
Unequal Realities: Forced Evictions in Five Indian Cities (2016)
Upload
khangminh22Category
view
1download
0
Vaccine 20 (2002) 3068–3087
Review
Realities and enigmas of human viral influenza: pathogenesis,epidemiology and control
Maurice R. Hilleman∗Merck Institute for Vaccinology, 770 Sumneytown Pike, West Point, PA 19486, USA
Received 29 April 2002; accepted 8 May 2002
Abstract
Influenza A is a viral disease of global dimension, presenting with high morbidity and mortality in annual epidemics, and in pandemicswhich are of infrequent occurrence but which have very high attack rates. Influenza probes reveal a continuing battle for survival betweenhost and parasite in which the host population updates the specificity of its pool of humoral immunity by contact with and response toinfection with the most recent viruses which possess altered antigenic specificity in their hemagglutinin (HA) ligand. HA ligand binds thevirus to the cell to bring about infection. Viral survival relies on escape from host immunity through antigenic alterations in nature whicharise through genetic drift by point mutation principally of the HA gene, or through genetic shift by reassortment exchange of the HAligand with that of viruses retained in avian species. Partial control of influenza is by use of killed whole, subunit, or possible live virusvaccines, all of which rely on worldwide surveillance to provide early detection of the altered immunologic specificity of the next virus tocome. Future global surveillance may be aided by studies of sampled viral isolates in laboratories having capabilities for accelerated geneticsequencing and for automated rapid throughput analyses as well. Influenza vaccines of the future must be directed toward use of conservedgroup-specific viral antigens, such as are present in transitional proteins which are exposed during the fusion of virus to the host cell.Chemotherapy, though still primordial, must eventually provide the ultimate solution to vaccine failures. Probing the enigma of the severeinfluenza pandemic of 1918–1919 is an exciting contemporary venture in which genetic reconstruction of the viral genome from survivingarchival RNA is being conducted with great success. Present evidence reveals successive recycling in pandemics, of only 3 of the 15 possibleavian viral HAs. Pandemics are believed, conventionally, to be derived solely by rare events in which wild viruses of man acquire a newHA ligand of avian origin. There might be an alternative possibility involving a periodicity in selective control by the host population itself,in its receptivity or rejection at a particular time of particular reassortant viruses which might be created more frequently in nature than weare presently aware. This hypothesis, though remote, provides a different way to view and to probe the enigma of pandemic influenza.© 2002 Elsevier Science Ltd. All rights reserved.
Keywords:Influenza hypothesis; Influenza epidemiology; Influenza
1. Introduction and background
Human viral influenza[1–3] is an acute respiratory dis-ease which is rooted in the distant past and is one of themost important infectious diseases of mankind. The diseaseis highly contagious, with person to person spread by aerosoldroplets which mainly infect the epithelial cells of the res-piratory tract. The means to understand influenza and itsepidemiology was provided in the late 1920s by the break-through discovery of transmissibility of swine influenza inpigs by a filterable agent[4], and was followed by the dis-coveries of human influenza type A in 1933[5], of B in1940[6], and of C in 1950[7]. All human influenza viruses[1,8,9] belong to the familyOrthomyxoviridae. There are
∗ Tel.: +1-215-652-8913; fax:+1-215-652-2154.E-mail address:[email protected] (M.R. Hilleman).
three genera, corresponding to influenza types A, B, andC. Influenza A viruses, uniquely, are divided into subtypesbased on major antigenic specificities of their hemagglutinin(HA) and neuraminidase (NA) proteins.
Type A is the most important of the influenza viruses,causing alternate annual outbreaks and epidemics during thewinter seasons of the northern and the southern hemispheres[10]. Virus types B and C are of lesser importance thanA, and will not be discussed here. Pandemics of influenzaA have occurred about three times per century since 1700[11] and were manifested by worldwide spread of the dis-ease, typically with high morbidity and mortality. The mostextreme pandemic in the 20th century was that of Spanishinfluenza of 1918–1919, which killed an estimated 20–40million persons throughout the world[11,12–14].
Influenza A, which is discussed in this review, is a dis-ease of immense complexity in which the occurrence and
0264-410X/02/$ – see front matter © 2002 Elsevier Science Ltd. All rights reserved.PII: S0264-410X(02)00254-2

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3069
the outcome of the illness depends on interactions betweena virus of diverse and changing genetic and antigenic com-position, and the multifaceted immune system of the hostwhich may not be able to respond adequately within a re-stricted time frame. The principal determining factor forwhether influenza outbreaks occur is that of degree of matchin specificities (complementarity) between the surface anti-gens (HA and NA) of the virus and the antibodies againstthem which are current in the population.
The record of the quest, by science, to conquer influenzais one of trial, triumph, tribulation, and even fiasco, in itsattempts to define the disease in all its complexities, andto develop and apply effective vaccines and drugs for itsprevention and control. Understanding is best achieved inreview of the virus and the disease in both its historic andcontemporary perspectives.
2. Clinical influenza
The importance of influenza A virus infection is centeredon its high morbidity and mortality during epidemics andpandemics. The disease[15,16], itself, is of sudden onsetfollowing a short incubation period. Local and systemicreactions may be prominent, and may include high fever,chills, headache, myalgia, dry cough and diarrhea. Infectionof the respiratory tract may give rise to primary pneumoni-tis and this may lead to streptococcal (pneumococcus andbeta hemolytic group A), staphylococcal andHaemophilusinfluenzaeinfections with severe pulmonary complications
Fig. 1. Simplified cartoon showing structure of influenza virion.
and death from pneumonia. Cardiac, muscle (myositis),central nervous system (Reye’s syndrome, Guillain-Barrésyndrome, encephalitis), and renal complications may alsooccur, though infrequently. Notable neurasthenia and fatigueof variable duration, accompanied by transient immunode-ficiency, may be present following recovery from the acutedisease[15–17].
3. Influenza virus
3.1. Structure and functions[2,3,18]
The pleomorphic 80–120 nm virus particle of influenzavirus is illustrated in the cartoon inFig. 1, and the knownand purported functions of the virus-encoded proteins arelisted inTable 1. The envelope of the virus consists of a lipidbilayer from which the spike-form HA and the NA glyco-proteins project. The HA binds the virus to the host cell andbrings about fusion with it. The NA prevents viral aggrega-tion and assists in release of newly minted virus from theinfected cell. These glycoproteins are the principal determi-nants for cell entry in infection (HA) and for exit from thecell after virus replication. Additionally, they are the prin-cipal antigens against which humoral (antibody) immuneresponses of the host are directed. Both the extent of anti-genic change and of protection afforded by NA antibody areminor compared with those of HA. Matrix protein M1 liesjust within the viral envelope where it interacts with the cellgenome and the nuclear export factor, and assists in viral
3070 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
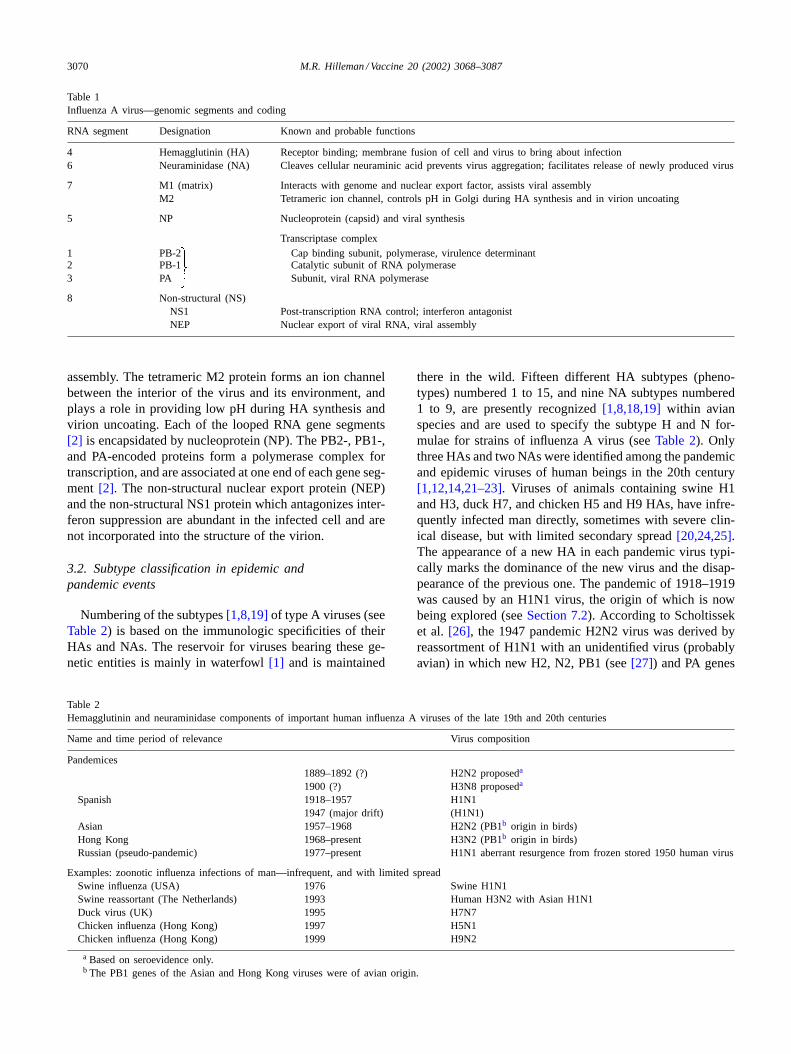
Table 1Influenza A virus—genomic segments and coding
RNA segment Designation Known and probable functions
4 Hemagglutinin (HA) Receptor binding; membrane fusion of cell and virus to bring about infection6 Neuraminidase (NA) Cleaves cellular neuraminic acid prevents virus aggregation; facilitates release of newly produced virus
7 M1 (matrix) Interacts with genome and nuclear export factor, assists viral assemblyM2 Tetrameric ion channel, controls pH in Golgi during HA synthesis and in virion uncoating
5 NP Nucleoprotein (capsid) and viral synthesis
Transcriptase complex1 PB-2 Cap binding subunit, polymerase, virulence determinant2 PB-1 Catalytic subunit of RNA polymerase3 PA Subunit, viral RNA polymerase
8 Non-structural (NS)NS1 Post-transcription RNA control; interferon antagonistNEP Nuclear export of viral RNA, viral assembly
assembly. The tetrameric M2 protein forms an ion channelbetween the interior of the virus and its environment, andplays a role in providing low pH during HA synthesis andvirion uncoating. Each of the looped RNA gene segments[2] is encapsidated by nucleoprotein (NP). The PB2-, PB1-,and PA-encoded proteins form a polymerase complex fortranscription, and are associated at one end of each gene seg-ment [2]. The non-structural nuclear export protein (NEP)and the non-structural NS1 protein which antagonizes inter-feron suppression are abundant in the infected cell and arenot incorporated into the structure of the virion.
3.2. Subtype classification in epidemic andpandemic events
Numbering of the subtypes[1,8,19]of type A viruses (seeTable 2) is based on the immunologic specificities of theirHAs and NAs. The reservoir for viruses bearing these ge-netic entities is mainly in waterfowl[1] and is maintained
Table 2Hemagglutinin and neuraminidase components of important human influenza A viruses of the late 19th and 20th centuries
Name and time period of relevance Virus composition
Pandemices1889–1892 (?) H2N2 proposeda
1900 (?) H3N8 proposeda
Spanish 1918–1957 H1N11947 (major drift) (H1N1)
Asian 1957–1968 H2N2 (PB1b origin in birds)Hong Kong 1968–present H3N2 (PB1b origin in birds)Russian (pseudo-pandemic) 1977–present H1N1 aberrant resurgence from frozen stored 1950 human virus
Examples: zoonotic influenza infections of man—infrequent, and with limited spreadSwine influenza (USA) 1976 Swine H1N1Swine reassortant (The Netherlands) 1993 Human H3N2 with Asian H1N1Duck virus (UK) 1995 H7N7Chicken influenza (Hong Kong) 1997 H5N1Chicken influenza (Hong Kong) 1999 H9N2
a Based on seroevidence only.b The PB1 genes of the Asian and Hong Kong viruses were of avian origin.
there in the wild. Fifteen different HA subtypes (pheno-types) numbered 1 to 15, and nine NA subtypes numbered1 to 9, are presently recognized[1,8,18,19]within avianspecies and are used to specify the subtype H and N for-mulae for strains of influenza A virus (seeTable 2). Onlythree HAs and two NAs were identified among the pandemicand epidemic viruses of human beings in the 20th century[1,12,14,21–23]. Viruses of animals containing swine H1and H3, duck H7, and chicken H5 and H9 HAs, have infre-quently infected man directly, sometimes with severe clin-ical disease, but with limited secondary spread[20,24,25].The appearance of a new HA in each pandemic virus typi-cally marks the dominance of the new virus and the disap-pearance of the previous one. The pandemic of 1918–1919was caused by an H1N1 virus, the origin of which is nowbeing explored (seeSection 7.2). According to Scholtisseket al. [26], the 1947 pandemic H2N2 virus was derived byreassortment of H1N1 with an unidentified virus (probablyavian) in which new H2, N2, PB1 (see[27]) and PA genes

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3071
Fig. 2. Cartoon illustrating mechanism for reassortment transfer of a single wild virus HA gene segment to an egg-adapted vaccine production strain ofinfluenza virus.
were acquired. The 1968 pandemic H3N2 virus was derivedthrough reassortment exchange of the H3 component for theH2 of the parent H2N2 virus. The reappearance and world-wide spread of virus subtype H1N1, which was the cause forthe “Russian pandemic” virus of 1977, 20 years followingits disappearance after 1957, is considered to be both aber-rant and remarkable[28,29]. Genetic analyses showed the1977 Russian isolate[28,29] to be of near genetic identitywith the H1N1 viruses that were circulating in 1950. Thisindicates that the virus had been preserved at some unde-fined location, probably in frozen storage[28]. The “Rus-sian pandemic”, as might be expected, was benign and thedisease was restricted mostly to persons who were born af-ter the late 1950s. This H1N1 subtype, once extinguishedbut reintroduced in 1977, now co-circulates with the 1968H3N2 virus, at the expense of the world population. It isnow proposed, based on seroepidemiologic data[22,23,30]that the pandemics of 1889 and 1900 were caused by H2N2and H3N8 viruses, respectively.
3.3. Influenza virus alteration by accumulation of pointmutation, by recombination, and by reassortment
The multigenic influenza A virus undergoes antigenicchanges through an accumulation of point mutations duringpropagation in the human host species[3]. These changesprovide a means for immunologic escape and for propaga-tive success in partially and variably immune populationsin which there are continuous selection pressures resultingfrom constraints imposed by developing herd immunity. Ge-netic recombination between different viral strains may alsobe a source for antigenic alteration, though this is not a com-mon event[1,19,31]. The most abrupt changes in antigenicspecificity occur through HA and NA gene reassortment,first noted by Hirst and Gotlieb[32], in the process of dualinfection with different influenza viruses in single cells inculture. This process, followed by immune selection in thehuman host in nature or on contrived reassortment in the lab-oratory [33,34], opened the door to understanding geneticexchanges between influenza viruses. Genetic reassortment
to create new human pandemic viruses, such as those of1957 and 1968, involved acquisition of new HA or HA andNA genes from a zoonotic virus by a virulent wild humanvirus [35].
The process by which reassortment occurs in nature[32–34] is illustrated inFig. 2 as an example which showspurposeful exchange of gene segments in the laboratorythrough dual infection of a wild influenza virus with anegg-adapted virus in embryonated hen’s eggs. The purposeis to obtain seed viruses which give high virus yields inembryonated hen’s eggs for vaccine production. The mixedpropagation of the two viruses in egg culture results in dualinfection in many cells and yields a diversity in the geneticcomposition of the reassortant progeny that appear. Im-munologic and other means of viral selection accomplishesrecovery of the wanted egg-adapted reassortant virus whichbears the new HA and NA genes, but retains the remaininggenes from the parent virus. Reassortment among differenthuman influenza viruses, in the venue of nature, rests on thesame basic principles as those used to prepare seed virusfor vaccine, and discussed earlier.
4. Viral infection and pathogenetic mechanisms
4.1. Infective process
Wild influenza viruses, following recovery in mammals,in embryonated hen’s eggs, or in cell cultures can be adaptedto growth in a number of different kinds of mammalian andavian cells in vitro[3,36–38]. Many different species ofenveloped RNA viruses share a common need to achieveboth attachment of virus to host cell membranes and fusionof virus and host cell membranes to provide for entry ofthe viral genome into the cell where replication occurs. Ex-amples (see[39]) include the Orthomyxoviruses that causeinfluenza; the Paramyxoviruses to which measles virus, res-piratory syncytial virus, simian virus 5, and parainfluenzavirus belong; the Filoviruses which include the Ebola andMarburg agents; and the Retroviruses of which human

3072 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
leukemia and the human immunodeficiency agent (HIV)are members. The basic mechanism of cell fusion amongthe mentioned agents proceeds by a common theme whichis being studied intensively at this time.
4.2. Virus–cell fusion
The mechanism for fusion of virus and cell has been pur-sued vigorously for more than a decade by several investiga-tors[39–43]. The findings by Kim and coworkers[39,44–46]may be the most advanced, because of their extensive en-gagement of crystallographic analysis and their modeling ofproteins which may interfere with fusion. The Kim findingshave focused on influenza virus, respiratory syncytial virus,and HIV agents, and are appropriate for review in relationto influenza.
In influenza A virus infection, the HA glycoprotein playsthe pivotal role in viral infection, and the NA aids in free-ing newly-formed virus from the cell membrane where it isassembled and buds with the formation of particles.
The native precursor HA0 monomers of mammalianviruses are cleaved extracellularly by host proteases atthe position of a conserved arginine residue giving sub-units, HA1 and HA2. The infective process carried outby the HA0 ligand starts with binding of the HA to sialicacid-containing cell receptors. This initiates endocytosisand formation of the endosome in which the HA2 proteinof the virus fuses with both the host and viral membranes.In the process, precursor HA HA0 monomers associatenon-covalently to form homotrimers[42]. Cleavage of theHA0 protein is brought about by cellular proteases and newN termini for the HA2 fusion peptide (F) are exposed. LowpH within the endosome triggers conformational changes
Fig. 3. Cartoon illustrating sequence of fusion of influenza virus to host cell with resultant infection.
in HA2, which bring about fusion of the virus and cellmembranes.
Fig. 3 is adapted from the Kim and coworkers’ model[39,44–46]and is intended only to illustrate an overall likelyprocess for cell fusion by influenza virus. Following bind-ing to the cell membrane, the virus is endocytosed and iscontained within an endosome. Low pH in the endosomeinitiates the fusion process. The C-terminal end (F1) ofHA2 fusion domain remains attached to the virus while theN-terminal segment comprises the fusion peptide. Betweenthe N and C elements of F1, are segments that resemble acoiled coil structure[45,47], containing what are called theHRN and HRC regions. The coiled coils of the two regionsassemble in anti-parallel mode and form pairs of pre-hairpinstructures. Once HRC associates with HRN, the three dimerssnap into hairpin-like structures which pull the lipid cell andviral membranes together and fuse them. This allows entryof the viral gene segments into the cell where viral replica-tion takes place.
4.2.1. Fusion blockadeConformational changes in peptides which are involved
and exposed during cell fusion appear to be highly conservedamong certain related groups of viruses such as HIV, thatare otherwise diverse. This finding has stimulated much lab-oratory research to develop group-specific vaccines[48–51]and small protein viral inhibitors[52,53]. Of special currentinterest is the discovery by Fouts et al.[49] of a commonantigenic determinant among HIV strains which resides ina single chain chimeric gp120–CD4 protein. This is a primecandidate for a group-specific vaccine against HIV. The re-markable findings in studies of the fusion process in HIVhave stimulated the search for inhibitors of fusion. Inhibitors

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3073
may prevent CD4 attachment to HIV, may inhibit co-receptorinteractions, or may target the viral gp41 itself[47]. Progresshas been made and some candidate substances[54–56]havebeen placed in clinical trials. Kim and coworkers[52] syn-thesized a small antiviral protein called 5-helix which in-hibits fusion of diverse HIV variants.
4.3. Fusion and virulence
The occurrence of epidemics and pandemics ordinarilyrepresents increase in numbers of cases of influenza withoutreference to virulence. Epidemic disease may be mild tosevere, depending on the extent of viral proliferation in thehost and other attributes which are resident within the virusitself. In some instances, pandemic virus may be no morevirulent than ordinary virulent strains of epidemic virus.
Steinhauer[42] conducted studies of highly virulent andapathogenic fowl influenza viruses to determine the basis forvirulence which might provide clues for human influenza.He found that the HA cleavage site is in a loop projectingfrom the HA0. Apathogenic viruses had but a single arginineresidue at the cleavage site and very few proteases were ableto cleave it. By contrast, the virulent strains were mutated,and contained multiple arginine and other basic residuesat the site. Cleavage could be archived by a wide rangeof host proteases which lent the capability for pantropicsystemic infection in a wide range of cells and tissues outsiderespiratory tract. The basis for differences in virulence forpoultry was of great scientific interest but provided no cluesto explain pandemic influenza of 1918–1919[12].
In his publication, Steinhauer[42] discussed the role thatco-infection of influenza virus with bacteria may play. Bac-teria can supply proteases which may bring about cleavagebeyond that which normally occurs in the respiratory epithe-lium thereby extending the cell tropisms of the virus.
4.4. Immunity and recovery from infections
Much of what is known about the immunology of in-fluenza is described in excellent reviews[1,3,57]. Only, abrief overview is given here. The basic pathogenesis andimmunology of influenza, in many respects, is like that formeasles virus infection[58]. The primary marker for resis-tance to and recovery from influenza virus infection is thatof humoral antibodies which are of complementary speci-ficity to the HA and NA antigens of the virus. HA antibodiesrestrict the virus by neutralization of infectivity and the NAantibodies restrict virus spread by interfering with release ofnewly replicated virus from the host cell. Loss of comple-mentary antibody through natural subsidence or by antigenicdrift accounts for reduction in resident humoral immunity.
Cell-mediated immunity[17,57,59–62]in man againstinfluenza is less well defined than that of humoral immu-nity. It is based on class I CD8+ cytotoxic T cell responseswhich usually appear within 3–4 days after infection.CD8+ cytotoxic T cells detect and lyse virus-infected host
cells and their specificity may be directed against HA, NP,M and PB2 epitopes which are more highly conserved thanthose concerned with humoral immunity. Class II CD4+ Thelper cells are of signal importance in facilitating bothhumoral and cellular immune responses and, in addition,they can exert cytolytic effects, though of lesser effect thanCD8+ cytotoxic T cells.
There is initial activation of innate immunity by patternrecognition of the virus components which brings aboutearly release of IL6 and IFN� [1] from epithelial cells. Stim-ulation of natural killer cells is of seminal importance in theprimary immune response. The ability of NS1 viral protein(Table 1) to inhibit the antiviral activity of interferon atteststo the essential role of this cytokine which normally upregu-lates MHC presentation of fragments of antigens by antigenpresenting cells.
As for measles, influenza virus infection causes transientdepression of delayed hypersensitivity through reduction ofblastogenesis of immune cells and probably through abortiveinfection of lymphocytes as well[1,17,57]. Such immuno-suppression might play a role in reducing resistance againstbacterial opportunists as is found with measles[58] andAIDS.
In overview, it is evident that both humoral andcell-mediated immunities have critically important roles toplay in control of influenza virus infection. Antibodies re-duce virus load and restrict re-infection. Cytotoxic T cellsdestroy virus-infected cells and may also provide cytokinesuppression. The specifics of cell-mediated immunity areclearly lacking, especially as relates to broad-spectrum im-mune responses to core antigens. The relative importanceof each of the two effector arms of the immune system isnot well defined for influenza.
5. Epidemiology of influenza A in historical andcontemporary perspective
Influenza is most commonly lethal in very young andelderly persons, transcribing a V-shaped age distributioncurve. During the 1918–1919 Spanish influenza pandemic,there was uniquely major lethality among persons 20–40years of age as well as in the very young and the old, tran-scribing a W-shaped curve (see[12,14,63]). Such patternhas not been prominent since that time.
Influenza occurrence is presently defined in terms of in-creased deaths from influenza and pneumonia which is inexcess of a pre-determined norm for all seasons of the year[11]. The surrogate influenza death rates are calculated bysubtraction of the numbers in the norm from the total deathsfor influenza and pneumonia.
Potter[11] determined that 10 pandemic influenza events,defined clinically and epidemiologically occurred during thepast 300 years, with an average of three for each century orone per 33 years. The pandemic of 1918–1919 was one of themost destructive events in medical history, and is estimated

3074 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
to have killed 20–40 million persons worldwide[8,13,14].The origin of the pandemic is unknown since it did appearin North America, Asia, and Europe at about the same time[12]. The pandemic disease[14,64], in its first wave, wasmild but changed to a very severe and lethal disease in itssecond wave. During the latter time, the virus appears to haveinfected pigs in which species it is retained to the presenttime. There were about 500,000 excess deaths in people inthe US during the 1918–1919 pandemic[65]. The pandemichas been shown to have been caused by H1N1 virus and isbeing investigated intensively at the present time[12]. Thepandemics of 1957 and 1968 are believed to have originatedin China [1,18,66] where ducks, pigs and human beingslived in close proximity. The PB1 as well as the HA andNA genes of the 1957 Asian virus were of avian origin[27].It was calculated[65] that the influenza-associated mortal-ity in the US numbered 69,800 persons in the 1957–1958pandemic event, and 33,800 persons in the 1968–1969 pan-demic. Influenza-associated deaths in the inter-pandemicyears is often cited at about 21,000 per year. The greatly in-creased numbers of deaths in pandemic years attract muchattention, but totaling the numbers[65] for the interpan-demic years of 1972–1992 in the USA, amounted to 426,000deaths, a cumulative mortality which greatly exceeds thetotal for the 1957 and 1968 pandemic years together. Eco-nomic costs are also very large. Kavet[67] estimated thatthe costs for influenza A in the epidemics of 1962–1963,1965–1966, and 1968–1969 in the USA amounted to US$3.1, 1.7, and 3.9 billion, respectively.
5.1. Beginnings that fueled the future
The discovery of influenza A virus in 1933[5] and thedevelopment and proof of efficacy of a prophylactic vaccineby the Commission on Influenza[68,69] during World WarII, marked the gradual transition of influenza epidemiologyfrom the clinic to the laboratory where cases of influenzacould be diagnosed based on specific viral etiology.
A prominent happening during the H1N1 virus era wasthe 1946–1947 outbreak of influenza virus A in which therewas lack of protection by the existing vaccine[70,71]. Thiswas to provide the impetus for a new program for influenzaresearch at the Walter Reed Army Institute of Research.
5.2. Scientific discoveries at the Walter Reed ArmyInstitute of Research
The author was recruited in 1948 to join the staff at WalterReed with the assignment “to study viral influenza in all itsaspects and to discover means to avert the next pandemicof influenza”. With the assignment came, the responsibilityto serve as the central research and surveillance laboratoryfor influenza which covered the worldwide deployment oftroops by the total US military establishment. Attention wasfocused immediately on whether the 1946–1947 event wasa pandemic or an epidemic, and on study of the cause for
failure of the then current vaccine to prevent the disease.The clue lay in the observation described earlier[70,71],that the antigenic specificity of the 1946–1947 virus differedmarkedly from that of the viral antigens in the vaccine. It hadbeen shown in numerous studies during previous years thatthere were antigenic differences between influenza A virusstrains, based on the findings in hemagglutination–inhibitioncomparisons, primarily, using ferret antisera[72–74], butthere was no attempt to provide a systematic investigationof the phenomenon.
It was noted in my laboratory that the measured viral anti-genic specificity, as determined using individual ferret antis-era, reflected, in part, differences between individual animalsin which the antisera were made. For preparing our antis-era, we substituted chickens which were injected intraperi-toneally using virus grown in embryonated hen’s eggs. Theviruses did not proliferate in the chickens but gave brisk an-tibody responses of high titer and were not biased in speci-ficity which had been noted between different ferret antisera.
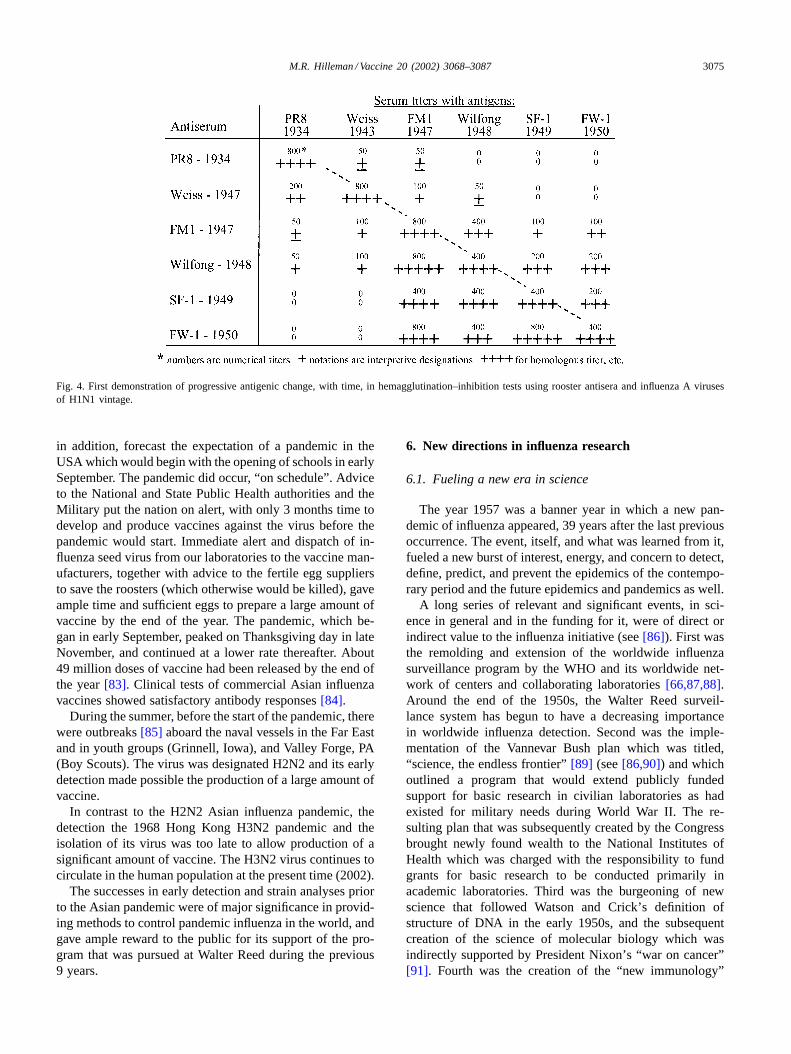
Our first studies[75,76] of immunologic specificitiesamong a number of influenza virus strains recovered indifferent years gave the breakthrough discovery (Fig. 4)that there was non-random progressive antigenic changein influenza A viruses isolated in successive years. This isnow referred to as antigenic drift, and has since shown tobe caused by an accumulation of point mutations[1,3,77].Further, studies[78] of influenza viruses recovered overextended time periods confirmed the drift phenomenon andprovided an early means to define and classify individualviruses [76]. Pioneering seroepidemiologic investigations[79] in our laboratories using sera from persons of differentages or collected in different years were consistent with andconfirmed the findings of progressive antigenic change ofinfluenza virus isolates with time. These studies providedtechnologies for study of influenza viruses in general andprovided the technologic means for rapid analysis to de-tect early pandemic occurrences as related in the followingsections.
In mid April of 1957, there were no unusual events ininfluenza detected by either our US military surveillanceoperation, or by the World Health Organization (WHO).By chance, the author read a report[80] in the 17 April1957, New York Times reporting an estimated 250,000 casesof respiratory disease among inhabitants of Hong Kong inwhich infected infants had glassy-eyed stare which was in-terpreted to be due to high fever. This seemed a clear alertfor pandemic influenza. An immediate dispatch to the Army406th Medical General Laboratory in Tokyo requesting pa-tient specimens netted virus by 17 May. Within 5 days, thevirus was defined and was shown to be antigenically differ-ent from anything seen before[81,82]. In addition, the seraof persons who had not been infected were shown to be de-void of antibody against the 1957 virus.
Worldwide press dispatches from Walter Reed on 22 Mayannounced the discovery of the new virus (first called “FarEast” and later called “Asian” influenza virus)[80–82]and,
M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3075
Fig. 4. First demonstration of progressive antigenic change, with time, in hemagglutination–inhibition tests using rooster antisera and influenzaA virusesof H1N1 vintage.
in addition, forecast the expectation of a pandemic in theUSA which would begin with the opening of schools in earlySeptember. The pandemic did occur, “on schedule”. Adviceto the National and State Public Health authorities and theMilitary put the nation on alert, with only 3 months time todevelop and produce vaccines against the virus before thepandemic would start. Immediate alert and dispatch of in-fluenza seed virus from our laboratories to the vaccine man-ufacturers, together with advice to the fertile egg suppliersto save the roosters (which otherwise would be killed), gaveample time and sufficient eggs to prepare a large amount ofvaccine by the end of the year. The pandemic, which be-gan in early September, peaked on Thanksgiving day in lateNovember, and continued at a lower rate thereafter. About49 million doses of vaccine had been released by the end ofthe year[83]. Clinical tests of commercial Asian influenzavaccines showed satisfactory antibody responses[84].
During the summer, before the start of the pandemic, therewere outbreaks[85] aboard the naval vessels in the Far Eastand in youth groups (Grinnell, Iowa), and Valley Forge, PA(Boy Scouts). The virus was designated H2N2 and its earlydetection made possible the production of a large amount ofvaccine.
In contrast to the H2N2 Asian influenza pandemic, thedetection the 1968 Hong Kong H3N2 pandemic and theisolation of its virus was too late to allow production of asignificant amount of vaccine. The H3N2 virus continues tocirculate in the human population at the present time (2002).
The successes in early detection and strain analyses priorto the Asian pandemic were of major significance in provid-ing methods to control pandemic influenza in the world, andgave ample reward to the public for its support of the pro-gram that was pursued at Walter Reed during the previous9 years.
6. New directions in influenza research
6.1. Fueling a new era in science
The year 1957 was a banner year in which a new pan-demic of influenza appeared, 39 years after the last previousoccurrence. The event, itself, and what was learned from it,fueled a new burst of interest, energy, and concern to detect,define, predict, and prevent the epidemics of the contempo-rary period and the future epidemics and pandemics as well.
A long series of relevant and significant events, in sci-ence in general and in the funding for it, were of direct orindirect value to the influenza initiative (see[86]). First wasthe remolding and extension of the worldwide influenzasurveillance program by the WHO and its worldwide net-work of centers and collaborating laboratories[66,87,88].Around the end of the 1950s, the Walter Reed surveil-lance system has begun to have a decreasing importancein worldwide influenza detection. Second was the imple-mentation of the Vannevar Bush plan which was titled,“science, the endless frontier”[89] (see[86,90]) and whichoutlined a program that would extend publicly fundedsupport for basic research in civilian laboratories as hadexisted for military needs during World War II. The re-sulting plan that was subsequently created by the Congressbrought newly found wealth to the National Institutes ofHealth which was charged with the responsibility to fundgrants for basic research to be conducted primarily inacademic laboratories. Third was the burgeoning of newscience that followed Watson and Crick’s definition ofstructure of DNA in the early 1950s, and the subsequentcreation of the science of molecular biology which wasindirectly supported by President Nixon’s “war on cancer”[91]. Fourth was the creation of the “new immunology”

3076 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
that has been funded for pursuit of the war against AIDSand has brought breakthroughs and understandings to ascience which had been long obscured in a black box ofmystery. These happenings were seminal to promoting so-phisticated scientific and technologic advances in generaland were utilized in creating an advanced understanding ofinfluenza.
6.2. New horizons
Closure of the decade of the 1980s’ found influenza re-searchers with a depth of understanding of the roles playedby drift and shift in HA and NA antigens of influenza viruseswhich provide the fuel for epidemic and pandemic disease.In recapitulation (Table 2), the H1N1 viruses persisted fromthe time of their discovery in 1933 to the Asian pandemicof 1957 and then disappeared from man, while continuingto infect pigs. The reemerging 1977 H1N1 Russian pan-demic virus was found to be the same virus as was circu-lating in 1950 and is believed to be an escapee from frozenstorage. The 1957 virus was comprised of a substitution ofH2 and N2 for the H1 and N1 components of the previousH1N1 virus[26]. The PB1 components of the 1957 and the1968 viruses were also derived from an avian virus[27].The 1968 Hong Kong H3N2 pandemic virus showed sub-stitution only of H3 for H2 of the 1957 agent[26]. H3N2viruses have shown very rapid antigenic drift since 1968[92].
What was left in 1990 was the enigma of the 1918–1919virus itself. Would the next pandemic be caused by a re-assortant as in 1957 and 1968 or would there be a farmore devastating alteration causing a pandemic such asin 1918? This became the new focus for influenza virusresearch.
6.3. Avian awakening
It has been long recognized that direct influenza virusinfections of zoonotic origin and, without reassortment,can occur with low frequency in man. These zoonoticagents derive mainly from pigs, poultry and horses andtypically show low virulence for man in all, but a smallproportion of individuals. Additionally, there is lack of se-rial communicability, as was amply illustrated in the smalloutbreak of swine influenza at Fort Dix, New Jersey, in1976 (see[93]) and in the avian influenza outbreak of 1997[94].
Renewed concern for zoonotic infections in man began in1997 when an epidemic of highly pathogenic H5N1 avianinfluenza occurred among domestic fowl in Hong Kong[94–96]. Special alarm was raised when 18 cases, with sixdeaths in man, were identified which occurred by direct in-fection from chickens, causing surprisingly severe disease.There was neither adaptation nor reassortment in man. Amassive campaign was waged by health officials to controlthe outbreak by slaughter of 1.4 million poultry in Hong
Kong. This ended the epidemic in poultry with no furtheroccurrence in human beings. Genetic probes[95,96] of thevirus revealed coding for polybasic amino acids at the HAcleavage site which has been found previously to be relatedto high level virulence of influenza in chickens. Discoveryof the H5N1 virus and its elimination by slaughter of thechicken population was seminal to the world of influenzacontrol.
Following elimination of H5N1 agent in 1997, the H9N2virus became widespread among poultry in Hong Kong[97–99]. In 1999, two cases of influenza caused by H9N2were found in man, but both were mild and self-limiting[97]and it was perceived not to give a public health threat. How-ever, human infections with zoonotic agents such as H5N1and H9N2 always raises the specter of becoming dangerousfor human beings if there is adaptation to man, or recombi-nation or reassortment with human influenza viruses. Sincethe reservoir for influenza subtypes is aquatic waterfowl, itmay be that domestic poultry, like pigs, may serve as anintermediary carrier to man[35].
7. Paleovirology of the 1918–1919 pandemic influenzavirus
7.1. Early expeditions to permafrost country
In spite of indirect knowledge and speculation which de-veloped over the decades as to what caused the pandemicof 1918–1919, the discovery of the virus itself became theholy grail for influenza scientists. It was deemed importantto define the composition of the 1918 virus itself, and todetermine why it was so virulent.
As related in Gina Kolata’s book—FLU[100], Drs.William Hale and John Hultin originated and pursuedthe concept that the virus which caused the pandemic of1918–1919 might be found in the bodies of humans whodied in the pandemic, and who, by chance, were buried inthe permafrost. Permafrost is found in the soil in areas, suchas in Alaska, where the ground remains frozen, even duringthe warm season of the year. Government maps existed[100] which told where Alaskan permafrost was known toexist.
These concepts led to expeditions to find and to exhumethe frozen bodies of victims of the 1918–1919 pandemic. Dr.John Hultin of the University of Iowa became the true pio-neer for “resurrection” of infected tissues from permafrost.His efforts were rewarded by the finding of suitable bodiesin his exhumations, in the summer of 1951, at Brevig Mis-sion in the Seward Peninsula of Alaska. A second such ex-pedition, carried out during the same summer, was the USMilitary Project George that was based at Nome and con-ducted as a joint scientific effort of the Harvard UniversitySchool of Public health, the Department of Epidemiologyof the University of Michigan, and the author’s laboratoryat the Walter Reed Army Institute of Research. The author

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3077
was the responsible laboratory officer for the project. Thecollections of tissue samples by both the Hultin and Wal-ter Reed expeditions failed to reveal viable influenza virus.Having no foresight or premonition of science to come, thesamples were destroyed.
7.2. Taubenberger’s triumph
First, resurrection and characterization of genes of the1918–1919 pandemic virus was made by Taubenbergerand coworkers[12,101] of the US Armed Forces Instituteof Pathology (AFIP). These workers searched the archivalfiles of formalin-fixed and paraffin-embedded autopsy tis-sues from 1918 influenza victims which were stored in theNational Tissue Repository of AFIP. Focus was on two sol-diers who died in the USA. A second source of virus wasfrom tissues newly collected by Hultin from a frozen bodyof a 1918 influenza victim at Brevig Mission in Alaska.Reconstruction of the genes obtained by PCR amplifica-tion and sequencing from the HA, NA, and NS segmentsshowed that the pandemic virus was indeed H1N1 as hadbeen anticipated from seroepidemologic investigations[22].The findings however did not provide answers to the ques-tions about virulence for man. There were no alterationseither in coding for the basic amino acid composition ofthe HA cleavage site[23] or for the �146 mutations thatmight have altered the tissue tropism of the virus. Therewas no evidence for alteration in the NS gene which wouldhave enhanced its anti interferon activity. As Taubenbergerand coworkers point out[12,23,101], virulence of influenzavirus is probably polygenic and new and interesting find-ings may derive from sequencing of the remaining five genesegments of the virus that is now in progress. Also, valu-able information may be revealed in comparative sequencestudies of N5N1 and other viruses.
The literature is replete with alternate interpretations ofthe Taubenberger data. All would agree with the H1N1 ge-netic identification of the 1918–1919 pandemic influenzavirus, but there are differences in explanations for the originsand the virulence of the virus[102,103]. Taubenberger[101]points out that while the pandemic virus is linked to avianviruses, the viral HA gene probably did not pass directly intoman in its pandemic form. Instead, it spent time in adaptingto an unidentified mammalian host through an unidentifiedmechanism. Webster[102], to the contrary, suggests suc-cessful direct transmission from birds to man in the mannerof the H5N1 outbreak in Hong Kong. Gibbs et al.[104] pro-poses that following an original switch of host from bird tomammal, the virus was adapted to both pigs and man. Ac-cording to Gibbs, there was a recombination event in whichthe H1 segment of the HA gene from pig virus was recom-bined with the H2 of the human virus. It was proposed thatsuch event was responsible for the sudden shift from the mildfirst wave to the severe second wave of the pandemic. Theenigma of the severity of the second pandemic wave may oc-cupy the attention of molecular biologists for years to come.
8. Vaccines
The evolution of vaccines, both killed and live, againstinfluenza of man were made possible by the discovery of thevirus itself in 1933[5]. Early experimental vaccines[105],both killed and live, were prepared using crude preparationsof infected mouse lung and chick embryo tissues, and weretested in human volunteers both in the US and the UK, withsuccesses.
8.1. Killed virus vaccines
8.1.1. Whole virusThe development of licensed killed whole virus vac-
cines against influenza relied on cultivation and purificationof the agent grown in the allantoic sac of embryonatedhen’s eggs[106]. Expanding on what was stated earlier,the first such vaccine was prepared by the Commissionon Influenza of the US Armed Forces EpidemiologicalBoard[68,69]. The virus was purified and concentrated byabsorption to and elution from red blood cells[157] andwas inactivated with formaldehyde. Though efficacious, thecrude preparation was soon replaced by Stanley’s[107]sharples centrifuge-purified vaccine which was the standardfor many years of routine production. Great improvementin purity of virus was achieved following developmentby Reimer et al.[108] of the continuous flow KII ultra-centrifuge with modification to permit density gradientcentrifugation. Filter membrane-purified and density gradi-ent centrifugation-purified vaccines were granted productlicenses to Merck in 1969 and 1970[80], respectively.
Killed virus vaccines[15,16] have been very successfulin preventing influenza. Present vaccine contains 15�g perdose each of H1N1 virus and of H3N2 type A virus, plusinfluenza B virus. Killed vaccine is considered[15] to have60–90% protective efficacy in children and adults with lesserprotection afforded in the elderly. These vaccines provedsafe and well tolerated. Unfortunately, they are underutilizedeven though there is a good rationale for use in underdevel-oped as well as developed countries[109]. All vaccines thatare given routinely are prepared using embryonated eggsproduced for hatching chicks and which derive from hensthat are ubiquitously infected with avian leukosis virus. Pres-ence of leukemia virus is not a problem for killed virus vac-cine since it is readily inactivated by the formaldehyde usedin the production process. Recent studies have shown thetechnical feasibility to replace egg-grown with mammaliancell culture (MDCK) propagated virus to prepare killed in-fluenza vaccine[110].
Because of unpredictable drift and shift in the HA andNA antigens, world wide influenza surveillance is now car-ried out principally by the WHO and its Global InfluenzaSurveillance Systems[111] with the purpose, as of now, ofselecting one H1N1 and one H3N2 influenza A virus eachyear which reflect the likely drift strain for the followingseason plus one influenza B virus which is also included

3078 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
in the trivalent vaccine. Selection of viruses for manufac-turing is made between February and April of each year,allowing sufficient time to produce vaccine and to deliverproduct by September or October of the same year. As re-lated earlier, in the pandemic year 1957, the fortunate earlydetection, outside the usual surveillance system[80–82],permitted a large amount of vaccine to be made and usedbefore the pandemic was over. In 1968, detection was toolate to prepare vaccine in significant amounts. The effortsthat were made to produce vaccine which would ward offthe “pseudo-pandemic” of swine influenza of 1976[93]is discussed later (Section 9.4). To facilitate routine vac-cine manufacture, the chosen viruses are reassorted[33,34]to provide an egg-adapted virus parent strain bearing thedesired HA and NA genes as described earlier.
8.1.2. Subunit vaccines
8.1.2.1. Split vaccines.Before means were developed toachieve high level purity of the killed influenza virus vaccine,some manufacturers developed ether- or detergent-split viralproducts which clearly were of reduced toxigenicity for man[112]. Such vaccine is now used widely throughout the worldand is especially recommended for use in children less than12 years of age.
Subunit vaccine given in a single dose is adequate foruse in immunizing persons who are primed or who haveimmunologic memory for antigens or epitopes of virusesagainst which they are expected to protect. To the contrary,split vaccines are very poorly immunogenic in personswho have not been primed through previous vaccinationor infection with homologous or antigenically related virus[83,112–115]and may require that two doses be given.
A cogent example of reduced immunogenicity resultingfrom the split process was provided by our own clinicalstudies in 1976 of whole and either split vaccines against
Table 3Comparison of hemagglutination–inhibition antibody responses in young children who were initially devoid of antibody and who were given a singledose of vaccine against H1N1 swine influenza virus, either whole or split subunit virus vaccine, in 1976
Age of subjects Vaccine Response
Kind Source Antigen doseCCA units
No. of percent with>1:20 antibody titer
Geometricmean titer
6–10 years Whole Merck & Co. 100 55/72 (76%) 33Whole Merrill National 100 57/86 (66%) 22
Split Wyeth 100 5/45 (11%) 6Split Wyeth 400 12/71 (17%) 8Split Parke Davis 100 2/50 (4%) 6Split Parke Davis 400 3/48 (6%) 6
3–5 years Whole Merck & Co. 100 14/15 (93%) 89Whole Merrill National 100 26.40 (65%) 23
Split Wyeth 100 2/29 (7%) 6Split Wyeth 200 1/10 (10%) 6Split Parke Davis 100 2/33 (6%) 6Split Parke Davis 200 0/9 (5%) 5
Based on data published by Hilleman[113].
the Fort Dix H1N1 swine virus (Table 3) [113]. There wasan overall progressively lesser antibody response to splitvaccine with decreasing age in the range of 18 years to6.5 months. This was most evident in children 10 years ofage or less, and especially at 6.5 months to 3 years of age.These findings were corroborated by Parkman et al.[114]in clinical studies.
The deficiencies in immune response to split vaccine inimmunologically inexperienced persons has largely goneunheeded because young children are not given influenzavaccine routinely. They do predict a likely lack of immuneresponse following split vaccine when protection againsta new shift pandemic virus is urgently needed. Becauseof these portents for future failures in pandemic threats,our laboratories continued to manufacture whole virus vac-cine. The health community, perhaps unaware of the factsand following recommendations for use of split vaccine inchildren less than 12 years of age, chose to use the splitvaccine which were of somewhat less reactogenicity[114].Our laboratories, not wishing to participate in marketing alikely inadequate vaccine in time of pandemic, withdrewfrom vaccine manufacture.
8.1.2.2. Conserved M2 protein vaccine.It is of great im-portance to influenza vaccine to develop means for circum-venting loss of effectiveness through antigenic drift and shift.The M2 protein of the virus is highly conserved antigenicallyand it has been shown that homologous antibody against itprevents infection in vitro and in mice in vivo[116,117].A promising approach to a universal influenza vaccine hasbeen made by immunizing mice with M2 protein[118].
8.1.3. DNA vaccineSlightly more than a decade past, Wolff et al.[119]
discovered that recombinant DNA plasmids bearing in-serted reporter gene sequences expressed foreign proteins

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3079
on injection into the muscle of experimental animals. Ap-plication of this discovery to a variety of different diseaseagents has produced a very large number of publicationsthat promise important practical applications in the develop-ment of DNA vaccines for use in human beings[120–123].Most notably, the DNA vaccine expresses its antigens inthe cytosol of antigen presenting cells, the fragments ofwhich can be displayed to the immune system by both theclasses I and II immunologic pathways. This field has beenextensively reviewed elsewhere[120–122]and will not beconsidered in detail here.
The potential importance of the DNA vector to influenzaA vaccine was shown in studies by Liu and coworkers[120]and Ulmer et al.[123] who demonstrated that mice devel-oped both humoral and cellular immunities following injec-tion of recombinant plasmids bearing gene sequences of theHA of the 1934 H1N1 PR8 virus, and the NP as well. Lethalintranasal challenge with virulent homologous PR8 virusshowed solid protection. More importantly, mice which wereinjected with recombinant DNA bearing the highly con-served NP of the H1N1 PR8 virus were near totally protectedagainst death when challenged intranasally with H3N2 pan-demic virus of 1968. The achievement of protection by 1934H1N1 DNA vaccine against 1968 pandemic H3N2 virus isvery promising and points a way toward possible develop-ment of a single vaccine containing the codon for conservedNP that might afford protection against all influenza Aviruses of man, present, past, and future. Such protection re-mains to be demonstrated for man and the licensure of DNAvaccines for humans must cross many safety hurdles beforeacceptance by the national regulatory authorities. Of specialinterest to influenza vaccine is the recent demonstration byOkuda et al.[124] that DNA vaccine expressing M geneproteins induced cell-mediated protective immunity in miceagainst homologous and heterologous influenza viruses.
8.1.4. Attenuated live influenza virus vaccinesadministered intranasally
Killed virus vaccine is the sole influenza immunogen thatis presently licensed for use in the USA. Alternative live in-fluenza virus vaccines have been used for many years in theformer Soviet Union[125] with apparent success. Intensiveresearch has been pursued in the USA for many years to de-velop similar live attenuated vaccines. The most importantadvance in this objective was made by Maassab et al.[16]at the University of Michigan who developed a cold adaptedattenuated reassortant, live influenza vaccine (CAIV) intowhich any desired HA or NA can be inserted. The vaccineis now under commercial pursuit with consideration for li-censure and dominates much of the present attention beinggiven toward the development of new influenza vaccines.
The Maassab trivalent live influenza A and B virus vac-cine is comprised of cold adapted attenuated influenza mas-ter strains in which HA and NA genes of contemporarysignificance replace the corresponding genes of the masterstrain by reassortment exchange. Attenuation of the master
strains of influenza A and B, in which growth is restrictedto the upper respiratory tract of man, was achieved by adap-tive propagation in eggs at low temperature (25◦C) whichbrought about stable alterations (mutations) in all three poly-merase genes, viz. PA, PB1, and PB2. Clinical studies inchildren, adults, and the elderly of live vaccine producedin embryonated hen’s eggs have generally been promising[126–129]see with respect to efficacy and safety when ap-plied to use in young children. Simplicity in administrationby the nasal route with high level efficacy in young childrenare favorable attributes.
There are many pros and cons in considerations for gen-eral use of the live vaccine. Recently, Pfleiderer et al.[130]of the German Regulatory Authority at the Paul Ehrlich In-stitute in Langen have published an assessment of the livevaccine from the view of risk-benefit, pointing out that qual-ity, efficacy, safety and practicality are of tantamount impor-tance and need to be considered in comparison with thoseof killed vaccine. Some highlights of their discussion pointto: the need to ascertain safety of the live vaccine in theimmunosuppressed; to measure cell-mediated as well as hu-moral responses; to assure storage stability for infectivity ofthe product; to ascertain whether there is interference amongthe vaccine strains that may affect the individual immune re-sponse in vaccines; to determine whether there is retentionof the genetic integrity of the viruses following prolifera-tion in the vaccinated person; and to measure whether theimmune response engendered following repeated use of thesame master strains from year to year will be restricted bydevelopment of immunity against the six remaining compo-nents (not HA or NA) of the vaccine.
Safety issues include assurance of freedom from in-digenous chicken viruses in vaccine produced using hens’eggs; allergic reactions from and allergic sensitization tochick proteins; breakdown of the mucosal barrier allowingsystemic entry of resident microbial pathogens; and thequestion of whether there is or can be genetic reversion invirulence of the vaccine components on passage in vaccinesand their contacts. Practical issues exist which relate tothe time required to proceed from wild virus identificationto vaccine production and release, especially if extensivegenetic studies of virus recovered from human recipientsof the vaccine will be required annually and if vaccines ofnew reassortant composition will be subject to new licensecertification each year. Important also will be the determi-nation of the role that live vaccine might play in private useand in public health application in various age groups.
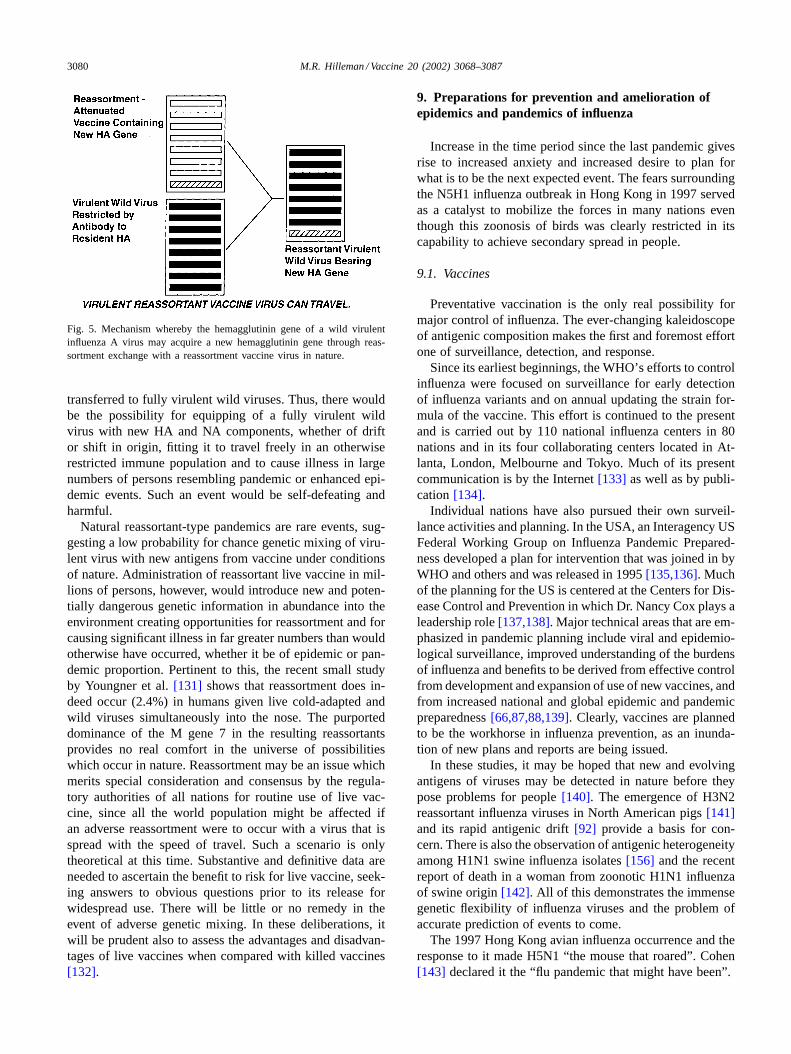
A significant issue concerning safety was raised in thePfleiderer publication[130] which relates to possible ad-verse effects of simultaneous infection in a person with bothwild virulent virus and live vaccine virus which results ingenetic exchange by the same reassortment mechanism bywhich the Asian in 1957 and the Hong Kong in 1968 in-fluenza pandemics were created (seeFig. 5). In an adverseevent, the reassortant vaccine itself might be the source forthe new drift or shift HA and NA genes which would be
3080 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
Fig. 5. Mechanism whereby the hemagglutinin gene of a wild virulentinfluenza A virus may acquire a new hemagglutinin gene through reas-sortment exchange with a reassortment vaccine virus in nature.
transferred to fully virulent wild viruses. Thus, there wouldbe the possibility for equipping of a fully virulent wildvirus with new HA and NA components, whether of driftor shift in origin, fitting it to travel freely in an otherwiserestricted immune population and to cause illness in largenumbers of persons resembling pandemic or enhanced epi-demic events. Such an event would be self-defeating andharmful.
Natural reassortant-type pandemics are rare events, sug-gesting a low probability for chance genetic mixing of viru-lent virus with new antigens from vaccine under conditionsof nature. Administration of reassortant live vaccine in mil-lions of persons, however, would introduce new and poten-tially dangerous genetic information in abundance into theenvironment creating opportunities for reassortment and forcausing significant illness in far greater numbers than wouldotherwise have occurred, whether it be of epidemic or pan-demic proportion. Pertinent to this, the recent small studyby Youngner et al.[131] shows that reassortment does in-deed occur (2.4%) in humans given live cold-adapted andwild viruses simultaneously into the nose. The purporteddominance of the M gene 7 in the resulting reassortantsprovides no real comfort in the universe of possibilitieswhich occur in nature. Reassortment may be an issue whichmerits special consideration and consensus by the regula-tory authorities of all nations for routine use of live vac-cine, since all the world population might be affected ifan adverse reassortment were to occur with a virus that isspread with the speed of travel. Such a scenario is onlytheoretical at this time. Substantive and definitive data areneeded to ascertain the benefit to risk for live vaccine, seek-ing answers to obvious questions prior to its release forwidespread use. There will be little or no remedy in theevent of adverse genetic mixing. In these deliberations, itwill be prudent also to assess the advantages and disadvan-tages of live vaccines when compared with killed vaccines[132].
9. Preparations for prevention and amelioration ofepidemics and pandemics of influenza
Increase in the time period since the last pandemic givesrise to increased anxiety and increased desire to plan forwhat is to be the next expected event. The fears surroundingthe N5H1 influenza outbreak in Hong Kong in 1997 servedas a catalyst to mobilize the forces in many nations eventhough this zoonosis of birds was clearly restricted in itscapability to achieve secondary spread in people.
9.1. Vaccines
Preventative vaccination is the only real possibility formajor control of influenza. The ever-changing kaleidoscopeof antigenic composition makes the first and foremost effortone of surveillance, detection, and response.
Since its earliest beginnings, the WHO’s efforts to controlinfluenza were focused on surveillance for early detectionof influenza variants and on annual updating the strain for-mula of the vaccine. This effort is continued to the presentand is carried out by 110 national influenza centers in 80nations and in its four collaborating centers located in At-lanta, London, Melbourne and Tokyo. Much of its presentcommunication is by the Internet[133] as well as by publi-cation[134].
Individual nations have also pursued their own surveil-lance activities and planning. In the USA, an Interagency USFederal Working Group on Influenza Pandemic Prepared-ness developed a plan for intervention that was joined in byWHO and others and was released in 1995[135,136]. Muchof the planning for the US is centered at the Centers for Dis-ease Control and Prevention in which Dr. Nancy Cox plays aleadership role[137,138]. Major technical areas that are em-phasized in pandemic planning include viral and epidemio-logical surveillance, improved understanding of the burdensof influenza and benefits to be derived from effective controlfrom development and expansion of use of new vaccines, andfrom increased national and global epidemic and pandemicpreparedness[66,87,88,139]. Clearly, vaccines are plannedto be the workhorse in influenza prevention, as an inunda-tion of new plans and reports are being issued.
In these studies, it may be hoped that new and evolvingantigens of viruses may be detected in nature before theypose problems for people[140]. The emergence of H3N2reassortant influenza viruses in North American pigs[141]and its rapid antigenic drift[92] provide a basis for con-cern. There is also the observation of antigenic heterogeneityamong H1N1 swine influenza isolates[156] and the recentreport of death in a woman from zoonotic H1N1 influenzaof swine origin[142]. All of this demonstrates the immensegenetic flexibility of influenza viruses and the problem ofaccurate prediction of events to come.
The 1997 Hong Kong avian influenza occurrence and theresponse to it made H5N1 “the mouse that roared”. Cohen[143] declared it the “flu pandemic that might have been”.

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3081
9.2. Global laboratory against influenza
The rapid advances in genomics and in technologies forrapid throughput analysis of the genetic composition of mi-crobes has reached into the program for influenza surveil-lance. This new twist[144] would make it feasible to includesurveillance in animals as well as in people to detect whatmay be taking place in nature and to get an advance noticefar before a pandemic event.
A very important recent development in influenzapreparedness came at a meeting on “automation in threatreduction and in infectious disease research”, which wasorganized by Layne et al.[145] and was held at the US Na-tional Academy of Sciences during April of 1999. A specialmeeting was convened by Dr. Scott Layne, who is a physi-cist, physician, and public health specialist, to structure aproposed global laboratory for forensic detection of humanpathogens[111,146,147], especially influenza. It was deter-mined that such effort would bring a quantum leap in globalinfluenza surveillance and vaccine strain selection whichwould harness automated laboratory and bioinformationfirepower. This rapid throughput laboratory would focus ondetermining epidemiologic phenotypic and genotypic fea-tures of a huge number of current influenza viruses, withthe targeted purpose to apprehend new and important viralstrains for the purpose to notify, and to develop vaccines wellin advance of a serious event. Such a laboratory could be ofextreme importance in detection and analysis of a number ofemerging infectious disease threats, and most importantly,in its application to bioweapons detection and bioterror-ism. Influenza surveillance could fit very well into such aprogram.
The proposed global laboratory network would be expen-sive, but influenza might only be included in a major worldeffort that would allow for forensic detection of clandes-tine operations and for combat both against biowarfare andbioterrorism and emerging diseases as well. A likely initialprogram might be to develop and equip a single forensic andanalytic laboratory that would serve as a model for eventualevolvement of a worldwide networking operation.
9.3. Chemotherapy in controlling influenza
It is clear that the aim for preventing of pandemic in-fluenza by vaccines cannot be relied upon as a means foreffective control of influenza, especially because of the shorttime between strain detection and selection and need for theproduct. Short of successful development of a vectored sin-gle vaccine that will protect against all subtypes, such as apromised by the experimental DNA vaccine which expressesconserved core antigens[120,123], or by an M2 channelantigen vaccine[116–118]which is highly conserved anti-genically other approaches will be needed. An alternativemay lie with antiviral drug research which shows promiseas a broad-spectrum means to control influenza. Four dif-ferent chemotherapeutic substances now exist[2,148–151],
Amantadine and Rimantadine[2] function by blocking theM2 ion channel protein of the virus. Both drugs may beof substantive benefit in treating influenza A but not with-out side effects on the central nervous system, the liver andkidneys and by emergence of drug resistance. More usefuldrugs are Zanamivir and Osteltamivir which block the ac-tion of NA to prevent release of newly formed virus fromthe infected host cell and its spread within the host. Lessis known about the efficacy and safety of the NA inhibitorsthan the Amantadines. These four drugs are clearly not theend in chemotherapy of influenza, but serve as examples andmodels in the search for new and improved drugs which be-come increasingly possible as the distinctions between viralmetabolism and cellular metabolism[152] are defined.
9.4. Response to the “pandemic” of swine influenza of1976 which did not occur: Zeal, Flawed Science, andMisguided Public Policy
An opportunity to exercise due diligence and good judg-ment in the face of a threatened pandemic of influenza (see[93] for review) came with the occurrence of a very smalloutbreak of H1N1 swine influenza among military person-nel at Fort Dix, New Jersey[153], in February 1976. A sin-gle case of influenza of zoonotic swine exposure occurredin a new trainee with resulting unusual occurrence of sec-ondary cases under the conditions of crowding and stress atthe training center. The finding, though infrequent, of anti-bodies against the swine virus in certain of the human pop-ulations on the post, and at large, was believed by some toindicate that there was a widespread seeding of the virus inthe US in spite of the fact that H1N1 virus had circulated upto 1957 and in spite of the fact that some military personnelduring WWII were immunized with killed vaccine whichcontained the swine virus. The findings in surveillance forcases and for the virus indicated that the agent had not spreadbeyond Fort Dix and that need for aggressive planning andimplementation was not indicated.
In the absence of adequate information during the earlytime period following the Fort Dix event, and with thespecter of a repeat of the pandemic influenza of 1918–1919,it was not unreasonable for scientists and government agen-cies to seek the high ground and to declare that prudent ac-tion should be taken to avert a possible, but uncertain, majorpandemic threat through rapid development and stockpilingof vaccine. It was believed that governmental leadership andimplementation were needed for successful pursuit of theinitiative.
A small group of influenza scientists, including the author,met with President Ford in April of 1976 to discuss thematter. This was followed by a presidential announcementof an intended mass vaccination initiative. Money requestedfor vaccine production was appropriated by the Congress.In retrospect, and in view of what ultimately happened, thescientific group had mislead the President through its failureto state that it was important to stockpile but not to use the
3082 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
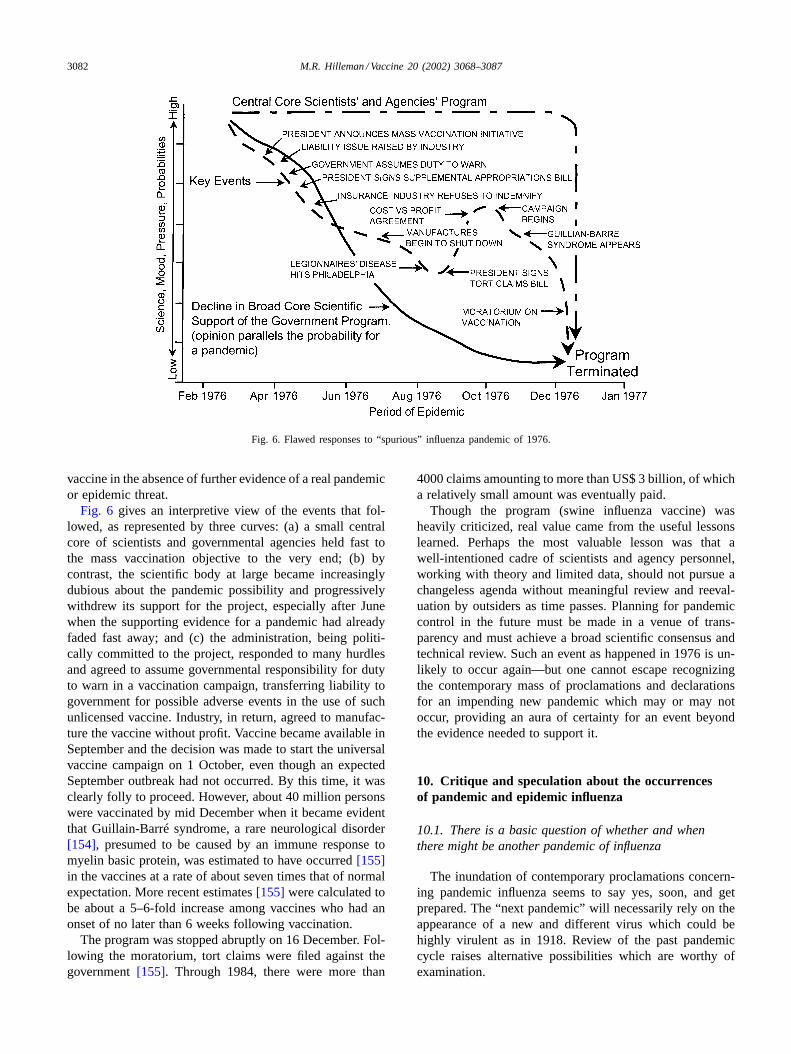
Fig. 6. Flawed responses to “spurious” influenza pandemic of 1976.
vaccine in the absence of further evidence of a real pandemicor epidemic threat.
Fig. 6 gives an interpretive view of the events that fol-lowed, as represented by three curves: (a) a small centralcore of scientists and governmental agencies held fast tothe mass vaccination objective to the very end; (b) bycontrast, the scientific body at large became increasinglydubious about the pandemic possibility and progressivelywithdrew its support for the project, especially after Junewhen the supporting evidence for a pandemic had alreadyfaded fast away; and (c) the administration, being politi-cally committed to the project, responded to many hurdlesand agreed to assume governmental responsibility for dutyto warn in a vaccination campaign, transferring liability togovernment for possible adverse events in the use of suchunlicensed vaccine. Industry, in return, agreed to manufac-ture the vaccine without profit. Vaccine became available inSeptember and the decision was made to start the universalvaccine campaign on 1 October, even though an expectedSeptember outbreak had not occurred. By this time, it wasclearly folly to proceed. However, about 40 million personswere vaccinated by mid December when it became evidentthat Guillain-Barré syndrome, a rare neurological disorder[154], presumed to be caused by an immune response tomyelin basic protein, was estimated to have occurred[155]in the vaccines at a rate of about seven times that of normalexpectation. More recent estimates[155] were calculated tobe about a 5–6-fold increase among vaccines who had anonset of no later than 6 weeks following vaccination.
The program was stopped abruptly on 16 December. Fol-lowing the moratorium, tort claims were filed against thegovernment[155]. Through 1984, there were more than
4000 claims amounting to more than US$ 3 billion, of whicha relatively small amount was eventually paid.
Though the program (swine influenza vaccine) washeavily criticized, real value came from the useful lessonslearned. Perhaps the most valuable lesson was that awell-intentioned cadre of scientists and agency personnel,working with theory and limited data, should not pursue achangeless agenda without meaningful review and reeval-uation by outsiders as time passes. Planning for pandemiccontrol in the future must be made in a venue of trans-parency and must achieve a broad scientific consensus andtechnical review. Such an event as happened in 1976 is un-likely to occur again—but one cannot escape recognizingthe contemporary mass of proclamations and declarationsfor an impending new pandemic which may or may notoccur, providing an aura of certainty for an event beyondthe evidence needed to support it.
10. Critique and speculation about the occurrencesof pandemic and epidemic influenza
10.1. There is a basic question of whether and whenthere might be another pandemic of influenza
The inundation of contemporary proclamations concern-ing pandemic influenza seems to say yes, soon, and getprepared. The “next pandemic” will necessarily rely on theappearance of a new and different virus which could behighly virulent as in 1918. Review of the past pandemiccycle raises alternative possibilities which are worthy ofexamination.

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3083
Considerations for pandemic occurrence might be wellserved first by looking at viral composition and virulence ofthe viruses of the past. Within the 20th century there werethree true pandemics of influenza. It was only the 1918–1919pandemic that was exceedingly lethal in contrast to the 1957and 1968 outbreaks which were not highly lethal, and weredistinguished mainly by increases in numbers of cases overexpected annual epidemic prevalence. The 1957 and 1968pandemics were caused by reassortant viruses; while theorigin of the 1918–1919 is still to be determined. The pan-demics of 1889 and 1900, respectively, were proposed tobe caused by H2N2 and H3N8 based on seroepidemiologicfindings[22,23].
The 1918 pandemic was likely caused by a unique recom-bination virus (seeSection 7.2) which included both avianand mammalian genetic elements and which may have re-quired several years residence in an unknown mammalianspecies before significant infections occurred in man. Withinthe small universe of experience, the 1918 virus was uniquein its virulence that was of chance occurrence. There is noevidence to support a belief that its unusual composition andhigh level toxicity will be duplicated in future pandemics.The H5N1chicken influenza threat may be more perceivedthan real since, short of reassortment with a human virus,zoonotic infections are of highly limited progression evenfrom the primary person to a second person.
10.2. Recycling of pandemic viruses
Review of both substantiated and speculative information(seeTable 2andSections 3.2 and 10.1) about past pandemicsled to assembly ofTable 4, which might be somewhatmore evidence-based and of greater predictive reliabilitythan either of the writings of Nostradamus or the Farmers’Almanac. Assuming that the seroepidemiologic definitionsrelating to the 1889 and 1900 pandemics are correct, it can beconcluded that pandemic influenza of man during the more
Table 4Concepts and speculations relating to the dynamics for the appearance, disappearance and reappearance of pandemic H subtypes in the light of recyclingH1, H2, and H3 subtypes
Pandemic introduction Years prevalent Years from initial appearanceto reappearancesa
Comment
Year Subtype
1889 H2N2 11 Seroepidemiology1900 H3N8 18 Seroepidemiology1918 H1N1 391957 H2N2 11 681968 H3N2 >34 681977 H1N1 >25 59 Calculation based on premature reintroduction of H1N1
from stored source. Expected appearance was for 1986a
1986 H1N1 >16 68
Predictive speculation for reappearance2025 H2N2 682036 H3N2 682054 H1N1 68 Calculation based on reappearance in 1986
a According to the 68-year cycle concept from appearance to reappearance.
than 110 years from 1889 to 2002 was restricted to threeHA subtypes, H1, H2, and H3. Interpreting that informationas revealed inTable 4, there is a speculative possibility thatan interval of about 68 years must pass before a particular Hserotype can reappear as the cause for a new pandemic. Bycoincidence, this is about the length of the contemporaryhuman lifespan. A 68-year recurrence restriction, if real,would suggest that there may need to be a correspondingapproximate 68-year period before sufficient subsidence ofhost immunity, humoral, cellular or whatever, before a pastvirus can regain access and become established as a “new”human influenza A virus in the human population.
There was a complicating factor in creating the conceptof a 68-year cycle for development of host population recep-tivity. This complication arose from the “premature” reap-pearance of the H1N1 virus in 1977 rather than in 1986, 9years before it would be expected according to the 68-yearconcept. It will be recalled that the reappearance of H1N1in 1977 was not a natural event, but, rather, the result of analmost certain escape of 1950 H1N1 virus from frozen stor-age[28] in an icebox (seeSection 3.2). Such virus was likelyto have been able to establish itself in a sufficiently largepopulation of young persons who had been born since 1957.Looking at the big picture, historically, it is both atypicaland remarkable for influenza A that two H subtypes, H3N2and H1N1 have been able to circulate together since 1977and to infect large numbers of persons to the present time.
If the 68-year, or thereabouts, concept for recycling fromappearance to reappearance of a particular subtype is to beconsidered real, then it is evident that H2N2 should be thecause for the next pandemic and the banner year shouldbe 34 years hence in 2025. By the same reasoning, H3N2should follow in 2036. H1N1, after an absence, should reap-pear in 2052 assuming its normal time for reappearance was1986, having been skewed by an artificial reappearance in1977. The bottom line in these speculations may be that thehost population itself may be as important as the antigenic

3084 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
differences among the H1, H2, and H3 viruses themselves indefining the possibility for reappearances in new pandemics.Clearly, there may be multifactorial “extra ingredients” fac-tors such as a role for cell-mediated immunity which, atpresent, must have an importance which has not been dis-closed to date.
The occurrence in nature of new reassortant viruses mightbe far more frequent than we commonly think. These maynot find a foothold since a permissive host population, basedon more than we know at present, might be required inorder for them to reappear in epidemic or pandemic events.Such speculation, irrespective of lack of definitive evidence,does give a different way to view the pandemic virus cycleand provides a new approach to predictive epidemiology ofinfluenza. Persons who expect to be alive in 2036 may bepresently amused but will also be the people who will be ableto determine whether the concept is one of sense or nonsense.
10.3. Assessing the importance of pandemic andepidemic disease
As noted inSection 5, the sum total of excess deathsresulting from interpandemic epidemics greatly exceeds thetotal that occur from pandemic events. The importance ofthis observation is readily confirmed in the events whichhave followed 1930, for example. If one assumes an ex-cess mortality of a conservative 20,000 deaths per yearin interpandemic years from 1930 to 2000, there wereabout 1,300,000 excess deaths. By contrast, there were nomore than 130,000 excess deaths in the collective years of1957–1958, 1968–1969 and 1977–1978.
This would indicate an approximate 10-fold greater im-portance of collective epidemic influenza compared with thecollective pandemic disease and would point to where re-sources and emphasis on control of pandemic influenza byvaccine might be placed. There is the low probability ofbeing able to control pandemic disease by vaccines as con-trasted with proven control of epidemic influenza. The 1957event was a partial success for pandemic control and 1968was a failure. Because of this, there is grave need for break-throughs and for successes in the development of safe andeffective antiviral drugs (seeSection 9.3) against influenza.It will be important also that influenza be included in anyof the systems proposed for surveillance against biologicalweapons and spontaneous emerging infectious agents (seeSection 9.2).
References
[1] Wright PF, Webster RG. Orthomyxoviruses. In: Knipe DM, HowleyPM, editors. Fields virology, vol. 1. 4th ed. Philadelphia: LippincottWilliams and Wilkins, 2001. p. 1533–79.
[2] Lamb RA, Krug RM. Orthomyxoviridae: the viruses and theirreplication. In: Knipe DM, Howley PM, editors. Fields virology,vol. 1. 4th ed. Philadelphia: Lippincott Williams and Wilkins, 2001.p. 1487–531.
[3] Kilbourne ED. A history of influenza virology. In: KoprowskiH, Oldstone MBA, editors. Microbe hunters—then and now.Bloomington (IL): Medi-Ed Press, 1996. p. 187–204.
[4] Shope RE. Swine influenza. Part III. Filtration experiments andetiology. J Exp Med 1931;54:373–85.
[5] Smith W, Andrewes CH, Laidlaw PP. A virus obtained frominfluenza patients. Lancet 1933;2:66–8.
[6] Francis Jr T. A new type of virus from epidemic influenza. Science1940;92:405–8.
[7] Francis Jr T, Quilligan Jr JJ, Minuse E. Identification of anotherepidemic respiratory disease. Science 1950;112:495–7.
[8] WHO Memorandum. A revised system of nomenclature forinfluenza viruses. Bull. WHO 1980;58:585–91.
[9] Webster RG. Influenza viruses. In: Webster RG, Granoff A, editors.Encyclopedia of virology, vol. 2. New York: Academic Press, 1994.p. 709–15.
[10] Simonsen L. The global impact of influenza on morbidity andmortality. Vaccine 1999;17(Suppl):S3–S10.
[11] Potter CW. A history of influenza. J Appl Microbiol 2001;91:572–9.[12] Taubenberger JK, Reid AH, Janczewski TA, Fanning TG. Integrating
historical, clinical and molecular genetic data in order to explainthe origin and virulence of the 1918 Spanish influenza virus. PhilTrans R Soc Lond B 2001;356:1829–39.
[13] Crosby AW. Flu and the American expeditionary force. In: Epidemicand peace, 1918. Wesiport (CT): Greenwood Press, 1976. p. 145–70.
[14] Crosby AW. America’s forgotten pandemic: the influenza of 1918.Cambridge: Cambridge University Press, 1989. 329 p.
[15] Cox NJ, Subbarao K. Influenza. Lancet 1999;354:1277–82.[16] Maassab HR, Herlocher ML, Bryant ML. Live influenza vaccines.
In: Plotkin SA, Orenstein WA, editors. Vaccines. 3rd ed.Philadelphia: Saunders, 1999. p. 909–27.
[17] Kantzler GH, Lauteria SF, Cusumano CL, Lee JD, Ganguly R,Waldman RH. Immunosuppression during influenza virus infection.Infect Immun 1974;10:996–1002.
[18] Webster RG, Bean WJ, Goren OT, Chambers TM, Kawaoka Y.Evolution and ecology of influenza A viruses. Microbiol Rev1992;56:153–79.
[19] Röhm C, Zhou N, Suss J, Mackenzie J, Webster RG.Characterization of a novel influenza hemagglutinin, H15. Criteriafor determination of influenza A subtypes. Virology 1996;217:508–16.
[20] Zhou NN, Kennedy F, Shortridge KF, Claas ECJ, Kraus SL, WebsterRG. Rapid evolution of H1N1 influenza viruses in chickens in HongKong. J Virology 1999;73:3366–74.
[21] Snacken R, Kendal AP, Haaheim LR, Wood JM. The next influenzapandemic: lessons from Hong Kong, 1997. Emerg Infect Dis1999;5:195–203.
[22] Dowdle WR. Influenza A virus recycling revisited. Bull Who1999;77:820–8.
[23] Reid AH, Taubenberger JK. The 1918 flu and other pandemics:over there and back again. Lab Invest 1999;79:95–101.
[24] Docso CC, Couch RB, Six HR, Young JF, Quarles JM. Sporadicoccurrence of zoonotic swine influenza virus infection. J ClinMicrobiol 1984;20:833–5.
[25] Ito T, Kawaoka Y. Host range barrier of influenza A viruses. VetMicrobiol 2000;74:71–5.
[26] Scholtissek C, Rohde W, von Hoyningen V, Rott R. On the originof the human influenza virus subtypes H2N2 and H3N2. Virology1978;87:13–20.
[27] Kawaoka Y, Krauss S, Webster RG. Avian-to-human transmissionof the PB1 gene of influenza A viruses in the 1957 and 1968pandemics. J Virol 1989;63:4603–8.
[28] Nakajima K, Desselberger U, Palese P. Recent human influenza Aviruses are closely related genetically to strains isolated in 1950.Nature 1978;274:334–9.
[29] Palese P, Young JF. Molecular epidemiology of influenza virus.In: Palese P, Kingsbury DW, editors. Genetics of influenza viruses.New York: Springer, 1983. p. 321–36.

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3085
[30] Davenport FM, Hennessy AV, Francis Jr T. Epidemiologicand immunologic significance of age distribution of antibody toantigenic variants of influenza virus. J Exp Med 1953;98:641–56.
[31] Rohde W, Scholtissek C. On the origin of the gene coding for aninfluenza A virus nucleocapsid protein. Arch Virol 1980;64:213–33.
[32] Hirst GK, Gotlieb T. The experimental production of combinationforms of virus. Part II. A study of serial passage in the allantoicsac of agents that combine the antigens of two distinct influenzaA strains. J Exp Med 1953;98:53–70.
[33] Kilbourne ED, Murphy JS. Genetic Studies of influenza viruses.Part I. Viral morphology and growth capacity in exchangeablegenetic traits. Rapid inovo adaptation of early passage Asian strainby combination with PR8. J Exp Med 1960;11:387–406.
[34] Kilbourne ED. Future influenza vaccines and the use of geneticrecombinants. Bull Who 1969;41:643–5.
[35] Webby RJ, Webster RG. Emergence of influenza A viruses. PhilTrans R Soc Lond B 2001;356:1817–28.
[36] Memoranda. Cell culture as a substitute for the production ofinfluenza vaccines: memorandum from a WHO meeting. Bull Who1995;73:431–5.
[37] Kistner O, Barrett PN, Mundt W, Reiter M, Schober-BendixenS, Dörner F. Development of a mammalian cell (Vero) derivedcandidate influenza virus vaccine. Vaccine 1998;16:960–8.
[38] Govorkova EA, Kaverin NV, Guboreva LV, Meignier B, WebsterRG. Replication of influenza A viruses in a Green Monkey kidneycontinuous cell line (Vero). J Infect Dis 1995;172:250–3.
[39] Zhao K, Singh M, Malaskevitch CH, Kim P. Structuralcharacterization of the human respiratory syncytial virus fusionprotein core. Proc Natl Acad Sci USA 2000;97:14172–7.
[40] Merz DC, Scheid A, Choppin PW. Importance of antibodies to thefusion glycoprotein of paramyxoviruses in the prevention of spreadof infection. J Exp Med 1980;151:275–88.
[41] Chen J, Skehel JJ, Wiley DC. N- and C-terminal residues combinein the fusion—pH influenza HA2 subunit to form an N capthat terminates the triple stranded coil. Proc Natl Acad Sci USA1999;96:8967–72.
[42] Steinhauer DA. Role of hemagglutinin cleavage for thepathogenicity of influenza virus. Virology 1999;258:1–20.
[43] Dutch RE, Hagglund RN, Nagel MA, Paterson RG, Lamb RA.Paramyxovirus fusion (F) protein: a conformational change oncleavage activation. Virology 2001;281:138–50.
[44] Chan DC, Kim PS. HIV entry and its inhibition. Cell 1998;93:681–4.
[45] Carr CM, Chaudhry C, Kim PS. Influenza hemagglutinin isspring-loaded by a metastable native conformation. Proc Natl AcadSci USA 1997;94:14306–13.
[46] Eckert DM, Malashkevich VN, Kim PS. Crystal structure ofGCN4-plQl, a trimeric coiled coil with buried polar residues. J MolBiol 1998;284:859–65.
[47] Oakley MG, Hollenbeck JJ. The design of anti-parallel coiled coils.Curr Opin Struct Biol 2001;11:450–7.
[48] La Casse RA, Follis KE, Scarborough JD, Littman DR, NunbergJH. Fusion competent vaccines: broad neutralization of primaryisolates of HIV. Science 1999;283:357–62.
[49] Fouts TR, Tuscan R, Godfrey K, Reitz M, Hone D, Lewis G, etal. Expression and characterization of a single-chain polypeptideanalogue of the human immunodeficiency virus type 1, gp120–CD4receptor complex. J Virol 2000;74:11427–36.
[50] Binley JM, Sanders RW, Clas B, Schuelke N, Master A, Guo Y, etal. A recombinant human immunodeficiency virus type 1 envelopeglycoprotein complex stabilized by an intermolecular disulfide bondbetween gp120 and gp41 subunits is an antigenic mimic of thetrivirion-associated structure. J Virol 2000;74:627–43.
[51] Peters BS. HIV immunotherapeutic vaccines. Antiviral ChemChemother 2000;11:311–20.
[52] Root MJ, Kay MS, Kim PS. Protein design of an HIV entryinhibitor. Science 2001;291:884–8.
[53] Turpin JA, Howard Z. Inhibitors of HIV cellular fusion. Exp OpinTher Pat 2000;10:1899–909.
[54] LaBranche CC, Galasso G, Moore JP, Bolognesi DP, Hirsch MS,Hammer SM. HIV fusion and its inhibition. Antivir Res 2001;50:95–115.
[55] Brelot A, Alizon M. HIV-1 entry and how to block it. AIDS2001;15(Suppl):S3–5.
[56] Buzko OV, Shokat SM. Blocking HIV entry. Nat Struct Biol1999;6:906–8.
[57] Karzon DT. Cytotoxic T cells in influenza immunity. Semin Virol1996;7:265–71.
[58] Hilleman MR. Current overview of the pathogenesis andprophylaxis of measles with focus on practical implications. Vaccine2002;20:652–65.
[59] Reed WP, Olds JW, Kisch AL. Decreased skin-test reactivityassociated with influenza. J Infect Dis 1972;125:398–402.
[60] Reed WP, Olds JW, Kisch AI. Decreased skin test reactivityassociated with influenza. J Infect Dis 1972;125:398–402.
[61] Lamb JR, McMichael AJ, Rothbard JB. T cell recognition ofinfluenza viral antigens. Human Immunol 1987;19:79–89.
[62] Townsend ARM, Rothbard J, Gotch FM, Bahadur G, Wraith D,McMichael AJ. The epitopes of influenza nucleoprotein recognizedby cytotoxic T lymphocytes can be defined with short syntheticpeptides. Cell 1986;44:959–66.
[63] Luk J, Gross P, Thompson WW. Observations on mortality duringthe 1918 influenza pandemic. Clin Infect Dis 2001;33:1375–8.
[64] Lauer WG, Bischofberger N, Webster RG. The origin and controlof pandemic influenza. Persp Biol Med 2000;43:173–92.
[65] Klimov A, Simonsen L, Fukuda K, Cox N. Surveillance and impactof influenza in the United States. Vaccine 1999;17(Suppl):S42–6.
[66] Shortridge KF. The next pandemic influenza virus. Lancet1995;346:1210–2.
[67] Kavet J. A perspective on the significance of pandemic influenza.Am J Publ Health 1977;66:1063–70.
[68] Francis Jr T. Influenza. In: Rivers TM, Horsfall Jr FL, editors. Viraland rickettsial infections of man. 3rd ed. Philadelphia: LippincottWilliams and Wilkins, 1959. p. 633–72.
[69] Francis Jr T. Vaccination against influenza. Bull Who 1953;8:725–41.
[70] Sartwell PE, Long AP. The army experience with influenza1946–1947. Part I. Epidemiological aspects. Am J Hyg 1948;47:135–41.
[71] Rasmussen Jr AF, Stokes JC, Smadel JE. The army experiencewith influenza 1946–1947. Part II. Laboratory aspects. Am J Hyg1948;47:142–9.
[72] Smith W, Andrewes CH. Serological races of influenza virus. BritJ Exp Pathol 1938;19:293–314.
[73] Hirst GK. Comparison of influenza virus strains from threeepidemics. J Exp Med 1947;86:367–81.
[74] Magill TP, Francis Jr T. Antigenic differences in strains of epidemicinfluenza virus. Part I. Cross-neutralization tests in mice. Brit J ExpPathol 1938;19:273–84.
[75] Hilleman MR, Mason RP, Rogers NG. Laboratory studies on the1950 outbreak of influenza. Publ Health Rep 1950;65:771–7.
[76] Hilleman MR, Mason RP, Buescher EL. Antigenic pattern of strainsof influenza A and B. Proc Soc Exp Biol Med 1950;75:829–35.
[77] Webster RG, Laver WG, Air GM. Antigenic variation amonginfluenza type A influenza viruses. In: Palese P, Kingsbury DW,editors. Genetics of influenza viruses. New York: Springer, 1983.p. 127–68.
[78] Hilleman MR. System for measuring and designating antigeniccomponents of influenza viruses with analysis of recently isolatedstrains. Proc Soc Exp Biol Med 1951;78:208–15.
[79] Hilleman MR, Werner JH, Gauld RL. Influenza antibodies in thepopulation of the USA. Bull Who 1953;8:613–31.
[80] Hilleman MR. Personal historical chronicle of six decades of basicand applied research in virology, immunology, and vaccinology.Immunol Rev 1999;170:7–27.

3086 M.R. Hilleman / Vaccine 20 (2002) 3068–3087
[81] Hilleman MR. Asian influenza: initial identification of Asiatic virusand antibody response in volunteers to vaccination. In: Proceedingsof the Special Conference on Influenza. US Department of HealthEducation and Welfare, 1957. p. 17–24.
[82] Meyer Jr HM, Hilleman MR, Miesse ML, Crawford IP, BankheadAS. New antigenic variant in Far East influenza epidemic. ProcSoc Exp Biol Med 1957;95:609–16.
[83] Wood JM. Developing vaccines against pandemic influenza. PhilTrans R Soc Lond B 2001;356:1953–60.
[84] Hilleman MR, Flatley FJ, Anderson SA, Leuking ML. Antibodyresponse in volunteers to Asian influenza vaccine. J Am Med Assoc1958;166:1134–40.
[85] Langmuir AD, Pizzi M, Trotter WY, Dunn F. Asian influenzasurveillance. Publ Health Rep 1958;73:114–20.
[86] Hilleman MR. Overview of vaccinology in historic and futureperspective. The whence and whither of a dynamic sciencewith complex dimensions. In: Ertl HCJ, editor. DNA vaccines.Georgetown (TX): Landes Bioscience, 2001, p. 1–40 [in press].
[87] Hampson AW, Cox NJ. Global surveillance for pandemic influenza:are we prepared. In: Brown LE, Hampson AW, Webster RD, editors.Options for the control of influenza. Amsterdam: Elsevier, 1996.p. 50–9.
[88] Lavanchy D. The importance of global surveillance of influenza.Vaccine 1999;17(Suppl):S24–5.
[89] Bush V. Science, the endless frontier. A report to the President ona Program for Post-war Scientific Research, July 1945. Reprintedby the National Science Foundation. Washington, DC, 1990.
[90] Hilleman MR. The business of science and the science of businessin the quest for an AIDS vaccine. Vaccine 1997;17:1211–22.
[91] Sporn MB. The war on cancer. Lancet 1996;347:1377–81.[92] Hoy AJ, Gregory V, Douglas AR, Lin YP. The evolution of human
influenza viruses. Phil Trans R Soc Lond B 2001;356:1861–70.[93] Hilleman MR. Cooperation between government and industry
in combating a perceived emerging pandemic. The 1976 swineinfluenza vaccination program. J Am Med Assoc 1996;275:241–3.
[94] Shortridge KF. Poultry and the H5N1 outbreak in HongKong, 1997. Abridged chronology and virus isolation. Vaccine1999;17(Suppl):S26–9.
[95] Claas EC, Osterhaus ADME, van Beek R, DeJong JC, RimelzwaanGF, Senne DA, et al. Human influenza A H5N1 virus related to ahighly pathogenic avian influenza virus. Lancet 1996;351:472–7.
[96] Subbarao K, Klimov A, Katz J, Regenery H, Lim W, Hall H, et al.Characterization of an avian influenza A (H5N1) virus isolated froma child with a fatal respiratory illness. Science 1998;279:393–6.
[97] Peiris M, Yuen KY, Leung CW, Chan KH, Ip PLS, Lai RWM.Human infection with influenza H9N2. Lancet 1999;354:916–7.
[98] Guan YI, Shortridge KF, Krauss S, Webster RG. Molecularcharacterization of H9N2 influenza viruses: were they the donorsof the internal genes of H5N1 viruses in Hong Kong. Proc NatlAcad Sci USA 1999;96:9363–7.
[99] Uyeki TM, Chong YH, Katz JM, Lim W, Ho YY, Wang SS, etal. Lack of evidence for human-to-human transmission of avianinfluenza A (H9N2) viruses in Hong Kong, China, 1999. EmergInfect Dis 2002;8:154–9.
[100] Kolata G. Flu: the story of the great influenza pandemic of 1918and the search for the virus that caused it. New York: Farrar Strausand Giroux, 1999. 330 p.
[101] Taubenberger JK. Sequencing influenza A from the 1918 pandemic,investigating its virulence, and averting future outbreaks. In: LayneSP, Beugelsdijk TJ, Patel CKN, editors. Firepower in the lab.Washington (DC): Joseph Henry Press, 2001. p. 123–30 [Chapter11].
[102] Webster RG. Spanish influenza: the secrets remain. Proc Natl AcadSci USA 1999;96:1164–6.
[103] Webster RG. A molecular whodunit. Science 2001;293:1774–5.[104] Gibbs MJ, Armstrong JS, Gibbs AJ. Recombination in the
hemagglutinin gene of the 1918 Spanish flu. Science 2001;293:1842–5.
[105] Kilbourne ED. A race with evolution—a history of influenzavaccines. In: Plotkin S, Fantini B, editors. Vaccine, vaccination andvaccinology: Jenner, Pasteur and their successors. Paris: Elsevier,1996. p. 183–8.
[106] Burnet FM. Growth of influenza virus in the allantoic cavity of thechick embryo. Aust J Exp Biol Med Sci 1941;19:291–5.
[107] Stanley WM. The preparation and properties of influenza virusvaccine concentrated and purified by differential centrifugation. JExp Med 1945;81:193–211.
[108] Reimer CB, Baker RS, van Frank RM, Newlin TE, Cline GB,Anderson NG. Purification of large quantities influenza virus bydensity gradient centrifugation. J Virol 1967;1:1207–16.
[109] Poland GA, Rottinghaus ST, Jacobson RM. Influenza vaccines:a review and rationale for use in developed and underdevelopedcountries. Vaccine 2001;19:2216–20.
[110] Halperin SA, Smith B, Mabrouk T, Germain M, Trépanier P, HassellT, et al. Safety and immunogenicity of a trivalent, inactivated,mammalian cell-culture derived influenza vaccine in healthy adults,seniors, and children. Vaccine 2002;20:1240–7.
[111] Cox N. Expanding the worldwide influenza surveillance systemand improving the selection of strains for vaccines. In: Layne SP,Beugelsijk TJ, Patel CKN, editors. Firepower in the lab. Washington(DC): Joseph Henry Press, 2001. p. 47–53.
[112] Kilbourne ED, Arden NH. Inactivated influenza vaccines. In: PlotkinSA, Orenstein WA, editors. Vaccines. Philadelphia: Saunders, 1999.p. 531–51.
[113] Hilleman MR. Serologic responses to split and whole swineinfluenza virus vaccines in the light of the next influenza pandemic.J Infect Dis 1977;136(Suppl):S683–5.
[114] Parkman PD, Hopps HE, Rastogi SC, Meyer Jr H. Summary ofclinical trials of influenza virus vaccines in adults. J Infect Dis1977;136(Suppl):S722–30.
[115] Barry DW, Mayner RE, Staton E, Dunlap R, Rastogi JE, HannahJE, et al. Comparative trial of influenza vaccines in man. Am JEpidem 1976;104:34–46.
[116] Zebedee FL, Lamb RA. Influenza A virus M2 protein: monoclonalantibody restriction of virus growth and detection of M2 in virions.J Virol 1988;62:2762–72.
[117] Treanor JJ, Tierney EL, Zebedee SL, Lamb RL, Murphy BR.Passively transferred monoclonal antibody to the M2 protein inhibitsinfluenza A virus replication in mice. J Virol 1990;64:1375–7.
[118] Neirynck S, Deroo T, Saelens X, Vanlandschoon P, Jou WM, FiersW. A universal influenza vaccine based on the extracellular domainof the M2 protein. Nat Med 1999;5:1157–63.
[119] Wolff JA, Malone RW, Williams P, Chang W, Acsadi G, Jani A,et al. Direct gene transfer into mouse muscle, in vivo. Science1990;247:1465–8.
[120] Donnelly JJ, Ulmer JB, Shiver JW, Liu MA. DNA vaccines. AnnuRev Immunol 1997;15:617–48.
[121] Webster RG, Robinson HL. DNA vaccines: a review ofdevelopments. Biol Drugs 1997;8:273–92.
[122] Schultz J, Dollenmaier G, Mölling K. Update on antiviral DNAvaccine research (1998–2000). Intervirology 2000;43:197–217.
[123] Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, DwarkiVJ. Heterologous protection against influenza by injection of DNAencoding a viral protein. Science 1993;259:1745–9.
[124] Okuda K, Ihata A, Watabe S, Okada E, Yamakawa T, HamasimaK, et al. Protective immunity against influenza A virus inducedby immunization with DNA plasmid containing influenza M gene.Vaccine 2001;19:3681–91.
[125] Kendal AP. Cold-adapted live attenuated influenza vaccinesdeveloped in Russia: can they contribute to meeting the needs forinfluenza control in other countries. Eur J Epidemiol 1997;13:591–609.
[126] Edwards KM, Dupont WD, Westrich MK, Plummer Jr WD, PalmerPS, Wright PF. A randomized controlled trial of cold-adapted andinactivated vaccines for the prevention of influenza A disease. JInfect Dis 1994;169:68–76.

M.R. Hilleman / Vaccine 20 (2002) 3068–3087 3087
[127] Belshe RB, Mendelman PM, Treanor J, King JD, Gruber WC,Piedra P, et al. The efficacy of live attenuated cold-adapted trivalent,intranasal influenza virus vaccine in children. N Engl J Med1998;338:1405–12.
[128] Mendelman PM, Cordova J, Cho I. Safety, efficacy and effectivenessof the influenza virus vaccine, trivalent, types A and B, types Aand B, live, cold-adapted (CAIV-T) in healthy children and healthyadults. Vaccine 2001;19:2221–6.
[129] Nichol KL. Live attenuated virus vaccines: new options for theprevention of influenza. Vaccine 2001;19:4373–7.
[130] Pfleiderer M, Löwer J, Kurth R. Cold-attenuated live influenzavaccines, a risk-benefit assessment. Vaccine 2002;20:886–94.
[131] Youngner JS, Treanor JJ, Betts RF, Whitaker-Dowling P. Effect ofsimultaneous administration of cold-adapted and wild-type influenzainfections in humans. J Clin Microbiol 1994;32:750–4.
[132] Beyer WEP, Palache AM, de Jong JC, Osterhaus ADME.Cold-adapted live influenza vaccine versus inactivated vaccine:systemic vaccine reactions, local and systemic antibody response,and vaccine efficacy. A meta analysis. Vaccine 2002;20:1340–53.
[133] World Health Organization. Influenza Pandemic Plan. The role ofWHO and Guidelines for National and Regional Planning. Geneva,April 1999 (http://www.who.int/emc).
[134] Gust ID, Hampson AW, Lavanchy D. Planning for the next pandemicof influenza. Rev Med Virol 2001;11:59–70.
[135] Pennisi E. Planning for the next flu pandemic. Science1995;270:1916–7.
[136] Marwick C. Readiness is all: public health experts draftplan outlining pandemic influenza response. J Am Med Assoc1996;275:179–80.
[137] Klimov A, Simonsen L, Fukuda K, Cox NJ. Surveillance and impactof influenza in the United States. Vaccine 1999;17(Suppl):S42–6.
[138] Patriarca PA, Cox NJ. Influenza pandemic preparedness plan forthe United States. J Infect Dis 1997;176(Suppl):S4–7.
[139] Jack DB. Getting ready for the next influenza pandemic. Lancet1996;347:1252.
[140] Fitch WM, Bush RM, Bender CA, Cox N. Long-term trends in theevolution of the H (3) HA1 human influenza type A. Proc NatlAcad Sci USA 1997;94:7712–8.
[141] Zhou NN, Senne DA, Landgraf JS, Swenson SL, Erickson G,Rossow K. Emergence of H3N2 reassortant influenza A viruses inNorth American pigs. Vet Microbiol 2000;74:47–58.
[142] Rota PA, Rocha EP, Harmon MW, Hinshaw VS, Sheerar MG,Kawaoka Y, et al. Laboratory characterization of a swine influenzavirus isolated from a fatal case of human influenza. J Clin Microbiol1989;27:1413–6.
[143] Cohen J. The flu pandemic that might have been. Science1997;277:1600–1.
[144] Oxford JS, Daniels RS. The holy grail of influenza virologists. NatMed 1999;5:484–5.
[145] Layne SP, Beugelsdijk TJ, Patel CKN, editors. Firepower in the lab:automation in the fight against infectious diseases and bioterrorism.Washington (DC): Joseph Henry Press, 2001. 299 p.
[146] Layne SP, Beugelsdijk TJ, Patel CKN, Taubenberger JK, CoxNJ, Gust ID. A global lab against influenza. Science 2001;293:1729.
[147] Abbott A. The flu HQ. Nature 2001;414:10–1.[148] Gubareva LV, Kaiser L, Hayden FG. Influenza virus neuraminidase
inhibitors. Lancet 2000;355:827–35.[149] Hayden FG. Perspectives on antiviral use during pandemic influenza.
Phil Trans R Soc Lond B 2001;353:1877–84.[150] Roberts N. Treatment of influenza with neuraminidase inhibitors:
virological implications. Phil Trans R Soc Lond B 2001;356:1895–7.
[151] Fleming DM. Influenza diagnosis and treatment: a view fromclinical practice. Phil Trans R Soc Lond B 2001;356:1933–43.
[152] Haseltine WA. Chicken Soup. Sci Am 2001;285:56–63.[153] Goldfield M, Bartley JD, Pizzuti W, Black HC, Altman R, Halperin
WE. Influenza in New Jersey in 1976: isolations of influenza A/NewJersey/76 virus at Fort Dix. J Infect Dis 1977;136(Suppl):S347–55.
[154] Hahn AF. Guillain-Barré syndrome. Lancet 1998;352:635–41.[155] Langmuir AD, Bregman DJ, Kurland LT, Nathanson N, Victor
M. An epidemiologic and clinical evaluation of Guillain-Barrésyndrome reported in association with the administration of swineinfluenza vaccines. Am J Epidem 1984;119:841–79.
[156] deJong JC, Heinen PP, Loeffen WLA, van Nieuwstadt AP,Claas ECJ, Bestebroer TM, et al. Antigenic and molecularheterogeneity in recent swine influenza A (H1N1) virus isolates withpossible implications for vaccination policy. Vaccine 2001;19:4452–64.
[157] Hirst GK. Adsorption of influenza hemagglutinins and virus by redblood cells. J Exp Med 1942;76:195–209.
Copyright © 2022 FDOKUMEN




















