
Profiles of Purine Metabolism in Leaves and Roots of ...
14
To determine the metabolic profiles of purine nucleotides and related compounds in leaves and roots of tea ( Camellia sinensis), we studied the in situ metabolic fate of 10 different 14 C-labeled precursors in segments from tea seedlings. The activities of key enzymes in tea leaf extracts were also investigated. The rates of uptake of purine precursors were greater in leaf segments than in root segments. Adenine and adenosine were taken up more rapidly than other purine bases and nucleosides. Xanthosine was slowest. Some adenosine, guanosine and inosine was converted to nucleotides by adenosine kinase and inosine/guanosine kinase, but these compounds were easily hydrolyzed, and adenine, guanine and hypoxanthine were generated. These purine bases were salvaged by adenine phosphoribosyltransferase and hypoxanthine/ guanine phosphoribosyltransferase. Salvage activity of adenine and adenosine was high, and they were converted exclusively to nucleotides. Inosine and hypoxanthine were salvaged to a lesser extent. In situ 14 C-tracer experiments revealed that xanthosine and xanthine were not salvaged, although xanthine phosphoribosyltransferase activity was found in tea extracts. Only some deoxyadenosine and deoxyguanosine was salvaged and utilized for DNA synthesis. However, most of these deoxynucleosides were hydrolyzed to adenine and guanine and then utilized for RNA synthesis. Purine alkaloid biosynthesis in leaves is much greater than in roots. In situ experiments indicate that adenosine, adenine, guanosine, guanine and inosine are better precursors than xanthosine, which is a direct precursor of a major pathway of caffeine biosynthesis. Based on these results, possible routes of purine metabolism are discussed. Keywords: Adenosine • Caffeine • Guanosine • Inosine • Tea ( Camellia sinensis) • Xanthosine. Abbreviations: DTT, dithiothreitol; IMP, inosine 5 ′- monophosphate; PCA, perchloric acid; PRPP, 5-phosphoribosyl- 1-pyrophosphate; TLC, thin-layer chromatography; XMP, xanthosine 5 ′-monophosphate. Introduction Many organisms, including plants, produce purine nucleotides by two distinct routes, the de novo pathway and also several salvage pathways (Moffatt and Ashihara 2002, Stasolla et al. 2003, Zrenner et al. 2006, Zrenner and Ashihara 2011). There is a de novo synthesis of purine nucleotides which utilizes 5-phosphoribosyl-1-pyrophosphate (PRPP), glutamine, glycine, aspartate, 10-formyltetrahydrofolate and HCO 3 − as building blocks, and is found in all plant species including leaves of tea (Ito and Ashihara 1999). In contrast to the de novo pathway, salvage pathways of nucleotide biosynthesis are more diverse and are less well understood (Sugiura and Takeda 2000). In some physiological circumstances the salvage pathways are stimulated but not the de novo pathways (Katahira and Ashihara 2006a). Purine salvage, degradation and secondary metabolic pathways derived from purine nucleotides appear to differ between plant species, and even between organs and tissues of the same species. Tea ( Camellia sinensis) is one of the most important eco- nomic plants used in beverages (Crozier et al. 2006). Tea plants produce unique nitrogen compounds such as caffeine and theanine. Many reports exist on the content of these com- pounds in commercial tea beverages. Caffeine typically com- prises 3–4 % of the dry weight of tea leaf products, and theanine 2–5 % (Crozier et al. 2006). The effect of these nitro- gen compounds on human health has been studied in detail (Weisburger 2004). Basic studies of the metabolism of nitrogen compounds in tea plants are relatively sparse, however. In the present work, we examined the metabolic fate of 10 distinct 14 C-labeled purine nucleosides and nucleobases in the segments of leaves and roots from 3-month-old tea seedlings. The data obtained from these in situ 14 C-tracer experiments were sup- plemented by in vitro measurements of the activity of some key enzymes relating to purine salvage in extracts from young tea leaves. We discuss the purine nucleotide recycling system and the major and minor pathways of purine metabolism in the leaves and roots of tea plants. Profiles of Purine Metabolism in Leaves and Roots of Camellia sinensis Seedlings Wei-Wei Deng and Hiroshi Ashihara* Department of Biological Sciences, Graduate School of Humanities and Sciences, Ochanomizu University, 2-1-1, Otsuka, Bunkyo-ku, Tokyo, 11-8610 Japan *Corresponding author: E-mail, [email protected]; Fax, +81-3-5978-5358 (Received October 13, 2010; Accepted November 2, 2010) Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175, available online at www.pcp.oxfordjournals.org © The Author 2010. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists. All rights reserved. For permissions, please email: [email protected] 2105 Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010. Regular Paper Downloaded from https://academic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 March 2022
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of Profiles of Purine Metabolism in Leaves and Roots of ...

To determine the metabolic profi les of purine nucleotides and related compounds in leaves and roots of tea ( Camellia
sinensis ), we studied the in situ metabolic fate of 10 different 14 C-labeled precursors in segments from tea seedlings. The activities of key enzymes in tea leaf extracts were also investigated. The rates of uptake of purine precursors were greater in leaf segments than in root segments. Adenine and adenosine were taken up more rapidly than other purine bases and nucleosides. Xanthosine was slowest. Some adenosine, guanosine and inosine was converted to nucleotides by adenosine kinase and inosine/guanosine kinase, but these compounds were easily hydrolyzed, and adenine, guanine and hypoxanthine were generated. These purine bases were salvaged by adenine phosphoribosyltransferase and hypoxanthine/guanine phosphoribosyltransferase. Salvage activity of adenine and adenosine was high, and they were converted exclusively to nucleotides. Inosine and hypoxanthine were salvaged to a lesser extent. In situ 14 C-tracer experiments revealed that xanthosine and xanthine were not salvaged, although xanthine phosphoribosyltransferase activity was found in tea extracts. Only some deoxyadenosine and deoxyguanosine was salvaged and utilized for DNA synthesis. However, most of these deoxynucleosides were hydrolyzed to adenine and guanine and then utilized for RNA synthesis. Purine alkaloid biosynthesis in leaves is much greater than in roots. In situ experiments indicate that adenosine, adenine, guanosine, guanine and inosine are better precursors than xanthosine, which is a direct precursor of a major pathway of caffeine biosynthesis. Based on these results, possible routes of purine metabolism are discussed.
Keywords: Adenosine • Caffeine • Guanosine • Inosine • Tea ( Camellia sinensis ) • Xanthosine .
Abbreviations : DTT , dithiothreitol ; IMP , inosine 5 ′ -monophosphate ; PCA , perchloric acid ; PRPP , 5-phosphoribosyl-1-pyrophosphate ; TLC , thin-layer chromatography ; XMP , xanthosine 5 ′ -monophosphate.
Introduction
Many organisms, including plants, produce purine nucleotides by two distinct routes, the de novo pathway and also several salvage pathways ( Moffatt and Ashihara 2002 , Stasolla et al. 2003 , Zrenner et al. 2006 , Zrenner and Ashihara 2011 ). There is a de novo synthesis of purine nucleotides which utilizes 5-phosphoribosyl-1-pyrophosphate (PRPP), glutamine, glycine, aspartate, 10-formyltetrahydrofolate and HCO 3 − as building blocks, and is found in all plant species including leaves of tea ( Ito and Ashihara 1999 ). In contrast to the de novo pathway, salvage pathways of nucleotide biosynthesis are more diverse and are less well understood ( Sugiura and Takeda 2000 ). In some physiological circumstances the salvage pathways are stimulated but not the de novo pathways ( Katahira and Ashihara 2006a ). Purine salvage, degradation and secondary metabolic pathways derived from purine nucleotides appear to differ between plant species, and even between organs and tissues of the same species.
Tea ( Camellia sinensis ) is one of the most important eco-nomic plants used in beverages ( Crozier et al. 2006 ). Tea plants produce unique nitrogen compounds such as caffeine and theanine. Many reports exist on the content of these com-pounds in commercial tea beverages. Caffeine typically com-prises 3–4 % of the dry weight of tea leaf products, and theanine 2–5 % ( Crozier et al. 2006 ). The effect of these nitro-gen compounds on human health has been studied in detail ( Weisburger 2004 ). Basic studies of the metabolism of nitrogen compounds in tea plants are relatively sparse, however. In the present work, we examined the metabolic fate of 10 distinct 14 C-labeled purine nucleosides and nucleobases in the segments of leaves and roots from 3-month-old tea seedlings. The data obtained from these in situ 14 C-tracer experiments were sup-plemented by in vitro measurements of the activity of some key enzymes relating to purine salvage in extracts from young tea leaves. We discuss the purine nucleotide recycling system and the major and minor pathways of purine metabolism in the leaves and roots of tea plants.
Profi les of Purine Metabolism in Leaves and Roots of Camellia sinensis Seedlings Wei-Wei Deng and Hiroshi Ashihara * Department of Biological Sciences, Graduate School of Humanities and Sciences, Ochanomizu University, 2-1-1, Otsuka, Bunkyo-ku, Tokyo, 11-8610 Japan *Corresponding author: E-mail, [email protected] ; Fax, + 81-3-5978-5358 (Received October 13, 2010; Accepted November 2, 2010)
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175, available online at www.pcp.oxfordjournals.org© The Author 2010. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
2105Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Regular PaperD
ownloaded from
https://academic.oup.com
/pcp/article/51/12/2105/1944539 by guest on 17 March 2022

Results
Uptake of 14 C-labeled purine compounds by seedling segments Total uptake of precursors by the seedling segments was calcu-lated by adding the radioactivity found in perchloric acid (PCA)-soluble and PCA-insoluble fractions and in CO 2 . In the case of deoxyribonucleoside metabolism, PCA was replaced by 80 % methanol containing an antioxidant (see Materials and Methods). Inspection of labeled metabolites in the incubation medium showed that there was negligible release of [ 14 C]metabolites from segments to the medium, at least during the 10 h incubation period. Purine nucleosides and bases were taken up at differing rates by the segments of leaves and the roots of tea seedlings ( Fig. 1 ). Uptake by leaves was generally faster than that by roots. In leaves and in roots, the uptake of some compounds, including adenine, adenosine and hypoxan-thine, was much faster than for xanthosine.
Metabolism of adenosine and adenine Adenosine and adenine are not intermediates of the de novo synthesis of purine nucleotides, but are catabolites of nucle-otides and nucleic acids ( Stasolla et al. 2003 , Zrenner et al. 2006 ,
0 2 4 6 8 10
Deoxyguanosine
Deoxyadenosine
Xanthosine
Xanthine
Inosine
Hypoxanthine
Guanosine
Guanine
Adenosine
Adenine
Leaf
Root
Uptake [nmol (100 mg FW)-1(10 h)-1]
Fig. 1 Uptake of [ 14 C]precursors by segments of leaves and roots from tea seedlings. Segments ( ∼ 100 mg FW) were incubated in the presence of labeled precursors in 2.0 ml of 30 mM potassium phosphate buffer (pH 5.6) containing 10 mM sucrose and 1 % sodium ascorbate for 10 h at 27 ° C in darkness. The total uptake of precursors by the segments was calculated by adding the radioactivity found in all cellular metabolites and CO 2 . Rates of uptake are expressed as nmol (100 mg FW) − 1 (10 h) − 1 ± SD.
A
B
Nucleotid
esRNA
Adenin
e
Adenosin
e
Xanth
osine
7-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Theophyll
ine
Xanth
ine
Allanto
inCO 2
0
20
40
60
80
G
A66%
G
A69%
Leaf
Root
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)
Nucleotid
esRNA
Adenin
e
Adenosin
e
Xanth
osine
7-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Theophyll
ine
Xanth
ine
Allanto
inCO 2
0
20
40
60
80
Leaf
Root
GA
67%
G
A81%
nd
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)
Fig. 2 Metabolic fate of [8- 14 C]adenosine (A) and [8- 14 C]adenine (B) in the segments of leaves and roots from tea seedlings. Segments ( ∼ 100 mg FW) were incubated in the presence of [8- 14 C]adenosine or [8- 14 C]adenine in 2.0 ml of 30 mM potassium phosphate buffer (pH 5.6) containing 10 mM sucrose and 1 % sodium ascorbate for 10 h at 27 ° C in darkness. Distributions of radioactivity are expressed as a percentage of the total radioactivity taken up by the samples ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) were as follows: (A) leaf, 12.9 ± 1.1; root, 2.1 ± 0.1; (B) leaf, 16.9 ± 1.3; root, 4.6 ± 0.8. The inset shows the percentage distribution of radioactivity in adenine (A) and guanine residues (G) of RNA.
Zrenner and Ashihara 2011 ). The metabolic fate of [8- 14 C]ade-nosine and [8- 14 C]adenine was studied using segments of leaves and roots, 10 h after the administration of labeled compounds ( Fig. 2A, B ). Values shown as bars are the percentage of total
2106
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

radioactivity taken up by the tissues; the values sum to 100 % . Absolute values of total radioactivity are shown in the fi gure legend.
Since only small amounts of [8- 14 C]adenosine and [8- 14 C]adenine were detected in the segments, exogenously supplied precursors must be rapidly metabolized to other compounds. Substantial amounts of radioactivity from [8- 14 C]adenosine and [8- 14 C]adenine were found in the salvage products: nucleotides and RNA. It follows that adenine and adenosine were converted to AMP and then entered the adenine nucle-otide pool comprising AMP, ADP and ATP. Some ATP was utilized for RNA synthesis. Only small differences were found between the metabolic fate of [8- 14 C]adenosine and [8- 14 C]adenine ( Fig. 2A, B ). When [8- 14 C]adenosine was used, 34 % of the radioactivity in RNA was recovered as guanine residues in leaves and 31 % in roots. Similar conversion was found when [8- 14 C]adenine was supplied. Consequently, an AMP → IMP → XMP → GMP pathway appears to be operating in tea seedlings. Radioactivity was also found in xanthosine,
7-methylxanthine, theobromine and caffeine. This suggests that XMP produced from AMP is utilized in the biosynthesis of purine alkaloids. Incorporation of radioactivity into theobro-mine and caffeine was signifi cantly greater in leaves than in roots. Much more radioactivity was found in theobromine than in caffeine. Although caffeine is the end-product, the precur-sors probably could not be transformed into caffeine during the incubation time. Smaller amounts of radioactivity were detected in theophylline, allantoin and CO 2 . Theophylline might be produced by the minor pathway of purine alkaloid biosynthesis, and allantoin and CO 2 were formed by the purine catabolic pathway starting from xanthine ( Ashihara et al. 2008 , Ashihara et. al. 2010a ). The allantoin fraction described herein contained small amounts of allantoic acid.
Fig. 3 shows the results of time-course studies of [8- 14 C]adenine. The data indicate that exogenously supplied adenine was rapidly metabolized in leaves and roots; the radioactivity was initially found in nucleotides (mainly adenine nucleotides), then in RNA and purine alkaloids including 7-methylxanthine,
0 6 12 18 240
20
40
60
80
100
0
2
4
6
8
10
12
14
16A
B
NucleotidesRNAAdenineXanthosine
Purine alkaloidsAllantoinCO2
Xanthine
Dis
tribu
tion
of ra
dioa
ctiv
ity (%
)
Ade
nine
, nu
cleo
tides
and
pur
ine
alka
loid
s
Oth
er m
etab
olite
s
Incubation time (h)
0 6 12 18 240
20
40
60
80
100
0
2
4
6
8
10
12
14
16
Nucleotides
RNA
Xanthine
Purine alkaloids
CO2
XanthosineOth
er m
etab
olite
s
Adenine
Allantoin
Ade
nine
, nu
cleo
tides
and
pur
ine
alka
loid
s
Incubation time (h)
Dis
tribu
tion
of ra
dioa
ctiv
ity (%
)
Fig. 3 Time-course studies of [8- 14 C]adenine in segments of leaves (A) and roots (B) from tea seedlings. Experimental conditions are the same as shown in the legend for Fig. 2 except for the incubation time and concentration of [ 14 C]precursors. The distribution of radioactivity is expressed as a percentage of the total radioactivity ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) after 1, 3, 6 and 24 h were, respectively, (A) leaf: 2.9 ± 0.2, 4.0 ± 0.1, 10.6 ± 0.3 and 32.2 ± 2.3; (B) root: 2.2 ± 0.2, 3.7 ± 0.1, 5.8 ± 0.1 and 12.3 ± 0.9.
2107
Profi les of purine metabolism in tea
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

theobromine, caffeine and theophylline. In leaves, more radio-activity was incorporated into RNA than into purine alkaloids until 6 h after incubation, so that nucleotides formed from adenine appear to be preferentially utilized for nucleic acid synthesis. Radioactivity in the purine alkaloids increased at 24 h, showing that formation of purine alkaloids is slower than RNA synthesis. Purine alkaloid synthesis was also detected in roots, but the rate of incorporation was lower than in leaves. Release of 14 CO 2 was observed in both leaves and roots.
Metabolism of guanosine and guanine Radioactivity from [8- 14 C]guanosine and [8- 14 C]guanine was also incorporated into purine nucleotides, alkaloids and catabolites ( Fig. 4A, B ). Salvage activity of guanosine and guanine is lower than that of adenosine and adenine. Considerable amounts of [8- 14 C]guanosine were converted to [8- 14 C]guanine, which accumulated in tea tissues. In contrast to 14 C-labeled adenosine and adenine, more radioactivity from [8- 14 C]guanine and [8- 14 C]guanosine was found in RNA than in nucleotides ( Fig. 4A, B ). This is probably because there is a smaller cellular pool size of guanine nucleotides than of ade-nine nucleotides ( Ashihara and Crozier 1999 , Koshiishi et al. 2001a ). Most of the radioactivity from [8- 14 C]guanosine and [8- 14 C]guanine was recovered in guanine residues, but recog-nizable amounts were also found in adenine residues of RNA. This contrasts with results from potato tubers, in which no adenine residues of RNA were labeled ( Katahira and Ashihara 2006b ). The discrepancy might be due to the longer incubation time (4 h for potatoes and 10 h for tea), but it is possible that this conversion is characteristic of tea metabolism. In bacteria and mammals, GMP is converted to IMP by the action of GMP reductase ( Mager and Magasanik 1960 , Stephens and Whittaker 1973 , Renart and Sillero 1974 ). No evidence for the presence of a GMP reductase in higher plants has yet been found, however ( Le Floc'h et al. 1982 ). Radioactivity from [8- 14 C]guanosine and [8- 14 C]guanine was incorporated into 7-methylxanthine, theobromine and caffeine. There was more radioactivity in these purine alkaloids in leaves than in roots. The radioactivity was also distributed in allantoin and CO 2 .
Metabolism of inosine and hypoxanthine Inosine and hypoxanthine are derived from IMP ( Stasolla et al. 2003 , Zrenner et al. 2006 ). IMP is both an intermediate of the de novo pathway and a catabolite of adenine nucleotides. Different patterns of inosine metabolism were found in leaves and roots ( Fig. 5A, B ). In leaves, 12 % of the radioactivity was found in the salvage products, 11 % was in purine alkaloids and nearly 70 % of the radioactivity was distributed in degrada-tion metabolites. It follows that almost 20 % of [8- 14 C]inosine was converted to IMP, and most IMP was further metabolized to ATP and GTP and then incorporated into RNA. The rest of the [8- 14 C]IMP was converted to XMP, and xanthosine was then formed. Xanthosine was utilized for the synthesis of theo-bromine and caffeine. Substantial amounts of inosine were hydrolyzed to allantoin and CO 2 . There are several possible
A
Nucleotid
esRNA
Guanin
e
Guanosin
e
3-Met
hylxan
thin
e
7-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Theophyll
ine
Allanto
in 2CO
0
20
40
60
80
G98%
G
85%
A
nd
Leaf
Root
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)B
Nucleotid
esRNA
Guanin
e
3-Met
hylxan
thin
e
7-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Theophyll
ine
Allanto
in 2CO
0
20
40
60
80
LeafG95%
G98% Root
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)
Fig. 4 Metabolic fate of [8- 14 C]guanosine (A) and [8- 14 C]guanine (B) in segments of leaves and roots from tea seedlings. The experimental conditions are the same as in Fig. 2 . Values are expressed as a percentage of the total radioactivity taken up by the samples ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) were as follows: (A) leaf, 2.9 ± 0.2; root, 1.0 ± 0.1; (B) leaf, 4.5 ± 0.4; root, 1.2 ± 0.1. The inset shows the percentage distribution of radioactivity in adenine (A) and guanine residues (G) of RNA.
routes of inosine catabolism, but the route starting inosine → hypoxanthine → xanthine is the most likely, since inosine nucleosidase activity was 12–25 times higher than the activity of the hypoxanthine and inosine salvage enzymes (see Table 1 ). Unlike in leaves, degradation of inosine was lim-ited in roots, and > 40 % of radioactivity was recovered as sal-vage products. The most radioactivity was recovered in RNA; 60 % of the radioactivity was distributed in guanine residues
2108
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

and 40 % in adenine residues. There was greater accumulation of inosine and xanthine in roots than in leaves.
Salvage of [8- 14 C]hypoxanthine for nucleotide and RNA synthesis was less than for [8- 14 C]inosine, especially in leaves ( Fig. 5B ). Only 4 % of total radioactivity was recovered in these salvage products in leaves, and 14 % in roots. In RNA, > 90 % of the radioactivity was found in guanine residues. High catabolic activity of [8- 14 C]hypoxanthine was observed in tea leaves;
A
B
Nucleotid
esRNA
Inosin
e
Hypoxa
nthin
e
Theobro
min
e
Caffe
ine
Xanth
ine
Allanto
inCO 2
0
20
40
60
80
G80%
A
Leaf
Root
AG
63%
Inco
rpor
atio
n (%
of
tota
l rad
ioac
tivi
ty)
Nucleotid
esRNA
Hypoxa
nthin
e
Xanth
osine
3-Met
hylxan
thin
e
7-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Theophyl
line
Xanth
ine
Allanto
inCO 2
0
20
40
60
80
LeafG
89%
G
91% Root
Inco
rpor
atio
n (%
of
tota
l rad
ioac
tivi
ty)
Fig. 5 Metabolic fate of [8- 14 C]inosine (A) and [8- 14 C]hypoxanthine (B) in segments of leaves and roots from tea seedlings. The experimental conditions are the same as in Fig. 2 . Values are expressed as a percentage of the total radioactivity taken up by the samples ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) were as follows: (A) leaf, 3.8 ± 0.2; root, 0.85 ± 0.06, (B) leaf, 7.8 ± 0.1; root, 1.8 ± 0.1. The inset shows the percentage distribution of radioactivity in adenine (A) and guanine residues (G) of RNA.
approximately 60 % of the radioactivity was released as 14 CO 2 . Only small amounts of purine alkaloids were produced from [8- 14 C]hypoxanthine.
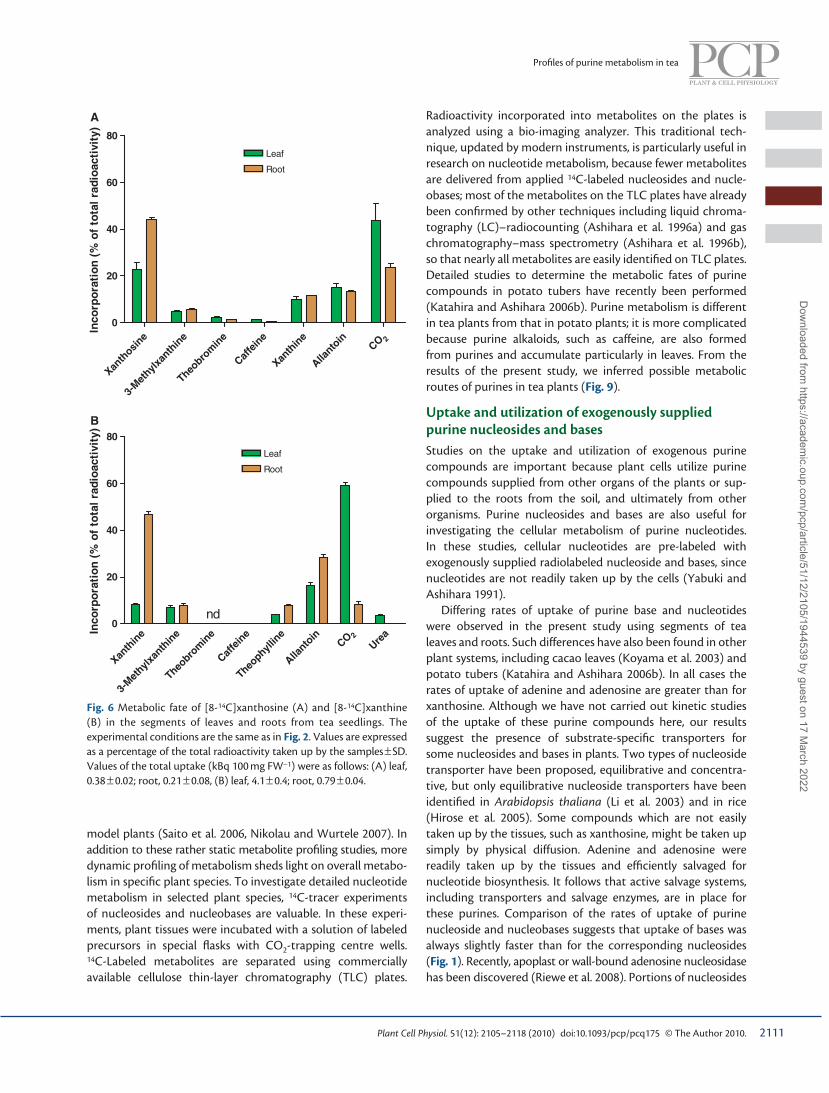
Metabolism of xanthosine and xanthine Xanthosine and xanthine are catabolites of purine nucleotides. Xanthosine is involved in the catabolic pathway of GMP, but AMP catabolism by-passes xanthosine ( Stasolla et al. 2003 ). Xanthosine is also a precursor of the main biosynthetic path-way of purine alkaloids in tea and coffee plants ( Ashihara et al. 2008 , Ashihara et al. 2010a , Ashihara et al. 2011 ). No salvage products were labeled from [8- 14 C]xanthosine ( Fig. 6A ). When [8- 14 C]xanthosine was supplied, a substantial amount of radio-activity was found in xanthosine and xanthine. Considerable amounts of 14 C from [8- 14 C]xanthosine were recovered as allantoin and CO 2 . Less than 4 % of the radioactivity from [8- 14 C]xanthosine was found in theobromine and caffeine in leaves. This incorporation rate was much lower than the rate found in [8- 14 C]adenosine-administered leaves, in which 26 % of radioactivity was observed in these purine alkaloids.
More than 90 % of [8- 14 C]xanthine taken up by the leaf segments was metabolized in leaves ( Fig. 6B ), but nearly half (47 % ) of the [8- 14 C]xanthine in the roots remained unaltered. Radioactivity from [8- 14 C]xanthine was incorporated into 3-methylxanthine and theophylline, but theobromine and caf-feine were not found to be labeled. As with [8- 14 C]xanthosine, incorporation of 14 C into CO 2 was greater in leaves than in roots. Radioactivity was also found in allantoin. To confi rm the metabolic fate of xanthine, we performed time-course studies of [8- 14 C]xanthine in leaves and roots ( Fig. 7A, B ). The radioac-tivity was found in allantoin and 3-methylxanthine at 1 and 3 h after administration of labeled compounds. Distribution in these compounds fell sharply with time in leaves, but remained almost constant in roots. Release of 14 CO 2 from [8- 14 C]xanthine was markedly increased at 6 h after administration in both leaves and roots. [8- 14 C]Xanthine was metabolized much faster in leaves than in roots. In leaves alone, up to 8 % of the radioac-tivity was observed in urea ( Fig. 7A ). This metabolite was not detected in roots ( Fig. 7B ).
Negligible amounts of 14 C were incorporated into theobro-mine and caffeine in leaves and roots. Most [8- 14 C]xanthine was catabolized via the purine catabolic pathway.
Metabolism of deoxyadenosine and deoxyguanosine The purine deoxynucleosides, deoxyadenosine and deox-yguanosine, are produced as catabolites of DNA. The concen-trations of these nucleosides appeared to be much lower than those of purine ribonucleosides in plant cells ( Nygaard 1972 ), because DNA turnover is slower than RNA turnover. Nearly 90 % of [8- 14 C]deoxyadenosine taken up by the leaf and root segments was metabolized, and almost 70 % of the total radio-activity was found in the salvage products (nucleotides and nucleic acids); 9 % of the radioactivity was located in caffeine and 1 % in theophylline in leaves. It was diffi cult to estimate the incorporation of radioactivity into theobromine, because
2109
Profi les of purine metabolism in tea
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

it was diffi cult to separate theobromine from deoxyadenosine. Some radioactivity in deoxyadenosine might therefore be due to [ 14 C]theobromine contamination. The rest of the radioactiv-ity was recovered as the degradation products, allantoin and CO 2 ( Fig. 8A, B ). We did not distinguish incorporation of 14 C into individual nucleotides using our methods, but the fraction may contain both ribo- and deoxyribonucleotides. Limited amounts of deoxyadenosine were salvaged to deoxyadenine nucleotides and utilized for DNA synthesis, because only part of the radioactivity from [8- 14 C]deoxyadenosine was found in DNA. More radioactivity was distributed in RNA. Approximately 66–68 % of the radioactivity in RNA from [8- 14 C]deoxyadenosine was recovered as adenine residues, and the rest ( ∼ 33 % ) as guanine residues. This pattern is similar to the results obtained with [8- 14 C]adenosine. Metabolism of [8- 14 C]deoxyguanosine was slow, and > 38 % of the radioactivity was found in unmetabolized deoxyguanosine in leaves and 66 % in unmetabolized deoxyguanosine in roots. Relative to deoxyadenosine there was low incorporation into nucleotides and nucleic acids. The radioactivity in RNA was distributed only to guanine residues. In leaves, approximately 26 % of the total radioactivity was recovered as purine alkaloids.
Activity of enzymes involved in purine salvage and nucleoside hydrolysis Table 1 shows the in vitro activity of enzymes of purine salvage and nucleoside hydrolysis, measured in protein extracts from young tea leaves. For maximum catalytic activity of enzymes in tissue extracts, the tissues were disrupted completely using extraction buffer, and the degree of disruption was checked
using a microscope. Consequently, the extracts contained enzymes distributed in the stroma of chloroplasts and the matrix of mitochondria. In preliminary experiments we did not detect any of the enzyme activities listed in Table 1 in the pellet that resulted after centrifuging at 30,000 × g .
Purine nucleoside is salvaged by two kinases, adenosine kinase and inosine/guanosine kinase. The activity of adenosine kinase was 100–250 times higher than that of inosine/guanosine kinase. No xanthosine kinase activity was observed. The nucleo-side is hydrolyzed to nucleobases and salvage. Three enzymes that are involved in purine nucleoside hydrolysis are adenine nucleosidase, inosine/guanosine nucleosidase and purine (xanthosine) nucleosidase. Adenosine hydrolysis activity was nearly 10 times greater than that of inosine/guanosine nucleo-sidase. There was relatively high xanthosine hydrolyzing activ-ity. There are two known purine base salvaging reactions in higher plants. Adenine phosphoribosyltransferase has the greatest activity of the enzymes studied here. Hypoxanthine/guanine phosphoribosyltransferase activity was low. There was xanthine phosphoribosyltransferase activity. This is the fi rst report to fi nd this enzyme in plants, although the activity was very low. In Lactobacillus casei , a distinct xanthine phosphori-bosyltransferase has been observed ( Krenitsky et al. 1970 ).
Discussion
Dynamic profi ling of purine metabolism Systematic studies of metabolites (referred to as metabolomics) have recently been performed in several useful plants, as well as
Table 1 Profi le of activity of enzymes involved in purine salvage and nucleoside hydrolysis in tea leaf extracts
Reaction Enzyme (EC) Enzyme activity
Nucleoside salvage
Adenosine a + ATP → AMP a + ADP Adenosine kinase (2.7.1.20) 34.7 ± 1.0
Guanosine a + ATP → GMP a + ADP Inosine/guanosine kinase (2.7.1.73) 0.13 ± 0.02
Inosine a + ATP → IMP a + ADP Inosine/guanosine kinase (2.7.1.73) 0.37 ± 0.01
Xanthosine a + ATP → XMP a + ADP Inosine/guanosine kinase (2.7.1.73) ND
Nucleoside hydrolysis
Adenosine a → Adenine a + Ribose Adenosine nucleosidase (3.2.2.7) 84.9 ± 0.6
Guanosine a → Guanine a + Ribose Inosine/guanosine nucleosidase (3.2.2.2) 8.53 ± 0.08
Inosine a → Hypoxanthine a + Ribose Inosine/guanosine nucleosidase (3.2.2.2) 9.21 ± 0.74
Xanthosine a → Xanthine a + Ribose Purine nucleosidase (3.2.2.1) 29.1 ± 0.4
Nucleobase salvage
Adenine a + PRPP → AMP a + PPi Adenine phosphoribosyltransferase (2.4.2.7) 96.9 ± 2.2
Guanine a + PRPP → GMP* + PPi Hypoxanthine/guanine phosphoribosyltransferase (2.4.2.8) 1.26 ± 0.01
Hypoxanthine a + PRPP → IMP a + PPi Hypoxanthine/guanine phosphoribosyltransferase (2.4.2.8) 0.79 ± 0.19
Xanthine a + PRPP → XMP a + PPi Xanthine phosphoribosyltransferase (2.4.2.22) 0.22 ± 0.04
Activities are expressed as pkat (mg protein) − 1 a Labeled substrates and products. ND, no enzyme activity detected.
2110
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022
model plants ( Saito et al. 2006 , Nikolau and Wurtele 2007 ). In addition to these rather static metabolite profi ling studies, more dynamic profi ling of metabolism sheds light on overall metabo-lism in specifi c plant species. To investigate detailed nucleotide metabolism in selected plant species, 14 C-tracer experiments of nucleosides and nucleobases are valuable. In these experi-ments, plant tissues were incubated with a solution of labeled precursors in special fl asks with CO 2 -trapping centre wells. 14 C-Labeled metabolites are separated using commercially available cellulose thin-layer chromatography (TLC) plates.
A
B
Xanth
osine
3-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Xanth
ine
Allanto
inCO 2
0
20
40
60
80
Leaf
Root
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)
Xanth
ine
3-Met
hylxan
thin
e
Theobro
min
e
Caffe
ine
Theophyll
ine
Allanto
in 2CO
Urea
0
20
40
60
80
Leaf
nd
Root
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)
Fig. 6 Metabolic fate of [8- 14 C]xanthosine (A) and [8- 14 C]xanthine (B) in the segments of leaves and roots from tea seedlings. The experimental conditions are the same as in Fig. 2 . Values are expressed as a percentage of the total radioactivity taken up by the samples ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) were as follows: (A) leaf, 0.38 ± 0.02; root, 0.21 ± 0.08, (B) leaf, 4.1 ± 0.4; root, 0.79 ± 0.04.
Radioactivity incorporated into metabolites on the plates is analyzed using a bio-imaging analyzer. This traditional tech-nique, updated by modern instruments, is particularly useful in research on nucleotide metabolism, because fewer metabolites are delivered from applied 14 C-labeled nucleosides and nucle-obases; most of the metabolites on the TLC plates have already been confi rmed by other techniques including liquid chroma-tography (LC)–radiocounting ( Ashihara et al. 1996a ) and gas chromatography–mass spectrometry ( Ashihara et al. 1996b ), so that nearly all metabolites are easily identifi ed on TLC plates. Detailed studies to determine the metabolic fates of purine compounds in potato tubers have recently been performed ( Katahira and Ashihara 2006b ). Purine metabolism is different in tea plants from that in potato plants; it is more complicated because purine alkaloids, such as caffeine, are also formed from purines and accumulate particularly in leaves. From the results of the present study, we inferred possible metabolic routes of purines in tea plants ( Fig. 9 ).
Uptake and utilization of exogenously supplied purine nucleosides and bases Studies on the uptake and utilization of exogenous purine compounds are important because plant cells utilize purine compounds supplied from other organs of the plants or sup-plied to the roots from the soil, and ultimately from other organisms. Purine nucleosides and bases are also useful for investigating the cellular metabolism of purine nucleotides. In these studies, cellular nucleotides are pre-labeled with exogenously supplied radiolabeled nucleoside and bases, since nucleotides are not readily taken up by the cells ( Yabuki and Ashihara 1991 ).
Differing rates of uptake of purine base and nucleotides were observed in the present study using segments of tea leaves and roots. Such differences have also been found in other plant systems, including cacao leaves ( Koyama et al. 2003 ) and potato tubers ( Katahira and Ashihara 2006b ). In all cases the rates of uptake of adenine and adenosine are greater than for xanthosine. Although we have not carried out kinetic studies of the uptake of these purine compounds here, our results suggest the presence of substrate-specifi c transporters for some nucleosides and bases in plants. Two types of nucleoside transporter have been proposed, equilibrative and concentra-tive, but only equilibrative nucleoside transporters have been identifi ed in Arabidopsis thaliana ( Li et al. 2003 ) and in rice ( Hirose et al. 2005 ). Some compounds which are not easily taken up by the tissues, such as xanthosine, might be taken up simply by physical diffusion. Adenine and adenosine were readily taken up by the tissues and effi ciently salvaged for nucleotide biosynthesis. It follows that active salvage systems, including transporters and salvage enzymes, are in place for these purines. Comparison of the rates of uptake of purine nucleoside and nucleobases suggests that uptake of bases was always slightly faster than for the corresponding nucleosides ( Fig. 1 ). Recently, apoplast or wall-bound adenosine nucleosidase has been discovered ( Riewe et al. 2008 ). Portions of nucleosides
2111
Profi les of purine metabolism in tea
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

applied may therefore be hydrolyzed to bases outside cells, and then taken up by the cells. The transport systems of other nucleoside and bases remain undetermined.
Table 1 shows that high activity of adenosine kinase (step 1 in Fig. 9 ) and adenine phosphoribosyltransferase (step 3) was found in the enzyme extracts, so that adenosine and adenine were utilized for AMP synthesis by these salvage enzymes in tea plants. Even if adenosine is taken up by the cells, it may be also hydrolyzed to adenine within the cells by adenosine nucleosidase (step 2) and then salvaged by adenine phosphori-bosyltransferase (step 3), because a high activity of adenosine nucleosidase was observed as a water-soluble protein in tea extracts ( Table 1 ). The diversity of adenosine salvage in plant species is striking. Only the adenosine kinase route (step 1) is operative in white spruce embryos, because no adenosine nucleosidase activity was detected ( Ashihara et al. 2001 ). In contrast, the adenosine nucleosidase and adenine phospho-ribosyltransferase route (steps 2 and 3) appears to be responsi-ble for adenosine salvage in shoots of Helianthus tuberosus ,
in which no adenosine kinase activity was observed ( Le Floc'h and Lafl euriel 1981 ). Deoxyadenosine appears to be converted to deoxyadenosine 5 ′ -monophosphates by deoxyadenosine kinase (step 4), but the amounts were limited. Deoxyadenosine is hydrolyzed possibly by adenosine nucleosidase (step 2), and the resulting adenine is salvaged by adenine phosphoribosyl-transferase (step 3).
In many organisms, inosine and guanosine are salvaged by a single enzyme, inosine/guanosine kinase (step 6), whereas hypoxanthine and guanine are salvaged by a hypoxanthine/guanine phosphoribosyltransferase (step 7). In plants, especially tea plants, the activity of adenine phosphoribosyltransferase and adenosine kinase is always much greater than the activity of inosine/guanosine kinase and hypoxanthine/guanine phos-phoribosyltransferase. As with deoxyadenosine, a very small amount of deoxyguanosine is converted to deoxyguanosine 5 ′ -monophosphates by deoxyguanosine kinase (step 5). Inosine and hypoxanthine are converted to IMP by salvage enzymes. IMP is a branch point of AMP and GMP synthesis, but IMP
A
B
0 6 12 18 240
20
40
60
80
100
Xanthine3-Methylxanthne
Other purine alkaloids
Allantoin
0
5
10
15
20
Dis
tribu
tion
of ra
dioa
ctiv
ity (%
)
Xan
thin
e, A
llant
oin
and
CO
2
Urea
Oth
er m
etab
olite
s
CO2
Incubation time (h)
0 6 12 18 240
20
40
60
80
100
Xanthine3-MethylxanthineOther purine alkaloids
Allantoin
0
5
10
15
20
Dis
tribu
tion
of ra
dioa
ctiv
ity (%
)
Xan
thin
e, A
llant
oin
and
CO
2
Oth
er m
etab
olite
s
CO2
Incubation time (h)
Fig. 7 Time-course studies of [8- 14 C]xanthine in segments of leaves (A) and roots (B) from tea seedlings. The experimental conditions are the same as in Fig. 3 . The distribution of radioactivity is expressed as a percentage of the total radioactivity ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) after 1, 3, 6 and 24 h were, respectively, (A) leaf: 0.67 ± 0.02, 1.4 ± 0.1, 5.9 ± 0.7 and 16.2 ± 1.1; (B) root: 0.77 ± 0.08, 0.81 ± 0.05, 1.6 ± 0.1 and 3.0 ± 0.2.
2112
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

synthesized from inosine and hypoxanthine was incorporated preferentially into guanine residues of RNA. This may be because the pool size of free GTP is smaller than that of ATP ( Ashihara and Crozier 1999 , Koshiishi et al. 2001a ). In fact, the salvage enzymes for inosine and hypoxanthine, i.e. inosine/guanosine kinase and hypoxanthine/guanine phosphoribosyl-transfease, are the same enzymes. If synthetic pathways for guanine nucleotides and adenine nucleotides are located
A
B
Nucleotid
esRNA
DNA
Adenin
e
Deoxy
aden
osine
Caffe
ine
Theophyll
ine
Allanto
in 2CO
0
20
40
60
80
AG
66%
AG
68% Root
Leaf
Inco
rpo
rati
on
(%
of
tota
l ra
dio
acti
vity
)
Nucleotid
esRNA
DNA
Guanin
e
Deoxy
guanosin
e
Theobro
min
e
Caffe
ine
Allanto
in 2CO
0
20
40
60
80
RootLeafGG GGG
Inco
rpo
rati
on (
% o
f to
tal
rad
ioac
tivi
ty)
Fig. 8 Metabolic fate of [8- 14 C]deoxyadenosine (A) and [8- 14 C]deoxyguanosine (B) in segments of leaves and roots from tea seedlings. The experimental conditions are the same as in Fig. 2 . Values are expressed as a percentage of the total radioactivity taken up by the samples ± SD. Values of the total uptake (kBq 100 mg FW − 1 ) were as follows: (A) leaf, 5.1 ± 1.0; root, 0.61 ± 0.01, (B) leaf, 1.8 ± 0.2; root, 1.3 ± 0.1. The inset shows the percentage distribution of radioactivity in adenine (A) and guanine residues (G) of RNA.
in different compartments, the salvage system for inosine and hypoxanthine may be associated with the guanine nucleotide-synthesizing compartment, which is distinct from the routes for adenine nucleotide synthesis. A study using A. thaliana indicated that the cDNAs encoding the enzymes involved in AMP synthesis contain likely plastid transit sequences, but the sequences of genes for GMP synthesis do not contain N-terminal transit peptides ( van der Graaff 2004 ). In tea leaves, a portion of the activities of adenine phosphoribosyltransferase, adenosine kinase and adenosine nucleosidase was located in chloroplasts, but > 60 % of activity was distributed in the cytosol ( Koshiishi et al. 2001b ). Localization of adenine phosphoribosyl-transferase and inosine/guanosine kinase in mitochondria has been reported ( Hirose and Ashihara 1982 , Le Floc'h and Lafl euriel 1983 , Ukaji and Ashihara 1986 , Combes et al. 1989 ). Nuclear localization of adenine phosphoribosyltransferase has also been suggested ( Allen et al. 2002 ). It follows that enzymes of purine salvage and interconversion are distributed in several compartments of plant cells. Detailed studies into these phenomena are needed.
Unlike in animals, hydrolysis of nucleosides by phosphory-lases is not detectable in plants; instead, at least three different purine nucleoside hydrolytic activities are demon-strated ( Guranowski 1982 , Stasolla et al. 2003 ): adenosine nucleosidase (step 2), inosine/guanosine nucleosidase (step 8) and purine (xanthosine) nucleosidase (step 9), as shown in Table 1 . Purine nucleosides are therefore readily hydrolyzed without phosphate, and produce the corresponding bases. In in situ tracer experiments, no evidence of xanthosine and xanthine salvage was found ( Fig. 6A, B ), although low activity of xanthine phosphoribosyltransferase was detected in tea extracts ( Table 1 ). This tells us that purine structures cannot be salvaged for nucleotide synthesis if they are converted to xanthine (step 17a) or xanthosine (step 10).
Catabolism of nucleotides in plants and the route of recycling Adenine nucleotides appear to be degraded to adenosine and adenine (steps 10 and 2), but these compounds are directly utilized for the regeneration of adenine nucleotides (steps 1 and 3). Unlike in bacteria and animals, no adenosine deaminase or adenine deaminase is present in plants ( Stasolla et al. 2003 , Zrenner et al. 2006 ), and deamination of adenine molecules is performed by AMP deaminase ( Yabuki and Ashihara 1991 , Koshiishi et al. 2001b ) (step 11). The product, IMP, is converted to inosine by 5 ′ -nucleotidase and/or various phosphatases (step 10). Although some inosine is salvaged to IMP synthesis (step 6), the rest is changed to hypoxanthine (step 8). Hypoxanthine can be salvaged by hypoxanthine/guanine phosphoribosyltransferase (step 7), but is mostly oxidized to xanthine and then uric acid by xanthine dehydrogenase (step 17); these catabolites are then no longer available for nucleotide synthesis. Guanine nucleotides are catabolized, and guanosine and guanine are formed (steps 10 and 8). These catabolites can be utilized for nucleotide synthesis.
2113
Profi les of purine metabolism in tea
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

In contrast to AMP, GMP is not easily converted to IMP (step 14), although the in situ experiments suggested this conversion, because adenine residues of RNA were labeled when [8- 14 C]guanosine and [8- 14 C]guanine were administered. Guanosine deaminase (step 15) and guanine deaminase activity (step 16) has been demonstrated in tea extracts ( Negishi et al. 1994 ). As shown below, some xanthosine and xanthine is utilized for purine alkaloid synthesis, but the rest is catabolized by the purine catabolic pathway (steps 17–21).
Adenosine, adenine, inosine, hypoxanthine, guanosine and guanine produced as degradation products in the cells can be recycled for nucleotide synthesis. Our present results suggest that tea plants have an effi cient salvage system of adenine and
adenosine relative to other purine nucleosides and bases. Inosine is located in positions which permit the direction of metabolism, i.e. salvage and catabolism. A clear difference in the metabolic fate of [8- 14 C]inosine was found in roots and leaves. Inosine salvage (step 6) is greater in roots, whereas catabolism (steps 8 and 17–21) is greater in leaves.
Details of the purine catabolic pathway cannot be evaluated in the present experiments, because only [8- 14 C]precursors were used. Alternative routes have been proposed for allantoic acid degradation in higher plants ( Winkler et al. 1988 , Schubert and Boland 1990 ). However, our results suggest that the tradi-tional pathway including allantoicase is operative. In this path-way, the radioactivity from [8- 14 C]purines appears to be located
Fig. 9 Metabolic pathways of purine compounds in leaves and roots of tea plants inferred from the fate of exogenously supplied [8- 14 C]purine nucleosides and nucleobases. Reactions and enzymes involved in purine salvage, degradation and alkaloid synthesis are shown with red, blue and green arrows and numbers, respectively. Numbers indicate the reaction steps catalyzed by the enzymes shown below. 1 , Adenosine kinase (EC 2.7.1.20); 2 , adenosine nucleosidase (3.2.2.7); 3 , adenine phosphoribosyltransferase (2.4.2.7); 4 , deoxyadenosine kinase (2.7.1.76); 5 , deoxyguanosine kinase (2.7.1.113); 6 , inosine/guanosine kinase (2.7.1.73); 7 , hypoxanthine/guanine phosphoribosyltransferase (2.4.2.8); 8 , inosine/guanosine nucleosidase (3.2.2.2); 9 , purine (xanthosine) nucleosidase (3.2.2.1); 10 , 5 ′ -nucleotidase (3.1.3.5) or various phosphatases (3.1.3.-); 11 , AMP deaminase (3.5.4.6); 12 , IMP dehydrogenase (1.1.1.205); 13 , GMP synthase (6.3.5.2); 14 , GMP reductase (1.7.1.7) (this enzyme has not yet been discovered in any plants); 15 , guanosine deaminase (3.5.4.15); 16 , guanine deaminase (3.5.4.3); 17 , xanthine dehydrogenase (1.1.1.204); 18 , uricase (1.7.3.3); 19 , allantoinase (3.5.2.5); 20 , allantoicase (3.5.3.4); 21 , ureidoglycolate lyase (4.3.2.3); 22 , urease (3.5.1.5); 23a , 7-methylxanthosine synthase (2.1.1.158); 23b , N -methylnucleosidase (3.2.2.25); 24a–d , caffeine synthase (2.1.1.160); 24a, c , theobromine synthase (2.1.1.159) (the gene for this enzyme has not yet been found in tea plants).
2114
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

in urea, when allantoic acid is degraded to urea and ureidogly-colate (step 20). Ureidoglycolate is further degraded to glyoxylate and urea (step 21). The resulting [ 14 C]urea is further hydrolyzed to ammonia and 14 CO 2 by urease (step 22). In this study we found the radioactivity in allantoin, urea and CO 2 , so that this pathway is operative and is the main route for purine catabolism in tea plants. Catabolic activity of purines in leaves is greater than in roots, because release of 14 CO 2 from [8- 14 C]xanthine was considerably higher in leaves.
Biosynthesis of purine alkaloids in tea leaves and roots Purine alkaloid biosynthesis in leaves is much greater than in roots. The radioactivity in theobromine is greater than in caffeine 10 h after administration of [ 14 C]precursors, but the proportion in caffeine increased after longer incubation. Consequently, the production of caffeine is time-consuming, and conversion of theobromine to caffeine (step 24b) is slow. Conversion of 7-methylxanthine to caffeine (steps 24a and 24b) appears to be catalyzed by dual-functional caffeine synthase ( Kato et al. 1999 , Kato et al. 2000 ). N 1 methylation of the xanthine structure by this enzyme is much slower than N 3 methylation.
It has been established that the major caffeine biosynthetic pathway is xanthosine → 7-methyxanthosine → 7-methylxanthine → theobromine → caffeine (steps 23a–24b). Methylation of xanthosine is catalyzed by 7-methylxanthosine synthase (step 23a), and 7-methylxanthine was formed by N -methylnu-cleosidase (step 23b) ( Negishi et al. 1988 ). However, structural studies on 7-methylxanthosine synthase in coffee plants suggested that the methyl transfer and nucleoside cleavage (steps 23a and 23b) are coupled and both are catalyzed by a single enzyme ( McCarthy and McCarthy 2007 ). The steps from 7-methylxanthine to caffeine (steps 24a and 24b) are catalyzed by dual-functional caffeine synthase ( Kato et al. 1999 , Kato et al. 2000 ), but a separate enzyme, theobromine synthase (step 24a), has also been found in some plants (see Ashihara et al. 2008 , Ashihara et al. 2010b ).
In situ experiments indicate that adenosine, adenine, guanosine, guanine, inosine and hypoxanthine are precursors of caffeine biosynthesis. Xanthosine, the precursor which is com-mitted to caffeine biosynthesis, is produced by several path-ways. Adenine and adenosine are converted to AMP, and IMP is then produced by AMP deaminase (steps 1–3 and 11). Inosine and hypoxanthine are converted to IMP (steps 6–7) and joined to the caffeine biosynthetic pathway. XMP is produced from IMP by IMP dehydrogenase (step 12), and xanthosine is formed by 5 ′ -nucleotidase (step 10). The amino group of guanosine is released by guanosine deaminase, and xanthosine is formed (step 15). Guanine appears to be salvaged to GMP (step 7), then guanosine is produced by 5 ′ -nucleotidase (step 10).
In situ 14 C-tracer experiments indicated that exogenously supplied 14 C-labeled adenosine, adenine, guanosine, guanine and inosine are better precursors of caffeine than [ 14 C]xanthosine, which is a direct precursor of the major pathway of caffeine
biosynthesis ( Ashihara et al. 2008 ). This is probably because most of the exogenously supplied xanthosine is hydrolyzed to xanthine, and most xanthine molecules enter the purine catabolic pathway. Time-course studies of [8- 14 C]xanthine indicate that some xanthine is used for theophylline biosynthe-sis by the xanthine → 3-methylxanthine → theophylline path-way (steps 24c and 24d). Since small amounts of radioactivity from other precursors were also detected in theophylline, this route may act as an alternative pathway for purine alka-loids. Large amounts of theophylline were not present in tea leaves, however ( Ashihara and Kubota 1986 ), so that this pathway is only a minor route of purine alkaloid biosynthesis in tea.
Some differences emerge in purine metabolism between tea leaves and roots. Purine metabolism in leaves is characterized as higher purine alkaloid synthesis and higher purine catabolism, whereas purine salvage is predominant in roots. The physiologi-cal signifi cance of these differences is unknown, but perhaps caffeine accumulation in young leaves and shoots acts as a chemical defense to protect young soft tissue from pathogens and herbivores ( Ashihara et al. 2008 ). The turnover of purine nucleotides which produce purine catabolites appears to be faster in leaves than in roots. The end-products of purine compounds, NH 3 , CO 2 and glyoxylic acid, are readily utilized for biosynthesis in leaves. In some cases, more radioactivity was located in allantoin in roots than in leaves. Allantoin produced from purines in roots might act to transport nitrogen to leaves from roots, as occurs in some leguminous plants ( Thomas and Schrader 1981 ).
Materials and Methods
Plant materials Seeds of tea ( C. sinensis , cv. Yabukita) were supplied in November 2008 from the Makurazaki Tea Research Station, National Institute of Vegetable and Tea Science, Japan. The seeds were stored in a cold room (4 ° C) for 1 month. Dormancy-broken seeds were sterilized with sodium hypochlorite and the seed coats were removed. They were then cultured in vermiculite for 3 months without additional nutrients. Culture conditions were the same as in a previous study ( Ashihara et al. 2010b ). In this study we used seedlings having three leaves, and a main root with some lateral roots. Leaves ( ∼ 130 mg FW, 40 mm long and 30 mm wide) and roots ( ∼ 200 mg FW, 50 mm long) were separated and used as experimental materials.
Radiochemicals The following radiochemicals were obtained from Moravek Biochemicals Inc.: [8- 14 C]adenine (specifi c activity, 1.9 GBq mmol − 1 ), [8- 14 C]adenosine (2.0 GBq mmol − 1 ), [8- 14 C]guanine (2.0 GBq mmol − 1 ), [8- 14 C]guanosine (1.9 GBq mmol − 1 ), [8- 14 C]hypoxanthine (2.1 GBq mmol − 1 ), [8- 14 C]inosine (1.9 GBq mmol − 1 ), [8- 14 C]xanthine (2.0 GBq mmol − 1 ), [8- 14 C]xanthosine
2115
Profi les of purine metabolism in tea
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

(2.1 GBq mmol − 1 ), [8- 14 C]deoxyadenosine (2.0 GBq mmol − 1 ) and [8- 14 C]deoxyguanosine (1.1 GBq mmol − 1 ).
Administration of labeled precursors The labeled compounds were administered essentially as described in previous studies ( Ashihara et al. 1996a , Katahira and Ashihara 2006b ). In brief, segments of leaves and roots were sterilized with 1 % sodium hypochlorite solution (Wako Pure Chemical Industries, Ltd.) for 10 min. The samples were washed with sterilized distilled water, and segments of leaves ( ∼ 4 mm x 4 mm disks) and roots ( ∼ 2 mm sections) were prepared asepti-cally. Sample segments ( ∼ 100 mg FW), and 2.0 ml of 30 mM potassium phosphate buffer (pH 5.6) containing 10 mM sucrose and 1 % sodium ascorbate, were placed in the main compart-ment of a 30 ml Erlenmeyer fl ask fi tted with a glass tube con-taining a piece of fi lter paper impregnated with 0.1 ml of 20 % KOH in the center well. Each reaction was started by adding 18.5 kBq (10 h incubation) or 37.0 kBq (time-course studies) of one of the radioactive purine compounds. The fl asks were incubated in an oscillating water bath at 27 ° C in the dark. After incubation, the glass tube was removed from the center well and placed in a 50 ml Erlenmeyer fl ask containing 10 ml of distilled water. At the same time, the segments of leaves and roots were separated from the incubation medium by fi ltering through a tea strainer; radioactive compounds attached to the surface of segments were removed completely with approxi-mately 50 ml of running tap water. The samples were washed with distilled water and then frozen with liquid N 2 and stored at − 80 ° C until extraction. The KH 14 CO 3 that had been absorbed by the fi lter paper was allowed to diffuse overnight into the distilled water, and 0.5 ml aliquots of the resulting solution were used for the determination of radioactivity incorporated into CO 2 . Aliquots of the incubation medium were concen-trated and the metabolites were analyzed by TLC as described below. Radioactivity was measured with a liquid scintillation counter (Beckman, LS 6500) using ACS-II scintillation fl uid (GE Healthcare Japan Ltd.).
Analysis of radiolabeled metabolites The labeled metabolites were analyzed by the methods described in our previous papers ( Katahira and Ashihara 2002 , Katahira and Ashihara 2006b ). Metabolites from 14 C-labeled purine nucleosides and nucleobases were extracted successively with 4 ml of cold 6 % PCA solution containing 0.1 mM AMP (twice). Lipids in the insoluble materials were removed with a mixture of ethanol and diethylether (1 : 1, v/v) at 50 ° C for 15 min. The precipitate was washed with ethanol and diethyle-ther (1 : 1, v/v), then with distilled water. For [8- 14 C]deoxyade-nosine and [8- 14 C]deoxyguanosine, cold 6 % PCA was replaced by 80 % methanol containing 20 mM sodium diethyldithiocar-bamate, since these compounds are easily hydrolyzed to purine bases by 6 % PCA ( Katahira and Ashihara 2006b ). RNA in the fraction insoluble in the ethanol and diethylether mixture was hydrolyzed with 0.3 M KOH at 37 ° C for 18 h, and was adjusted to pH 2 with PCA. After the mixture had been centrifuged
at 10,000 × g for 5 min, the supernatant (RNA hydrolysates) was collected and the DNA in the precipitate was hydrolyzed with 6 % PCA at 100 ° C for 20 min. In some experiments, RNA and DNA were simultaneously hydrolyzed with 6 % PCA at 100 ° C for 20 min. The PCA-soluble metabolites and hydro-lysates of nucleic acids were neutralized with KOH; radioactive metabolites were then separated by TLC using cellulose plates (Merck) and the following solvent systems: system I, n -butanol/acetic acid/water (4/1/2, by vol.); system II, ethyl acetate/methanol/water (100/13.5/10, by vol.); system III, distilled water. Radioactivity of liquid samples was determined using a multipurpose scintillation counter. Radioactivity of 14 C on the TLC plates was analyzed using a bio-imaging analyzer (Type FLA-2000, Fuji Photo Film Co. Ltd.).
Enzyme preparation For the determination of enzyme activity, young leaves were obtained from fl ush shoots from trees of C. sinensis (cv. Saemidori) in March 2010 at an estate of the Makurazaki Tea Research Station, National Institute of Vegetable and Tea Science, Japan. Segments of leaves ( ∼ 1 g FW) were homogenized in a chilled extraction medium containing 50 mM HEPES-NaOH buffer (pH 7.6), 2 mM Na-EDTA, 2 mM dithiothreitol (DTT), 0.5 % sodium ascorbate and 2.5 % polyvinylpolypyrrolidone, using a mortar and pestle on ice. The homogenate was centri-fuged at 30,000 × g for 20 min at 2 ° C. The supernatant was collected and treated with fi nely ground solid (NH 4 ) 2 SO 4 . The protein fraction that precipitated with 80 % saturation was collected by centrifuging (30,000 × g for 10 min), and was dissolved in 2.5 ml of 50 mM HEPES-NaOH buffer (pH 7.6) and applied on a column of PD-10 (GE Healthcare Bioscience Co.) equilibrated with the buffer. The protein fraction eluted with 3.5 ml of the buffer was used as the enzyme preparation.
Determination of enzyme activities All enzyme activities in this study were determined using 14 C-labeled substrates according to the method described in a previous report ( Ashihara et al. 2000 ). To obtain proper assay conditions, various preliminary experiments were carried out before determining the activity of each enzyme. Proportionality of the reaction velocity to the amount of enzyme was con-fi rmed by plotting initial velocities against at least three different amounts of the enzyme preparation.
The total volume of the reaction mixture was 100 µl, and incubation was performed at 30 ° C. The enzyme reactions were terminated by adding 10 µl of 60 % PCA. The reaction mixture was neutralized with 20 % KOH. After the precipitate had been removed, the neutralized samples were evaporated to dryness and the pellets were dissolved in 50 % (v/v) ethanol. The samples were then loaded onto cellulose TLC plates (Merck). The labeled substrates and products were separated using solvent systems I and III.
The compositions of the reaction mixtures for the enzyme assays were slightly different from those in a previous report ( Katahira and Ashihara 2006b ). The reaction mixtures for
2116
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

nucleoside kinases were 30 mM HEPES-NaOH buffer (pH 7.6), 10 mM MgCl 2 , 1 mM DTT, 3.75 mM ATP and 55 µM [8- 14 C]ade-nosine (0.65 GBq mmol − 1 ), [8- 14 C]guanosine (0.70 GBq mmol − 1 ), [8- 14 C]inosine (0.66 GBq mmol − 1 ) or [8- 14 C]xanthosine (0.70 GBq mmol − 1 ). The reaction mixtures for nucleosidases were the same as for nucleoside kinases except that ATP was omitted. The reaction mixtures for adenine-, hypoxanthine/guanine- and xanthine phosphoribosyltransferases were 30 mM HEPES-NaOH buffer (pH 7.6), 10 mM MgCl 2 , 1 mM DTT, 0.6 mM PRPP and 100 µM [8- 14 C]adenine (1.9 GBq mmol − 1 ), [8- 14 C]guanine (2.0 GBq mmol − 1 ), [8- 14 C]hypoxanthine (2.1 GBq mmol − 1 ) or [8- 14 C]xanthine (2.0 GBq mmol − 1 ).
Acknowledgements
We thank Dr. Junichi Tanaka, National Institute of Vegetable and Tea Science, Japan, for his generous supply of plant materials and helpful advice for preparation of tea seedlings.
References
Allen , M. , Qin , W. , Moreau , F. and Moffatt , B. ( 2002 ) Adenine phosphoribosyltransferase isoforms of Arabidopsis and their potential contributions to adenine and cytokinin metabolism . Physiol. Plant. 115 : 56 – 68 .
Ashihara , H. and Crozier , A. ( 1999 ) Biosynthesis and metabolism of caffeine and related purine alkaloids in plants . Adv. Bot. Res. 30 : 117 – 205 .
Ashihara , H. , Deng , W.W. , Mullen , W. and Crozier , A. ( 2010b ) Distribution and biosynthesis of fl avan-3-ols in Camellia sinensis seedlings and expression of genes encoding biosynthetic enzymes . Phytochemistry 71 : 559 – 566 .
Ashihara , H. , Kato , M. and Crozier , A. ( 2010a ) Distribution, biosynthesis and catabolism of methylxanthins in plants . In Handbook of Experimental Pharmacology Vol. 200 (Methylxanthines) . Edited by Fredholm , B.B. pp. 11 – 31 . Springer-Verlag , Berlin .
Ashihara , H. and Kubota , H. ( 1986 ) Patterns of adenine metabolism and caffeine biosynthesis in different parts of tea seedlings . Physiol.
Plant. 68 : 275 – 281 . Ashihara , H. , Monteiro , A.M. , Gillies , F.M. and Crozier , A. ( 1996a )
Biosynthesis of caffeine in leaves of coffee . Plant Physiol. 111 : 747 – 753 .
Ashihara , H. , Monteiro , A.M. , Moritz , T. , Gillies , F.M. and Crozier , A. ( 1996b ) Catabolism of caffeine and related purine alkaloids in leaves of Coffea arabica L . Planta 198 : 334 – 339 .
Ashihara , H. , Ogita , S. and Crozier , A. ( 2011 ) Purine alkaloid biosynthesis . In Plant Metabolism and Biotechnology . Edited by Ashihara , H. , Crozier , A. and Komamine , A. Wiley-Blackwell , Oxford (in press) .
Ashihara , H. , Sano , H. and Crozier , A. ( 2008 ) Caffeine and related purine alkaloids: biosynthesis, catabolism, function and genetic engineering . Phytochemistry 69 : 841 – 856 .
Ashihara , H. , Stasolla , C. , Loukanina , N. and Thorpe , T.A. ( 2000 ) Purine and pyrimidine metabolism in cultured white spruce ( Picea glauca ) cells: metabolic fate of 14 C-labeled precursors and activity of key enzymes . Physiol. Plant. 108 : 25 – 33 .
Ashihara , H. , Stasolla , C. , Loukanina , N. and Thorpe , T.A. ( 2001 ) Purine metabolism during white spruce somatic embryo development: salvage of adenine, adenosine, and inosine . Plant Sci. 160 : 647 – 657 .
Combes , A. , Lafl euriel , J. and Le Floc'h , F. ( 1989 ) The inosine–guanosine kinase activity of mitochondria in tubers of Jerusalem artichoke . Plant Physiol. Biochem. 27 : 729 – 736 .
Crozier , A. , Yokota , T. , Jaganath , I.B. , Marks , S.C. , Saltmarsh , M. and Clifford , M.N. ( 2006 ) Secondary metabolites in fruits, vegetables, beverages and other plant-based dietary components . In Plant Secondary Metabolites . Edited by Crozier , A. , Clifford , M.N. and Ashihara , H. pp. 208 – 302 . Blackwell , Oxford .
Guranowski , A. ( 1982 ) Purine catabolism in plants. Purifi cation and some properties of inosine nucleosidase from yellow lupin ( Lupinus
leteus L.) seeds . Plant Physiol. 70 : 344 – 349 . Hirose , F. and Ashihara , H. ( 1982 ) Adenine phosphoribosyltransferase
activity in mitochondria of Catharanthus roseus cells . Z. Naturforsch. 37c : 1288 – 1289 .
Hirose , N. , Makita , N. , Yamaya , T. and Sakakibara , H. ( 2005 ) Functional characterization and expression analysis of a gene, OsENT2 , encoding an equilibrative nucleoside transporter in rice suggest a function in cytokinin transport . Plant Physiol. 138 : 196 – 206 .
Ito , E. and Ashihara , H. ( 1999 ) Contribution of purine nucleotide biosynthesis de novo to the formation of caffeine in young tea ( Camellia sinencis ) leaves . J. Plant Physiol. 154 : 145 – 151 .
Katahira , R. and Ashihara , H. ( 2002 ) Profi les of pyrimidine biosynthesis, salvage and degradation in disks of potato ( Solanum tuberosum L.) tubers . Planta 215 : 821 – 828 .
Katahira , R. and Ashihara , H. ( 2006a ) Role of adenosine salvage in wound-induced adenylate biosynthesis in potato tuber slices . Plant
Physiol. Biochem. 44 : 551 – 555 . Katahira , R. and Ashihara , H. ( 2006b ) Profi les of purine biosynthesis,
salvage and degradation in disks of potato ( Solanum tuberosum L.) tubers . Planta 225 : 115 – 126 .
Kato , M. , Mizuno , K. , Crozier , A. , Fujimura , T. and Ashihara , H. ( 2000 ) Caffeine synthase gene from tea leaves . Nature 406 : 956 – 957 .
Kato , M. , Mizuno , K. , Fujimura , T. , Iwama , M. , Irie , M. , Crozier , A. , et al . ( 1999 ) Purifi cation and characterization of caffeine synthase from tea leaves . Plant Physiol. 120 : 579 – 586 .
Koshiishi , C. , Crozier , A. and Ashihara , H. ( 2001a ) Profi les of purine and pyrimidine nucleotides in fresh and manufactured tea leaves . J. Agric. Food Chem. 49 : 4378 – 4382 .
Koshiishi , C. , Kato , A. , Yama , S. , Crozier , A. and Ashihara , H. ( 2001b ) A new caffeine biosynthetic pathway in tea leaves: utilisation of adenosine released from the S -adenosyl- L -methionine cycle . FEBS
Lett. 499 : 50 – 54 . Koyama , Y. , Tomoda , Y. , Kato , M. and Ashihara , H. ( 2003 )
Metabolism of purine bases, nucleosides and alkaloids in theobromine-forming Theobroma cacao leaves . Plant Physiol.
Biochem. 41 : 977 – 984 . Krenitsky , T.A. , Neil , S.M. and Miller , R.L. ( 1970 ) Guanine and xanthine
phosphoribosyltransfer activities of Lactobacillus casei and Escherichia coli . Their relationship to hypoxanthine and adenine phosphoribosyltransfer activities . J. Biol. Chem. 245 : 2605 – 2611 .
Le Floc'h , F. and Lafl euriel , J. ( 1981 ) The purine nucleosidases of Jerusalem artichoke shoots . Phytochemistry 20 : 2127 – 2129 .
Le Floc'h , F. and Lafl euriel , J. ( 1983 ) The role of mitochondria in the recycling of adenine into purine nucleotides in the Jerusalem artichoke ( Heliunthus tuberosus L.) . Z. Pfl anzenphysiol. 113 : 61 – 71 .
Le Floc'h , F. , Lafl euriel , J. and Guillot , A. ( 1982 ) Interconversion of purine nucleotides in Jerusalem artichoke shoots . Plant Sci. Lett. 27 : 309 – 316 .
Li , G. , Liu , K. , Baldwin , S.A. and Wang , D. ( 2003 ) Equilibrative nucleoside transporters of Arabidopsis thaliana . cDNA cloning, expression
2117
Profi les of purine metabolism in tea
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022

pattern, and analysis of transport activities . J. Biol. Chem. 278 : 35732 – 35742 .
Mager , J. and Magasanik , B. ( 1960 ) Guanosine 5 ′ -phosphate reductase and its role in the interconversion of purine nucleotides . J. Biol.
Chem. 235 : 1474 – 1478 . McCarthy , A.A. and McCarthy , J.G. ( 2007 ) The structure of two
N -methyltransferases from the caffeine biosynthetic pathway . Plant Physiol. 144 : 879 – 889 .
Moffatt , B.A. and Ashihara , H. ( 2002 ) Purine and pyrimidine nucleotide synthesis and metabolism . In The Arabidopsis Book . Edited by Somerville , C. and Meyerowitz , E. American Society of Plant Biologists , Rockville, MD . doi/10.1199/tab.0018. http://www.aspb.org/publications/arabidopsis/
Negishi , O. , Ozawa , T. and Imagawa , H. ( 1994 ) Guanosine deaminase and guanine deaminase from tea leaves . Biosci. Biotech. Biochem. 58 : 1277 – 1281 .
Negishi , O. , Ozawa , T. and Imagawa , H. ( 1988 ) N -methylnucleosidase from tea leaves . Agric. Biol. Chem. 52 : 169 – 175 .
Nygaard , P. ( 1972 ) Deoxyribonucleotide pools in plant tissue cultures . Physiol. Plant. 26 : 29 – 33 .
Nikolau , B.J. and Wurtele , E.S. ( 2007 ) Concepts in Plant Metabolomics . Springer , Berlin .
Renart , M.F. and Sillero , A. ( 1974 ) GMP reductase in Artemia salina . Biochim. Biophys. Acta 341 : 178 – 186 .
Riewe , D. , Grosman , L. , Fernie , A.R. , Zauber , H. , Wucke , C. and Geigenberger , P. ( 2008 ) A cell wall-bound adenosine nucleosidase is involved in the salvage of extracellular ATP in Solanum tuberosum . Plant Cell Physiol. 49 : 1572 – 1579 .
Saito , K. , Dixon , R.A. and Willmitzer , L. ( 2006 ) Plant Metabolomics (Biotechnology in Agriculture and Forestry) . Springer , Berlin .
Schubert , K.R. and Boland , M.J. ( 1990 ) The ureides . In The Biochemistry of Plants . Edited by Stump , P.K. and Conn , E.E. pp. 197 – 282 . Academic Press , San Diego .
Stasolla , C. , Katahira , R. , Thorpe , T.A. and Ashihara , H. ( 2003 ) Purine and pyrimidine nucleotide metabolism in higher plants . J. Plant
Physiol. 160 : 1271 – 1295 . Stephens , R.W. and Whittaker , V.K. ( 1973 ) Calf thymus GMP
reductase: control by XMP . Biochem. Biophys. Res. Commun. 53 : 975 – 981 .
Sugiura , M. and Takeda , Y. ( 2000 ) Nucleic acids . In Biochemistry and Molecular Biology of Plants . Edited by Buchanan , B.B. , Gruissen , W. and Jones , R.L. pp. 260 – 310 . American Society of Plant Physiologists , Rockville, MD .
Thomas , R.J. and Schrader , L.E. ( 1981 ) Ureide metabolism in higher plants . Phytochemistry 20 : 361 – 371 .
Ukaji , T. and Ashihara , H. ( 1986 ) Purine salvage in mitochondria of cultured Catharanthus roseus cells . J. Plant Physiol. 125 : 191 – 197 .
Van der Graff , E. , Hooykaas , P. , Lein , W. , Lerchl , J. , Kunze , G. , Sonnewald , U. , et al . ( 2004 ) Molecular analysis of ‘de novo’ purine biosynthesis in solanaceous species and in Arabidopsis thaliana . Front. Biosci. 9 : 1803 – 1816 .
Weisburger , J.H. ( 2004 ) Tea and health . In Herbal and Traditional Medicine: Molecular Aspects of Health . Edited by Packer , L. , Ong , C.N. and Halliwell , B. pp. 117 – 144 . Marcel Dekker , New York .
Winkler , R.G. , Blevins , D.G. and Randall , D.D. ( 1988 ) Ureide catabolism in soybeans . Plant Physiol. 86 : 1084 – 1088 .
Yabuki , N. and Ashihara , H. ( 1991 ) Catabolism of adenine nucleotides in suspension-cultured plant cells . Biochim. Biophys. Acta 1073 : 474 – 480 .
Zrenner , R. and Ashihara , H. ( 2011 ) Nucleotide metabolism . In Plant Metabolism and Biotechnology . Edited by Ashihara , H. , Crozier , A. and Komamine , A. Wiley-Blackwell , Oxford (in press) .
Zrenner , R. , Stitt , M. , Sonnewald , U. and Boldt , R. ( 2006 ) Pyrimidine and purine biosynthesis and degradation in plants . Annu. Rev. Plant
Biol. 57 : 805 – 836 .
2118
W.-W. Deng and H. Ashihara
Plant Cell Physiol. 51(12): 2105–2118 (2010) doi:10.1093/pcp/pcq175 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2105/1944539 by guest on 17 M
arch 2022




















