
Perspectives in Plant Ecology, Evolution and Systematics
10
Contents lists available at ScienceDirect Perspectives in Plant Ecology, Evolution and Systematics journal homepage: www.elsevier.com/locate/ppees Shrubs mediate forest start-up and patch dynamics in a semiarid landscape P. Macek a,b, ⁎ , C. Schöb c , F.I. Pugnaire b , M. Núñez-Ávila d,e , J.J. Armesto d,e a Faculty of Science, University of South Bohemia, Branišovská 1760, CZ-37005, České Budějovice, Czech Republic b LINCGlobal, Estación Experimental de Zonas Áridas, Consejo Superior de Investigaciones Científicas, Ctra. Sacramento s/n, 04120, La Cañada, Almería, Spain c Department of Environmental Systems Science, Swiss Federal Institute of Technology, ETH Zurich, Tannenstrasse 1, CH-8092, Zürich, Switzerland d LINCGlobal, Departamento de Ecología, Facultad Ciencias Biológicas, Pontificia Universidad Católica de Chile, Casilla 114-D, Santiago, Chile e Institute of Ecology and Biodiversity, Santiago, Chile ARTICLE INFO Keywords: Community dynamics Facilitation-competition balance Fog interception Plant-plant interactions Semiarid environment Spatial pattern ABSTRACT Plant spatial patterns in semiarid environments are often driven by a patchy availability of resources. We ex- amined here drivers of forest patterns in a landscape where tree and shrub patches coexist in an herbaceous matrix on fog-inundated coastal hilltops in semiarid Chile, aiming to identify factors that set off forest patch formation and development. We hypothesized that a successional process driven by plant-plant interactions would start when tree seedlings establish in shrub patches, eventually leading to resource accumulation and increased diversity. Within shrub patches, soil moisture increased with increasing cover of a common shrub, Baccharis vernalis, unlike another shrub species, Griselinia scandens. Soil nutrients increased with patch size, but soil moisture remained similar among patches. Forest patches showed differences regarding soil moisture be- tween wet windward and dry leeward edges, and these were associated with differences in community com- position. While Baccharis patches were similar to windward forest edges, leeward edges were more similar to open areas. Patterns of species richness and diversity showed a shift in plant-plant interactions from positive to negative along successional transition from Baccharis to forest patches. Outside forest patches, tree species are rare but become established and survive under Baccharis vernalis, most likely thanks to a combination of fog- interception capacity, soil nutrient availability and low competition. Therefore, forest patch formation is strongly dependent on fog water interception and community composition. While some shrubs like Baccharis vernalis facilitate tree establishment and eventually initiate new forest patches, others compete with tree seed- lings preventing establishment. Hence, a facilitation-competition balance between shrubs and trees explains the spatial arrangement of the forest-shrub patch mosaic in this semiarid environment. 1. Introduction Ecologists have long been interested in the mechanisms that explain spatial patterns of vegetation (Watt, 1947). Understanding the different processes leading to large-scale vegetation patterns continues to be a critical and challenging issue in plant ecology (Fortin and Dale, 2005; Getzin et al., 2016). Some drivers of large-scale vegetation patterns are well known, such as differences in topography, substrate, soil resources, or disturbances (Aguiar and Sala, 1999; Greig-Smith, 1979; Rietkerk and van de Koppel, 2008; Wetzel, 2002). In particular, spatial hetero- geneity in limiting resources creates locally species filters and therefore differentially assembled communities leading to non-random vegeta- tion patterns (Macek et al., 2009a; Seabloom et al., 2005; Tirado and Pugnaire, 2005). Because water is essential for plants, its directional supply may result in non-random community structure and pronounced changes in species composition (Stanton et al., 2013). For example, water flowing downslope may lead to linear vegetation patterns (Borthagaray et al., 2010; White, 1971). Water provided by fog is important for plants able to use such water source (Karbulková et al., 2008; Macková et al., 2013; Martorell and Ezcurra, 2007; Rundel and Dillon, 1998). Fog intercep- tion by leaves can lead to a significant increase in soil moisture beneath fog-exposed crowns or branches (Cereceda et al., 2008; Shmida et al., 1986; Uclés et al., 2014), sometimes supplying extra water to nearby plants (Macek et al., 2018). Indeed, whole ecosystems may, to a large extent, be dependent on fog interception (Dawson, 1998; Huntley et al., 1997; Stanton et al., 2013). Fog is especially important in coastal arid regions (Ewing et al., 2009; Garreaud et al., 2010; Gutiérrez et al. 1998; Johnstone and Dawson, 2010; Rundel and Dillon, 1998; Uclés et al., 2014), arising when cold oceanic currents interact with warm air https://doi.org/10.1016/j.ppees.2018.09.002 Received 14 June 2017; Received in revised form 31 August 2018; Accepted 17 September 2018 ⁎ Corresponding author at: Faculty of Science, University of South Bohemia, Na Zlaté stoce 3, CZ-37005, České Budějovice, Czech Republic. E-mail address: [email protected] (P. Macek). Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149 Available online 25 September 2018 1433-8319/ © 2018 Elsevier GmbH. All rights reserved. T
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Perspectives in Plant Ecology, Evolution and Systematics

Contents lists available at ScienceDirect
Perspectives in Plant Ecology, Evolution and Systematics
journal homepage: www.elsevier.com/locate/ppees
Shrubs mediate forest start-up and patch dynamics in a semiarid landscapeP. Maceka,b,⁎, C. Schöbc, F.I. Pugnaireb, M. Núñez-Ávilad,e, J.J. Armestod,ea Faculty of Science, University of South Bohemia, Branišovská 1760, CZ-37005, České Budějovice, Czech Republicb LINCGlobal, Estación Experimental de Zonas Áridas, Consejo Superior de Investigaciones Científicas, Ctra. Sacramento s/n, 04120, La Cañada, Almería, SpaincDepartment of Environmental Systems Science, Swiss Federal Institute of Technology, ETH Zurich, Tannenstrasse 1, CH-8092, Zürich, Switzerlandd LINCGlobal, Departamento de Ecología, Facultad Ciencias Biológicas, Pontificia Universidad Católica de Chile, Casilla 114-D, Santiago, Chilee Institute of Ecology and Biodiversity, Santiago, Chile
A R T I C L E I N F O
Keywords:Community dynamicsFacilitation-competition balanceFog interceptionPlant-plant interactionsSemiarid environmentSpatial pattern
A B S T R A C T
Plant spatial patterns in semiarid environments are often driven by a patchy availability of resources. We ex-amined here drivers of forest patterns in a landscape where tree and shrub patches coexist in an herbaceousmatrix on fog-inundated coastal hilltops in semiarid Chile, aiming to identify factors that set off forest patchformation and development. We hypothesized that a successional process driven by plant-plant interactionswould start when tree seedlings establish in shrub patches, eventually leading to resource accumulation andincreased diversity. Within shrub patches, soil moisture increased with increasing cover of a common shrub,Baccharis vernalis, unlike another shrub species, Griselinia scandens. Soil nutrients increased with patch size, butsoil moisture remained similar among patches. Forest patches showed differences regarding soil moisture be-tween wet windward and dry leeward edges, and these were associated with differences in community com-position. While Baccharis patches were similar to windward forest edges, leeward edges were more similar toopen areas. Patterns of species richness and diversity showed a shift in plant-plant interactions from positive tonegative along successional transition from Baccharis to forest patches. Outside forest patches, tree species arerare but become established and survive under Baccharis vernalis, most likely thanks to a combination of fog-interception capacity, soil nutrient availability and low competition. Therefore, forest patch formation isstrongly dependent on fog water interception and community composition. While some shrubs like Baccharisvernalis facilitate tree establishment and eventually initiate new forest patches, others compete with tree seed-lings preventing establishment. Hence, a facilitation-competition balance between shrubs and trees explains thespatial arrangement of the forest-shrub patch mosaic in this semiarid environment.
1. Introduction
Ecologists have long been interested in the mechanisms that explainspatial patterns of vegetation (Watt, 1947). Understanding the differentprocesses leading to large-scale vegetation patterns continues to be acritical and challenging issue in plant ecology (Fortin and Dale, 2005;Getzin et al., 2016). Some drivers of large-scale vegetation patterns arewell known, such as differences in topography, substrate, soil resources,or disturbances (Aguiar and Sala, 1999; Greig-Smith, 1979; Rietkerkand van de Koppel, 2008; Wetzel, 2002). In particular, spatial hetero-geneity in limiting resources creates locally species filters and thereforedifferentially assembled communities leading to non-random vegeta-tion patterns (Macek et al., 2009a; Seabloom et al., 2005; Tirado andPugnaire, 2005).
Because water is essential for plants, its directional supply may
result in non-random community structure and pronounced changes inspecies composition (Stanton et al., 2013). For example, water flowingdownslope may lead to linear vegetation patterns (Borthagaray et al.,2010; White, 1971). Water provided by fog is important for plants ableto use such water source (Karbulková et al., 2008; Macková et al., 2013;Martorell and Ezcurra, 2007; Rundel and Dillon, 1998). Fog intercep-tion by leaves can lead to a significant increase in soil moisture beneathfog-exposed crowns or branches (Cereceda et al., 2008; Shmida et al.,1986; Uclés et al., 2014), sometimes supplying extra water to nearbyplants (Macek et al., 2018). Indeed, whole ecosystems may, to a largeextent, be dependent on fog interception (Dawson, 1998; Huntley et al.,1997; Stanton et al., 2013). Fog is especially important in coastal aridregions (Ewing et al., 2009; Garreaud et al., 2010; Gutiérrez et al. 1998;Johnstone and Dawson, 2010; Rundel and Dillon, 1998; Uclés et al.,2014), arising when cold oceanic currents interact with warm air
https://doi.org/10.1016/j.ppees.2018.09.002Received 14 June 2017; Received in revised form 31 August 2018; Accepted 17 September 2018
⁎ Corresponding author at: Faculty of Science, University of South Bohemia, Na Zlaté stoce 3, CZ-37005, České Budějovice, Czech Republic.E-mail address: [email protected] (P. Macek).
Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
Available online 25 September 20181433-8319/ © 2018 Elsevier GmbH. All rights reserved.
T

inland. Due to temperature inversion, air becomes moistened from thecold ocean by advection, eventually forming a thick mantle of stratusclouds which are pushed by wind and then expand as fog over the landsurface (Cereceda et al., 2002; Garreaud et al., 2008).
The role of plant-plant interactions in forest patch formation hasreceived little attention in fog-dependent systems. Plant-plant interac-tions are acknowledged as important forces shaping spatial patterns(Fibich et al., 2013; Haase et al., 1996; Schöb et al., 2008; Tirado andPugnaire, 2005; White, 1971). In arid regions, the positive effect ofsome shrub species on the herbaceous understory has often been linkedto amelioration of soil conditions (Moro et al., 1997; Maestre et al.,2003; Prieto et al., 2011; Pugnaire et al., 2004, 2011). Consistent withthese and other studies, van Zonneveld et al. (2012) suggested thatshrubs could have a facilitative effect on woody seedlings of forestspecies. Shrub-driven amelioration of soil conditions, including in-creased soil moisture through fog interception, can provide a relevantmechanism to stimulate tree regeneration, a process that could even-tually give rise to new forest patches in dry environments.
The forest-shrubland mosaic on coastal hilltops of Fray Jorge ForestNational Park (hereafter Fray Jorge), in semiarid north-central Chile,provides an outstanding model to address the importance of both bioticand abiotic processes in spatial pattern dynamics. Fog is continuouslyarriving from the Pacific Ocean (Garreaud et al., 2008) and forest patchpersistence relies largely on fog interception by wind-exposed tree ca-nopies (del-Val et al., 2006). Located at a subtropical latitude (30 °S)Fray Jorge hosts remnants of coastal temperate rain forests which havebeen gradually fragmented due to progressive aridization during theQuaternary (Núñez-Ávila and Armesto, 2006). Nowadays, forest pat-ches are scattered like islands within a sparse shrubland matrix,forming the northernmost Chilean lowland forest. These forest patchesare slowly growing on the windward edge and decaying sharply at theleeward edge due to declining water interception by the canopy (del-Val et al., 2006). However, the presence of variable patch sizes, rangingfrom a few square meters and a single generally young tree to patches ofover 1 ha with numerous trees (Núñez-Ávila et al., 2013), suggests thatthe forest patch mosaic is dynamic and new patches may occasionallybe formed in the predominant shrubland matrix.
Here we analyze abiotic and biotic factors driving forest patch in-itiation in the forest-shrubland mosaic of Fray Jorge. Specifically, weaimed at identifying the potential drivers of tree establishment near andaway forest edges, with a particular focus on plant-plant interactionsand fog water subsidies (Table 1). Our main hypothesis was that onlythe presence of a facilitator species and directional fog-water inputsmay lead to the establishment of the main forest tree (Aextoxiconpunctatum), conditions which are fulfilled in 1) forest windward edges,and 2) solitary shrub patches scattered within the open areas. For thispurpose, we contrasted the processes occurring on windward versusleeward edges of established forest patches and also those occurring inshrub patches located away (12–52m) from forests. We seek to identifykey shrub species with a critical role in initiating forest patch formationand to assess the main mechanisms of community change, particularlyfocusing on plant-plant interactions.
Based on a previous study (Macek et al., 2018), we hypothesized
that 1) shrubs both at the windward edge and away from forest patcheswill promote a successional transition from a herbaceous vegetation toa forest patch, where isolated shrub patches will be similar in speciescomposition to windward edges of forests; 2) this gradient translatesinto varying environmental conditions and species diversity; we hy-pothesized that resources will accumulate along the gradient eventuallyleading to greater diversity at later stages; in detail, we hypothesizedthat the presence of certain shrub species would increase soil resourcelevels, and promote associated plant species diversity, including theestablishment of forest trees, and finally, 3) that species-specific plant-plant interactions in different habitats will lead to the presence of foresttree species in shrub patches, while leeward forest edges and open areaswill lack the conditions for forest tree growth. In a companion study weprovided strong experimental evidence of a contrasting survival of treespecies in the presence versus absence of shrubs (Macek et al., 2018),while this study places this facilitative effect of shrubs on tree seedlingsin a community level context to explain forest patch formation.
2. Material and methods
2.1. Study site
Fray Jorge Forest National Park, a World Biosphere Reserve at acoastal location in north-central Chile (30°38′S, 71°40′W), includessemiarid shrublands in inland areas and scattered forests on fog-in-undated coastal hilltops (500–600m). Annual rainfall averages 147mm(1983–2003), with 95% occurring in the austral winter months, June-August (López-Cortés and López, 2004). Fog moving inland from thePacific Ocean is a permanent feature, especially during spring andsummer months. Fog-loaded winds blow predominantly from thesouthwest all year round (Macek et al., 2018) bringing humidity tocoastal hills. Forests are dominated by the broadleaved evergreen treespecies Aextoxicon punctatum Ruiz et Pav. (Aextoxicaceae), with pre-sence of Myrceugenia correifolia Hook et Arn. (Myrtaceae) and Drimyswinteri J.R. Forst. et G. Forst. (Winteraceae) (Gutiérrez et al., 2008;Salgado-Negret et al., 2013), all of which extend their distributionssouthwards into the Mediterranean and temperate climate region ofsouthern South America receiving a higher rainfall (Salgado-Negretet al., 2013). At the landscape level, forest patches form a mosaic withshrublands and open (shrub free) areas (Fig. 1) with isolated, scatteredtiny shrub patches. Shrublands are often dominated by the two wide-spread species Baccharis vernalis F.H. Hellwig (Asteraceae), and Aris-teguietia salvia (Colla) R. M. King. et H. Rob. (Asteraceae); both speciescan tolerate dry conditions outside forests and thrive in open (non-forest) areas intermingled with other shrub species.
2.2. Solitary Baccharis patches
Shrub patches of varying size (ranging 0.9‒70.7m2; n= 50) weresampled around established forest patches. Shrub patch distance to thenearest forest fragment was measured. Shrub patches always containedBaccharis vernalis and are hence referred to as Baccharis patches (Fig.S1). Baccharis patches were sorted into three size groups, i.e. < 9m2,
Table 1Conceptual sampling design and predictions of presence (+) or absence (-) of seedlings or juveniles of forest tree species (Aextoxicon punctatum), according toenvironmental abiotic and biotic conditions.
Forest fragments Baccharis patches Open areas
Baccharis present Baccharis absent Baccharis present Baccharis absent
Fog present + – + –*
Fog absent – – n.a. –*
* Hypotheses already tested elsewhere (Macek et al., 2018), resulting in no survival of trees planted in open areas, regardless of the presence of water providingfog collectors.
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
141

10–18m2, and> 18 m2, referred to as ‘small’, ‘intermediate’ and ‘large’patches, respectively. We recorded all perennial plant species withinthe whole patch, and in an equivalent open area (without shrub cover)located in its close vicinity (edges of both plots less than 3m away).Three soil samples per each plot were collected for soil water content(SWC) and soil organic matter content (SOM) determinations. Lightconditions (PAR) were recorded at ground level using Li-1600 (LiCor,Nebraska, USA) during a cloudless day.
2.3. Shrubs at the edge of forest patches
We furthermore selected 21 rather small forest patches (range:0.04–0.45 ha; remaining of the 180 forest patches on the surroundinghills are between 0.2 and 22 ha (del-Val et al., 2006) interspersed withshrublands and open grassland on a flat area on coastal hilltops. Smallforest patch sizes are the most common in this flat environment. Pat-ches had an ellipsoid shape with longer sides facing south (windward)and north (leeward; Fig. 1). From the center of each forest patch, twotransects were run, one in the windward and the other in the leewarddirection. Each transect was divided into two 25m2 (5m x 5m) plotsstarting 3-m from the edge of the forest patch (defined here as the lastbranch of> 5m tall trees), thus reaching 13m outside the forest ca-nopy. These plots, at both windward and leeward edges, were thencalled forest patch edge plots, and were further identified based on theirdistance to the forest patch (i.e. ‘near’ and ‘far’). All perennial plantspecies were identified in each plot (Table 3).
Soil samples (three subsamples per plot) were collected at bothwindward and leeward forest sides (forest patch edge plots, near only)for SWC and SOM determinations. In addition, at the same sites we
collected fresh leaves (youngest mature leaves) of the nearestAextoxicon punctatum tree to estimate foliar nutrient (nitrogen, N, andcarbon, C) content and their isotopic composition (δ15N and δ13C).
2.4. Environmental measurements, nutrient analyses and isotopiccomposition
Soil samples (collected in February 2012 in forest sides, under shrubpatches and in open areas) devoid of roots were weighed, dried at105 °C, and weighed again to determine SWC. The soil samples werethen ground and analyzed for SOM using the carbon loss by ignitionmethod. Soils were then analyzed for total C, N, and P contents (CHNSElemental Analyzer vario MICRO cube; Elementar AnalysensystemeGmbH, Germany; and FIAstar; FOSS, Denmark). Leaves of Aextoxiconpunctatum were dried at 60 °C for 48 h, ground, and analyzed for totalN, C and isotopes (δ13C and δ15N) with a PDZ Europa ANCA-GSL ele-mental analyzer and mass spectrometer (Sercon Ltd., Cheshire, UK).The 13C/12C ratios (Rp) were referenced to 13C/12C ratios of standardVPDB (Vienna-Pee-Dee Belemnite; Rs) and calculated as δ13C = (Rp/Rs-1)*1000 in ‰. We expressed δ15N for 15N/14N ratios (referenced toatmospheric N2) in a similar way. The standard deviation of δ13C andδ15N determination in standard samples was lower than 0.1‰ and0.2‰, respectively. Isotope ratios of C and N were used to assess wateruse efficiency, and to identify the possible N-source for Aextoxiconpunctatum trees in patches, respectively. Less negative values of δ13Chave been assumed to indicate increased water use efficiency(Ehleringer et al., 1993), whereas values of δ15N have often been usedto assess the main N source (Robinson, 2001).
Fig. 1. The study site located within Fray Jorge Forest National Park. Left: Scattered forest fragments intermingled with shrublands, isolated Baccharis patches andshrub-free open areas. Rectangle indicates inset (Right) with transect plots around a forest patch (squares) and Baccharis patches (arrows).
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
142

2.5. Data analyses
To compare plant species composition among wind- and leewardforest patch edges, shrublands and open areas, we compared relativespecies cover using multivariate ordination methods. We first comparedvegetation of forest patch edges, solitary Baccharis patches, and openareas using a Principal Component Analysis (PCA). Then, we comparedspecies composition of Baccharis patches with windward-edge forestpatch edge plots by PCA. Finally, to test for differences in communitycomposition among plots in opposite forest edges and different dis-tances to forests, Redundancy Analysis (RDA, with standardized vari-ables) was used. RDA can be considered an extension of multivariatelinear regression for a multivariate response variable (Lepš andŠmilauer, 2003), with the parametric test replaced by Monte Carlopermutation test (499 permutations used). We used log-transformedcover data because of the skewed dataset (few species with large covervalues). Data from transects were further organized in a split plot de-sign, with edge orientation and distance to forest patch as categoricalfactors, and the main plot (forest fragment ID) had therefore four splitplots (a combination of the two factors each with two levels). All or-dinations were carried out using Canoco 4.5 and CanoDraw 4 (ter Braakand Šmilauer, 2002). The frequency of occurrence of Aextoxicon punc-tatum in different communities at different distances to forest patcheswas tested using deviance analysis (residual deviance test based on χ2
distribution).Due to unequal sample sizes for Baccharis patches (divided into
three size classes) and open areas (altogether), we used sample-basedrarefaction techniques to compare species richness among habitats(Gotelli and Colwell, 2001). For each Baccharis patch size class and foropen areas, 500 random samples with replacement were drawn (i.e.from one sample up to as many samples as we had for each Baccharispatch size class or open area). We then drew species accumulationcurves with means and 95% confidence intervals for the three sizeclasses, and for all open areas pooled. Rarefaction analyses were per-formed in R (R Core Team, 2012) using the specaccum function and the‘exact’ method of the vegan package (Oksanen et al., 2012).
For differences in soil and Aextoxicon punctatum leaf traits betweenleeward and windward edge of forest patches we used dependent t-tests. For differences in soil properties and Shannon diversity index, weused one-way ANOVA. Alternative models of linear and second-orderpolynomial regressions were fitted to the relationship between SWC,nutrient contents (and SOM) and dominant species cover in Baccharispatches; model selection followed the Akaike Information Criterion
(Sakamoto et al., 1986). Linear and logistic regressions were used torelate the presence of Aextoxicon punctatum or nutrient contents inshrub patches with the distance to the nearest forest patch.
3. Results
3.1. Species composition patterns
Plant species composition was clearly different among forest patchedges, solitary Baccharis patches, and open areas, the latter dominatedby Happlopappus hirtus and several graminoid and annual species(Fig. 2a). Open areas showed the largest dissimilarity to other habitats,represented by the first canonical axis (explaining 30.8% of variability).Solitary Baccharis patches were similar to the shrubby vegetation in thewindward edge of forest patches, sharing species such as Baccharisvernalis itself and Senecio sinuatilobus, but dissimilar with the leewardforest edge; this was represented by the second axis (explaining addi-tional 21.9% of variability).
We examined in more detail differences among Baccharis patches ofthree size classes and compared them to the most similar vegetation ofwindward forest edges (Fig. 2b). The first axis, explaining 27.7% ofvariability, distinguished vegetation in windward forest patch edgesfrom Baccharis patches. The second PCA axis (17.4% of the variability)differentiated between small and large Baccharis patches. Median scoreson the second axis were: Windward edge of forest fragments: -0.25(far), 0.61 (near); Baccharis patches: -0.39 (small), -0.15 (intermediate),0.12 (large). Smaller Baccharis patches and plots located “far” fromforest patch edges resembled each other and were dominated by gra-minoids and small shrubs. Large solitary Baccharis patches and vege-tation closer to forest edges, on the other hand, were dominated by tallshrubs and tree species (e.g. Griselinia scandens, Baccharis vernalis,Myeceugenia correifolia, Aextoxicon punctatum). Species richness anddiversity depended also on the size class of patches. On the one hand,the smallest Baccharis patches were similar to open areas in speciesrichness, while the two larger patch classes were significantly richer(Fig. S2). On the other hand, Shannon’s diversity index was the highestin intermediate Baccharis patches and gradually decreased with in-creasing patch size towards forest patches (Fig. 3). Finally, we found anegative correlation between the cover of the most dominant tree offorest patches (Aextoxicon punctatum) in Baccharis patches and thedistance to the nearest forest patch (p=0.047), with probability offinding Aextoxicon punctatum decreasing below 50% when shrub patchwas further than 30.9m away from a forest patch.
Fig. 2. Results of PCA analyses showing (a) the vegetation composition of both edges of forest fragments (empty symbols), Baccharis patches (full symbols) and openareas (cross symbols) altogether, and (b) only the vegetation composition of the windward edge of forest fragments and Baccharis patches. Individual habitats wereenclosed into envelopes showing their similarity. First axes explained (a) 30.8% and (b) 27.7% of variability, both first and second axes explained (a) 52.7% and (b)45.1% of variability in species composition. For species acronyms see Table 3.
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
143
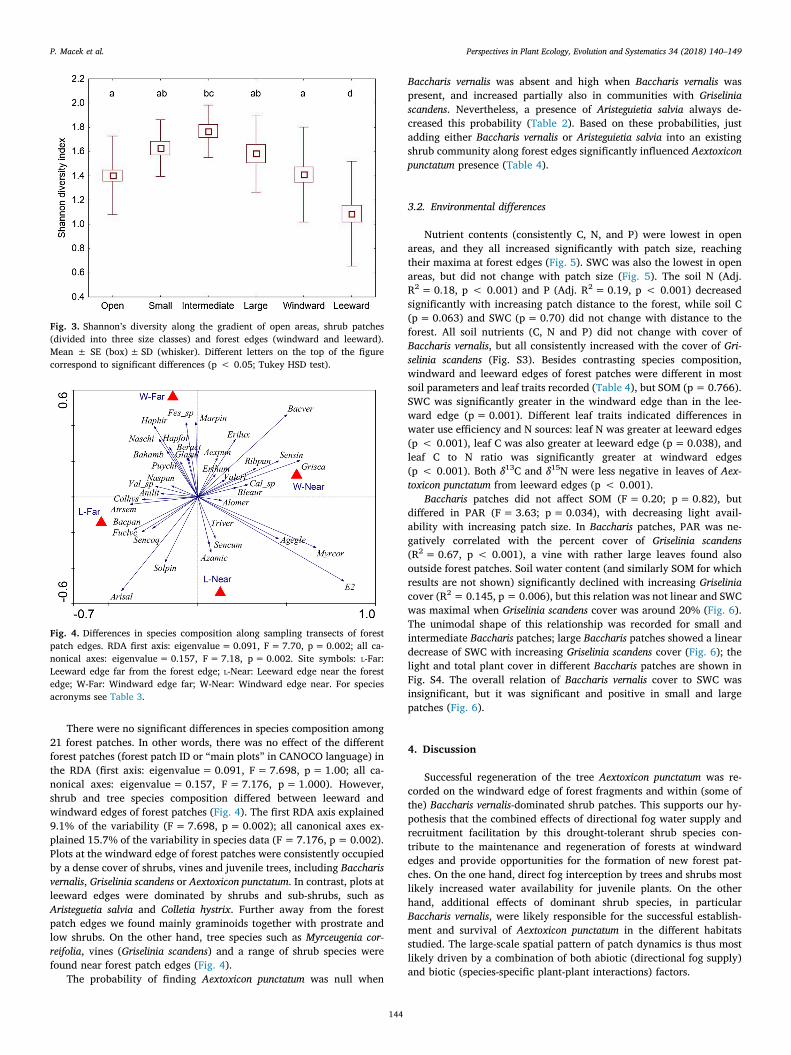
There were no significant differences in species composition among21 forest patches. In other words, there was no effect of the differentforest patches (forest patch ID or “main plots” in CANOCO language) inthe RDA (first axis: eigenvalue= 0.091, F= 7.698, p= 1.00; all ca-nonical axes: eigenvalue=0.157, F=7.176, p=1.000). However,shrub and tree species composition differed between leeward andwindward edges of forest patches (Fig. 4). The first RDA axis explained9.1% of the variability (F= 7.698, p= 0.002); all canonical axes ex-plained 15.7% of the variability in species data (F=7.176, p= 0.002).Plots at the windward edge of forest patches were consistently occupiedby a dense cover of shrubs, vines and juvenile trees, including Baccharisvernalis, Griselinia scandens or Aextoxicon punctatum. In contrast, plots atleeward edges were dominated by shrubs and sub-shrubs, such asAristeguetia salvia and Colletia hystrix. Further away from the forestpatch edges we found mainly graminoids together with prostrate andlow shrubs. On the other hand, tree species such as Myrceugenia cor-reifolia, vines (Griselinia scandens) and a range of shrub species werefound near forest patch edges (Fig. 4).
The probability of finding Aextoxicon punctatum was null when
Baccharis vernalis was absent and high when Baccharis vernalis waspresent, and increased partially also in communities with Griseliniascandens. Nevertheless, a presence of Aristeguietia salvia always de-creased this probability (Table 2). Based on these probabilities, justadding either Baccharis vernalis or Aristeguietia salvia into an existingshrub community along forest edges significantly influenced Aextoxiconpunctatum presence (Table 4).
3.2. Environmental differences
Nutrient contents (consistently C, N, and P) were lowest in openareas, and they all increased significantly with patch size, reachingtheir maxima at forest edges (Fig. 5). SWC was also the lowest in openareas, but did not change with patch size (Fig. 5). The soil N (Adj.R2= 0.18, p < 0.001) and P (Adj. R2= 0.19, p < 0.001) decreasedsignificantly with increasing patch distance to the forest, while soil C(p= 0.063) and SWC (p= 0.70) did not change with distance to theforest. All soil nutrients (C, N and P) did not change with cover ofBaccharis vernalis, but all consistently increased with the cover of Gri-selinia scandens (Fig. S3). Besides contrasting species composition,windward and leeward edges of forest patches were different in mostsoil parameters and leaf traits recorded (Table 4), but SOM (p=0.766).SWC was significantly greater in the windward edge than in the lee-ward edge (p= 0.001). Different leaf traits indicated differences inwater use efficiency and N sources: leaf N was greater at leeward edges(p < 0.001), leaf C was also greater at leeward edge (p=0.038), andleaf C to N ratio was significantly greater at windward edges(p < 0.001). Both δ13C and δ15N were less negative in leaves of Aex-toxicon punctatum from leeward edges (p < 0.001).
Baccharis patches did not affect SOM (F=0.20; p=0.82), butdiffered in PAR (F=3.63; p=0.034), with decreasing light avail-ability with increasing patch size. In Baccharis patches, PAR was ne-gatively correlated with the percent cover of Griselinia scandens(R2= 0.67, p < 0.001), a vine with rather large leaves found alsooutside forest patches. Soil water content (and similarly SOM for whichresults are not shown) significantly declined with increasing Griseliniacover (R2=0.145, p=0.006), but this relation was not linear and SWCwas maximal when Griselinia scandens cover was around 20% (Fig. 6).The unimodal shape of this relationship was recorded for small andintermediate Baccharis patches; large Baccharis patches showed a lineardecrease of SWC with increasing Griselinia scandens cover (Fig. 6); thelight and total plant cover in different Baccharis patches are shown inFig. S4. The overall relation of Baccharis vernalis cover to SWC wasinsignificant, but it was significant and positive in small and largepatches (Fig. 6).
4. Discussion
Successful regeneration of the tree Aextoxicon punctatum was re-corded on the windward edge of forest fragments and within (some ofthe) Baccharis vernalis-dominated shrub patches. This supports our hy-pothesis that the combined effects of directional fog water supply andrecruitment facilitation by this drought-tolerant shrub species con-tribute to the maintenance and regeneration of forests at windwardedges and provide opportunities for the formation of new forest pat-ches. On the one hand, direct fog interception by trees and shrubs mostlikely increased water availability for juvenile plants. On the otherhand, additional effects of dominant shrub species, in particularBaccharis vernalis, were likely responsible for the successful establish-ment and survival of Aextoxicon punctatum in the different habitatsstudied. The large-scale spatial pattern of patch dynamics is thus mostlikely driven by a combination of both abiotic (directional fog supply)and biotic (species-specific plant-plant interactions) factors.
Fig. 3. Shannon’s diversity along the gradient of open areas, shrub patches(divided into three size classes) and forest edges (windward and leeward).Mean ± SE (box)± SD (whisker). Different letters on the top of the figurecorrespond to significant differences (p < 0.05; Tukey HSD test).
Fig. 4. Differences in species composition along sampling transects of forestpatch edges. RDA first axis: eigenvalue= 0.091, F= 7.70, p= 0.002; all ca-nonical axes: eigenvalue= 0.157, F= 7.18, p= 0.002. Site symbols: L-Far:Leeward edge far from the forest edge; L-Near: Leeward edge near the forestedge; W-Far: Windward edge far; W-Near: Windward edge near. For speciesacronyms see Table 3.
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
144

4.1. Differences in plant communities
Species composition of solitary Baccharis patches was similar to thatof windward forest edges, but it clearly deviated from the shrubby as-semblages found in the leeward forest edges. Furthermore, the largerthe Baccharis patch the higher the likelihood of finding Aextoxiconpunctatum saplings and other species of windward forest edges. Whilespecies richness increased with patch size, species diversity decreased.
Hence, smaller Baccharis patches provided sites with diverse nichesthanks to an increase of the limiting resource and less negative plant-plant interactions. We assume that competition then gains importancewhen patches grow and together with species dominance it may haveresulted in a decrease of species diversity.
Within forest patches, tall trees create steep soil moisture gradients(cf. Salgado-Negret et al., 2013), which resulted in species differencesbetween leeward and windward edges, as evidenced by the abundanceof the two dominant shrub species; Aristeguietia salvia was dominant atthe leeward edge while Baccharis vernalis dominated the windwardedge. Environmental conditions led to higher diversity at windwardforest edges compared to leeward edges and together with the con-trasting community composition support the prevailing idea of forestpatch growth at windward side and patch recession at the leeward side.We hence suggest that windward edges of forest patches promote di-versity, in this case through canopy dripping, as they do in other sys-tems (Macek et al., 2009a; Troxler Gann et al., 2005). Asymmetry be-tween windward and leeward sides has been documented earlier withinforest parches (del-Val et al., 2006; Stanton, 2011; Stanton et al., 2013).Yet, our data expand this asymmetry beyond the forest fragment, to-wards its edges and deeply into the surrounding non-forest areas, wheretree regeneration may take place.
Within Baccharis patches, contrasting light and soil water contentpatterns related to species dominance: while soil was wetter at siteswhere Baccharis vernalis dominated, the cover of Griselinia scandens wasnegatively related to soil moisture. This may be related to the Griseliniascandens architecture, with large leaves (up to 10 cm) that should havehigh transpiration rates. Dense stands of Griselinia scandens may haveimposed light stress to tree seedlings, so that we can hypothesize thatGriselinia scandens is a strong competitor for both water and light,which can be a serious constraint for Aextoxicon punctatum establish-ment. In contrast, Baccharis vernalis has small (up to 2.5 cm) and fewerleaves, which do not cast deep shade. Interception of fog takes placefrequently at stems of Baccharis vernalis densely covered by epiphyticlichens and mosses (P. Macek & C. Schöb, personal observation). In
Table 2Effect of community composition (i.e. species present in model communities) of shrub patches on the occurrence of the tree Aextoxicon punctatum tested using adecrease of deviance test. The effect of added species (species whose new presences are tested compared to model communities composition) on Aextoxicon punctatumoccurrence is shown (in bold when significant; p < 0.05). Furthermore, probabilities of Aextoxicon punctatum occurrence in final communities are shown (samplesize in parenthesis). For species acronyms, see Table 3.
Community composition Added species n Deviance p (χ2) Effect Probability (n)
– Bacver 80 4.49 0.034 Positive 0.10 (52)Bacver Arisal 52 6.26 0.012 Negative 0.00 (23)Bacver Grisca 52 2.67 0.103 Positive 0.17 (24)Bacver Myrcor 52 0.08 0.771 Positive 0.11 (28)Bacver+Arisal Grisca 23 0 1 Positive 0.00 (4)Bacver+Arisal Myrcor 23 0 1 Negative 0.11 (12)Bacver+Arisal Ribpun 23 0 1 Negative 0.13 (15)Bacver+Arisal+Myrcor Grisca 12 0 1 Positive 0.00 (3)Bacver+Arisal+Myrcor Ribes 12 0 1 Negative 0.00 (9)Bacver+Grisca Myrcor 24 0.31 0.577 Negative 0.13 (15)
Table 3List of species full names and abbreviations used in the ordination diagrams.Nomenclature follows Tropicos.org.
Abbrev. Scientific name
Aexpun Aextoxicon punctatum (juv.) Ruiz & Pav.Agegle Ageratina glechonophylla (Less.) R.M. King & H. Rob.Alomer Alonsoa meridionalis (L. f.) KuntzeAnilit Anisomeria littoralis (Poepp. & Endl.) Moq.Annuals Annual species coverArisal Aristeguietia salvia (Colla) R.M. King & H. Rob.Atrsem Atriplex semibaccata R. Br.Azamic Azara microphylla Hook. f.Bacpan Baccharis paniculata DC.Bacver Baccharis vernalis HellwigBahamb Bahia ambrosioides Lag.Beract Berberis actinacantha Mart.Bleaur Blechnum auriculatum Cav.Cal_sp Calceolaria sp.Colhys Colletia hystrix ClosE2 Tree coverErilux Erigeron luxurians (Skottsb.) SolbrigErihum Eryngium humboldtii F. DelarocheFesmyu Festuca myuros L.Fuclyc Fuchsia lycioides AndrewsGal sp Galium sp.Glasul Glandularia sulphurea (D. Don) Schnack & CovasGnahet Gnaphalium heterotrichum Phil.Grisca Griselinia scandens (Ruiz & Pav.) Taub.Hapfol Haplopappus foliosus DC.Haphir Haplopappus hirtellus Phil.Jarplu Jarava plumosula (Nees ex Steud.) F. RojasMarpin Margyricarpus pinnatus (Lam.) KuntzeMyrcor Myrceugenia correifolia (Hook. & Arn.) O. BergNaschi Nassella chilensis (Trin.) E. Desv.Naspun Nassella pungens E. Desv.Puychi Puya chilensis MolinaRibpun Ribes punctatum Ruiz & Pav.Sencoq Senecio coquimbensis Phil.Sencum Senna cumingii (Hook. & Arn.) H.S. Irwin & BarnebySensin Senecio sinuatilobus DC.Solpin Solanum pinnatum Cav.Total Total plant coverTriver Tristerix verticillatus (Ruiz & Pav.) Barlow & WiensVal_sp Valeriana sp.Valcri Valeriana crispa Ruiz & Pav.
Table 4Mean (± S.D.) carbon and nitrogen concentration, C to N ratio, and isotopic(δ13C and δ15N) composition of Aextoxicon punctatum leaves at opposing forestfragment edges. Differences between leeward and windward edges were testedby dependent t-tests.
Windward Leeward t-value p
Foliar N (μg g−1) 31.4 ± 2.7 37.7 ± 4.8 4.79 ***Foliar C (μg g−1) 1432 ± 51.3 1464 ± 50.6 2.22 *Foliar C/N 45.9 ± 3.2 39.4 ± 4.3 5.07 ***δ13C (‰) −28.4 ± 0.8 −26.3 ± 1.3 6.93 ***δ15N (‰) −4.8 ± 0.7 −2.4 ± 1.1 8.08 ***
*p < 0.05; *** p < 0.001.
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
145

Fig. 5. The nutrient contents in soils and soil water content (SWC) along the gradient of open areas, shrub patches (divided into three size classes) and forest edges(windward and leeward). Mean ± SE (box)± SD (whisker). Different letters on the top of the figure correspond to significant differences in nutrient content or SWC(p < 0.05; Tukey HSD test; in case of SWC the different letters in parenthesis correspond to significant differences between leeward and windward forest side basedon dependent t-test; t= 3.68, p=0.001).
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
146

short, while both species are efficient in fog capture, Griselinia scandensmay consume most of it while Baccharis vernalis leaves a greater part toother species in the form of wetter soil underneath its canopy. ThereforeBaccharis vernalis acts as patch-forming species, playing a key role in theestablishment of Aextoxicon punctatum and other forest species, being acrucial driver of long-term patch dynamics.
4.2. Environmental differences
A continuous increase of soil resources from open areas to forestpatches corresponds to community changes and indicates a resourceaccumulation with a patch growth. Moreover, Baccharis patches andwindward edges of forest fragments had similar soil moisture, sug-gesting a similar microenvironment. As rainfall in this semiarid regionis highly variable among years, and generally insufficient for forestestablishment (less than 150mm), fog supply is crucial for long-termforest survival (del-Val et al., 2006). Fog is usually delivered from thesouth (Macek et al., 2018), and this directionality of water supplysupports the formation of linear patterns similar to other arid systems(Aguiar and Sala, 1999). The asymmetric water supply determines asharp gradient across forest fragments in all moisture-related and nu-trient content variables (Salgado-Negret et al., 2013; Stanton et al.,2013). Indeed, less negative δ13C values of leaves that grew at theleeward side indicate stomatal closure, which is under the conditions inFray Jorge most likely imposed due to drought stress (Ehleringer et al.,1993; Farquhar et al., 1989). This greater water use efficiency of treesat the leeward edge confirms contrasting water supply through time(Macek et al., 2009b), and indicates its importance for supporting ad-ditional biodiversity in forest fragments (del-Val et al., 2006). Greater Ncontent in Aextoxicon leaves at the leeward side can be explained by theconcurrence of two factors. First, as N brought by fog accumulates withtime, it resulted in greater leaf N at leeward edges, where older trees are(del-Val et al., 2006; Gutiérrez et al., 2008). Therefore, we can rule outN shortage as the primary cause of tree mortality observed at the lee-ward edge of forest fragments (Gutiérrez et al., 2008). Second, differ-ences in δ15N indicate different N sources. The δ15N of plants unable tofix N is usually greater than 0‰, while δ15N of N-fixing species is be-tween -2‰ and 0‰ (Evans and Ehleringer, 1994; Männel et al., 2007).Therefore the δ15N values of plants at leeward side of forest patchessuggest that their N partly originates from N fixation. However, thedepleted δ15N signal indicates a direct input from the fog (Peterson andFry, 1987), in contrast with nearby Atacama Desert, where plants donot show a significant N input from fog (Evans and Ehleringer, 1994).
4.3. Dynamics of forest patches
Baccharis vernalis is a drought-tolerant species that can establish inopen habitats in Fray Jorge coastal hills. Its wind-exposed individualshence act as ecosystem engineers (sensu Jones et al., 1997) by supplyingwater to other species eventually leading to changes in nutrient contentand species diversity and becoming islands of diversity in the sur-rounding matrix. Similar facilitation effects have been reported nearbyconcerning other non-forest, woody species (van Zonneveld et al.,2012), and elsewhere in semiarid systems (e.g., Butterfield and Briggs,2009; Pugnaire et al., 1996). Successional changes in shrub patchesmay be related to the accumulation of resources over time, and speciessuch as Aextoxicon punctatum, Myrceugenia correifolia and Griseliniascandens are facilitated but end up shading out the facilitator (Baccharisvernalis), which is, indeed, absent from inside forest patches (Stantonet al., 2013). A critical Baccharis patch size probably lies between smalland intermediate sizes, as small patches had lower nutrients and morethan half the Aextoxicon punctatum incidence as compared to bothlarger ones. Similar pattern formation, driven by a succession fromscattered shrubs to forests, has been reported for example from wetlandecosystems (Wetzel, 2002). Besides, shrubs are crucial in forest for-mation and regeneration dynamics at windward forest edges. Never-theless, our results provided evidence that Aextoxicon punctatum cannotrecruit in the leeward edge of forest fragments, mainly due to waterscarcity. Furthermore, the presence of Aextoxicon punctatum saplingswas strongly negatively correlated with the cover of Aristeguietia salvia,negative interactions may thus also occur between these two speciesalong edges. Aristeguietia salvia could either act as strong competitor(e.g. for light, or space through dead leaves, as it forms very densestands), or through changing soil chemistry, since Aristeguietia speciesare known to produce allelopathic substances (Morikawa et al., 2012).Therefore, shrub species composition at windward and leeward edgesof forest fragments likely play a crucial role in the patterns of estab-lishment of Aextoxicon punctatum; further research on the possible al-lelopathic is still needed.
A negative correlation between Aextoxicon punctatum cover inBaccharis patches and the distance to nearest forest fragment indicatesthat, although Baccharis vernalis may establish everywhere in thelandscape, Aextoxicon punctatum shows dispersal limitation and cannotbecome established beyond 30–35m from an existing fragment.Baccharis shrubs attract seed-dispersing birds (Cornelius et al., 2000;Núñez-Ávila et al., 2013) that use shrubs as perches (Toh et al., 1999).Furthermore, genetic analyses are consistent with our findings as theyrevealed strongly leptocuric distribution of seeds from maternal tree
Fig. 6. Regressions between soil water content (SWC) and cover of two dominant species in Baccharis patches, Baccharis vernalis (left), and Griselinia scandens (right).The patches are divided into three groups according to their size, and the best fitting regression selected using AIC is shown; in the case of SWC in intermediatepatches and Baccharis vernalis cover, no appropriate fit was found.
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
147

(Aextoxicon is a dioecious species), very rarely exceeding 50m (Núñez-Ávila et al., 2013). We therefore recognize that seed dispersal limitationmay explain scanty tree establishment outside forests, and we furtherstress the importance of solitary shrubs for tree regeneration there.
5. Conclusion
We gathered strong evidence that shrub species play crucial roles inthe shrub‒forest patch dynamics in the Fray Jorge. By modifying soilproperties, and enhancing water supply derived from incoming oceanicfog, Baccharis vernalis shrubs represent a key element in forest nuclea-tion. Forest fragments should grow towards the windward edge by fogcapture and by biotic interactions, which are favorable for tree estab-lishment. Away from forest patches, Baccharis vernalis promotes newforest patch initiation and successional change through tree establish-ment in its understory. If this mechanism of tree recruitment is oper-ating in Fray Jorge, presumably at slow pace, this may be an argumentagainst the prevailing idea that forest patches are dying because ofreductions in water supply. New forest patches may be starting atvarious locations in the landscape inundated by the fog.
Acknowledgements
We thank CONAF for permission to work at Fray Jorge Forest NPand Jana Macková and two anonymous reviewers for constructivecomments. PM was supported by JAE-Doc Program (Junta para laAmpliación de Estudios) co-financed by ESF and MSMTLM2015078;C.S. was financially supported by the Swiss National Foundation(PAOOP3_136474; PZ00P3_148261). F.I.P. was partly supported byMINECO (grant CGL2014-59010-R). This publication is also part of theresearch program of the Long-term Socio-Ecological Research (LTSER)Network-Chile, supported by CONICYT PIA APOYO CCTEAFB170008grant, and by Millennium Scientific Initiative grant P05-002, fromGovernment of Chile.
Appendix A. Supplementary data
Supplementary material related to this article can be found, in theonline version, at doi:https://doi.org/10.1016/j.ppees.2018.09.002.
References
Aguiar, M.R., Sala, O.E., 1999. Patch structure, dynamics and implications for the func-tioning of arid ecosystems. Trends Ecol. Evol. 14, 273–277.
Borthagaray, A.I., Fuentes, M.A., Marquet, P.A., 2010. Vegetation pattern formation in afog-dependent ecosystem. J. Theor. Biol. 265, 18–26.
Butterfield, B.J., Briggs, J.M., 2009. Patch dynamics of soil biotic feedbacks in theSonoran Desert. J. Arid Environ. 73, 96–102.
Cereceda, P., Osses, P., Larrain, H., Farias, M., Lagos, M., Pinto, R., Schemenauer, R.S.,2002. Advective, orographic and radiation fog in the Tarapaca region. Chile. Atmos.Res. 64, 261–271.
Cereceda, P., Larrain, H., Osses, P., Farias, M., Egana, I., 2008. The climate of the coastand fog zone in the Tarapaca Region, Atacama Desert, Chile. Atmos. Res. 87,301–311.
Cornelius, C., Cofre, H., Marquet, P.A., 2000. Effects of habitat fragmentation on birdspecies in a relict temperate forest in semiarid Chile. Conserv. Biol. 14, 534–543.
Dawson, T.E., 1998. Fog in the California redwood forest: ecosystem inputs and use byplants. Oecologia 117, 476–485.
del-Val, E., Armesto, J.J., Barbosa, O., Christie, D.A., Gutiérrez, A.G., Jones, C.G.,Marquet, P.A., Weathers, K.C., 2006. Rain forest islands in the chilean semiarid re-gion: Fog-dependency, ecosystem persistence and tree regeneration. Ecosystems 9,598–608.
Ehleringer, J.R., Hall, A.E., Farquhar, G.D., 1993. Stable Isotopes and Plant Carbon–WaterRelations. Academic Press, London, UK.
Evans, R.D., Ehleringer, J.R., 1994. Plant δ15N values along a fog gradient in the Atacamadesert. Chile. J. Arid Environ. 28, 189–193.
Ewing, H.A., Weathers, K.C., Templer, P.H., Dawson, T.E., Firestone, M.K., Elliott, A.M.,Boukili, V.K.S., 2009. Fog water and ecosystem function: heterogeneity in aCalifornia redwood forest. Ecosystems 12, 417–433.
Farquhar, G.D., Ehleringer, J.R., Hubbick, K.T., 1989. Carbon isotope discrimination andphotosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 503–537.
Fibich, P., Vítová, A., Macek, P., Lepš, J., 2013. Establishment and spatial associations ofrecruits in meadow gaps. J. Veg. Sci. 24, 496–505.
Fortin, M.J., Dale, M.R.T., 2005. Spatial Analysis: A Guide for Ecologists. CambridgeUniversity Press, Cambridge, UK.
Garreaud, R., Barichivich, J., Christie, D.A., Maldonado, A., 2008. Interannual variabilityof the coastal fog at Fray Jorge relict forests in semiarid Chile. J. Geophys. Res.-Biogeosci. 113.
Garreaud, R.D., Molina, A., Farias, M., 2010. Andean uplift, ocean cooling and Atacamahyperaridity: a climate modeling perspective. Earth Planet. Sci. Lett. 292, 39–50.
Getzin, S., Yizhaq, H., Bell, B., Erickson, T.E., Postle, A.C., Katra, I., Tzuk, O., Zelnik, Y.R.,Wiegand, K., Wiegand, T., Meron, E., 2016. Discovery of fairy circles in Australiasupports self-organization theory. Proc. Natl. Acad. Sci. 113, 3551–3556.
Gotelli, N.J., Colwell, R.K., 2001. Quantifying biodiversity: procedures and pitfalls in themeasurement and comparison of species richness. Ecol. Lett. 4, 379–391.
Greig-Smith, P., 1979. Pattern in vegetation. J. Ecol. 67, 755–779.Gutiérrez, A.G., Barbosa, O., Christie, D.A., del-Val, E., Ewing, H.A., Jones, C.G., Marquet,
P.A., Weathers, K.C., Armesto, J.J., 2008. Regeneration patterns and persistence ofthe fog-dependent Fray Jorge forest in semiarid Chile during the past two centuries.Glob. Change Biol. Bioenergy 14, 161–176.
Haase, P., Pugnaire, F.I., Clark, S.C., Incoll, L.D., 1996. Spatial patterns in a two-tieredsemi-arid shrubland in southeastern Spain. J. Veg. Sci. 7, 527–534.
Huntley, L.B., Doley, D., Yayes, D.J., Boonsaner, A., 1997. Water balance of an Australiansubtropical rainforest at altitude: the ecological and physiological significance ofintercepted cloud and fog. Aust. J. Bot. 45, 311–329.
Johnstone, J.A., Dawson, T.E., 2010. Climatic context and ecological implications ofsummer fog decline in the coast redwood region. Proc. Natl. Acad. Sci. U. S. A. 107,4533–4538.
Jones, C.G., Lawton, J.H., Shachak, M., 1997. Positive and negative effects of organismsas physical ecosystem engineers. Ecology 78, 1946–1957.
Karbulková, J., Schreiber, L., Macek, P., Šantrůček, J., 2008. Differences between waterpermeability of astomatous and stomatous cuticular membranes: effects of air hu-midity in two species of contrasting drought-resistance strategy. J. Exp. Bot. 59,3987–3995.
Lepš, J., Šmilauer, P., 2003. Multivariate Analysis of Ecological Data Using CANOCO.Cambridge University Press, Cambridge.
López-Cortés, F., López, D., 2004. Antecedentes bioclimáticos del Parque Nacional BosqueFray Jorge. In: Squeo, F.A., Gutiérrez, J.R., Hernández, I.R. (Eds.), Historia Naturaldel Parque Nacional Bosque Fray Jorge. Ediciones Universidad de La Serena, LaSerena, Chile, pp. 45–60.
Macek, P., Rejmánková, E., Fuchs, R., 2009a. Biological activities as patchiness drivingforces in wetlands of northern Belize. Oikos 118, 1687–1694.
Macek, P., Macková, J., de Bello, F., 2009b. Morphological and ecophysiological traitsshaping altitudinal distribution of three Polylepis treeline species in the dry tropicalAndes. Acta Oecol. 35, 778–785.
Macek, P., Schöb, C., Núñez-Ávila, M.C., Hernández Gentina, I.R., Pugnaire, F.I., Armesto,J.J., 2018. Shrub facilitation drives tree establishment in a semiarid fog-dependentecosystem. App. Veg. Sci. 21, 113–120.
Macková, J., Vašková, M., Macek, P., Hronková, M., Schreiber, L., Šantrůček, J., 2013.Plant response to drought stress simulated by ABA application: changes in chemicalcomposition of cuticular waxes. Environ. Exp. Bot. 86, 70–75.
Maestre, F.T., Bautista, S., Cortina, J., 2003. Positive, negative, and net effects in grass-shrub interactions in mediterranean semiarid grasslands. Ecology 84, 3186–3197.
Männel, T.T., Auerswald, K., Schnyder, H., 2007. Altitudinal gradients of grasslandcarbon and nitrogen isotope composition are recorded in the hair of grazers. GlobalEcol. Biogeogr. 16, 583–592.
Martorell, C., Ezcurra, E., 2007. The narrow-leaf syndrome: a functional and evolutionaryapproach to the form of fog-harvesting rosette plants. Oecologia 151, 561–573.
Morikawa, C.I.O., Miyaura, R., Figueroa, M.D.T.Y., Salgado, E.L.R., Fujii, Y., 2012.Screening of 170 Peruvian plant species for allelopathic activity by using theSandwich Method. Weed Biol. Manag. 12, 1–11.
Moro, M.J., Pugnaire, F.I., Haase, P., Puigdefabregas, J., 1997. Mechanisms of interactionbetween a leguminous shrub and its understorey in a semi-arid environment.Ecography 20, 175–184.
Núñez-Ávila, M.C., Armesto, J.J., 2006. Relict islands of the temperate rainforest treeAextoxicon punctatum (Aextoxicaceae) in semi-arid Chile: genetic diversity and bio-geographic history. Aust. J. Bot. 54, 733–743.
Núñez-Ávila, M.C., Uriarte, M., Marquet, P.A., Armesto, J.J., 2013. Decomposing re-cruitment limitation for an avian-dispersed rain forest tree in an anciently frag-mented landscape. J. Ecol. 101, 1439–1448.
Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R., O’Hara, R.B., Simpson,G.L., Solymos, P., Henry, M., Stevens, H., Wagner, H., 2012. Vegan: CommunityEcology Package. R Package Version 2.0-5. http://CRAN. project.org/pack-age=vegan. .
Peterson, B.J., Fry, B., 1987. Stable isotopes in ecosystem studies. Ann. Rev. Ecol. Syst.18, 293–320.
Prieto, I., Padilla, F.M., Armas, C., Pugnaire, F.I., 2011. The role of hydraulic lift onseedling establishment under a nurse plant species in a semi-arid environment.Perspect. Plant Ecol. Evol. Syst. 13, 181–187.
Pugnaire, F.I., Haase, P., Puigdefábregas, J., Cueto, M., Clark, S.C., Incoll, L.D., 1996.Facilitation and succession under the canopy of a leguminous shrub, Retamasphaerocarpa, in a semi-arid environment in south-east Spain. Oikos 76, 455–464.
Pugnaire, F.I., Armas, C., Valladares, F., 2004. Soil as a mediator in plant-plant interac-tions in a semi-arid community. J. Veg. Sci. 15, 85–92.
Pugnaire, F.I., Armas, C., Maestre, F.T., 2011. Positive plant interactions in the IberianSoutheast: mechanisms, environmental gradients, and ecosystem function. J. AridEnviron. 75, 1310–1320.
R Core Team, 2012. R: A Language and Environment for Statistical Computing. 3-900051-07-0, URL. R Foundation for Statistical Computing, Vienna, Austria. http://
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
148

www.R-project.org/.Rietkerk, M., Van de Koppel, J., 2008. Regular pattern formation in real ecosystems.
Trends Ecol. Evol. 23, 169–175.Robinson, D., 2001. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 16,
153–162.Rundel, P.W., Dillon, M.O., 1998. Ecological patterns in the Bromeliaceae of the lomas
formations of Coastal Chile and Peru. Plant Syst. Evol. 212, 261–278.Sakamoto, Y., Ishiguro, M., Kitagawa, G., 1986. Akaike Information Criterion Statistics.
Reidel Publishing Company, Dordrecht, The Netherlands.Salgado-Negret, B., Pérez, F., Markesteijn, L., Castillo, M.J., Armesto, J.J., 2013.
Diverging drought-tolerance strategies explain tree species distribution along a fog-dependent moisture gradient in a temperate rain forest. Oecologia 173, 625–635.
Schöb, C., Kammer, P.M., Kikvidze, Z., Choler, P., Veit, H., 2008. Changes in speciescomposition in alpine snowbeds with climate change inferred from small-scale spatialpatterns. Web Ecol. 8, 142–159.
Seabloom, E.W., Bjornstad, O.N., Bolker, B.M., Reichman, O.J., 2005. Spatial signature ofenvironmental heterogeneity, dispersal, and competition in successional grasslands.Ecol. Monogr. 75, 199–214.
Shmida, A., Evenari, M., Noy-Meir, I., 1986. Hot desert ecosystems: an integrated view.In: Evenari, M., Noy-Meir, I., Goodall, D.W. (Eds.), Hot Deserts and Arid Shrublands.Elsevier Pub, Amsterdam.
Stanton, D., 2011. Filter Feeding Forests: Epiphytes and Fog in the Self-Organization andFunction of Ecosystems. Princeton University, USA.
Stanton, D.E., Negret, B.S., Armesto, J.J., Hedin, L.O., 2013. Forest patch symmetry de-pends on direction of limiting resource delivery. Ecosphere 4.
ter Braak, C.J.F., Šmilauer, P., 2002. CANOCO Reference Manual and CanoDraw forWindows User’s Guide: Software for Canonical Community Ordination (version 4.5).Microcomputer Power, Ithaca, NY.
Tirado, R., Pugnaire, F.I., 2005. Community structure and positive interactions in con-straining environments. Oikos 111, 437–444.
Toh, I., Gillespie, M., Lamb, D., 1999. The role of isolated trees in facilitating tree seedlingrecruitment at a degraded sub-tropical rainforest site. Restor. Ecol. 7, 288–297.
Troxler Gann, T.G., Childers, D.L., Rondeau, D.N., 2005. Ecosystem structure, nutrientdynamics, and hydrologic relationships in tree islands of the southern Everglades,Florida. USA. Forest Ecol. Manag. 214, 11–27.
Uclés, O., Villagarcía, L., Moro, M., Cantón, Y., Domingo, F., 2014. Role of dewfall in thewater balance of a semiarid coastal steppe ecosystem. Hydrol. Process. 28,2271–2280.
van Zonneveld, M.J., Gutiérrez, J.R., Holmgren, M., 2012. Shrub facilitation increasesplant diversity along an arid scrubland-temperate rain forest boundary in SouthAmerica. J. Veg. Sci. 23, 541–551.
Watt, A.S., 1947. Pattern and process in the plant community. J. Ecol. 35, 1–22.Wetzel, P.R., 2002. Tree island ecosystems of the world. In: Sklar, F.H., van der Valk, A.G.
(Eds.), Tree Islands of the Everglades. Kluwer Academic Publishers, Dordrecht, TheNetherlands, pp. 19–69.
White, L.P., 1971. Vegetation stripes on sheet wash surfaces. J. Ecol. 59, 615–622.
P. Macek et al. Perspectives in Plant Ecology, Evolution and Systematics 34 (2018) 140–149
149




















