
Protection against cisplatin in calorie-restricted Saccharomyces cerevisiae is mediated by the...
13
RESEARCH ARTICLE Protection against cisplatin in calorie-restricted Saccharomyces cerevisiae is mediated by the nutrient-sensor proteins Ras2, Tor1, or Sch9 through its target Glutathione Diana Mariani 1 , Frederico A.V. Castro 1 , Luciana G. Almeida 1 , Fernanda L. Fonseca 2,3 & Marcos D. Pereira 1 1 Departamento de Bioqu ımica, Instituto de Qu ımica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil; 2 Centro de Desenvolvimento Tecnol ogico em Sa ude, CDTS Fiocruz, Rio de Janeiro, RJ, Brazil; and 3 Instituto de Microbiologia Professor Paulo de G oes, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil Correspondence: Marcos D. Pereira, Avenida Athos da Silveira Ramos, 149, Bloco A-5° andar, laborat orio 547, Cidade Universit aria, Rio de Janeiro, RJ, Brasil, CEP: 21941-909. Tel.: +55 21 2562 7735; fax: +55 21 2562 7266; e-mail: [email protected] Received 2 April 2014; revised 5 August 2014; accepted 2 September 2014. Final version published online 21 October 2014. DOI: 10.1111/1567-1364.12214 Editor: Ian Dawes Keywords calorie restriction; cisplatin; Ras2, Sch9, or Tor1; glutathione; Saccharomyces cerevisiae. Abstract There is substantial interest in developing alternative strategies for cancer che- motherapy aiming to increase drug specificity and prevent tumor resistance. Calorie restriction (CR) has been shown to render human cancer cells more susceptible to drugs than normal cells. Indeed, deficiency of nutrient signaling proteins mimics CR, which is sufficient to improve oxidative stress response and life expectancy only in healthy cells. Thus, although CR and reduction of nutrient signaling may play an important role in cellular response to chemo- therapy, the full underlying mechanisms are still not completely understood. Here, we investigate the relationship between the nutrient sensor proteins Ras2, Sch9, or Tor1 and the response of calorie-restricted Saccharomyces cerevisiae cells to cisplatin. Using wild-type and nutrient-sensing mutant strains, we show that deletion of any of these proteins mimics CR and is sufficient to increase cell protection. Moreover, we show that glutathione (GSH) is essential for proper CR protection of yeast cells under cisplatin chemotherapy. By measur- ing the survival rates and GSH levels, we found that cisplatin cytotoxicity leads to a decrease in GSH content reflecting in an increase of oxidative damage. Finally, investigating DNA fragmentation and apoptosis, we conclude that GSH contributes to CR-mediated cell survival. Introduction The search for alternative strategies during cancer chemo- therapy aiming to increase drug specificity and prevent tumor resistance has generated substantial interest. One of these approaches, calorie restriction (CR), makes human cancer cells more susceptible to drugs if compared with normal cells (Lee et al., 1998, 2012; Raffaghello et al., 2008). For instance, the combination of a chemo- therapy drug, for example, cisplatin, with 2-deoxy-D-glu- cose (2DG), a glucose analog which cannot be further metabolized by cells, enhances cytotoxicity of cancer cells by a mechanism related to oxidative stress (Ahmad et al., 2005; Andringa et al., 2006; Simons et al., 2007). Such findings indicate a new horizon for cancer treatment using CR as an alternative for sensitizing cancer rather than normal cells. The process of CR is characterized by the reduction of calorie intake, generally, c. 20–40%, without compromis- ing the normal daily diet. It has been suggested that CR is the best alternative intervention, which improves cellu- lar function of many species (Dilova et al., 2007; Fontana et al., 2010). From yeast to humans, CR triggers complex signaling pathways that promote healthy extension of life due to the enhancement of reproductive time, oxidative stress response, and resistance to abnormal toxic proteins (Roth et al., 2002; Bordone & Guarente, 2005; Dilova et al., 2007). Recently, CR has also been related to pre- vention of several human pathologies such as obesity, dia- betes, hypertension, and atherosclerosis, protection and FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved YEAST RESEARCH
Transcript of Protection against cisplatin in calorie-restricted Saccharomyces cerevisiae is mediated by the...

R E S EA RCH AR T I C L E
Protection against cisplatin in calorie-restricted Saccharomycescerevisiae is mediated by the nutrient-sensor proteins Ras2,
Tor1, or Sch9 through its target Glutathione
Diana Mariani1, Frederico A.V. Castro1, Luciana G. Almeida1, Fernanda L. Fonseca2,3
& Marcos D. Pereira1
1Departamento de Bioqu�ımica, Instituto de Qu�ımica, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil; 2Centro de
Desenvolvimento Tecnol�ogico em Sa�ude, CDTS Fiocruz, Rio de Janeiro, RJ, Brazil; and 3Instituto de Microbiologia Professor Paulo de G�oes,
Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil
Correspondence: Marcos D. Pereira,
Avenida Athos da Silveira Ramos, 149, Bloco
A - 5° andar, laborat�orio 547, Cidade
Universit�aria, Rio de Janeiro, RJ, Brasil, CEP:
21941-909. Tel.: +55 21 2562 7735;
fax: +55 21 2562 7266;
e-mail: [email protected]
Received 2 April 2014; revised 5 August
2014; accepted 2 September 2014. Final
version published online 21 October 2014.
DOI: 10.1111/1567-1364.12214
Editor: Ian Dawes
Keywords
calorie restriction; cisplatin; Ras2, Sch9, or
Tor1; glutathione; Saccharomyces cerevisiae.
Abstract
There is substantial interest in developing alternative strategies for cancer che-
motherapy aiming to increase drug specificity and prevent tumor resistance.
Calorie restriction (CR) has been shown to render human cancer cells more
susceptible to drugs than normal cells. Indeed, deficiency of nutrient signaling
proteins mimics CR, which is sufficient to improve oxidative stress response
and life expectancy only in healthy cells. Thus, although CR and reduction of
nutrient signaling may play an important role in cellular response to chemo-
therapy, the full underlying mechanisms are still not completely understood.
Here, we investigate the relationship between the nutrient sensor proteins Ras2,
Sch9, or Tor1 and the response of calorie-restricted Saccharomyces cerevisiae
cells to cisplatin. Using wild-type and nutrient-sensing mutant strains, we show
that deletion of any of these proteins mimics CR and is sufficient to increase
cell protection. Moreover, we show that glutathione (GSH) is essential for
proper CR protection of yeast cells under cisplatin chemotherapy. By measur-
ing the survival rates and GSH levels, we found that cisplatin cytotoxicity leads
to a decrease in GSH content reflecting in an increase of oxidative damage.
Finally, investigating DNA fragmentation and apoptosis, we conclude that GSH
contributes to CR-mediated cell survival.
Introduction
The search for alternative strategies during cancer chemo-
therapy aiming to increase drug specificity and prevent
tumor resistance has generated substantial interest. One
of these approaches, calorie restriction (CR), makes
human cancer cells more susceptible to drugs if compared
with normal cells (Lee et al., 1998, 2012; Raffaghello
et al., 2008). For instance, the combination of a chemo-
therapy drug, for example, cisplatin, with 2-deoxy-D-glu-
cose (2DG), a glucose analog which cannot be further
metabolized by cells, enhances cytotoxicity of cancer cells
by a mechanism related to oxidative stress (Ahmad et al.,
2005; Andringa et al., 2006; Simons et al., 2007). Such
findings indicate a new horizon for cancer treatment
using CR as an alternative for sensitizing cancer rather
than normal cells.
The process of CR is characterized by the reduction of
calorie intake, generally, c. 20–40%, without compromis-
ing the normal daily diet. It has been suggested that CR
is the best alternative intervention, which improves cellu-
lar function of many species (Dilova et al., 2007; Fontana
et al., 2010). From yeast to humans, CR triggers complex
signaling pathways that promote healthy extension of life
due to the enhancement of reproductive time, oxidative
stress response, and resistance to abnormal toxic proteins
(Roth et al., 2002; Bordone & Guarente, 2005; Dilova
et al., 2007). Recently, CR has also been related to pre-
vention of several human pathologies such as obesity, dia-
betes, hypertension, and atherosclerosis, protection and
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
YEA
ST R
ESEA
RC
H

reduction risk for factors for cancer and cardiovascular
disease, and last but not least, reduce neurodegeneration
(Gross & Dreyfuss, 1990; Roth et al., 2002; Hursting
et al., 2003; Bordone & Guarente, 2005; Dilova et al.,
2007). Although much attention has been dedicated to
understanding the effects of CR, the exact mechanism by
which it exerts such benefits is not completely clear. It is
possible that the lower availability of nutrients during CR
is detected by sensors and effectors, triggering signaling
cascades that account for beneficial effects. Indeed, previ-
ous studies indicated that Ras2, Tor1, and Sch9 are nutri-
ent-responsive proteins related to signaling cascades that
modulate cell metabolism in response to nutrients
(Thevelein et al., 2000; Pedruzzi et al., 2003; Roosen
et al., 2005). These regulatory proteins positively regulate
the expression of common effectors involved in several
processes related to cell growth, such as nutrient uptake,
biogenesis of ribosomes, and protein synthesis (Thevelein
et al., 2000; Martin et al., 2004; Roosen et al., 2005;
Wullschleger et al., 2006). Moreover, they negatively reg-
ulate other important processes, such as stress response
and autophagy (Rolland et al., 2002; Minina et al., 2013).
Interestingly, the decrease (or deletion) of these protein
sensor activities increases replicative life span as well as
oxidative stress response and has been proposed as a
model of CR intervention (Rolland et al., 2002; Dilova
et al., 2007; Fontana et al., 2010; Minina et al., 2013).
Although it has been suggested that CR and reduction of
signaling mediated by nutrient-sensing proteins may play
an important role in cellular response to chemotherapy
(Lee et al., 1998, 2012; Raffaghello et al., 2008), the full
underlying mechanisms are still not completely under-
stood.
Here, we investigate the relationship between nutrient
sensor proteins and the response of yeast cells to chemo-
therapy drugs, for instance cisplatin. Our results show
that deletion of any single nutrient sensor protein tested,
that is, Ras2, Tor1, or Sch9, mimics CR and is sufficient
to increase cell protection against cisplatin. Analysis of
protein–protein interaction (PPI) networks demonstrates
that several transcription factors (TFs) involved in stress
response are negatively regulated by direct or indirect
interaction with Ras2, Tor1, and Sch9. Furthermore,
among the protective factors positively controlled by the
TFs found in PPI, the tripeptide glutathione (c-glutamyl-
cysteinyl-glycine, GSH) has a pivotal role in cells sub-
jected to the effects of cisplatin, possibly acting in the
control of redox status, with consequences to oxidative
stress balance and apoptosis.
Material and methods
Yeast strains and growth conditions
Saccharomyces cerevisiae strains used in this work are
listed in Table 1. Stocks of all strains were maintained on
solid 2% YPD (2% glucose, 1% yeast extract, 2% pep-
tone, and 2% agar) at 4 °C to avoid the selection of
petites or suppressors. For all experiments using the
BY4741 background, cells were grown up to the early
exponential phase (0.8 mg dry weight mL�1) in either
liquid 2% YPD (2% glucose, 1% yeast extract, and 2%
peptone) or 0.5% YPD (1% yeast extract, 0.5% glucose,
and 2% peptone) medium using an orbital shaker at
28 °C and 160 r.p.m., with the ratio of flask volume:
medium of 5 : 1. The wild-type and isogenic strains from
DBY476 background were also grown up to the early
exponential phase but in liquid 2% SD (2% glucose,
0.67% nitrogen base without amino acids) or 0.5% SD
(2% glucose and 0.67% nitrogen base without amino
acids) supplemented with required amino acids (see
genotypes in Table 1).
Evaluation of cisplatin toxicity
Cytotoxicity was analyzed by direct addition of cisplatin
(80 or 450 lM) to 10 mL of the cell culture for 1, 2, and
24 h at 28 °C/160 r.p.m. Cellular viability was monitored
before and after cisplatin exposition by plating, after
proper dilution (10009 in distilled water) in triplicate,
on solidified 2% YPD medium. Then, the plates
were incubated at 28 °C for 72 h and the colonies
counted. Cellular viability was expressed as percentage of
survival.
Table 1. Strains of the yeast Saccharomyces cerevisiae used in this work
Strains Genotype Phenotype Source
DBY476 MATa, leu 2-3.112, his 3Δ1, trp 1-289, ura 3-52, GAL+ Wild type *
sch9D Isogenic to DBY746 except SCH9::URA Sch9 deficient *
ras2D Isogenic to DBY746 except RAS2::LEU Ras2 deficient *
tor1D Isogenic to DBY746 except TOR1::HIS Tor1 deficient *
BY4741 MATa; his3; leu2; met15; ura3 Wild type †
gsh1D Isogenic to BY4741 except GSH1::KanMX4 GSH deficient †
*Andrus Gerontology Center, Department of Biological Sciences, and Norris Cancer Center, University of Southern California, Los Angeles, USA.†Euroscarf, Institute of Molecular Biosciences, Johann Wolfgang Goethe-University Frankfurt, Frankfurt, Germany.
FEMS Yeast Res 14 (2014) 1147–1159ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
1148 D. Mariani et al.

Protein–protein interaction (PPI) network
design and gene ontology (GO) analysis
PPI network between the nutrient-sensing proteins Ras2,
Tor1, and Sch9 and stress responsive proteins as well as
GO was essentially performed as described by Bonatto
(2007). For PPI network analysis, a multiple name search
was performed using STRING 9.05 database (http://
string-db.org/) with limited number of interactors and
high confidence. The obtained PPI networks were
recorded and analyzed using the freeware CYTOSCAPE ver-
sion 3.0.1 available at http://www.cytoscape.org/plugins2.
php. Analysis of GO was carried out with all proteins
obtained in the nutrient-sensing protein PPI network
designed by CYTOSCAPE. Statistical evaluation of GO was
determined by Biological Network Gene Ontology (BINGO)
using the actual annotations of the Gene Ontology Con-
sortium (http://www.geneontology.org).
Determination of glutathione contents
Glutathione, reduced (GSH) or oxidized (GSSG) forms,
was determined spectrophotometrically, in neutralized
10% trichloroacetic acid (TCA) extracts obtained from
cells subjected or not to cisplatin (80 or 450 lM). GSH
and GSSG were measured according to Bernt & Bergmey-
er (1974).
Analysis of lipid peroxidation
For lipid peroxidation determination, 50 mg of cells sub-
jected to cisplatin (450 lM/2 h) at 28 °C was cooled on
ice, harvested by centrifugation, and washed twice with
20 mM Tris-HCl buffer, pH 7.4. Cells were resuspended
in 500 lL of the same buffer, and 1.5 g of glass beads
was added. The samples were lysed by three cycles of 1-
min agitation on a vortex mixer followed by 1 min on
ice and the extracts were used for detection of malondial-
dehyde-MDA as described by Steels et al. (1994).
Evaluation of aconitase inactivation
Aconitase activity was determined spectrophotometrically
before and after exposition of cells to 450 lM cisplatin
for 2 h using an isocitrate dehydrogenase NADPH-depen-
dent coupled reaction, as previously reported by Gardner
et al. (1995).
Analysis of chromatin fragmentation and
apoptosis
Determination of chromatin fragmentation and apoptosis
was performed before and after exposition to cisplatin
(450 lM/2 h). For chromatin fragmentation, cells were
washed two times with PBS, incubated with 5.0 lg mL�1
DAPI (Sigma-Aldrich, St. Louis) in PBS for 30 min and
then washed three times with PBS. Apoptosis was ana-
lyzed by exposition of phosphatidylserine, which was
detected by the reaction with FITC-coupled annexin V
(Annexin V-FITC Apoptosis Detection Kit – APOAF,
Sigma-Aldrich), essentially as described by Madeo et al.
(1997). In both assays, the yeast cells were applied to
microscope slides and observed using an Axioplan 2 fluo-
rescence microscope (Zeiss, Germany). Images were
acquired using a Color View SX digital camera and were
processed with the ANALYSIS software system (Soft Imaging
System) prior to processing with IMAGEJ software (pro-
vided by NIH, http://rsb.info.nih.gov/ij/). To determine
the frequencies of morphological phenotypes (DAPI and
annexinV), at least 300 cells of three independent experi-
ments were evaluated.
Data analysis
The results represent the mean � SD of at least three
independent experiments. Statistical differences were
tested using Student’s t-test. The latter denotes homoge-
neity between experimental groups at P < 0.05.
Results and discussion
Nutrient-sensing proteins affect the response
of S. cerevisiae to chemotherapy by interacting
with TFs
In several biological systems, the nutrient-sensing signal-
ing pathways modulate life span extension and resistance
to oxidative stress (Droge, 2005; Powers et al., 2006; Wei
et al., 2009). In fact, deficiency in a single nutrient-sens-
ing protein such as Ras2, Sch9, or Tor1 is sufficient to
improve oxidative stress response and life expectancy
(Kale & Jazwinski, 1996; Fabrizio et al., 2003; Powers
et al., 2006). Interestingly, deletion of genes that code for
these proteins mimics CR, which explains life span exten-
sion in several species (Fabrizio et al., 2003; Hursting
et al., 2003; Bordone & Guarente, 2005; Powers et al.,
2006; Dilova et al., 2007; Wei et al., 2009).
To investigate whether nutrient-sensing proteins are
related to chemotherapy response, we subjected the wild-
type, DBY746, and Ras2, Tor1, and Sch9 mutant cells to
cisplatin. The absence of one of the nutrient sensor proteins
increased survival of cells after cisplatin treatment (Fig. 1a).
We also tested the performance of these mutant cells to
other chemotherapy drugs such as carboplatin, doxorubi-
cin, etoposide, and 5-fluorouracil. Similarly, all mutant
cells were extremely resistant against chemotherapy
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Protection against cisplatin 1149

(Supporting Information, Fig. S1). Our results indicate that
a common protective mechanism against these chemother-
apeutic drugs seems to be triggered due to the reduction or
impairment of nutrient-sensing signaling through Ras2,
Tor1, and Sch9. The physical interaction between these
nutrient sensors may explain why the absence of a single
nutrient sensor can disrupt nutrient signaling. We have
also confirmed that mutation in these nutrient sensors can
mimic CR. Accordingly, calorie-restricted wild-type strain
presented the same survival profile of mutants when
exposed to cisplatin (Fig. 1b). Taken together, these results
indicate that CR and mutations in nutrient-sensing path-
ways share the same survival mechanisms during cisplatin
chemotherapy.
System biology is a powerful tool widely used to inte-
grate the high amount of physical protein–protein inter-
actions (PPPI) of biological systems solidly grounded on
molecular-level understanding. We performed PPI net-
work analysis between the nutrient-sensing proteins
Ras2, Tor1, and Sch9 and responsive stress targets. As
expected, PPPI network revealed a direct association
between these nutrient-sensing proteins (Fig. 2a and b).
In addition, GO clustering also showed that the most
relevant biological categories were mostly related to sig-
naling transduction, response to nutrient signals, cellular
metabolism related to cell proliferation, and response to
stress (Table 2).
At a molecular level, analysis of PPPI network indicates
that although nutrient-sensing proteins act through dis-
tinct signaling pathways, they overlap regarding stress-
related TFs controlled by Ras2, Tor1, and Sch9 (Fig. 2a
and b). For example, Tor and PKA signaling cascades
negatively regulate Rim15 contributing to repression of
stress genes and stimulating cellular proliferation (Ped-
ruzzi et al., 2003; Swinnen et al., 2006). Rim15 is a ser-
ine/threonine protein kinase repressed by glucose that
plays a crucial function in the upregulation of stress
responsive genes as well as G0 entrance by activating
Msn2/4 and Gis1 (Pedruzzi et al., 2003; Cameroni et al.,
2004; Swinnen et al., 2006;). Furthermore, MCODE analysis
of our PPPI network revealed three subnetworks with
high score and strong significance (5.533; 2.923; and
1.917). These three networks were merged using the CYTO-
SCAPE core Merge Networks plug-in, and the results
obtained indicated the presence of the most relevant fac-
tors related to stress response: heat shock factor 1 (Hsf1),
multicopy suppressor of SNF1 mutation 2 and 4 (Msn2
and Msn4, respectively), suppressor of Kre Null 7 (Skn7),
Glg1-2 suppressor (Gis1), UREidosuccinate transport
(Ure2), and the regulator of IME2 (Rim15) (Fig. 2b).
To find new insights regarding the involvement of
nutrient-sensing proteins and response to stress, we fur-
ther explored within the PPPI network the possible inter-
actions between Ras2, Tor1, and Sch9 and stress-related
TFs. Among the thirteen TFs obtained in our PPPI net-
work, six factors presented high relevance for stress sig-
naling: the well-studied Hsf1, Msn2 and Msn4, Yap1,
Skn7, and Gis1 (Table 3). TFs regulate the expression of
several genes including some coding for major protective
factors (heat shock proteins, antioxidant enzymes, and
glutathione) involved in cell maintenance during stress
conditions (Schmitt & McEntee, 1996; Lee et al., 1999;
Raitt et al., 2000; Hahn et al., 2004; Harbison et al.,
2004). A cross-talk between some of these TFs and nutri-
ent sensors has been reported, strengthening the connec-
tion between them and the appropriate cellular response
under stress conditions (Powers et al., 2006; Swinnen
et al., 2006).
To the best of our knowledge, protective factors such
as superoxide dismutases, catalases, and GSH are under
regulation by the TFs obtained in our PPPI networks
(Table 4). Although those proteins are highly relevant for
cell maintenance under stress conditions, only the absence
of GSH was critical for cell survival under cisplatin expo-
sure (Fig. S2). GSH, synthesized by a coupled reaction
(a)
(b)
Fig. 1. Nutrient-sensing genes modulate survival of Saccharomyces
cerevisiae subjected to cisplatin. (a) The wild-type (DBY746) and
nutrient-sensing mutants (ras2D, tor1D, and sch9D) cells were grown
to early exponential phase in 2% SD media, subjected directly to
80 lM cisplatin for 24 h and then plated on solid 2% YPD. (b) The
wild-type cells were grown to early exponential phase in 2% SD or
0.5% SD media, subjected directly to 80 lM cisplatin for 24 h and
then plated on solid 2% YPD. Cellular survival was determined as the
percentage of viable cells after cisplatin exposition. The results
represent the mean � SD of at least three independent experiments
(*means different results at P < 0.05).
FEMS Yeast Res 14 (2014) 1147–1159ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
1150 D. Mariani et al.

(a)
(b)
Fig. 2. A protein–protein interaction network between the nutrient-sensing proteins Ras2, Tor1, and Sch9. These three initial proteins (black)
were used to create a network of physical protein–protein interactions (60 interactors) whose information was obtained from Saccharomyces
cerevisiae database (a). MCODE analysis of PPPI network revealed three subnetwork merged using the CYTOSCAPE core Merge Networks plug-in (b).
TFs involved in stress response are also highlighted (gray circles). All edges represent first (one-way) interactions.
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Protection against cisplatin 1151

catalyzed by c-glutamylcysteine synthetase (Gsh1) and
glutathione synthetase (Gsh2), is the most abundant
intracellular low molecular weight thiol within cells. It
may have an outstanding function because it is involved
in several physiological processes such as the control of
intracellular redox status by reacting with ROS or bind-
ing to sulfhydryl groups on proteins (glutathiolation),
drug metabolism, DNA synthesis, protein stabilization,
mitochondrial function and integrity, cell proliferation,
and apoptosis (Grant et al., 1996; Ballatori et al., 2009;
Pallard�o et al., 2009). GSH is also a substrate for enzymes
such as glutathione transferases, which catalyze the nucle-
ophilic attack on several electrophilic xenobiotics favoring
the efflux of conjugates from cells (Hayes et al., 2005).
Interestingly, cisplatin can be detoxified through this
mechanism of drug efflux as glutathione-Pt complex, Pt
Table 3. TFs obtained from the PPI network under the regulation of Ras2, Tor1, and Sch9 nutrient protein sensors
Name Biochemical function*
Negatively regulated
Gln3 Positively regulating genes that are subject to nitrogen catabolite repression
Skn7 Activates gene expression in response to heat, oxidative, osmotic, and hypoxic stress conditions
Msn2 Activates cellular general stress response under adverse conditions
Msn4 Activates cellular general stress response under adverse conditions
Hsf1 Activates multiple genes in response to highly diverse stresses, mainly heat stress
Gis1 Activator or repressor involved in the expression of genes involved in postdiauxic shift
Yap1 Activator involved in oxidative stress response and redox homeostasis
Positively regulated
Mbp1 Stimulates proliferation by inducing the expression of genes related to transition from G1 to S phase
Gcn4 Transcriptional activator of amino acid biosynthetic genes. Activated by amino acid starvation prior to entering cell cycle progression
Sfp1 Positive regulator of genes related to ribosome biogenesis
Tec1 Positively controls cell division and vegetative adhesion
Maf1 Negative regulator of RNA polymerase III (pol III) transcription
Ste12 Induces gene expression to mating or pseudohyphal/invasive growth pathways
*Biochemical function was extracted from S. cerevisiae genome database http://www.yeastgenome.org/.
Table 2. Analysis of GO performed using all proteins from the protein–protein network generated by CYTOSCAPE
Biological process (GO category) GO No. P value Corrected P value* k† f‡
Signal transmission 23060 6.51 9 10�25 4.96 9 10�22 33 262
Biological regulation 65007 1.91 9 10�20 2.56 9 10�18 58 1792
Aging 7568 1.93 9 10�18 1.42 9 10�16 19 74
Ras protein signal transduction 7265 2.76 9 10�12 1.85 9 10�10 12 34
Response to stimulus 50896 7.88 9 10�9 4.53 9 10�7 35 965
Response to chemical stimulus 42221 3.86 9 10�8 1.83 9 10�6 23 399
Second-messenger-mediated signaling 19932 7.28 9 10�8 3.25 9 10�6 9 29
Regulation of cell size 8361 3.06 9 10�7 1.30 9 10�5 13 104
Regulation of cAMP biosynthetic process 30817 1.08 9 10�6 3.48 9 10�5 5 5
Regulation of cyclic nucleotide biosynthetic process 30802 1.08 9 10�6 3.48 9 10�5 5 5
Developmental process 32502 2.45 9 10�6 7.04 9 10�5 21 399
Regulation of molecular function 65009 3.49 9 10�6 9.03 9 10�5 14 153
Regulation of catalytic activity 50790 3.59 9 10�6 9.03 9 10�5 13 126
Regulation of cellular metabolic process 31323 1.40 9 10�5 2.89 9 10�4 32 1045
Protein amino acid phosphorylation 6468 1.61 9 10�5 3.17 9 10�4 13 142
Regulation of transcription, DNA-dependent 6355 6.72 9 10�5 1.13 9 10�3 21 478
TOR signaling cascade 31929 7.71 9 10�5 1.27 9 10�3 6 16
Regulation of RNA metabolic process 51252 1.01 9 10�4 1.61 9 10�3 21 489
Response to abiotic stimulus 9628 1.03 9 10�4 1.61 9 10�3 13 165
Regulation of cell growth 1558 4.75 9 10�4 6.48 9 10�3 5 11
Response to stress 6950 5.99 9 10�4 7.65 9 10�3 24 704
Cellular response to nutrient levels 31669 8.15 9 10�3 8.48 9 10�2 8 74
*P values calculated after FDR application.†Number of proteins found in the network that integrate a specific GO.‡Total number of proteins belonging to a specific GO.
FEMS Yeast Res 14 (2014) 1147–1159ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
1152 D. Mariani et al.
(GS)2 (Hayes et al., 2005; Kuo et al., 2007), which may
be of relevance in understanding the relationship between
GSH and the protection it confers to this chemotherapy
drug. Indeed, analyzing the levels of GSH in the wild-type
DBY746, we have observed that cisplatin chemotherapy
caused a significant reduction (�fourfold) in the levels of
GSH (Fig. 3a). On the other hand, in the nutrient-sensing
mutant strains Tor1 and Sch9, the same treatment with
cisplatin did not affect the levels of GSH (Fig. 3a).
Regarding to Ras2 mutant strain, the 1.5-fold reduction
did not seem to be relevant to promote cell susceptibility
during cisplatin exposure. Furthermore, analyzing the
GSH levels in calorie-restricted wild-type (DBY746)
strain, we can observe the same GSH profile as presented
by the Ras2 mutant (Fig. 4). This result combined with
survival of Ras2 and calorie-restricted wild type indicates
that may exist a threshold in GSH levels necessary to pro-
tect cells during cisplatin chemotherapy. In accordance,
the treatment of cells with BSO (buthionine sulfoximine),
an inhibitor of GSH synthesis, conferred a strong sensitiv-
ity in all cells studied (Fig. S3) Interestingly, GSSG levels
remained unchanged in all strains (Figs 3b and 4b), indi-
cating that the decrease of GSH levels is mainly due to Pt
(GS)2 generation. In addition, these results also show that
Ras2, Tor1, and Sch9 single deletions may be advanta-
geous for GSH metabolism.
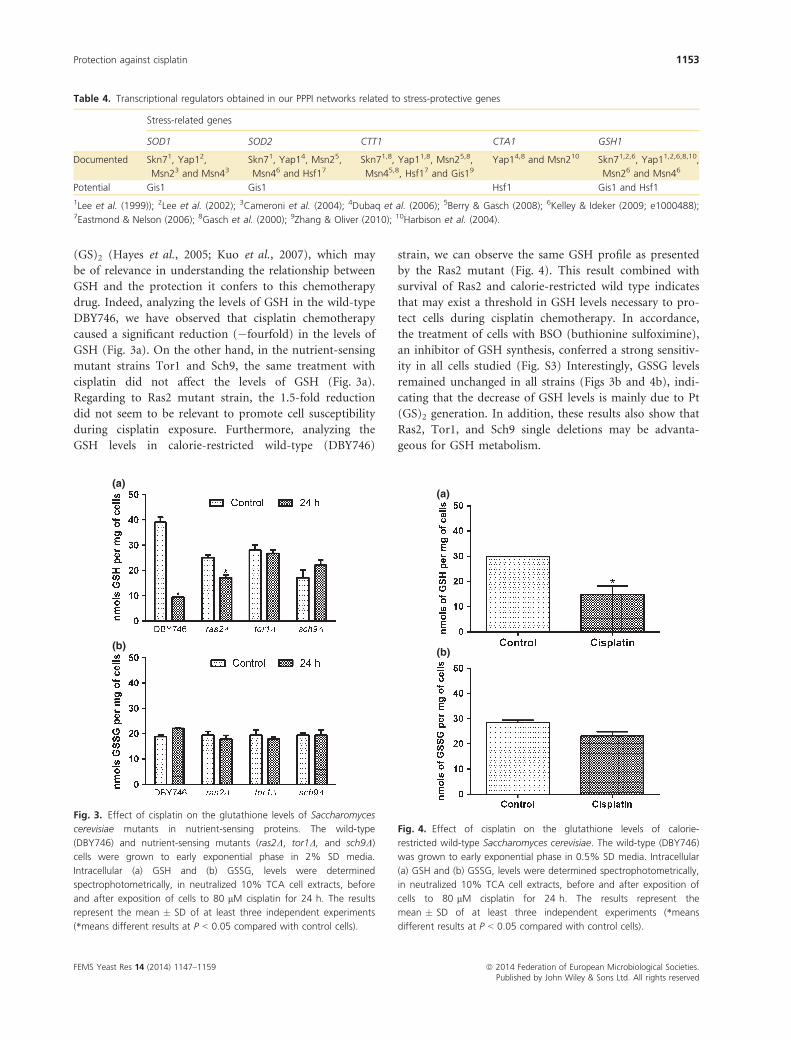
Table 4. Transcriptional regulators obtained in our PPPI networks related to stress-protective genes
Stress-related genes
SOD1 SOD2 CTT1 CTA1 GSH1
Documented Skn71, Yap12,
Msn23 and Msn43Skn71, Yap14, Msn25,
Msn46 and Hsf17Skn71,8, Yap11,8, Msn25,8,
Msn45,8, Hsf17 and Gis19Yap14,8 and Msn210 Skn71,2,6, Yap11,2,6,8,10,
Msn26 and Msn46
Potential Gis1 Gis1 Hsf1 Gis1 and Hsf1
1Lee et al. (1999)); 2Lee et al. (2002); 3Cameroni et al. (2004); 4Dubaq et al. (2006); 5Berry & Gasch (2008); 6Kelley & Ideker (2009; e1000488);7Eastmond & Nelson (2006); 8Gasch et al. (2000); 9Zhang & Oliver (2010); 10Harbison et al. (2004).
(a)
(b)
Fig. 3. Effect of cisplatin on the glutathione levels of Saccharomyces
cerevisiae mutants in nutrient-sensing proteins. The wild-type
(DBY746) and nutrient-sensing mutants (ras2D, tor1D, and sch9D)
cells were grown to early exponential phase in 2% SD media.
Intracellular (a) GSH and (b) GSSG, levels were determined
spectrophotometrically, in neutralized 10% TCA cell extracts, before
and after exposition of cells to 80 lM cisplatin for 24 h. The results
represent the mean � SD of at least three independent experiments
(*means different results at P < 0.05 compared with control cells).
(a)
(b)
Fig. 4. Effect of cisplatin on the glutathione levels of calorie-
restricted wild-type Saccharomyces cerevisiae. The wild-type (DBY746)
was grown to early exponential phase in 0.5% SD media. Intracellular
(a) GSH and (b) GSSG, levels were determined spectrophotometrically,
in neutralized 10% TCA cell extracts, before and after exposition of
cells to 80 lM cisplatin for 24 h. The results represent the
mean � SD of at least three independent experiments (*means
different results at P < 0.05 compared with control cells).
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Protection against cisplatin 1153

GSH is important for the fully operating CR
protection mechanism against cisplatin in
S. cerevisiae
Given the observed connection between nutrient-sensing
signaling pathways and GSH, we decided to further investi-
gate the involvement of GSH in CR benefits under cisplatin
treatment. Thus, using the wild-type BY4741 and the iso-
genic mutant strain deficient in Gsh1, we analyzed the
effect of GSH absence in calorie-restricted cell response
under cisplatin chemotherapy. As expected, cells of the
wild-type strain BY4741, growing on 2% glucose medium,
showed increased cisplatin susceptibility over time
(Fig. 5a). On the other hand, when calorie-restricted cells
were subjected to cisplatin treatment, resistance was
observed (Fig. 5b). This result confirms that regulating the
calorie uptake, by nutrient sensor modulation or even
nutrient intake, benefits cell response against cisplatin che-
motherapy. Accordingly, previous findings described that
dietary restriction protects from toxicities of
chemotherapeutic agents (Raffaghello et al., 2008; Lee
et al., 2012). However, the possibility of regulating calorie
intake to protect humans against the harmful effects of che-
motherapeutic agents is still under investigation (Safdie
et al., 2009). Interestingly, unlike the wild-type strain, the
gsh1 mutant lost the ability to respond to cisplatin even
when cells were cultured under CR (Fig. 5a and b). The
maintenance of GSH content in calorie-restricted cells
exposed to cisplatin, as revealed in Fig. 6, seems to account
for this improved survival. Indeed, the preservation of GSH
levels in calorie-restricted cells, after cisplatin exposure,
contributed to their high survival, whereas cells cultured in
high-glucose culture experienced a drop in GSH content
and decreased survival. These results point to a central role
of GSH for survival and cellular response during cisplatin
chemotherapy. Although there are several findings showing
the contribution of GSH for cellular protection, this is the
first evidence for a role of GSH in cellular protection con-
ferred by the CR regimen against cisplatin.
As cisplatin exposure induced a decrease in the GSH
content in cells grown in media containing 2% glucose,
we measured the extent of oxidative damage generated by(a)
(b)
Fig. 5. Intracellular glutathione is important for survival of
Saccharomyces cerevisiae cells subjected to cisplatin. The wild-type
(BY4741) and its isogenic mutant gsh1D were grown to early
exponential phase in 2% YPD (a) or 0.5% YPD (b) media, subjected
directly to 450 lM cisplatin for 24 h and then plated on solid 2%
YPD. Cellular survival was determined, at appropriate time, as the
percentage of viable cells after cisplatin exposition. The results
represent the mean � SD of at least three independent experiments
(*means different results at P < 0.05 compared with the wild-type
strain; **means different results at P < 0.05 compared with cells
cultured in 2% SD).
(a)
(b)
Fig. 6. Effect of cisplatin exposition on glutathione levels in
Saccharomyces cerevisiae cells growing on rich (2% YPD) or calorie-
restricted (0.5% YPD) media composition. The wild-type (BY4741)
was grown to early exponential phase in 2% YPD (a) or 0.5% YPD
(b) media. Intracellular GSH and GSSG, levels were determined
spectrophotometrically, in neutralized 10% TCA cell extracts, before
and after exposition of cells to 450 lM cisplatin for 2 h. The results
represent the mean � SD of at least three independent experiments
(*means different results at P < 0.05 compared with control cells;
**means different results at P < 0.05 compared with cells cultured in
2% SD).
FEMS Yeast Res 14 (2014) 1147–1159ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
1154 D. Mariani et al.

cisplatin treatment. The relevance of GSH-dependent CR
protection was also investigated. During oxidative stress,
lipids and intracellular proteins are the main targets of
ROS. While lipid peroxidation is a chain reaction causing
the loss of membrane properties and the production of
more toxic by-products, such as MDA (Steels et al.,
1994), the oxidative inactivation of proteins, such as
aconitase, occurs mainly due to the presence of iron–sul-fur (Fe-S) cluster that is susceptible to inactivation by
superoxide radicals (Gardner et al., 1995). Although cis-
platin halved the levels of GSH in the wild-type strain
(cultured in high glucose), this reduction was not fol-
lowed by damage to oxidative stress markers (Fig. 7a and
b). However, when the GSH synthesis was abolished in
the mutant strain gsh1D, we observed increased damage
in lipid peroxidation and inactivation of aconitase
(Fig. 7a and b). On the other hand, when calorie-
restricted cells were subjected to cisplatin, two distinct
aspects were observed; (1) an increase in the levels of
lipid peroxidation in the mutant gsh1D; and (2) a four-
fold increase in aconitase activity in the wild type in rela-
tion to the GSH-deficient strain. Indeed, the oxidative
damages were mainly observed in the gsh1D strain. It has
been described that depletion of GSH is related to defects
in iron homeostasis leading to an impairment of cellular
function. Furthermore, the authors stated that GSH is
not effective by itself to control cellular thiol-redox func-
tions contributing to enhanced cytotoxicity observed in
the mutant gsh1D (Kumar et al., 2011). Taken together,
besides cisplatin cytotoxicity, the abolishment of GSH
synthesis causes cellular disorders that put in check the
fully operating CR mechanism of protection.
Several data highlight the essential role of GSH for reg-
ulation of apoptosis signals (Rudin et al., 2003; P�ocsi
et al., 2004; Anathy et al., 2012). In addition, it has been
described that enhancement of intracellular ROS is deci-
sive to trigger apoptosis signals in yeast (P�ocsi et al.,
2004). Thus, an efficient antioxidant system is required
for the accurate balance between generation and elimina-
tion of intracellular ROS preventing the induction of the
hallmarks of programmed cell death. To investigate
whether cisplatin activates apoptosis in gsh1 mutant calo-
rie-restricted cells, we analyzed the levels of DNA frag-
mentation and the phosphatidylserine externalization by
DAPI and annexin staining, respectively. As expected, cis-
platin exposure induced DNA fragmentation in both yeast
strains growing in high-glucose medium; however, the
extent of DNA damage was significantly higher in cells
harboring GSH deficiency (Fig. 8a and b). This result is
consistent with certain data from the literature showing
that the lack of GSH, caused mainly by chemical stresses,
leads to the activation of apoptosis (Rudin et al., 2003;
P�ocsi et al., 2004; Anathy et al., 2012). The induction of
apoptosis has multiple features but the most outstanding
event during apoptosis is the strikingly depletion of GSH
levels without any dependence of ROS generation (Franco
(a) (b)
(c) (d)
Fig. 7. Cisplatin induces oxidative stress in glutathione-deficient Saccharomyces cerevisiae cells. Lipid peroxidation was measured by the TBARS
method in the early exponential wild type and its isogenic gsh1D mutant cells grown in 2% YPD (a and b) or 0.5% YPD (c and d) and then
subjected to 450 lM cisplatin for 2 h. Controls are cells that were not subjected to cisplatin. Aconitase activity was determined
spectrophotometrically in S. cerevisiae cells subjected to the same conditions described in lipid peroxidation. The results represent the mean � SD
of at least three independent experiments (*means different results at P < 0.05 compared with control; **means different results at P < 0.05
compared with 2% SD).
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Protection against cisplatin 1155

et al., 2007). Interestingly, the DNA damage observed in
the mutant strain was associated with the increase in
cytotoxicity and oxidative stress markers indicating that
during cisplatin exposure cells might be affected by both
alkylation and oxidative stress-induced apoptosis. Regard-
ing the effects of CR in DNA fragmentation, we can
observe an increase in the number of wild-type cells pre-
senting DAPI staining, contrasting with the reduction in
the percentage of DNA fragmentation in cells harboring
deficiency in GSH synthesis. This result suggests that CR
could be activating a self-defense system to protect the
cell population through apoptosis. As shown in Fig. 8c,
the higher number of annexin-positive cells in the gsh1Dcells suggests that deficiency in GSH metabolism contrib-
utes to apoptosis. On the other hand, due to the reduc-
tion in annexin staining, GSH seems to be essential for
activating apoptosis in calorie-restricted cells. Remarkably,
in calorie-restricted gsh1 mutant cells, the apoptotic pro-
file was quite different from the wild-type cells, suggesting
that for apoptosis activation in CR regimen, the presence
of GSH is vital.
At the best of our knowledge, cancer cells show altered
metabolism in comparison with its healthy counterparts.
Overall, cancer cells display increased rates of fermenta-
tive metabolism with enhancement of lactate production
(Warburg, 1956). Indeed, the reduced rates of respiration
associated with high rates of glycolysis suggest that cancer
cells depend mainly on ‘aerobic fermentation’, also
known as ‘Warburg effect’ (Brand, 1997; Gatenby & Gil-
lies, 2004). Interestingly, although the exact mechanism
by how cancer cells undergo this metabolic shift are still
unclear, this phenomenon may indicate an advantage for
cancer cells allowing them to proliferate in microenviron-
ments (hypoxic areas) usually encountered in solid
tumors (Gatenby & Gillies, 2004). As a Crabtree-positive
cell, S. cerevisiae has the capacity to alter its metabolism
according to nutrients availability. Thus, when glucose is
abundant S. cerevisiae uses fermentative metabolism as its
main energy supply source; however, when this nutrient
is depleted this yeast has the ability to shift to respiratory
metabolism (Entian et al., 1984; Thevelein, 1994). There-
fore, due to similar metabolic features of cancer cells,
S. cerevisiae has proved to be an excellent cellular
model for the screening of metabolism-targeted drugs
and alternative anticancer therapy (D�ıaz-Ruiz et al., 2009,
2011).
(a)
(b) (c)
Fig. 8. Impact on the chromatin fragmentation and apoptosis of glutathione-deficient S. cerevisiae cells subjected to cisplatin. (a) Representative
photographs of the wild-type (BY4741) and its isogenic mutant gsh1D grown to early exponential phase in 2% YPD or 0.5% YPD media,
subjected directly to 450 lM cisplatin for indicated 2 h and then stained with DAPI for chromatin fragmentation. Percentage of chromatin
fragmentation (b) and annexin V (c) in S. cerevisiae cells following exposition with cisplatin (450 lM/2 h) (*means different results at P < 0.05
between cells cultured in 2% SD and 0.5% SD ; **means different results at P < 0.05 between gsh1D cells cultured in 2% SD and 0.5% SD).
FEMS Yeast Res 14 (2014) 1147–1159ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
1156 D. Mariani et al.

CR or glucose deprivation has proven to generate a
metabolic shift from fermentative to respiratory pathways
(Lin et al., 2002; Oliveira et al., 2008). Several studies
have proved that CR makes human cancer cells more sus-
ceptible to drugs if compared with normal (healthy) cells
due to the induction of ROS generation and subse-
quently, oxidative stress (Lee et al., 1998, 2012; Raffag-
hello et al., 2008). In this context, the combination of CR
with chemotherapy agents (e.g. cisplatin) seems to be a
potential alternative intervention to induce cancer cell
death. The rational of this therapy is to promote a dual
mode of cytotoxicity, which leads to the induction of oxi-
dative damages through CR or glucose deprivation (2DG
administration) and by cisplatin (chemotherapy agents)
mode of action, both acting together to enhance cell
death (Simons et al., 2007). Unfortunately, the efficacy of
this therapy falls on the need to evaluate the redox state
of healthy cells before the start of the therapy; otherwise
this alternative treatment tends to be also highly cytotoxic
for normal cells. In this work using S. cerevisiae as a
model of healthy cells, we showed that CR induces
cellular protection against chemotherapy but it strikingly
dependent of GSH contents. Our results showed that in
healthy calorie-restricted cells, the absence of GSH
imposed critical dysfunctions leading to a dramatic
decrease in survival associated with oxidative damages
(e.g. lipid peroxidation and aconitase) and a significant
reduction in apoptosis. Furthermore, the critical role
of GSH for CR benefits is clearly shown when nutrient-
sensing mutants, which mimic CR, were treated with
BSO.
Taken together, the benefits of controlling calorie
intake for cells subjected to chemotherapy drugs are evi-
dent, but in this work, we highlight for the first time a
role of GSH for cellular protection against cisplatin. In
fact, the lack of GSH is critical for proper cellular
response, even if cells are in calorie-restrictive conditions.
Although improvement of cell response can be achieved
by CR, it seems to be very important to continue the
efforts to evaluate the real redox status of biological
systems before starting a CR-related chemotherapy.
Acknowledgement
This work was supported by grants from FAPERJ,
CAPES, and CNPq.
References
Ahmad IM, Aykin-Burns N, Sim JE et al. (2005)
Mitochondrial O�2 and H2O2 mediate glucose
deprivation-induced cytotoxicity and oxidative stress in
human cancer cells. J Biol Chem 280: 4254–4263.
Anathy V, Roberson EC, Guala AS, Godburn KE, Budd RC &
Janssen-Heininger YM (2012) Redox-based regulation of
apoptosis: S-glutathionylation as a regulatory mechanism to
control cell death. Antioxid Redox Signal 16: 496–505.Andringa KK, Coleman MC, Aykin-Burns N, Hitchler MJ,
Walsh SA, Domann FE & Spitz DR (2006) Inhibition of
glutamate cysteine ligase (GCL) activity sensitizes human
breast cancer cells to the toxicity of 2-deoxy-D-glucose.
Cancer Res 66: 1605–1610.Ballatori N, Krance SM, Notenboom S, Shi S, Tieu K &
Hammond CL (2009) Glutathione dysregulation and the
etiology and progression of human diseases. Biol Chem 390:
191–214.Bernt E & Bergmeyer U (1974) Glutathione. Methods Enz Anal
4: 1643–1647.Berry DB & Gasch AP (2008) Stress-activated genomic
expression changes serve a preparative role for impending
stress in yeast. Mol Biol Cell 19: 4580–4587.Bonatto D (2007) A systems biology analysis of protein–
protein interactions between yeast superoxide dismutases
and DNA repair pathways. Free Radic Biol Med 43: 557–567.Bordone L & Guarente L (2005) Calorie restriction, SIRT1 and
metabolism: understanding longevity. Nat Rev Mol Cell Biol
6: 298–305.Brand K (1997) Aerobic glycolysis by proliferating cells:
protection against oxidative stress at the expense of energy
yield. J Bioenerg Biomembr 29: 355–364.Cameroni E, Hulo N, Roosen J, Winderickx J & De Virgilio C
(2004) The novel yeast PAS kinase Rim 15 orchestrates
G0-associated antioxidant defense mechanisms. Cell Cycle 3:
462–468.D�ıaz-Ruiz R, Uribe-Carvajal S, Devin A & Rigoulet M (2009)
Tumor cell energy metabolism and its common features
with yeast metabolism. Biochim Biophys Acta 1796: 252–265.
D�ıaz-Ruiz R, Rigoulet M & Devin A (2011) The Warburg and
Crabtree effects: on the origin of cancer cell energy
metabolism. Biochim Biophys Acta 1807: 568–576.Dilova I, Easlon E & Lin S-J (2007) Calorie restriction and the
nutrient sensing signaling pathways. Cell Mol Life Sci 64:
752–767.Droge W (2005) Oxidative aging and insulin receptor
signaling. J Gerontol A Biol Sci Med Sci 60: 1378–1385.Dubaq C, Chavalier A, Courbeyrette R, Petat C, Gidrol X &
Mann C (2006) Role of the iron mobilization and oxidative
stress regulons in the genomic response of yeast to
hydroxyurea. Mol Genet Genomics 275: 114–124.Eastmond DL & Nelson HC (2006) Genome-wide analysis
reveals new roles for the activation domains of the
Saccharomyces cerevisiae heat shock transcription factor
(Hsf1) during the transient heat shock response. J Biol
Chem 281: 32909–32921.Entian KD, Frohlich KU & Mecke D (1984) Regulation of
enzymes and isoenzymes of carbohydrate metabolism in the
yeast Saccharomyces cerevisiae. Biochim Biophys Acta 799:
181–186.
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Protection against cisplatin 1157

Fabrizio P, Liou LL, Moy VN, Diaspro A, Valentine JS, Gralla
EB & Longo VD (2003) Sod2 functions downstream of Sch9
to extend longevity in yeast. Genetics 163: 35–46.Fontana L, Partridge L & Longo VD (2010) Extending healthy
life span-from yeast to humans. Science 328: 321–326.Franco R, Panayiotidis MI & Cidlowski JA (2007) Glutathione
depletion is necessary for apoptosis in lymphoid cells
independent of reactive oxygen species formation. J Biol
Chem 282: 30452–30465.Gardner PR, Raineri I, Epstein LB & White CW (1995)
Superoxide radical and iron modulate aconitase activity in
mammalian cells. J Biol Chem 270: 13399–13405.Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB,
Storz G, Botstein D & Brown PO (2000) Genomic
expression programs in the response of yeast cells to
environmental changes. Mol Biol Cell 11: 4241–4257.Gatenby RA & Gillies RJ (2004) Why do cancers have high
aerobic glycolysis? Nat Rev Cancer 4: 891–899.Grant CM, MacIver FH & Dawes IW (1996) Glutathione is an
essential metabolite required for resistance to oxidative
stress in the yeast Saccharomyces cerevisiae. Curr Genet 29:
511–515.Gross L & Dreyfuss Y (1990) Prevention of spontaneous and
radiation-induced tumors in rats by reduction of food
intake. P Natl Acad Sci USA 87: 6795–6797.Hahn J-S, Hu Z, Thiele DJ & Iyer VR (2004) Genome-wide
analysis of the biology of stress responses through heat
shock transcription factor. Mol Cell Biol 24: 5249–5256.Harbison CT, Gordon DB, Lee TI et al. (2004) Transcriptional
regulatory code of a eukaryotic genome. Nature 431: 99–104.Hayes JD, Flanagan JU & Jowsey IR (2005) Glutathione
transferases. Annu Rev Pharmacol Toxicol 45: 51–88.Hursting SD, Lavigne JA, Berrigan D, Perkins SN & Barrett JC
(2003) Calorie restriction, aging, and cancer prevention:
mechanisms of action and applicability to humans. Annu
Rev Med 54: 131–152.Kale SP & Jazwinski SM (1996) Differential response to UV
stress and DNA damage during the yeast replicative life
span. Dev Genet 18: 154–160.Kelley R & Ideker T (2009) Genome-wide fitness and
expression profiling implicate Mga2 in adaptation to
hydrogen peroxide. PLoS Genet 5: e1000488.
Kumar C, Igbaria A, D0autreaux B, Planson A-G, Junot C,
Godat E, Bachhawat AK, Delaunay-Moisan A & Toledano
MB (2011) Glutathione revisited: a vital function in iron
metabolism and ancillary role in thiol-redox control. EMBO
J 30: 2044–2056.Kuo MT, Chen HHW, Song IS, Savaraj N & Ishikawa T
(2007) The roles of copper transporters in cisplatin
resistance. Cancer Metastasis Rev 26: 71–83.Lee YJ, Galoforo SS, Berns CM, Chen JC, Davis BH, Si JE,
Corry PM & Spitz DR (1998) Glucose deprivation-induced
cytotoxicity and alterations in mitogen-activated protein
kinase activation are mediated by oxidative stress in
multidrug-resistant human breast carcinoma cells. J Biol
Chem 273: 5294–5299.
Lee J, Godon C, Lagniel G, Spector D, Garini J, Labarre J &
Toledano MB (1999) Yap1 and Skn7 control two specialized
oxidative stress response regulons in yeast. J Biol Chem 274:
16040–16046.Lee Tl, Rinaldi NJ, Robert F et al. (2002) Transcriptional
regulatory networks in Saccharomyces cerevisiae. Science 298:
799–804.Lee C, Raffaghello L, Brandhorst S et al. (2012) Fasting cycles
retard growth of tumors and sensitize a range of cancer cell
types to chemotherapy. Sci Transl Med 4: 124ra27.
Lin S-J, Kaeberlein M, Andalis AA, Sturtz LA, Defossez P-A,
Culotta VC, Fink GR & Guarente L (2002) Calorie
restriction extends Saccharomyces cerevisiae lifespan by
increasing respiration. Nature 418: 344–348.Madeo F, Fr€ohlich E & Fr€ohlich K-U (1997) J Cell Biol 139:
729–734.Martin DE, Soulard A & Hall MN (2004) TOR regulates
ribosomal protein gene expression via PKA and the
forkhead transcription factor FHL1. Cell 119: 969–979.Minina EA, Sanchez-Vera V, Moschou PN, Suarez MF,
Sundberg E, Weih M & Bozhkov PV (2013) Autophagy
mediates caloric restriction-induced lifespan extension in
Arabidopsis. Ageing Cell 12: 327–329.Oliveira GA, Tahara EB, Gombert AK, Barros MH &
Kowaltowski AJ (2008) Increased aerobic metabolism is
essential for the beneficial effects of caloric restriction on
yeast life span. J Bioenerg Biomembr 40: 381–388.Pallard�o FV, Markovic J, Garc�ıa JL & Vi~na J (2009) Role of
nuclear glutathione as a key regulator of cell proliferation.
Mol Aspects Med 30: 77–85.Pedruzzi I, Dubouloz F, Cameroni E, Wanke V, Roosen J,
Winderickx J & DeVirgilio C (2003) TOR and PKA
signaling pathways converge on the protein kinase Rim15 to
control entry into G0. Mol Cell 12: 1607–1613.P�ocsi I, Prade RA & Penninckx MJ (2004) Glutathione,
altruistic metabolite in fungi. Adv Microb Physiol 49: 1–76.Powers RW, Kaeberlein M, Caldwell SD, Kennedy BK &
Fields S (2006) Extension of chronological life span in
yeast by decreased TOR pathway signaling. Genes Dev 20:
174–184.Raffaghello L, Lee C, Safdie FM, Wei M, Madia F, Bianchi G
& Longo VD (2008) Starvation-dependent differential stress
resistance protects normal but not cancer cells against
high-dose chemotherapy. P Natl Acad Sci USA 105:
8215–8220.Raitt DC, Johnson AL, Erkine AM, Makino K, Morgan B,
Gross DS & Johnston LH (2000) The Skn7 response
regulator of Saccharomyces cerevisiae interacts with Hsf1 in
vivo and is required for the induction of heat shock genes
by oxidative stress. Mol Biol Cell 11: 2335–2347.Rolland F, Winderickx J & Thevelein JM (2002)
Glucose-sensing and -signalling mechanisms in Yeast. FEMS
Yeast Res 2: 183–201.Roosen J, Engelen K, Marchal K, Mathys J, Griffioen G,
Cameroni E, Thevelein JM, DeVirgilio C, DeMoor B &
Winderickx J (2005) PKA and Sch9 control a molecular
FEMS Yeast Res 14 (2014) 1147–1159ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
1158 D. Mariani et al.

switch important for the proper adaptation to nutrient
availability. Mol Microbiol 55: 862–880.Roth GS, Lane MA, Ingram DK, Mattison JA, Elahi D, Tobin
JD, Muller D & Metter EJ (2002) Biomarkers of caloric
restriction may predict longevity in humans. Science 297: 811.
Rudin CM, Yang Z, Schumaker LM, Vander Weele DJ,
Newkirk K, Egorin MJ, Zuhowski EG & Cullen KJ (2003)
Inhibition of glutathione synthesis reverses Bcl-2-mediated
cisplatin resistance. Cancer Res 63: 312–318.Safdie FM, Dorff T, Quinn D, Fontana L, Wei M, Lee C,
Cohen P & Longo VD (2009) Fasting and cancer treatment
in humans: a case series report. Aging 1: 988–1007.Schmitt AP & McEntee K (1996) Msn2p, a zinc finger
DNA-binding protein, is the transcriptional activator of the
multistress response in Saccharomyces cerevisiae. P Natl Acad
Sci USA 93: 5777–5782.Simons Al, Ahmad IM, Mattson DM, Dornfeld KJ & Spitz DR
(2007) 2-Deoxy-d-glucose combined with cisplatin enhances
cytotoxicity via metabolic oxidative stress in human head
and neck cancer cells. Cancer Res 67: 3364–3370.Steels EL, Learmonth RP & Watson K (1994) Stress tolerance
and membrane lipid unsaturation in Saccharomyces
cerevisiae grown aerobically or anaerobically. Microbiology
140: 569–576.Swinnen E, Wanke V, Roosen J, Smets B, Dubouloz F,
Pedruzzi I, Cameroni E, De Virgilio C & Winderickx J
(2006) Rim15 and the crossroads of nutrient signalling
pathways in Saccharomyces cerevisiae. Cell Div 3: 1–3.Thevelein JM (1994) Signal transduction in yeast. Yeast 10:
1753–1790.
Thevelein JM, Cauwenberg L, Colombo S et al. (2000)
Nutrient-induced signal transduction through the protein
kinase A pathway and its role in the control of metabolism,
stress resistance, and growth in yeast. Enzyme Microb
Technol 26: 819–825.Warburg O (1956) On the origin of cancer cells. Science 123:
309–314.Wei M, Fabrizio P, Madia F, Hu J, Ge H, Li LM & Longo VD
(2009) Tor1/Sch9-regulated carbon source substitution is as
effective as calorie restriction in life span extension. PLoS
Genet 5: e1000467.
Wullschleger S, Loewith R & Hall MN (2006) TOR signaling
in growth and metabolism. Cell 124: 471–484.Zhang N & Oliver SG (2010) The transcription activity of Gis1
is negatively modulated by proteasome-mediated limited
proteolysis. J Biol Chem 285: 6465–6476.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Fig. S1. Nutrient-sensing genes modulate survival of Sac-
charomyces cerevisiae subjected to chemotherapy drugs.
Fig. S2. Survival of Saccharomyces cerevisiae cells sub-
jected to cisplatin.
Fig. S3. Dependence of GSH for nutrient-sensing proteins
modulate survival of Saccharomyces cerevisiae subjected to
cisplatin.
FEMS Yeast Res 14 (2014) 1147–1159 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Protection against cisplatin 1159




















