Protein-DNA Interactions Determine the Shapes of DNA Toroids Condensed in Virus Capsids
15
Title page Protein-DNA interactions determine the shapes of DNA toroids condensed in virus capsids Authors: Amélie Leforestier * , Antonio Šiber † , Françoise Livolant * and Rudolf Podgornik ‡,§ * Laboratoire de Physique des Solides, Centre National de la Recherche Scientifique UMR 8502, Université Paris-Sud, Bâtiment 510, 91405 Orsay Cedex, France † Institute of Physics, Bijenička cesta 46, 10000 Zagreb, Croatia ‡ Theoretical Physics Department, J. Stefan Institute, SI-1000 Ljubljana, Slovenia § Institute of Biophysics, School of Medicine and Department of Physics, Faculty of Mathematics and Physics, University of Ljubljana, SI-1000 Ljubljana, Slovenia Keywords: DNA elasticity, DNA free energy, DNA condensation, viral encapsidation
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Protein-DNA Interactions Determine the Shapes of DNA Toroids Condensed in Virus Capsids

Title page Protein-DNA interactions determine the shapes of DNA toroids condensed in virus capsids
Authors:
Amélie Leforestier*, Antonio Šiber†, Franc oise Livolant* and Rudolf Podgornik‡,§
*Laboratoire de Physique des Solides, Centre National de la Recherche Scientifique UMR 8502, Université Paris-Sud, Ba timent 510, 91405 Orsay Cedex, France
†Institute of Physics, Bijenic ka cesta 46, 10000 Zagreb, Croatia
‡ Theoretical Physics Department, J. Stefan Institute, SI-1000 Ljubljana, Slovenia §Institute of Biophysics, School of Medicine and Department of Physics, Faculty of Mathematics and Physics, University of Ljubljana, SI-1000 Ljubljana, Slovenia
Keywords: DNA elasticity, DNA free energy, DNA condensation, viral encapsidation

Abstract page DNA toroids formed inside the bacteriophage capsid present different shapes according to whether they are formed by addition of spermine or PEG to the bathing solution. Spermine-DNA toroids present a convex facetted section with no or minor distortions of the DNA interstrand spacing with respect to those observed in the bulk, whereas PEG-induced toroids are flattened to the capsid inner surface and show a crescent-like, non-convex shape. By modelling the energetics of the DNA toroid using a free energy functional composed of energy contributions related to the elasticity of the wound DNA, exposed surface DNA energy and adhesion between the DNA and the capsid, we established that the crescent-shape of the toroidal DNA section comes from attractive DNA interactions between DNA and the capsid. Such attractive interactions seem specific to the PEG condensation process and they are not observed in the case of spermine-induced DNA condensation.

Introduction
The control of DNA condensation plays a crucial role in both bacteria as well as eukaryotic cells at various stages of their cell cycle (1). Similarly DNA is tightly packed also inside viral capsids (2) with DNA densities almost approaching those of close packing. Because of the difficulties in investigation of the structure of these dense and complex DNA arrays without destroying them, and also with the hope to unravel the mechanisms governing the condensation and decondensation processes at work in the context of living cells, much interest was generated for the understanding of simplified systems where all parameters can be precisely controlled. Indeed, DNA condensation can be reproduced in vitro and DNA has been observed to undergo a drastic volume change on addition of various condensing agents such as polyvalent cations - polyamines (spermidine and spermine), Co(NH3)6
3+, polylysine, histone H1 - but even in the presence of monovalent cations with crowding agents such as poly-ethyleneglycol (PEG) (3).
A most interesting case of DNA condensation is presented by the DNA toroid formation that can be observed under specific conditions, namely very low DNA concentrations. Various images of toroids formed by multivalent cations and basic proteins can be found in the literature and their characteristics and dimensions have been extensively discussed (3). By comparison, only a few micrographs of individual DNA chains collapsed by PEG have been presented (4), probably because of the preferred spherical or ellipsoidal conformation of the PEG-induced DNA globules (5). Ever since it has been observed, the regular and highly ordered form of the toroidal condensate has been attracting the interest of theorists (6,7). The new cryoEM methods renewed this interest by enabling a detailed quantitative analysis of their internal structure and explicit visualization of the local DNA ordering inside the toroid (3).
The formation of toroidal globules of single DNA chains trapped inside a confining volume has been reported recently, where the confining space is provided by a nanochannel (8), a liquid droplet (9), a liposome (10) or a virus capsid itself (11, 12, 13), either upon addition of spermine (Spm4+) or in a monovalent salt buffer containing PEG. These experiments raise additional theoretical and biological questions. Indeed DNA is always confined in vivo, whatever the system under consideration: each DNA chain with its associated proteins forming chromatin is confined in a chromosomal territory inside the nucleus of the eukaryotic cell (14), itself limited by the nuclear envelope; the bacterial nucleoid is confined in the central volume of the bacteria by the macromolecular crowding effect arising from other macromolecules (15) and the viral genome is confined within the protein capsid (16). This confinement is particularly important in this latter case because the capsid is rigid and entraps the entire genome under a very dense and still unknown conformation. It has become clear recently that after a partial ejection of the viral DNA, the fragment of the chain that is kept inside can be collapsed into a toroid (11, 12, 13). This transition between a confined coil and a confined toroid, triggered by changes in the environment of the virus, raises the interest to understand how the capsid can influence the organization of the hedged in DNA chain.
In what follows we focus on the formation of toroidal condensates of a single DNA chain inside an intact viral capsid, with corresponding DNA densities comparable to previous observations. We compare toroids formed by two different mechanisms: the addition of Spm4+ or the addition of PEG to the outside bathing solution of the capsids. The toroidal DNA condensate can furthermore interact specifically with the confining capsid wall to the extent that its shape can be substantially modified by this interaction. We observe this kind of strong DNA - capsid wall interaction only on addition of PEG, where the shape of the toroid is drastically modified. Compared to this case the addition of spermine shows no modification in the shape of the DNA, indicating that there is only a weak interaction of the DNA toroid with the inner capsid wall.
Constrained DNA toroid formation inside viral capsids
The T5 is a tailed bacteriophage composed of a protein capsid hedging in the DNA genome, and a long tail through which the double stranded DNA (dsDNA) chain is ejected upon interaction with the phage receptor inserted in the external membrane of the host bacteria, E. coli. The portal complex of

the bacteriophage is located at one apex of the icosahedral capsid and connects the capsid to the tail. Using protocols detailed in Material and Methods, we prepared viruses whose capsids contain a unique fragment of their initial genome of length 113.9 kbp i.e. 38.7 µm. The length of the fragment varies from capsid to capsid and can only be determined a posteriori.
In Figure 1 we present a selection of capsids containing a DNA fragment about 50-55,000 bp long (contour length ∼ 17µm). Depending on the conditions of preparation of the capsids, the DNA fragments may either occupy the entire volume of the capsid (Figure 1a-a’) or be condensed into toroidal globules (Figure 1b-b”, c-c”).
Figure 1a’ shows a capsid immersed in a classical buffer (10mM Tris-Cl- containing monovalent salt, 100mM NaCl, with traces of divalent cations required to maintain the integrity of the protein capsid, 1mM MgCl2 and 1mM CaCl2). The DNA fragment obviously occupies the total volume of the capsid. From the estimated interhelical distance (∼ 3.9 nm), we calculate the DNA concentration that falls within the range of the bulk cholesteric phase. Since the length of the encapsidated DNA can not be controlled, it has to be determined from the value of the interaxial spacing and the volume of the capsid itself. With the resolution of the cryoEM, we do not observe any specific interaction with the protein capsid under these particular conditions: neither preferential adsorption to the surface, nor any significant exclusion from peripheral volume close to the inner capsid wall. In fact DNA appears to be uniformly distributed within the whole capsid volume available without any detectable density variation (17). Concomitantly we also never observe a pronounced depletion region with axial symmetry in the DNA density, that is sometimes predicted to exist on theoretical grounds (18). In fact, one can actually construct a robust theoretical argument that the axial depletion region should be in fact quite small (19) and may thus not be detectable by the cryo-EM method.
Figure 1.
Toroids presented in Figure 1b-b” and c-c” are formed by collapsing a similar DNA fragment by adding to the bathing solution of viruses either a tetravalent cation, spermine (Spm4+) (Figure 1b-b”) or a neutral polymer, polyethyleneglycol (Figure 1c-c”), while keeping all other solution conditions fixed. Details of the protocols are given in the Material and Methods section.
When polyvalent Spm4+ cations, which are small enough and can diffuse through the capsid wall, are added to the solution, they trigger the DNA coil – toroidal globule transition. As detailed earlier, DNA is hexagonally packed in the toroid and the interhelical distances (aH) is fixed by the balance of forces conferred by multivalent and monovalent cations (20). The measured value of the interaxial spacing under these conditions is aH= 2.88±0.12 nm. The attractive component to the total force is provided by the polycations, whereas the monovalent counterions engender repulsive interactions. The balance of attraction and repulsion gives rise to a van der Waals – like behavior of the osmotic pressure (21,22) that at high enough polyvalent cation concentration leads to a minimum and an onset of condensation. The perpendicular section of the toroidal DNA globule often shows pronounced faceting that follows the reticular planes of the hexagonal lattice. We noted previously how one of the reticular planes usually aligns itself parallel to the faces of the capsid (also seen on Figure 1b”). The external diameter of the toroid may be smaller than the diameter of the capsid (13) but the toroid nevertheless always sticks to one of the inner wall capsid faces.
On the other hand, the formation of the DNA toroidal condensate presented in Figure 1c’-c”, is due to the presence of PEG outside the capsid (23). PEG is well known to induce condensation of DNA in the presence of monovalent salts. This phenomenon is referred as PSI-DNA-condensation (24, 25). The protein capsid behaves here as a semi-permeable membrane that allows exchanges of water and ions from one side to the other, while keeping PEG and DNA separated. Compared to a standard dialysis membrane, the protein capsid is different in the sense that it is rigid and does not deform under the osmotic pressure of PEG at concentrations investigated. Indeed, the volume of the capsid stays roughly constant (within the resolution of our observations). The perpendicular section of the toroid in this case deviates significantly from a circle, and adopts a crescent-like shape with a markedly concave inner surface. This deformation of the cross-section of the DNA condensate obviously increases the

surface of interaction between DNA and the internal surface wall of the capsid. The local DNA packing is less regular in this case and shows an interaxial spacing of aH=3±0.2 nm. At concentrations used in the experiments (a 15% solution of PEG 6000), polyethyleneglycol exerts an osmotic pressure of e.g. 3.2 atm (26). The capsids can withstand such an osmotic pressure without any trouble. In fact osmotically induced elastic collapse of viral shells, recently observed in the analogous case of frozen cationic lipid vesicles (27), happens at larger values of osmotic pressure, typically above 10 atm, in the case of empty viral shells (28) and for even higher values for DNA filled capsids. These estimates, however, depend on the exact values of elastic parameters characterizing the capsid shell.
The experimentally observed shapes of the DNA condensates show variation in the case of Spm4+ as well as PEG condensation. One could argue that the temperature quench used in cryomicroscopy should show an equilibrium distribution of shapes corresponding to the free energies of the different shapes. If there would be a well defined ground-state of minimal energy, separated from other shapes by a finite energy gap, one could expect that this ground-state shape would dominate the observed shape space spectrum. The broadness of this spectrum on the other hand, would indicate that relatively different shapes share similar free energies and should thus show up in the quenched distribution provided by cryomicroscopy. The experiments typically do not show non-crescent like shapes in the case of PEG condensation and thus our singling out of this shape as a consequence of the pronounced attractive DNA condensate –capsid wall interactions seems to carry some weight.
We want to make clear that here we do not study how the presence of PEG outside the capsid interacts with the ejection mechanism itself. We focus on capsids containing a fragment of their initial DNA genome mimicking capsids at an intermediate state of DNA encapsidation or ejection. PEG has indeed been used to oppose the ejection of DNA from the capsid of Lambda, T5 and Spp1 bacteriophages. Contrary to Lambda (29) and Spp1 (30), where the fraction of ejected DNA is directly related to the applied pressure, in T5, the remaining encapsidated length of DNA cannot be controlled by varying the external osmotic pressure (31). The mechanism is more tricky and not fully elucidated, and for each applied pressure capsids are found, containing varying amounts of encapsidated DNA. Uncompletely filled capsids can also be prepared without any osmotic pressure opposing the ejection, just by stopping the ejection before its completion (17).
Theory of a DNA toroid constrained in a sticky sphere
The experiments investigate a very complicated situation with myriad of interactions present. It is almost hopeless and probably not very useful to approach the problem by microscopic representation of all the involved entities and interactions. We thus simplify the situation by modelling the condensed DNA toroid using a continuum theory that accounts for the elastic energy of the wound-up DNA ( ), the energy of the exposed surface of the toroid ( , is the effective surface tension of the unconstrained DNA toroid and is its surface area) and an attractive adhesion energy of the toroid-capsid interaction. The free energy functional of our model is
(1)
where the first two terms, the surface free energy of the toroid, and the elastic energy of the wound-up DNA, are the same as in the model proposed by Ubbink and Odijk (UO, 32). The third term complicates the model and makes it less tractable due to the introduction of surface energy parameter,
, that depends on the strength of the effective short-range DNA-capsid interaction. The area of the contact region between the DNA condensate and the inside capsid wall is denoted by . This is an unknown quantity whose value is obtained only after the minimization of the functional. We consider the free energy functional on a hyperplane of constant DNA volume, , the same as in the UO case. In our case, we additionally restrict the problem by the introduction of the capsid which acts as a constraint, allowing only solutions contained within the sphere of radius .

In UO model, the solution for the shape of the DNA toroid depends only on two parameters: the volume and an adimensional parameter αt that weights the relative importance of surface and elastic energies
(2)
where and are the contour and persistence lengths of the DNA, respectively, while l was defined above in terms of V. In our case, the solution in addition depends on ratios and . When
, corresponding to an unconstrained torus inside an infinitely large capsid, and , our model reduces to the one studied by UO, and we have checked that this is indeed so by examining our results in this limit. The experiments restrict the freedom of our theoretical model since they fix the constraint (capsid) radius and the DNA volume. The DNA volume and the radius of the capsid interior can be obtained from experimental images of phages (see Figure 1). In this way we determine
nm3 (with interhelical distance equal to 3 nm and base-pair separation of 0.34 nm, this corresponds to 53 kbp) and nm.
Though faceting in the perpendicular cross-section of the DNA condensate can be sometimes observed in our experiments, the continuum model of UO (33) does not take into account the discrete nature of DNA lattice within the condensate, and thus can not account for discrete features such as toroid faceting. Formally the faceting is an effect of the orientational dependence of the surface energy, σ in our case, which we assume to be a constant, and can be obtained via a Wulff construction as is the case in the theory of equilibrium shapes of crystals (34). In a similar vein we also do not take into account the polyhedral structure of the capsid wall and even less its discrete protein architecture, but treat it as being ideally and uniformly spherical. The inner polyhedral faces can in fact promote a preferred interaction between the facets of the DNA condensate and the centers of the polyhedral faces leading to preferred connected interaction patches along the inner capsid surface.
Numerical analysis of the model
To compare with experiments, we examine the model on a remaining, two-dimensional plane of (unknown) parameters, and . The toroid shapes are fully determined by their cross-sections. These were first represented in a discrete form, i.e. by points, where is typically of the order of 500. Only half of the cross-section needs to be represented, the other half can be obtained by reflection. The free energy functional is then represented in terms of the coordinates of these points, so that it becomes a function of variables. This function is numerically minimized using a suitable variant of the conjugate gradient optimization. The constraints of the fixed volume and the impenetrable capsid wall were implemented via energy-penalty contributions to the functional. The inclusion of the short-range attraction in the functional requires a specification of the critical separation between the toroid patch and the surface. Below the critical separation the toroid patch is considered to be in contact with the capsid and only then its area contributes to the total area of the contact region, . The critical separation should be made as small as possible so as to be by far the smallest length in the model and that the solutions do not depend on its exact choice.
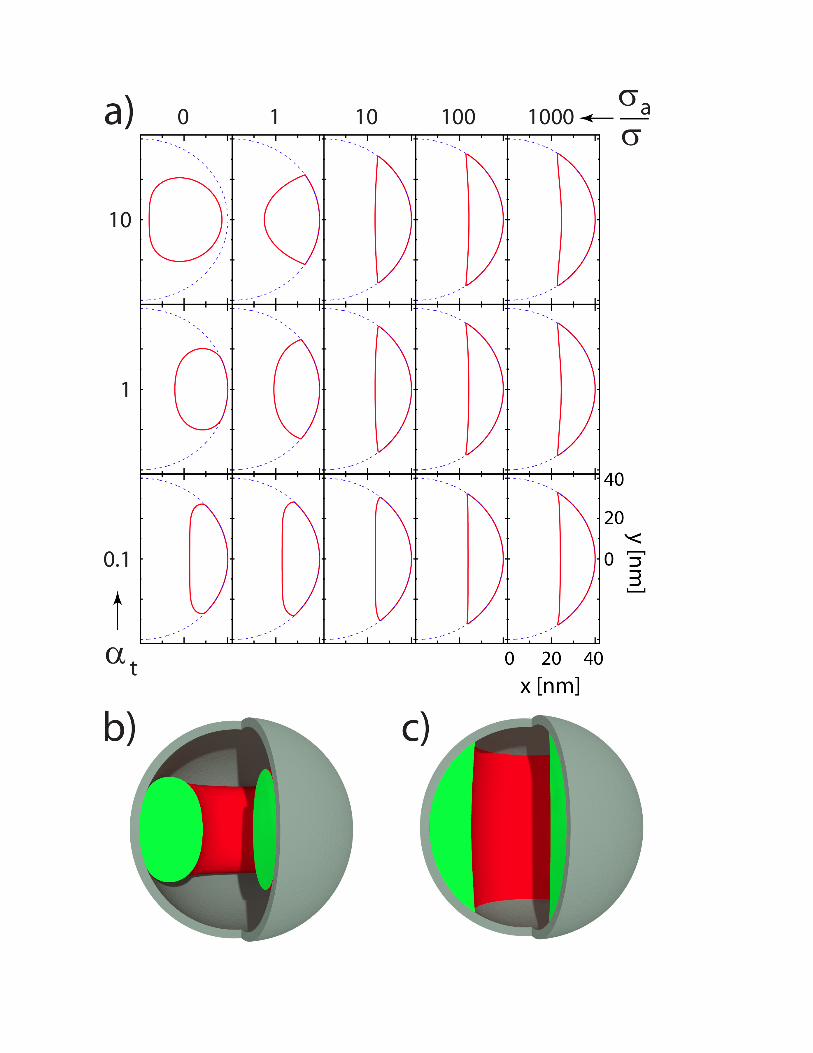
Figure 2.
A shape phase-diagram of the problem representing the optimal toroid cross-sections for particular combination of and parameters is shown in Figure 2. Columns of cross-sections corresponding to case, i.e. without attraction between the DNA and the capsid, are presented in Figure 2a). Note that the top-left shape ( and ) is the same as would be obtained in the UO model since the toroid does not touch the capsid in this case, i.e. the capsid impenetrability constraint is not active (35). For sufficiently large adhesion energy parameter (last column of images in

Figure 2a), the cross-section of the DNA toroid only weakly depends on . Obviously the presence of attractive interactions between the capsid wall and the toroidal condensate leads to a deformation of its cross-section that becomes markedly concave, crescent-like, Figure 2c). By analyzing numerical solutions for very small values of the parameter, without any attraction between the capsid wall and the DNA toroid ( ), one finds that the limiting shape for small values of is a plan-convex lens. This leads to the conclusion that the concavity of the inner surface of the DNA toroid section can be observed only when the attraction between the DNA and the capsid is present. Otherwise the cross-sections are always flat or convex i.e. the shape constraint in combination with elasticity and the surface energy minimization cannot produce a concave shape. The crescent-like form of the condensate cross-section thus appears to be the distinguishing feature of the attractive interaction between the DNA condensate and the capsid wall.
The experiment shows two markedly different types of toroid shapes (see Figure 1). In numerical calculations the characteristic crescent-like shapes of the cross-sections with concave inner surface (see Figure 2), similar to those found in experiments with PEG, are obtained only in the case of a sufficiently strong attractive interaction between the DNA condensate and the inner capsid wall as codified by the parameter . These distinguishing features are certainly observed in the case of PEG condensation but are absent in the case of Spm4+ condensation. In that case one can identify more circle-like cross-sections consistent with model calculations for and small enough .
The comparison of the model with the experiments thus suggests quite different energetics of the system in cases when the solution contains PEG and Spm4+. Spm4+ ions apparently condense DNA and do not induce significant DNA-capsid attraction, while PEG molecules do, although the capsid is impermeable to them.
Discussion
There are a few features of our result on DNA condensation in capsidā that we need to put in perspective.
It follows from our observations and the corresponding model analysis that the strength of DNA toroid – capsid wall interactions is much larger in the case of PEG condensation than in the case of Spm4+ condensation. In the latter case there is a tendency that the (faceted) condensate as a whole will interact with the capsid surface, or more correctly with its polyhedral faces, but this surface interaction does not appear to be large enough to deform the shape of the condensate. This interaction can become substantial when the toroid outer radius is larger than the inner radius of the capsid. In that case the confinement interaction of the DNA toroid forced to reside inside the capsid dominates (36), but the confinement alone, without any attractive interactions between the DNA condensate and the capsid wall, can only modify the shape of the toroidal condensate in such a way that the inner surface of the condensate becomes and remains flat (in the limit when -> 0), while the outer surface is pushed against the inner surface of the capsid. Thus confinement in and of itself can not simulate attractions between the DNA condensate and the capsid wall.
The UO model used to analyze the possible shapes of a confined toroidal condensate, with or without surface specific interactions with the confining wall, is based on three effective parameters, viz. ,
and the effective radius of the capsid. All numerical results that we presented above in fact depend on the combination of these three parameters characterizing the model of the confined DNA toroidal condensate. Short range microscopic interactions between capsid wall protein moieties and DNA are clearly the dominant contribution to the ratio. At least partly, these interactions have to be electrostatic in nature, as both DNA as well as capsid proteins contain a lot of charges, but more specific protein binding to DNA can not be excluded. The parameter obviously depends on whatever interactions set the equilibrium spacing between DNA molecules in the condensate, elasticity of the molecule and its total length. The surface tension of the DNA toroid can also be modified by the condensing agent and probably is. The attractive branch of these interactions is generally assumed to be due at least in part to an electrostatic correlation effect (24) but should also

depend on specific properties of the polyvalent condensing agents (23) since even equally charged condensing cations do not induce DNA condensation to the same extent. The specific properties of ions differentiating different types of DNA condensing cations are relatively poorly known and still inadequately understood 37. The two parameters of the continuum model that guided our analysis of the PEG and Spm4+ induced DNA condensation in vitro can be estimated very roughly to be ~ 1 and ~ 0 for Spm4+ induced aggregation and concurrent shape of the confined DNA toroidal, while ~ 1 and ~ 100 would be more appropriate for the PEG induced aggregation.
What are the physical reasons for the fundamental difference between PEG and Spm4+ in the observed behavior of DNA condensation induced in capsidā? The experiment itself does not give a definitive answer to this question. Nevertheless we may try to patch together a string of arguments that might shed some light on this problem. PEG itself does not induce a direct interaction between DNA segments. In fact in DNA condensation experiments that are based on long fragments of DNA, DNA and PEG spontaneously demix. What PEG always induces is a modification in the chemical potential of water, implying that the properties of water inside the capsid are changed and that its chemical potential adjusts to the same new value as is set by the concentration of PEG in the bathing solution. Once the chemical potential of water inside the capsid is changed, DNA self-condenses in order to counter this new value of chemical potential or, what amounts to the same thing, the new value of osmotic pressure. That the state of DNA depends on the water chemical potential or its osmotic pressure was amply demonstrated in experiments of Rau, Parsegian and coworkers (38) where the DNA condensation was induced directly by changes in the PEG concentration in the bathing solution (39). The same is true here as the DNA interhelix distance goes down from about 3.9 nm to 3 nm after addition of PEG. The chemical potential of water in the PEG bathing solution thus sets the chemical potential of water inside the capsid and DNA reacts to this change by condensing. The PEG induced condensation appears quite different from the Spm4+ induced condensation, where the condensing agent gives rise to explicit attractive interactions between DNA molecules, that could be rationalized as either mediated by attractive electrostatic correlation interactions (40) or attractive hydration interactions (41), that lead to condensation. The main difference between the PEG-induced and the polyvalent counterion induced condensation of DNA would thus be the indirect vs. the direct routes to DNA condensation.
The main difference between our PEG experiments and the experiments of Rau, Parsegian et al. is that in our case, the semi-permeable capsid is rigid. The consequence of this rigidity is that the volume of the capsid is kept to a good approximation a constant. Therefore, after being collapsed, DNA occupies only a fraction of the capsid volume and two compartments are created inside the capsid: one containing DNA with its associated water and counterions and another one devoid of DNA. In their experiments, Rau, Parsegian et al. mostly used no semi-permeable membrane and therefore there were only two compartments: the DNA solution and the PEG solution. The existence of the third compartment could possibly be a consequence of the negative hydrostatic pressure inside the capsid caused by its impenetrability to the external PEG but we can not say anything more definitive at this point. We thus leave the PEG induced condensation in capsidā as a challenge for the theory.
Our results could also have broader ramifications when considered together with some other related experimental investigations. One may wonder whether the formation of toroids that has been only exceptionally reported in the bulk in the presence of crowding agents (see Ref. 4) may be facilitated by the confinement inside the capsid. Indeed, Zhang et al (42) recently observed how the confinement of the DNA chain in a nanochannel facilitates compaction with a neutral crowding agent at low ionic strength. The observation by Hou et al (43) that toroids can be formed inside confined droplets adsorbed on mica surfaces, in the absence of any condensing agent, just by geometry confinement, would further support such an hypothesis. Additionally, we may also wonder whether DNA condensation may be assisted and directed by the internal surface of the capsid itself, as is apparently the case for a mica surface (53,44) or on the phospholipid membrane of a giant vesicle (45).
The PEG, via the changes in water chemical potential or its osmotic pressure, could not only induce DNA aggregation but could also effect conformational changes in the proteins of the capsid wall. This assertion is of course speculative since we do not have any direct indications that it is true.

Nevertheless we know that PEG via its effect on the chemical potential of water can affect conformational changes in e.g. haemoglobin and various membrane (channel) proteins (46). Similarly it is not preposterous to hypothesize that PEG could affect also the conformation of capsid proteins which in its turn could lead to a more pronounced interaction with the DNA. This interaction can again be either very specific or quite generic i.e. electrostatic. It would be difficult to assert more at this point but maybe a separate investigation of the conformation of capsid proteins as a function of PEG osmotic pressure could settle some of the subtler points of our speculation.
In fact, we do not know exactly what is the distribution of charges on the protein surface interacting with DNA because the structure of the capsid has not been solved yet. Nevertheless, T5 belongs to the Siphoviridae family of viruses, like lambda or HK97. The structure of the capsid was elucidated at atomic resolution only for HK97 (47). The proteins pb8 (the main component of the T5 capsid) and gp5 (that constitutes the capsid of HK97) share sequence simarities and also the same fold (except from some loops involved in the HK97 interlinked rings - catenanes) (48). We can thus rely on HK97 to have an idea of the distribution of aminoacids (and charges) on the internal surface of the T5 capsid. A few positive charges could favour attractive interactions with the encapsidated DNA together with a comparable number of negative charges that may also constitute possible loci for electrostatic links to the negative DNA phosphate charges through the intermediate bridging of divalent cations that are present (and required) in the buffer.
Whatever the details of this PEG induced attractive interaction between DNA and the capsid, the fact that DNA in capsidā condenses onto the internal protein surface creates an empty space in the centre of the capsid. This effect should also exist in newly synthesized capsids inside the bacteria. The bacterial cytoplasm being highly crowded, should favour condensation of the entering DNA into the capsid (as does PEG in our experiments) and provide an empty space in the center of the capsid, thus supporting the encapsidation process and facilitate the work of the portal motor.
Material and methods (Experimental part; T5 in 15% PEG)
Biochemical purification. T5 st(0) bacteriophages were produced in E. coli, purified, and concentrated at 2.5 1012 or 5 1012 phages/ml in 10mM Tris-Cl-, pH 7.6, 100mM NaCl, 1mM CaCl2 and 1mM MgCl2. The protein receptor FhUA was purified as described in (Boulanger et al, 1996) and stored at 4°C in 25mM Tris-Cl, pH 8.0, 250mM NaCl and 1% octyl glucoside (w:v) at a concentration of 4mg/ml.
Partial ejection and DNA condensation. Capsids containing only part of their genome were obtained in two different ways: either by freezing the specimen before the entire DNA molecule is ejected (time resolved experiments, 17), or by applying an external osmotic pressure (31). The phage solution was mixed with an excess of FhuA (phage/protein ratio = 1/1000), in the absence or presence of an osmolyte (PEG MW 6000, Fluka), and immediately transferred to 37°C to trigger the ejection. 0.03% lauryldimethylamine N-oxide (LDAO) was present to ensure the stability and solubility of the receptor protein. DNAse I (Sigma) was added at a concentration of 0.1 U/µl to digest the ejected DNA. The ejection was let to proceed for 10 minutes (absence of PEG) or 3 hours (presence of PEG). Two alternative protocoles were used to condense the DNA segment remaining inside the capsid: i) the addition of spermine at a final concentration of 5 mM (followed, when needed, by dilution of the sample to decrease the PEG concentration below 1% while keeping constant the ionic conditions), ii) DNA ejection in the presence of 15% PEG 6000 followed by direct freezing of the sample in the PEG solution.
Cryo-EM. A drop of the phage solution was deposited on an EM grid covered with a holey carbon film (Quantifoil R2/2) previously treated by plasma glow discharge. Grids were blotted to remove the excess of material and immersed in liquid ethane cooled down by liquid nitrogen. All the process was carried out in a home-made plunging device to control the temperature and hygrometry, and prevent any dehydration or any change of the ionic concentrations of the sample. The preparation of thin films was more difficult in the presence of PEG that increases the viscosity of the solution and induces phage aggregation (visible at low magnification). The grids were transferred to a JEOL 2010 LaB6 or a

JEOL 2010-FEG TEM operated at 200kV. Images were recorded with a direct magnification of x50,000 under low dose conditions of imaging (10-20 e-/Å2). The defocus was set at 850nm to optimize the imaging of the DNA lattice spacing in the capsid.
Acknowledgments
R.P. acknowledges funds from the Agency for Research and Development of Slovenia (Grant P1- 0055(C)).
A.Š. acknowledges support by the Ministry of Science, Education, and Sports of Republic of Croatia (Project No. 035-0352828-2837).
We thank Gerard Pehau-Arnaudet (Institut Pasteur, Paris) for his support on the federative FEG microscope and Madalena Renouard for the biochemical purification of the protein receptor FhuA.
We also acknowledge M. Castelnovo who draw FL’s attention to the likely presence of toroids in Fig. 5 of ref 13.
Figure legends
Figure 1
T5 bacteriophage capsids containing a ds DNA fragment of about 17µm (50,000bp) after partial ejection of their genome. In the presence of monovalent cations (a,a’) the DNA chain occupies the entire volume of the capsid (a’). DNA toroids are formed by addition of polyamines (Spm4+) (b-b”)) or polyethylene glycol (c-c”) in the buffer. The polycations can go through the capsid whereas PEG stays outside. For each conditions, we present top views (b’,c’) and side views (b”, c”) of the DNA toroids. The section of the toroid is underlined on Fig. a” and b”. Scale bar= 20nm.
Figure 2
a) The shapes of DNA toroids in [ , ] parameter space. Blue dashed lines in a) indicate the inner surface of the spherical container with nm. The volume of the DNA in these calculations was fixed to nm3. b) Three-dimensional rendering of the DNA toroid with , c) Three-dimensional rendering of the DNA toroid with ,
References
1 Wolffe, A. P., Chromatin structure, Advances in Genome Biology 5, 363-414 (1998). 2 Cerritelli, M., Cheng, N., Rosenberg, A., McPherson, C., Booy, F. & Steven, A., Encapsidated Conformation of Bacteriophage T7 DNA, Cell 91, 271–280 (1997). 3 Hud, N.V. and I. D. Vilfan, Toroidal DNA Condensates: Unraveling the Fine Structure and the Role of Nucleation in Determining Size. Annu. Rev. Biophys. Biomol. Struct. 34, 295–318 (2005). 4 Evdokimov Yu.M., Platonov, A.L., Tikhonenko, A.S., Varshavsky, Ya.M. et al, A compact form of double stranded DNA in solution. FEBS Lett. 23, 180-184 (1972) 5 Laemmli, U.K., Characterization of DNA condensates induced by poly(ethylene oxide) and polylysine. Proc. Natl. Acad. Sci., 72, 4288-4292 (1975) 6 Grosberg, A.Yu.,and A.V. Zhestkov, On thecompactformoflinear duplexDNA: globular states of the uniform elastic (persistent) macro-molecule. J. Biomol. Struct. Dyn. 3, 859-872 (1986).

7 Ubbink, J. and Odijk T., Deformation of toroidal DNA condensates under surface stress, Europhys. Lett., 33, 353-358 (1996) 8 Zhang C., Shao, P.G., van Kan J.A., van der Maarel, R.C. Macromolecular crowding induced elongation and compaction in a naochannel. Biophys. J. 97, 1678-1686 (2009). 9 Hou X.-M., Li W., Dou S.-X., Zhang L.-Y., Xie P., Wang W.-C., Wang P.Y. Formation of DNA toroids Inside confined droplets adsorbed on mica surfaces. Phys. Rev. E 79, 051912 (2009). 10 Lambert O., Letellier L., Gelbart W.M., Rigaud J.-L., DNA delivery by phage as a strategy for encapsulating toroïdal condensates of arbitrary size into liposomes, Proc. Natl. Acad. Sci. 97, 7248-7253 (2000). 11 Evilevitch A. (2006) Effect of condensing agents and nuclease on the extent of ejection from phage λ. J. Phys. Chem. B 110, 22261-22265. 12 Leforestier, A. and Livolant, F., Structure of toroidal DNA collapsed inside the phage capsid, Proc. Natl. Acad. Sci. 106, 9157-9162 (2009). 13 Jeembaeva, M., Castelnovo, M., Larsson, F., Evilevitch, A. Osmotic pressure : resisting of promoting DNA ejection from phage λ. J. Mol. Biol. 381, 310-323 (2008). A careful examination of Figure 5 led us to recognize the presence of toroids in partially DNA-filled Lambda phage particles. 14 Cremer, T. and Cremer, C., Chromosome territories, nuclear architecture and gene regulation in mammalian cells, Nature Reviews Genetics 2, 292-301 (2001). 15 Cunha, S. , Odijk, T. , Su leymanoglu, E., Woldringh, C. L. , Isolation of the Escherichia coli nucleoid, Biochimie 83 149−154 (2001). 16 Marenduzzo, D., Micheletti, C., Orlandini, E., Biopolymer organization upon confinement. J. Phys. Condens. Matter 22, 283102 (2010). 17 Leforestier A., Livolant F. The bacteriophage genome undergoes a succession of intracapsid phase transitions upon DNA ejection. J. Mol. Biol. 396, 384-395 (2010). 18 Purohit P.K., Kondev J., and Phillips, R., Mechanics of DNA packaging in viruses, PNAS 100 3173-3178 (2003). 19 Šiber, A., Dragar, M., Parsegian, V.A. and Podgornik R., Packing nanomechanics of viral genome, Eur. Phys. J. E 26 317–325 (2008). 20 Raspaud, E., Durand, D. and Livolant, F. , Interhelical Spacing in Liquid Crystalline Spermine and Spermidine-DNA Precipitates, Biophys. J. 88, 392-403 (2005) 21 Rau D. C. and Parsegian V. A., Direct Measurement of the Intermolecular Forces Between Counterion-condensed DNA Double Helices. Evidence for Long Range Attractive Hydration Forces., Biophys. J. 61 246- 259 (1992). Rau D. C. and Parsegian V. A., Direct measurement of temperature-dependent solvation forces between DNA double helices., Biophys. J. 61, 260-271 (1992). 22 Kanduc,M., Naji, A., Forsman, J., and Podgornik, R., Dressed counterions: Strong electrostatic coupling in the presence of salt, J. Chem. Phys. 132 124701-15 (2010). 23 We selected examples from the two protocols but a larger diversity exists in each set of experiments. 24 Lerman, L.S., A Transition to a Compact Form of DNA in Polymer Solutions, PNAS. 68 1886-1890 (1971). 25 Maniatis, T., Venable, J.H., Lerman, L .S., The structure of ψ DNA, J. Mol. Biol. 84, 37-64 (1974). 26 Cohen, J. A. , Podgornik, R. , Hansen, P. L. ,and Parsegian, V. A. , A Phenomenological One-Parameter Equation of State for Osmotic Pressures of PEG and Other Neutral Flexible Polymers in Good Solvents, J. Phys. Chem. B 113 3709-3714 (2009).

27 Béalle, G., Jestinc J. and Carrière, D., Osmotically induced deformation of capsid-like icosahedral vesicles, Soft Matter, Advance Article, DOI: 10.1039/C0SM00490A (2011). 28 Šiber A. and Podgornik R., Stability of elastic icosadeltahedral shells under uniform external pressure: application to viruses under osmotic pressure, Phys. Rev. E 79, 011919 (2009). 29 Evilevitch A. , Gober J.W., Phillips M., Knobler C.M., and Gelbart W.M. Measurements of DNA Lengths remaining in a viral capsid after osmotically suppressed partial ejection. Biophys. J. 88, 751–756 (2005). 30 São-José C.., de Frutos M., Raspaud E., Santos M.A., and Tavares P., Pressure built by DNA packing Inside virions: enough to drive DNA ejection in vitro, largely insufficient for delivery into the bacterial cytoplasm. J.Mol. Biol. 374, 346-355 (2007). 31 Leforestier A., S. Brasilès, M. de Frutos, E. Raspaud, L. Letellier, P. Tavares and F. Livolant Bacteriophage T5 DNA eéjection under pressure. J. Mol. Biol. 384, 730-739 (2008). 32 Ubbink, J. and Odijk T., Deformation of toroidal DNA condensates under surface stress, Europhys. Lett., 33, 353-358 (1996) 33 Or any continuum model for that matter. 34 Landau, L. D., Lifshitz, E. M.: Statistical physics, Vol. 1. Oxford: Pergamon Press (1980). 35 One may wonder whether there is a need to examine the behaviour of the model for negative values of ratio. This would correspond to repulsion between the DNA and the capsid. However, for short-ranged repulsions this would simply result in an effective renormalization of the constraint radius i.e. an appearance of the void spherical shell between the toroid and the capsid whose thickness, would correspond to the effective range of the repulsion. The toroid would thus behave as constrained in a sphere of radius . 36 Tzlil S, Kindt JT, Gelbart WM, Ben-Shaul A, Forces and pressures in DNA packaging and release from viral capsids. Biophys J 84:1616–1627 (2003). 37 Ben-Yaakov, D., Andelman, D., Harries, D. and Podgornik, R., Beyond standard Poisson–Boltzmann theory: ion-specific interactions in aqueous solutions, J. Phys.: Condens. Matter 21 424106 (2009). 38 Papers cited in Ref. 13. 39 One should realize that in that case the DNA subphase and the bathing solution were (sometimes) separated by a semipermeable flexible membrane. The membrane is rigid in our case but we do not think that this invalidates our argument. 40 M. Kanduč, A. Naji, Y. S. Jho, P. A. Pincus and R. Podgornik, The role of multipoles in counterion-mediated interactions between charged surfaces: strong and weak coupling, J. Phys.: Condens. Matter 21 424103 (2009). 41 Rau D.C. and V.A. Parsegian, Direct measurement of the intermolecular forces between counterion-condensed DNA double helices. Evidence for long range attractive hydration forces., Biophys. J. 61 246-259 (1992). 42 Zhang C., Shao, P.G., van Kan J.A., van der Maarel, R.C. Macromolecular crowding induced elongation and compaction in a naochannel. Biophys. J. 97, 1678-1686 (2009). 43 Hou X.-M., Li W., Dou S.-X., Zhang L.-Y., Xie P., Wang W.-C., Wang P.-Y. Formation of DNA toroids Inside confined droplets adsorbed on mica surfaces. Phys. Rev. E 79, 051912 (2009). 44 Fang Y., Hoh J.H., Surface-directed DNA condensation in the absence of soluble multivalent cations. Nucl. Acids Res. 26, 588-593 (1998). 45 Kato A., Sakaue T., Tsuji A., Yoshikawa K. Conformational transition of giant DNA in a confined space surrounded by a phospholipid membrane. Biophys. J.97, 1678-1686 (2009). 46 Parsegian, V.A., Rand R.P . and D.C. Rau, Macromolecules and water: Probing with osmotic stress, Methods in Enzymology 259 43-94 (1995).

47 Helgstrand C., Wikoff W.R., Duda R.L., Hendreix R.W., Johnson J.E., Liljas L. The refined structure of a protein catenane: the HK97 bacteriophage capsid at 3.44 A resolution J. Mol. Biol. 334, 885-899, PDB 1OHG (2003). 48 Effantin G., P. Boulanger, E. Neumann, L. Letellier and J.F. Conway. Bacteriophage T5 Structure Reveals Similarities with HK97 and T4 Suggesting Evolutionary Relationships. J. Mol. Biol. 361, 993-1002 (2006).