
Reproduction and feeding of Sagitta enflata in the Humboldt Current system off Chile
10
Reproduction and feeding of Sagitta enflata in the Humboldt Current system off Chile Ricardo Giesecke and Humberto E. Gonza ´lez Giesecke, R., and Gonza ´lez, H. E. 2008. Reproduction and feeding of Sagitta enflata in the Humboldt Current system off Chile. – ICES Journal of Marine Science, 65: 361–370. This study is based on 3 years (August 2002 – July 2005) of monthly zooplankton sampling at a fixed station located 18 km off Coliumo Bay (368S), Chile. The reproduction of Sagitta enflata, its feeding rate, specific daily ration, and prey selectivity were analysed and related to several environmental variables: temperature, salinity, dissolved oxygen, chlorophyll a concentrations, and both meso- and micro- zooplankton abundance. The main predatory activity of S. enflata was centred on the copepods Paracalanus parvus, Oithona spp., and Calanus chilensis. These three species were consumed at different rates, depending mainly on the maturity of the S. enflata population. When Stages I and II individuals dominated, predation focused on the small P. parvus and Oithona spp., whereas more mature populations (Stages III and IV) preyed selectively on C. chilensis. The mean specific daily ration of 0.1 d 21 increased to 0.5 d 21 before and during matu- ration. The reproductive phase of S. enflata was closely coupled with the abundance of nauplii, suggesting that chaetognath reproduction paralleled that of copepods, most likely to diminish the mortality of its offspring resulting from starvation. The relationships between these were included in a conceptual model and their ecological significance is discussed. Keywords: chaetognaths, Humboldt Current system, reproduction, Sagitta enflata. Received 11 July 2007; accepted 30 January 2008; advance access publication 18 March 2008. R. Giesecke: Centre for Oceanographic Research in the Eastern South Pacific (COPAS), Concepcio ´n, Chile, and Graduate Programme in Oceanography, Department of Oceanography, Universidad de Concepcio ´n, Concepcio ´n, Chile. H. E. Gonza ´lez: Centre for Oceanographic Research in the Eastern South Pacific (COPAS), Concepcio ´n, Chile, and Institute of Marine Biology “Ju ¨rgen Winter”, Universidad Austral de Chile, Valdivia, Chile. Correspondence to R. Giesecke: tel: þ56 63 22 1559; fax: þ56 63 22 1455; e-mail: [email protected]. Introduction Many studies demonstrate that fecundity and development in chaetognaths are functions of temperature (Sameoto, 1971; Conway and Williams, 1986). Therefore, latitude may be a major factor in determining the number of generations produced in an annual cycle (Ghirardelli, 1968). In general, the number of gener- ations increases as the distance from the poles becomes greater (Owre, 1960). The association between these two variables can only exist under conditions in which food resources—both in terms of quality and quantity—are not limiting. Poor food quality or quantity would lead to a low or negative growth rate (Reeve, 1970) as a result of the lack of carbon or nitrogen resources available for sustaining the metabolic demands of growth and reproduction (e.g. the starvation of mature Sagitta hispida results in a cessation of reproductive activity and weight loss; Reeve and Cosper, 1975). Despite a significant effort to determine the reproductive pulses of chaetognaths (Alvarin ˜o, 1965), no attempt has been made to explain the processes favouring repro- duction. The latter is crucially important because it could also explain the interannual discrepancies in periodicity and duration of the chaetognath breeding and life cycles observed in several studies (McLaren, 1966, 1969; Khan and Williamson, 1970; Conway and Williams, 1986; Sameoto, 1987). There is some evi- dence that reproduction of chaetognaths may be linked to the increase of copepod abundance during spring and summer (Zo, 1973; King, 1979; Ohman, 1986), suggesting a dependence on prey for reproduction. The processes involved in chaetognath maturation and breed- ing are key to understanding their population dynamics; such information may allow some predictability of chaetognath popu- lations. The goal of this paper is to identify and evaluate the abiotic (temperature, salinity, dissolved oxygen concentrations) and biotic (prey availability, feeding behaviour) variables respon- sible for the maturation of chaetognaths in the central Humboldt Current system (HCS). Sagitta enflata was selected for this purpose. It is the dominant species in the HCS and one of the most abundant and widely distributed epipelagic chaetognath species in the world oceans (Alvarin ˜o, 1965). The study was conducted in the HCS, a major eastern boundary current system with one of the highest primary production rates measured for coastal environments (4 –20 g C m 22 d 21 ; Daneri et al., 2000) and high seasonal heterogeneity. The biological pro- ductivity of this system is mainly supported by the intrusion of high-nutrient and oxygen-depleted equatorial subsurface water (ESSW) during austral spring and summer when northerly winds prevail. During austral autumn and winter, increased river run-off and decreased upwelling periodicity promote a system dominated by small diatoms and net heterotrophy. These characteristics make this system especially interesting for studying biological associations and dynamics that are coupled # 2008 International Council for the Exploration of the Sea. Published by Oxford Journals. All rights reserved. For Permissions, please email: [email protected] 361
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Reproduction and feeding of Sagitta enflata in the Humboldt Current system off Chile

Reproduction and feeding of Sagitta enflata in the HumboldtCurrent system off Chile
Ricardo Giesecke and Humberto E. Gonzalez
Giesecke, R., and Gonzalez, H. E. 2008. Reproduction and feeding of Sagitta enflata in the Humboldt Current system off Chile. – ICES Journal ofMarine Science, 65: 361–370.
This study is based on 3 years (August 2002 –July 2005) of monthly zooplankton sampling at a fixed station located 18 km off ColiumoBay (368S), Chile. The reproduction of Sagitta enflata, its feeding rate, specific daily ration, and prey selectivity were analysed and relatedto several environmental variables: temperature, salinity, dissolved oxygen, chlorophyll a concentrations, and both meso- and micro-zooplankton abundance. The main predatory activity of S. enflata was centred on the copepods Paracalanus parvus, Oithona spp.,and Calanus chilensis. These three species were consumed at different rates, depending mainly on the maturity of the S. enflata population.When Stages I and II individuals dominated, predation focused on the small P. parvus and Oithona spp., whereas more mature populations(Stages III and IV) preyed selectively on C. chilensis. The mean specific daily ration of 0.1 d21 increased to 0.5 d21 before and during matu-ration. The reproductive phase of S. enflata was closely coupled with the abundance of nauplii, suggesting that chaetognath reproductionparalleled that of copepods, most likely to diminish the mortality of its offspring resulting from starvation. The relationships between thesewere included in a conceptual model and their ecological significance is discussed.
Keywords: chaetognaths, Humboldt Current system, reproduction, Sagitta enflata.
Received 11 July 2007; accepted 30 January 2008; advance access publication 18 March 2008.
R. Giesecke: Centre for Oceanographic Research in the Eastern South Pacific (COPAS), Concepcion, Chile, and Graduate Programme inOceanography, Department of Oceanography, Universidad de Concepcion, Concepcion, Chile. H. E. Gonzalez: Centre for OceanographicResearch in the Eastern South Pacific (COPAS), Concepcion, Chile, and Institute of Marine Biology “Jurgen Winter”, Universidad Austral deChile, Valdivia, Chile. Correspondence to R. Giesecke: tel: þ56 63 22 1559; fax: þ56 63 22 1455; e-mail: [email protected].
IntroductionMany studies demonstrate that fecundity and development inchaetognaths are functions of temperature (Sameoto, 1971;Conway and Williams, 1986). Therefore, latitude may be a majorfactor in determining the number of generations produced in anannual cycle (Ghirardelli, 1968). In general, the number of gener-ations increases as the distance from the poles becomes greater(Owre, 1960). The association between these two variables canonly exist under conditions in which food resources—both interms of quality and quantity—are not limiting. Poor foodquality or quantity would lead to a low or negative growth rate(Reeve, 1970) as a result of the lack of carbon or nitrogen resourcesavailable for sustaining the metabolic demands of growth andreproduction (e.g. the starvation of mature Sagitta hispidaresults in a cessation of reproductive activity and weight loss;Reeve and Cosper, 1975). Despite a significant effort to determinethe reproductive pulses of chaetognaths (Alvarino, 1965), noattempt has been made to explain the processes favouring repro-duction. The latter is crucially important because it could alsoexplain the interannual discrepancies in periodicity and durationof the chaetognath breeding and life cycles observed in severalstudies (McLaren, 1966, 1969; Khan and Williamson, 1970;Conway and Williams, 1986; Sameoto, 1987). There is some evi-dence that reproduction of chaetognaths may be linked to theincrease of copepod abundance during spring and summer (Zo,
1973; King, 1979; Ohman, 1986), suggesting a dependence onprey for reproduction.
The processes involved in chaetognath maturation and breed-ing are key to understanding their population dynamics; suchinformation may allow some predictability of chaetognath popu-lations. The goal of this paper is to identify and evaluate theabiotic (temperature, salinity, dissolved oxygen concentrations)and biotic (prey availability, feeding behaviour) variables respon-sible for the maturation of chaetognaths in the central HumboldtCurrent system (HCS). Sagitta enflata was selected for thispurpose. It is the dominant species in the HCS and one of themost abundant and widely distributed epipelagic chaetognathspecies in the world oceans (Alvarino, 1965). The study wasconducted in the HCS, a major eastern boundary currentsystem with one of the highest primary production ratesmeasured for coastal environments (4–20 g C m22 d21; Daneriet al., 2000) and high seasonal heterogeneity. The biological pro-ductivity of this system is mainly supported by the intrusion ofhigh-nutrient and oxygen-depleted equatorial subsurface water(ESSW) during austral spring and summer when northerlywinds prevail. During austral autumn and winter, increasedriver run-off and decreased upwelling periodicity promote asystem dominated by small diatoms and net heterotrophy.These characteristics make this system especially interesting forstudying biological associations and dynamics that are coupled
# 2008 International Council for the Exploration of the Sea. Published by Oxford Journals. All rights reserved.For Permissions, please email: [email protected]
361

to large-scale responses such as the El Nino Southern Oscillationand the Pacific Decadal Oscillation, and can provide some insightinto possible responses to ongoing climate change.
Material and methodsOceanographic propertiesThe study was conducted at Station 18 (St. 18) off Coliumo Bay(36830’S 73807’W; Figure 1). Sampling was conducted approxi-mately monthly between August 2002 and July 2005. Sampleswere taken twice a month during favourable weather (September2002 to April 2004) and partially halted at the end of australautumn and beginning of winter owing to unfavourable weather.For each sample, water temperature, conductivity, and oxygenprofiles were obtained with a CTD-O (SeaBird SBE 19 Plus).Variables such as temperature, salinity, and dissolved oxygen con-centrations were analysed as mean values of the integrated watercolumn (0–80 m). Upwelling events were identified using theBakun upwelling index (Bakun, 1978). Wind data (velocity, direc-tion), recorded four times a day every 6 h (from 02:00 to 20:00),were obtained from the weather station at Carriel Sur airport.The mean daily zonal wind intensity (m s21) was used to calculatethe daily mean wind drag coefficient and windstress according toWu (1982). The mean daily Ekman offshore transport (Bakunupwelling index, Mx) was calculated for the 100 m coastal line as:
Mx ¼ ðt y=rf Þ � 100;
where ty is windstress, r the water density, and f the Coriolisparameter.
The Bakun upwelling index was calculated for each samplingdate using 5 d of mean daily water transport (m3 s21) databefore the sampling date. Only positive upwelling events thatexceeded 10 m3 s21 were considered (Parada et al., 2001).
Zooplankton collection and analysisZooplankton samples were collected using a Tucker trawl zoo-plankton net (200 mm mesh size; 1 m2 mouth area); a WP2(once) or a Hensen net (six times) was used when the Tuckernet malfunctioned. All the samples were collected during the dayso that feeding patterns would not be biased by differential dayand night feeding preferences. In all, 38 integrated oblique nettows were done from the bottom to the surface (ca. 0–80 m),and the volume filtered was quantified using a calibrated digital
flowmeter. Zooplankton samples were fixed almost immediatelyin 10% tetraborate buffered formaldehyde-seawater to minimizethe loss of prey in the chaetognath guts as a result of regurgitationor defaecation (Baier and Purcell, 1997). The fixed samples wereanalysed to estimate copepod and chaetognath abundance; thewhole sample was usually analysed although, in some cases, sub-samples were taken with a Folsom splitter. Sagitta enflata weresorted from the samples, measured (head to tail, excluding thecaudal fin) to the nearest 0.2 mm, and the development stageidentified based on Alvarino’s (1967) classification (Table 1).The proportion of S. enflata containing prey (FCR) and numberof prey per S. enflata (NPC) were also noted (Feigenbaum andMaris, 1984). To account for possible codend feeding, all preyfound in the foregut (the upper quarter of the gut) wereomitted from the gut content analysis (Feigenbaum and Maris,1984). Gut contents of S. enflata were assessed by dissectionunder a stereomicroscope and were further analysed under aninverted microscope (�400 magnification) following the method-ology of Øresland (1987). In almost all cases (ca. 90%), foodremains were unrecognizable; the hard parts of copepod prey wereidentified using the mandible blades (Giesecke and Gonzalez,2004a). Appendicularians and chaetognaths in gut contents wererecognized by faecal pellets and grasping spines, respectively.Copepod prey sizes were estimated from the relationship betweenmandible width (MW) and prosome length (PL), as used byGiesecke and Gonzalez (2004a). On some occasions, large, uniden-tified mandible blades were found in chaetognath guts; these werelater identified as belonging to Calanus chilensis. The relationshipbetween MW and PL of this species was determined to estimatethe length of the C. chilensis ingested by S. enflata.
The feeding rate on total and individual prey by species itemwas calculated from the equation proposed by Bajkov (1935):
FR ¼ ðNPC� 24ÞDT
;
where the daily feeding rate, or the mean number of prey ingestedper 24 h (FR), is calculated from the mean number of preyingested per chaetognath (NPC) and the digestion time (DT) inhours.
. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Table 1. Maturity stage classification of S. enflata according toAlvarino (1967).
Male gonads Female gonads
Stage I No visible testes orvesicles
Ovaries reaching quarter or halfthe length of the posterior finsalong the trunk
StageII
Easily visible testes;seminal vesicles beginningto appear
Ovaries reaching to aboutmid-length of the extent ofposterior fins on the trunk ornear the anterior end of theposterior fins
StageIII
Testes filled with sperm;seminal vesicles beginningto fill
Ovaries nearly reaching theanterior end of the posteriorfins or the posterior end of theanterior fins
StageIV
Seminal vesicles full,bursting, or already empty
Ovaries reaching the anteriorend of the posterior finsFigure 1. Location of time-series Station 18.
362 R. Giesecke and H. E. Gonzalez

The DT of S. enflata was estimated from the equation proposedby Ohman (1986):
DT ¼ 10:48e�0:086T ;
where T is water temperature (8C), estimated, in this case, as theaverage of the integrated water column (0–80 m) for eachsampling date.
The specific daily ration (SDR) of S. enflata was calculated as
SDRd ¼ FRcd=Wsd;
where FRcd (mg C ind.21 d21) is the mean feeding rate in carbonweight of S. enflata during each sampling period (d), and Wsd
is the mean carbon weight of S. enflata at each sampling(mg C ind.21). The SDR was only estimated in cases in whichthe number of animals with food in their guts exceeded ten indi-viduals per sample.
The mean feeding rate in carbon weight (FRcd) was estimatedfrom
FRcd ¼ FRnd
XfdjWpj;
where FRnd is the mean feeding rate during each sampling (d;prey ind.21 d21), fdj the frequency of prey ( j) in the gut of S.enflata on date (d), and Wpj the carbon weight of prey ( j; Saitoand Kiørboe, 2001).
The mean carbon content per copepod prey item was esti-mated, based on the PL to carbon conversion proposed by Uye(1991) for Paracalanus parvus, Uye (1982) for Centropages brachia-tus, Escribano et al. (1997) for C. chilensis, Berggreen et al. (1988)for Acartia tonsa, Sabatini and Kiørboe (1994) for Oithona spp.,and Satapoomin (1999) for Oncaea spp. and Corycaeus spp. Themean value of the PL of each copepod species was obtainedfrom Hidalgo and Escribano (2007) and P. Hidalgo (pers.comm.). For large, unidentified copepods, we assumed a carboncontent of 51 mg, whereas, for small unidentified copepods, weassumed a carbon content equivalent to that of a 550 mm lengthP. parvus. We used a carbon content of 5.17 mg for Oikopleuraspp., based on the mean trunk length of 971 mm (I. Andrade,pers. comm.) and a carbon:length conversion proposed by Kinget al. (1980).
Dry weight and carbon content of S. enflataSpecimens of S. enflata in good condition with empty gutswere isolated from natural samples according to their develop-mental stage, placed on preweighed GFF filters, and dried at608C for 24 h to measure dry weight (DW) with a precision of+10 mg. Carbon content was estimated according to the carbon:dry weight ratio of Batistic (2003) as
mgC ¼ DWðmgÞ � 0:35:
SelectivitySelectivity of S. enflata for ingested prey was estimated using thePearre’s selectivity index C (Pearre, 1982). This index relies onx2 analysis of the number of individual prey items in predatorguts, compared with those available in the environment; the
index was calculated as
C ¼+ðjadbe � bdaej � n=2Þ2
abde
" #1=2
:
The terms of the equation are defined in Table 2.The selectivity C values were calculated from the relative
abundance (percentage) of each zooplankton prey item ingested inrelation to the total prey standing stock, where a and b represent therelative abundance of a particular species (A in Table 2) and allothers, respectively, and subscripts d and e indicate the diet and theenvironment. The sign of the selectivity index is given by the inspec-tion of adbe2aebd. The index ranges from 21 to 1. Positive valuesindicate selection, negative values indicate avoidance, and a value ofzero indicates no selection. Statistical significance was tested withthe x2 statistic with one degree of freedom (Pearre, 1982) at a signifi-cance level of 0.05. For the analysis, we only considered sampling dateson which at least ten prey were identified in chaetognath guts.
Maturity and reproductionWe considered that reproduction took place when Stages III andIV were present, meaning that maximal oocyte and testis develop-ment was achieved. The percentage of Stages III and IV of theentire population was used as a reproduction index and comparedwith environmental variables (temperature, salinity, dissolvedoxygen) through Spearman correlations. Biological variablesincluded total prey abundance (including all prey items), chloro-phyll a concentration—Chl a (mg m22, used as a productivityproxy), unicellular microzooplankton abundance, and naupliiabundance (ind. m22). The effect of feeding activity on reproduc-tion, expressed as FCR and NPC, was also considered.
Abundance of prey for S. enflata offspringMicrozooplankton samples were collected before each zooplank-ton net haul at three depths (10, 30, and 80 m) using 30 l Niskinbottles. These samples were concentrated (10–30 l) by passingthem through a 20 mm sieve (final volume of 100 ml). Theywere preserved in buffered formalin (4%) and later analysed forgeneral taxonomic composition and abundance using an invertedmicroscope Olympus CK2 at �400 (Utermohl, 1958). As bufferedformalin was used to preserve the samples, the microzooplanktonmay have been underestimated because of labile protozoandisintegration.
We considered that the timing of recently spawned chaetognathspecimens ready to feed on microzooplankton is closely relatedto the maturity cycle displayed by S. enflata, because the age offirst-feeding of S. enflata occurs shortly after hatching (8–9 d;Doncaster, 1902, after Pearre, 1991).
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Table 2. Terms used to calculate Pearre’s C selectivity index.
Species
A Others Total
Diet ad bd ad þ bd = d
Environment ae be ae þ be = e
Total ad þ ae = a bd þ be = b ad þ ae þ bd þ be = n
Reproduction and feeding of Sagitta enflata 363
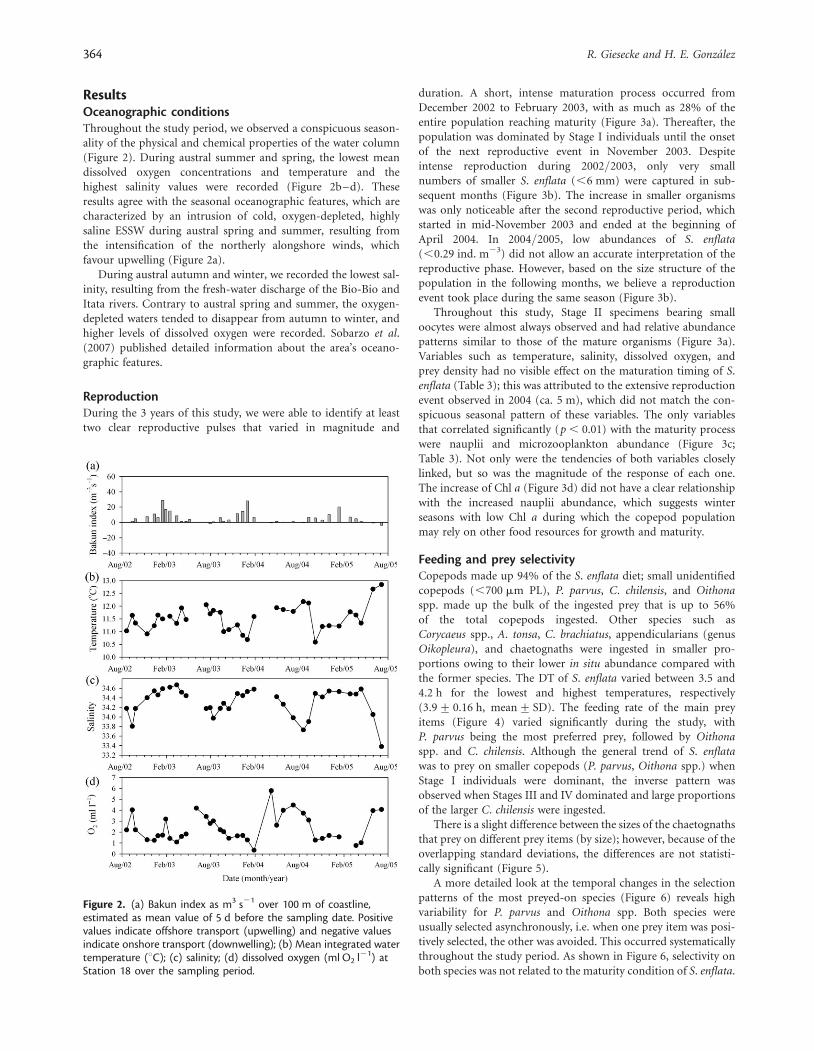
ResultsOceanographic conditionsThroughout the study period, we observed a conspicuous season-ality of the physical and chemical properties of the water column(Figure 2). During austral summer and spring, the lowest meandissolved oxygen concentrations and temperature and thehighest salinity values were recorded (Figure 2b–d). Theseresults agree with the seasonal oceanographic features, which arecharacterized by an intrusion of cold, oxygen-depleted, highlysaline ESSW during austral spring and summer, resulting fromthe intensification of the northerly alongshore winds, whichfavour upwelling (Figure 2a).
During austral autumn and winter, we recorded the lowest sal-inity, resulting from the fresh-water discharge of the Bio-Bio andItata rivers. Contrary to austral spring and summer, the oxygen-depleted waters tended to disappear from autumn to winter, andhigher levels of dissolved oxygen were recorded. Sobarzo et al.(2007) published detailed information about the area’s oceano-graphic features.
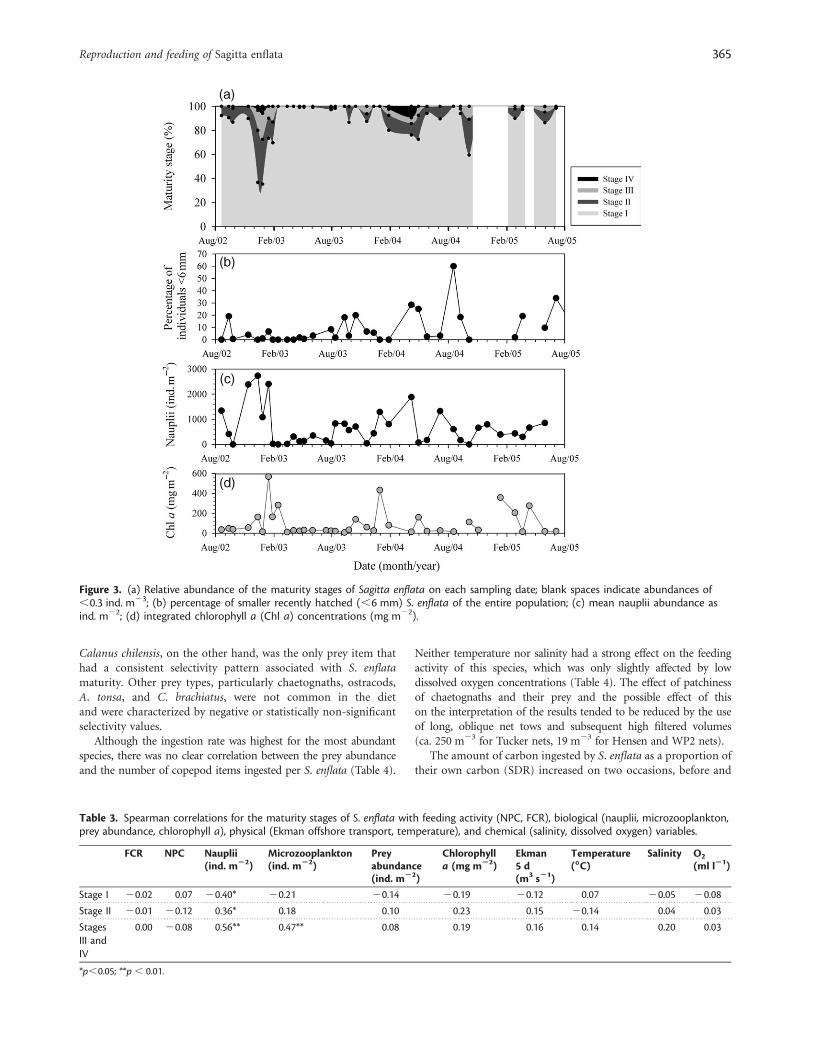
ReproductionDuring the 3 years of this study, we were able to identify at leasttwo clear reproductive pulses that varied in magnitude and
duration. A short, intense maturation process occurred fromDecember 2002 to February 2003, with as much as 28% of theentire population reaching maturity (Figure 3a). Thereafter, thepopulation was dominated by Stage I individuals until the onsetof the next reproductive event in November 2003. Despiteintense reproduction during 2002/2003, only very smallnumbers of smaller S. enflata (,6 mm) were captured in sub-sequent months (Figure 3b). The increase in smaller organismswas only noticeable after the second reproductive period, whichstarted in mid-November 2003 and ended at the beginning ofApril 2004. In 2004/2005, low abundances of S. enflata(,0.29 ind. m23) did not allow an accurate interpretation of thereproductive phase. However, based on the size structure of thepopulation in the following months, we believe a reproductionevent took place during the same season (Figure 3b).
Throughout this study, Stage II specimens bearing smalloocytes were almost always observed and had relative abundancepatterns similar to those of the mature organisms (Figure 3a).Variables such as temperature, salinity, dissolved oxygen, andprey density had no visible effect on the maturation timing of S.enflata (Table 3); this was attributed to the extensive reproductionevent observed in 2004 (ca. 5 m), which did not match the con-spicuous seasonal pattern of these variables. The only variablesthat correlated significantly (p , 0.01) with the maturity processwere nauplii and microzooplankton abundance (Figure 3c;Table 3). Not only were the tendencies of both variables closelylinked, but so was the magnitude of the response of each one.The increase of Chl a (Figure 3d) did not have a clear relationshipwith the increased nauplii abundance, which suggests winterseasons with low Chl a during which the copepod populationmay rely on other food resources for growth and maturity.
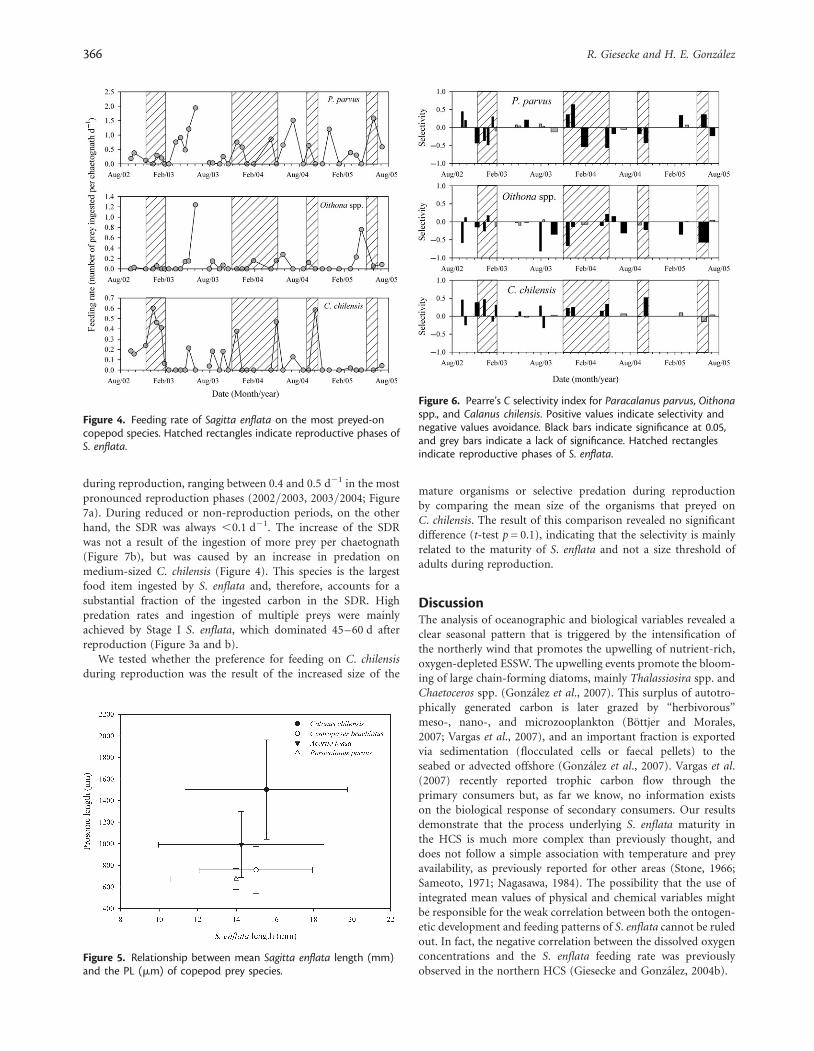
Feeding and prey selectivityCopepods made up 94% of the S. enflata diet; small unidentifiedcopepods (,700 mm PL), P. parvus, C. chilensis, and Oithonaspp. made up the bulk of the ingested prey that is up to 56%of the total copepods ingested. Other species such asCorycaeus spp., A. tonsa, C. brachiatus, appendicularians (genusOikopleura), and chaetognaths were ingested in smaller pro-portions owing to their lower in situ abundance compared withthe former species. The DT of S. enflata varied between 3.5 and4.2 h for the lowest and highest temperatures, respectively(3.9+ 0.16 h, mean+ SD). The feeding rate of the main preyitems (Figure 4) varied significantly during the study, withP. parvus being the most preferred prey, followed by Oithonaspp. and C. chilensis. Although the general trend of S. enflatawas to prey on smaller copepods (P. parvus, Oithona spp.) whenStage I individuals were dominant, the inverse pattern wasobserved when Stages III and IV dominated and large proportionsof the larger C. chilensis were ingested.
There is a slight difference between the sizes of the chaetognathsthat prey on different prey items (by size); however, because of theoverlapping standard deviations, the differences are not statisti-cally significant (Figure 5).
A more detailed look at the temporal changes in the selectionpatterns of the most preyed-on species (Figure 6) reveals highvariability for P. parvus and Oithona spp. Both species wereusually selected asynchronously, i.e. when one prey item was posi-tively selected, the other was avoided. This occurred systematicallythroughout the study period. As shown in Figure 6, selectivity onboth species was not related to the maturity condition of S. enflata.
Figure 2. (a) Bakun index as m3 s21 over 100 m of coastline,estimated as mean value of 5 d before the sampling date. Positivevalues indicate offshore transport (upwelling) and negative valuesindicate onshore transport (downwelling); (b) Mean integrated watertemperature (8C); (c) salinity; (d) dissolved oxygen (ml O2 l21) atStation 18 over the sampling period.
364 R. Giesecke and H. E. Gonzalez
Calanus chilensis, on the other hand, was the only prey item thathad a consistent selectivity pattern associated with S. enflatamaturity. Other prey types, particularly chaetognaths, ostracods,A. tonsa, and C. brachiatus, were not common in the dietand were characterized by negative or statistically non-significantselectivity values.
Although the ingestion rate was highest for the most abundantspecies, there was no clear correlation between the prey abundanceand the number of copepod items ingested per S. enflata (Table 4).
Neither temperature nor salinity had a strong effect on the feedingactivity of this species, which was only slightly affected by lowdissolved oxygen concentrations (Table 4). The effect of patchinessof chaetognaths and their prey and the possible effect of thison the interpretation of the results tended to be reduced by the useof long, oblique net tows and subsequent high filtered volumes(ca. 250 m23 for Tucker nets, 19 m23 for Hensen and WP2 nets).
The amount of carbon ingested by S. enflata as a proportion oftheir own carbon (SDR) increased on two occasions, before and
Figure 3. (a) Relative abundance of the maturity stages of Sagitta enflata on each sampling date; blank spaces indicate abundances of,0.3 ind. m23; (b) percentage of smaller recently hatched (,6 mm) S. enflata of the entire population; (c) mean nauplii abundance asind. m22; (d) integrated chlorophyll a (Chl a) concentrations (mg m22).
. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . .
Table 3. Spearman correlations for the maturity stages of S. enflata with feeding activity (NPC, FCR), biological (nauplii, microzooplankton,prey abundance, chlorophyll a), physical (Ekman offshore transport, temperature), and chemical (salinity, dissolved oxygen) variables.
FCR NPC Nauplii(ind. m22)
Microzooplankton(ind. m22)
Preyabundance(ind. m22)
Chlorophylla (mg m22)
Ekman5 d(m3 s21)
Temperature(88888C)
Salinity O2
(ml l21)
Stage I 20.02 0.07 20.40* 20.21 20.14 20.19 20.12 0.07 20.05 20.08
Stage II 20.01 20.12 0.36* 0.18 0.10 0.23 0.15 20.14 0.04 0.03
StagesIII andIV
0.00 20.08 0.56** 0.47** 0.08 0.19 0.16 0.14 0.20 0.03
*p,0.05; **p , 0.01.
Reproduction and feeding of Sagitta enflata 365
during reproduction, ranging between 0.4 and 0.5 d21 in the mostpronounced reproduction phases (2002/2003, 2003/2004; Figure7a). During reduced or non-reproduction periods, on the otherhand, the SDR was always ,0.1 d21. The increase of the SDRwas not a result of the ingestion of more prey per chaetognath(Figure 7b), but was caused by an increase in predation onmedium-sized C. chilensis (Figure 4). This species is the largestfood item ingested by S. enflata and, therefore, accounts for asubstantial fraction of the ingested carbon in the SDR. Highpredation rates and ingestion of multiple preys were mainlyachieved by Stage I S. enflata, which dominated 45–60 d afterreproduction (Figure 3a and b).
We tested whether the preference for feeding on C. chilensisduring reproduction was the result of the increased size of the
mature organisms or selective predation during reproductionby comparing the mean size of the organisms that preyed onC. chilensis. The result of this comparison revealed no significantdifference (t-test p = 0.1), indicating that the selectivity is mainlyrelated to the maturity of S. enflata and not a size threshold ofadults during reproduction.
DiscussionThe analysis of oceanographic and biological variables revealed aclear seasonal pattern that is triggered by the intensification ofthe northerly wind that promotes the upwelling of nutrient-rich,oxygen-depleted ESSW. The upwelling events promote the bloom-ing of large chain-forming diatoms, mainly Thalassiosira spp. andChaetoceros spp. (Gonzalez et al., 2007). This surplus of autotro-phically generated carbon is later grazed by “herbivorous”meso-, nano-, and microzooplankton (Bottjer and Morales,2007; Vargas et al., 2007), and an important fraction is exportedvia sedimentation (flocculated cells or faecal pellets) to theseabed or advected offshore (Gonzalez et al., 2007). Vargas et al.(2007) recently reported trophic carbon flow through theprimary consumers but, as far we know, no information existson the biological response of secondary consumers. Our resultsdemonstrate that the process underlying S. enflata maturity inthe HCS is much more complex than previously thought, anddoes not follow a simple association with temperature and preyavailability, as previously reported for other areas (Stone, 1966;Sameoto, 1971; Nagasawa, 1984). The possibility that the use ofintegrated mean values of physical and chemical variables mightbe responsible for the weak correlation between both the ontogen-etic development and feeding patterns of S. enflata cannot be ruledout. In fact, the negative correlation between the dissolved oxygenconcentrations and the S. enflata feeding rate was previouslyobserved in the northern HCS (Giesecke and Gonzalez, 2004b).
Figure 4. Feeding rate of Sagitta enflata on the most preyed-oncopepod species. Hatched rectangles indicate reproductive phases ofS. enflata.
Figure 5. Relationship between mean Sagitta enflata length (mm)and the PL (mm) of copepod prey species.
Figure 6. Pearre’s C selectivity index for Paracalanus parvus, Oithonaspp., and Calanus chilensis. Positive values indicate selectivity andnegative values avoidance. Black bars indicate significance at 0.05,and grey bars indicate a lack of significance. Hatched rectanglesindicate reproductive phases of S. enflata.
366 R. Giesecke and H. E. Gonzalez

During the two maturity events that could be investigated ade-quately, a clear correlation was recorded between the number ofmature organisms and copepod nauplii abundance. The relation-ship between these can be explained by an indirect associationbetween both, rather than a direct predator–prey relationship,because nauplii were not found in S. enflata guts. Moreover,nauplii are not expected to contribute substantially to thecarbon requirements of S. enflata. We were not able to determinewhether S. enflata smaller than 5 mm preyed on nauplii, becausethe nets used undersampled small chaetognaths.
Increased nauplii abundance, together with the matureS. enflata population previously reported by McLaren (1969),occurred simultaneously with enhanced predation (as FR) onC. chilensis, regardless of its relative abundance (increased selectiv-ity). This, on some occasions, resulted in an increase in the SDRbefore and during reproduction, similar to the observationsmade by Kehayias et al. (1996). The selective ingestion of largecopepods might account for the surplus energy necessary tocope with the growth and maturation requirements of thisspecies. However, as reported by Reeve (1970), the low capacityof almost all chaetognaths to store lipids (4% of their dryweight) make them “totally dependent” on external lipid avail-ability (Conover and Corner, 1968). Therefore, chaetognathsmight need some lipid surplus to fulfil their own vitellogenesis.Reeve (1970) estimated that 41% of the ingested nitrogen isutilized for gamete production, which is equivalent to the ratioincorporated into its own body tissue during early growth. Thissurplus of energy and nitrogen may be obtained from the ingestionof lipid-bearing organisms that are, in this case, highly availableduring reproduction of their prey (copepods). If we considernauplii abundance to be a proxy for copepod reproductionbecause of the quick egg–nauplii transition (1–2 d at in situ temp-eratures; Tang et al., 1998), then C. chilensis might account com-pletely or partially for the required amount of lipids necessary topromote oogenesis in S. enflata. The latter suggestion is alsosupported by Hidalgo and Escribano (2007), who reported high
reproductive pulses of C. chilensis during austral spring andsummer and intermittent reproduction during winter at thesame station (St. 18). The reproduction of some copepodsduring austral winter was eventually supported by the ingestionof prey other than diatoms, such as micro- and nanoplankton,which may dominate during these seasons (Bottjer and Morales,2007; Gonzalez et al., 2007).
A dependence on external lipids, especially polyunsaturatedfatty acids (PUFA), for maturation and reproduction has beenextensively reported for crustacean zooplankton in marine andfresh-water environments (reviewed by Brett and Muller-Navarra, 1997; Lee et al., 2006). In the HCS, high abundances ofchain-forming diatoms usually promote the egg production ratein calanoid copepods (Peterson et al., 1988), mainly associatedwith the increase of highly saturated fatty acids in their diet(Vargas et al., 2006). The subsequent accumulation of triacylgly-cerols and wax esters in oil sacs is characteristic of the copepodgenera Calanus, Euchaeta, Paraeuchaeta, Eucalanus, Rhincalanus,and Pseudocalanus (Lee et al., 2006). The transfer of lipids fromprimary consumers to higher trophic levels has received less atten-tion, except for a few examples of natural fish and fish larvae(reviewed in Dalsgaard et al., 2003; Varpe et al., 2005). By usingfatty acids as trophic markers, St John and Lund (1996) demon-strated that the diatom-based foodweb transfer significantlyimproves the condition of cod larvae in natural environments,owing to the transport of PUFA through the foodweb.
The only work that deals with the temporal developmentof total lipid concentrations in chaetognaths (Reeve et al., 1970)was unable to explain the seasonal oscillations by means of temp-erature, salinity, or food concentrations. Alvarino (1965) observedmajor discrepancies in reproduction frequency within species insimilar environments. Perhaps the limiting of external lipids tofulfil maturity might be one of the main processes controllingthe reproductive pulses in chaetognaths and might explain suchdiscrepancies. However, further work is needed to determine theexact mechanisms coupling prey and predator reproduction.
On the other hand, the environment in terms of food quantityand quality, in which the S. enflata offspring develop after largereproductive pulses, might be highly favourable. As shown inFigure 3, the time required for S. enflata to reach maturity(from Stage II to III or IV) took a maximum of 1.5 months(from 0% on 12 November 2002 to 27% Stage III–IV on 27December 2002), whereas the hatching time after reproductionranged from 19 to 48 h depending on the temperature (reviewedin Pearre, 1991). Small S. enflata juveniles begin active feeding8–9 d after hatching (Doncaster, 1902 after Pearre, 1991), whenlarge numbers of copepod nauplii are still available in the environ-ment for the small chaetognaths (,6 mm) to feed on. Potentialprey ingested by small S. enflata (,6 mm) should not exceedhead width. Therefore, we expect the prey ingested by newlyhatched S. enflata to range between a maximum width of 227and 445 mm for specimens 3 and 6 mm long, respectively, taking
. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .
Table 4. Spearman’s correlations for Sagitta enflata feeding activity with biological (prey abundance, chlorophyll a), physical (Ekmanoffshore transport, temperature), and chemical (salinity, dissolved oxygen) variables.
Prey abundance (ind. m22) Chlorophyll a (mg m22) Ekman 5 d (m3 s21) Temperature (88888C) Salinity O2 (ml l21)
FCR 20.07 20.07 20.13 0.08 20.15 0.24
NPC 20.11 20.32 20.31 0.26 20.17 0.34*
*p , 0.05.
Figure 7. (a) Specific daily ratio (SDR) as a proportion of carboningested daily by the population of Sagitta enflata in relation to itsown body carbon; (b) Comparison between the food containingratio (FCR) and number of prey ingested (NPC) for S. enflata.Hatched rectangles indicate reproductive phases of S. enflata.
Reproduction and feeding of Sagitta enflata 367

into account a head-width–body-length ratio of 0.0758 for S.enflata (Pearre, 1980). These sizes agree with the sizes of thesmall calanoid copepod nauplii (116+ 40 mm) collected duringthis study (Aparicio, 2006), and C. chilensis NI and NII stages(220–330 mm; R. Escribano, pers. comm.).
The presence of a large number of nauplii would, in turn,favour growth of newly hatched S. enflata and so diminish theirsize-dependent vulnerability to predation. However, as shown inFigure 3, the successful establishment of smaller chaetognathsdid not seem to be related to the reproduction intensity but tothe temporal extension of the reproductive event that coincideswith the occurrence of large numbers of nauplii. After the strong2002/2003 spawning event, the abundance of recently hatched S.enflata was low in subsequent months, possibly associated withthe recruitment failure that was likely caused by the poor avail-ability of food for the small juveniles. The near absence ofsmaller S. enflata after the 2002/2003 reproductive pulse seemsnot to be a consequence of the limited sampling procedure(monthly). This was determined because, not only were smallsizes (,6 mm) scarce, but large sizes (7–9 mm) were also scarcein the following 2 months (R. Giesecke, unpublished). Duringthe two later reproductive events (2003/2004, 2004/2005), inwhich an extended breeding period was achieved and a continuousabundance of nauplii available, a significant proportion of theentire population was made up of newly hatched chaetognaths.We cannot, however, dismiss the possibility that the low abun-dance of small chaetognaths in the following months after thefirst peak in reproduction might be a consequence of advectionprocesses of small organisms transported offshore during theprevailing upwelling conditions in austral spring and summer.
Based on the results of this study, we propose a conceptualmodel (Figure 8) in which the onset of favourable food conditionsfor oogenesis and reproduction in copepods triggers a chain reac-tion that starts with the selective predation of S. enflata on largelipid-bearing copepods (such as C. chilensis) while favouringlipid storage in S. enflata Stages I and II for reaching maturity(Stages III and IV). Once mature organisms have reproducedand the hatched offspring start to feed, the considerable number
of copepod nauplii remaining in the environment as a result ofthe copepod reproduction that triggered chaetognath maturitycreate favourable food conditions for newly hatched chaetognaths.
AcknowledgementsWe thank FONDAP-COPAS Research Programme 3, which kindlyprovided the copepod abundance data, and Ruben Escribano,Pamela Hidalgo, and Alvaro Araneda who collected the samplesincluded in this study. We thank the COPAS time-series teamand the crew of the RV “Kay Kay”. Two anonymous reviewersgreatly improved an earlier version of the manuscript. The studywas supported by a CONICYT Doctoral fellowship grant andthe FONDAP-COPAS Centre funded by FONDAP-Chile grant15010007.
ReferencesAlvarino, A. 1965. Chaetognaths. Oceanography and Marine Biology.
An Annual Review, 3: 115–194.
Alvarino, A. 1967. The chaetognaths of the NAGA expedition (1959–1961) in the South China Sea and the Gulf of Thailand. Part 1.Systematics. NAGA Report, 4: 1–197.
Aparicio, C. 2006. Variabilidad espacial y temporal de la abundancia ycomposicion del microzooplankton y detritus en una estacioncostera fija en la plataforma continental de Concepcion (368S),durante el perıodo 2002–2005. MSc thesis, Universidad Australde Chile. 73 pp.
Baier, C. T., and Purcell, J. E. 1997. Effects of sampling and preser-vation on apparent feeding by chaetognaths. Marine EcologyProgress Series, 146: 37–42.
Bajkov, A. D. 1935. How to estimate the daily food consumption offish under natural conditions. Transactions of the AmericanFisheries Society, 65: 288–289.
Bakun, A. 1978. Daily and weekly upwelling indices, west coast ofNorth America, 1967–73. NOAA Technical Reports, NMFSSSRS-693. 103 pp.
Batistic, M. 2003. Abundance, biomass, C and N content ofFlaccisagitta enflata and Mesosagita minima (Chaetognatha).Marine Ecology Publicazioni della Stazione Zoologica di Napoli,24: 1–13.
Berggreen, U., Hansen, B., and Kiørboe, T. 1988. Food size spectra,ingestion and growth of the copepod Acartia tonsa during develop-ment: implications for determination of copepod production.Marine Biology, 99: 341–352.
Bottjer, D., and Morales, C. E. 2007. Nanoplanktonic assemblages inthe upwelling area off Concepcion (368S), central Chile: abun-dance, biomass, and grazing potential during the annual cycle.Progress in Oceanography, 75: 415–434.
Brett, M. T., and Muller-Navarra, D. C. 1997. The role of highly unsa-turated fatty acids in aquatic foodweb processes. FreshwaterBiology, 38: 483–499.
Conover, R. J., and Corner, E. D. S. 1968. Respiration and nitrogenexcretion by some marine zooplankton in relation to their lifecycles. Journal of the Marine Biological Association of the UK,48: 49–57.
Conway, D. V. P., and Williams, R. 1986. Seasonal population struc-ture, vertical distribution and migration of the chaetognathSagitta elegans in the Celtic Sea. Marine Biology, 93: 377–387.
Dalsgaard, J., St John, M., Kattner, G., Muller-Navarra, D., and Hagen,W. 2003. Fatty acid trophic markers in the pelagic marine environ-ment. Advances in Marine Biology, 46: 225–340.
Daneri, G., Dellarossa, V., Quinones, R., Jacob, B., Montero, P., andUlloa, O. 2000. Primary production and community respirationin the Humboldt Current system off Chile and associated oceanicareas. Marine Ecology Progress Series, 197: 41–49.
Figure 8. Conceptual scheme of the associations between copepodmaturity and the feeding activity and oogenesis of Sagitta enflata andthe possible association between their recently hatched offspring.Favourable food conditions for copepod reproduction are linkedwith selective predation of S. enflata upon large oil-sac-bearingcopepods (Calanus chilensis). This process may allow theincorporation of lipids by S. enflata, which are later used for theirown oogenesis. The offspring produced during these reproductivepulses are released into an environment with large numbers ofpotential prey, which would favour their development.
368 R. Giesecke and H. E. Gonzalez

Doncaster, L. 1902. On the development of Sagitta: with notes on theanatomy of the adult. Quarterly Journal of Microscopical Science,46: 351–398.
Escribano, R., Irribarren, C., and Rodrıguez, L. 1997. Influence of foodquantity and temperature on development and growth of marinecopepod Calanus chilensis from northern Chile. Marine Biology,128: 281–288.
Feigenbaum, D. L., and Maris, R. C. 1984. Feeding in theChaetognatha. Oceanography and Marine Biology. An AnnualReview, 22: 343–392.
Ghirardelli, E. 1968. Some aspects of the biology of the chaetognaths.Advances in Marine Biology, 6: 271–375.
Giesecke, R., and Gonzalez, H. E. 2004a. Mandible characteristics andallometric relations in copepods: a reliable method to estimate preysize and composition from mandible occurrence in predator guts.Revista Chilena de Historia Natural, 77: 607–616.
Giesecke, R., and Gonzalez, H. E. 2004b. Feeding of Sagitta enflata andvertical distribution of chaetognaths in relation to low oxygen con-centrations. Journal of Plankton Research, 26: 475–486.
Gonzalez, H. E., Menschel, E., Aparicio, C., and Barria, C. 2007.Spatial and temporal variability of microplankton and detritus,and their export to the shelf sediments in the upwelling area offConcepcion, Chile (368S), during 2002–2005. Progress inOceanography, 75: 435–451.
Hidalgo, P., and Escribano, R. 2007. Coupling of life cycles of thecopepods Calanus chilensis and Centropages brachiatus to upwellinginduced variability in the central-southern region of Chile.Progress in Oceanography, 75: 501–517.
Kehayias, G., Lykakis, J., and Fragopoulu, N. 1996. The diets of thechaetognaths Sagitta enflata, S. serratodentata atlantica, and S.bipunctata at different seasons in eastern Mediterranean coastalwaters. ICES Journal of Marine Science, 53: 837–846.
Khan, M. A., and Williamson, D. I. 1970. Seasonal changes in thedistribution of Chaetognatha and other plankton in the IrishSea. Journal of Experimental Marine Biology and Ecology, 5:285–303.
King, K. R. 1979. The life history and vertical distribution of thechaetognath, Sagitta elegans, in Dabob Bay, Washington. Journalof Plankton Research, 1: 153–167.
King, K. R., Hollibaugh, J. T., and Azam, F. 1980. Predator–preyinteractions between the larvacean Oikopleura dioica andbacterioplankton in enclosed water columns. Marine Biology, 56:49–57.
Lee, R. F., Hagen, W., and Kattner, G. 2006. Lipid storage in marinezooplankton. Marine Ecology Progress Series, 307: 273–306.
McLaren, I. A. 1966. Adaptive significance of large size and long life ofthe chaetognath Sagitta elegans in the Arctic. Ecology, 47: 852–855.
McLaren, I. A. 1969. Population and production ecology of zooplank-ton in Ogac Lake, a landlocked fiord on Baffin Island. Journal ofthe Fisheries Research Board Canada, 26: 1485–1559.
Nagasawa, S. 1984. Laboratory feeding and egg production in thechaetognath Sagitta crassa Tokioka. Journal of ExperimentalMarine Biology and Ecology, 76: 51–65.
Ohman, M. D. 1986. Predator-limited population growth of thecopepod Pseudocalanus sp. Journal of Plankton Research, 8:673–713.
Øresland, V. 1987. Feeding of the chaetognaths Sagitta elegans andSagitta setosa at different seasons in Gullmarsfjorden, Sweden.Marine Ecology Progress Series, 39: 69–79.
Owre, H. B. 1960. Plankton of the Florida Current. Part VI. TheChaetognatha. Bulletin of Marine Science, 10: 255–322.
Parada, C. E., Sobrazo, M., Figueroa, D., and Castro, L. 2001.Circulacion del Golfo de Arauco en un perıodo de transicionestacional: un nuevo enfoque. Investigaciones Marinas, Valparaıso,29: 11–23.
Pearre, S. 1980. Feeding by Chaetognatha: the relation of prey size topredator size in several species. Marine Ecology Progress Series,3: 125–134.
Pearre, S. 1982. Estimating prey preference by predators: uses ofvarious indices and a proposal of another based on x2. CanadianJournal of Fisheries and Aquatic Sciences, 39: 914–923.
Pearre, S. 1991. Growth and reproduction. In The Biology ofChaetognaths, pp. 61–75. Ed. by Q. Bone, H. Kapp, and A. C.Pierrot-Bults. Oxford University Press, New York. 173 pp.
Peterson, W., Arcos, D., McManus, G., Dam, H., Bellantoni, D.,Johnson, T., and Tiselius, P. 1988. The nearshore zone duringcoastal upwelling: daily variability and coupling between primaryand secondary production off Central Chile. Progress inOceanography, 20: 1–40.
Reeve, M. R. 1970. The biology of Chaetognatha. I. Quantitativeaspects of growth and egg production in Sagitta hispida. InMarine Food Chains, pp. 169–189. Ed. by J. H. Steele. Oliverand Boyd, Edinburgh. 552 pp.
Reeve, M. R., and Cosper, T. C. 1975. Chaetognatha. In Reproductionof Marine Invertebrates, pp. 157–184. Ed. by A. C. Giese, and J. S.Pearse. Academic Press, New York. 343 pp.
Reeve, M. R., Raymont, J. E. G., and Raymont, J. K. B. 1970. Seasonalbiochemical composition and energy sources of Sagitta hispida.Marine Biology, 6: 357–364.
Sabatini, M., and Kiørboe, T. 1994. Egg production, growth and devel-opment of the cyclopoid copepod Oithona similis. Journal ofPlankton Research, 16: 1329–1351.
Saito, H., and Kiørboe, T. 2001. Feeding rates in the chaetognath S.elegans: effects of prey size, prey swimming behaviour andsmall-scale turbulence. Journal of Plankton Research, 23:1385–1398.
Sameoto, D. D. 1971. Life history, ecosystem production and anempirical mathematical model of the population of Sagittaelegans in St Margaret’s Bay, Nova Scotia. Journal of the FisheriesResearch Board of Canada, 28: 971–985.
Sameoto, D. D. 1987. Vertical distribution and ecological significanceof chaetognaths in the Arctic environment of Baffin Bay. PolarBiology, 7: 317–328.
Satapoomin, S. 1999. Carbon content of some common tropicalAndaman Sea copepods. Journal of Plankton Research, 21:2117–2123.
Sobarzo, M., Bravo, L., Donoso, D., Garces-Vargas, J., and Schneider,W. 2007. Coastal upwelling and seasonal cycles that influencethe water column over the continental shelf off central Chile.Progress in Oceanography, 75: 363–382.
St John, M. A., and Lund, T. 1996. Lipid biomarkers: linking theutilization of frontal plankton biomass to enhanced condition ofjuvenile North Sea cod. Marine Ecology Progress Series, 131:75–85.
Stone, J. H. 1966. The distribution and fecundity of Sagitta enflataGrassi in the Agulhas current. Journal of Animal Ecology, 35:533–541.
Tang, K. W., Dam, H. G., and Feinberg, L. R. 1998. The relativeimportance of egg production rate, hatching success, hatchingduration and egg sinking in population recruitment of twospecies of marine copepods. Journal of Plankton Research, 20:1971–1987.
Utermohl, H. 1958. Vervollkommnung der quantitativenPhytoplankton-Methodik. Mitteilungen Internationale VereinigungTheorie Angewandte Limnologie, 9: 1–38.
Uye, S. 1982. Length-weight relationships of important zooplanktonfrom the inland Sea of Japan. Journal of the OceanographySociety of Japan, 38: 149–158.
Uye, S. 1991. Temperature-dependent development and growth of theplanktonic copepod Paracalanus sp. in the laboratory. Bulletin ofthe Plankton Society of Japan, Special Volume: 627–636.
Reproduction and feeding of Sagitta enflata 369

Vargas, C. A., Escribano, R., and Poulet, S. 2006. Phytoplankton diver-sity determines time-windows for successful zooplankton repro-ductive pulses. Ecology, 87: 2992–2999.
Vargas, C. A., Martınez, R., Cuevas, L. A., Pavez, M. A., Cartes, C.,Gonzalez, H. E., Escribano, R., et al. 2007. The relative importanceof microbial and classical foodwebs in a highly productivecoastal upwelling area. Limnology and Oceanography, 52:1495–1510.
Varpe, Ø., Fiksen, Ø., and Slottte, A. 2005. Meta-ecosystem and bio-logical energy transport from ocean to coast: the ecological import-ance of herring migration. Oecologia, 146: 443–451.
Wu, J. 1982. Wind-stress coefficients over sea surface from breeze tohurricane. Journal of Geophysical Research, 82: 9704–9706.
Zo, Z. 1973. Breeding and growth of the chaetognath Sagitta elegans inBedford Basin. Limnology and Oceanography, 18: 750–756.
doi:10.1093/icesjms/fsn030
370 R. Giesecke and H. E. Gonzalez




















