
Antennal responses of the western pine beetle, Dendroctonus ...
Upload
ucriversideCategory
view
0download
0
Journal of Chemical Ecology, Vol. 17, No. 11, 1991
RESPONSE OF Dendroctonus brevicomis AND Ips paraconfusus (COLEOPTERA: SCOLYTIDAE) TO COMBINATIONS OF SYNTHETIC PHEROMONE
ATTRACTANTS AND INHIBITORS VERBENONE AND IPSDIENOL
T . D . P A I N E * and C . C . H A N L O N
Department of Entomology University of California
Riverside, California 92521
(Received February 8, 1991; accepted July 8, 1991)
Abstract--Dendroctonus brevicomis and Ips paraconfusus are sympatric bark beetle species colonizing Pinus ponderosa in western North America. Inter- specific and intraspecific competition for resources is, in part, mediated through semiochemicals. The response of D. brevicomis to its attractant pher- omone was significantly reduced by simultaneous release of either verbenone or racemic ipsdienol. Trap catch was significantly further reduced by includ- ing both inhibitors with the attractant pheromones. However, although the response of Ips paraconfusus was significantly inhibited with the addition of either verbenone or racemic ipsdienol, both compounds together did not result in a significant further reduction in trap catch. There was a trend for greater reduction in response ofD. brevicomis to attractant pheromones with increased release rates of either 69 % ( +)-/31% ( -)-verbenone or 84 % ( -)- /16 % ( +)- verbenone. Response of associates to attractants and inhibitor combinations was also determined.
Key Words--Dendroctonus brevicomis, Ips paraconfusus, bark beetle, ver- benone, ipsdienol, pheromones, inhibitors, Coleoptera, Scolytidae.
INTRODUCTION
T h e w e s t e r n p ine bee t l e , Dendroctonus brevicomis L e C o n t e , uses a c o m b i n a -
t ion o f b e e t l e - p r o d u c e d p h e r o m o n e s [ ( + ) - e x o - b r e v i c o m i n { (+)-exo-7-ethyl-5- m e t h y l - 6 , 8 - d i o x a b i c y c l o [ 3 . 2 . 1 ] o c t a n e } and ( - ) - f r o n t a l i n { ( - ) - l , 5 - d i m e t h y l -
6 , 8 - d i o x a b i o c y c l o [ 3 . 2 . 1 ] o c t a n e } ] (S i lve r s t e in et a l . , 1968; K i n z e r et a l . , 1969;
* To whom correspondence should be addressed.
2163
0098-0331/91/1100-2163506,50/0 �9 1991 Plenum Publishing Corporation

2164 PAINE AND HANLON
Wood et al., 1976) and host odors, primarily myrcene (Bedard et al., 1969), during the aggregation phase of tree colonization (Wood, 1972). The engraver beetles, Ips paraconfusus Lanier and L pini (Say), are sympatric with D. brev- icomis in California and may colonize the same host material. Ips paraconfusus pheromone consists of three components: (-)-ipsenol[(-)-2-methyl-6-meth- ylene-7-octen-4-ol], (+)-cis-verbenol, and (+)-ipsdienol[(+)-2-methyl-6- methylene-2,7-octadien-4-ol] (Silverstein et al., 1966). The pheromone of / . pini in California was initially described as a single component pheromone, (-)-ipsdienol (Birch et al., 1980a, b); however, there is some suggestion that a second component may be important to complete the pheromone blend (S.A. Teale, personal communication).
The pheromones produced have interspecific effects in addition to the intraspecific effect. Attraction ofL pini to its pheromone is inhibited by racemic ipsenol (Birch and Light, 1977) and to (+)-ipsdienol (Birch et al., 1980a, b). These compounds are components of L paraconfusus aggregation pheromone. Similarly, attraction of L paraeonfusus to its pheromone is inhibited by ( - ) - ipsdienol (Light and Birch, 1979), a component of the aggregation pheromone of L pini. Dendroctonus brevicomis was inhibited by actively boring L para- confusus (Byers and Wood, 1980). Subsequent studies by Byers and Wood (1981a) demonstrated that at least two of the constituents of L paraconfusus pheromone must be present for the inhibitory effect.
Although the constituents of D. brevicomis pheromone function to attract individuals, high release rates of exo-brevicomin or myrcene may also function to interrupt the response (Tilden et al., 1981). Oxidation of tree-produced ot-pinene by either beetles or by microorganisms results in production of ver- benone. This compound found in male D. brevicomis (Byers and Wood, 1980) interrupts the response of D. brevicomis (Bedard et al., 1980) to its attractant pheromone. In addition, verbenone similarly inhibits the attraction response of I. paraconfusus (Byers and Wood, 1980, 1981a).
Colonization of conifers usually occurs as a community process. Although there may be a temporal separation, it would be expected that each species in the community would produce the components of its pheromone with the result- ing potential for interspecific and intraspecific interactions. The objective of the study presented here was to determine how the compounds associated with inhi- bition would effect aggregation of D. brevicomis and L paraconfusus if pre- sented singly or in combination.
METHODS AND MATERIALS
The behavioral responses of D. brevicomis and L paraconfusus to inter- specific and intraspecific chemical signals were examined at two locations dur- ing July and August 1990. Each site had obvious tree mortality, and examination

RESPONSE TO ATTRACTANTS AND INHIBITORS 2165
of the recently killed trees indicated that populations of both D. brevicomis and L paraconfusus were high. While both sites were located within Yellow Pine Forest ecosystems (Munz and Keck, 1973), one site (Julian, San Diego County) was at 1300 m elevation, dominated by Pinus couheri D. Don. and Q. agrifolia Nee near the interface with the Chaparral ecosystem, while the other site (Pal- omar Mountain, San Diego County) was at 1500 m elevation and dominated by mixed conifers.
One of 12 combinations of D. brevicomis attractant [Db = (+)-exo-brev- icomin (released at 2.6 mg/24 hr at 25~ (-)-frontalin (2.0 mg/24 hr), and myrcene (300 mg/24 hr)], L paraconfusus pheromone lip = 50% (+)-/50% (-)-ipsenol (0.2 mg/24 hr), cis-verbenol (2 mg/24 hr), and 95% (+)-/5% ( - ) - ipsdienol (0.2 mg/24 hr)], 16% (+)-/84% (-)-verbenone (10 rag/24 hr), and/ or 50% (+)-/50% (-)-ipsdienol (10 mg/24 hr) was placed on a Lindgren funnel trap (trap and all synthetic compounds from Phero Tech Inc., Delta, British Columbia, Canada). Inhibitors were always tested in combination with one or more of the attractant pheromones. An unbaited trap was also tested as a con- trol. A 3-cm z piece of DDVP-impregnated plastic was placed in the collection cup of each trap to kill any captured beetles. A line of traps spaced at 20-m intervals containing one of each of the 13 treatments constituted a block. Three lines of traps were located in the Julian site and two lines of traps were located at Mount Palomar in two separate tests. Beetles were collected from the traps daily for 12 days, each day constituting a replicate. After each collection, the chemical baits were shifted in a predetermined sequence to a new position within each line. The trapped insects were brought into the laboratory and the numbers of L paraconfusus and D. brevicomis were counted. We also counted Den- droctonus valens LeConte and the predators Enocleris lecontii (Wolcott) (Coleoptera: Cleridae) and Temnochila chlorodia (Mannerheim) (Coleoptera: Trogositidae).
Verbenone has been reported as an inhibitor of both L paraconfusus and D. brevicomis. However it is not clear whether either or both enantiomers of verbenone are effective at inhibiting the response to the attractant pheromones. We tested a mixture of 16% (+)-/84% (-)-verbenone released at five elution rates at the Mount Palomar site during August 1990. Funnel traps were baited with one of six treatments: D. brevicomis pheromone [exo-brevicomin (released at 2.6 mg/24 hr at 24~ frontalin (2.0 mg/24 hr), and myrcene (300 mg/24 hr)] alone and D. brevicomis pheromone plus the verbenone enantiomers eluted at 0.01 mg/24 hr (at 25~ 0.18 mg/24 hr, 1.8 mg/24 hr, 5 mg/24 hr, and 20 mg/24 hr. As in the previous test, an unbaited trap served as a control. Each of the seven treatments was replicated 10 times. Beetles were collected from the traps three times at 72-hr intervals. An identical test was subsequently con- ducted using 69 % (+)-/31% (-)-verbenone in the same location.
Differences in trap catches among the 13 treatments at each location for

2166 PAINE AND HANLON
each beetle species, as well as the differences in the response of D. brevicomis among release rate treatments for an enantiomeric blend were determined using General Linear Modeling procedures (PROC GLM) and Duncan's new multi- pie-range test on SAS for microcomputers (SAS Institute, 1988).
RESULTS
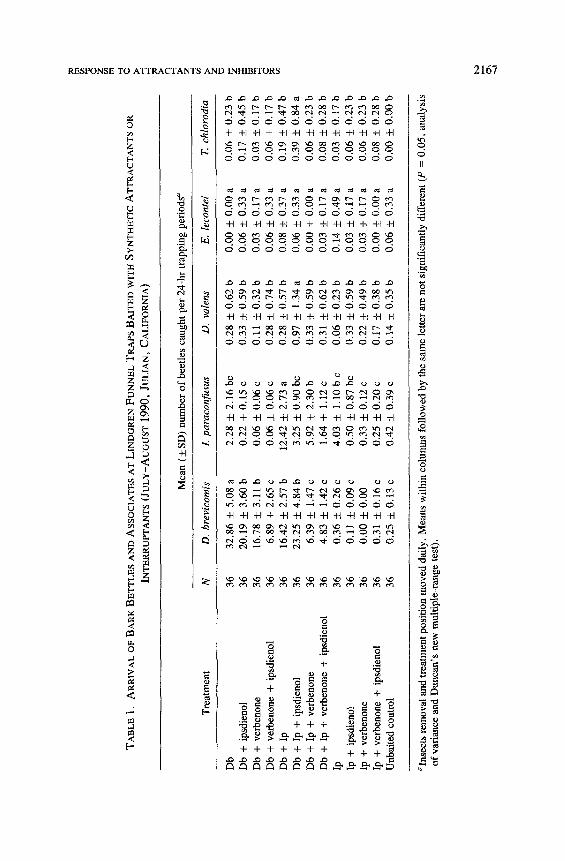
At both study sites, the traps baited with D. brevicomis pheromone (Db) alone caught significantly more of that species than any of the other treatments [Julian: F = 18.41, d f = 12, P = 0.0001 (Table 1); Palomar: F = 13.99, d f = 12, P = 0.0001 (Table 2)]. Four treatments [Db and verbenone, Db and ipsdienol, Db and Ips paraconfusus pheromone (Ip) and Db and Ip and ips- dienol] at both sites inhibited the response of D. brevicomis to the attractant pheromone, but still caught more beetles than the blank control traps. The three remaining treatments (Db and Ip and verbenone, Db and verbenone and ips- dienol, and Db and Ip and verbenone and ipsdienol) further reduced catches of D. brevicomis to levels not significantly different from either the unbaited con- trol or to those treatments lacking the aggregation pheromone (Db) of this spe- cies.
The arrival of L paraconfusus at traps at the Mount Palomar location was significantly inhibited (F = 8.34, d f = 12, P = 0.0001) by the addition of verbenone and ipsdienol to treatments that included the aggregation pheromones (Table 2). The aggregation pheromones of this species alone and in combination with the aggregation pheromones from D. brevicomis were the only two treat- ments that were significantly different from the responses to the unbaited con- trol. Although the response of L paraconfusus was significantly greater (F = 6.87, dd = 12, P = 0.0001) to the aggregation pheromones of both L para- confusus and D. brevicomis together, the response to the aggregation phero- mone of L paraconfusus alone was not different from the unbaited control or any of the inhibitor treatments at the Julian location (Table 1).
While there were some beetles caught in the traps, there were no significant differences in the arrival of E. lecontii (F = 0.76, d f = 12, P = 0.2799) to any of the 13 treatments tested in the first study at Julian, but the response of both D. valens (F = 4.27, d f = 12, P = 0.0001) and T. chlorodia (F = 3.25, d f = 12, P = 0.0002) was significantly greater to the E-M-F and [50% (+)- / 50 % ( -)- ipsenol , cis-verbenol, and 95 % (+)- /5 % ( -)-ipsdienol] and ipsdienol treatment (Table 1). The response of D. valens at the Mount Palomar location was not significantly different to any treatment (F = 0.90, d f = 12, P = 0.5519). However, there were significant differences observed for the response of the other two associates to the same treatments at the Mount Palomar site [E. lecontii (F = 2.06, d f = 12, P = 0.0197), T. chlorodia (F = 2.12, d f =
TA
BL
E 1
. A
RR
IVA
L O
F B
AR
K B
ET
TL
ES
AN
D A
SSO
CIA
TE
S A
T L
IND
GR
EN
FU
NN
EL
TR
APS
BA
ITE
D W
ITH
SY
NT
HE
TIC
AT
TR
AC
TA
NT
S O
R
INT
ER
RU
PTA
NT
S (J
uL
Y-A
UG
US
T 1
99
0,
JUL
IAN
, C
AL
IFO
RN
IA)
~o
�9
z -]
O
>
,d
Mea
n (+
SD
) n
um
ber
of
beet
les
caug
ht p
er 2
4-hr
tra
ppin
g pe
riod
s"
Tre
atm
ent
N
D.
brev
icom
is
L pa
raco
nfus
us
D.
vale
ns
E.
leco
ntei
T.
chl
orod
ia
r ,q
>
z >
Db
36
32.8
6 +
5.
08 a
2.
28
+
2.16
be
0.28
+
0.
62 b
0.
00
+
0.00
a
0.06
+
0.
23 b
D
b +
ip
sdie
nol
36
20.1
9 __
+ 3.
60 b
0.
22
+
0.15
c
0.33
+
0.
59 b
0.
06
+
0.33
a
0.17
+
0.
45 b
D
b +
ver
beno
ne
36
16.7
8 +
3,
11 b
0,
06 +
0.
06 c
0.
11
+
0.32
b
0.03
+
0.
17 a
0.
03
+
0.17
b
Db
+
verb
enon
e +
ip
sdie
nol
36
6.89
+
2.
65 c
0.
06
• 0.
06 c
0.
28
• 0.
74 b
0.
06
+
0.33
a
0.06
+
0.
17 b
D
b +
Ip
36
16.4
2 •
2.57
b
12.4
2 +
2.
73 a
0.
28
5:0
.57
b
0.08
•
0.37
a
0.19
•
0.47
b
Db
+
Ip +
ip
sdie
nol
36
23.2
5 •
4.84
b
3.25
•
0.90
bc
0.97
+
1.
34 a
0.
06
• 0.
33 a
0.
39
• 0.
84 a
D
b +
Ip
+
verb
enon
e 36
6.
39
5:1
.47
c
5.92
5
:2.3
0 b
0.
33
• 0.
59 b
0.
00
+
0.00
a
0.06
5
:0.2
3 b
D
b +
Ip
+
verb
enon
e +
ip
sdie
nol
36
4.83
•
1.42
c
1.64
+
1.
12 c
0.
31
+
0.62
b
0.03
+
0.
17 a
0.
08
• 0.
28 b
Ip
36
0
.36
• 4
.03
• 1
.10
bc
0
.06
• 0
.14
• 0.
03
• Ip
+
ipsd
ieno
l 36
0.
11
• 0.
09 c
0.
50
5:0
.87
bc
0.33
5
:0.5
9 b
0.
03
• 0.
17 a
0.
06 +
0.
23 b
Ip
+
verb
enon
e 36
0.
00 5
:0.0
0
0.33
•
0.12
c
0.22
5
:0.4
9 b
0.
03
• 0.
17 a
0.
06
• 0.
23 b
Ip
+
verb
enon
e +
ip
sdie
nol
36
0,31
•
0.16
c
0.25
5
:0.2
0 c
0.
17
+
0.38
b
0.00
•
0.00
a
0.08
•
0.28
b
Unb
aite
d co
ntro
l 36
0.
25
• 0,
13 c
0.
42
• 0.
39 c
0.
14 •
0.
35 b
0.
06
• 0.
33 a
0.
00
• 0.
00 b
-r
~In
sect
s re
mov
al a
nd t
reat
men
t po
siti
on m
ov
ed d
aily
. M
eans
wit
hin
colu
mns
fol
low
ed b
y th
e sa
me
lett
er a
re n
ot s
igni
fica
ntly
dif
fere
nt (
P =
0.0
5, a
naly
sis
of
vari
ance
and
Du
nca
n's
new
mul
tipl
e-ra
nge
test
).
to

t~
oQ
TA
BL
E 2
. A
RR
IVA
L O
F B
AR
K B
EE
TL
ES
AN
D A
SSO
CIA
TE
S A
T L
IND
GR
EN
FU
NN
EL
TR
APS
BA
ITE
D W
ITH
SYN
THE
TIC
AT
TR
AC
TA
NT
S O
R
INT
ER
RU
PTA
NT
S (J
uL
Y-A
uG
uS
T 1
99
0,
MT
. PA
LO
MA
R,
CA
LIF
OR
NIA
)
Mea
n (
• S
D)
nu
mb
er o
f b
ettl
es c
aug
ht
per
24-
h tr
app
ing
per
iod
a
Tre
atm
ent
N
D.
brev
icom
is
1. p
arac
onfu
sus
D.
vale
ns
E.
leco
ntei
T.
chl
orod
ia
Db
24
23
5.3
3
+
43
,58
a
0.0
0 _
+ 0
.00
b
0.4
6 +
0
.93
a
0.0
0 +
0
.00
b
0.2
9
• 0
.75
a b
c
Db
+
ipsd
ien
ol
24
72.2
1 +_
_ 16
.09
b c
0.2
5
+
0.21
b
0.5
0
• 1.
29 a
0
.08
•
0.2
8 b
0
.46
•
0.8
3 a
b
Db
+
ver
ben
on
e 23
7
3.9
6 +
3
4,1
4 b
c 0
.00
+
0.0
0 b
0
.18
•
0.6
6 a
0
.05
•
0.21
b
0.0
9
__. 0
.29
c
Db
+ v
erb
eno
ne
+
ipsd
ien
ol
22
16.0
9 •
2.7
2 d
0
.14
•
0.1
4 b
0
.27
+
0
.88
a
0.1
8 +
0
.59
b
0.41
+
1.
05 a
b c
Db
+ I
p 24
10
6.42
+
2
3.4
6 b
11
.54
+
3.6
2 a
0
.38
+
1.
44 a
0
.08
•
0.2
8 b
0
.83
•
1,31
a
Db
+
Ip +
ip
sdie
no
l 2
4
73
.92
•
12.5
2 b
c 1.
79 •
0
.54
b
0.7
9
• 1.
35 a
0
.13
•
0.3
4 b
0
.67
•
1.09
a b
Db
+
Ip +
v
erb
eno
ne
24
2
4.1
7 _
_+ 5
.75
cd
3.5
0 •
0
.85
b
0.3
8
+
1.44
a
0.0
4 •
0
.20
b
0.5
4 •
0
.93
a b
c
Db
+
Ip +
v
erb
eno
ne
+
ipsd
ien
ol
23
20
.78
+
5
.14
cd
1.57
•
0.6
2 b
0
.57
+
0
.95
a
0.0
0 +
0
.00
b
0.3
0 •
0
.56
a b
c
Ip
24
0
.79
+
0.
31 d
12
,50
• 3
.64
a
0.0
8
• 0
.28
a
0.0
0 •
0
,00
b
0.2
9
+
0.8
6 a
b c
Ip +
ip
sdie
no
l 23
0.
61
• 0
.26
d
1.13
•
0.6
2 b
0
.35
•
1.78
a
0.2
2
___
0.6
0 b
0
.17
•
0.3
9 b
c
Ip +
ve
rben
one
20
1.
05
+
0.7
0 d
1
,60
•
0.7
4 b
0
.70
•
2.6
8 a
0
.55
•
1.43
a
0.5
0 •
1.
00 a
b c
Ip
+
ver
ben
on
e +
ip
sdie
nol
23
0.1
7
• 0
.10
d
0.1
7
• 0
.14
b
0.0
9
• 0
.29
a
0.2
2
+__ 0
.60
b
0.0
0 •
0
.00
c
Unb
aite
d co
ntr
ol
24
1.17
•
0.3
2 d
0
,08
•
0.0
8 b
0.
21
• 0.
51 a
0
.04
•
0.2
0 b
0
.08
•
0.2
8 c
a Ins
ects
rem
ov
al a
nd
tre
atm
ent p
osi
tio
n m
ov
ed d
aily
. M
ean
s w
ith
in c
olu
mn
s fo
llo
wed
by
the
sam
e le
tter
are
no
t si
gn
ific
antl
y d
iffe
rent
(P
=
0.0
5,
anal
ysi
s of
var
ian
ce a
nd D
un
can
's n
ew m
ult
iple
-ran
ge
test
).
r~
7.
z z

RESPONSE TO ATTRACTANTS AND INHIBITORS 2169
12, P = 0.0159), Table 2]. Individuals of these three species were also caught in traps baited with D. brevicomis aggregation pheromone and different release rates of verbenone. The response of D. valens was significantly less (F = 3.60, df = 6, P = 0.0021) when any 69% (+)-/31% (-) -verbenone treatment was included, but there were no significant differences among treatments for E. lecontii (F = 0.96, df = 6, P = 0.4519) or T. chlorodia (F = 1.71, df = 6, P = 0.1199) (Table 3). In contrast, when 16% (+)- /84% (-) -verbenone was tested, catch of D. valens was significantly greater than the unbaited control (F = 2.37, df = 6, P = 0.0311) for only the lowest release rate (Table 4). Response of E. lecontii was significantly different from the control (F = 2.72, df = 6, P = 0.0147) at the highest release rate, and response of T. chlorodia was not different to any verbenone treatment (F = 1.37, df = 6, P = 0.2287).
The response of D. brevicomis to the aggregation pheromone alone was significantly greater (F = 15.06, df = 6, P = 0.0001) than to any of the treatments containing the 69 % (+)- /31% (- ) -verbenone (Table 3). There was no clear dose-response relationship, but all treatments attracted more beetles than did the unbaited control. In contrast, the responses ofD. brevicomis to the two lowest release rates of 16% (+)- /84% (-) -verbenone were not signifi- cantly different (F = 17.57, df = 6, P = 0.0001) than the response to the aggregation pheromone alone (Table 4). However, traps baited with the lowest release rate of this enantiomeric blend of verbenone plus aggregation phero- mone caught significantly more beetles than did the treatment with the next lowest release rate. As in the previous test of enantiomers, all treatments attracted more beetles than did the unbaited control traps.
D I S C U S S I O N
Our results confirm that the response of D. brevicomis to its aggregation pheromone is reduced by the presence of L paraconfusus pheromone, racemic ipsdienol, or verbenone, but also demonstrate that the presence of multiple inhibitor compounds further reduces the response. While it is not clear why the response of L paraconfusus to its aggregation pheromone was not different (P = 0.05) from the control treatment at the Julian site, the reduction in the response at the Mt. Palomar location was not significantly enhanced by the addition of more than one inhibitory compound. However, at the Julian loca- tion, there was a tendency for the treatments with either D. brevicomis or L paraconfusus pheromones alone or both pheromones together plus only a single inhibitor to have higher trap catches. The response of the associates was gen- erally very low to any of the treatments. However, Wood et al. (1968) dem- onstrated that E. lecontii responds to pheromone constituents of L paraconfusus, while Vit6 and Pitman (1969) found that T. chlorodia responded to D. brevi- comis pheromone.

TA
BL
E 3
. A
RR
IVA
L O
F B
AR
K B
EE
TL
ES
AN
D A
SSO
CIA
TE
S A
T L
IND
GR
EN
FU
NN
EL
TR
APS
BA
ITE
D W
ITH
FR
ON
TA
LIN
, ex
o-BR
EVIC
OM
IN, A
ND
M
YR
CE
NE
, PL
US
DIF
FER
EN
T R
EL
EA
SE R
AT
ES
OF
69 %
(+
)-/3
1%
(--)
-VE
RB
EN
ON
E (
AuG
uST
-SE
PTE
MB
ER
19
90
, M
T.
PA
LO
MA
R,
CA
LIF
OR
NIA
)
Mea
n (
+
SD
) n
um
ber
of
bee
tles
cau
gh
t p
er 3
6-h
r tr
app
ing
per
iod
a
Tre
atm
ent
N
D.
brev
icom
is
L pa
raco
nfus
us
D.
vale
ns
E.
leco
ntei
T.
chl
orod
ia
Db
3O
Db
+ v
erb
eno
ne
at 0
.01
rag
/24
hr
29
Db
+
verb
enon
e at
0.1
8 r
ag/2
4 h
r 3
0
Db
+
ver
ben
on
e at
1.8
mg
/24
hr
30
Db
+ v
erb
eno
ne
at 5
rag
/24
hrs
29
Db
+
ver
ben
on
e at
20
rag
/24
hr
30
Unb
aint
ed c
ontr
ol
28
193.
90 +
2
9.4
3 a
0
.03
_
0.1
8 a
1.
27 +
2
.75
a
0.4
7
+
2.0
3 a
0
.07
+_
0.2
5 a
106.
03
+
15.1
2 b
0.0
7 +
0
.26
a
0.4
8 +
0
.95
b
0.1
7 +
0
.47
a
0.1
7
+
0.4
7 a
94
.47
+
15
.58
bc
0.1
3
+
0.7
3 a
0
.37
+
1.
00 b
0
.10
+
0.3
1 a
0
.07
+
0.2
5 a
120.
10 +
2
1.2
3 b
0
.23
+
1.
10 a
0
.10
+
0.31
b
0.1
0 +
0.
31 a
0
.03
+
0
.18
a
53
.66
+
i0
.26
c
0.6
6+
1
.72
a 0.
31
4- 0
.71
b 0
.03
+
0
.19
a
0.0
3
+
0.1
9a
98
.50
4-
19.3
5 b
c 0
.63
4-
1.9
6 a
0.0
7
+
0.2
5 b
0
.07
4-
0.2
5 a
0
.00
4-
0.0
0 a
0
.39
+
0.11
d
0.6
4 _
_+_ 1
.97
a 0
.07
_
0.2
6 b
0
.07
+
0
.26
a
0.0
0 +
0
.00
a
'~In
sect
s re
mo
ved
eve
ry t
hree
day
s fr
om
10
loca
tio
ns
per
tre
atm
ent.
Nu
mb
ers
wit
hin
co
lum
ns
foll
ow
ed b
y th
e sa
me
lett
ers
are
not
sig
nif
ican
tly
dif
fere
nt
(P =
0
.05
, an
aly
sis
of
var
ian
ce a
nd
Du
nca
n's
mu
ltip
le-r
ang
e te
st).
z >
z >
z r-'
�9

TA
BL
E 4.
A
RR
IVA
L O
F B
AR
K B
EE
TL
ES
AN
D A
SSO
CIA
TE
S A
T L
IND
GR
EN
FU
NN
EL
TR
AP
S B
AIT
ED
WIT
H F
RO
NT
AL
IN,
exo-
BR
EV
ICO
MIN
, AN
D
MY
RC
EN
E,
PL
US
DIF
FE
RE
NT
RE
LE
AS
E R
AT
ES
OF
16 %
(+)-
/84
%
(--)
-(A
uG
usT
-SE
PT
EM
BE
R 1
99
0,
MT
. P
AL
OM
AR
, C
AL
IFO
RN
IA)
Mea
n (
+
SD
) n
um
ber
of
bee
tles
cau
gh
t p
er 3
6-hr
tra
pp
ing
per
iod
a
Tre
atm
ent
N
D.
brev
icom
is
L pa
raco
nfus
us
D.
vale
ns
E, l
econ
tei
T. c
hlor
odia
Db
3O
Db
+
ver
ben
on
e at
0.0
1 m
g/2
4 h
r 30
Db
+
verb
enon
e at
0,1
8 m
g/2
4 h
r 30
Db
+
verb
enon
e at
1,8
mg
/24
hr
30
Db
+
verb
enon
e at
5 m
g/2
4 h
r 30
Db
+
ver
ben
on
e at
20
mg
/24
hr
30
Un
bai
nte
d c
ontr
ol
30
27
8.2
7 _
+ 32
.95
ab
0.0
3
_ 0
.18
a
1,20
+
1,81
ab
0.3
3
+
0.61
ab
0.2
7
+
0.4
5 a
3
10
.60
+
37.1
5 a
0.0
0 _
0
.00
a
1.90
+
3.5
4 a
0
.30
•
0.6
5 a
b 0
.33
+
0
.80
a
21
0.7
3
_ 30
.72
bc
0.0
0 +
0
.00
a
1.20
+
2.7
3 a
b 0
.07
•
0.2
5 b
0
,23
+
0
.50
a
148.
07
• 2
1.4
4 c
0
.00
•
0.0
0 a
0
.87
•
1.50
ab
0.3
3
• 0
.99
ab
0.1
7
• 0
.46
a
171.
30 •
31
.38
c 0
.20
+
1.10
a
0,6
3
+
1.07
b
0,1
0 :
t: 0.
31 b
0
.17
•
0.3
8 a
163,
53
• 2
2.2
4 c
0
.20
+
0.9
2 a
0
.83
+
1.
64 a
b 0
.60
+
1.
07 a
0
.10
__
_ 0.
31 a
0.9
3
+
0.3
7 d
0
.00
•
0.0
0 a
0
.03
+
0
.18
b
0.0
3
• 0
.18
b
0,0
3
_ 0
.18
a
aln
sect
s re
mo
ved
ev
ery
thr
ee d
ays
fro
m 1
0 lo
cati
on
s p
er t
reat
men
t. N
um
ber
s w
ith
in c
olu
mn
s fo
llo
wed
by
the
sam
e le
tter
s ar
e n
ot
sig
nif
ican
tly
dif
fere
nt
(P 0
.05,
an
aly
sis
of
var
ian
ce a
nd
Du
nca
n's
mu
ltip
le-r
ang
e te
st).
b.9
-,,,3

2172 PAINE AND HANLON
The process of tree colonization by D. brevicomis takes place within a very short time and is mediated by attractant semiochemicals (Wood, 1972). How- ever, there is the potential for competition among the arriving beetles for the limited amount of available resource (Alcock, 1982; Berryman et al., 1985). One of the mechanisms used by the beetles appears to be a complex coordina- tion of timing in release of attractants and production of inhibitory compounds (Byers et al., 1984). Consequently, the arriving beetles are spaced to maximize individual fitness (Schlyter et al., 1987; DeJong and Sabelis, 1988).
The recognition of verbenone as an inhibitor occurs in several species within the Dendroctonus and Ips genera, Pitman et al. (1969) identified ver- benone from D. brevicomis, D. frontalis Zimmermann, and D. ponderosae Hopkins. Subsequent investigations demonstrated that the compound inhibits arrival of beetles to attractive sources (Bedard et al., 1980; Ryker and Yandell, 1983; Payne and Billings, 1988). Ips paraconfusus and L typographus (L.) were also shown to be inhibited by verbenone (Byers and Wood, 1980, 1981a; Bakke, 1981).
It is not common to observe individuals of a single bark beetle species colonizing a tree. Rather it is usually a community of species (Paine et al., 1980). The interactions among the members of the community may be mediated by the pheromone communication system (Birch et al., 1980a, b; Smith et al., 1990). However, several beetle species may produce the same chemical com- pounds and exploit the inhibitor message to reduce competition. Male D. brev- icomis and male D. ponderosae produce (+)-ipsdienol from myrcene (Renwick et al., 1976; Byers, 1982; Hunt et al., 1986). This enantiomer inhibits conspe- cifics (Byers, 1982; Hunt and Borden, 1988) as well as L pini (Birch et al., 1980a, b), a potential competitor of either species. Thus, D. brevicomis and D. ponderosae release two compounds (verbenone and ipsdienol) that have inter- specific and intraspecific inhibitory effects. The effect of releasing the inhibitors simultaneously has not been observed previously for a Dendroctonus species. There was a suggestion from the studies of Bakke (1981) that the release of two inhibitory compounds was more effective for inhibiting the response of L typo- graphus than the release of either compound alone, but subsequent work could not confirm those results (Schlyter et al., 1988).
The chemical components of the pheromone blends have multifunctional effects; they may function as attractant pheromones, inhibitory pheromones, or as synomones. The insects may produce the compounds, or production may be the result of microbial action on either ingested substrate or exposure to volatile substrates. Brand et al. (1975) isolated a bacterium from L paraconfusus that utilized ot-pinene to synthesize cis- and trans-verbenol. Byers and Wood (1981b) reported that streptomycin treatments of L paraconfusus prevented the conver- sion of myrcene to ipsenol and ipsdienol. In contrast, L typographus produces cis- and trans-verbenol but requires associated yeasts to convert these com- pounds to verbenone (Leufven et al., 1984; Leufven and Birgersson, 1987).

RESPONSE TO A T T R A C T A N T S AND INHIBITORS 2173
The yeasts were unable to produce either the verbenols or verbenone directly from a-pinene (Leufven et al., 1988). Similarly, D. ponderosae requires a gut microorganism to produce verbenone (Hunt and Borden, 1989).
There is an obvious ecological advantage to both arriving and colonizing beetles to avoid either marginal resources or intense competition. Verbenone, derived from ot-pinene through the intermediate verbenols by microbial action in several systems, appears to be a common signal that the resource may be attacked and potentially unsuitable for further colonization (Schlyter et al., 1988). The terpene alcohols, ipsenol and ipsdienol, may also be interspecific and intraspecific inhibitors that serve to reduce detrimental competition. Light et al. (1983) demonstrated that interspecific competition between L paracon- fusus and L pini produced greater negative effects than intraspecific competition and speculated that these competitive interactions may be the selective force that resulted in mutual pheromone inhibition between the species. The geo- graphic range of D. brevicomis is within the range of Pinus ponderosa Lawson. Ips pini and D. brevicomis are sympatric in much of that range, and the enan- tiomer of ipsdienol produced by D. brevicomis is an effective inhibitor of I. pini where sympatry occurs (Birch et al., 1980a, b). It is, however, a constituent of the attractant pheromone of L paraconfusus that overlaps with D. brevicomis in only a portion of the geographic range.
While parallel studies have not been completed for L paraconfusus and D. brevicomis, the competitive interactions may be similarly detrimental. How- ever, it is not clear why the presence of multiple inhibitors produced a greater effect on D. brevicomis than L paraconfusus. It is possible that this Ips species is more sensitive to the presence of the inhibitors or the described pheromone blend for this species is incomplete, and as a result, the maximum reduction was observed to any single compound. There is a difference between L para- confusus and L typographus. The results of Bakke (1981) suggest that attraction of L typographus is inhibited more by a combination of ipsenol plus verbenone than by either compound alone. However, in the European spruce system, this species may be more of an ecological homologue to D. brevicomis than to L paraconfusus in westem North America.
Acknowledgments--We thank J. Trumble, R. Redak, P. Luft, T. Grasswitz, P. Leddy, S. Bertram, and M. Moratorio for helpful reviews of the manuscript; Phero Tech Inc. (Delta, British Columbia, Canada) for the release devices used in the enantiomeric dose test; and D. Messore, S. Gilbert, M. Goeden, and S. Bertram for data collection. This study was supported in part by a grant from the California Department of Forestry and Fire Protection.
REFERENCES
ALCOCK, J. 1982. Natural selection and communication among bark beetles. Fla. Entomol. 65:17- 32.
BAKKE, A. 1981. Inhibition of the response in Ips typographus to the aggregation pheromone; field evaluation of verbenone and ipsenol. Z. Angew. Entomol. 92:172-177.

2174 PAINE AND HANLON
BEDARD, W.D., TILDEN, P.E., WOOD, D.L., SILVERSTEIN, R.M., BROWNLEE, R.G., and RODIN, J.O. 1969. Western pine beetle: Field responses to its sex pheromone and a synergistic host terpene, myrcene. Science 164:1284-1285.
BEDARD, W.D., TILDEN, P.E., LINDAHL, K.Q., JR., WOOD, D.L., and RAUCH, P.A. 1980. Effects of verbenone and trans-verbenol on the response of Dendroctonus brevicomis to natural and synthetic attractant in the field. J. Chem. Ecol. 6:997-1013.
BERRYMAN, A.A., DENNIS, B., RAFFA, K.F., and STENSETH, N.C. 1985. Evolution of optimal group attack with particular reference to bark beetles (Coleoptera: Scolytidae). Ecology 66:898- 903.
BIRCH, M.C., and LIGHT, D.M. 1977. Inhibition of the attractant pheromone response in lps pini and I. paraconfusus (Coleoptera: Scolytidae): Field evaluation of ipsenol and linalool. J. Chem. Ecol. 3:257-267.
BIRCH, M.C., LIGHT, D.M., WOOD, D.L., BROWNE, L.E., SILVERSTEIN, R.M., BERGOT, B.J., OHLOFF, G., WEST, J.R., and YOUNG, J.C. 1980a. Pheromonal attraction and allomonal inter- ruption of lps pini in California by the two enantiomers of ipsdienol. J. Chem. Ecol. 6:703- 717.
BIRCH, M.C., SVIHRA, P., PAINE, T.D., and MILLER, J.C. 1980b. Influence of chemically mediated behavior on host tree colonization by four cohabiting species of bark beetles. J. Chem. Ecol. 6:395-414.
BRAND, J.M., BRACKE, J.W., MARKOVETZ, A.J., WOOD, D.L., and BROWNE, L.E. 1975. Produc- tion of verbenol pheromone by bacterium isolated from bark beetles. Nature 254:136-137.
BYERS, J.A. 1982. Male-specific conversion of the host plant compound, myrcene, to the phero- mone, (+)-ipsdienol, in the bark beetle, Dendroctonus brevicomis. J. Chem. Ecol. 8:363- 371.
BYERS, J.A., and WOOD, D.L. 1980. Interspecific inhibition of the response of the bark beetles, Dendroctonus brevicornis and Ips paraconfusus, to their pheromones in the field. J. Chem. Ecol. 6:149-163.
BYERS, J.A., and WOOD, D.L. 1981a. Interspecific effects of pheromones on the attraction of the bark beetles, Dendroctonus brevicomis and Ips paraconfusus, in the laboratory. J. Chem. Ecol. 7:9-18.
BYERS, J.A., and WOOD, D.L. 1981b. Antibiotic-induced inhibition of pheromone synthesis in a bark beetle. Science 213:763-764.
BYERS, J.A., WOOD, D.L., CRAIG, J., and HENDRY, L.B. 1984. Attractive and inhibitory phero- mones produced in the bark beetle, Dendroctonus brevicomis, during host colonization: Reg- ulation of inter- and intraspecific competition. J. Chem. Ecol. 10:861-877.
DEJONG, M.C.M., and SABELIS, M.W. 1988. HOW bark beetles avoid interference with squatters: an ESS for colonization by lps typographus. Oikos 51:88-96.
HUNT, D.W.A., and BORDEN, J.H. 1988. Response of mountain pine beetle, Dendroctonus pon- derosae Hopkins, and the pine engraver, Ips pini (Say), to ipsdienol in southwestern British Columbia. J. Chem. Ecol. 14:277-293.
HUNT, D.W.A., and BORDEN, J.H. 1989. Terpene alcohol pheromone production by Dendroctonus ponderosae and lps paraconfusus (Coleoptera: Scolytidae) in the absence of readily culturable microorganisms. J. Chem. Ecol. 15:1433-1463.
HUNT, D.W.A., BORDEN, J.H., PIERCE, H.D., JR., SLESSOR, K.N., KING, G.G.S., and CZY- ZEWSKA, E.K. 1986. Sex-specific production of ipsdienol and myrcenol by Dendroctonus pon- derosae (Coleoptera: Scolytidae) exposed to myrcene vapors. J. Chem. Ecol. 12:1579-1586.
KINZER, G.W., FENTIMAN, A.F., PAGE, T.F., FOLTZ, R.L., VIT~, J.P., and PITMAN, G.B. 1969. Bark beetle attractants: Identification, synthesis, and field bioassay of a new compound iso- lated from Dendroctonus. Nature 221:477-478.

RESPONSE TO ATTRACTANTS AND INHIBITORS 2175
LIGHT, D.M., and BIRCH, M.C. 1979. Inhibition of the attractant pheromone response in Ips par- aconfusus by (R)-(-)-ipsdienol. Naturwissenschafien 66:159-160.
LIGHT, D.M., BIRCH, M.C., and PAINE, T.D. 1983. Laboratory study of intraspecific and inter- specific competition within and between two sympatric bark beetle species, Ips pini and L paraconfusus. Z. Angew. Entomol. 96:233-241.
LEUEVEN, A., and BIRGERSSON, G. 1987. Quantitative variation of different monoterpenes around galleries of Ips typographus (Coleoptera: Scolytidae) attacking Norway spruce. Can. J. Bot. 65:1038-1044.
LEUFVEN, A., BERGSTROM, G., and FALSEN, E. 1984. Interconversion of verbenols and verbenone by identified yeasts isolated from the spruce bark beetle lps typographus. J. Chem. Ecol. 10:1349-1361.
LEUFVEN, A., BERGSTROM, G., and FALSEN, E. 1988. Oxygenated monoterpenes produced by yeasts, isolated from Ips typographus (Coleoptera: Scolytidae) and grown in phloem medium. J. Chem. Ecol. 14:353-362.
MUNZ, P.A., and KECK, D.A. 1973. A California Flora. University of California Press, Berkeley. 1681 pp.
PAINE, T.D., BIRCH, M.C., and SVIHRA, P. 1980. Niche breadth and resource partitioning by four sympatric species of bark beetles (Coleoptem: Scolytidae). Oecologia 48:1-6.
PAYNE, T.L., and BILLINGS, R.F. 1988. Effect of pheromone and pheromone plus tree cutting on suppression of southern pine beetle infestations, pp. 267-273, in T.L. Payne and H. Saaren- maa (eds.). Integrated Control of Scolytid Bark Beetles. Virginia Polytechnic Institute and State University, Blacksburg, Virginia.
PITMAN, G.B., VIT~, J.P., KINZER, G.W., and FENTIMAN, A.F. 1969. Specificity of population- aggregating pheromones in Dendroctonus. J. Insect Physiol. 15:363-366.
RENWICK, J.A.A., HUGHES, P.R., PITMAN, G.B., and VITE, J.P. 1976. Oxidation products of terpenes identified from Dendroctonus and Ips bark beetles. J. Insect Physiol. 22:725-727.
RYKER, L.C., and YANDELL, K.L. 1983. Effect of verbenone on aggregation of Dendroctonus ponderosae Hopkins (Coleoptera: Scolytidae) to synthetic attractant. Z. Angew. Entomol. 96:452-459.
SAS Institute. 1988. SAS/STAT User's Guide, Release 6.03 Edition. Cary, North Carolina. 1028
PP. SCHLYTER, F., BYERS, J.A., and LOFQVIST, J. 1987. Attraction to pheromone sources of different
quantity, quality, and spacing: Density-regulation mechanisms in bark beetle Ips typographus. J. Chem. Ecol. 13:1503-1523.
SCHLYTER, F., BYERS, J.A., LOFQVlST, J., LEUFVEN, A., and BIRGERSSON, G. 1988. Reduction of attack density of bark beetles Ips typographus and Tomicus piniperda on host bark by verben- one inhibition of attraction to pheromone and host kairomone, pp. 53-68, in T.L. Payne and H. Saarenmaa (eds.). Integrated Control of Scolytid Bark Beetles. Virginia Polytechnic Insti- tute and State University, Blacksburg, Virginia.
SILVERSTEIN, R.M., RODIN, J.O., and WOOD, D.L. 1966. Sex attractants in frass produced by male lps confusus in ponderosa pine. Science 154:509-510.
SILVERSTEIN, R.M., BROWNLEE, R.G., BELLAS, T.E., WOOD, D.L., and BROWNE, L.E. 1968. Brevicomin: Principal sex attractant to the frass of the female western pine beetle. Science 159:889-890.
SMITH, M.T., PAYNE, T.L., and BIRCH, M.C. 1990. Olfactory-based behavioral interactions among five species in the southern pine bark beetle group. J. Chem. Ecol. 16:3317-3331.
TILDEN, P.E., BEDARD, W.D., WOOD, D.L., and STUBBS, H.A. 1981. Interruption of response of Dendroctonus brevicomis to its attractive pheromone by components of the pheromone. J. Chem. Ecol. 7:183-196.

2176 PAINE AND HANLON
VIT~, J.P., and PITMAN, G.B. 1969. Insect and host odors in the aggregation of the western pine beetle. Can. Entomol. 101:113-117.
WOOD, D.L. 1972. Selection and colonization of ponderosa pine by bark beetles, pp. 101-117, in H.F. van Emden (ed.). Insect/Plant Relationships. Symposium, Royal Entomology Society, London. No. 6.
WOOD, D.L., BROWNE, L.E., BEDARD, W.O., TILDEN, P.E., SILVERSTEIN, R.M., and RODIN, J.O. 1968. Response of Ips confusus to synthetic sex pheromones in nature. Science 159:1373- 1374.
WOOD, D.L., BROWNE, L.E., EWING, B., LINDAHL, K., BEDARD, W.D., TILDEN, P.E., MORI, K., PITMAN, G.B., and HUGHES, P.R. 1976. Western pine beetle: Specificity among enantiomers of male and female components of an attractant pheromone. Science 192:896-898.
Copyright © 2022 FDOKUMEN














