
Plasma triglyceride levels correlate with body weight, age and gender influence body weight changes...
11
Camp. B&hem. Physiol. Vol. 113A, No. 3, pp. 255-265, 1996 Copyright 8 1996 ElsevierScience Inc. ELSEVIER ISSN 0300-9629/96/$15.00 SSDI 0300-9629(96)ooO32-5 Plasma Triglyceride Levels Correlate with Body Weight, Age and Gender Influence Body Weight Changes in Spemtophilus Zateralis during Prehibemation and Hibernation 1. Yeh,’ S. Tsui,’ E. Catuiru, f T. T. Le, f V. Pupu, f IL.. Pena, f M. Vuscpez,f C. Vu, f S. Wang, f G. A. Lopez,f and C. F. Turn’ DEPARTMENTS OF ‘HEALTH AND NUTRITIONAL SCIENCES,AND BIOLOGY AND MICROBIOLOGY, CALIFORNIA STATE UNIVERSITY, Los ANGELES, 5151 STATE UNIVERSITY DRIVE, Los ANGELES, CA 90032, USA ABSTRACT’. Our experimental groups consisted of a total 16 male (M) and 15 female (F), juvenile and adult S~philus lareralis. This study evaluated weight gain rate (WGR) and mass-specific rate of gain (MSRG) during prehibemation, and weight loss rate (WLR), mass-specific rate of loss (MSRL) plus changes in the levels of plasma triglyceride (TG) during hibernation in these squirrels (SQ). Between January and March, the plasma samples were obtained from 26 SQ. PIasma TG was determined enzymatically, and it averaged 189 mg% with no significant difference in gender. Correlations were observed between plasma TG and sacrificed body weight (T = 0.441, p C 0.05), and between plasma TG and MSRL (r = 0.409, p < O.OS), although plasma TG was not correlated with WLR during hibernation. Juvenile female SQ had a higher WGR than adult female SQ, but no significant difference was found between juvenile and adult males. Both juvenile males and females showed a significantly higher MSRG than adult male and female SQ. Conversely, adult male SQ had a higher WLR and MSRL than juvenile male SQ, but no significant difference was observed between adult and juvenile females. Moreover, juvenile male SQ showed a significantly lower WLR but not MSRL than juvenile females. For these animals, our results suggest that the age factor influences on WGR and MSRG during prehibemation, and also influences on WLR and MSRL during torpor. COMP BIOCHEM PHYSIOL 113A;3:255-265, 1996. KEY WORDS. Adult, female, golden-mantled ground squirrel, hibernators, juvenile, male INTRODUCTION Hibernating animals are in a state of starvation (11) and undergo an extended period of inactivity (38). They expe- rience a change in lean body mass, in utilization of fat reserves, and in maintenance of glycogen in tissues during hibernation (14). The golden-mantled ground squirrel, Spennot>hilus (= Citellus) lateralis, exhibits endogenous rhythms of body weight changes (27,5) during its yearly cycle. The gain of body weight during summer activity is primarily due to changes in total body fat which increases threefold from the month of August to the month of Sep- Address reprint requests to: Dr. CF. Tam, Departmentof Health and Nutritional Sciences, CSULA, 5151 State University Dr., Los Angeles, CA 90032. Abbreviutkm-A = Adult, F = Female, J = Juvenile, LD = Light- dark, M = Male, MSRG = Mass-specific rate of gain, MSRL = Mass- specific rate of loss, OWT = Original body weight, PHWT = Prior to hibernation body weight, SQ = Squirrels, SWT = Sacrificed body weight, TG = Triglycerides, WGR = Weight gain rate, WLR = Weight loss rate. Received 1 Februav 1996. tember just prior to hibernation (15). The extra energy that is required for increasing body weight, specifically for the addition of adipose tissue mass, comes from overeating (hyperphagia) and reducing energy expenditure (inactiv- ity) as hibernation nears (16,25,26). The loss of body weight of Spermo~hilus lateralis during hibernation occurs in winter. During the period of winter dormancy, the squirrels do not eat at all or eat sparingly, and survive by greatly reducing energy expenditure and by a slow regulated depletion of accumulated fat stores in deep torpor (9). During hibernation, of the roused animals’ ef- forts, a very small percentage of time ,is devoted to eating or drinking (34). They eat only when they rouse rhythmi- cally, and their mean daily caloric intake is about 1 Kcal (16). The torpid state itself costs the animal very little amount of energy (14). In spite of the relatively very small number of hours (only 7.4% of total hibernation hours) with periodic arousals, the expenditure of energy already reaches about 90% of the total energy consumption in hi- bernating Cite&u citellur (18) and Spermophilus richhonii (37). In Spemophilus saturatus, the two-thirds of the year
-
Upload
www-cmac-sullana -
Category
Documents
-
view
2 -
download
0
Transcript of Plasma triglyceride levels correlate with body weight, age and gender influence body weight changes...

Camp. B&hem. Physiol. Vol. 113A, No. 3, pp. 255-265, 1996 Copyright 8 1996 Elsevier Science Inc.
ELSEVIER
ISSN 0300-9629/96/$15.00 SSDI 0300-9629(96)ooO32-5
Plasma Triglyceride Levels Correlate with Body Weight, Age and Gender Influence Body Weight
Changes in Spemtophilus Zateralis during Prehibemation and Hibernation
1. Yeh,’ S. Tsui,’ E. Catuiru, f T. T. Le, f V. Pupu, f IL.. Pena, f M. Vuscpez, f C. Vu, f S. Wang, f G. A. Lopez, f and C. F. Turn’
DEPARTMENTS OF ‘HEALTH AND NUTRITIONAL SCIENCES, AND BIOLOGY AND MICROBIOLOGY, CALIFORNIA STATE UNIVERSITY, Los ANGELES, 5151 STATE UNIVERSITY DRIVE, Los ANGELES, CA 90032, USA
ABSTRACT’. Our experimental groups consisted of a total 16 male (M) and 15 female (F), juvenile and adult S~philus lareralis. This study evaluated weight gain rate (WGR) and mass-specific rate of gain (MSRG) during prehibemation, and weight loss rate (WLR), mass-specific rate of loss (MSRL) plus changes in the levels of plasma triglyceride (TG) during hibernation in these squirrels (SQ). Between January and March, the plasma samples were obtained from 26 SQ. PI asma TG was determined enzymatically, and it averaged 189 mg% with no significant difference in gender. Correlations were observed between plasma TG and sacrificed body weight (T = 0.441, p C 0.05), and between plasma TG and MSRL (r = 0.409, p < O.OS), although plasma TG was not correlated with WLR during hibernation. Juvenile female SQ had a higher WGR than adult female SQ, but no significant difference was found between juvenile and adult males. Both juvenile males and females showed a significantly higher MSRG than adult male and female SQ. Conversely, adult male SQ had a higher WLR and MSRL than juvenile male SQ, but no significant difference was observed between adult and juvenile females. Moreover, juvenile male SQ showed a significantly lower WLR but not MSRL than juvenile females. For these animals, our results suggest that the age factor influences on WGR and MSRG during prehibemation, and also influences on WLR and MSRL during torpor. COMP BIOCHEM
PHYSIOL 113A;3:255-265, 1996.
KEY WORDS. Adult, female, golden-mantled ground squirrel, hibernators, juvenile, male
INTRODUCTION
Hibernating animals are in a state of starvation (11) and
undergo an extended period of inactivity (38). They expe- rience a change in lean body mass, in utilization of fat reserves, and in maintenance of glycogen in tissues during hibernation (14). The golden-mantled ground squirrel, Spennot>hilus (= Citellus) lateralis, exhibits endogenous rhythms of body weight changes (27,5) during its yearly cycle. The gain of body weight during summer activity is primarily due to changes in total body fat which increases threefold from the month of August to the month of Sep-
Address reprint requests to: Dr. CF. Tam, Department of Health and Nutritional Sciences, CSULA, 5151 State University Dr., Los Angeles, CA 90032.
Abbreviutkm-A = Adult, F = Female, J = Juvenile, LD = Light- dark, M = Male, MSRG = Mass-specific rate of gain, MSRL = Mass- specific rate of loss, OWT = Original body weight, PHWT = Prior to hibernation body weight, SQ = Squirrels, SWT = Sacrificed body weight, TG = Triglycerides, WGR = Weight gain rate, WLR = Weight loss rate.
Received 1 Februav 1996.
tember just prior to hibernation (15). The extra energy that is required for increasing body weight, specifically for the addition of adipose tissue mass, comes from overeating (hyperphagia) and reducing energy expenditure (inactiv- ity) as hibernation nears (16,25,26).
The loss of body weight of Spermo~hilus lateralis during hibernation occurs in winter. During the period of winter dormancy, the squirrels do not eat at all or eat sparingly,
and survive by greatly reducing energy expenditure and by a slow regulated depletion of accumulated fat stores in deep
torpor (9). During hibernation, of the roused animals’ ef- forts, a very small percentage of time ,is devoted to eating or drinking (34). They eat only when they rouse rhythmi- cally, and their mean daily caloric intake is about 1 Kcal
(16). The torpid state itself costs the animal very little amount of energy (14). In spite of the relatively very small number of hours (only 7.4% of total hibernation hours) with periodic arousals, the expenditure of energy already reaches about 90% of the total energy consumption in hi- bernating Cite&u citellur (18) and Spermophilus richhonii (37). In Spemophilus saturatus, the two-thirds of the year

256 I. Yeh et al.
spent in hibernation is only associated with 13-17% of annual energy expenditure (19). The reduction in body mass of S. later& during the weight loss phase is almost entirely due to an 84% decrease in total lipid reserves and reflects a decrease in adipocyte size but not number (9).
Although gender and age differences in weight gain and weight loss have been observed in both Spermophilus satura- tus (19) and Sgennophilus richurdsonii (23) during prehiber- nation activity and hibernation, most importantly, age and gender differences in the rate of weight gain and the rate of weight loss have not been tested in SPermophilus lateralis during prehibemation and hibernation.
The golden-mantled ground squirrel, Spermophilus later- alis, undergoes dramatic weight changes mainly from a re- duction of total lipid reserves during its annual cycle (9). During prehibemation in the summer, ground squirrels in- crease in plasma insulin and glucocorticoids such as corti- costerone and cortisol, which are found to be at peak levels prior to hibernation (31,6). Insulin is well known for its role in glycogenesis, lipogenesis, and protein synthesis. Similarly, glucocorticoids which are associated with prehi- bemating fattening (31,1) serve to stimulate lipogenic en- zymes (4), maintain obesity in laboratory rodents (20)) and possibly promote feeding behavior (12). Human obesity is proposed to be due to endogenous factors, such as insulin and glucocorticoids (33,41,13,29,2,7). Hence, this animal can be considered an excellent animal model for the study of the interrelationships between the regulation of obesity, adipose tissue mass, and lipid metabolism.
In view of the lack of studies done to explore the rela- tionship between plasma triglyceride levels and body weight changes in the golden-mantled ground squirrel dur- ing hibernation, the rationale for doing this study is:
1. to test the influences of age and gender on the rate of weight gain and the rate of weight loss in S. later&, during the period of prehibemation and hibernation, and
2. to examine the correlations among plasma triglyceride level, weight loss rate, mass-specific rate of loss and sacrificed body weight in the golden-mantled ground squirrel during the hibernating phase.
MATERIALS AND METHODS 1. Experimental Aninuds
The wild ground squirrels enter hibernation when snow falls and rouse from hibernation (emergence) when snow melts (8,22). No squirrels are observed to be present from late November to early March at 5,000 ft elevation or to April at 6,000 ft elevation in years of normal snowfall (22). It has been reported that the emergence time of golden- mantled ground squirrels trapped from four different eleva- tions demonstrates that wild adult squirrels emerge in late March to late May, whereas juvenile squirrels emerge in late June to early August (8). During the months of January
and March, the ground squirrel is in the deep torpid stage and near the emergent stage. We designed our experimen- tal environment as closely as possible to the environment of the wild for our animals. Under our experimental design, by placing the animals in a cold room between the months of November and March, they should have completed their normal hibernation cycle similar to living in the wild.
In the month of August, a total of 16 male and 15 female golden-mantled ground squirrels, Spermo~hilus lateralis, were trapped from the mountains near Redding, CA, and housed individually in wire cages in an animal room with a light-dark (LD) cycle of 12 : 12 at 22°C and ambient relative humidity. Distilled water and Wayne Rodent Blox (24% protein, 6% fat, 4.5% fiber, and approximately 2.2 mEq Na) were available ad Ii&turn during the prehibemat- ing phase. Age identification was made on the first day the animals arrived according to body mass, body length, and appearance (10,22). Animals were considered to be juve- niles until they reached their first reproductive season (April-May) (22). After arrival, animals older than 1 year were grouped collectively as adults. Of the 31 squirrels, there were 7 juveniles and 9 adults in the male group, and 4 juveniles and 9 adults in the female group, whereas 2 female squirrels cannot be identified with certainty as be- longing to either the juvenile or the adult groups. These 2 female squirrels were excluded from our data analyses pertaining to the influences of age. In addition, we ex- cluded 2 squirrels: 1 adult female which did not become torpid and 1 adult male which awoke shortly before sacri- ficing, from our data analyses for body weight prior to hi- bernation. In the fall, the squirrels in their individual wire cages were moved to a cold room at 6°C with an LD cycle of 0: 24. Distilled water and Wayne Rodent Blox were also available ad libitum during hibernating period. Cotton was made available for nesting. We finished the sacrifice of all animals by late March.
2. Measurements of Body Weight
Body weight was measured to the nearest 0.1 g on the first day the animals arrived on campus in the month of August (original body weight, OWT), on the day prior to being placed in the cold room in mid-November (prior to hiber- nation body weight, PHWT), and on the days of sacrificing between the month of January and the month of March (sacrificed body weight, SWT). The weight gain rate (WGR; g/day) for animals in the active, prehibemation period was obtained from the difference in body weight between OWT and PHWT divided by the number of days elapsed. The mass-specific rate of gain (MSRG; %/day) was calculated by the percent change of body weight gain between OWT and PHWT divided by the number of days elapsed. The weight loss rate (WLR; g/day) for hibernating animals was similarly calculated from the difference in body weight between PHWT and SWT divided by the number

Triglycerides and Body Weight in Squirrels 257
of days elapsed. The mass-specific rate of loss (MSRL; %/ day) as described by Michener and Locklear (23) was also calculated by the percent change of body weight loss be- tween PHWT and SWT divided by the number of days elapsed. Twente and Twente (35) reported that the han- dling of animals can cause differences between the sexes with respect to their hibernating behavior. In our experi- ments, we therefore measured body weight at limited time points in order to minimize the influences of handling in our pre-hibernating and hibernating animals.
TABLE 1. The mean original body weight (OWT) of four subgroups, juvenile male and female, adult male and fe- male, and of combined gender of Spemophib lateralis
Age OWT’ gender OWTT
Gender N (31) (8) (a)
Juvenile Male 7 113.7 + 3.1 112.1 * 2.2 Female 4 109.2 + 2.2
Adult Male 9 152.3 * 3.9 154.6 + 2.7 Female 9 156.8 + 3.8
Uncertain Female 2 137.4 2 1.7
3. Measurement of Plasma Triglycerides
Between the months of January and March, some deeply torpid animals were selected by their gender, age, and weight-matched and then sacrificed by decapitation in weekly intervals. No animal was aroused during the han- dling of the weight-matched procedure to minimize the influences of the frequency of handling that the animals received during the hibernating period before they were sacrificed. The plasma samples were obtained from 26 ani- mals (13 males and 13 females). The blood was collected immediately following decapitation and placed in vials containing sodium heparin. The blood sample was then centrifuged at 5,000 rpm for 15 min and the plasma portion was removed using the Pasteur pipet. The plasma samples were immediately frozen at - 20°C until assays and re- centrifuged at 5,000 rpm for 5 min before assays.
The plasma triglycerides (TG) concentration was deter- mined enzymatically as described by Sigma Diagnostics Procedure No. 337-B (St Louis, MO), which was a modi- fied method of McGowan et al. (21). Each sample was assayed in duplicate at 30°C.
4. Analyses of Data
Data were analyzed using a commercial statistical program SPSS for Windows v. 6.1 (SPSS, Inc., 1994). All results were expressed as means +- SE. Statistical comparisons for the differences in WGR, MSRG, WLR, MSRL, and plasma TG levels of the experimental animals as influenced by gender or age were performed by the Student’s unpaired t-test. Differences in WLR, MSRL, and the plasma con- centration of triglycerides among the months of January, February, and March during hibernation were tested by one-way ANOVA. Using two-way ANOVA, we further analyzed whether the factor of age: juvenile and adult are equally effective, and the factor of gender: male and female are also equally effective in influencing the rate of gain or loss in body weight during prehibemation or hibernation, and how the factors of age and gender combine or interact to influence our response measurement such as WGR, MSRG, WLR, or MSRL (32). The Pearson correlation coefficient was used to test for any bivariate relationship among the variables. Statistical differences were considered to be significant at a level of fi < 0.05 or lower.
Values are means f SE. ‘p < 0.001 Juvenile male vs. adult male; juvenile male vs. adult female;
juvenile female vs. adult female; and juvenile female vs. adult male. ?p < 0.001 Juvenile vs. adult.
RESULTS
Of the initial 31 experimental animals, 2 females were excluded from our data analyses regarding the effects of age because of the uncertainty of their age. The results of mean original body weight (OWT) of the four subgroups, juve- nile male and female, adult male and female, of S@rmophi- 1u.s lateralis are shown in Table 1. In general, the mean OWT of the adult squirrels was significantly higher than that of the juveniles in the month of August. The rankings of the cohorts from the heaviest to the lightest mean OWT showed the sequence as such: adult females 2 adult males > juvenile males 2 juvenile females. Two of the remaining 29 experimental animals were not included in our final data analyses because they either did not become torpid (1 adult female) or awoke shortly before sacrifice (1 adult male). By the time the squirrels (N = 27) entered hibema- tion, both juvenile and adult groups had attained body weights of similar magnitude (Table 2). The rankings of the cohorts from the heaviest to the lightest mean prior hibernation body weight (PHWT) showed the sequence as such: juvenile females 2 adult males > adult females 2 juvenile males (Table 2). Between males and females, there were no significant differences in OWT, PHWT, and SWT of these animals (Table 3).
1. Weight @in Rate (WGR) and Mass-Specijic Rate of &in (MSR@
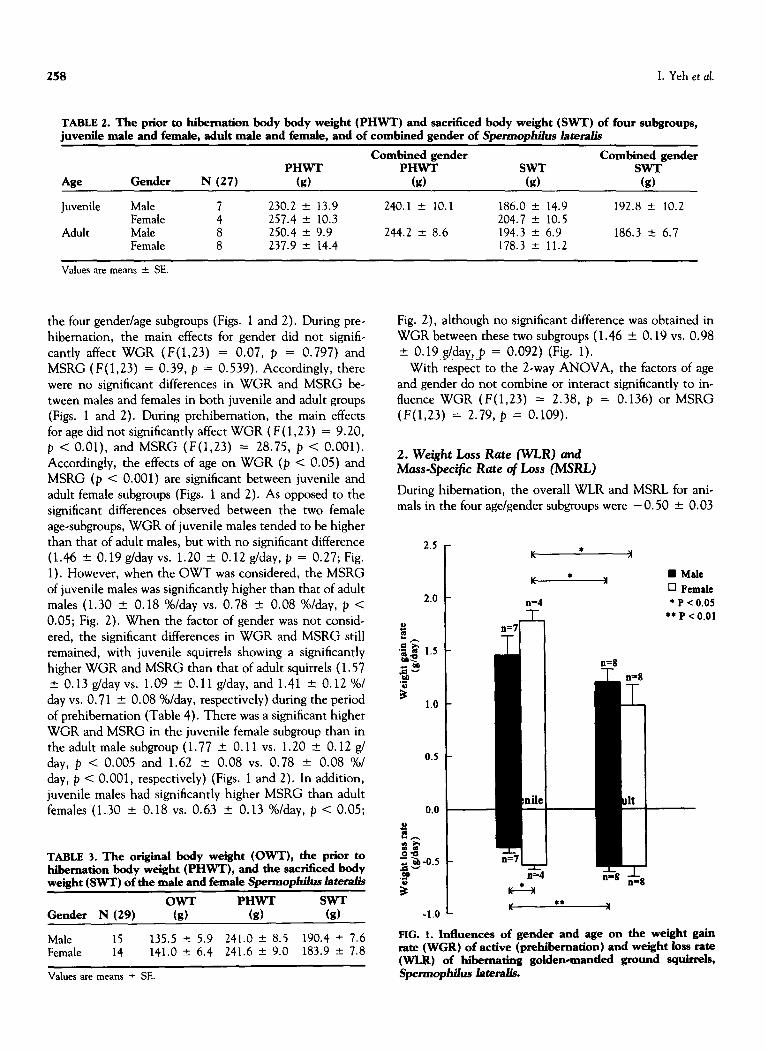
Overall, active golden-mantled ground squirrels (Sgertno- p/&s later&) of all subgroups had mean WGR of 1.27 + 0.09 grams per day (range 0.35 to 2.45 g/day) and mean MSRG of 0.98 2 0.09 %/day (range 0.21-2.20%/day) during the period of prehibemation from the month of August to the month of November.
The juvenile female squirrels had the highest WGR (1.77 + 0.11 g/day) and MSRG (1.62 2 0.08 %/day), while the adult female squirrels had the lowest WGR (0.98 f 0.19 g/day) and MSRG (0.63 + 0.13 %/day) among
258 I. Yeh et al.
TABLE 2. The prior to hibernation body body weight (PHWT) and sacrificed body weight (SWT) of four subgroups, juvenile male and female, adult male and female, and of combined gender of Spemophihs hterafis
Combined gender Combined gender PHWT PHWT SWT SWT
Gender N (27) (9) (g) (g) (g)
Juvenile Male 7 230.2 k 13.9 240.1 k 10.1 186.0 t 14.9 192.8 k 10.2 Female 4 257.4 2 10.3 204.7 2 10.5
Adult Male 8 250.4 * 9.9 244.2 * 8.6 194.3 k 6.9 186.3 2 6.7 Female 8 237.9 * 14.4 178.3 * 11.2
Values are means f SE.
the four gender/age subgroups (Figs. 1 and 2). During pre- hibernation, the main effects for gender did not signifi-
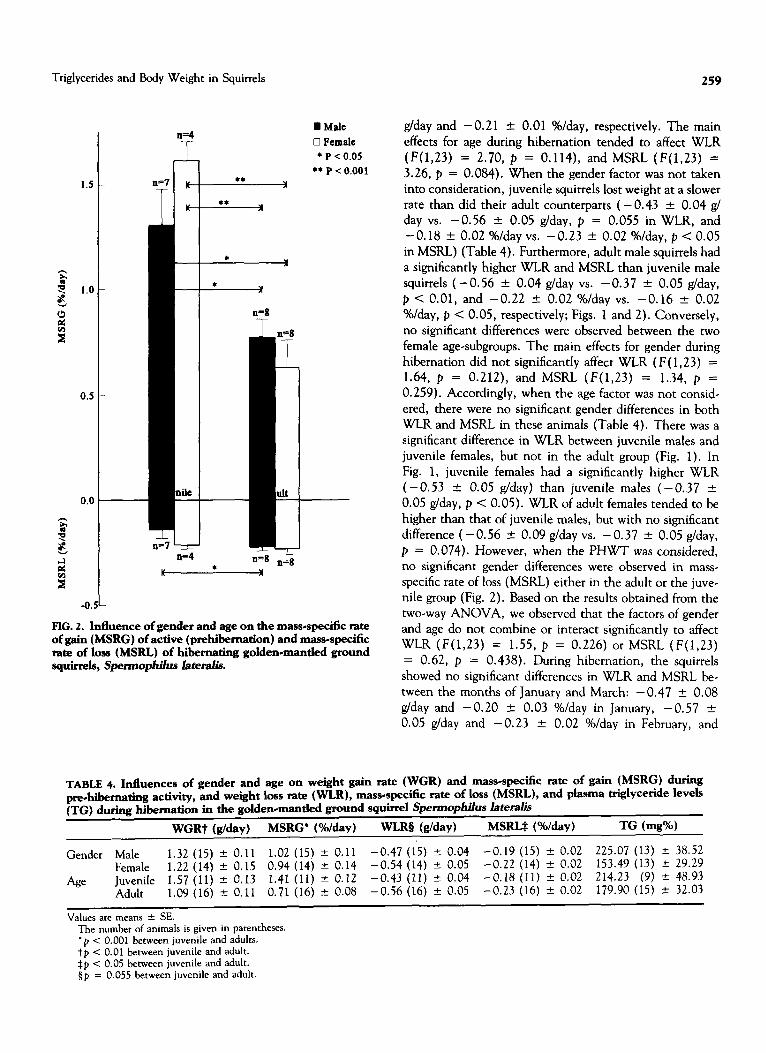
cantly affect WGR (F(1,23) = 0.07, p = 0.797) and MSRG (F(1,23) = 0.39, p = 0.539). Accordingly, there were no significant differences in WGR and MSRG be- tween males and females in both juvenile and adult groups (Figs. 1 and 2). During prehibemation, the main effects for age did not significantly affect WGR ( F( 1,23) = 9.20, p < O.Ol), and MSRG (F(1,23) = 28.75, p < 0.001). Accordingly, the effects of age on WGR (p < 0.05) and MSRG (p < 0.001) are significant between juvenile and adult female subgroups (Figs. 1 and 2). As opposed to the significant differences observed between the two female age-subgroups, WGR of juvenile males tended to be higher than that of adult males, but with no significant difference (1.46 t O.l9g/dayvs. 1.20 k O.l2g/day,p = 0.27;Fig. 1). However, when the OWT was considered, the MSRG of juvenile males was significantly higher than that of adult males (1.30 + 0.18 %/day vs. 0.78 + 0.08 %/day, p < 0.05; Fig. 2). When the factor of gender was not consid- ered, the significant differences in WGR and MSRG still remained, with juvenile squirrels showing a significantly higher WGR and MSRG than that of adult squirrels ( 1.57 ‘-c 0.13 g/day vs. 1.09 f 0.11 g/day, and 1.41 + 0.12 %/ day vs. 0.7 1 k 0.08 %/day, respectively) during the period of prehibemation (Table 4). There was a significant higher WGR and MSRG in the juvenile female subgroup than in the adult male subgroup (1.77 2 0.11 vs. 1.20 “- 0.12 g/ day, p < 0.005 and 1.62 2 0.08 vs. 0.78 +- 0.08 %/ day, p < 0.001, respectively) (Figs. 1 and 2). In addition, juvenile males had significantly higher MSRG than adult females (1.30 + 0.18 vs. 0.63 k 0.13 %/day, p < 0.05;
TABLE 3. The original body weight (OWT), the prior to hibernation body weight (PHWT), and the sacrificed body weight (SWT) of the male and female Spermophirus ~teralis
OWT PHWT SWT Gender N (29) (g) (8) (g)
Male 15 135.5 k 5.9 241.0 2 8.5 190.4 ? 7.6 Female 14 141.0 k 6.4 241.6 + 9.0 183.9 2 7.8
Values are means 2 SE.
Fig. 2), although no significant difference was obtained in WGR between these two subgroups (1.46 t 0.19 vs. 0.98 k 0.19 g/day, p = 0.092) (Fig. 1).
With respect to the 2-way ANOVA, the factors of age and gender do not combine or interact significantly to in- fluence WGR (F(1,23) = 2.38, p = 0.136) or MSRG (F(1,23) = 2.79, p = 0.109).
2. Weight Loss Rate IWLR) and Mass-Specijic Rate of Loss (MARL)
During hibernation, the overall WLR and MSRL for ani- mals in the four age/gender subgroups were - 0.50 k 0.03
2.5
2.0
u P i
*
: n1
*
a=4
II=7 l-5 ii_ $$j 1.5 - 00. 25! .f$
3 1.0 -
0.5 -
0.0
s t, “,P
pJo.s -
.9
i
-1.0 -
ll=S
n-8 c 1t
a-8 ,,-8
K l *
;I
n Male
q Female l P < 0.05 l * P < 0.01
FIG. 1. Influences of gender and age on the weight gain rate (WGR) of active (prehibemation) and weight loss rate (WL.R) of hibernating golden-mantled ground squirrels, spemlophilus lateralis.
Triglycerides and Body Weight in Squirrels
n Male
0 Female l P < 0.05
l * P < 0.001 .*
)I
** 1
l 1
r8
T
* n=8 ,,,8
.I
FIG. 2. Influence of gender and age on the mass-specific rate of gain (MSRG) of active (prehibernation) and mass-specific rate of loss (MSRL) of hibernating golden-mantled ground squirrels, spermophiius lateralis.
259
g/day and - 0.21 of: 0.01 %/day, respectively. The main effects for age during hibernation tended to affect WLR
(F(1,23) = 2.70, p = 0.114), and MSRL (F(1,23) = 3.26, p = 0.084). When the gender factor was not taken into consideration, juvenile squirrels lost weight at a slower rate than did their adult counterparts ( - 0.43 * 0.04 g/ day vs. -0.56 + 0.05 g/day, p = 0.055 in WLR, and - 0.18 + 0.02 %/day vs. -0.23 2 0.02 %/day, p < 0.05 in MSRL) (Table 4). Furthermore, adult male squirrels had a significantly higher WLR and MSRL than juvenile male squirrels (-0.56 + 0.04 g/day vs. -0.37 + 0.05 g/day, p < 0.01, and -0.22 + 0.02 %/day vs. -0.16 + 0.02 Oh/day, p < 0.05, respectively; Figs. 1 and 2). Conversely, no significant differences were observed between the two female age-subgroups. The main effects for gender during hibernation did not significantly affect WLR (F( 1,23) = 1.64, p = 0.212), and MSRL (F(1,23) = 1.34, p = 0.259). Accordingly, when the age factor was not consid- ered, there were no significant gender differences in both WLR and MSRL in these animals (Table 4). There was a significant difference in WLR between juvenile males and juvenile females, but not in the adult group (Fig. 1). In Fig. 1, juvenile females had a significantly higher WLR (-0.53 -t 0.05 g/day) than juvenile males (-0.37 * 0.05 g/day, p < 0.05). WLR of adult females tended to be higher than that of juvenile males, but with no significant difference (-0.56 * 0.09 g/day vs. -0.37 _t 0.05 g/day, p = 0.074). However, when the PHWT was considered, no significant gender differences were observed in mass- specific rate of loss (MSRL) either in the adult or the juve- nile group (Fig. 2). Based on the results obtained from the two-way ANOVA, we observed that the factors of gender and age do not combine or interact significantly to affect WLR (F(1,23) = 1.55, p = 0.226) or MSRL (F(1,23) = 0.62, p = 0.438). During hibernation, the squirrels showed no significant differences in WLR and MSRL be- tween the months of January and March: - 0.47 + 0.08 g/day and -0.20 2 0.03 %/day in January, -0.57 f 0.05 g/day and - 0.23 * 0.02 %/day in February, and
TABLE 4. Influences of gender and age on weight gain rate (WGR) and mass-specific rate of gain (MSRG) during pm.Ebmahg activity, and weight loss rate (WLR), mass*specific rate of loss (MSRL), and plasma triglyceride levels (TG) during hibernation in the golden-mantled ground squirrel spemophirus h~terafh
WGRt (g/day) MSRG’ (%/day) wLR9 (g/day) MSRU (%/day) TG (mg%)
Gender Male 1.32 (15) * 0.11 1.02 (15) * 0.11 -0.47 (15) t 0.04 -0.19 (15) 2 0.02 225.07 (13) 2 38.52 Female 1.22 (14) f 0.15 0.94 (14) + 0.14 -0.54 (14) * 0.05 -0.22 (14) k 0.02 153.49 (13) 2 29.29
Age Juvenile 1.57 (11) -C 0.13 1.41 (11) k 0.12 -0.43 (11) * 0.04 -0.18 (11) c 0.02 214.23 (9) -t 48.93 Adult 1.09 (16) r 0.11 0.71 (16) + 0.08 -0.56 (16) 2 0.05 -0.23 (16) 2 0.02 179.90 (15) k 32.03
Values are means f SE. nhe number of animals is given in parentheses. ‘p < 0.001 between juvenile and adults. tp < 0.01 between juvenile and adult. $p < 0.05 between juvenile and adult. @p = 0.055 between juvenile and adult.
260 I. Yeh et al.
3oc
25a
50
.
0
t-
,-
I
100 I I
200 300
Plasma triglycerides concentration (mg%)
I
400
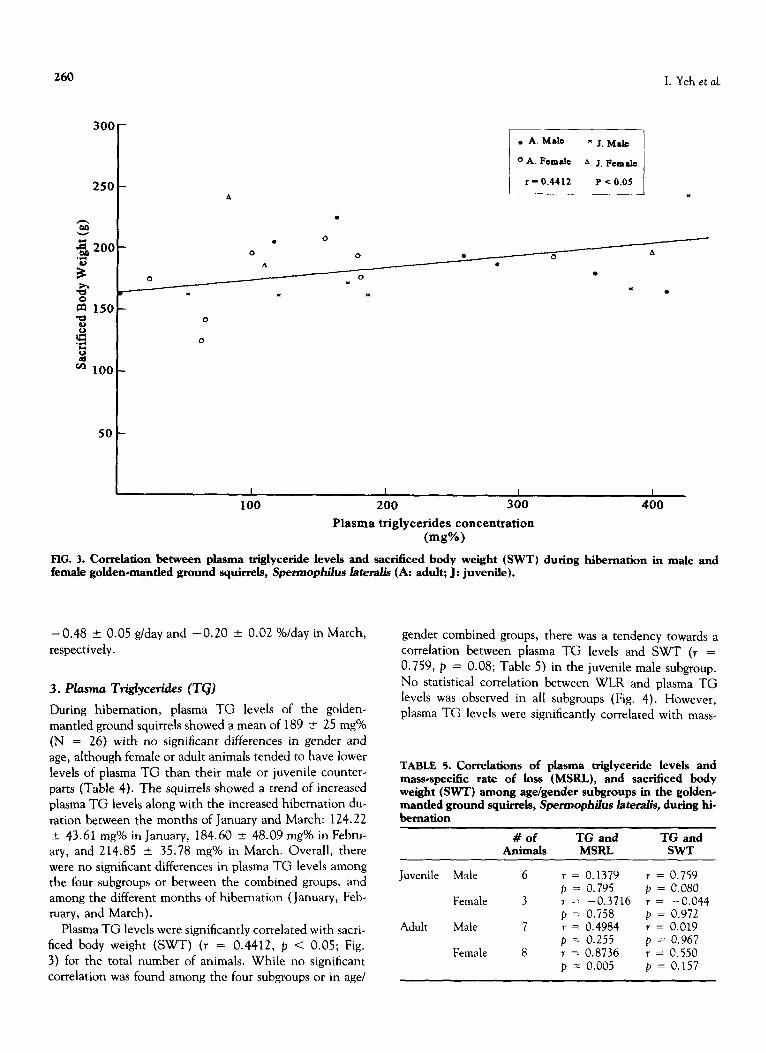
FIG. 3. Correlation between plasma triglyceride levels and sacrificed body weight (SWT) during hibernation in male and female golden*mantled ground squirrels, Spermophilus lateralis (A: adult; J: juvenile).
-0.48 +- 0.05 g/day and -0.20 k 0.02 %/day in March,
respectively.
3. Pkma Triglycerides (TGI
During hibernation, plasma TG levels of the golden- mantled ground squirrels showed a mean of 189 * 25 mg% (N = 26) with no significant differences in gender and age, although female or adult animals tended to have lower levels of plasma TG than their male or juvenile counter- parts (Table 4). The squirrels showed a trend of increased plasma TG levels along with the increased hibernation du- ration between the months of January and March: 124.22 -C 43.61 mg% in January, 184.60 k 48.09 mg% in Febru- ary, and 214.85 + 35.78 mg% in March. Overall, there were no significant differences in plasma TG levels among the four subgroups or between the combined groups, and among the different months of hibernation (January, Feb- ruary, and March).
Plasma TG levels were significantly correlated with sacri- ficed body weight (SWT) (r = 0.4412, p < 0.05; Fig. 3) for the total number of animals. While no significant correlation was found among the four subgroups or in age/
gender combined groups, there was a tendency towards a correlation between plasma TG levels and SWT (r = 0.759, p = 0.08; Table 5) in the juvenile male subgroup. No statistical correlation between WLR and plasma TG levels was observed in all subgroups (Fig. 4). However, plasma TG levels were significantly correlated with mass-
TABLE 5. Correlations of plasma triglyceride levels and mass*specitic rate of loss (MSRL), and sacrificed body weight (SWT) among age/gender subgroups in the golden- mantled ground squirrels, Spermophilus laterals, during hi- bernation
# of TG and Animals MSRL
TG and SWT
Juvenile Male 6 r = 0.1379 I = 0.759 p = 0.795 p = 0.080
Female 3 I = -0.3716 r = -0.044 p = 0.758 p = 0.972
Adult Male 7 r = 0.4984 r = 0.019 p = 0.255 p = 0.967
Female 8 r = 0.8736 r = 0.550 p = 0.005 p = 0.157
Triglycerides and Body Weight in Squirrels 261
0
-0.2
-0.8
-1.0
r 100
I
Plasma triglycerides concentration
(ms% 200 300
I 400
x
0
A *
0
A.
0
*
. ‘J * .
0
IL
.
*
A .
. 0
*A. Malo ” J.Mdo
0 A. Fsmolo A J. Fando r = 0.236 P = 0.246
I
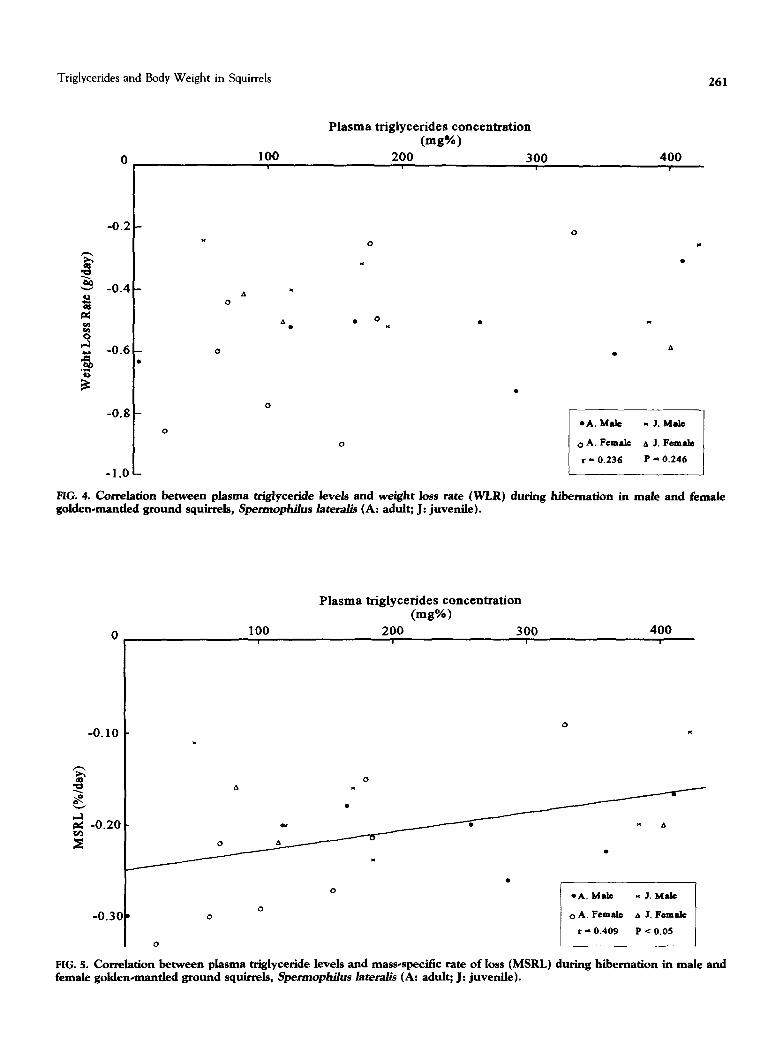
FIG. 4. Correlation between plasma triglyceride levels and weight loss rate (WLR) during hibernation in male and female
golden-mantled ground squirrels, Spermophiius lateralis (A: adult; J: juvenile).
0
x
-0.31
100
Plasma triglycerides concentration (mg%)
200 300 I I
400
0 x
A 0
*
.
* A
.
*
0
0 I 0
0
.
.A. Mdo * J.Mdo
0 A. Fomalo A J. Fern&
r - 0.409 P < 0.05
FIG. 5. Correlation between plasma triglyceride levels and mass-specific rate of loss (MSRL) during hibernation in male and female golden-mantled ground squirrels, Spermophihs lateral% (A: adult; J: juvenile).

262 I. Yeh et al.
specific rate of loss (MSRL) (T = 0.409, p < 0.05; Fig. 5) for the total number of animals. This correlation was also found in the adult female squirrels (Table 5).
DISCUSSION
It is noteworthy that the influences of age and gender on weight gain rate (WGR), mass-specific rate of gain (MSRG), weight loss rate (WLR), and mass-specific rate of loss (MSRL) have never been tested in Spermophilw lateralis during prehibemation and hibernation. The mean of OWT of the juveniles were found to be lower when compared to that of the adult ground squirrels (Table 1). Moreover, our results of PHWT of adult males and females (Table 2) were consistent with that of McKeever’s (22) study. In our present study, adult males tended to be heav- ier in PHWT than adult females. Conversely, juvenile fe- male squirrels were heavier than juvenile males in PHWT, although no significant difference was found (Table 2). Our results were consistent with other observations of body weights at peak values (224.6 g and 231.0 g) in young golden-mantled ground squirrels obtained from two coun- ties of California (28). However, there was no report on body weight in adult squirrels at all in Phillips’ (28) study. We further observed that the juvenile squirrels reached similar body weight of the adult squirrels (240.1 t 10.1 and 244.2 * 8.6 g, respectively) on entry into hibernation (Table 2). When age was not considered, there were no significant differences between female and male animals in OWT, PHWT, and SWT (Table 3).
1. Weight @sin Rate (WGR) and Mass-Specific Rate of Gain (MSRCj)
The factor of age had a strong influence on the weight gain of S. lateralis during prehibemating activity in this study, for WGR, F(l, 23) = 9.20 (p < 0.01) for MSRG, F(l, 23) = 28.75 (p < 0.001). With respect to the effect of age, the mean WGR and the mean MSRG of juveniles, including male and female, was higher than that of adult animals (Table 4).
The range of WGR of juvenile animals, including male and female, observed in our study was between 0.85 to 2.45 g/day which was lower than 1.2 to 3.7 g/day of juve- nile animals raised under laboratory conditions (22), whereas our age and gender-pooled mean WGR 1.27 g/day was less than that of 3.2 g/day observed in McKeever’s (22) study. McKeever mentioned that the average gain 3.2 g/ day for 30 animals was similar to the average gain of juve- nile squirrels raised in captivity after they began eating solid foods. The difference in age and gender-pooled WGR between McKeever’s and our study may be due to the inclu- sion of a different proportion of juvenile squirrels in rela- tion to adult animals as well as different living environ- ments. With respect to McKeever’s study, both Labration
(a product of Diablo Laboratories, San Francisco) and rolled oats were provided each animal ad l&turn. We fed our animals with Wayne Rodent Blox (24% protein, 6% fat, 4.5% fiber, and approximately 2.2 mEq Na) ad libitum. In addition, McKeever’s laboratory condition may not be similar to ours. Such differences of WGR could be contrib- uted by different diets and laboratory conditions. We fur- ther analyzed the differences in WGR between male and female, and juvenile and adult (Table 4) which were never shown in McKeever’s study. Juvenile squirrels were gener- ally less heavy than adults (Table 1). Since they continue to grow and gradually increase their basic body weight until they enter hibernation (15), this may account for the higher WGR in juveniles than in adults observed in our study (Table 4 and Fig. 1). It is noteworthy that WGR of juvenile males was not significantly higher than that of adult males (Fig. l), however, when the original body weight (OWT) was considered, MSRG of juvenile males was significantly higher than that of adult males (Fig. 2). It is important, therefore, to include mass-specific rate of gain (MSRG) for the evaluation of the rate of gain in these animals.
The gender factor had no significant influence on WGR in the present study (F(1,23) = 0.07, p = 0.797). The effect of gender was the same for juvenile and adult animals in relation to WGR and MSRG during prehibemation. Furthermore, according to t-test, the mean WGR and the mean MSRG were similar between male and female ani- mals (Table 4).
In our study, juvenile males and females did increase in weight by 102% and 135.6%, respectively, whereas adult males and females did increase in weight by 62% and 51%, respectively. This was similar to McKeever’s (22) observa- tion in adult squirrels. McKeever showed increases in WGR of 68% in adult males and 59% in adult females, but did not show the rate of weight gain in juvenile squirrels.
The significant difference in WGR or MSRG found be- tween the juvenile and adult animals in this study might be mainly contributed by the juvenile female squirrels (Ta- ble 4). This may be reflective of age and gender-influenced variations in intermediary metabolism, in addition to gen- eralized growth, during the juvenile period.
2. Weight Loss Rate (WLR) and Mass-Specific Rate of Loss (MSRL)
During hibernation, the overall WLR (-0.50 f. 0.03 g/ day) for animals in the four age/gender subgroups in this study was again similar to previous studies (36,22). Wade (36) studied WLR in a combination of three species, C. tridecemlineatus, C. frankinii, and C. late&is. Our study em- phasized on the WGR, MSRG, WLR, and MSRL in S. lateralis ( = C. later&s). We futher compared differences in four subgroups (juvenile male, juvenile female, adult male, and adult female) (Figs. 1 and 2) which were not

Triglycerides and Body Weight in Squirrels 263
observed by McKeever (22). WLR during hibernation for all juveniles in the present study was -0.43 g/day (Table 4), which was similar to the WLR of - 0.44 g/day reported by Phillips’ (28) study. In addition, the WLR of juvenile was less than the adult group ( - 0.43 g/day vs. - 0.56 g/ day, p = 0.055) (Table 4). It is noteworthy that there was no report on WLR of adult hibernating squirrels in Phillips’
(28) study. The rankings of the cohorts from the heaviest to the
lightest mean SWT showed the sequence to be: juvenile females > juvenile males > adult males > adult females (Table 2). Based on our study, the effect of age was pro- nounced on WLR and MSRL of S. later& during hibema- tion, whereas the factor of gender had no significant influ- ence on the rate of body weight loss (Table 4). This corresponded with our ANOVA results on the main effect of age or gender. In the juvenile group, males had a signifi- cantly lower WLR than their female counterparts (Fig. 1). Males arise from hibernation earlier (22,8) and the ener- getic cost of mating for males is higher (24). The juvenile male lost weight slower than the other three subgroups (Fig. 1 and 2), suggesting the fact that hibernating juvenile males may retain more weight and/or residual fat ready to be used for the more energetic costs of mating efforts after hibernation, while juvenile females may use energy derived from body fat to attain reproductive capability earlier than their male counterparts. A previous study of the Richard- son’s ground squirrels done by Michener and Locklear (23) demonstrates that juvenile males have a markedly lower rate of over-winter loss of mass relative to structural mass than other subgroups. Similarly, from our study of Spermo- philus later&, we found that juvenile males had a markedly lower WLR and MSRL than the other three subgroups, in particular, when compared to male adult squirrels (Figs. 1 and 2). Our results suggest that juvenile male S. lateralis could have a lower metabolic rate than the others during hibernation, therefore, they would maintain comparable mass for growth to compete with the older males for mating in the wild.
In addition, the greater WLR and MSRL observed in the adult animals in this study (Table 4) may reflect the fact that adults, having completed growth, can better man- age the energy for maintenance of fat deposition only, whereas the juveniles must still partition energy between somatic growth and fattening (19,23).
3. Pbsmu Triglycerides (Tcj) and Sacrificed Body Weight (SWT] during Hibernation
There were no significant age or gender influences on plasma TG levels in different subgroups. From January to March, the squirrels tended to have an increase in plasma TG levels along with the increase of hibernation time. It might reflect the fact that most plasma TG levels were progressively being released into the plasma by these ani- mals during hibernation.
When either the age or gender factor was not con- sidered, we observed no significant correlation between plasma TG levels and sacrificed body weight (SWT) among all subgroups due to small sample size (Table 5). However, a significant correlation between SWT and plasma triglyc- eride levels was observed when we combined the total number of animals (Fig. 3). Our finding was consistent with a report of increased circulating level of triglycer- ides, related to higher body weight, in either genetic or hypothalamic obese rats (3). We also observed a cor- relation between MSRL and plasma triglyceride levels in the total group of animals (Fig. 5). This result may indicate that the lower the MSRL (the higher body weight hence maintained), the higher the amount of TG is being released into the plasma by these animals during hibernation. In view of the fact that we observed the markedly lower WLR and MSRL in juvenile males during hibernation (Figs. 1 and Z), as well as a tendency toward a correlation between their SWT and plasma TG levels (Table 5), our results may imply that these juvenile males could deplete their triglyceride reserves at a slower rate than the others, which may, in turn, contribute to or account for a lower metabolic inten- sity (23).
It has been suggested that plasma insulin functions to signal the amount of body fat to the central nervous system (40,30), and to affect the gene expression of enzymes re- sponsible for fat metabolism (39). Similarly, glucocorti- coids which are associated with prehibemating fattening (3 1,1) serves to stimulate lipogenic enzymes (4), maintain obesity in laboratory rodents (20), and possibly promote feeding behavior (12). These hormones play similar roles in humans such that it may also induce weight gain attrib- utable to fat (7). The plasma insulin levels were highly correlated with body weight in hibernating S. later& (42). Since controlling feeding behavior and fat storage are criti- cal to the survival of hibernating ground squirrels, espe- cially during the summer, the discovery or determination of the controlling factor may lead to the ability to control satiety and obesity in humans. S. lateralis succeed in protein sparing on a very low calorie diet where obese people fail, suggesting obese people should reduce food intake pro- gressively from a low calorie diet to a very low calorie diet similar to what ground squirrels do spontaneously (17). It is worthy to investigate whether this animal that spontane- ously overeats and fasts as part of its natural life cycle is able to achieve more efficient regulation of body weight than humans.
In conclusion, for the first time, we have reported that in S. later&, the factor of age had influence on WGR and MSRG during prehibemation activity, and also on WLR and MSRL during torpor. During hibernation, plasma tri- glyceride levels and SWT as well as plasma TG and MSRL were significantly correlated in the golden-mantled ground squirrels.

264 I. Yeh et al.
This work was supported in part by NIGMS-MBRS grant #08101. We would like to eeess our sincere thanks to Genevieve Duncan for her editorial commentS of this paper.
20.
21.
King, B.M. Glucocorticoids and hypothalamic obesity. Neu- rosci Biobehav. Rev., 12:29-37;1988. McGowan, M. W.; Artiss, J.D.; Standbergh, D.R.; Zak, B. A peroxidase-coupled method for the calorimetric determi- nation of serum triglycerides. Clin. Chem. 29:538-542; 1983.
References 1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
Armitage, K. B. Factors affecting corticosteroid concentra- tions in yellow-bellied marmots. Comp. Biochem. Physiol. 98A:47-54;1991. Bailey, C. J.; Day, C.; Lipson, L. ; Bray, G. A. Role of adrenal glands in the development of abnormal glucose and insulin homeostasis in genetically obese (ob/ob) mice. Horm. Meta- bol. Res. 18:357-360;1987. Barry, W.S.; Bray, G.A. Plasma triglycerides in genetically obese rats. Metabolism 18:833-839;1969. Berdanier, C.D. Role of glucocorticoids in the regulation of lipogenesis. FASEB J 3:2179-2183;1989. Blake, B.H. The annual cycle and fat storage in two popula- tions of golden-mantled ground squirrels. J. Mamma. 53: 157-167;1972. Boswell, T.; Woods, SC.; Kenagy, G.J. Seasonal changes in body mass, insulin, and glucocorticoids of free-living golden-mantled ground squirrels. Gen. Comp. Endocrinol. 96:339-346;1994. Bray, G.A. Obesity, a disorder of nutrient partitioning: the MONA LISA hypothesis. J. Nutr. 121:1146-1162;1991. Bronson, M.T. Altitudinal variation in emergence time of golden-mantled ground squirrels (Spermophilus lateralis). J. Mamma. 61:124-126;1980. Dark, J.; Stem, J.S.; Zucker, I. Adipose tissue dynamics dur- ing cyclic weight loss and weight gain of ground squirrels. Am. J. Physiol. 256:R1286-1292;1989. Davis, D.E. Pitfalls in in the use of ground squirrels for re- search on hibernation. Can. J. Zool. 62:1656-1658;1984. Green, C. J.; Brosnan, J .T.; Fuller, B.J.; Lowry, M.; Stubbs. M.; Ross, B.D. Effect of hibernation on liver and kidney metabolism in 13-lined ground squirrels. Comp. Biochem. Physiol. 79B:167-17131984. Green, P.K.; Wilkinson, C.W.; Woods, S.C. lntraventricu- lar corticosterone increases the rate of body weight gain in underweight adrenalectomized rats. Endocrinol. 130:269- 275;1992. Hansen, F.M.; Nilsson, P.; Hustvedt, B.E.; Nilsson-Ehle, P.; Lovo, A. Significance of hyperinsulinemia in ventromedial hypothalamus-lesion rats. Am. J. Physiol. 244:E203-E208; 1983. Hochachka, P.W.; Somero, G.N. Chapter 7, off-switches in metabolism: from anhydrobiosis to hibernation. In: Ho- chachka, P.W.; Somero, G.N., eds. Biochemical adaptation. Princeton, NJ: Princeton University Press; 1984:204-249. Jameson, E.W., Jr., and Mead, R.A. Seasonal changes in body fat, water and basic weight in Citellus lateralis, Eucamius speciosus, and E. amoenus. J. Mamma. 45:359-365;1964. Jameson, E.W., Jr. Food consumption of hibernating and nonhibernating CiteUus lateralis. J. Mamma. 45:634-640; 1965. Karmann, H.; Mrosovsky, N.; Heitz, A.; Le Maho, Y. Pro- tein sparing on very low calorie diets: ground squirrels suc- ceed where obese people fail. Itnl. J. Obesity 18(5):351-353; 1994. Kayser, C. Nutrition. In: Kayser, C., ed. The physiology of natural hibernation. London: Pergamon Press; 1961: 133- 165. Kenagy, G.J.; Sharbaugh, S.M.; Nagy, K.A. Annual cycle of energy and time expenditure in a golden-mantled ground squirrel population. Oecologia, 78:269-282;1989.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
McKeever, S. The biology of the golden-mantled ground squirrel. Ecologic Monogr. 34:383-401;1964. Michener, G.R.; Locklear, L.L. Over-winter weight loss by Richardson’s ground squirrels in relation to sexual differences in mating effort. J. Mamma. 71:489-499;1990-a. Michener, G.R.; Locklear, L.L. Differential costs of repro- ductive effort for male and female Richardson’s ground squir- rels. Ecology 71:855-868;1990-b. Mrosovsky, N. The medial hypothalamus and prehibemation obesity: a theory on behavioral tests. In: Morosovsky, N., ed. Hibernation and the hypothalamus. Appleton-Century Crofts Education Division, Meredith Corporation, New York; 1971: 25-102. Muchlinski, A.E.; Mariano, R.H. Three digestive tract en- docrine peptides and the circannual cycle ot body weight in Spermophilus lateralis. Comp. Biochem. Physiol. 88A:183- 186;1987. Pengeiley, E.T.; Asmundson, S.M. Free-running periods of endogenous circannian rhythms in the golden-mantled ground squirrel, Citellus lateralis. Comp. Biochem. Physiol. 30:177-183;1969. Phillips, J.A. Growth and its relationship to the initial an- nual cycle of the golden-mantled ground squirrel, Spermophi- lus lateralis. Can. J. Zool. 59:865-871;1981. Saito, M.; Bray, G.A. Adrenalectomy and food restriction in the genetically obese (ob/ob) mouse. Am. J. Physiol. 246: R20-R25;1984. Schwartz, M.W.; Figlewicz, D.P.; Baskin, D.G.; Woods, S.C.; Porte, D., Jr. Insulin in the brain: A hormonal regula- tion of energy balance. Endocrine Rev. 13(3):387-414;1992. Shivatcheva, T.M.; Ankov, V.K.; Hadjioloff, A.I. Circan- nual fluctuations of the serum cortisol in the European ground squirrel, Citdlus citellus L. Comp. Biochem. Physiol. 90A: 515-518:1988. Spence, J.T.; Cotton, J.T.; Underwood, B.J.; Duncan, C.P. Two-way analysis of variance. In: Spence, J.T.; Cotton, J.T.; Underwood, B.J.; Duncan, C.P., eds. Elementary statistics. Englewood Cliffs, New Jersey: Prentice Hall; 1992: 271-294. Stauffacher, W.; Orci, L. ,; Cameron, D.; Burr, L.; Renold, E. Spontaneous hyperglycemia and/or obesity in laboratory rodents: an example of the possible usefulness of animal dis- ease models with both genetic and environmental compo- nents. Recent Prog. Harm. Res. 27:41-95;1971. Torke, K.G.; Twente, J.W. Behavior of Spermophilus lateralis between periods of hibernation. J. Mamma. 58:385-390; 1977. Twente, J.W.; Twente, J.A. Seasonal variation in the hiber- nating behavior of Citellus lateralis. In: Fisher, K.C.; Dawn, A.R.; Lyman, C.P.; Schonbum, E.; South, F.E., Jr., eds. Mammalian hibernation III. New York: Elsevier; 1967: 47-63. Wade, 0. The behavior of certain Spermophiks with special reference to aestivation and hibernation. J. Mamma. 11: 160-188;1930. Wang, L.C.H. Energetic and field aspects of mammalian tor- por: the Richardson’s ground squirrel. In: Wang, L.C.H.; Hudson, J. W., eds. Strategies in cold-natural torpidity and thermogenesis. New York: Academic Press; 1978: 109-145. Wickler, S.J.; Hoyt, D.F.; van Breukelen, F. Disuse atrophy in the hibernating golden-mantled ground squirrel, Spermo- philus lateralis. Am. J. Physiol. 261:R1214-1217;1991.

Triglycerides and Body Weight in Squirrels
39. Wilson, B.E.; Deeb, S.; Florant, G.L. Seasonal changes in hormone-sensitive and lipoprotein lipase mRNA concentra- tions in marmot white adipose tissue. Am. J. Physiol. 262: R177-R181;1992.
40. Woods, S.F.; Figlewicz Latteman, D.P.; Schwartz, M. W.; Porte, D., Jr. A reassessment of the regulation of adiposity and appetite by the brain insulin system. Iml. J. Obesity, 14(suppl 3):69-76;1990.
265
41. Yukimura, Y.; Bray, G.A. Effects of adrenalectomy on body weight and the size and number of fat cells in the Zucker rat. Endocr. Res. Commun. 5: 189-198;1978.
42. Yeh, I.; Tam, CF.; Catuira, E.; Le, T.T.; Papa, V.; Pena, L.; Vasquez, M. ; Vu, C.; Wang, S. ; Lopez, G.A. Changes in various plasma lipid components, glucose, and insulin in SpermoI&Ius later& during hibernation. Comp. Biochem. Physiol. 1 llB:651-663;1995.




















