
Probing structural elements of thermal stability in bacterial oligomeric alcohol dehydrogenases. I....
10
I~1 Lettersin Peptide Science 5: 399-408, 1998. ~ KLUWER/ESCOM 1998 Kluwer AcademicPublishers. Printedin the Netherlands. 399 LIPS 261 Probing structural elements of thermal stability in bacterial oligomeric alcohol dehydrogenases. I. Construction and characterization of chimeras consisting of secondary ADHs from Thermoanaerobacter brockii and Clostridium be~ierinckii Oren Bogin, Moshe Peretz & Yigal Burstein* Department of Organic Chemistry, The Weizmann Institute of Science, Rehovot 76100, Israel Received 1 July 1997;Accepted25 August 1997 Key words: ADH, chimera, mutagenesis, protein engineering, thermal stability Summary Two tetrameric secondary alcohol dehydrogenases (ADHs), one from the mesophile Clostridium beijerinckii (CBADH) and the other from the extreme thermophile Thermoanaerobacter brockii (TBADH), share 75% se- quence identity but differ by 26 ~ in thermal stability. To explore the role of linear segments of these similar enzymes in maintaining the thermal stability of the thermostable TBADH, a series of 12 CBadh and TBadh chimeric genes and the two parental wild-type genes were expressed in Escherichia coli, and the enzymes were isolated, purified and characterized. The thermal stability of each chimeric enzyme was approximately exponen- tially proportional to the content of the amino acid sequence of the thermophilic enzyme, indicating that the amino acid residues contributing to the thermal stability of TBADH are distributed along the whole protein molecule. It is suggested that major structural elements of thermal stability may reside among the nine discrepant amino acid residues between the N-terminal 50-amino acid residues of TBADH and CBADH. Abbreviations: ADH, alcohol dehydrogenase; TBADH, Thermoanaerobacter brockii ADH; CBADH, Clostrid- ium beijerinckii ADH; DTT, d, l dithiothreitol; Gdn-HC1, guanidine hydrochloride; PAGE, polyacrylamide gel electrophoresis; SDS, sodium dodecyl sulfate. Introduction The ubiquitous alcohol dehydrogenases (ADHs) are found not only in bacteria [1] but also in eukaryotes ranging from yeast to man [1-5]. ADHs are oxidore- ductases, requiring either NADH or NADPH as coen- zyme, that react with primary and secondary, linear and branched-chain, aliphatic and aromatic alcohols and with their corresponding aldehydes and ketones. Most ADHs contain zinc at the active site. The Zn- ADHs are either dimers, usually found in higher plants and mammals, or tetramers, such as those present in yeast and bacteria. The ADHs have been exten- sively studied and there is a comprehensive database * To whomcorrespondence shouldbe addressed. on their kinetics and sequences. Nevertheless, the de- tailed structure of only the horse liver [6] and the human/31#1 [7,8] ADHs, both eukaryotic, dimeric NAD-linked ADHs, has been determined by X-ray crystallography. No three-dimensional (3D) structure of a prokaryotic or tetrameric or NADP-linked ADH has been published. We have been studying ADHs from the ex- treme thermophile Thermoanaerobacter brockii and the mesophile Clostridium beijerinckii. Both the T. brockii ADH (TBADH) and the C. beijerinckii ADH (CBADH) are homotetramers comprising "-~38 kDa subunits. They are medium-chain, zinc-containing en- zymes with similarly broad substrate ranges. The two secondary ADHs are NADP(H)-linked, but have a
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Probing structural elements of thermal stability in bacterial oligomeric alcohol dehydrogenases. I....

I~1 Letters in Peptide Science 5: 399-408, 1998. ~ KLUWER/ESCOM
�9 1998 Kluwer Academic Publishers. Printed in the Netherlands. 399
LIPS 261
Probing structural elements of thermal stability in bacterial oligomeric alcohol dehydrogenases. I. Construction and characterization of chimeras consisting of secondary ADHs from Thermoanaerobacter brockii and Clostridium be~ierinckii
Oren Bogin, Moshe Peretz & Yigal Burstein* Department of Organic Chemistry, The Weizmann Institute of Science, Rehovot 76100, Israel
Received 1 July 1997; Accepted 25 August 1997
Key words: ADH, chimera, mutagenesis, protein engineering, thermal stability
Summary
Two tetrameric secondary alcohol dehydrogenases (ADHs), one from the mesophile Clostridium beijerinckii (CBADH) and the other from the extreme thermophile Thermoanaerobacter brockii (TBADH), share 75% se- quence identity but differ by 26 ~ in thermal stability. To explore the role of linear segments of these similar enzymes in maintaining the thermal stability of the thermostable TBADH, a series of 12 CBadh and TBadh chimeric genes and the two parental wild-type genes were expressed in Escherichia coli, and the enzymes were isolated, purified and characterized. The thermal stability of each chimeric enzyme was approximately exponen- tially proportional to the content of the amino acid sequence of the thermophilic enzyme, indicating that the amino acid residues contributing to the thermal stability of TBADH are distributed along the whole protein molecule. It is suggested that major structural elements of thermal stability may reside among the nine discrepant amino acid residues between the N-terminal 50-amino acid residues of TBADH and CBADH.
Abbreviations: ADH, alcohol dehydrogenase; TBADH, Thermoanaerobacter brockii ADH; CBADH, Clostrid- ium beijerinckii ADH; DTT, d, l dithiothreitol; Gdn-HC1, guanidine hydrochloride; PAGE, polyacrylamide gel electrophoresis; SDS, sodium dodecyl sulfate.
Introduction
The ubiquitous alcohol dehydrogenases (ADHs) are found not only in bacteria [1] but also in eukaryotes ranging from yeast to man [1-5]. ADHs are oxidore- ductases, requiring either NADH or NADPH as coen- zyme, that react with primary and secondary, linear and branched-chain, aliphatic and aromatic alcohols and with their corresponding aldehydes and ketones. Most ADHs contain zinc at the active site. The Zn- ADHs are either dimers, usually found in higher plants and mammals, or tetramers, such as those present in yeast and bacteria. The ADHs have been exten- sively studied and there is a comprehensive database
* To whom correspondence should be addressed.
on their kinetics and sequences. Nevertheless, the de- tailed structure of only the horse liver [6] and the human/31#1 [7,8] ADHs, both eukaryotic, dimeric NAD-linked ADHs, has been determined by X-ray crystallography. No three-dimensional (3D) structure of a prokaryotic or tetrameric or NADP-linked ADH has been published.
We have been studying ADHs from the ex- treme thermophile Thermoanaerobacter brockii and the mesophile Clostridium beijerinckii. Both the T. brockii ADH (TBADH) and the C. beijerinckii ADH (CBADH) are homotetramers comprising "-~38 kDa subunits. They are medium-chain, zinc-containing en- zymes with similarly broad substrate ranges. The two secondary ADHs are NADP(H)-linked, but have a

400
NdeI 20 40 NarI 601
I : : ] : CBADH: G-N-L R-VA-SY S L-D-K
TBADH: MKGFAMLSIGKVGWIEKEKPAPGPFDAIVRPLAVAPCTSDIHTVFEGAIGERHNMILGHE
CBADH:
TBADH:
80 i00 120 : : :
,--,,--'---,,---,-,,---. F
AVGEVVEVGSEVKDFKPGDRVVVPAITPDWRTSEVQRGYHQHSGGMLAGWKFSNVKDGVF
CBADH:
TBADH:
140 160 180
--Y I---DM---N .... T .... S QM-SS-V-I---A---
GEFFHVNDADMNLAHLPKEIPLEAAVMIPDMMTTGFHGAELADIELGATVAVLGIGPVGL
Accl 220 240 I: : :
CBADH: -GI-- G ..... .--.---.--.---.---,-,-,-,-.--.-,---,,-
TBADH: MAVAGAKLRGAGRIIAVGSRPVCVDAAKYYGATDIVNYKDGPIESQIMNLTEGKGVDAAI
CBADH:
TBADH:
260 280 Aatll
: : I : M---GSETLSQ--SM ..... I-S-I--H-S-DA-LI--V
IAGGNAD IMATAVK IVKPGGT IANVNYFGEGEVL PVPRL EWGCGMAHKT I KGGLC PGGRL
320 340 BamHI
: : I CBADH : -A-M-R-M-V-N- --L YH---H--E-LL- A .... *
TBADH : RMERLIDLVFYKRVDPSKLVTHVFRGFDNIEKAFMLMKDKPKDLIKPVVILA*
Figure 1. Amino acid sequence alfgnment of alcohol dehydrogenases from the thermophile T. brockii, (TBADH) and the mesophile C. beijer- inckii (CBADH). Identical residues in CBADH are marked by a bar. The positions of the corresponding restriction sites in the adh genes are marked by vertical bars. The BamHI site is located at the termination codon.
very low level of NAD(H)-linked activity. In previ- ous research, we purified TBADH to homogeneity and determined its primary structure [9] and stere- ospecificity [10], and identified the catalytic metal zinc and its amino acid ligands [11]. CBADH was purified to homogeneity from C. beijerinckii strain NRRL B593 [12]. We have also cloned, sequenced and overexpressed both enzymes [13]. Although they share 75% sequence identity (Figure 1), an unusually high degree of similarity for proteins from different organisms [13], they differ greatly in their thermosta- bility. TBADH reversibly catalyzes the oxidation of secondary alcohols to the corresponding ketones with a --1/2T60 mi~ of 93 ~ (the temperature at which 50% of the enzymatic activity is lost after 60 min), while
T60 min ~ C CBADH has a -1/2 of 67 [14]. Comparing the detailed structure of these similar enzymes could shed light on the principles governing protein thermostabil-
ity and consequently aid in the design of structural elements of thermal stability in engineered enzymes. Such enzymes with enhanced thermostability would be useful catalysts in biotechnology in general and in biotransformation reactions in particular [15-17].
Although general methods of increasing protein thermostability are lacking, several approaches have been suggested to serve as guidelines for this purpose. These were based on comparisons of primary, sec- ondary and tertiary structures of mainly monomeric and several oligomeric mesophilic proteins and their thermophilic counterparts. The studies point to al- most every interaction in proteins as a possible struc- tural determinant of thermostability. These include stabilization of structure and dipole moments of or- helices [18-21], additional prolines in r and external loops [22,23], salt bridges [24-27], increase in the number of interactions between charged side

401
chains and neutral partners ('charged-neutral' hydro- gen bonds) [28], and improved hydrophobic interac- tions and oligomeric rearrangements that fill existing cavities and lead to tighter molecular packing [29- 31]. Salt-bridge networks that are energetically more favorable than isolated ion pairs [32] have been noted as prominent structural features that can explain the unusual properties of hyperthermophiles [27,33,34]. It seems that there is no single suggested property that makes a protein withstand elevated temperatures. Rather, as predicted by Matthews et al. [35], protein stabilization is likely accomplished through a com- bination of subtle changes specific to each protein. Effects of the above structural determinants on the thermal stability of several individual proteins have been tested by site-directed mutagenesis [36-40], but no generalization that would explain the underlying principles of stabilization has emerged from these studies.
A good strategy for understanding the thermosta- bility of proteins would be to analyze families of uni- versal, oligomeric enzymes with different thermosta- bilities. Members of these enzyme families should have high sequence homology and eventually de- tailed 3D structures for both the thermophilic and the mesophilic proteins. However, even in the ab- sence of a detailed 3D structure of the mesophilic and thermophilic enzymes, construction and analysis of chimeric enzymes should provide essential infor- mation about the location of structural elements of thermostability in the thermostable protein. Such stud- ies were performed with several chimeras and hybrids of glyceraldehyde-3-phosphate dehydrogenase [41], r [42] and isopropylmalate dehydrogenase [43,44].
We report here the construction and expression of the chimeric T. brockii and C. beijerinckii adh genes, and the isolation and characterization of their gene products, chimeric alcohol dehydrogenases.
Materials and methods
Materials
Enzymes for DNA cloning, sequencing and ampli- fication were from Amersham (Amersham, U.K.), GIBCO-BRL (Uxbridge, U.K.), New England Bio- labs (Beverley, MA, U.S.A.) or Promega (Madison, WI, U.S.A.). [F-32p]ATP was from Amersham or DuPont NEN (Boston, MA, U.S.A.). Oligonucleotides for cloning, sequencing and site-directed mutagenesis
of the T. brockii and Ci beijerinckii adh genes were synthesized by the WIS Chemical Synthesis Labora- tory (Rehovot, Israel). DEAE-cellulose DE-52 was a product of Whatman Chemicals (Maidstone, U.K.). Red Sepharose-CL-6B was from Pharmacia (Uppsala, Sweden). All other chemicals were of analytical grade.
Recombinant DNA techniques
The construction and expression of the genes encod- ing TBADH and CBADH was performed according to Peretz et al. essentially as described elsewhere [13]. Site-directed mutagenesis was performed according to Kunkel et al. [45] and the mutations were verified by DNA sequencing of the relevant plasmids.
Synthetic oligonucleotides used for generating or aborting restriction sites are listed below and the ex- changed bases are underlined. In the C. brockii adh gene (SacI-BamHI fragment):
for aborting the Nde169 site 5/-GTACAATAGCATCATAC GAACCCGCAACTG-3 /
for the NdeI 1 site 5 / - CA'ITG CAAAACCTTTCATA T GATATGTTCCTC-3 t
for the BamH11060 site 5 r - G TA G.__G_G ATCCTCCAT GATCTATTATG - 3 t
In pBS-P89CBADH-l:
for the NarI139site 5~ -CTTCCTATCTCCAAGG GC G CCCTCAAAAACAG-3 I
for the Acc1596 site 51-GCCTCAACACAAATCGGTCTACTCCCCACTCC-3 ~
for the AatI1892 site 5 / .CATTTCTGCTCTCAGACGTCCCCCAGGAC_3 I
Colonies containing hybrid gene plasmids were first selected by hybridization with the following short oligonucleotide probes overlapping the spliced re- gions:
NarI site TB/CB ! l 5 - GAAG G C G C C CTTGGAG-3
NarI site CB/TB
5 I-GAGGGCGCCATTGGCG-3 I
AccI site TB/CB
5'-CGTAGGCAG T A G A CCGA-3 I
AccI site CB/TB
5 f-GAGTGGGGAG T A GA CCAG-3'
AatII site TB/CB t ! 5 -CGGTGG A C G T CTGAGAGCAG-3
AatlI site CB/TB
Y-CTGGGGG A C G T CTAAGAATG-3 r
The hybrid products were then verified by DNA se-

402
quencing of the entire coding region in the relevant plasmids.
pBluescript II SK (+) was modified to accommo- date the adh genes by eliminating theAcc1675 site from the polylinker. This was accomplished by first cleav- ing the vector with SalI (at the common restriction site Sal[674/AccI675), followed by trimming with DNA polymerase and ligation.
Protein expression and purification All the recombinant plasmids harboring TBADH, CBADH and the Chimeric ADHs were transformed into E. coli strain TG-1 [46]. The tetrameric recombi- nant proteins were purified according to a modification of the procedure described by Peretz and Burstein [9]. Briefly, the transformed cells were cultured in 2YT with Ampicillin (100 /zg/ml) for 14-17 h at 37 ~ harvested by centrifugation at 8000 x g for 20 min at 4 ~ and resuspended in 25 mM Tris-HCl, 0.1 mM DTT, 0.1 mM EDTA, 1 mM benzamidine, 0.02% sodium azide (pH 7.3) (buffer A). Cells were dis- rupted for 5 min by intermittent sonication (30 s, with 1 min intermissions) on ice, followed by centrifuga- tion at 23 000 x g for 15 min, to remove cell debris. The supernatant was then heat-treated at 65 ~ for 3 min and centrifuged again for 15 rain at 12 000 x g. The clear supernatant was applied onto a DEAE-52- cellulose column (13 x 3 cm), pre-equilibrated with buffer A at 4 ~ and was extensively washed with the same buffer until no protein was eluted (measured according to Bradford [47]). The recombinant pro- teins were eluted from the column with a solution of 0.1 mM NaC1 in buffer A. The enzymatically ac- tive fractions were pooled and dialyzed extensively against buffer A for 12 h, and then applied to a short Red-Sepharose column (7 x 3 cm). The recom- binant enzyme was eluted from the column with a linear gradient of NaC1 (0.1-2 M) in buffer A. Frac- tions containing enzymatic activity were collected and concentrated by ultrafiltration (Amicon YM-30), and stored at 4 ~ All the enzymes used in this study were purified to homogeneity as judged by SDS-PAGE (data not shown).
Enzyme assay and kinetic experiments Enzymatic activity of the ADHs was measured at 40 ~ by following the reduction of NADP + (and monitoring the formation of NADPH) at 340 nm (e340 = 6.2/mM/cm). The standard assay mixture contained 150 mM 2-butanol, 0.5 mM NADP + and 100 mM Tris-HC1 (pH 9.0) in a total volume of 1 ml. One
unit of ADH is defined as the amount of enzyme that catalyzes the oxidation of I/xmol of 2-butanol/min un- der initial velocity at the above-mentioned conditions. Enzymatic activity was measured with a Beckman DU-7500 spectrophotometer equipped with a Multi- component/SCA/Kinetics Plus software package and a thermostat water circulating bath. Values are the means of three experiments and the individual mea- surements were within 10% of the quoted mean.
(it60 rain) analysis Thermal inactivation 1/2 The thermal stability of the ADHs was determined by monitoring their residual enzymatic activity after a 60 rain incubation in buffer A (see above) at different temperatures. Enzymatic activity was assayed by spec- trophotometric analysis of the formation of NADPH at 340 nm, as above. Values are the means of three exper- iments and the individual measurements were within 10% of the quoted mean.
Analytical procedures DNA was extracted by the alkaline lysis procedure [46], separated by agarose gel (1%, w/v) elec- trophoresis, and then purified to homogeneity using GenecleanII kit (Bio 101). DNA sequencing was per- formed using an Applied Biosystems Model 373A DNA sequencer by the dideoxy method [48] and ap- propriate primers. SDS-PAGE was performed on 12% slab gels and 5% stacking gels [49], using a Bio- Rad MiniProtean II system. The gels were stained with Coomassie brilliant blue. Protein concentrations were determined using Bradford [47] and Lowry [50] reagents, with bovine serum albumin as the standard.
Results and discussion
Construction and expression of plasmids encoding CBADH and TBADH The methods used for constructing the expression vec- tors for the T. brockii (TB)adh and the C. beijerinckii (CB)adh genes and their chimeric genes, in pBlue- script II SK (+), are outlined in Figure 2. The wild- type TBadh gene contains three unique restriction sites: Nar1139, Acc1595 and AatIIS92[13 ]. In a previ- ous study, three additional restriction sites, Sac1-89, NdeI 1 and BamHI 1~176 were generated at nucleotide -89 in the 5' untranslated region, at the initiation codon, and following the termination codon, respec- tively [13]. The TBadh gene (1168-nucleotide SacI- BamHI fragment) was cloned into the SacI-BamHI
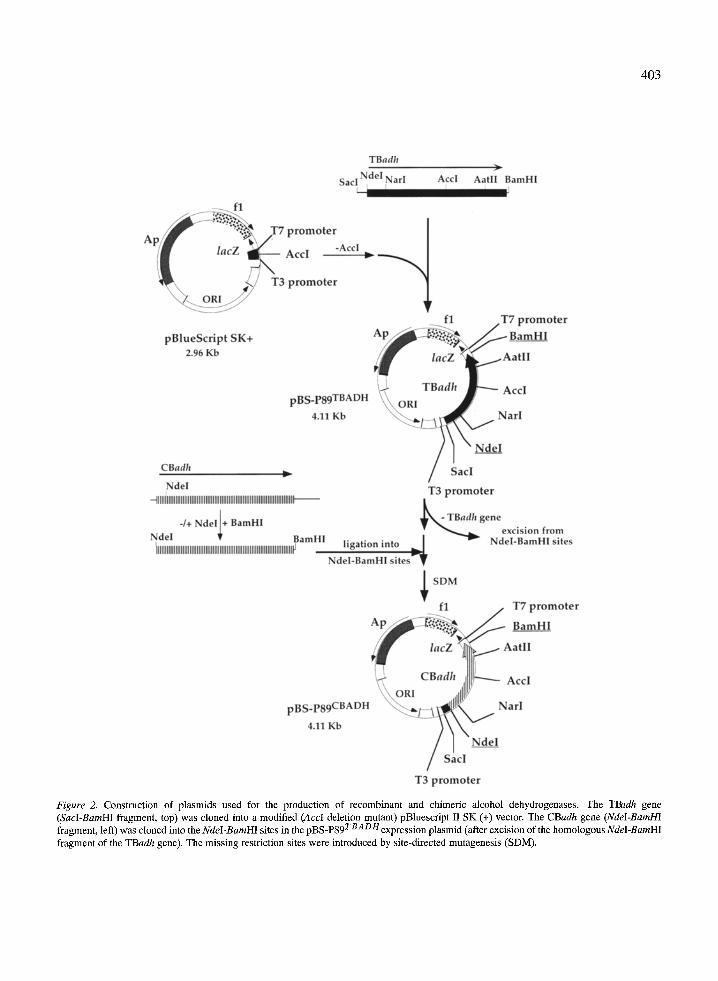
403
Figure 2. Construction of plasmids used for the production of recombinant and chimeric alcohol dehydrogenases. The TBadh gene (SacI-BamHI fragment, top) was cloned into a modified (AccI deletion mutant) pBluescript II SK (+) vector. The CBadh gene (NdeI-BamHI
TBADH fragment, left) was cloned into the NdeI-BamHI sites in the pBS-P89 expression plasmid (after excision of the homologous NdeI-BamHI fragment of the TBadh gene), The missing restriction sites were introduced by site-directed mutagenesis (SDM).

404
adh
NdeI NarI
C B A D H C1
),_
AccI AatII R a m H I
C2,4 C5-6 C7
T B A D H 11111111111111111111~1.1~1411111111111111~11111111~111 T1 T2-4 T5-6 T7
Figure 3. Schematic representation and nomenclature of the CBadh and the TBadh wild-type genes and their respective gene prod- ucts CBADH and TBADH. The five restriction sites (NdeI, NarI, AccI, AatII and BamHI) divide the adh coding gene region into four contiguous fragments consisting of multiples (xl, x3, x2 and xl) of approximately 50 nucleotides each. Accordingly, in the translated ADH, the N-terminal ~50-amino acid residue segment (coded by the NdeI1-Nar1139 fragment) is denoted 'l'; the adjacent ~150-residue segment (coded by the NarI139-Acc1595 fragment) is denoted '2.3.4'; the next ~100-residue segment (coded by the AccI595-AatI1892 fragment) is denoted '5.6'; and the C-terminal ~50-residue segment, (coded by theAatII892-BamH11060 fragment) is denoted '7'. The prefix T (for TBADH) or C (for CBADH) in- dicates the origin of the fragment. Open boxes, CBADH; hatched boxes: TBADH.
cloning sites of pBluescript II SK (+) (with the adh gene at the same orientation as the lacZ gene) to form the plasmid pBS-P89 rBADt/. The CBadh gene con- tained only one of the above restriction sites (NdeI69), though not in the desired position of the coding re- gion. This NdeI site was eliminated from position 69 and introduced at the initiation codon by silent mu- tagenesis. A BamHI site was then constructed at the 3 ~ end, and the 1078-nucleotide NdeI-BamHI frag- ment was cloned into the Ndel-BamHI sites of the vector pBS-P89 TBADH (see above). In this plasmid, denoted pBS-P89 cRApe- l , the CBadh gene replaced the TBadh gene, thus forming an expression vector for CBADH. In both plasmids pBS-P89 TBADH and pBS-P89 cBADH-1, the adh genes were oriented in the same direction.
The three restriction sites Nar1139, Acc1595 and AatIIS92present in pBS-P89 tRAp/4 have been intro- duced into identical positions in pBS-P89 cBaDH-1, using single or double mutations that preserved the primary structure of the enzyme (see the Materials and Methods section). The plasmid thus formed, denoted pBS-P89 CBaDH, and the parent non-mutated plas- mid pBS-P89 cBADH-1, both directed the synthesis of recombinant CBADH in essentially identical yields (~50 mg enzyme/l) and specific activities (180 u/mg).
Construction, nomenclature and expression of TB- and CB-adh chimeric genes
Although the properties of chimeric proteins can provide useful information and definite conclusions about structure-function, structure-thermal stability and other relationships, the unknown effects of seg- mental substitutions on adjacent and distant structural features of the protein constrain the information that can be derived by such an approach [51,52]. In the case of chimeric enzymes that are evaluated on the basis of thermal stability, several potential outcomes can be an- ticipated: (ia) the chimeric enzyme exhibits enzymatic activity that conforms to the wild-type parental activ- ity; (ib) the chimera lacks enzymatic activity; (ic) the enzymatic activity of the chimera is altered; (iia) the thermal stability of the chimeric enzymes is similar to the stability of one of the parental wild-type enzymes; (iib) the thermal stability of the chimeric enzymes dif- fers from those of the parental wild-type enzymes. As will be shown, several combinations of the above experimental outcomes occurred.
The monomeric subunits of CBADH and TBADH are 351 [13] and 352 [9] residues long, respectively (Figure 1). The unique restriction sites NdeI 1, Nar1139, Acc1595, AatI1892 and BamH11060 (present in both plasmids pBS-P89 cRAb/4 and pBS-P89 TBaDH) are positioned at translated amino acid residue numbers 1, 47, 199, 299 and beyond the C-terminal residue, respectively (Figure 1). These restriction sites split the adh genes into four segments encoding approxi- mately 50, 150, 100 and 50 residues, respectively (see Figure 1). For nomenclature purposes, the ADH mole- cules ('-~350 residues each) were divided into seven contiguous segments of approximately 50 residues each (Figure 3).
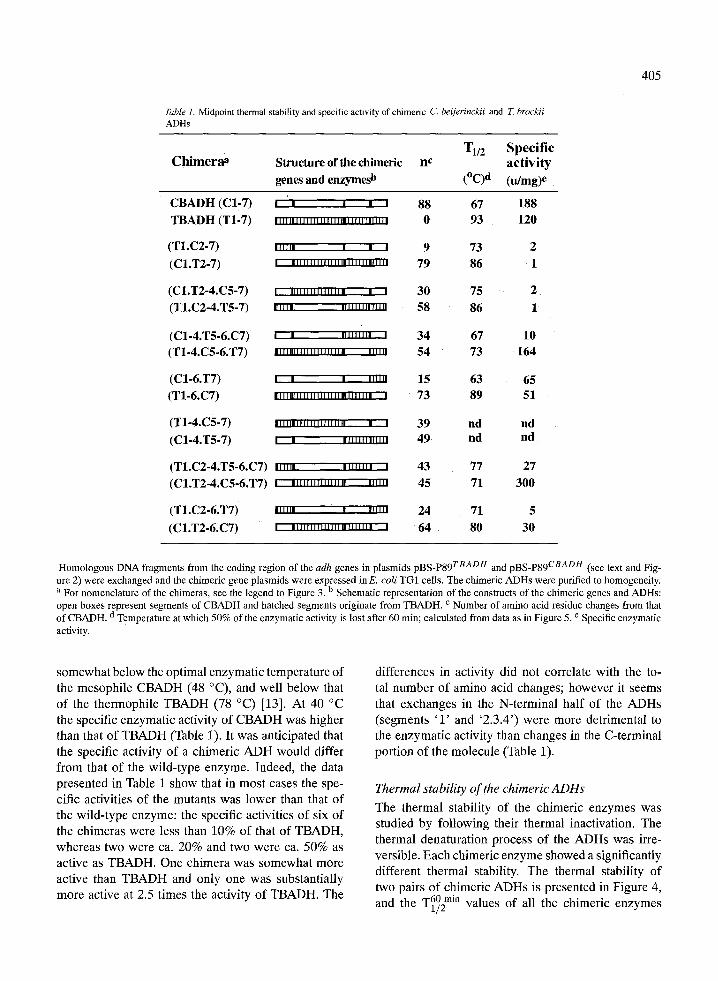
As shown in Table 1 (column 2), all 14 differ- ent combinations and permutations of the chimeric genes were constructed utilizing the available five splice sites. Homologous DNA fragments were ex- changed between the TB- and the CB-adh genes forming complementary pairs of hybrid-daughter adh genes. Four pairs of single fragment exchanges and three pairs of double fragment exchanges were suc- cessfully expressed and the recombinant chimeric en- zymes were isolated in varying yields. Attempts to express chimeras T1-4.C5-7 and C1-4.T5-7 (Table 1) were unsuccessful and only minute quantities of the chimeric enzymes could be obtained. The chimeric enzymes that were obtained were purified to homo- geneity and their specific enzymatic activities were determined using 2-butanol as the substrate at 40 ~
405
Table 1. Midpoint thermal stability and specific activity of chimeric C. beijerinckii and Z brockii ADHs
C h i m e r a a Structure of the chimeric nc
genes and enzymesb
T1/2 Specif ic ac t iv i ty
(~ (idmg)e
CBADH (C1-7) ' ' ' ' ' 88 67 188 TBADH (T1-7) . . . . . , . . . . . . , , . . . , . . . , . . r . . . , . 0 93 120
(T1.C2-7) . l . l . . ' ' ' 9 73 2 (C1.T2-7) ' ' " ' " " " " " ' " ' " " " " ' 79 86 1
(C1.T2-4.C5-7) I " " ' " " " ' " " . ' 30 75 2 (T1.C2-4.T5-7) . I . . . " " " " ' " ' " 58 86 1
(C1-4.T5-6.C7) I t I " ' " " ' 34 67 10 (T1-4.C5-6.T7) ' " " " " ' ' " . . . . . " " " 54 73 164
(C1-6.T7) I I ' "" f' 15 63 65 (T1-6.C7) ,lll,B,,,,,l,,,,,,,,,,,l,,l, , 73 89 51
(T1-4.C5-7) I,,,,,,,,,,l,,.,,., ' " ' 39 nd nd (C1-4.T5-7) I �9 .I.,,,...... 49 nd nd
(T1.C2-4.T5-6.C7) ,,,,,, ,,,,,,,, , 43 77 27 (C1.T2-4.C5-6.T7) , , , , , , , , l , , . , , , , ,.,~, 45 71 300
(T1.C2-6.T7) ,. , , , , I.,l, 24 71 5
(C1.T2-6.C7) | ...,,,..r...I...,,..... . 64 80 30
Homologous DNA fragments from the coding region of the adh genes in plasmids pBS-P89 TBADH and pBS-P89 CBADH (see text and Fig- ure 2) were exchanged and the chimeric gene plasmids were expressed inE. coli TG1 cells. The chimeric ADHs were purified to homogeneity. a For nomenclature of the chimeras, see the legend to Figure 3. b Schematic representation of the constructs of the chimeric genes and ADHs: open boxes represent segments of CBADH and hatched segments originate from TBADH. c Number of amino acid residue changes from that
d of CBADH. Temperature at which 50% of the enzymatic activity is lost after 60 min; calculated from data as in Figure 5. e Specific enzymatic activity.
somewhat below the optimal enzymatic temperature of the mesophile CBADH (48 ~ and well below that of the thermophile TBADH (78 ~ [13]. At 40 ~ the specific enzymatic activity of CBADH was higher than that of TBADH (Table 1). It was anticipated that the specific activity of a chimeric ADH would differ from that of the wild-type enzyme. Indeed, the data presented in Table 1 show that in most cases the spe- cific activities of the mutants was lower than that of the wild-type enzyme: the specific activities of six of the chimeras were less than 10% of that of TBADH, whereas two were ca. 20% and two were ca. 50% as active as TBADH. One chimera was somewhat more active than TBADH and only one was substantially more active at 2.5 times the activity of TBADH. The
differences in activity did not correlate with the to- tal number of amino acid changes; however it seems that exchanges in the N-terminal half of the ADHs (segments ' 1 ' and '2.3.4') were more detrimental to the enzymatic activity than changes in the C-terminal portion of the molecule (Table 1).
Thermal stability of the chimeric ADHs
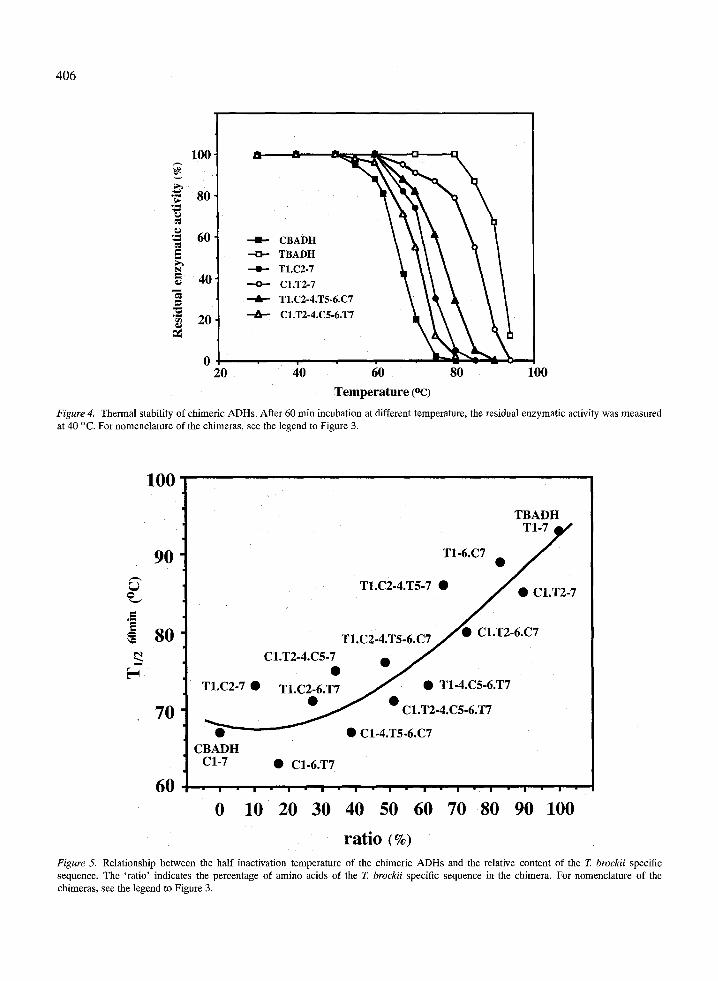
The thermal stability of the chimeric enzymes was studied by following their thermal inactivation. The thermal denaturation process of the ADHs was irre- versible. Each chimeric enzyme showed a significantly different thermal stability. The thermal stability of two pairs of chimeric ADHs is presented in Figure 4,
T60 rain and the ~1/2 values of all the chimeric enzymes
406
100 t
�9 ~ 8o
'~ 61)
=~ 40 --~- Cl.T2-7 "~ \ \ \ \ / _.-~ TLC2-4.T5-6.C7 \ ~ ~ ~
"~ ~ 20 + C1.T2-4.C5-6.T7 ~ ~ , , ~
20 40 60 80 100 Temperature (oc)
Figure 4. Thermal stability of chimeric ADHs. After 60 rain incubation at different temperature, the residual enzymatic activity was measured at 40 ~ For nomenclature of the chimeras, see the legend to Figure 3.
e =
100
90
80
70
60
TBADH TI-~ j~ t
T1-6.C7 �9 /
T1.C2-4.T5-7 �9 / �9 C1.T2-7
T 1 . C 2 . 4 . T 5 . 6 . C T / � 9 C1.T2-6.C7
C1.T2-4.C5-~ � 9
T1.C2.7 �9 T1 C2.6 T7 J �9 T1-4.C5-6.T7
~ _ _ y �9 C1.T2-4.C5-6.T7
�9 C1-4.T5-6.C7 CBADH
C1-7 �9 C1-6.T7
I I I I I I I I I I I
0 10 20 30 40 50 60 70 80 90 100
ratio ( % )
Figure 5. Relationship between the half inactivation temperature of the chimeric ADHs and the relative content of the Z brockii specific sequence. The 'ratio' indicates the percentage of amino acids of the T. brockii specific sequence in the chimera. For nomenclature of the chimeras, see the legend to Figure 3.

(determined from data such as in Figure 4) are pre- sented in Table 1 (column 4). Figure 5 shows the re- lationship between the half-denaturation temperature T60 min~ --1/2 j and the relative content of the T. brockii ADH
sequence of the chimeric ADHs. A rather smooth exponential relationship between the half denatura- tion temperature and the relative content of TBADH is observed, indicating that the amino acid residues contributing to the thermal stability of TBADH are distributed along the whole protein molecule. A simi- lar phenomenon was observed in chimeric isopropyl- malate dehydrogenase [44]. A more careful inspection of the data reveals that almost all values ' above ' the polynomial curve belong to chimeras containing the T1 segment, suggesting that major structural elements of thermal stability may re s ide among the nine dis- crepant amino acid residues between the T1 and C1 segments of TBADH and CBADH. This suggestion will be tested once the crystal structure of TBADH and CBADH [53] is determined.
Conclusions
We have demonstrated that chimeric genes constructed from alternating linear segments of T. brockii and C. beijerinckii adh genes can be overexpressed in E. coli, and the chimeric ADHs can be isolated in high yields. The specific activity of the majori ty of the chimeric ADHs was lower than that of either parent wild- type ADH, indicating that segmental substitu- tions may have affected adjacent or distant structural features involved in enzymatic activity. The thermal stability of the chimeric ADHs indicated that the amino acid residues contributing to the thermal stabil- ity of TBADH are distributed along the whole protein molecule, and it is suggested that major structural elements of thermal stability may reside among the first nine discrepant amino acid residues between the N-terminal segments of TBADH and CBADH.
Several of the chimeric ADHs will be crystallized, their crystal structure determined and the roles of linear segments of the enzymes in maintaining their thermal stability will be analyzed.
Acknowledgements
This work was supported by a research grant from the Henri and Francoise Glasberg Foundation. Y.B.
407
is the Maynard I. and Elaine Wishner Professor for Bioorganic Chemistry.
References
1. Reid, M.E and Fewson, C.A., Crit. Rev. Microbiol., 20 (1994) 13.
2. Yasunami, M., Chen, C.S. and Yoshida, A., Proc. Natl. Acad. Sci. USA, 88 (1991) 7610.
3. Danielsson, O., Atrian, S., Luque, T., Hjelmqvist, L., Gonzalez-Duarte, R. and J6rnvall, H., Proc. Natl. Acad. Sci. USA, 91 (1994) 4980.
4. Sun, H.W. and Plapp, B.V., J. Mol. Evol., 34 (1992) 522. 5. Satre, M.A., Zgombic, K.M. and Duester, G., J. Biol. Chem.,
269 (1994) 15606. 6. Br~ind6n, C.I., Eklund, H., Nordstrom, B., Boiwe, T., Soder-
lund, G., Zeppezauer, E., Ohlsson, I. and Keson, A., Proc. Natl. Acad. Sci. USA, 70 (1973) 2439.
7. Hurley, T.D., Bosron, W.E, Hamilton, J.A. and Amzel, L.M., Proc. Natl. Acad. Sci. USA, 88 (199l) 8149.
8. Yang, Z.N., Borson, W.F. and Hurley, T.D., J. Mol. Biol., 265 (1997) 330.
9. Peretz, M. and Burstein, Y., Biochemistry, 28 (1989) 6549. 10. Peretz, M., Bogin, O., Keinan, E. and Burstein, Y., Int. J. Pept.
Protein Res., 42 (1993) 490. 11. Bogin, O., Peretz, M. and Burstein, Y., Protein Sci., 6 (1997)
450. 12. Ismaiel, A.A., Zhu, C.X., Colby, G.D. and Chen, J.S., J.
Bacteriol., 175 (1993) 5097. 13. Peretz, M., Bogin, O., Tel-Or, S., Cohen, A., Li, G., Chen,
J.-S. and Burstein, Y., Anaerobe, 3 (1997) 259. 14. Bogin, O., Peretz, M. and Burstein, Y., in Geisow, M.J. (Ed.)
Perspectives on Protein Engineering: From Folds to Functions, Vol. 3, BIODIGM, Nottingham, 1996, p. 157.
15. Cowan, D.A., Biochem. Soc. Symp., 58 (1992) 149. 16. Bergquist, P.L. and Morgan, H.W., In Herbert, R.A. and
Sharp, R.S. (Eds.) Molecular Biology and Biotechnology of Extrernophiles, Blackie, Glasgow, 1993, pp. 44--75.
17. Vieille, C., Burdette, D.S. and Zeikus, J.G., Biotcchnol. Annu. Rev., 2 (1996) 1.
18. Hol, W.G.J., van Duijnen, P.T. and Berendsen, H.J.C., Nature, 273 (1978) 443.
19. Argos, P., Rossmann, M.G., Grau, U.M., Suborn, H., Frank, G. and Tratschin, J.D., Biochemistry, 18 (1979) 5698.
20. Davies, GJ., Gamblin, S.J., Littlechild, J.A. and Watson, H.C., Proteins, 15 (1993) 283.
21. Russell, R.J., Hough, D.W., Danson, M.J. and Taylor, G.L., Structure, 2 (1994) 1157.
22. Matthews, B.W., Nicholson, H. and Becktel, W.J., Proc. Natl. Acad. Sci. USA, 84 (1987) 6663.
23. Watanabe, K., Chishiro, K., Kitamura, K. and Suzuki, Y., J. Biol. Chem., 266 (1991) 24287.
24. Perutz, M.E and Raidt, H., Nature, 255 (1975) 256. 25. Perutz, M.E, Science, 201 (1978) 1187. 26. Walker, J.E., Wonacott, A.J. and Harris, J.I., Eur. J. Biochem.,
108 (1980) 581. 27. Hennig, M., Darimont, B., Sterner, R., Kirschner, K. and
Jansonius, J.N., Structure, 3 (1995) 1295. 28. Tanner, J.J., Hecht, R.M. and Kranse, K.L., Biochemistry, 35
(1996) 2597.

408
29. Kifino, H., Aoki, M., Aoshima, M., Hayashi, Y., Ohba, M., Yamagishi, A., Wakagi, T. and Oshima, T., Eur. J. Biochem., 220 (1994) 275.
30. Delboni, L.E, Mande, S.C., Rentierdelrue, F., Mainffoid, V., Turley, S., Vellieux, F.M.D., Martial, J.A. and Hol, W.G.J., Protein Sci., 4 (1995) 2594.
31. Salminen, T., Teplyakov, A., Kankare, J., Cooperman, B.S., Lahti, R. and Goldman, A., Protein Sci., 5 (1996) 1014.
32. Horovitz, A., Serrano, L., Avron, B., Bycrofl, M. and Fersht, A.R., J. Mol. Biol., 216 (1990) 1031.
33. Yip, K.S.P., Stillman, T.J., Britton, K.L., A_rtymiuk, P.J., Baker, P.J., Sedelnikova, S.E., Engel, P.C., Pasquo, A., Chiaraluce, R., Consalvi, V., Scandurra, R. and Rice, D.W., Structure, 3 (1995) 1147.
34. Korndorfer, I., Steipe, B., Huber, R., Tomschy, A. and Jaenicke, R., J. Mol. Biol., 246 (1995) 511.
35. Matthews, B.W., Weaver, LH. and Kester, W.R., J. Biol. Chem., 249 (1974) 8030.
36. Kraulis, P., J. Appl. Crystallogr., 24 (1991) 946. 37. Hurley, J.H., Baase, W.A. and Matthews, B.W., J. Mol. Biol.,
224 (1992) 1143. 38. Tomschy, A., B6hm, G. and Jaenicke, R., Protein Eng., 7
(1994) 1471. 39. Watanabe, K., Masuda, T., Ohashi, H., Mihara, H. and Suzuki,
Y., Eur. J. Biochem., 226 (1994) 277. 40. Moriyama, H., Onodera, K., Sakurai, M., Tanaka, N., Kirino,
K.H., Oshima, T. and Katsube, Y., J. Biochem. (Tokyo), 117 (1995) 408.
41. Biro, J., Fabry, S., Dietmaier, W., Bogedain, C. and Hensel, R., FEBS Lett., 275 (1990) 130.
42. Olsen, O., Borriss, R., Simon, O. and Thomsen, K.K., Mol. Gen. Genet., 225 (1991) 177.
43. Onodera, K., Sakurai, M., Moriyama, H., Tanaka, N., Numata, K., Oshima, T., Sato, M. and Katsube, Y., Protein Eng., 7 (1994) 453.
44. Numata, K., Muro, M., Akutsu, N., Nosoh, Y., Yamagishi, A. and Oshima, T., Protein Eng., 8 (1995) 39.
45. Kunkel, T.A., Roberts, J.D. and Zakour, R.A., Methods Enzymol., 154 (1987) 367.
46. Sambrook, J., Fritch, E.F. and Maniatis, T., Molecular Cloning: a Laboratory Manual, Cold Spring Harbor Labora- tory, Cold Spring Harbor, NY, 1989.
47. Bradford, M.M., Anal. Biochem., 72 (1976) 248. 48. Sanger, E and Coulson, A.R., J. Mol. Biol., 94 (1975) 441. 49. Laemmli, U.K., Nature, 227 (1970) 680. 50. Peterson, G.L., Anal. Biochem., 100 (1979) 201. 51. Wales, M.E. and Wild, J.R., Methods Enzymol., 202 (1991)
687. 52. Hjelmstad, R.H., Morash, S.C., McMaster, C.R. and Bell,
R.M., J. Biol. Chem., 269 (1994) 20995. 53. Korkhin, Y., Frolow, E, Bogin, O., Peretz, M., Kalb (Gilboa),
A.J. and Burstein, Y., Acta Crystallogr., D52 (1996) 882.




















