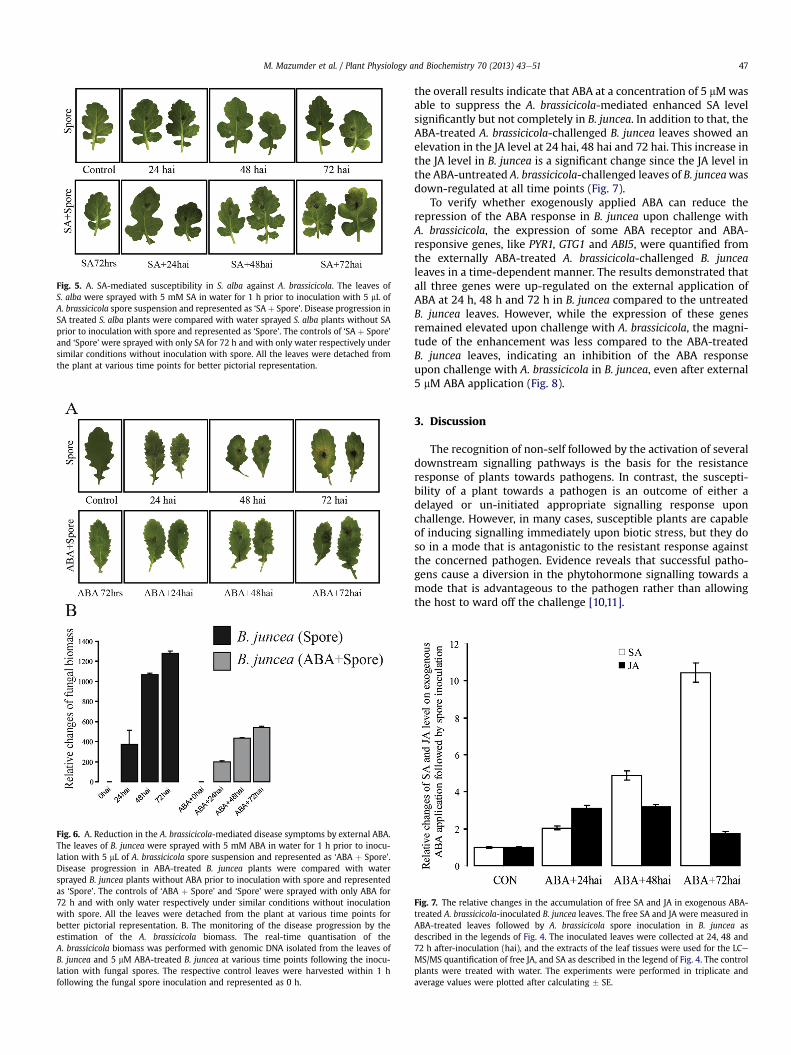
Fig. 6. A. Reduction in the A. brassicicola-mediated disease symptoms by external ABA.The leaves of B. juncea were sprayed with 5 mM ABA in water for 1 h prior to inocu-lation with 5 mL of A. brassicicola spore suspension and represented as ‘ABA þ Spore’.Disease progression in ABA-treated B. juncea plants were compared with watersprayed B. juncea plants without ABA prior to inoculation with spore and representedas ‘Spore’. The controls of ‘ABA þ Spore’ and ‘Spore’ were sprayed with only ABA for72 h and with only water respectively under similar conditions without inoculationwith spore. All the leaves were detached from the plant at various time points forbetter pictorial representation. B. The monitoring of the disease progression by theestimation of the A. brassicicola biomass. The real-time quantisation of theA. brassicicola biomass was performed with genomic DNA isolated from the leaves ofB. juncea and 5 mM ABA-treated B. juncea at various time points following the inocu-lation with fungal spores. The respective control leaves were harvested within 1 hfollowing the fungal spore inoculation and represented as 0 h.
Fig. 5. A. SA-mediated susceptibility in S. alba against A. brassicicola. The leaves ofS. alba were sprayed with 5 mM SA in water for 1 h prior to inoculation with 5 mL ofA. brassicicola spore suspension and represented as ‘SAþ Spore’. Disease progression inSA treated S. alba plants were compared with water sprayed S. alba plants without SAprior to inoculation with spore and represented as ‘Spore’. The controls of ‘SA þ Spore’and ‘Spore’ were sprayed with only SA for 72 h and with only water respectively undersimilar conditions without inoculation with spore. All the leaves were detached fromthe plant at various time points for better pictorial representation.
M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e51 47
the overall results indicate that ABA at a concentration of 5 mMwasable to suppress the A. brassicicola-mediated enhanced SA levelsignificantly but not completely in B. juncea. In addition to that, theABA-treated A. brassicicola-challenged B. juncea leaves showed anelevation in the JA level at 24 hai, 48 hai and 72 hai. This increase inthe JA level in B. juncea is a significant change since the JA level inthe ABA-untreated A. brassicicola-challenged leaves of B. junceawasdown-regulated at all time points (Fig. 7).
To verify whether exogenously applied ABA can reduce therepression of the ABA response in B. juncea upon challenge withA. brassicicola, the expression of some ABA receptor and ABA-responsive genes, like PYR1, GTG1 and ABI5, were quantified fromthe externally ABA-treated A. brassicicola-challenged B. juncealeaves in a time-dependent manner. The results demonstrated thatall three genes were up-regulated on the external application ofABA at 24 h, 48 h and 72 h in B. juncea compared to the untreatedB. juncea leaves. However, while the expression of these genesremained elevated upon challenge with A. brassicicola, the magni-tude of the enhancement was less compared to the ABA-treatedB. juncea leaves, indicating an inhibition of the ABA responseupon challenge with A. brassicicola in B. juncea, even after external5 mM ABA application (Fig. 8).
3. Discussion
The recognition of non-self followed by the activation of severaldownstream signalling pathways is the basis for the resistanceresponse of plants towards pathogens. In contrast, the suscepti-bility of a plant towards a pathogen is an outcome of either adelayed or un-initiated appropriate signalling response uponchallenge. However, in many cases, susceptible plants are capableof inducing signalling immediately upon biotic stress, but they doso in a mode that is antagonistic to the resistant response againstthe concerned pathogen. Evidence reveals that successful patho-gens cause a diversion in the phytohormone signalling towards amode that is advantageous to the pathogen rather than allowingthe host to ward off the challenge [10,11].
Fig. 7. The relative changes in the accumulation of free SA and JA in exogenous ABA-treated A. brassicicola-inoculated B. juncea leaves. The free SA and JA were measured inABA-treated leaves followed by A. brassicicola spore inoculation in B. juncea asdescribed in the legends of Fig. 4. The inoculated leaves were collected at 24, 48 and72 h after-inoculation (hai), and the extracts of the leaf tissues were used for the LCeMS/MS quantification of free JA, and SA as described in the legend of Fig. 4. The controlplants were treated with water. The experiments were performed in triplicate andaverage values were plotted after calculating � SE.
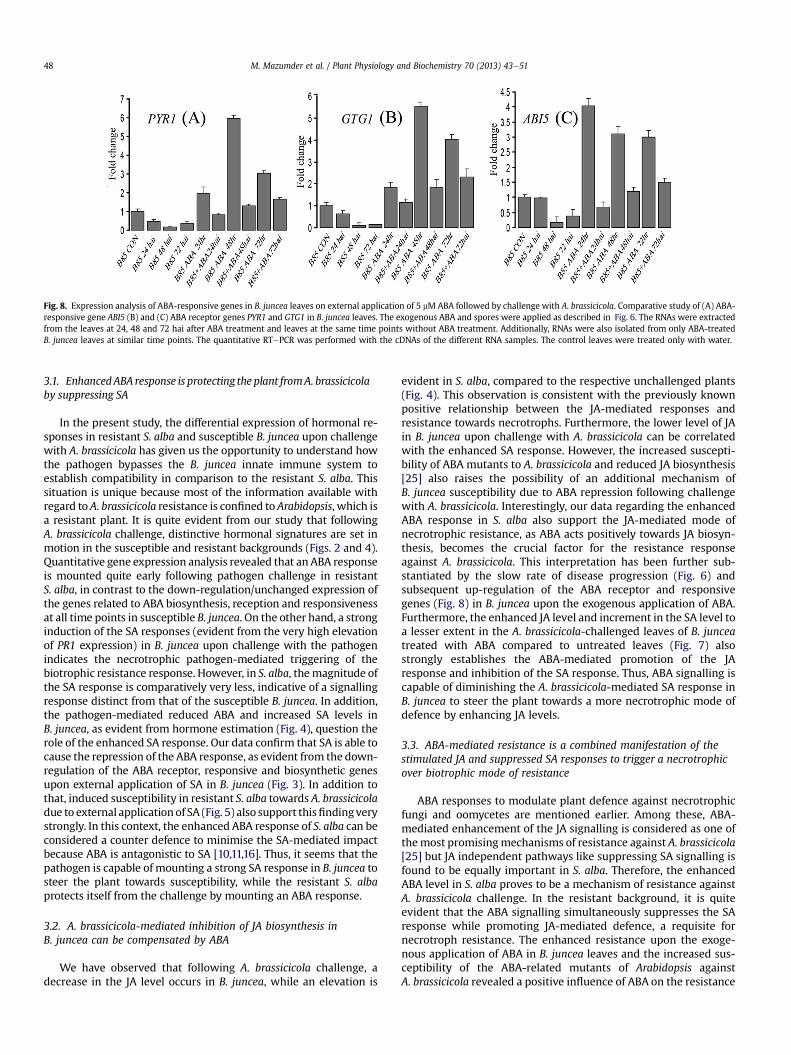
Fig. 8. Expression analysis of ABA-responsive genes in B. juncea leaves on external application of 5 mM ABA followed by challenge with A. brassicicola. Comparative study of (A) ABA-responsive gene ABI5 (B) and (C) ABA receptor genes PYR1 and GTG1 in B. juncea leaves. The exogenous ABA and spores were applied as described in Fig. 6. The RNAs were extractedfrom the leaves at 24, 48 and 72 hai after ABA treatment and leaves at the same time points without ABA treatment. Additionally, RNAs were also isolated from only ABA-treatedB. juncea leaves at similar time points. The quantitative RTePCR was performed with the cDNAs of the different RNA samples. The control leaves were treated only with water.
M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e5148
3.1. EnhancedABA response is protecting the plant fromA. brassicicolaby suppressing SA
In the present study, the differential expression of hormonal re-sponses in resistant S. alba and susceptible B. juncea upon challengewith A. brassicicola has given us the opportunity to understand howthe pathogen bypasses the B. juncea innate immune system toestablish compatibility in comparison to the resistant S. alba. Thissituation is unique because most of the information available withregard to A. brassicicola resistance is confined to Arabidopsis, which isa resistant plant. It is quite evident from our study that followingA. brassicicola challenge, distinctive hormonal signatures are set inmotion in the susceptible and resistant backgrounds (Figs. 2 and 4).Quantitative gene expression analysis revealed that an ABA responseis mounted quite early following pathogen challenge in resistantS. alba, in contrast to the down-regulation/unchanged expression ofthe genes related to ABA biosynthesis, reception and responsivenessat all time points in susceptible B. juncea. On the other hand, a stronginduction of the SA responses (evident from the very high elevationof PR1 expression) in B. juncea upon challenge with the pathogenindicates the necrotrophic pathogen-mediated triggering of thebiotrophic resistance response. However, in S. alba, themagnitude ofthe SA response is comparatively very less, indicative of a signallingresponse distinct from that of the susceptible B. juncea. In addition,the pathogen-mediated reduced ABA and increased SA levels inB. juncea, as evident from hormone estimation (Fig. 4), question therole of the enhanced SA response. Our data confirm that SA is able tocause the repression of the ABA response, as evident from the down-regulation of the ABA receptor, responsive and biosynthetic genesupon external application of SA in B. juncea (Fig. 3). In addition tothat, induced susceptibility in resistant S. alba towards A. brassicicoladue to external applicationof SA (Fig. 5) also support thisfindingverystrongly. In this context, the enhanced ABA response of S. alba can beconsidered a counter defence to minimise the SA-mediated impactbecause ABA is antagonistic to SA [10,11,16]. Thus, it seems that thepathogen is capable of mounting a strong SA response in B. juncea tosteer the plant towards susceptibility, while the resistant S. albaprotects itself from the challenge by mounting an ABA response.
3.2. A. brassicicola-mediated inhibition of JA biosynthesis inB. juncea can be compensated by ABA
We have observed that following A. brassicicola challenge, adecrease in the JA level occurs in B. juncea, while an elevation is
evident in S. alba, compared to the respective unchallenged plants(Fig. 4). This observation is consistent with the previously knownpositive relationship between the JA-mediated responses andresistance towards necrotrophs. Furthermore, the lower level of JAin B. juncea upon challenge with A. brassicicola can be correlatedwith the enhanced SA response. However, the increased suscepti-bility of ABA mutants to A. brassicicola and reduced JA biosynthesis[25] also raises the possibility of an additional mechanism ofB. juncea susceptibility due to ABA repression following challengewith A. brassicicola. Interestingly, our data regarding the enhancedABA response in S. alba also support the JA-mediated mode ofnecrotrophic resistance, as ABA acts positively towards JA biosyn-thesis, becomes the crucial factor for the resistance responseagainst A. brassicicola. This interpretation has been further sub-stantiated by the slow rate of disease progression (Fig. 6) andsubsequent up-regulation of the ABA receptor and responsivegenes (Fig. 8) in B. juncea upon the exogenous application of ABA.Furthermore, the enhanced JA level and increment in the SA level toa lesser extent in the A. brassicicola-challenged leaves of B. junceatreated with ABA compared to untreated leaves (Fig. 7) alsostrongly establishes the ABA-mediated promotion of the JAresponse and inhibition of the SA response. Thus, ABA signalling iscapable of diminishing the A. brassicicola-mediated SA response inB. juncea to steer the plant towards a more necrotrophic mode ofdefence by enhancing JA levels.
3.3. ABA-mediated resistance is a combined manifestation of thestimulated JA and suppressed SA responses to trigger a necrotrophicover biotrophic mode of resistance
ABA responses to modulate plant defence against necrotrophicfungi and oomycetes are mentioned earlier. Among these, ABA-mediated enhancement of the JA signalling is considered as one ofthemost promisingmechanisms of resistance against A. brassicicola[25] but JA independent pathways like suppressing SA signalling isfound to be equally important in S. alba. Therefore, the enhancedABA level in S. alba proves to be a mechanism of resistance againstA. brassicicola challenge. In the resistant background, it is quiteevident that the ABA signalling simultaneously suppresses the SAresponse while promoting JA-mediated defence, a requisite fornecrotroph resistance. The enhanced resistance upon the exoge-nous application of ABA in B. juncea leaves and the increased sus-ceptibility of the ABA-related mutants of Arabidopsis againstA. brassicicola revealed a positive influence of ABA on the resistance

M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e51 49
against necrotrophs [25,26]. Therefore, ABA-mediated resistance inA. brassicicola is a simultaneous expression of both JA-dependentand JA independent pathways as represented in Fig. 9. Further-more, in Arabidopsis no effect of COI1 mutant, a very importantcomponent of JA signalling, on ABA dependent BABA-inducedresistance to A. brassicicola [27] indicate JA independent path-ways are also responsible for resistance response.
It seems that necrotrophic A. brassicicola very cleverly modu-late the phytohormone signalling and diverts it towards the SA(i.e., biotrophic) mode to cause a successful infection in B. juncea.This diversion is in conjunction with the observation wherechallenge with the biotrophic P. syringae that induces SA-mediated defence renders the plants more susceptible to thenecrotrophic A. brassicicola by suppression of the JA signallingpathway [4]. In all likelihood, A. brassicicola deceives the receptor-recognition and, thus, the plant immune system in susceptiblebackgrounds by evoking inappropriate phytohormone signallingto facilitate colonisation. This deception also explains the obser-vation of [10], where even the ectopic expression of a singlebacterial effector (AvrPtoB) of P. syringae induces ABA biosynthesisand signalling (instead of SA signalling) in Arabidopsis to suppressthe host immunity and promote colonisation. Thus, it seemsthat the resistivity/susceptibility to A. brassicicola is ultimatelycontrolled by the regulation of the trade-off and fine tuning be-tween the SA-dependent resistance to biotrophs and the JA-dependent resistance to necrotrophs. This trade-off between thebiotrophic and necrotrophic modes of resistance in plants hasalready been reported in the studies of various pathogens[4,10,11,25].
This indicates operation of a functional ETS (Effectors TriggerSensitivity) by the unknown effector molecule of A. brassicicola topromote SA response in B. juncea is a very promising possibility. Onthe contrary, enhanced ABA response is comparable with the ETI(Effector Trigger Immunity) in S. albawhich allows the recognition/modification of the effectors of A. brassicicola for initiating the SAresponse and simultaneous triggering of ABA response.
Fig. 9. Model illustrating how ABA, SA and JA play a role in the resistance and sus-ceptibility against A. brassicicola. The differential responses in susceptible B. juncea andresistant S. alba on challenge with A. brassicicola, steer the SA response towards sus-ceptibility in B. juncea while providing ABA-mediated resistance to S. alba. The boldarrow indicates the accumulation of the compound in a higher amount. Sharp, solidlines represent stimulatory effects and blunted lines depict antagonistic interactions.
Our observations shed new light on the interplay betweenand balance of phytohormones in the defence response againstA. brassicicola operating in the Brassicaceae family. This study, forthe first time, has functionally demonstrated that the mounting ofthe SA-mediated signalling pathways by A. brassicicola is capable ofrendering a host susceptible. In future, this will enable us toformulate strategies by engineering the function of some of thesecomponents to invigorate the susceptible oilseed mustard tominimise yield loss due to A. brassicicola-mediated black spotdisease.
4. Methods
4.1. Plant and fungal growth conditions
S. alba and B. junceawere grown in a controlled environment ina green house at 24 �C with a relative humidity of 60% under long-day conditions (16 h light and 8 h dark). Spores of A. brassicicolastrain MUCL 20297 (obtained from Willem Broekaert, KatholiekeUniversiteit Leuven, Belgium) were grown in half-strength potatodextrose agar at 25 �C for 20 days [24].
4.2. Fungal inoculation procedure
Plants were inoculated with spores of the fungal pathogenA. brassicicola as described previously by Ghose et al. [24]. Leaves of3-week-old plants were inoculated with 5 ml of spore suspension inwater at a concentration of 4 � 105 spores/mL and kept inside atransparent box at 28 �C at relatively high humidity for the obser-vance of the disease progression and tissue harvesting. The controlplants were treated only with water. The leaves were detachedfrom the plants at different time points after inoculation forphotographic representation of disease progression in bothB. juncea and S. alba.
4.3. External hormone treatment
Water solution of 5 mM and 5 mM of ABA and SA respectivelywere prepared from the 1 M methanolic stock of the respectivehormone from sigma. The water solutions were sprayed to thewhole plant and kept in a control environment as described in theplant growth condition section.
4.4. Fungal biomass estimation
The amount of fungal biomass was quantified by quantitativereal-time PCR using the primers Forward 50-CGGATCTCTTGGTTCTGGCA-30 and Reverse 50-AATGACGCTCGAACAGGCAT-30, designedfrom the genomic sequence of Alternaria sp. 5.8S rRNA region, asdescribed by Kariola et al. [28].
4.5. Transcript level analysis
Transcript levels were analysed by reverse transcription (RT)PCR or by quantitative real-time PCR. Total RNA was isolated fromthe leaves of S. alba and B. juncea using the NucleoSpin RNA Plantkit (MachereyeNagel, Germany), as per the manufacturer’s in-structions. Total RNA was treated extensively with RNase-freeDNaseI provided in the kit to remove any contaminating genomicDNA. The first strand cDNA was synthesised from 2 mg of total RNAusing SuperScript III Reverse Transcriptase (Invitrogen) in a reac-tion volume of 20 ml with the oligo d(T) primer according to themanufacturer’s protocol.

M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e5150
4.6. Identification of the cDNA sequences of oilseed mustardcorresponding to Arabidopsis orthologues from the database andprimer designing
The full-length cDNA sequences of the Arabidopsis orthologueswere used for BLASTX analysis in the Brassica database (www.brassicadb.org/brad/), and the cDNA sequences that showed morethan 70% identity were chosen for the primer design for each gene.In Supplementary Table 1, the accession numbers of individualtranscripts both from the TAIR and Brassica databases arementioned along with the percentage of identity. Primers werechecked through the RT-PCR-mediated amplification of a singleband of appropriate size. The real-time quantitation of the tran-scripts by PCR was performed with the respective primers(Supplementary Table 1) using the appropriately diluted cDNA ofthe control and treated leaf samples.
The real-time PCR was conducted in 96 well blocks with anApplied Biosystem 7500 machine using the SYBR Green I mastermix (Applied Biosystem) according to the manufacturer’s in-structions. The fold change in the expression was calculated fromthe threshold cycle (CT) values by the DDCTmethod as prescribed bythe vendor. The reactions were performed in triplicate in each run,and the level of the ACTIN transcript was used to normalise eachsample. Three independent biological replicates were consideredfor each treatment condition, and the values of each treatment setthat showed statistical significance after calculating SE were usedfor further analysis. All three independent biological replicatesfollowed the same trends of expression change, of which one suchvalue has been represented in this study. The statistical significanceof the values was calculated using Student’s t-test (P < 0.05). Mostof the values were found to be significant; however, in a few cases,they were in the range P � 0.05 � 0.05. The bars in the graphsrepresent the average fold change in expression�mean SE of threereplicates. The changes in expression that were less than two-foldwere not considered significant.
4.7. Hormone estimation
For hormone estimation, leaves of 3-week-old plants wereinoculated with an A. brassicicola spore suspension as described byGhose et al. [24]. The tissues (2 g) from 10 young plants wereharvested at 0 hai, 24 hai, 48 hai and 72 hai. The entire experimentwas performed in triplicate. To estimate the hormone levels in ABA-treated A. brassicicola-challenged plants, the same procedure wasadopted following pre-treatment with 5 mM ABA. The hormoneswere extracted following the protocol of [29].
The stock solutions of the individual PGR standards were pre-pared by accurately weighing 10 � 0.1 mg of each analyte involumetric flasks (certified A class) and dissolving them in 10 mL ofmethanol. These solutions were stored in dark vials at 4 �C. Aworking standardmixture of 1 mg L�1 was prepared by appropriatedilution of the stock solution, fromwhich the calibration standards(0.001e0.25 mg mL�1) were prepared by serial dilutions withmethanol:water (1:1, v/v).
The analysis of the samples was performed using an LCeMS/MSsystem that included Shimadzu UFLCXR HPLC connected to an API5500 Q trap (AB Sciex, Toronto, Canada) mass spectrometer. Elec-trospray ionisation (ESI) in positive and negative polarity modeswas employed in themultiple reactionmonitoring (MRM)mode forthe determination of the PGRs. The source parameters were opti-mised to obtain the molecular ions [M þ H]þ in the positive ion-isation mode and [M � H]- in the negative ionisation mode, whichwas further fragmented in the collision-induced dissociation (CID)mode to obtain the daughter ions. The estimation of JA was per-formed in the positive mode (scan time¼ 50 ms), whereas ABA and
SA were estimated in the negative mode (scan time ¼ 50 ms). Thedetailed mass parameters are given in Supplementary Table 2.
The chromatographic separation was performed on the AtlantisT3 column (100 mm � 2.1 mm ID, 3 mm, Waters India Ltd., Banga-lore). Themobile phasewas composed of (A)methanol:water:formicacid (99:900:1 v/v/v) and (B) methanol:water:formic acid (900:99:1v/v/v); gradient 0.01e1min98%A,1e7min98-5%Aphase, 7e10min5% A phase, 10e11 min 5e98% A phase and 11e15 min 98% A phase.The column oven temperature and mobile phase flow rate weremaintained at 25 �C and 0.4 mL min�1, respectively. An aliquot of10 mL was injected using the Sil-20ACXR prominence auto sampler.
The changes in the hormone levels in ng per gm of fresh tissueswere calculated from the standard curve prepared from theparticular hormone standard. The experiments were performedthree times, and the values of each treatment set that showedstatistical significance after calculating the SE were used forfurther analysis. All three sets of experiments showed similartrends of expression change. The statistical significance of thevalues was calculated using Student’s t-test (P < 0.05). Most of thevalues were found to be significant; however, in a few cases, theywere in the range P � 0.05 � 0.05. The relative changes of thehormone level of each biological replicates were found out bycalculating the ratio of the absolute values of the individual timepoint of each treatment with the absolute values of the respectivecontrol. The graphs represent the average fold change � mean SEof three replicates.
Acknowledgements
The authors are grateful to Dept. of Biotechnology, India forfinancial support and the Director of Bose Institute, Kolkata forproviding the infrastructure and financial assistance to carry outresearch work.
Appendix A. Supplementary data
Supplementary data related to this article can be found athttp://dx.doi.org/10.1016/j.plaphy.2013.04.025.
References
[1] R. Bari, J.D. Jones, Role of plant hormones in plant defence responses, Plant.Mol. Biol. 69 (2009) 473e488.
[2] M.R. Grant, J.D. Jones, Hormone (dis)harmony moulds plant health and dis-ease, Science 324 (2009) 750e752.
[3] C.M.J. Pieterse, D. van der Does, C. Zamioudis, A. Leon-Reyes, S.C.M. van Wees,Hormonal modulation of plant immunity, Annu. Rev. Cell. Dev. Biol. 28 (2012)489e521.
[4] S.H. Spoel, J.S. Johnson, X. Dong, Regulation of tradeoffs between plant de-fenses against pathogens with different lifestyles, Proc. Natl. Acad. Sci. U S A104 (2007) 18842e18847.
[5] J. Glazebrook, Contrasting mechanisms of defense against biotrophic andnecrotrophic pathogens, Annu. Rev. Phytopathol. 43 (2005) 205e227.
[6] J.D. Jones, J.L. Dangl, The plant immune system, Nature 444 (2006) 323e329.[7] B.P.H.J. Thomma, K. Eggermont, I.A.M.A. Penninckx, B. Mauch-Mani,
R. Vogelsang, B.P.A. Cammue, W.F. Broekaert, Separate jasmonate-dependentand salicylate-dependent defense-response pathways in Arabidopsis areessential for resistance to distinct microbial pathogens, Proc. Natl. Acad. Sci. US A 95 (1998) 15107e15111.
[8] A. Koornneef, C.M.J. Pieterse, Cross talk in defense signaling, Plant. Physiol.146 (2008) 839e844.
[9] Z. Chen, J.L. Agnew, J.D. Cohen, P. He, L. Shan, J. Sheen, B.N. Kunkel, Pseudo-monas syringae type III effector AvrRpt2 alters Arabidopsis thaliana auxinphysiology, Proc. Natl. Acad. Sci. U S A 104 (2007) 20131e20136.
[10] M. de Torres-Zabala, W. Truman, M.H. Bennett, G. Lafforgue, J.W. Mansfield,E.P. Rodriguez, L. Bogre, M. Grant, Pseudomonas syringae pv. tomato hijacksthe Arabidopsis abscisic acid signaling pathway to cause disease, EMBO. J. 26(2007) 1434e1443.
[11] M. de Torres-Zabala, M.H. Bennett, W.H. Truman, M.R. Grant, Antagonismbetween salicylic and abscisic acid reflects early host-pathogen conflict andmoulds plant defense responses, Plant. J. 59 (2009) 375e386.

M. Mazumder et al. / Plant Physiology and Biochemistry 70 (2013) 43e51 51
[12] M. de Torres, J.W. Mansfield, N. Grabov, I.R. Brown, H. Ammouneh, G. Tsiamis,A. Forsyth, S. Robatzek, M. Grant, J. Boch, Pseudomonas syringae effectorAvrPtoB suppresses basal defense in Arabidopsis, Plant. J. 47 (2006) 368e382.
[13] A. Robert-Seilaniantz, M. Gran, J.D.G. Jones, Hormone cross-talk in plant dis-ease and defense: more than just JASMONATEeSALICYLATE antagonism,Annu. Rev. Phytopathol. 49 (2011) 317e343.
[14] X.Y. Zheng, N.W. Spivey, W. Zeng, P.P. Liu, Z.Q. Fu, D.F. Klessig, S.Y. He, X. Dong,Coronatine promotes Pseudomonas syringae virulence in plants by activating asignaling cascade that inhibits salicylic acid accumulation, Cell Host Microbe11 (2012) 587e596.
[15] K. Audenaert, G.B. de Meyer, M.M. Hofte, Abscisic acid determines basalsusceptibility of tomato to Botrytis cinerea and suppresses salicylic acid-dependent signaling mechanisms, Plant. Physiol. 128 (2002) 491e501.
[16] P.G. Mohr, D.M. Cahill, Suppression by ABA of salicylic acid and lignin accu-mulation and the expression of multiple genes, in Arabidopsis infected withPseudomonas syringae pv. tomato, Funct. Integr. Genomics. 7 (2007) 181e191.
[17] J. Fan, L. Hill, C. Crooks, P. Doemer, C. Lamb, Abscisic acid has a key role inmodulating diverse plant-pathogen interactions, Plant. Physiol. 150 (2009)1750e1761.
[18] M.Melotto,W.Underwood, J. Koczan, K. Nomura, S.Y. He, Plant Stomata functionin innate immunity against bacterial Invasion, Cell 126 (2006) 969e980.
[19] M. Melotto, W. Underwood, S.Y. He, Role of Stomata in plant innate immunityand Foliar bacterial diseases, Annu. Rev. Phytopathol. 46 (2008) 101e122.
[20] N.C. Turner, A. Graniti, Fusicoccin: a fungal Toxin that opens Stomata, Nature223 (1969) 1070e1071.
[21] R.L. Guimaraes, H.U. Stotz, Oxalate production by Sclerotinia sclerotiorumDeregulates Guard cells during infection, Plant. Physiol. 136 (2004) 3703e3711.
[22] B. Asselbergh, K. Curvers, S.C. Francxa, K. Audenaert, M. Vuylsteke,F.V. Breusegem, M. Hofte, Resistance to Botrytis cinerea in Sitiens, an abscisicacid-deficient tomato mutant, involves timely production of hydrogenperoxide and cell wall modifications in the epidermis, Plant. Physiol. 144(2007) 1863e1877.
[23] S.C.M. van Wees, H.S. Chang, T. Zhu, J. Glazebrook, Characterization of theearly response of Arabidopsis to Alternaria brassicicola infection usingexpression profiling, Plant. Physiol. 132 (2003) 606e617.
[24] K. Ghose, S. Dey, H. Barton, G.J. Loake, D. Basu, Differential profiling of selecteddefense-related genes induced on challenge with Alternaria brassicicola inresistant white mustard and their comparative expression pattern in sus-ceptible India mustard, Mol. Plant Path 9 (2008) 763e775.
[25] B.A.T. Adie, J. Perez-Perez, M.M. Perez-Perez, M. Godoy, J.J. Sanchez-Serrano,E.A. Schmelz, R. Solano, ABA is an essential signal for plant resistance topathogens affecting JA biosynthesis and the activation of defenses in Arabi-dopsis, Plant. Cell. 19 (2007) 1665e1681.
[26] V. Flors, J. Ton, R. Doorn, G. Jakab, P. García-Agustín, B. Mauch-Mani, Interplaybetween JA, SA and ABA signaling during basal and induced resistance againstPseudomonas syringae and Alternaria brassicicola, Plant. J. 54 (2008) 81e92.
[27] J. Ton, B. Mauch-Mani, b-amino-butyric acid-induced resistance againstnecrotrophic pathogens is based on ABA-dependent priming for callose, Plant.J. 38 (2004) 119e130.
[28] T. Kariola, G. Brader, J. Li, E.T. Palva, Chlorophyllase 1, a damage controlenzyme, affects the balance between defense pathways in plants, Plant. Cell.17 (2005) 282e294.
[29] X. Pan, R. Welti, X. Wang, Quantitative analysis of major plant hormones incrude plant extracts by high-performance liquid chromatography-massspectrometry, Nat. Protoc. 5 (2010) 986e992.
Copyright © 2022 FDOKUMEN




















