
Multiple biomarkers of pollution effects in caged mussels on the Greek coastline
Upload
khangminh22Category
view
0download
0
Journal of Physiology (1991), 432, pp. 639-680 639With 18 figures
Printed in Great Britain
CROSS-BRIDGE KINETICS IN THE PRESENCE OF MgADPINVESTIGATED BY PHOTOLYSIS OF CAGED ATP IN RABBIT PSOAS
MUSCLE FIBRES
BY J. A. DANTZIG*, M. G. HIBBERDt, D. R. TRENTHAMtAND Y. E. GOLDMAN*
From the *Department of Physiology, University of Pennsylvania, Philadelphia,PA 19104, USA, the tNational Institute for Medical Research, Mill Hill, London
and the tMassachusetts General Hospital, Boston, MA 02146, USA
(Received 20 December 1989)
SUMMARY
1. The interaction between MgADP and rigor cross-bridges in glycerol-extractedsingle fibres from rabbit psoas muscle has been investigated using laser pulsephotolysis of caged ATP (P3-1(2-nitrophenyl)ethyladenosine 5'-triphosphate) in thepresence of MgADP and following small length changes applied to the rigor fibre.
2. Addition of 465 gM-MgADP to a rigor fibre caused rigor tension to decrease by15-3 + 0 7% (s.E.M., n = 24 trials in thirteen fibres). The half-saturation value for thistension reduction was 18+4 /SM (n = 23, thirteen fibres).
3. Relaxation from rigor by photolysis of caged ATP in the absence of Ca2` wasmarkedly slowed by inclusion of 20 /,M-2 mM-MgADP in the photolysis medium.
4. Four phases of tension relaxation occurred with MgADP in the medium: at, aquick partial relaxation (in pre-stretch fibres); bt, a slowing of relaxation or a rise intension for 50-100 ms; ct, a sudden acceleration of relaxation; and dt, a final, nearlyexponential relaxation.
5. Experiments at varied MgATP and MgADP concentrations suggested thatphase at is due to MgATP binding to nucleotide-free cross-bridges.
6. Phase bt was abbreviated by including 1-20 mM-orthophosphate (Pi) in thephotolysis medium, or by applying quick stretches before photolysis or during phasebt. These results suggest that phases bt and ct are complex processes involving ADPdissociation, cross-bridge reattachment and co-operative detachment involvingfilament sliding and the Ca2+-regulatory system.
7. Stretching relaxed muscle fibres to 3'2-3-4 ,um striation spacing followed byATP removal and release of the rigor fibre until tension fell below the relaxed levelallowed investigation of the strain dependence ofrelaxation in the regions ofnegativecross-bridge strain. In the presence of 50 /,M-2 mm-MgADP and either 10 mM-Pi or20 mM-2,3-butanedione monoxime, relaxation following photolysis of caged ATPwas 6- to 8-fold faster for negatively strained cross-bridges than for positivelystrained ones. This marked strain dependence of cross-bridge detachment ispredicted from the model of A. F. Huxley (1957).
8. In the presence of Ca2+, activation of contraction following photolysis of cagedMS 8160

J. A. DANTZIG AND OTHERS
ATP was slowed by inclusion of 20-500 /,M-MgADP in the medium. An initialdecrease in tension related to cross-bridge detachment by MgATP was markedlysuppressed in the presence of MgADP.
9. Ten millimolar Pi partly suppressed active tension generation in the presence ofMgADP. The tension transients obtained when MgADP, Ca2+ and Pi were includedin the photolysis medium showed an unexpected initial tension rise followingphotolysis of caged ATP and then a decrease to the steady tension level.
10. Computer simulations of the cross-bridge reactions involving ADP release,MgATP binding, detachment, and reattachment into force-generating intermediateswere fitted to the transients recorded following photolysis of caged ATP. In theabsence of ADP the time course of the transients could be simulated using a simplemodel without strain-dependent rate constants and assuming that attached cross-bridge states in rapid equilibrium with detached states ({AM.ATP} and { M ADP Piexerted zero force. However, in the presence of MgADP the transients simulatedwith these assumptions showed a deeper tension dip following photolysis of cagedATP than the experimental records.
11. Two explanations of this discrepancy are considered. In the first hypothesis,rigor cross-bridges are assumed to be distributed over a wide range of forces,including negative forces, and ADP dissociates more rapidly from negativelystrained cross-bridges than from positively strained ones. In the presence ofMgADP,rapid detachment of the negatively strained cross-bridges limits the magnitude oftension dip following ATP release.
12. In the second explanation, in the presence of MgADP, attached cross-bridgestates that are in rapid equilibrium with detached states ({.A} and P}) areconsidered to bear significant force. Force in these states limits the magnitude of the
IAI.ATP} AM.XDP Pinitial tension dip following release of ATP. Since {MAT} and { M.ADP.Pj,} cannotbind MgADP, the assignment of tension to these states with MgADP present impliesthat in this model, interactions between cross-bridges or between individual heads ofa myosin molecule occur in the presence ofMgADP. Kinetic simulations on the basisof both models approximated the experimental records in the absence and presenceof MgADP. However, the first model better accommodated data obtained in thepresence of Ca2 , MgADP and Pi.
INTRODUCTION
The cyclic interaction of myosin in the thick filaments of the sarcomere and actinin the thin filaments is widely accepted as the basis for striated muscle contraction(Huxley, 1969; Huxley, 1974; Taylor, 1979; Cooke, 1986). Attachment of myosinheads to actin, a structural change leading to generation of force or filament slidingand subsequent detachment of the heads, is thought to take place many timesasynchronously during a contraction. The kinetics of several of the elementarybiochemical steps of the actomyosin (AM) ATPase (Scheme I) associated with thischemical cycle have been determined, but the specific step controlling the rate of
640

MgADP AND CROSS-BRIDGE KINETICS
overall cross-bridge cycling and ATPase activity has yet to be experimentallydemonstrated (Hibberd & Trentham, 1986; Goldman, 1987b).
ATP Pi ADP
AM - AM.ATP 'AM. ADP. P AM.ADP AM. ADP AM
M.ATP >='.M.ADP.Pi (Scheme I)
Many of the cross-bridge reaction steps that have been identified in isometricconditions are too fast relative to overall ATPase activity to be rate-limiting:MgATP binding to rigor cross-bridges (AM, Scheme I), detachment of myosin-ATP(M.ATP) from actin, hydrolysis of protein-bound ATP to form protein-boundADP.Pi, cross-bridge attachment and the force-generating event itself (Goldman,Hibberd & Trentham, 1984a, b; Ferenezi, 1986). There is evidence that Pi releasefrom the AM.ADP.Pi state to form AM'.ADP is linked to the structural changeleading to the force-generating intermediate (Cooke & Pate, 1985; Hibberd, Dantzig,Trentham & Goldman, 1985; Webb, Hibberd, Goldman & Trentham, 1986). Theslow step that controls the cross-bridge cycling rate is likely to be one ofthe remainingsteps in Scheme I, an isomerization of AM'.ADP or the release of ADP fromAM.ADP.The rate of the reaction that limits cross-bridge cycling in activated isometric
muscle is likely to be increased when the filaments slide and the mechanical strain inthe cross-bridges is reduced, because total energy liberation (related to ATPase rate)increases with shortening velocity (the Fenn effect: Fenn, 1923; Hill, 1964). In his1957 model, A. F. Huxley was able to provide an explanation of the Fenn effect onthe hypothesis that the cross-bridge detachment step is relatively slow in activeisometric muscle, but detachment accelerates markedly when the filaments slide andcross-bridges move into the region of low or negative mechanical strain.
In the present work, we investigated the mechanics of the cross-bridge reactionsassociated with MgADP binding and dissociation. We initiated relaxation andcontraction of glycerol-extracted fibres of rabbit psoas muscle from the rigor state byphotolysis of caged ATP (Kaplan, Forbush & Hoffman, 1978) in the absence andpresence of steady MgADP concentrations. We expected that initiation of relaxationor contraction would be slowed if ADP bound at the ATPase active site would haveto dissociate before MgATP could bind and detach the cross-bridges. By altering themechanical strain in the rigor cross-bridges before the laser pulse, we tested the straindependence of the kinetics in the presence of MgADP. Some of the results have beenbriefly reported (Dantzig, Hibberd, Goldman & Trentham, 1984; Dantzig, Trentham& Goldman, 1986; Dantzig & Goldman, 1989).
METHODS
Glycerol-extracted single fibres from rabbit psoas muscle were prepared as described byGoldman et at. (1984a) and mounted in an apparatus capable of producing rapid changes in theATP concentration within a fibre by photolysis of a photolabile precursor, caged ATP (Kaplan
21 PHY 432
641

J. A. DANTZIG AND OTHERS
et al. 1978; Walker, Reid, McCray & Trentham, 1988). The ends of the fibre were attached byaluminium clips (Goldman & Simmons, 1984) to a strain-gauge tension transducer (Sensonor,Norway; Goldman & Simmons, 1984) and a piezoelectric stack (Physik Instrumente, Switzerland).The tension signal consisted of the force generated by the fibre superimposed on a sinusoidal
component due to a1 /sm, 500 Hz length oscillation imposed by the piezoelectric stack. The in-phase and 90 deg out-of-phase sinusoidal components were separated from the tension signal by alock-in amplifier. The component in phase with the length change provides qualitative informationon the stiffness or number of cross-bridges attached. The component out-of-phase with the lengthchange (quadrature stiffness) is related to the visco-elastic nature of the cross-bridges. Positivedeflection of this signal indicates the existence of active cross-bridges which exhibit quick stressrelaxation (Kawai & Brandt, 1980; Goldman et al. 1984a, b).
Purification of ADP
Commercially supplied ADP (Sigma Chemical Company, or Boehringer Mannheim Biochemicals)was purified (>99%) by eluting it from a DEAE-cellulose column with a 10-500 mm gradient oftriethylammonium bicarbonate buffer (TEAB); pH 7 4,4 'C. The ADP fractions were concentratedto dryness in vacuo and residual TEAB removed by repeated evaporation with methanol. Thetriethylammonium salt of ADP was dissolved in deionized water, titrated to pH 7-0, rapidly frozenwith liquid nitrogen and stored at -80 'C.
Solution exchangesThe constituents of the experimental solutions are listed in Table 1. All references to ADP in
the text signify MgADP except where methods for preparation of solutions are described. The usualmeans of exchanging solutions was to lower the trough assembly containing the experimentalsolutions, move it laterally to position a new trough below the fibre and then raise the trough toimmerse the fibre in the new solution. The fibre was suspended in air for less than 3 s. Experimentswere carried out at 19-21 'C except where indicated.
For experiments in which small changes in tension were measured on the addition of MgADP tofibres in rigor, we adopted a flow-through procedure that minimized trough-changing artifacts. Thesolution in the 60,ul trough was exchanged by flowing through 400,u1 of the new solution. A glasscannula with continuous suction was arranged above the strain gauge to collect the solutionoverflow and to maintain the meniscus in a constant position. Purified ADP was added withequimolar MgCl2 to a concentrated rigor stock solution (Table 1) together with an ATP-depletingsystem of040 mM-glucose, 10jig ml-' lyophilized yeast hexokinase (Calbiochem Corporation, USA)and 045 mm of the myokinase inhibitor diadenosine pentaphosphate (Ap5A, Sigma ChemicalCompany). A few preparations of hexokinase irreversibly diminished rigor tension presumably dueto contaminating proteases. Results from these experiments were excluded.
Experimental protocol for photolysis trials
Fibres were relaxed in a 0 1 mM-MgATP relaxing solution with low calcium (Table 1) and thentransferred to a rigor solution devoid of nucleotide and allowed to develop tension. The fibre wasequilibrated for 2 min in a trough containing a rigor solution with 10 mM-caged ATP and 0-2 mm-MgADP (Table 1). Up to 1 mM-ATP was liberated by a pulse of 347 nm radiation from a frequency-doubled ruby laser. To ensure complete diffusion of Ca2+ into the fibre for rigor-photo-lysis-activation trials it was immersed in an extra wash of rigor solution with 30 jiM-free Ca2+ thentransferred to the same solution with added 10 mM-caged ATP and 0-500 jiM-MgADP. Mechanicaltransients were digitized and stored on computer diskettes for further analysis. The concentrationof ATP liberated from caged ATP was determined by anion exchange (SAX) high-performanceliquid chromatography with a running buffer of 04 M-(NH4)H2P04 adjusted to pH 40 and mixedwith methanol (92/8 v/v).Except where indicated, the caged ATP used for the experiments was a mixture (approximately
50:50) of the two diastereoisomers, one of which binds to cross-bridges and slows relaxation(Dantzig, Goldman, Luttmann, Trentham & Woodward, 1989). This slowing of cross-bridgekinetics by caged ATP binding would not markedly affect the present experiments because the rateprocesses observed on addition of MgADP to the photolysis medium were slower than tension
642

MgADP AND CROSS-BRIDGE KINETICS
relaxation in the presence of the higher affinity isomer. Preliminary experiments with purifiedisomers confirmed that MgADP slows relaxation in experiments both with the racemic caged ATPmixture and with the lower affinity isomer.
Sarcomere length experimentsIn some experiments the sarcomere length of a central region of the fibre was measured at high
time resolution by a white light diffraction method that avoids the Bragg-angle artifact in laser
TABLE 1. Composition of solutions used during caged ATP photolysis trialsSolution MgADP MgCl2 Mg2+ EGTA HDTA TES GSH PiRigor 0 3-2 1 53 0 100 10 0Rigor with Pi 0 2-7 1 20 19 100 10 25Rigor with MgADP 2 4-1 1 20 26 100 10 0Rigor with Pi, MgADP 2 4-7 1 20 12 100 10 25Ca2+ rigor 0 1-3 1 20 33 100 10 0Ca2+ rigor with Pi 0 1.5 1 20 28 100 10 10Ca2+ rigor with MgADP 2 3-2 1 20 26 100 10 0Ca2+ rigor with Pi, MgADP 2 3-5 1 20 22 100 10 100-1 mM-MgATP relax 0 2-7 1 30 0 100 10 0
All concentrations are in mm. The ionic strength of solutions with no caged ATP was 0-2 M. Forphotolysis solutions, caged ATP, concentrated rigor stock solutions and H20 were combined toobtain the above concentrations plus 10 mM-caged ATP. No adjustment was made for the ionicstrength contribution of caged ATP. Solutions were made up at room temperature (18-23 °C) andbrought to pH 7-1 with KOH or HCI. All solutions contained - 140 mM-Na+ + K+. Intermediateconcentrations of ADP and Pi were obtained by mixing the appropriate solutions above. Insolutions with no added Ca2+, free Ca2+ < 10-8 M. In solutions with added Ca2+, free Ca2+ t 30 ,UM.Relaxing solution contained 22 mM-creatine phosphate and 1 mg ml-' creatine phosphokinase(100-150 units activity: 1 unit transfers 1.0 /smol phosphate from creatinine phosphate to ADP perminute at pH 7-4 and 30 °C).EGTA, ethyleneglycol-bis-(,8-aminoethylether)N,N,NN'-tetraacetic acid; HDTA, 1,6-diami-
nohexane-N,N,N'N'-tetraacetic acid; TES, N-tris(hydroxymethyl)methyl-2-aminoethanesulphonicacid; GSH, glutathione (reduced form).
diffraction experiments (Goldman, 1987 a). For these experiments, a pulsed xenon lamp(Chadwick-Helmuth) was used to photolyse the caged ATP. Quartz collecting optics and filters (U-340, Hoya, Japan and WG-305, Schott), transmitted 300-380 nm light to the fibre trough.
RESULTS
Rigor tension in the presence ofMgADPVarious effects on tension have been observed when MgADP is added to the
bathing medium of a fibre in rigor (Marston, Tregear, Rodger & Clark, 1979; Kawai,1986; Schoenberg & Eisenberg, 1987). We sought to determine the influence ofMgADP on rigor tension under conditions minimizing both low-level ATPcontamination and mechanical artifacts associated with the solution changes.Column-purified ADP, an ATP-depleting system of glucose, hexokinase and themyokinase inhibitor (Ap5A), together with a flow-through arrangement for solutionexchanges and effective wash-out of ATP from the troughs, were all required toobtain valid results.
Addition of 1 mM-MgCl2 and 1 mM-purified ADP (465 /,M-MgADP) to a rigor fibre21-2
643

J. A. DANTZIG AND OTHERS
in the presence of the ATP-depleting system caused rigor tension to decrease (Fig. 1,first ADP arrow) by 15-3+ 0-7% (s.E.M., n = 24 trials in thirteen fibres). If ATP waswashed out of the fibre effectively and the ATP-depleting system was adequate, littletension recovery was observed following a small length decrease (AL). Tension
xco
I- I
zcra: 2 < = a w L ''c
10 minFig. 1. Protocol for measuring the tension of cross-bridges in rigor in the presence of addedMgAMP and MgADP in the absence of 0a2+. A permeabilized fibre of rabbit psoas musclewas put into rigor (R) from 04 mM-MgATP relaxing solution (Table 1) and then washedtwice more (at R) with rigor solution to remove residual ATP. A flow-through solutionchange procedure as described in the Methods section was used to minimize mechanicalartifacts during solution changes. At the arrow labelled 'AMP' the solution was changedto rigor with 1 mM-MgCl2 and 1 mM-AMP. AL indicates a 082% release and subsequentrestretch to check for possible tension recovery due to cross-bridge cycling. AMP iswashed out at H. During the same rigor contraction, hexokinase (04a5 mg/ml), glucose(0w5 mM) and Ap5A (3R5 /M, final concentrations) were added at S. 1 mM-MgCl2 and 1mw -ADP were added at the first ADP arrow to give 465rgM-MgADP. After a length releaseand restretch (AL) and several washes inh, 465hsM-MgADP was introduced without theATP-depleting system (second ADP arrow). The fibre was reimmersed in rigor solutionand then relaxed. Sarcomere length =2m39 ,m; fibre dimensions = 229 mm x 5840,usm2.
recovered almost to the original rigor tension when ADP was removed (two washesin rigor solution, R) if allowance was made for the slow decline of rigor tension.
Introduction of the same concentration of MgADP without the ATP-depletingsystem caused an initial tension decrease (Fig. 1, second ADP atrow), but then overthe next 5 mi tension increased. The net effect of adding MgADP without the ATP-depleting system was an increase of tension. Also, when a length release was applied(AL), tension did not remain at the lower value indicating that enough ATP waspresent within the fibre to allow development of tension by cross-bridge cycling. ThisATP was probably produced within the fibre by endogenous myokinase, becausewith column-purified ADP, the tension rise could be prevented by adding Ap5Awithout glucose and hexokinase. Since adding low concentrations of MgATP to arigor fibre causes tension to rise (Goldman et al. 1984a, aand freences therein), thetension decrease on adding MgADP cannot be caused by ATP contaminating thesolutions or generated within the fibre.
Addition of 1 mM-MgCl2 and 1 mM-AMP to the fibre in rigor (Fig. 1, AMP arrow)did not cause a change in tension and force did not recover following a length release,suggesting that the tension reduction by MgADP reflects a specific interaction withthe nucleotide binding site of myosin.
644
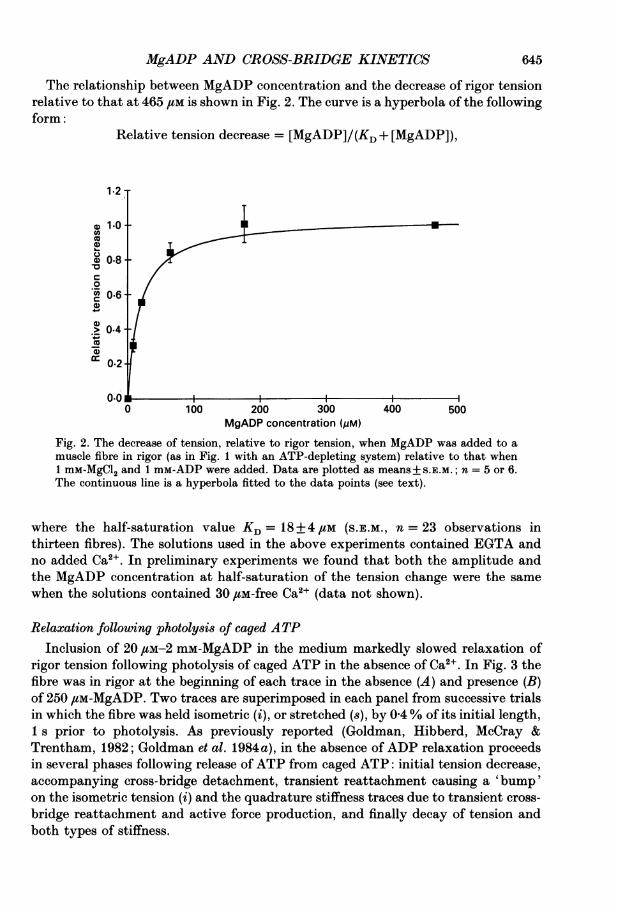
MgADP AND CROSS-BRIDGE KINETICS
The relationship between MgADP concentration and the decrease of rigor tensionrelative to that at 465 JM is shown in Fig. 2. The curve is a hyperbola of the followingform:
Relative tension decrease = [MgADP]/(KD + [MgADP]),
1.2
, 1.0
0.8
0C 0.6
>04-0.
0.2
0.00 100 200 300 400 500
MgADP concentration (#M)
Fig. 2. The decrease of tension, relative to rigor tension, when MgADP was added to amuscle fibre in rigor (as in Fig. 1 with an ATP-depleting system) relative to that when1 mM-MgCl2 and 1 mM-ADP were added. Data are plotted as means+S.E.M.; n = 5 or 6.The continuous line is a hyperbola fitted to the data points (see text).
where the half-saturation value KD= 18+ 4 /iM (S.E.M., n = 23 observations inthirteen fibres). The solutions used in the above experiments contained EGTA andno added Ca2 . In preliminary experiments we found that both the amplitude andthe MgADP concentration at half-saturation of the tension change were the samewhen the solutions contained 30 /tM-free Ca2+ (data not shown).
Relaxation following photolysis of caged ATPInclusion of 20 tM-2 mM-MgADP in the medium markedly slowed relaxation of
rigor tension following photolysis of caged ATP in the absence of Ca2 . In Fig. 3 thefibre was in rigor at the beginning of each trace in the absence (A) and presence (B)of 250 ,IM-MgADP. Two traces are superimposed in each panel from successive trialsin which the fibre was held isometric (i), or stretched (s), by 0 4% of its initial length,1 s prior to photolysis. As previously reported (Goldman, Hibberd, McCray &Trentham, 1982; Goldman et al. 1984a), in the absence of ADP relaxation proceedsin several phases following release of ATP from caged ATP: initial tension decrease,accompanying cross-bridge detachment, transient reattachment causing a 'bump'on the isometric tension (i) and the quadrature stiffness traces due to transient cross-bridge reattachment and active force production, and finally decay of tension andboth types of stiffness.
645

646 J. A. DANTZIG AND OTHERS
In the presence of 250 /tM-MgADP (Fig. 3B), tension rose or maintained a plateaufor - 75 ms and then suddenly relaxed with a time course similar to the relaxationin panel A. The positive quadrature signal was also larger and prolonged suggestinga maintenance of actively tension-generating cross-bridges during the plateau phase.
A 0 ADP B 250 iM-MgADP
0.2r Quadrature stiffness
In-phase stiffnessLs
C4 sWEz Tension
° s-i s-i.
100 ms 100 ms
Fig. 3. Effects of MgADP on tension and stiffness transients induced by the liberation ofATP in the absence of Ca2+. At the arrows, caged ATP was photolysed to liberate 700 uM-ATP. In A, no ADP was in the rigor solution (Table 1) and in B, 250 ftM-MgADP waspresent. Two traces are superimposed in each panel from successive trials in which thefibre was held isometric (i) and stretched (s), by 0-4% of its initial length 1 s prior tophotolysis. Simultaneously recorded quadrature and in-phase stiffness records are shownat the top as indicated. The upper in-phase and lower quadrature traces were recorded inthe pre-stretch trials. Baseline records (flat traces) were obtained in the photolysis troughabout 1 min after the relaxation had been completed. Some records show transientartifacts induced by the laser pulse, apparent as an instantaneous decrease in tension.These tension artifacts cause transient positive or negative deflections of some stiffnessand quadrature signals for 10-20 ms. The lowest traces are algebraic differences (s- i)between the pre-stretched and isometric records. Temperature = 20-2 'C.
In-phase stiffness decreased more slowly in the presence of MgADP than in itsabsence. During the time course of the tension plateau in the presence of MgADP,stiffness fell indicating that tension per attached cross-bridge was increasing. Tensionper unit stiffness (relative to rigor stiffness before the laser pulse) was 36+ 5%(S.E.M., n = 10) greater than that in the absence of ADP at 10-20 ms where the0 ADP trace underwent an initial minimum. At the same time point, stiffness(relative to rigor stiffness before the laser pulse) in MgADP was 115 + 3% (n = 10)of that at 0 ADP. In-phase stiffness recordings containing an artifactual deflectiondue to an instantaneous tension drop at the laser pulse (similar to the initialdeflection in the upper quadrature stiffness trace of Fig. 3B) were not used forquantitative analysis. Although tension was maintained or increased during theplateau, the falling stiffness indicates that net cross-bridge detachment was occurringthroughout the relaxation.To investigate the kinetics of the slowing of relaxation by MgADP, a series of
experiments was conducted (Figs 4, 5 and 6) in which tension transients were

MgADP AND CROSS-BRIDGE KINETICS
examined as a function of MgADP and released ATP concentrations in the absenceof Ca2". In both the absence and presence of MgADP, tension traces starting fromdiffering levels of positive rigor tension converged well before they reached thebaseline, so that the final relaxation proceeded along a common time course. Tihis
A1 mM-ATP
bt
E i
LA
LOrs-
adi bdCd
100 ms
0.4 mm-ATP4
S
s-i
100 Ms
B 0.2 mM-ATP4I
i
s-i
100 Ms
D0.9 mM-ATP
4S
i
s-i
100 msFig. 4. Tension transients following photo-release of various concentrations of ATP in afibre in the presence of 200 /tM-MgADP and absence of Ca2l (Table 1). 10 mM-caged ATPwas present throughout and various concentrations of ATP were released as indicated ineach panel by varying the intensity of the laser pulse. Two traces are superimposed ineach panel together with a baseline recording and difference trace as in Fig. 3. After threesets of traces (A, B and C) at successively lower [ATP], 900 ,uM-ATP was released in D tocheck that the fibre had not deteriorated during the experiment (compare with traces inA). Phases at, bt, ct, dt, ad bd and c are described in the text. Sarcomere length = 2 20 ,tm;fibre dimensions = 2-42 mm x 5044 ,sm2; temperature = 21-5 'C.
behaviour is illustrated by pairs of tension traces at various MgATP and MgADPconcentrations in Figs 3, 4, 5 and 7. The relaxation of tension could be characterizedby the four phases indicated in Fig. 4A: phase at, initial partial relaxation lasting5-20 ms; bt, a slowing of relaxation or a tension rise to a plateau for 50-100 ms; ct,a sudden acceleration of relaxation and convergence of pairs of traces starting atdifferent rigor tensions; and dt, final, nearly exponential relaxation when these pairsof traces were superimposed. The tension traces were concave downward duringphase bt, became concave upward at an inflexion point during phase ct and remainedconcave upward for the remainder of relaxation. The most prominent effect of addingMgADP to the photolysis medium was a 5- to 20-fold prolongation of phase btrelative to the duration of the corresponding delay or 'bump' in the absence of ADP.
647

648 J. A. DANTZIG AND OTHERS
The kinetics of phases ct and dt did not depend on MgADP concentration (Figs 3and 5). This was analysed by fitting the tension data (T) following the inflexion pointin phase ct with the sum of two exponential components of the form:
T = A exp (k1 t) +B exp (k2 t) + C.
A s 0 ADP B s 50 pM-MgADP
ZC4
S-i
LLJ
s-i s-i
100 ms 100 ms
C s 1001 M-MgADP D s 500 iM-MgADP
I m~~~~~~~~~I
s-i s-lV.
lOOms lOOms
Fig. 5. Tension transients following photolysis of caged ATP at various MgADPconcentrations. 650 4uM-ATP was released from 10 mM-caged ATP in a fibre using rigorsolutions in the absence of Ca2+ (Table 1) with MgADP concentrations indicated in eachpanel. Other details as in Fig. 4. Sarcomere length = 229,4um; fibre dimensions= 2-68 mm x 5580 4am2; temperature = 20*2 °C.
The traces were well fitted by this equation and the apparent rate constants (1c1 andk2) for final relaxation were 85-5+ 16s8 and 15-5±0s9 S-1 (S.E.M., n = 16) in thepresence of 100 /tM-MgADP and 95@2 + 6-2 and 15*2 ± 1e2 s-i (n = 6) in the absence ofADP. T.he slower of the two processes accounted for 59±9% of the relaxationfollowing the ct inflexion point at 100 ftM-MgADP and 68+2 % at 0 ADP.
In previous reports on relaxation of rigor muscle using caged ATP, we plotted thealgebraic difference between pairs of tension traces that started at different levels ofrigor force to quantify the half-time for convergence. This procedure was based onthe assumption that the convergence time indicates the initial detachment of rigorcross-bridges by MgATP (Goldman et al. 1984a). In the absence of ADP, the tensiondifference traces (8-i, Figs 3A and 5A) decayed promptly to the baseline with half-times of 6-10 ins. As reported earlier (Goldman et al. 1984a) this rate depended onthe released MgATP concentration.When 20 /LM-1 mM-MgADP was added to the medium the difference traces
decayed in several phases (Fig. 4A): ad, an initial decay lasting 5-30mis; bd, aslowing or lag phase lasting 50-100 ins; an abrupt transition into Cd, a more rapid

MgADP AND CROSS-BRIDGE KINETICS
further decay; and in most cases, an undershoot before the final zero level wasreached. During phase bd, an inflexion point marks a change in the difference tracefrom concave upward to concave downward. Phases ad, bd and cd of the tensiondifference traces are approximately coincident with phases at, bt and ct of the originaltension recordings. In the difference traces there is no analogue of phase dt, becausethe tension traces converge before final relaxation leading to a zero difference traceduring dt.The initial rapid decay phase (ad) of the tension difference trace was dependent on
the concentration of ATP released from caged ATP. Two hundred micromolarMgADP was present throughout the experiment shown in Fig. 4, and theconcentration of ATP liberated from caged ATP was varied by adjusting theintensity of the laser pulse. The final ATP concentration is shown in each panel. Theamplitude of the rise in tension was greater at the lower ATP concentrations, but theduration of the plateau phase (bt) did not depend strongly on [MgATP]. For thetension difference traces (s- i in each panel), phase ad was faster at higher [ATP], butthe amplitude and duration of the slower components bd and cd were not verysensitive to [MgATP]. These results suggest that phases at and ad are due to rapiddetachment of rigor cross-bridges without bound ADP and that the slower phasesresult from cross-bridges with ADP bound initially.The slower phases (bd and onward) of decay of the difference traces were separated
from phase ad by fitting a straight line to the difference trace near the inflexion pointin phase bd (Fig. 6A, inset). The back-extrapolation of that line to the time of thelaser pulse was taken as the amplitude (As) of the slow decay and the half-time (ti)of the slow components was measured as the time for the tension difference trace todecay half-way from As to the baseline (Fig. 6A, inset). When no slow phase waspresent in the absence of ADP, tL was defined as the halftime of decay of the entiretension difference.The ti of the slow phases of the difference traces increased markedly when
[MgADP] was raised up to 50 ,tM and then more gradually between 50 ,UM and 1 mM(Fig. 5 and the plot of Fig. 6A, closed symbols). The line fitted to the closed symbols(at [MgADP] > 20 /M) corresponds to ti (ms) = 51-2 + 48-5 [MgADP] where [MgADP]is in mm.For the simplified Scheme II:
ADP ATPk-
AM.ADP - AM Detached (Scheme II)'k+D
it can be shown that when ADP dissociation is slow compared to nucleotide binding,the expected half-time of the slow component of cross-bridge detachment isapproximated by tL = ln(2)(1/klD+ k+D[MgADP]/(k1 D/+T[MgATP])) (Bernasconi,1976). Substituting the slope and intercept into this equation we obtaink-D= 13-5 s-1 and k+T[MgATP]/k+D = 1 1 mM. [MgATP] is approximately 0-8 mMon average in these experiments and k+T - 2-5 x 105 M-1 s-1 (Goldman et al. 1984a).The value of k+D is calculated to be 1-5-3-8 x 105 M-1 s-1, similar to the rate of ATP-induced detachment. The ratio (klD/k+D) of these estimates gives 36-90 /IM as theapparent dissociation constant (KD) for MgADP. These values are reasonable but
649

650 J. A. DANTZIG AND OTHERS
A
801 t-
E 60 -
-ZL 40
0 200 400 600 800 1000MgADP concentration (jM)
B
0.6 50m(D~ ~ ~ ~ ~ I
0
a 0.20 __________________\____ms
0 0 20 40 0200 10
MgADP concentration (jim)
Fig. 6. Half-times (A, *@) and amplitudes (B) of the slow phases (bd and onward) of tensiondifference traces (obtained from records such as those shown in Fig. 5) plotted againstMgADP concentration. The insets show the methods by which the half-times (t1) andamplitudes (A8) of the slow components of tension difference traces relative to the totalamplitude (At) were obtained in the absence of Ca2+. The trace in the insets is thedifference trace from Fig. 4A. In A, the open symbols represent half-times of the decayof tension difference traces obtained in the presence of 30 ,uM-Ca2+ (records as in Figs12-15). Error bars indicate S.E.M. with n values between 3 and 15, except at 0 ADP wherethe error bars are within the size of the symbols and n = 106 (-Ca2+) and n = 17 (+Ca2+).In A, straight lines are fitted to the data at [MgADP] > 50 jiM. In B, a hyperbola is fittedto the data at [MgADP] > 20 /M. n values for B are between 3 and 11 except at 0 ADPwhere n = 60.
they should not be taken as accurate estimates of the rate constants because theshapes of the recorded relaxation and difference traces are not exponential as wouldbe expected from Scheme II.
IfADP dissociation is fast relative to nucleotide binding, then the slow componentof detachment is approximated by ti = ln(2)(1 + k+D[MgADP]/k-D)/(k+T[MgATP]).Based on the line fitted to the closed symbols in Fig. 6A, the ratio kD/lk+Dcorresponding to KD would be 806 /LM, a value much larger than the 18-50 /IM figures
MgADP AND CROSS-BRIDGE KINETICS
A s 4
(N FI_.I
o "'. 0 AP, P
.b0 s
s-i
100 ms
B s4fiS
oPi
S-i
100 MsD s 4
iS I
(N F
E ,z V 1 mM-Pi
0
S-IL
-S-i
I100 Ms
10 mM-pi
F150
100
25 mM-Pi lo
0
100 ms
s-i A
100 ms
I9
4
5 1 113
13
0 5 10 15 20 25Steady phosphate concentration (mM)
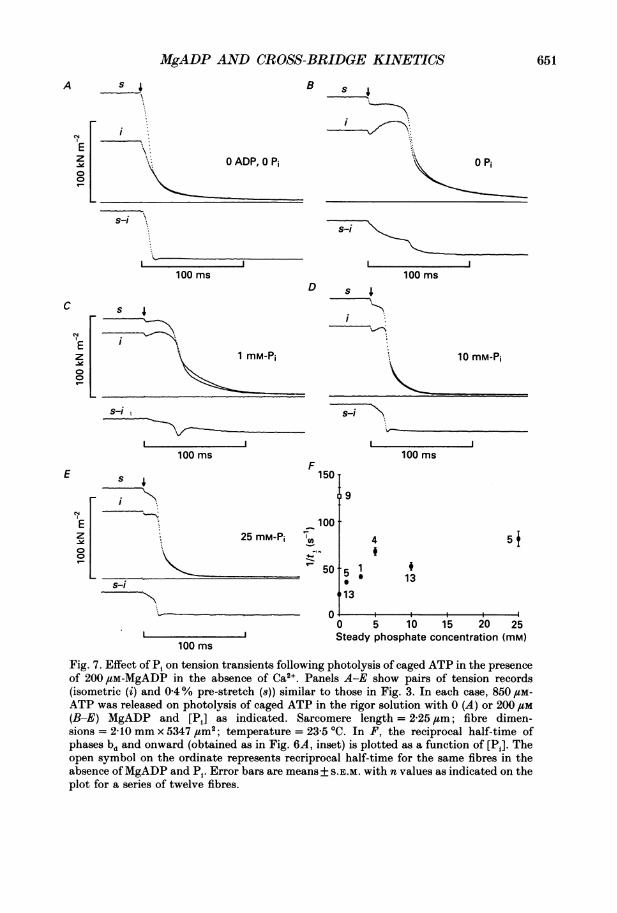
Fig. 7. Effect of Pi on tension transients following photolysis of caged ATP in the presenceof 200 /SM-MgADP in the absence of Ca2 . Panels A-E show pairs of tension records(isometric (i) and 0-4% pre-stretch (8)) similar to those in Fig. 3. In each case, 850 1cM-
ATP was released on photolysis of caged ATP in the rigor solution with 0 (A) or 200 /M(B-E) MgADP and [Pi] as indicated. Sarcomere length = 2-25 /m; fibre dimen-sions = 210 mm x 5347 um2; temperature = 23-5 'C. In F, the reciprocal half-time ofphases bd and onward (obtained as in Fig. 6A, inset) is plotted as a function of [Pi]. Theopen symbol on the ordinate represents recriprocal half-time for the same fibres in theabsence ofMgADP and Pi. Error bars are means+ S.E.M. with n values as indicated on theplot for a series of twelve fibres.
C
E
(N F
Elz01
s 4fs
S-i
651
I
lw

J. A. DANTZIG AND OTHERS
estimated from Figs 2 and 6B (see below). Therefore we interpret the slowing ofrelaxation on the basis of slow ADP release. The open symbols in Fig. 6A refer toconvergence half-times in the presence of Ca2+ and will be discussed later.
In the tension difference traces, the amplitude (As) of the slow component (phasesbd and onward) was sensitive to the MgADP concentration. A greater proportion ofthe overall decay of the difference trace was represented in the slow component at thehigher MgADP concentrations relative to the initial decrease (phase ad). The ratio of
As to the total amplitude of tension difference (At, Fig. 6B, inset) is plotted vs.[MgADP] in Fig. 6B. In Fig. 6, both thetL and AS/At points plotted at zero MgADPare elevated from typical values due to a subgroup of fibres used in this series ofexperiments. The data with MgADP present were well fitted by a hyperbola with ahalf-saturation value of 50 /tM and intercept on the ordinate at AS/At = 0 5.The results of altering MgADP and MgATP concentrations were highly
reproducible and did not depend on the sequence of applying the various testconcentrations as long as care was taken to wash ADP out of the fibres extensivelywhengoing from high to low or zero [MgADP]. The recordings in Fig. 4 were obtainedin the order A-D, with the last panel taken at high [MgATP] as a control record.Some slowing of the response is evident in panel D compared to panel A, but the mainchanges observed at lower [ATP] are not due to fibre deterioration.The data shown in Figs 4-6 suggest that phase ad represents ATP-induced
detachment of rigor cross-bridges that did not have ADP bound at the time of thelaser pulse. As the MgADP concentration was increased, the proportion of the cross-bridges in the AM state decreased, so the relative amplitude of phase ad decreased.The remainder of the decay, then, would be a more complex process of cross-bridgedetachment, presumably involving ADP dissociation at 10-15 s-', MgATP binding,detachment and possibly further reactions. Cross-bridge reattachment seems tocause the delay or 'bump' during relaxation in the absence of ADP, so the prolongedtension plateau (phase bt) may also involve cross-bridge reattachment and activeforce production. Since stiffness decay is also slowed during phase bt, maintenance ofthe derepressed condition of the Ca2+-regulatory system by the attached cross-bridges probably contributes to slowing of relaxation. To test this hypothesis weinvestigated the events that terminate phase bt.
Factors determining the duration of the plateau phaseIn the experiments shown in Figs 2-6, an abrupt transition occurred from slow
relaxation (phases bt and bd) to a much more rapid relaxation (phases ct and Cd),suggesting that a threshold or co-operative event rapidly increased the rate of cross-bridge detachment 50-100 ms after the laser pulse. Three experiments wereperformed to characterize the chemical and mechanical factors associated withtermination of the plateau phase: addition of orthophosphate (Pi) to the mediumalong with MgADP, measurement of striation spacing during the transients andsmall length changes to test the strain dependence.Although the in-phase stiffness declines during phase bt, implying net cross-bridge
detachment, some cross-bridges may be reattaching. We previously reported(Hibberd et al. 1985) that in the absence of ADP and Ca2 , inclusion of 1-25 mM-P1suppresses tension development of reattaching cross-bridges. If Pi also suppresses
652

MgADP AND CROSS-BRIDGE KINETICS
tension due to reattaching cross-bridges in the presence of MgADP, it might hastenthe onset of phase ct.
Addition of 1-25 mM-Pi in the presence of 200,tM-MgADP considerably ab-breviated the tension plateau (phase bt, Fig. 7B-E) and reduced the half-time of the
ATP ~~~~~Sarcomere lengthAT.?P
E\o t ~~~~~~Tension
100 ms
Fig. 8. Change of striation spacing during relaxation of a fibre following photo-release of- 600 4uM-ATP in the presence of 200 /SM-MgADP. Conditions as for Fig. 3 except that thephotolysis UV light source was a pulsed xenon flash lamp as described in Methods.
slow component (bd and onward) of the tension difference traces (Fig. 7F) asexpected on the hypothesis of cross-bridge reattachment. For comparison, I/ti forthe tension difference traces of the same series of fibres in the absence of ADP andPi is plotted as the open symbol in Fig. 7F. MgADP slowed relaxation andconvergence at all Pi concentrations tested and an abbreviated plateau of tensionremained up to the maximum Pi concentration (25 mM). The marked effect of Pi onconvergence of the tension traces in the presence ofMgADP contrasts with the verysmall effect of Pi on convergence reported in the absence of ADP (Hibberd et al.1985). These results suggest that reattaching cross-bridges contribute to maintenanceof the tension plateau following photolysis of caged ATP in the presence of MgADP.But even when the formation of AM'.ADP states was suppressed by Pi, MgADPslowed convergence.
Final relaxation following the plateau (phases ct and dt) was markedly acceleratedby Pi in the presence of MgADP as was previously reported by Hibberd et al. (1985)in the absence ofADP. This result indicates that Pi binding to AM'.ADP and reversalof cross-bridge attachment contributes to phases ct and dt of relaxation.To check whether filament sliding occurs during the relaxation, the photolysis
technique was combined with measurements of striation spacing using a white-lightdiffraction apparatus (Goldman, 1987 a). During the initial relaxation and theplateau phase of tension (phases at and bt), striation spacing was remarkablyconstant (Fig. 8). In five trials, striation spacing was constant to within 5 nm half-sarcomere-1 during the tension plateau. Just before phase ct, striation spacingchanged suddenly. In the experiment of Fig. 8 striation spacing suddenly increased
653

654 J. A. DANTZIG AND OTHERS
just prior to phase ct and continued to increase until relaxation was complete. Insome experiments, striation spacing was constant until the tension plateau ended,but then decreased briefly during phase ct and finally increased to the relaxed level.The length changes indicate that substantial axial motions can occur during phase
ATPA S _ B
AL
ATP
rL
100 ms 100 ms
Fig. 9. Influence of quick length changes before (A) and after (B) photo-release of ATP.In A, 700 1uM-ATP was liberated in the presence of 50 /uM-MgADP. A 0-6 % pre-stretch (s)and pre-release (r) were applied 1 s prior to the laser pulse. Sarcomere length = 2-25 4um;fibre dimensions = 2-88 mm x 6025 4um2. In B, 600 /LM-ATP was liberated in the presenceof 100 /SM-MgADP. A 0 5% stretch and release were done after the laser pulse during thetension plateau. Sarcomere length = 2-15 1sm; fibre dimensions = 2-52 mm x 6738 /m2;temperature = 21-9 'C.
ct of relaxation. Since stiffness is falling faster than tension during phase bt, tensionper attachment is increasing. The axial motions suggest that some sarcomeresrapidly elongate ('give') when tension per attachment reaches a value above themomentary tension-bearing capacity of the cross-bridges. The resulting axialmotions might either initiate the rapid relaxation or they might be a consequence ofnon-uniform relaxation along the fibre.To investigate further the relationship between the rapid phase of relaxation and
strain in the cross-bridges, length perturbations were applied. Figure 9A showsrecordings from an experiment in which an isometric trial (i) was compared to tracesfollowing a 06% stretch (s) or release (r) applied 1 s before the laser pulse. Releasingthe muscle fibre before the laser pulse prolonged the tension plateau and stretchingthe fibre before the laser pulse abbreviated the plateau.Length changes were also applied during the plateau period (Fig. 9B). Releasing
the muscle fibre during the transient prolonged the plateau phase, whereas stretchingthe muscle fibre abbreviated the plateau and immediately initiated rapid relaxation.The above results using length changes suggest that there is a maximum tension perattached cross-bridge during the plateau and when this tension threshold isexceeded, rapid relaxation is initiated.
Negatively strained cross-bridgesWe recently reported a method of applying negative strain to rigor cross-bridges
(Goldman, McCray & Vallette, 1988) and have used that technique to test the strain
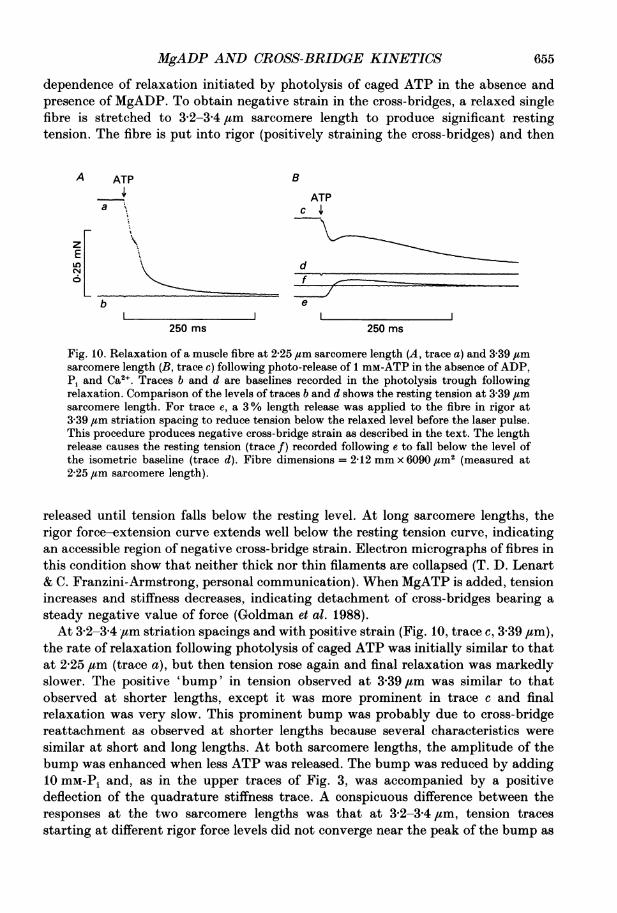
MgADP AND CROSS-BRIDGE KINETICS 655
dependence of relaxation initiated by photolysis of caged ATP in the absence andpresence of MgADP. To obtain negative strain in the cross-bridges, a relaxed singlefibre is stretched to 32-34 /am sarcomere length to produce significant restingtension. The fibre is put into rigor (positively straining the cross-bridges) and then
A ATP B
r__, ATPa \ c
Z:
CN6 L _ _ _ _ _ _ _ _ _ _ _
af _ -_
b e
250 ms 250 ms
Fig. 10. Relaxation of a muscle fibre at 2-25 um sarcomere length (A, trace a) and 3 39 /zmsarcomere length (B, trace c) following photo-release of 1 mM-ATP in the absence ofADP,Pi and Ca2". Traces b and d are baselines recorded in the photolysis trough followingrelaxation. Comparison of the levels of traces b and d shows the resting tension at 3-39 ,umsarcomere length. For trace e, a 3% length release was applied to the fibre in rigor at3 39 ,um striation spacing to reduce tension below the relaxed level before the laser pulse.This procedure produces negative cross-bridge strain as described in the text. The lengthrelease causes the resting tension (trace f) recorded following e to fall below the level ofthe isometric baseline (trace d). Fibre dimensions = 2-12 mm x 6090 ,um2 (measured at2 25 ,um sarcomere length).
released until tension falls below the resting level. At long sarcomere lengths, therigor force-extension curve extends well below the resting tension curve, indicatingan accessible region of negative cross-bridge strain. Electron micrographs of fibres inthis condition show that neither thick nor thin filaments are collapsed (T. D. Lenart& C. Franzini-Armstrong, personal communication). When MgATP is added, tensionincreases and stiffness decreases, indicating detachment of cross-bridges bearing asteady negative value of force (Goldman et al. 1988).At 3-2-3-4 ,um striation spacings and with positive strain (Fig. 10, trace c, 3 39 #um),
the rate of relaxation following photolysis of caged ATP was initially similar to thatat 2-25 ,tm (trace a), but then tension rose again and final relaxation was markedlyslower. The positive 'bump' in tension observed at 3-39 jum was similar to thatobserved at shorter lengths, except it was more prominent in trace c and finalrelaxation was very slow. This prominent bump was probably due to cross-bridgereattachment as observed at shorter lengths because several characteristics weresimilar at short and long lengths. At both sarcomere lengths, the amplitude of thebump was enhanced when less ATP was released. The bump was reduced by adding10 mM-Pi and, as in the upper traces of Fig. 3, was accompanied by a positivedeflection of the quadrature stiffness trace. A conspicuous difference between theresponses at the two sarcomere lengths was that at 3-2-34,um, tension tracesstarting at different rigor force levels did not converge near the peak of the bump as

J. A. DANTZIG AND OTHERS
occurred at the shorter lengths. Instead, tension traces following a pre-stretchmaintained a higher tension than in isometric trials until the traces met at therelaxed level.
For trace e in Fig. 10, the protocol described above to apply negative tension tothe cross-bridges was used, with the extra step of adding 10 mM-caged ATP to themedium 2 min before the mechanical release used to induce negative strain (see alsolegend to Fig. 10). Photolysis of the caged ATP (at the time of the arrow) resultedin rapid increase of tension to a value above the resting level. Tension then decayedto the resting level with a time course similar to that of the final relaxation in tracec. Thus, two processes contribute to the transient observed with negative cross-bridge tension: detachment of negatively strained cross-bridges and active forcegeneration of reattaching cross-bridges.
In order to isolate the component due to detachment of negatively strained cross-bridges, tension generation by reattaching cross-bridges was minimized by addingeither 25 mM-Pi (Hibberd et al. 1985) or 20 mM-2,3-butanedione monoxime (BDM;Horiuti, Higuchi, Umazume, Konishi, Okazaki & Kurihara, 1988; Dantzig &Goldman, 1989; Higuchi & Takemori, 1989; Lenart, Tanner & Goldman, 1989). Pi isthought to decrease force production per attached cross-bridge and BDM seems toreduce the rate constant for formation of force-generating cross-bridges (T. D.Lenart, J. W. Tanner & Y. E. Goldman, personal communication).
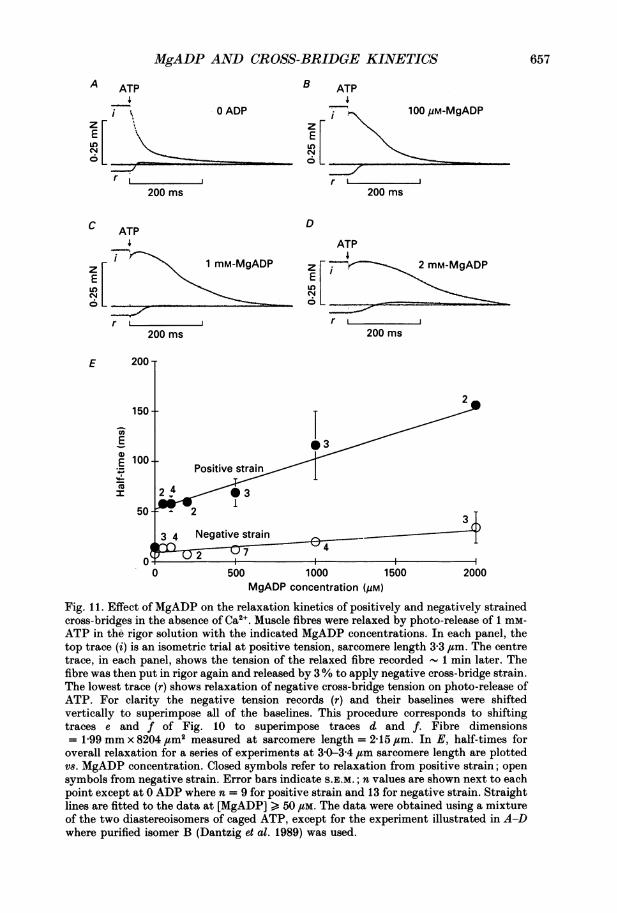
Figure 11 shows tension transients initiated from sarcomere length 3 30 4um, in thepresence of 20 mM-BDM, when the cross-bridges were negatively and positivelystrained in steady MgADP concentrations from 0 to 2 mm. Steady levels ofMgADPfrom 50 fM to 1 mm had much less effect on the rate of relaxation from negativetension than from positive tension. Two millimolar MgADP slowed relaxation fromboth positive and negative tensions, but the slowing was more marked from positivetension. Similar results were obtained using 25 mM-Pi instead of BDM to suppresstension of reattaching cross-bridges (data not shown).kD and k+D were estimated with stretched fibres in the presence of BDM using an
analysis similar to that discussed for Fig. 6A and Scheme II. A straight line wasfitted to a plot of ti values for overall relaxation of a series of seven fibres vs. MgADPconcentration in the range 50 JtM to 2 mm (Fig. IIE). Overall relaxation rather thanthe tension difference trace was used to estimate klD and k+D in this case becausetension transients from varying initial rigor levels did not converge at long sarcomerelengths, suggesting that positively strained, reattached cross-bridges contribute tothe tension difference traces. However, in the presence of Pi or BDM, the maincomponent of original tension recordings from negative strain was the tensionincrease due to detaching, negatively strained cross-bridges. The values obtainedfrom the ti analysis were k-D = 13 s-5 and 77 s-1 at positive and negative strainrespectively. k+D = 1-6-5-2 x 105 m-1 s-1 at positive and negative strain. These resultsindicate that in the presence of MgADP, relaxation from rigor is markedly straindependent in the direction to cause relaxation more rapidly from negatively strainedcross-bridges.
Ca2+ activation by photolysis of caged ATP in the presence ofMgADPPhotolysis of caged ATP in the presence of Ca2+ rapidly activates muscle fibres
from rigor. In the absence of added ADP, release of ATP (Fig. 12A, arrow) initially
656
MgADP AND CROSS-BRIDGE KINETICS
A ATP B4
I.zF
ElLo
C
IL6 L
E
0)
E-._
I
0 ADP
dCM
r,200 ms
ATP4
1 mM-MgADP
r 2 m200 ms
200 T
IU 1
ATP4
100 UM-MgADP
IzFEl
_AI
r L200 ms
D
ATP
z C j, , mM-MgADPEl
oL
r i2m200 ms
3 4 Negative strainnt ) i iS7 4
0 500 1000MgADP concentration (#M)
1500 2000
Fig. 11. Effect of MgADP on the relaxation kinetics of positively and negatively strainedcross-bridges in the absence of Ca2 . Muscle fibres were relaxed by photo-release of 1 mr-ATP in the rigor solution with the indicated MgADP concentrations. In each panel, thetop trace (i) is an isometric trial at positive tension, sarcomere length 3-3 Jim. The centretrace, in each panel, shows the tension of the relaxed fibre recorded 1 min later. Thefibre was then put in rigor again and released by 3% to apply negative cross-bridge strain.The lowest trace (r) shows relaxation of negative cross-bridge tension on photo-release ofATP. For clarity the negative tension records (r) and their baselines were shiftedvertically to superimpose all of the baselines. This procedure corresponds to shiftingtraces e and f of Fig. 10 to superimpose traces d and f. Fibre dimensions= 1-99 mm x 8204 #um2 measured at sarcomere length = 2 15 ,tm. In E, half-times foroverall relaxation for a series of experiments at 3 0-3 4 ,um sarcomere length are plottedvs. MgADP concentration. Closed symbols refer to relaxation from positive strain; opensymbols from negative strain. Error bars indicate S.E.M.; n values are shown next to eachpoint except at 0 ADP where n = 9 for positive strain and 13 for negative strain. Straightlines are fitted to the data at [MgADP] > 50 /tM. The data were obtained using a mixtureof the two diastereoisomers of caged ATP, except for the experiment illustrated in A-Dwhere purified isomer B (Dantzig et al. 1989) was used.
657
2
0 3
Positive strainI-T.
2 4 0 31
,, T
150
ioo0
:nE

J. A. DANTZIG AND OTHERS
caused tension to decrease due to cross-bridge detachment (Goldman et al. 1984b).Subsequent cross-bridge reattachment and force development resulted in a rise inforce to the steady active level. The in-phase stiffness decreased from the rigor valueto a moderate level, indicating that fewer cross-bridges are attached during an active
A 0 ADP B 200 gM-MgADP
0.25 [ s
0 n7,_, ess01 In-Quadrtue stiffness _ _ _ _ _ _ _
1
LO [ =? Tension
s-i s-i_~~~~~~~~~~~~~~~~~~~~~ _100 ms 100 ms
Fig. 12. Tension and stiffness transients obtained following photo-release of 1-5 mM-ATPin the presence of Ca2+ with 0 (A) and 200 /LM-MgADP (B). Tension traces labelled i areisometric and those labelled s were stretched by 0 4% 1 s before the laser pulse (arrows).Simultaneously recorded quadrature and in-phase stiffness records are shown at the topas indicated. The upper stiffness traces were recorded in the pre-stretch trial. The flattraces are baselines recorded in relaxing solution. The lowest traces (s- i) are algebraicdifferences for the pairs of tension traces. Sarcomere length = 2-31 ,um; fibre dimen-sions = 2-30 mm x 8672 /sm2; temperature = 18-6 'C.
contraction than in rigor. The quadrature stiffness increased to a steady positivelevel reflecting quick stress relaxation of the force-generating cross-bridges (Kawai &Brandt, 1980).MgADP slowed tension development following photolysis of caged ATP in the
presence of Ca2" (Fig. 12B) and reduced the amplitude of the initial tension decreaseobserved in the absence of ADP (Fig. 12A). The overall shapes of the stiffness traceswere similar to those obtained in the absence of added ADP although stiffnesschanged more slowly in the presence of MgADP.MgADP also slowed convergence of pairs of tension traces starting at different
rigor tension levels (Fig. 12). Half-times of the tension difference traces, in thepresence of Ca2+, are plotted as a function ofMgADP concentration in Fig. 6A (opensymbols). The line fitted to the open symbols in Fig. 6A corresponds toti (ms) = 19-6+24-3[MgADP] where [MgADP] is in mm. As discussed earlierregarding the corresponding data in the absence of Ca2+ and Scheme II, the line fittedto the data in the presence of Ca2+ provides estimates for ADP dissociation (klD) andassociation (kl+D) rate constants of kD = 35 s-1 and k+D = 2-5 x 105 M-1 s-, cor-responding to aKD of 70-170 /,M. k/+D is approximately equal to k/+T (MgATP-induceddissociation of rigor cross-bridges) and does not depend on Ca2+. k/D, estimated fromthe convergence half-times, is approximately 3-5-fold greater in the presence than inthe absence of Ca2+. This accounts for the more rapid convergence of the tensiontraces at [MgADP] > 20 /M in the presence of Ca2+ than at zero [Ca2+].
658
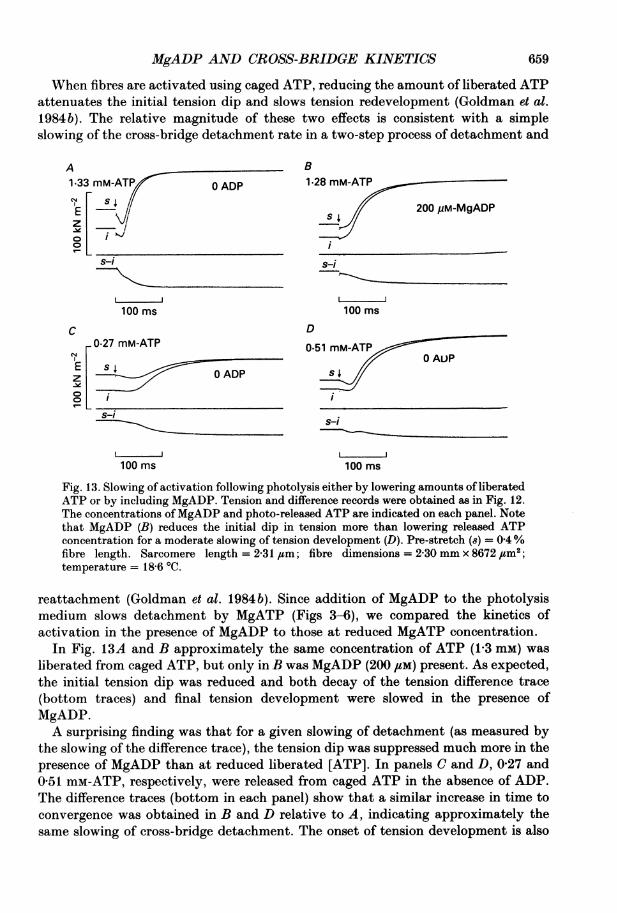
MgADP AND CROSS-BRIDGE KINETICS 659
When fibres are activated using caged ATP, reducing the amount of liberated ATPattenuates the initial tension dip and slows tension redevelopment (Goldman et al.1984b). The relative magnitude of these two effects is consistent with a simpleslowing of the cross-bridge detachment rate in a two-step process of detachment and
A B1.33 mM-AT/ 0 ADP 1 28 mM-ATP
E F 200 uM-MgADPz V
S-i S-i
100 ms 100 ms
C D0.27 mM-ATP 0-51 mM-ATP
0 AU)PE s | OADPz
s-isi
100 ms 100 ms
Fig. 13. Slowing of activation following photolysis either by lowering amounts of liberatedATP or by including MgADP. Tension and difference records were obtained as in Fig. 12.The concentrations of MgADP and photo-released ATP are indicated on each panel. Notethat MgADP (B) reduces the initial dip in tension more than lowering released ATPconcentration for a moderate slowing of tension development (D). Pre-stretch (8) = 0 4%fibre length. Sarcomere length = 2-31,um; fibre dimensions = 230 mm x 8672,#m2;temperature = 18 6 'C.
reattachment (Goldman et al. 1984 b). Since addition of MgADP to the photolysismedium slows detachment by MgATP (Figs 3-6), we compared the kinetics ofactivation in the presence of MgADP to those at reduced MgATP concentration.
In Fig. 13A and B approximately the same concentration of ATP (1-3 mM) wasliberated from caged ATP, but only in B was MgADP (200 ,UM) present. As expected,the initial tension dip was reduced and both decay of the tension difference trace(bottom traces) and final tension development were slowed in the presence ofMgADP.A surprising finding was that for a given slowing of detachment (as measured by
the slowing of the difference trace), the tension dip was suppressed much more in thepresence of MgADP than at reduced liberated [ATP]. In panels C and D, 0-27 and0-51 mM-ATP, respectively, were released from caged ATP in the absence of ADP.The difference traces (bottom in each panel) show that a similar increase in time toconvergence was obtained in B and D relative to A, indicating approximately thesame slowing of cross-bridge detachment. The onset of tension development is also

U60 J. A. DANTZIG AND OTHERS
similar in panels B and D. However, the transients in B and D are qualitativelydifferent in that the reduction of initial tension dip is more marked in B (200 gm-MgADP) than D (reduced MgATP). If the liberated [ATP] is reduced further (panelC), so that the attenuation of the dip is closer to that in the presence of MgADP, the
A
s4500 gM-MgADP
B
s 100uM-MgADP
Cs fi
20 pM-MgADP
D
r D MgADPE .z .
100 msFig. 14. Comparison of tension responses on photo-release of 770 ,SM-ATP in the presenceof Ca2+ and various MgADP concentrations as indicated. In each case pairs of recordsfrom a fibre in rigor (isometric, i) and stretched by 0 5% (s) were superimposed. In A,the flat trace is a baseline recording in relaxing solution. Pairs of traces in A, B and C wereshifted vertically for clarity. The short horizontal lines are the corresponding baselines.Sarcomere length = 2-26 ,tm; fibre dimensions = 2-26 mm x 4298 ,#m2.
decay of the difference trace and final tension development are much slower(compare B and C). Thus addition of MgADP to the medium causes a greaterattenuation of the tension dip than reducing the concentration of released ATP forthe same slowing of tension redevelopment. Also, as the steady MgADP con-centration was increased from 0 to 500 /M, the reduction of the tension dip was moremarked with little slowing of the subsequent tension redevelopment up toapproximately 100 #M (Fig. 14). Above 200 /tM-MgADP, slowing of force develop-ment became appreciable.The pronounced reduction in dip of the tension transient was unexpected even

MgADP AND CROSS-BRIDGE KINETICS 661
when allowance was made for a 15% drop in rigor tension when ADP is bound to thecross-bridges (Fig. 1). This observation suggests that the states populated at earlytimes following MgATP binding can support tension. To make the tension productionat early times following photolysis of caged ATP more evident, the steady value of
A ATP BTATP
E PP 200 pM-MgADP, 0 Pi
LLJs-i
100 ms
0APs 0 ADP, 10 mm-Pi
,j100 ms
D ATP
s4 200 pM-MgADP, 10 mM-Pi
s-i s-i
lOOms lOOms
Fig. 15. Effects ofMgADP and P, on tension responses following photo-release of 780 /M-
ATP in the presence of Ca2 . MgADP and Pi concentrations are indicated in each panel.Other conditions as in Fig. 12. The pre-stretch for traces labelled 8 was 04%. In thepresence of MgADP and Pi, tension initially rises following release of ATP, which is notobserved in the absence of ADP. Sarcomere length = 2'31 /sm; fibre dimensions= 2 54 mm x 6953 #um2; temperature = 22 4 'C.
tension was reduced by adding Pi to the photolysis solution. Pi is thought to suppresssteady tension by decreasing the proportion of force-producing AM'.ADP states(Cooke & Pate, 1985; Hibberd et al. 1985). Figure 15 shows that Pi suppressed finaltension to a similar level in the absence and presence of MgADP (panels C and D).Pi had little effect on the decay of tension difference traces (s- i). In the presence ofPi and MgADP, a transient increase in tension was observed following release ofATP(panel D) instead of a dip. This transient tension increase was absent when no ADPwas added (Fig. 15 C). The transient tension increase in Fig. 15D suggests that in thepresence of MgADP, states populated upon MgATP binding (AM.ATP and/orAM.ADP.Pi) bear tension or else that very rapid MgATP-induced detachment ofrigor cross-bridges bearing negative force increases net tension.
DISCUSSION
Kinetics of relaxation and activation, initiated by photolysis of caged ATP, havea complex dependence on MgADP concentration. The present results suggestinteractions between AM.ADP cross-bridges and neighbouring sites as well as
C As
E Iz _

J. A. DANTZIG AND OTHERS
interactions with the Ca2+-regulatory system. The rate of relaxation from rigor in thepresence of MgADP is markedly sensitive to the mechanical strain in the cross-bridge. The data presented here support the idea that ADP release has a role incontrolling the cross-bridge cycling rate.
Steady-state binding of MgADP to rigor cross-bridgesA small decrease of rigor tension was observed on adding MgADP to rigor fibres
(Figs1 and 2) when care was taken to reduce the level of contaminant ATP withinthe fibres. Marston et al. (1979) and Schoenberg & Eisenberg (1987) also observed thistension decrease. The MgADP concentration (KD) at half-saturation of the tensionreduction was found to be 18 /tM, which is below other estimates forKD in rigor musclefibres in the absence of Ca21: 40 gM ([14C]ADP binding to rigor fibres, Marston, 1973);60 /M (competition between ADP and either pyrophosphate or adenylyl-imi-dodiphosphate (AMPPNP) for binding to rigor fibres detected by the rate ofmechanical stress relaxation, Schoenberg & Eisenberg, 1987). The MgADPconcentration at half-saturation of the reduction of rigor tension may be lower thanthose other estimates for KD because the distribution of rigor strain among cross-bridges may be wide and the tension reduction is governed largely by the cross-bridges that exert the most tension. Ligands such as MgADP might bind moretightly to those cross-bridges than to ones with average strain (Webb et al. 1986).
These values for KD contrast with estimates at saturating levels of Ca2+:250-400,/M (Table 2 of the present study, see below); 200-300,UM (inhibition ofactive shortening velocity, Cooke & Pate, 1985); and 360 /tM (decrease in rateconstant for low-frequency sinusoidal response, Kawai & Halvorson, 1989).With isolated actomyosin, intermediate KD values have been obtained: 200,UM
(competitive inhibition by MgADP of MgATP-induced dissociation of actomyosinsubfragment-1 (acto S-1) and acto-heavy-meromyosin, White, 1977); 140,UM(dissociation of acto S-1, Greene & Eisenberg, 1980); 140 /IM (inhibition of acto S-IATPase activity, Biosca, Greene & Eisenberg, 1986); and 100 /tM ([3H]ADPbinding to myofibrils, Biosca, Greene & Eisenberg, 1988). Dissociation of thenucleotide analogue 1-N6-ethenoadenosine 5'-diphosphate from regulated acto S-1(with troponin and tropomyosin present) is 10- to 15-fold more rapid in the presencethan in the absence of Ca2+ (Rosenfeld & Taylor, 1987a, b), consistent with anapparent tendency for weaker MgADP binding when Ca2+ is bound to the contractileregulatory system.
In the present work, an apparent effect of Ca2+ on KD is observed in comparing fitsof a kinetic model to transient data in the presence of Ca2+ (Table 2,KD = 250-400 /tM) with titration of the MgADP-induced tension reduction in theabsence of Ca2+ (Fig. 2, KD = 18,UM). But a direct comparison of the presence andabsence of Ca2+ in the latter type experiment failed to reveal a clear effect of Ca2+ onKD. Further experiments will be required to determine if Ca2+ affects equilibriumMgADP binding to rigor cross-bridges.To explain the tension decrease on adding MgADP to a fibre in rigor we should
consider either cross-bridge detachment (possibly one or both heads) or else astructural change in the heads themselves. Tension recovered when ADP was washedfrom a fibre (Fig. 1), suggesting that either a reversible structural change occurredor that cross-bridges detached by MgADP had to rebind to the original actin
662

MgADP AND CROSS-BRIDGE KINETICS
monomers when ADP was removed. Electron paramagnetic resonance data fromexperiments with fibres spin-labelled at the reactive cysteine residue (SH-1) in themyosin head showed virtually no change of orientation when ADP was bound(Thomas, Svensson & Polnaszek, 1985; Fajer, Fajer, Matta & Thomas, 1989)indicating no detachment. Birefringence and X-ray diffraction measurements ofunlabelled fibres also indicated little detachment (Obiorah & Irving, 1989). Thus thereduction of rigor tension shown in Fig. 1 suggests that MgADP binding induces asmall structural change in attached cross-bridges.
If this inference is correct, we can estimate the size of the motion induced byMgADP binding on the assumption that the compliant part of the cross-bridge isstretched by 4 nm on average during isometric contractions (Ford, Huxley &Simmons, 1977), 75% of the cross-bridges attached in rigor are attached duringactive contractions (Haselgrove & Huxley, 1973; Matsubara, Yagi & Hashizume,1975; Goldman & Simmons, 1977; Fig. 12 of the present paper) and that theforce-extension curve at the cross-bridge is linear (Ford et al. 1977). Rigor force isapproximately 40 % of active force, so the average stretch on the compliance of therigor cross-bridge is 1P2 nm (0 40 x 0 75 x 4 nm). Since the force reduction on bindingMgADP averaged 15% of rigor force, the apparent motion in the cross-bridge is 15%of 1-2 nm or 0418 nm. This can be considered a small structural change compared tothe 4-12 nm motion estimated for the power stroke during active contractions (Fordet al. 1977; Huxley & Kress, 1985).
Relaxation from positive rigor tension in the presence ofMgADPFibres are expected to relax more slowly on photolysis of caged ATP when ADP
is bound to the ATPase sites of the cross-bridges than in the absence of ADP as wasobserved. However, the shape of tension traces obtained indicates that a complexinteraction between MgADP and the cross-bridges affects the kinetics of relaxation.In particular the plateau of tension maintenance (phase bt) and sudden transition toa more rapid relaxation rate (Figs 3-9) are striking departures from expectationbased on simple kinetic schemes. As presented in the Results section, the enhancedquadrature signal during the plateau and the effects of Pi and BDM suggest that thetension plateau is predominantly due to reattaching cross-bridges. Since stiffnessfalls continuously during the plateau and small stretches accelerate termination ofthe plateau, the sudden fall in tension seems to indicate a threshold for exceeding thelimiting force per attachment.
According to this limiting force hypothesis, the sequence of events duringrelaxation in the presence of MgADP would be as follows: before the laser pulse, thecross-bridges are distributed between AM and AM.ADP states according to theMgADP concentration and the dissociation constant (KD) for MgADP. The thinfilament regulatory system is derepressed due to the rigor attachments. When ATPis liberated, the AM cross-bridges detach promptly (Figs 4 and 5). AM.ADP cross-bridges detach more slowly maintaining the derepressed state of the thin filaments.Tension is further maintained or increased by reattaching cross-bridges, but thenumber of detaching cross-bridges exceeds the number reattaching, so stiffness fallscontinuously. Remaining cross-bridges are under steadily increasing stress until theforce becomes too great and some of them detach. The resulting decrease of force-bearing capacity leads to sarcomere elongation and even higher force per remaining
663

J. A. DANTZIG AND OTHERS
attachment. The process then becomes regenerative and relaxation acceleratesabruptly. The final tail of tension relaxation may indicate release of ADP fromand/or Pi binding to AM'.ADP cross-bridges. These postulated events are givenstrong support by the sudden length change detected by light diffraction at the timeof the transition to rapid relaxation (Fig. 8), the fact that the final relaxation hasessentially the same kinetics in the absence and presence of MgADP and that theduration of the plateau and final rate of relaxation are sensitive to Pi concentration.The period during which cross-bridges reattach is also extended when the rate of
cross-bridge detachment is reduced by lowering the concentration of released ATP(Goldman et al. 1984a); however, lowering released ATP does not lead to thethreshold phenomena discussed above for the experiments with MgADP in theabsence of Ca2 . Some mechanism for enhancing the force-bearing capacity ofreattaching cross-bridges is required to explain the dramatic prolongation of theplateau tension when MgADP is present despite net cross-bridge detachment.Several possible effects of MgADP on the cross-bridges and their relationship to theCa2+-regulatory system may be involved.
Since the highly non-exponential, threshold behaviour of Figs 2-9 was onlyobserved in the absence of Ca2+, the troponin-tropomyosin regulatory systemprobably takes part in the events during relaxation. At the transition to phase ct, theregulatory proteins may suddenly shut the thin filaments off contributing to fasterrelaxation.
In the presence of MgADP, cross-bridges may either be more likely to reattach tothe thin filaments or when they do reattach, they may generate more tension thanin the absence of ADP. Stiffness recordings suggest more force per cross-bridge andalso possibly increased reattachment. The myosin heads able to reattach and go onto produce force are presumably in the M.ATP or M.ADP.Pi state, so these effects ofMgADP in the medium would not operate directly on those particular heads. Thuswe are led to the idea that AM.ADP cross-bridges increase the likelihood ofreattachment or the force per reattachment of neighbouring myosin heads. Force perattachment could be enhanced if AM.ADP states constrain other myosin heads toreattach to the same actin monomer they bound before being detached by MgATP.This mechanism would cause maintenance of tension and also the slowed decay of thetension difference traces (Figs 3-9). These possible mechanisms for the effects ofMgADP on relaxation are not mutually exclusive.
In light of the complexities related to reattaching cross-bridges, it follows that thetension records in the absence of Ca2+ do not give a direct indication of the ADPdissociation rate constant. However, the extended plateau (50-100 ms) suggests thatADP is bound to cross-bridges throughout this period implying a net dissociationrate constant in isometric conditions of approximately 10-15 s-1 (Fig. 6A).The range of values reported by previous investigators for ADP release from
AM.ADP in the absence of Ca2+ is very broad. White (1977) and Siemankowski,Wiseman & White (1985) found that ADP dissociation from acto S-1 occurred at400 s-5 or greater. Several investigators have confirmed that ADP dissociation fromisolated AM.ADP is rapid (Greene & Eisenberg, 1980; Borejdo, Ando & Burghardt,1985; Rosenfeld & Taylor, 1987a, b). However, Geeves (1989) found that ADPdissociates from a subpopulation of acto S-1.ADP at only 2 s-1 in a process ascribed
664

MgADP AND CROSS-BRIDGE KINETICS
to an isomerization of AM.ADP. Thus ADP release from actomyosin may be slowunder some conditions.
Kinetics ofADP dissociation from negatively strained cross-bridgesRelaxation rate in the presence of MgADP depended on the level of rigor strain
(Figs 9 and 11). As mentioned in the Introduction, from the Fenn effect (1923) andits implications (Huxley, 1957), we expect some relatively slow cross-bridge reactionsto accelerate at low strains. The strain dependence of the rate of MgATP-inducedrelaxation observed from positive rigor tensions is opposite to this expectation (Fig.9 and Goldman et al. 1984 a) but the increased relaxation rate from negative tensionis in the expected direction.
It is apparent from the above discussion that cross-bridge reattachment precludesstraightforward measurements of ADP dissociation kinetics from tension recordedduring displacement of ADP by ATP in the absence of Ca2+. Therefore, tensionproduction by reattaching cross-bridges was suppressed by Pi or BDM. The resultanttension records obtained on MgADP displacement by MgATP were used to estimatethe influence of strain on the kinetics of ADP dissociation.
In the presence ofMgADP and either Pi or BDM, it was found that relaxation wasmarkedly faster from the negatively strained condition than when the cross-bridgeswere under the normal positive strain (Fig. 11). The simplest explanation for thisbehaviour is that ADP dissociation from AM.ADP is strain dependent, i.e. ADPdissociates faster from negatively strained than from positively strained AM.ADPstates. The direction of this strain dependence is appropriate for the expectationsfrom the Fenn effect and from the Huxley (1957) model: cross-bridges with negativestrain detach more rapidly than those with positive strain.From the tension recordings it is difficult to make an accurate estimate of the
enhancement of the ADP dissociation rate at negative tension because reattachingcross-bridges probably have some influence even when suppressed by Pi or BDM.When fibres were relaxed from negative tension, no indication of separate phases forMgATP-induced dissociation ofAM cross-bridges and slower dissociation ofAM.ADPcross-bridges (e.g. Fig. 4) was obtained. This result suggests either that negativelystrained cross-bridges are saturated with MgADP or that cross-bridges are in a rapidequilibrium with MgADP. If the former were the case, then the dissociation rateconstant ofADP is approximately 77 s-' from the ordinate intercept of a straight linefitted to the half-time data vs. [MgADP] (Fig. lIE, open symbols). In the latter case,KD would be 0O8 mm from the ratio of the intercept to the slope of the line fitted tothe half-time data. Assuming that k+D is similar to k+T (2-5 x 105 M-1 s-1), k/d(= KDk+D) is then estimated to be 160-400 s-1. These values for klD should becompared to the estimates for klD from positive strain: 13-5 s-' (short sarcomerelength, - Ca2+, -Pi, Fig. 6A, closed symbols), 46 s-1 (short sarcomere length,-Ca2 , +Pi, Fig. 7E), 35 s-' (short sarcomere length, +Ca2+,-P1, Fig. 6A, opensymbols) and 13 s- (long sarcomere length,-Ca2+, +Pi or +BDM, Fig. liE, closedsymbols).The data indicate that klD is strain dependent in a manner characteristic of the
rate-limiting step of the cross-bridge cycle as postulated by Huxley (1957). However,since a cross-bridge in rigor may not be the same as the nucleotide-free AM state of
665

J. A. DANTZIG AND OTHERS
the normal cross-bridge cycle, and because Ca21 is absent in the hegative strainexperiments, caution is needed in extrapolating this interpretation to activatedcross-bridges in contracting muscle.
Kinetics of intact fibre relaxationRelaxation of intact frog muscles or fibres is slower after prolonged tetanic
stimulation than after a twitch or short tetanus (Abbott, 1951). When the train of
A B
50 ms 50 ms
C D
50 ms 50 ms
Fig. 16. Simulations of effects of changing MgATP or MgADP concentrations in SchemeIII of the text. The rate constants for trace a in each panel were: k= 118 s-';k+:=2 x 105 M-1 s-1; k = 100 8-1; k = 150 s-1; and k-4 = 0 (absence of Pi). In thesimulation of trace a in each panel, 1 mM-MgATP was released in the absence of ADP. Inpanel A, traces b, c and d correspond to reducing photo-released MgATP concentrationsto 500, 250 and 125 /M respectively. In panels B, C and D traces b, c and d correspond torelease of 1 0 mM-ATP in the presence of 200 /LM-MgADP. In B, k+D is 2 x 105 M-1 s-I andk_D values are 80, 40 and 20 s-1 for traces b, c and d respectively. In panel C, k-D is set at40 s-1 and k+D values are 1, 2 and 4 x 105 M-1 s-I for trace b, c and d respectively. In panelD, trace b simulates release of 1 mM-MgATP in the presence of 200 /iM-MgADP withk+D = 2 x 105 M-1 s-1 and klD = 40 s-1. Trace c is the same except that tension (Te) in the{AMATP} and {AM.ADP.Pi} states was assigned 60% of rigor tension instead of zero as in all theother traces. T7 > 0 reduces the amplitude of the initial tension dip.
stimuli ends, tension falls slowly for 200-500 ms (at 0 °C), then a transition occurs toinitiate much faster relaxation. This behaviour is qualitatively similar to therelaxation we observe in skinned rabbit fibres on photolysis of caged ATP in thepresence of MgADP. In both cases, substantial length changes suddenly take placeat the transition to the final rapid relaxation phase.
666

MgADP AND CROSS-BRIDGE KINETICS
There is evidence that the slowing of relaxation following tetanic stimulation infrog muscles is the result of saturation of Ca2+ binding sites on parvalbumin, asoluble Ca2+-binding protein, leading to decreased Ca2+ uptake by parvalbumin(Gillis, 1985). In support of the parvalbumin hypothesis are measurements ofresidual free Ca21 levels above baseline (Cannell, 1986) and heat measurements(Peckham & Wolledge, 1986; Rall, 1989). However these data do not exclude acontribution to slowing of relaxation by accumulation ofMgADP or other metabolicchanges during the tetanus. If the MgADP concentration within a fibre increasesfrom 10 to 20 /tM during a prolonged tetanus, the accumulated MgADP would causesignificant slowing of relaxation, according to the behaviour of rabbit skinned fibres(Fig. 6A). The relationship between myoplasmic Ca21 levels and specific cross-bridgeevents during relaxation of intact muscles is not clear at present.
Caged ATP photolysis in the presence ofMgADP and Ca2+In caged ATP experiments in the presence of Ca2+, inclusion of MgADP slowed
activation and slowed convergence of pairs of tension traces starting from differentrigor tension levels as expected on the basis of competition between MgADP andMgATP for the nucleotide binding site. MgADP caused a dramatic reduction of theamplitude of the transient dip before active tension generation (Figs 12-16). Thisattenuation of the dip was more prominent than would be expected on the basis ofkinetic schemes in which the states AM.ATP and AM.ADP.Pi are not considered tobear significant force. In this type of scheme, major reduction of the tension dip isalways associated with more pronounced slowing of final tension development thanwas observed in the experiments (Fig. 16).To determine the expected relationship between the amplitude of the tension dip
and slowing of tension development the following kinetic model (Scheme III) wassimulated with various choices for the rate constants and nucleotide concentrationsto compare with the experimental results:
kiCaged ATP -l' ATP
ADP ATP PiAM . ADP AM
l {AM. ATP1 ff JAM. ADP. P1 AM'LADPAM.ADP~~~~~~~~~~~~~~~~-~~AM 1M.ATP J M.ADP.P1 --A' D
(Scheme III)
This scheme (III) is a simplified form of the biochemical model (Scheme I)presented in the Introduction, uniting the AM.ATP and M.ATP states into a singleentity and likewise combining AM.ADP.Pi with M.ADP.Pi on the assumption thatthe attachment-detachment kinetics of these states are rapid. For this discussion,the lumped states ({'m } and {AM.ADPPi}) are termed rapid equilibrium states andare initially considered not to support significant force. The kinetic model includedthe photolysis dark reaction rate (k1 = 118 s-1) for release of ATP from caged ATP(Goldman et al. 1984a).
In Fig. 16A, the effect of changing photo-liberated ATP concentration wassimulated in the absence of ADP. Traces a-d correspond to lowering released [ATP]
667

J. A. DANTZIG AND OTHERS
from 1 mm to 125 /tM. As [MgADP] is reduced, the initial tension dip and the rate offinal tension activation are both reduced as observed experimentally in the absenceof ADP (cf. Fig. 13, panels A, C and D, and Goldman et al. 1984b). Figure 16B andC shows control simulations in the absence ofADP (traces a) and the effect of adding200 /SM-MgADP (traces b-d) with various choices for k+D and k/D. In panel B, k+D isheld constant (2 x 105 M-1 s-1) and k/D is varied from 80 to 20 s-5. In panel C, klD isconstant (40 s-1) and k+D values are 1-4 x 105 M-1 s-' for traces b-d. Panels B andC are qualitatively similar to panel A; in each case, when MgADP is added, thereduction of the tension dip is accompanied by a more pronounced slowing of thefinal development of tension than was observed experimentally (Figs 12-16). Asnoted earlier, adding MgADP to the photolysis medium causes a striking reductionof the tension dip with less of a decrease in the activation rate than in the simulationsof Fig. 16B and C.A simple alteration to the simulation that does mimic the experimental behaviour
fAM.ATP AM.ADP.P1is shown in Fig. 16D, trace c. Tension in the {t MATP } and { M ADP Pi } states was setto 60% of rigor tension for trace c instead of zero as in panels A-C. In this case, thedip in tension is reduced without such marked slowing of tension development.
Assigning tension values to the rapid equilibrium states corresponds to incompleteloss of mechanical strain while cross-bridges are populating those states. To quantifythe tension required in {AM.ATpP} and {AMADP.Pi} to explain the results, sets of four toeight tension traces (absence and presence of MgADP; isometric and pre-stretch)such as those in Figs 12 and 14 were fitted simultaneously to Scheme III whileallowing the tension (Te) in the {AMATjP} and {AM.ADP.Pi} states (relative to rigortension) to vary as an adjustable parameter. States AM and AM.ADP were bothassigned the rigor tension value. Active tension and k-D, k+D, k+T, k3 and k4 were allallowed to vary during the parameter optimization, but one set of rate constants wasapplied to all four tension traces simultaneously. Te adopted separate values for thetraces in the absence and presence of MgADP.A set of tension traces simultaneously fitted to Scheme III (Fig. 17) shows that the
shapes of the tension records and difference traces are consistent with this kineticmodel. Tension in the {AM.ATP} and { M,ADP,Pi } states was zero for panel A and 46%of rigor tension for panel B.The experimental records do not uniquely determine all three of the fitted
parameters k+D, k-D and k+T. Therefore, further constraints were placed on therelationship between these values. The experimental records were fitted using threeslightly different procedures. The first method was based on the assumption that k+Dand k+T have the same value. For the second method of fitting, k+D and k+T wereallowed to adopt separate values during parameter optimization, but klD was heldconstant at a high value (500 s-1, Siemankowski et al. 1985). In the third method k+D,k-D and k+T were all allowed to vary independently. The square symbolssuperimposed on the recordings of Fig. 17 represent the fit to the traces using the firstmethod (k+D = k+T) and correspond well to the recordings. The other two methodsalso fit the traces well, although with the second method (kD fixed at 500 s-'), thecalculated values for k+D were approximately an order of magnitude higher than k+T.Contrary to the simpler model previously used by Goldman et al. (1984b), no delaywas necessary in these simulations to match the time for the minimum of the dip in
668

MgADP AND CROSS-BRIDGE KINETICS
tension because an extra free parameter was included to fit the tension dip andredevelopment. k3 and k4 correspond to all of the steps from ATP hydrolysis throughforce generation and, therefore, the values obtained (Table 2) are not applicablespecifically to the hydrolysis and Pi release steps respectively.
Table 2 shows estimates (using the three procedures) for the various rate constantsin Scheme III for a series of eleven fibres and for actomyosin subfragment- 1 in
TABLE 2. Rate constants from fitting procedures 1, 2 and 3 and fromactomyosin S-I in solution
Fitting 1 2 3 Actomyosinparameter (k+T = k+D) (k-D = 500 s'-) (all vary) S-i Ref.k-D (8-1) 126+48 500 129+53 > 500 1k+D (M-1 S-1) 2-6+06 x 105 1-7 +05 x 106 4-5+ 16x 105 2-5 x 106 2k+T (M-1 S-1) 2-6+006 x 105 2-5+0 8 x 105 3-1 +07 x 105 106-107 3, 4KD (#UM) 265+50 395+ 120 318+97 150-200 2, 5k3 (s1) 347 +90 278+44 426+ 145k4 (s1) 164+ 16 217 +30 140+32n 11 4 5
k-4 of Scheme III (Pi binding) was set to zero to simulate conditions with no added Pi.References for actomyosin S-1 rate constants: 1, Siemankowski et al. (1985); 2, White (1977); 3,
Taylor (1979); 4, Geeves (1989); 5, Greene & Eisenberg (1980).
solution for comparison. The apparent dissociation constant, KD estimated from thefitting procedure as the ratio of the ADP dissociation rate (k/D) and binding rateconstant (k+D) was 250-400 /LM, much higher than the 18 ftM value obtained in theabsence of Ca2+ from titrating the steady-state reduction of rigor force.The tension values (Te) assigned to the {MAT} and {MADPi} states by the fitting
procedure are plotted vs. the MgADP concentration in Fig. 17C. In the absence ofADP, Te was 2'5+4 7% (S.E.M., n = 11) of the rigor tension value, indicating that,following MgATP binding, the rapid equilibrium states do not bear tension. As theMgADP concentration increased, Te amounted to a significant percentage of rigortension, e.g. 48+5% (S.E.M., n = 6) at 200 ,tM-MgADP.The experiment shown in Fig. 15 provides further support for the idea that in the
presence of MgADP, tension develops very early (within 5 ms) upon MgATP binding.Assuming that Pi suppresses the tension due to cross-bridges in the AM'.ADP state(Hibberd et al. 1985; Goldman, 1987 b), the transient increase of tension on release ofATP in the presence of MgADP (Fig. 15D) and the absence of the transient increasein the absence of ADP (Fig. 15 C) suggest that the immediate tension generationoccurs only when MgADP is present. Two explanations of this phenomenon areconsidered: strain-dependent rate of ADP dissociation from AM.ADP and co-operative interactions between cross-bridges.
Simulation of strain-dependent rate of ADP dissociationWhen negatively strained cross-bridges detach, net tension generated by a fibre
increases (Figs 10 and 11). Cross-bridge detachment appears more rapid fornegatively strained cross-bridges than for positively strained ones (Fig. 11). If theinitial strain distribution of rigor cross-bridges includes very high forces as well as
669

J. A. DANTZIG AND OTHERS
B
200 uM-MgADP
s-i.
0100 ms
T
j. .
5000.0+ 1
0 100 200 300 400Steady MgADP concentration (jiM)
Fig. 17. Simulations based on text Scheme III fitted to tension records obtained followingphoto-release of 660,uM-ATP (+ Ca2+) in the absence (A) and in the presence (B) of200 /,M-MgADP. The pre-stretch for traces labelled 8 was 0-5 %. Other conditions as inFig. 12. A rapid-descent optimization method (Fletcher & Powell, 1963) was used tominimize the total sum of the squares of the residuals between the four simulated tracesand the four corresponding experimental tension records. The symbols representsimulated traces for Scheme III with the following rate constants:k+D +T= 6X 105 M-1 s-1 k D = 80-3 s-1; k3 = 166 s-1; k4 = 159 s-1; k-4 =0. Te (tensionin the { M }ATP } and {AM.AD p } states) was zero in A and 46% of rigor tension in B.
Sarcomere length = 2-20 ,um; fibre dimensions = 2-20 mm x 4211 ,sm2; temperature= 21 'C. In panel C, Te (relative to rigor tension) obtained by fitting Scheme III to setsof four to eight tension traces as in A and B is plotted against MgADP concentration inthe photolysis trials. Data are plotted as means+ S.E.M.; n values between 2 and 11. k+Twas constrained to equal k+D (fitting method 1 as described in the text). Note that Te atzero MgADP is not significantly different from zero. The other two text methods ofconstraining the parameters yielded 1T, values within 2% of the values plotted.
negative forces, then in the presence of MgADP, quick detachment of the negativelystrained cross-bridges would increase net tension before the slower detachment ofpositively strained cross-bridges. Simulations of a kinetic model based on these ideas
A
Ez
r-.
0 MgADP
mSKi
100 ms
1.0 T
0.8 -
0.6+
C
o0)0._
0
40
Co
0)'._n 0.4 +
ISIT
I
0.21+
670

MgADP AND CROSS-BRIDGE KINETICS
are shown in Fig. 18A-D (negative strain model). Details of the calculations aregiven in Appendix A, but the essential features are that a wide distribution of rigorcross-bridge strains is simulated by considering positively and negatively strainedcross-bridges independently according to Scheme III above and then the sum of theforces from the two groups is taken as the simulated fibre tension. Force valuesassigned to the positively and negatively strained rigor cross-bridges are given inAppendix A. Tension values for the {AIATTP} and {AM.ADP.jP'} states are set to zero. Therate constant for MgADP binding to AM (k+D in Scheme III) is assumed to be5 x 105 M-1 s-I irrespective of strain. ADP dissociation from AM.ADP (k-D) is 25 s-1for force > zero and 250 s-5 for force < zero. These values correspond to MgADPdissociation constants (KD) of 50 /M at positive strain and 500 /M at negative strain.The suppression of active tension by Pi is simulated by Pi binding to AM'.ADP at1.5 x 104 M-1 S-1 (Hibberd et al. 1985; Webb et al. 1986).Many features of the recordings in Fig. 15 are explained by the negative strain
model (Fig. 18A-D). Suppression of the tension dip following ATP release in thepresence of MgADP occurs in the simulations (panels B and D) because positivelystrained cross-bridges that are initially in the AM.ADP state detach relativelyslowly. In the presence of MgADP and Pi, tension rises briefly above the final activelevel because rapid detachment of negatively strained cross-bridges leads toincreased net tension. At 200 /LM-MgADP, most negatively strained cross-bridges areinitially in the AM state because their affinity for MgADP is low. In the presence ofMgADP an initial small tension dip before the tension rise is due to detachment ofthe positively strained cross-bridges that are in the AM state at the time of ATPrelease (1/5 at 200 /SM-ADP and KD = 50 sM). There is a hint of such a brief tensiondip in the recordings with MgADP present (Figs 12-15 and 17), but the instantaneoustension drop caused by the laser pulse in most experiments makes this aspect difficultto resolve.The main assumption of the negative strain model that is not based on
independent data is the wide distribution of rigor strains. For simplicity, two discretetension values were chosen for the simulations: AM states in the isometric conditionwere given tension values of 1-5 and - 025 relative to active tension. But a realisticdistribution would not be discrete and thus would need to be even wider to obtainthe amplitude of tension rise shown in Figs 15D and 18D. Width of the rigor straindistribution is determined by the mismatch of thick and thin filament periodicitiesand by possible filament sliding during induction of rigor after some rigor cross-bridges have formed.
Simulation of interactions between paired sitesAn alternative explanation of the decreased tension dip following ATP release in
the presence ofMgADP that does not require a wide distribution of rigor cross-bridgestrains is to assign tension to the rapid equilibrium states as in the simulations of Fig.17. But neither { M.ATP} nor { M.ADP.Pi } can bind MgADP, so it is not readily apparentwhy their capacity to sustain tension should depend on [MgADP]. A possible answeris that one head of a myosin molecule may influence the rate of attachment to actinor the tension of the other head of that molecule. Consider the situation when one ofthe heads of a pair detaches upon binding MgATP and the other head remains
671

J. A. DANTZIG AND OTHERS
Negative strain model
A 0 ADP, 0 Pi
S
C 0 ADP, 10 mM-Pis
i --.
V_,
200 ms
B 200 pM-MgADP, 0 Pi
S
D 200 jM-MgADP, 10 mM-Pi
s
j
0 m200 ms
Co-operative model
E 0 ADP
Ss I
G 400 pM-MgADP;altered stress
s
I .
20
200 ms
F 400 uM-MgADP;standard parameters
S
H 400 uM-MgADP;altered rate constant
S --)
s \/v
200 ms
Fig. 18. Tension transients simulated using kinetic models for the isometric (i) and pre-stretch (s) conditions. A-D were calculated using Scheme III as described in the text andAppendix A. Two groups of cross-bridges with either positive or negative strain weresimulated independently and then combined by algebraic addition of the time courses ofsimulated tension. Rate constants and tension values assigned to the intermediates aregiven in Appendix A. ADP dissociation is assumed to occur at 25 s-5 from positivelystrained cross-bridges and 250 s-5 from negatively strained ones. 1 mM-ATP was photo-released in the simulations in the absence and presence of MgADP and Pi as indicated ineach panel. Panels A-D should be compared to Fig. 15. Panels E-H were calculated usinga co-operative model (Scheme V) described in the text and Appendix B. Rate constantsand tension assignments for the model are given in Appendix B and Table 3. Panel E is acontrol set of simulated traces at 500 uM-released ATP and zero ADP. F shows the effectof including 400 #M-MgADP with no simulated co-operativity (i.e. the rate constants andtension assignments follow directly from the single-site Scheme IV). For G, co-operativitywas introduced by assigning extra tension to state LD (one cross-bridge (L) containing
672

MgADP AND CROSS-BRIDGE KINETICS
attached with MgADP bound. The detached head may have a restricted range ofactin sites for reattachment, resulting in net tension in the AM.ATP and AM.ADP.Pistates. It may also spend a higher proportion of time in attached configurations.Another, perhaps less likely, possibility is that if a head in a rapid equilibrium statehas a neighbouring head in the AM.ADP state, then that head is more rapidlytransformed into the activated tension-bearing state AM'.ADP.To test these ideas qualitatively, a model containing pairs of myosin cross-bridge
sites (possibly myosin heads) was devised. The detailed reaction scheme for thismodel is presented in Appendix B, but the essential features are that a simple four-state reaction scheme is used for the individual cross-bridges and sites are combinedinto pairs corresponding to combinations of the four states. The model considered fora single site is:
kiCaged ATP - ATP.ADP ATP RDk gk A(Scheme IV)
L R--R
k+D
where L is ligand-bound actomyosin formed from the rigor state (e.g. AM.ADP), Ris rigor (AM), D includes the detached states and corresponding rapid equilibriumattached states ({AM.ATP} and M.ADP.PiD}), and A is an actively force-generating cross-bridge (e.g. AM'.ADP). The ten possible pairs of these intermediates (e.g. LL, LR,LD, LA, RR, RD, etc.) were arranged on a reaction scheme diagram with the sixteenpair-wise reaction steps (LL -- LR + ADP, LR -- RR + ADP, LR + ATP -+ LD, etc.;Scheme V, shown in Appendix B). In the pair-wise model, tension values assigned toeach state and reaction rates follow from the reaction scheme for an individual site.The initial condition for each simulation was taken to be a distribution between RR,LR and LL appropriate to binding of MgADP and with the selected binding anddissociation rate constants k2, k3, k6 and k7. Initial tension values assigned to statesRR, LR and LL were in the ratio 1 0: 0-925: 0-85 (isometric traces, i) to simulate thesmall decrease of rigor tension obtained when MgADP bound to a rigor fibre (Fig. 1).Pre-stretch traces (s) were simulated by increasing the force assignment of each statecontaining R or L cross-bridges as listed in Appendix B with further details of thetension assignments and reaction rates.
Figure 18E-H (co-operative model) shows transients simulated with choices ofrate constants indicated in the figure legend and Appendix B. Panels E and F aretransients simulated in the absence and presence of 400/,M-MgADP without co-operativity between sites. Tension assigned to individual detached sites (D) is zero.As shown for the single-site model of Fig. 16, in the presence ofMgADP, the simulatedtension dip is not reduced as much as in the experimental records.
Panel G is the same as F (400 /tM-MgADP), except that interaction between oneAM.ADP cross-bridge of a pair and the tension generation of its partner is simulated
bound ADP and one cross-bridge in a state (D) with rapidly equilibrating attach-ment-detachment kinetics). For panel H, co-operativity was introduced by a threefoldincrease in the rate of conversion ofLD to LA (one site containing bound ADP, its partneractively generating force). G and H show less tension dip than F.
673
22 PHY 432

J. A. DANTZIG AND OTHERS
by setting the tension of state LD the same as LR. This change corresponds tohypothesizing that when one attached head has ADP bound, detachment of itspartner does not reduce strain of the pair. Panel H is similar, but instead of addingco-operativity to the system by increasing the tension of state LD, the rate of activeforce generation (k14, LD -+ LA) by a detached head with an AM.ADP partner isaccelerated threefold. Traces in G and H are qualitatively similar to the experimentalrecords in MgADP with marked reduction of the tension dip relative to F. Thusinteractions between cross-bridge sites can explain the reduction of tension dip in thepresence of MgADP. The co-operative model considered here will not explain thebrief tension rise of Fig. 15D unless state LD is given considerably more force thanstate LL, which seems unlikely.
Control of the cross-bridge cycleIn activated isometric psoas muscle fibres, the cross-bridge cycling rate (judged by
the rate of ATP hydrolysis) is approximately 3 s-1 per myosin head at 20 °C, 200 mMionic strength (Webb et al. 1986) and most cross-bridges are attached in activatedisometric mnuscle (Haselgrove & Huxley, 1973; Matsubara et al. 1975; Goldman &Simmons, 1977). Measurements of MgATP-induced cross-bridge dissociation, ATPhydrolysis, tension development and kinetics of Pi release (Goldman et al. 1984b;Hibberd et al. 1985; Ferenczi, 1986) have indicated that all of these steps are too fastto limit the overall cycling rate in activated isometric muscle. Taken together, theseresults suggest that in activated isometric muscle fibres, either release of ADP fromAM.ADP or an isomerization ofAM'.ADP is the rate-limiting step of the cross-bridgecycle.The 3 s-I turnover rate implies that cross-bridge life-times are of the order of
300 ms. In the present experiments, mechanical changes related to ADP dissociationfrom the cross-bridges required 50-100 ms in the absence and 20-40 ms in thepresence of Ca2+. In activated isometric muscle, these ADP dissociation rates wouldnot contribute strongly to limiting the cross-bridge cycling rate, but it should benoted that the AM.ADP state achieved by adding MgADP to rigor muscle is not thesame as the AM'.ADP stat.p populated during contraction and whether AM.ADP is anormal intermediate of the ATPase pathway is not known.The complex mechanical transients indicate a Ca2' dependence of cross-bridge
detachment from AM.ADP. Thus ADP release may be an important control point forthe Ca2+-regulatory system, particularly during relaxation following a twitch ortetanus.
Since energy liberation increases during shortening (Fenn effect), we expect therate-limiting step in isometric conditions to be markedly accelerated by decreasedstrain or filament sliding during active fibre shortening. If the range of interactionbetween a myosin head and a thin filament site is t 10 nm (Huxley & Simmons,1971), at a filament sliding velocity of 2,um/s, the maximum time a cross-bridgecould remain attached to a particular thin filament site would be only 5 ms. In thissituation, a 20-100 ms time period for cross-bridge detachment from the AM.ADPstate could not easily be incorporated into the reaction sequence. Thus the rate ofADP dissociation from AM.ADP is expected to depend on the mechanical stress orstrain in the cross-bridge. At high shortening velocities, ADP dissociation shouldoccur more rapidly than observed in our (isometric) photolysis experiments. In
674

MgADP AND CROSS-BRIDGE KINETICS
agreement with this hypothesis, biochemical experiments without mechanical strainindicate rapid dissociation (> 500 s-') of ADP from most AM.ADP sites (White,1977; Geeves, 1989). Siemankowski, Wiseman & White (1985) reported that the rateof ADP release from AM.ADP in vitro, using myosin from a variety of muscle types,correlates with the maximum shortening velocity over a wide range of muscle speeds.We observed strain dependence of relaxation in the absence of Ca2+ from the
AM.ADP state suggesting faster ADP dissociation from negative strains than frompositive strains. A kinetic model incorporating the same strain dependence of ADPdissociation explains many features of the data obtained in the presence of Ca2+.These observations provide support for the hypothesis that ADP dissociation (or anisomerization of AM'.ADP) contributes to the control of cross-bridge cycling rate.
APPENDIX A
We present the rate constants and tension assignments for states in the simulationof Fig. 18A-D (negative strain model). This model considers the effects of summingforces from cross-bridges with positive and negative strain and it incorporates straindependence of the ADP dissociation rate from AM.ADP. The reaction sequence isScheme III in the Discussion section. The reaction scheme was solved separately forpositive and negative strain conditions and then the forces from the two runs wereadded algebraically. The following rate constants were assigned to Scheme III:k= 118 s-1,klD = 25 s-5 for force in AM.ADP > 0,kD = 250 s-5 for force in AM.ADP < 0,k+D= 5x 105 M-1 S-1k+T = 2 x 105M 51S1,k3 = 100 S,k = 150 s-1,k_4=1P5x 104 M-1 S-1.Relative to the force during steady active isometric contraction (i.e. that of
AM'.ADP), the rigor tension values assigned to positively strained cross-bridges are1-5 for AM and 1-25 for AM.ADP (isometric) and 1-75 for AM and 1-5 for AM.ADPfollowing a rigor pre-stretch. The corresponding tension assignments for thenegatively strained population are -0-25 for AM and -0 5 for AM.ADP (isometricrigor) and 00 for AM and -0-25 for AM.ADP (pre-gtretch). The {AI.ATTP} andAAMD PPi states are assigned zero tension. These values give isometric rigor tension{M.ADP.Pi }at 200 /LM-MgADP as 80% of that in the absence of ADP.The differential equations for Scheme III were integrated using a fourth-order
Runge-Kutta algorithm implemented by a FORTRAN program on a Z80microprocessor. The initial conditions were set to the equilibrium distributionbetween AM and AM.ADP appropriate to the MgADP concentration and MgADPaffinity for AM. At 200 /,M-MgADP 80% of the positively strained cross-bridges butonly 29% of negatively strained cross-bridges bound ADP. Tension from equalpopulations of cross-bridges initially strained positively and negatively werecalculated separately and then added to produce the net fibre tension. Although thenumbers in the two populations of cross-bridges were assumed to be equal, rigortension was positive because the positively strained population was given a higher
22-2
675

J. A. DANTZIG AND OTHERS
absolute value of force. The traces in Fig. 18A-D were all calculated with 1 mm-photo-released ATP using the set of rate constants and tension assignments givenabove, but at various combinations of MgADP and Pi concentrations shown on eachpanel. The simulations are comparable to the data of Fig. 15.
APPENDIX B
The reaction scheme, differential equations and rate constants used for thesimulation of Fig. 18E-H (co-operative model) are presented. This kinetic schemeconsiders pairs of cross-bridge sites that can interact co-operatively. Tension andrate constants referring to one site of a pair depend on the state of that site's partner.The model considered for a single site is shown in the Discussion as Scheme IV. Thefollowing rate constants of Scheme IV were assigned as standard conditions:kA= 118s-1,k_D = 500 51,k+D= 2X lO6 M-1s-1k+T= x 10 5M-1-1k4= 50 s-.If active force corresponds to 10 tension unit per site then the tension values for
the four intermediates are: TA = 1P0, TR = 05, TD = 00 and TL = 0 425 (0-85 x TR)under isometric rigor conditions and TA = 1P0, TR = 0 75, TD = 0 0 and TL = 0 675 fortransients following rigor pre-stretch.Combining the four intermediates into pairs gives Scheme V:
Caged ATP - ATPADP ATP'2 < 0 14
LL -' LD- LA6 78 9
ADP 3ATPADP ADP 5RR>ATPA'RD-W 5R (Scheme V)
ATP ATP
12 1316
DD - DA
17
AAwhere, for example, RA corresponds to a pair of sites, one in the R (rigor) state andthe other in A (actively generating force), and where numbers refer to reaction rateconstants. For instance k13 is the rate constant for RA + ATP -. DA. Including cagedATP, MgATP and MgADP, there are thirteen kinetic species and seventeenelementary reactions, leading to differential equations of the following form:
d caged ATP k,[caged ATP],dt =-jae T]
676

MgADP AND CROSS-BRIDGE KINETICS
dATP=- k[caged ATP] -klo[LR][ATP]dt
-kl [RR][ATP] - k2[RD][ATP] - k13[RA][ATP],dLRdt = k2[LL] - k3[LR] -k6[LR][ADP] + k7[RR][ADP] - kj0[LR][ATP],
and similarly for ADP and the other paired cross-bridge intermediates.The rate constants in Scheme V were set to: k= 118 s-1; k2 = 1000 S-1; k k4,
k5 =5001; k6, k8, k9= 2 x 106 M- S1; k7= 4 x 106 M1 S-1; k1, k12,
TABLE 3. Tension assignments for simulations of Fig. 18E-GTension per reaction
intermediateStatenumber State Isometric Pre-stretch
1 LL 0 850 1-3502 LR 0-925 1-4253* LD 0-425 0-6754 LA 1-425 1-6755 RR 1.000 1P5006 RD 0 500 0 7507 RA 1-500 1-7508 DD 01000 0)0009 DA 1000 100010 AA 21000 2-000
* In the co-operative model of Fig. 18G, the tension of intermediate LD was set to 0-925 for theisometric and 1-425 for the pre-stretch simulations.
k13 = 106 M-1 S-1; k1l = 2x 106 M-1 S-1; k14, k15 k17= 50 s-1; k16 = 100 s-1. Note thatstates with matching pairs of sites (e.g. RR) have twice the probability of undergoinga specific reaction (e.g. binding MgATP) than unmatched pairs, so their rateconstants (k2, k7, ki, and kl,) are doubled.The tension assignments for each of the ten intermediates in the two-site model are
given in Table 3, which is generated by adding the tension assignments of each siteof the pair (e.g. for isometric simulations, TLA = TL (0 425) + TA (10) force units persite). The initial condition for each simulation was taken to be a distribution betweenLL, LR and RR appropriate to the equilibrium binding of MgADP using theassociation and dissociation rate constants k2, k3, k6and k7. The differential equationswere integrated using a fourth-order Runge-Kutta algorithm implemented by aFORTRAN program on a Z80 microprocessor. Traces in panels E and F of Fig. 18were calculated using the rate constants listed above and tension values of Table 3at 500 /M-photo-released ATP and 0 and 400 /LM steady MgADP concentrationsrespectively. To simulate maintenance of rigor tension by one D site when its partnersite is in the L state, TLD was set equal to TLR (0 925) for panel 0. To simulateincreased rate of tension development by a D state when its partner is in the L state,ki4 (LD -+ LA) was increased from the standard value of 50 s-1 to 150 s-'. In this caseTLD was at its standard value of 0-425 and the resulting simulated traces are shownin Fig. 18H.
677

J. A. DANTZIG AND OTHERS
We would like to thank Mr Mark A. Luttmann and Mr Douglas P. Vallette for technicalassistance, Mr Marcus G. Bell for skilful electronic design and computer programming and DrsMichael A. Geeves, Earl Homsher, Vincenzo Lombari and Neil A. Millar for helpful comments onan earlier version of the paper. This work was supported by NIH grants HL15835 to thePennsylvania Muscle Institute and AM00745 and by the Muscular Dystrophy Association. M. G. H.held a British-American Research Fellowship of the British Heart Foundation and the AmericanHeart Association.
REFERENCES
ABBOTT, B. C. (1951). The heat production associated with the maintenance of a prolongedcontraction and the extra heat produced during large shortening. Journal of Physiology 112,438-445.
BERNAScoNI, C. F. (1976). Relaxation Kinetics, chap. 3. Academic Press, New York.BIosCA, J. A., GREENE, L. E. & EISENBERG, E. (1986). Binding of ADP and ATP analogs to cross-
linked and non-cross-linked acto-S-1. Journal of Biological Chemistry 261, 9793-9800.BIOSCA, J. A., GREENE, L. E. & EISENBERG, E. (1988). Binding of ADP and 5'-adenylyl
imidodiphosphate to rabbit muscle myofibrils. Journal ofBiological Chemistry 263, 14231-14235.BOREJDO, J., ANDO, T. & BURGHARDT, T. P. (1985). The rate ofMgADP binding to and dissociationfrom acto-Si. Biochimica et Biophysica Acta 828, 172-176.
CANNELL, M. B. (1986). Effect of tetanus duration on the free calcium during the relaxation of frogskeletal muscle fibres. Journal of Physiology 376, 203-218.
COOKE, R. (1986). The mechanism of muscle contraction. CRC Critical Reviews in Biochemistry 21,53-118.
COOKE, R. & PATE, E. (1985). The effects of ADP and phosphate on the contraction of musclefibers. Biophysical Journal 48, 789-798.
DANTZIG, J. A. & GOLDMAN, Y. E. (1989). Photolysis of caged ATP in muscle fibers with negativelystrained cross-bridges indicates that relaxation in the presence of MgADP is highly straindependent. Biophysical Journal 55, 260a.
DANTZIG, J. A., GOLDMAN, Y. E., LUTTMANN, M. L., TRENTHAM, D. R. & WOODWARD, S. K. A.(1989). Binding of caged ATP diastereoisomers to rigor cross-bridges in glycerol-extracted fibresof rabbit psoas muscle. Journal of Physiology 418, 61P.
DANTZIG, J. A., HIBBERD, M. G., GOLDMAN, Y. E. & TRENTHAM, D. R. (1984). ADP slows cross-bridge detachment rate induced by photolysis of caged ATP in rabbit psoas muscle fibers.Biophysical Journal 45, 8a.
DANTZIG, J. A., TRENTHAM, D. R. & GOLDMAN, Y. E. (1986). Kinetics of activation of skeletalmuscle fibers by photolysis of caged ATP in the presence of MgADP. Biophysical Journal 49,268 a.
FAJER, P. G., FAJER, E. A., MATTA, J. J. & THOMAS, D. D. (1989). Myosin head orientation in thepresence of ADP. Biophysical Journal 55, 261 a.
FENN, W. 0. (1923). A quantitative comparison between the energy liberated and the workperformed by the isolated sartorius muscle of the frog. Journal of Physiology 58, 175-203.
FERENCZI, M. A. (1986). Phosphate burst in permeable muscle fibers of the rabbit. BiophysicalJournal 50, 471-477.
FLETCHER, R. & POWELL, M. J. D. (1963). A rapidly convergent descent method of minimization.Computer Journal 6, 163-168.
FORD, L. E., HUXLEY, A. F. & SIMMONS, R. M. (1977). Tension responses to sudden length changein stimulated frog muscle fibres near slack length. Journal of Physiology 269, 441-515.
GEEVES, M. A. (1989). Dynamic interaction between actin and myosin subfragment 1 in thepresence of ADP. Biochemistry 28, 5864-5871.
GILLIS, J. M. (1985). Relaxation of vertebrate skeletal muscle. A synthesis of the biochemical andphysiological approaches. Biochimica et Biophysica Acta 811, 97-145.
GOLDMAN, Y. E. (1987 a). Measurement of sarcomere shortening in skinned fibers from frog muscleby white light diffraction. Biophysical Journal 52, 57-68.
GOLDMAN, Y. E. (1987 b). Kinetics of the actomyosin ATPase in muscle fibers. Annual Reviews ofPhysiology 49, 637-654.
678

MgADP AND CROSS-BRIDGE KINETICS
GOLDMAN, Y. E., HIBBERD, M. G., MCCRAY, J. A. & TRENTHAM, D. R. (1982). Relaxation of musclefibres by photolysis of caged ATP. Nature 300, 701-705.
GOLDMAN, Y. E., HIBBERD, M. G. & TRENTHAM, D. R. (1984a). Relaxation of rabbit psoas musclefibres from rigor by photochemical generation of adenosine-5'-triphosphate. Journal ofPhysiology354, 577-604.
GOLDMAN, Y. E., HIBBERD, M. G. & TRENTHAM, D. R. (1984b). Initiation of active contraction byphotogeneration of adenosine-5'-triphosphate in rabbit psoas muscle fibres. Journal ofPhysiology354, 605-624.
GOLDMAN, Y. E., MCCRAY, J. A. & VALLETTE, D. P. (1988). Cross-bridges in rigor fibres of rabbitpsoas muscle support negative forces. Journal of Physiology 398, 72P.
GOLDMAN, Y. E. & SIMMONS, R. M. (1977). Active and rigor muscle stiffness. Journal of Physiology269, 55-57P.
GOLDMAN, Y. E. & SIMMoNS, R. M. (1984). Control of sarcomere length in skinned muscle fibres ofRana temporaria during mechanical transients. Journal of Physiology 350, 497-518.
GREENE, L. E. & EISENBERG, E. (1980). Dissociation of the actin'subfragment 1 complex byadenyl-5'-yl imidodiphosphate, ADP, and PP1. Journal of Biological Chemistry 255, 543-548.
HASELGROVE, J. C. & HUXLEY, H. E. (1973). X-ray evidence for radial cross-bridge movement andfor the sliding filament model in actively contracting skeletal muscle. Journal of MolecularBiology 77, 549-568.
HIBBERD, M. G., DANTZIG, J. A., TRENTHAM, D. R. & GOLDMAN, Y. E. (1985). Phosphate releaseand force generation in skeletal muscle fibers. Science 228, 1317-1319.
HIBBERD, M. G. & TRENTHAM, D. R. (1986). Relationship between chemical and mechanical eventsduring muscular contraction. Annual Reviews of Biophysics and Biophysical Chemistry 15,119-161.
HIGUCHI, H. & TAKEMORI, S. (1989). Butanedione monoxime suppresses contraction and ATPaseactivity of rabbit skeletal muscle. Journal of Biochemistry 105, 638-643.
HILL, A. V. (1964). The effect of load on the heat of shortening of muscle. Proceedings of the RoyalSociety B 159, 297-318.
HORIUTI, K., HIGUCHI, H., UMAZUME, Y., KONISHI, M., OKAZAKI, 0. & KURIHARA, S. (1988).Mechanism of action of 2,3-butanedione 2-monoxime on contraction of frog skeletal musclefibres. Journal of MUscle Research and Cell Motility 9, 156-164.
HUXLEY, A. F. (1957). Muscle structure and theories of contraction. Progress in Biophysics andBiophysical Chemistry 7, 255-318.
HUXLEY, A. F. (1974). Review lecture. Muscular contraction. Journal of Physiology 243, 1-43.HUXLEY, A. F. & SIMMONS, R. M. (1971). Proposed mechanism of force generation in striated
muscle. Nature 233, 533-538.HUXLEY, H. E. (1969). The mechanism of muscular contraction. Science 164, 1356-1366.HUXLEY, H. E. & KRESS, M. (1985). Crossbridge behaviour during muscle contraction. Journal ofMuscle Research and Cell Motility 6, 153-161.
KAPLAN, J. H., FORBUSH, B. III & HOFFMAN, J. F. (1978). Rapid photolytic release of adenosine5'-triphosphate from a protected analogue: utilization by the Na: K pump of human red bloodcell ghosts. Biochemistry 17, 1929-1935.
KAWAI, M. (1986). The role of orthophosphate in crossbridge kinetics in chemically skinned rabbitpsoas fibres as detected with sinusoidal and step length alterations. Journal of Muscle Researchand Cell Motility 7, 421-434.
KAWAI, M. & BRANDT, P. W. (1980). Sinusoidal analysis: a high resolution method for correlatingbiochemical reactions with physiological processes in activated skeletal muscles of rabbit, frogand crayfish. Journal of Muscle Research and Cell Motility 1, 279-303.
KAWAI, M. & HALVORSON, H. R. (1989). Role of MgATP and MgADP in the cross-bridge kineticsin chemically skinned rabbit psoas fibers. Study of a fast exponential process. BiophysicalJournal 55, 595-603.
LENART, T. D., TANNER, J. W. & GOLDMAN, Y. E. (1989). 2,3-Butanedione monoxime (BDM)suppresses cross-bridge reattachment following laser photolysis of caged ATP. BiophysicalJournal 55, 260a.
MARSTON, S. (1973). The nucleotide complexes of myosin in glycerol-extracted muscle fibres.Biochimica et Biophysica Acta 305, 397-412.
679

J. A. DANTZIG AND OTHERS
MARSTON, S. B., TREGEAR, R. T., RODGER, C. D. & CLARKE, M. L. (1979). Coupling between theenzymatic site of myosin and the mechanical output of muscle. Journal ofMolecular Biology 128,111-126.
MATSUBARA, I., YAGI, N. & HASHIZUME, H. (1975). Use of an X-ray television for diffraction of thefrog striated muscle. Nature 255, 728-729.
OBIORAH, 0. & IRVING, M. (1989). ADP binding to rigor crossbridges produces no major changein their conformation. Biophysical Journal 55, 9a.
PECKHAM, M. & WOLEDGE, R. C. (1986). Labile heat and changes in rate of relaxation of frogmuscles. Journal of Physiology 374, 123-135.
RALL, J. A. (1989). Relationship of isometric unexplained energy production to parvalbumincontent in frog skeletal muscle. In Muscle Energetics, ed. PAUL, R., pp. 117-126. Alan R. Liss,Inc., New York.
ROSENFELD, S. S. & TAYLOR, E. W. (1987a). The mechanism of regulation of actomyosinsubfragment 1 ATPase. Journal of Biological Chemistry 262, 9984-9993.
ROSENFELD, S. S. & TAYLOR, E. W. (1987 b). The dissociation of 1-N6-ethenoadenosine diphosphatefrom regulated actomyosin subfragment 1. Journal of Biological Chemistry 262, 9994-9999.
SCHOENBERG, M. & EISENBERG, E. (1987). ADP binding to myosin cross-bridges and its effects onthe cross-bridge detachment rate constants. Journal of General Physiology 89, 905-920.
SIEMANKOWSKI, R. F., WISEMAN, M. 0. & WHITE, H. D. (1985). ADP dissociation from actomyosinsubfragment 1 is sufficiently slow to limit the unloaded shortening velocity in vertebrate muscle.Proceedings of the National Academy of Sciences of the USA 82, 658-662.
TAYLOR, E. W. (1979). Mechanism of actomyosin ATPase and the problem of muscle contraction.CRC Critical Reviews in Biochemistry 6, 103-164.
THOMAS, D. D., SVENSSON, E. C. & POLNASZEK, C. F. (1985). ADP does not induce rigid axialrotation of myosin heads in rigor muscle fibers. Biophysical Journal 47, 380a.
WALKER, J. W., REID, G. P., MCCRAY, J. A. & TRENTHAM, D. R. (1988). Photolabile 1-(2-nitrophenyl)ethyl phosphate esters of adenine nucleotide analogues. Synthesis and mechanismof photolysis. Journal of the American Chemical Society 110, 7170-7177.
WEBB, M. R., HIBBERD, M. G., GOLDMAN, Y. E. & TRENTHAM, D. R. (1986). Oxygen exchangebetween P, in the medium and water during ATP hydrolysis mediated by skinned fibers fromrabbit skeletal muscle. Journal of Biological Chemistry 261, 15557-15564.
WHITE, H. D. (1977). Magnesium-ADP binding to acto-myosin-Si and acto-heavymeromyosin.Biophysical Journal 17, 40a.
680
Copyright © 2022 FDOKUMEN




















