
Peptides with Arginine at P2 Similarity to HLA-B27 that Binds with Only Limited Sequence Express an...
12
of January 3, 2014. This information is current as Peptides with Arginine at P2 Sequence Similarity to HLA-B27 that Binds an MHC Class I Molecule with Only Limited Gorillas with Spondyloarthropathies Express Sette and David I. Watkins Alessandro L. Neiffer, Bonnie Raphael, William Hildebrand, Lorentzen, Clare Shufflebotham, Edward J. Collins, Donald Prilliman, Todd M. Allen, Kevin J. Schwartz, David Julie A. Urvater, Heather Hickman, John L. Dzuris, Kiley http://www.jimmunol.org/content/166/5/3334 2001; 166:3334-3344; ; J Immunol References http://www.jimmunol.org/content/166/5/3334.full#ref-list-1 , 20 of which you can access for free at: cites 58 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2001 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on January 3, 2014 http://www.jimmunol.org/ Downloaded from by guest on January 3, 2014 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Peptides with Arginine at P2 Similarity to HLA-B27 that Binds with Only Limited Sequence Express an...

of January 3, 2014.This information is current as
Peptides with Arginine at P2Sequence Similarity to HLA-B27 that Bindsan MHC Class I Molecule with Only Limited Gorillas with Spondyloarthropathies Express
Sette and David I. WatkinsAlessandroL. Neiffer, Bonnie Raphael, William Hildebrand,
Lorentzen, Clare Shufflebotham, Edward J. Collins, DonaldPrilliman, Todd M. Allen, Kevin J. Schwartz, David Julie A. Urvater, Heather Hickman, John L. Dzuris, Kiley
http://www.jimmunol.org/content/166/5/33342001; 166:3334-3344; ;J Immunol
Referenceshttp://www.jimmunol.org/content/166/5/3334.full#ref-list-1
, 20 of which you can access for free at: cites 58 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 3, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Gorillas with Spondyloarthropathies Express an MHC Class IMolecule with Only Limited Sequence Similarity to HLA-B27that Binds Peptides with Arginine at P21
Julie A. Urvater,2* Heather Hickman,† John L. Dzuris,‡ Kiley Prilliman, † Todd M. Allen,*Kevin J. Schwartz,* David Lorentzen,§ Clare Shufflebotham,3* Edward J. Collins,¶
Donald L. Neiffer,4i Bonnie Raphael,§§ William Hildebrand, † Alessandro Sette,‡ andDavid I. Watkins 5*§
The human MHC class I gene,HLA-B27, is a strong risk factor for susceptibility to a group of disorders termed spondyloar-thropathies (SpAs).HLA-B27-transgenic rodents develop SpAs, implicatingHLA-B27 in the etiology of these disorders. Severalnonhuman primates, including gorillas, develop signs of SpAs indistinguishable from clinical signs of humans with SpAs. Todetermine whether SpAs in gorillas have a similarHLA-B27-related etiology, we analyzed the MHC class I molecules expressedin four affected gorillas. Gogo-B01, isolated from three of the animals, has only limited similarity to HLA-B27 at the end of thea1 domain. It differs by several residues in the B pocket, including differences at positions 45 and 67. However, the molecularmodel of Gogo-B*0101 is consistent with a requirement for positively charged residues at the second amino acid of peptides boundby the MHC class I molecule. Indeed, the peptide binding motif and sequence of individual ligands eluted from Gogo-B*0101demonstrate that, like HLA-B27, this gorilla MHC class I molecule binds peptides with arginine at the second amino acid positionof peptides bound by the MHC class I molecule. Furthermore, live cell binding assays show that Gogo-B*0101 can bind HLA-B27ligands. Therefore, although most gorillas that develop SpAs express an MHC class I molecule with striking differences toHLA-B27, this molecule binds peptides similar to those bound by HLA-B27. The Journal of Immunology,2001, 166: 3334–3344.
T he strongest known association between the MHC anddisease susceptibility is that ofHLA-B27and inflamma-tory spondyloarthropathies (SpAs)6 in humans. Several
theories have been proposed to explain the role of B27 in thedevelopment of SpAs and have been reviewed elsewhere (1–3).The primary function of MHC class I molecules is to present en-dogenously produced peptides to CTLs. Thus the most intuitive of
these theories, the arthritogenic peptide hypothesis, postulates thatB27 plays a direct role in pathogenesis by binding an arthritogenicpeptide (or peptides) and presenting it to autoreactive CTLs. Re-cent transgenic mouse and in vitro studies suggest that the role ofB27 in the mechanism of disease may be distinct from its primaryfunction as an Ag presentation and CD81 T cell restriction mol-ecule (4–7). However, the transgenic rat model provides evidencethat the specificity of peptides bound to B27 significantly influ-ences the prevalence of arthritis in these animals (8). Thus,whether the disease mechanism involves B27 functioning in itsconventional role of peptide binding molecule remains in question.
There are several features of B27 that make it unique amongMHC class I molecules. Crystal structures and molecular modelshave demonstrated that B27 is unique in its possession of an un-usually deep B pocket when compared with other MHC class Imolecules (9). Indeed, many groups have now eluted and se-quenced peptides bound to B27 and found that the peptides con-tained the bulky and positively charged amino acid, arginine, at thesecond position (10–12). Additionally, the combination of aminoacids that make up the B27 B pocket is unique to and conserved inall B27 subtypes (13), all of which bind peptides with arginine atthe second position. Of these residues, glutamic acid at position 45(E45), and cysteine at position 67 (C67) have been shown to becritical for peptide binding, cell surface expression, and CTL rec-ognition (13, 14). C67 has also been implicated in triggering au-toimmunity (15) and in the formation of uniqueb2-microglobulin(b2m)-free B27 heavy chain homodimers (7).
AlthoughB27 is the primary genetic factor determining suscep-tibility to SpAs, not allB27-positive individuals develop disease;this remains one of the mysteries of these disorders. In general,;0.2% of the general population will develop ankylosing spon-dylitis (AS), whereas 2% ofB27-positive individuals will develop
*Wisconsin Regional Primate Research Center, University of Wisconsin, Madison,WI 53715;†Department of Microbiology and Immunology, University of OklahomaHealth Sciences Center, Oklahoma City, OK 73190;‡Eppimune, San Diego, CA92121;§University of Wisconsin Histocompatibility Laboratory, Division of Labo-ratory Medicine, Department of Pathology and Laboratory Medicine, Madison, WI53792;¶Department of Microbiology and Immunology, University of North Carolina,Chapel Hill, NC 27599;iPittsburgh Zoo, Pittsburgh, PA 15206; and§§Wildlife HealthSciences, Bronx, NY 10460
Received for publication July 24, 2000. Accepted for publication December 11, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This research was supported by a Biomedical Science Award from the ArthritisFoundation and by National Institutes of Health Grant RR00167 (to the WisconsinRegional Primate Research Center).2 Current address: Fred Hutchinson Cancer Research Center, 1100 Fairview AvenueNorth, Seattle, WA 98109.3 Current address: Zeneca Pharmaceuticals, Mereside, Alderley Park, Macclesfield,Cheshire, SK10 4TG, U.K.4 Current address: Disney’s Animal Kingdom, P.O. Box 10000, Lake Buena Vista, FL32830.5 Address correspondence and reprint requests to Dr. David I. Watkins, WisconsinRegional Primate Research Center, University of Wisconsin, 1220 Capitol Court,Madison, WI 53715. E-mail address: [email protected] Abbreviations used in this paper: SpAs, spondyloarthropathies; AS, ankylosingspondylitis; ReA, reactive arthritis; P2, second amino acid of peptides bound by MHCclass I molecule; PBR, peptide binding region; nano-ES-MS/MS, nanoelectrospraytandem mass spectrometry.
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

the disease (16). Although there is no direct data showing whatpercentage ofB27-positive individuals will develop reactive ar-thritis (ReA), extrapolation from data summarized by Keat indicatethat 20–30% ofB27-positive individuals that contractShigellashould develop ReA (17). Sixty to 80% of ReA patients areB27-positive, a lower association than is seen in AS where as many as96% of patients areB27-positive. However, this allele is onlypresent in;7% of Caucasian populations.
Although SpAs are a common phenomenon in nonhuman primates,very little is known about the relationship between disease occurrenceand expression of MHC class I molecules in these species. Previously,other groups described AS in the gorilla, Beta (18), and post shigel-losis ReA in Holli and Husani (19). Another gorilla, Harry, also de-veloped ReA (D. Neiffer, manuscript in preparation). By examiningskeletal remains, Rothschild and Woods have also observed SpAs in20% of gorillas (20). Gorillas are one of humans closest relatives, lastsharing a common ancestor;10 million years ago (21, 22) and theyexpress homologs of theHLA-A, B, andC loci (23, 24). Given theassociation between B27 and SpAs and the remarkable similarity be-tween SpAs in gorillas and humans, we asked whether gorillas withSpAs expressed a MHC class I molecule with sequence and func-tional similarity to B27.
Materials and MethodsAnimals
Whole blood was obtained opportunistically by venipuncture from gorillasanesthetized for routine exams (Gorilla gorilla) at the Yerkes RegionalPrimate Research Center (Atlanta, GA), the Bronx Zoo (Wildlife Conser-vation Society, Bronx, NY), the Brookfield Zoo (Chicago Zoological So-ciety, Brookfield, IL), the Toledo Zoo (Toledo Zoological Society, Toledo,OH), and the Pittsburgh Zoo (Zoological Society of Pittsburgh, Pittsburgh,PA). Description of AS in Beta and ReA in Holli and Husani was reportedpreviously (18, 19). The clinical report of ReA in Harry is in preparation(D. Neiffer et al., manuscript in preparation).
Cell culture
PBL were separated from whole blood using Ficoll/diatrizoate gradientcentrifugation. These cells were cultured with 5mg/ml Con A (Sigma, St.Louis, MO) and 20 U/ml rIL-2 (a gift from Roche, Nutley, NJ). PBLs fromBeta and Holli were also transformed with EBV by culturing PBL withsupernatant of the B95-8 cell line (25) in the presence of cyclosporin A at0.2 mg/ml using a protocol modified from Lawlor et al. (26). Transformedand activated lymphocytes were cultured at 13 106 cells/ml in RPMI 1640medium (Life Technologies, Grand Island, NY) supplemented with 15%heat-inactivated FBS (Sterile Systems, Logan, UT), 2 mML-glutamine,5 3 1025 M 2-ME, 20 mM HEPES, 50 U/ml penicillin, and 50mg/mlstreptomycin.
DNA/RNA extraction, cDNA synthesis, and PCR
Total cellular RNA was extracted from 2–73 106 lymphocytes usingRNAzol (Tel-Test, Friendswood, TX). cDNA was synthesized from 0.1–1mg of RNA in a 20-ml reaction containing 50 mM Tris, pH 8.3, 5 mMMgCl2, 1 mM each of dATP, dGTP, dCTP, and dTTP (Gene AMP-Perkin-Elmer, Foster City, CA), 0.5mg random primers (Promega, Madison, WI),50 U of SuperScript II reverse transcriptase (Life Technologies, Gaithers-burg, MD), and 20 U of RNase inhibitor (Gene AMP-Perkin-Elmer).cDNA was synthesized at room temperature for 10 min, 42°C for 15 min,99° for 5 min, and 5° for 5 min in a Perkin-Elmer Cetus 9600 thermocycler(Norwalk, CT). PCR was then conducted in a Perkin-Elmer Cetus 9600using several sets of locus-specific primers. All of theB locus typing inTable III was conducted with theB locus-specific primer set GG59BXHO:59-CGGCCTCGAGATGCCTCCTCCTGCTGCTCTCGGC-39; GG39BH3:59-CGAAGCTTCCCTCACAACACAGCTGTCTCAGGCTTTT-39. For A,B, andC locus typing of Beta, Holli, and Husani, additional locus-specificprimer sets were used. They were as follows: The 59 generic primer,BETA2H33HO (59-CGCTCGAGGACTCAGAATCTCCCCAGACGCCGAG-39) paired with either 3PALOCH3 (59-CCGCAAGCTTTTGGGGAGGGAGCACAGGTCAGCGTGGGAAG-39), the B locus-specific primerBETA2H3, (59-CGAAGCTTGGAGGAAACACAGGTCAGCATGGGAAC-39), or 3PCLOCH3 (59-CCGCAAGCTTTCGGGGAGGGAACACAGGTCAGTGTGGGGAC-39). The latter four primers were derived
from primers in Lawlor et al. (23). We designed additional locus-specificprimer sets that were also used for these animals. These were: 59GGAXHO(59-CGGCCTCGAGATGGCGCCCCGAACCCTCSTCCTGCTA-39),39GGAH3 (59-CGGCAAGCTTCACACAAGGCAGCTGTCTCACACTTTA-39), GGCF (59-CKCCCCGAACCCTCA-39), and GGCR (59-AGGCTTTACAAGYGATGAGAGACT-39). Each primer was used at afinal concentration of 0.25mM. The PCR mixture contained 2 mM MgCl2,50 mM Tris, pH 8.3, and 2.5 U Amplitaq DNA polymerase (Perkin-ElmerCetus) in a final volume of 100ml. The reactions were denatured initiallyfor 2 min at 94°C followed by 30 cycles of 94°C for 1 min, 60°C for 1 min,72°C for 1.5 min, and a single final extension at 72°C for 10 min. RNA wasnot available for Harry, so MHC typing was performed on genomic DNAextracted from 200ml of blood using the QIAmp Blood kit (Qiagen, SantaClarita, CA). DNA (175 ng) was amplified in a 50-ml reaction using theInvitrogen PCR optimizer kit, Buffer F (Invitrogen, Carlsbad, CA) with theMHC class I generic primer NA1STAR1 (59-GCGAATTCGCTCYCACTCCWTGARGTATTTC-39) and theB friendly primer 39GGBA2(59-GTCCGCCGCGGTCCAGGAGCT-39) at a final concentration of0.5 mM.
Subcloning and sequencing
Amplified products were gel purified using QIAEX II suspension (Qiagen)and subcloned into the pCR2.1 vector using the TA cloning kit from In-vitrogen. Plasmid DNA (200 ng) was sequenced using the ABI PRISMDye Terminator Cycle Sequencing Ready Reaction Kit with AmpliTaqDNA Polymerase, FS, according to the manufacturer’s instructions (Per-kin-Elmer-Applied Biosystems, Foster City, CA). The sequencing primerused for screening, 62P: 59-GTGGGCTACGTGGACG-39, anneals to nu-cleotides 73–88 in exon 2 of MHC class I cDNAs. The sequence generatedby the 62P primer (near nucleotide 109 of exon 2 to nucleotide 216 of exon3) was aligned to a database of published gorilla MHC class I cDNAs usingMacVector (Scientific Imaging Systems, New Haven, CT). Sequences thatdid not match to the database were assigned a new allele name accordingto established convention (27). Briefly, the MHC of the lowland gorilla isdesignated by a four-letter abbreviation of the scientific names of the spe-cies,Gorilla gorilla, or Gogo.This is followed by the locus designationbased on homology to human class I loci. Allele numbers were assigned inthe order they were isolated. Three copies of each new MHC class I cDNAwere sequenced full length to minimize reporting of PCR-generated nu-cleotide substitutions and to confirm locus identity. Primers used for se-quencing were reported previously (28). Sequences of new cDNAs shownin this paper have been deposited with GenBank and have accession num-bers AF157406–AF157411.
Antibodies
The W6/32 hybridoma was obtained from the American Type Culture Col-lection (ATCC) (Manassas, VA). This mouse mAb is directed against hu-man MHC class I proteins. W6/32 from hybridoma supernatant was puri-fied over a Sepharose CL4B column (Sigma) for use in purifying MHCclass I molecules. The 949 (anti-class II Ab) hybridoma obtained fromATCC was used to generate mouse 949 ascites according to establishedprotocols (29).
Molecular modeling of Gogo-B*0101
A molecular model of Gogo-B01 was constructed using the crystallographiccoordinates of HLA-B27 (PDB1HSA) (30). Substitutions in the extracellularportion of B27 to make it into Gogo-B01 were made using the graphicalprogram O (31). The substituted side chains were placed in the rotamer loca-tion (32) that minimized steric clashes with neighboring residues. Local min-imization was performed to remove bad contacts, and the model was examinedvisually for poor interactions. The resulting model was subjected to 40 cyclesof energy minimization refinement using X-PLOR (33) with a small harmonicconstraint placed on thea carbon positions.
Stable transfection of Gogo-B*0101 into the 721.221 cell line
A clone containing the consensus cDNA for Gogo-B*0101 was subclonedinto the pKG5 expression vector (a gift from Andrew McMichael, OxfordUniversity, Oxford, U.K.). This vector was then electroporated into the721.221 cell line, a cloned EBV-transformed B lymphoblastoid cell line(BLCL) with homozygous deletions of the MHC class I loci (34). Cell line721.221 cells (7.53 106) were transfected in a 0.4-cm electroporationcuvette with 25mg of plasmid DNA. Electroporation was conducted witha Bio-Rad Gene Pulser II Electroporation System (Bio-Rad, Hercules, CA)at 200 V and a capacitance of 950mF. The cells were then put into 50 mlof RPMI 1640 culture medium supplemented with 50 U/ml penicillin, 50
3335The Journal of Immunology
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

mg/ml streptomycin, 2 mML-glutamine, 5% defined FBS (HyClone, Lo-gan, UT), and 10% defined/supplemented bovine calf serum (HyClone)and plated at 1 ml/well in 24-well plates. The cells were incubated for 2days at 37°C. On day 3, the cells were placed under selection by adding 1ml of culture medium containing G418 (Life Technologies, Gaithersburg,MD) for a final concentration of 650mg/ml. About 4 wk later, viabletransfectants were tested for MHC class I surface expression by flow cy-tometry with the W6/32 mAb directly conjugated to FITC (Sigma). Thetransfectant with the highest level of MHC class I expression was selectedto be grown up for peptide elution studies.
We also produced soluble Gogo-B*0101 transfectants to produce higheramounts of bound peptides for sequencing. This method was describedpreviously (35).
Affinity purification of Gogo-B*0101
MHC class I molecules were purified from 721.221 transfectants accordingto a modified protocol as previously described (36, 37). Briefly, 63 109
cells were washed in cold HBSS (Life Technologies), harvested, and fro-zen until needed. Thawed cells were then resuspended in 100 ml of 1%Nonidet P-40 lysis buffer containing 0.25% sodium deoxycholate, 174mg/ml PMSF, 5mg/ml aprotinin, 10mg/ml leupeptin, 10mg/ml pepstatinA, 20 mg/ml iodoacetamide, 0.2% sodium azide, and 0.003mg/ml EDTA.Cell lysates were incubated at 4°C for 1 h, centrifuged at 100,0003 g at4°C to remove cellular debris, and then filtered sequentially through 0.8-and 0.22-mm Nalgene filters to remove any remaining lipids. Filtered ly-sates were then passed twice over a 949-coupled protein A-Sepharose col-umn to preclear the lysate. The flowthrough was passed twice over twoconsecutive W6/32-coupled columns to specifically bind MHC class I mol-ecules. The protein A beads of the W6/32 columns were then washedseparately, twice with lysis buffer (without protease inhibitors), twice witha high salt buffer (1 M NaCl, 20 mM Tris pH 8.0), and twice with a no saltbuffer (20 mM Tris, pH 8.0). Purification of soluble Gogo-B*0101 was asdescribed previously (35).
Purification of Gogo-B*0101-bound peptides
Peptides were eluted from Gogo-B*0101 as described previously (36).Briefly, the protein A beads were incubated in 0.2 N acetic acid. The beadswere then briefly centrifuged and transferred to a new tube and the processwas repeated. Glacial acetic acid (100 ml) was added to each tube to allowfor dissociation of the MHC heavy chain/b2m/peptide complexes. MHCclass I heavy chains,b2m, and W6/32 Abs were then separated from thepeptides by centrifugation through an Ultrafree-CL filter (500 NMWL;Millipore, Bedford, MA). Peptide yields were determined by quantitationof Gogo-B*0101 heavy chain using SDS-PAGE. Purification of peptidesfrom soluble Gogo-B*0101 was as described (35).
HPLC fractionation and automated Edman degradationsequencing of peptides
The filtered peptide eluate was purified by reverse phase HPLC on a 1.03150 mm C18 column (Michrom Bioresources, Auburn, CA) using the fol-lowing gradient conditions at a flow rate of 40ml/min: 2–10% acetonitrilein 0.02 min, 10–60% acetonitrile in 2 min. The entire region correspond-ing to UV absorbance at 215 nm was collected during the gradient andsubjected to pooled Edman degradation on a model 492A pulsed liquidphase protein sequencer (Perkin-Elmer Applied Biosystems Division, Nor-walk, CT) with underivatized cysteine. Nonpeptide material, which copu-rified with the peptides in the first experiment (from the cell-bound mol-ecules), eluted as two peaks on the HPLC analysis. One of these peaksobscured threonine and aspartate for the majority of the cycles and glycinefor the first three cycles. Only data from the second run (peptides elutedfrom soluble molecules) is used for these amino acids. Raw data analysiswas performed according to established protocols (38–40). The averagerelative frequency table was generated according to Kubo et al. (38).
Nanoelectrospray tandem mass spectrometry (nanoES-MS/MS)Sequencing of individual peptide ligands was as previously described (41).Typical nanoES-MS/MS runs involved gating for an ion with the firstquadrupole and scanning a range with the third quadrupole of 30–1200m/zusing a step size of 0.2 atomic mass units and a dwell time of 1.5 ms withunderivatized lysine; the collision gas (Ar) was adjusted in each case tooptimize fragmentation for the ion examined. Nano-ES-MS/MS data wereevaluated and interpreted using the Predict Sequence algorithm (BioMul-tiView software; PE Sciex, Boston, MA) as well as PeptideSearch 3.0.2(42) in instances of low ligand ionization/concentration or poor fragmen-tation. Advanced Basic Local Alignment Search Tool searches were per-formed against databases available through the National Center for Bio-
technology Information (National Institutes of Health, Bethesda, MD) webserver to identify homology with currently catalogued sequences.
Peptides and125iodine labeling
Peptides were obtained as lyophilized products from Chiron Mimotopes(San Diego, CA) or synthesized at Epimmune using standard t-Boc orF-moc solid phase synthesis methods (43). Peptides were stored in stocksolutions at either 10 or 20 mg/ml in 100% DMSO and then diluted inRPMI 1640 for use in the cellular assays. HPLC-purified peptides wereradiolabeled with125I according to the chloramine-T method (44).
Live cell binding assays
Peptide binding to gorilla MHC class I molecules was measured by theability of test, unlabeled peptides to inhibit binding of a radiolabeledpeptide probe in live cell binding assays The live cell binding assay wasperformed as previously described (36, 45). Briefly, gorilla MHC classI-transfected 721.221 cells (106 cells/ml) were preincubated overnight inRPMI 1640 supplemented with 15% FBS,L-glutamine, penicillin (100IU/ml), and streptomycin (100mg/ml) at room temperature. Then, cellswere washed twice in RPMI 1640 without supplements and resuspended toa final concentration of 1.253 107 cells/ml in RPMI 1640 supplementedwith 3 mg/ml b2m (Scripps Clinic and Research Foundation, La Jolla, CA).
Aliquots of 2 3 106 cells/well were incubated in a 96-well U-bottommicrotiter plate with 105 cpm (10ml) of specific radiolabeled peptide andin the presence of a protease inhibitor cocktail (final concentrations of 250mg/ml PMSF, 1.07 mg/ml EDTA, 62.5mg/ml pepstatin A, 60mg/ml1-chloro-3-tosylamido-7-amino-2-heptanone-hydrochloride and 325mg/mlphenanthroline). Following a 4-h incubation at 20°C, unbound peptide wasremoved by washing three times in serum-free medium, followed by pas-sage through a FBS gradient. The amount of a bound, labeled peptide wasthen determined by counting pelleted cells on a gamma scintillationcounter. For inhibition assays, a dose range of unlabeled peptide inhibitorswas coincubated with the radiolabeled probe and the cells, and the con-centration of peptide yielding 50% inhibition of the binding of the radio-labeled probe peptide was calculated (IC50).
ResultsGorillas that develop SpAs express an MHC class I moleculewith limited similarity to B27
Gorillas develop signs of SpAs indistinguishable from clinical signsseen in humans with SpAs. The case description of Beta’s AS was thefirst in a nonhuman primate (18) (Table I). She was initially diagnosedwith arthritis in 1979 when radiographs were taken due to a stiffnessin her gait. Later her condition worsened further; she moved withgreat difficulty and used an awkward, rigid gait. Under anesthesia,board-like rigidity of the dorsal spine and extreme bilateral contrac-tures of the hips were noted. There was also evidence of chronicsynovitis of the larger peripheral joints, and swelling and/or contrac-tures involving the shoulders, elbows, wrist, knees, and ankles in asymmetric fashion. Radiographs showed hallmark and progressivechanges typical of AS, including advanced sacroiliitis and lumbarspondylitis. Beta’s son, Kwashi developed inflammatory synovitis ofthe right wrist (18) (Table I). Holli and Husani developed signs verytypical of ReA (19). Both developedShigella flexnerienteritis andsubsequently developed inflammatory joint disease. Harry also devel-oped signs of ReA following shigellosis (Table I).
Given the remarkable similarity between SpAs in gorillas andhumans and the close association ofB27 with these diseases inhumans, we cloned and sequenced all of the MHC class I mole-cules expressed in the four affected gorillas (Table II). Of all theMHC class I molecules isolated from these animals,Gogo-B01was the most similar toB27(Figs. 1 and 2). However, at the aminoacid level, Gogo-B*0101 is most similar to HLA-B*1513, differ-ing by 21 residues. HLA-B*2702 is the next most similar humanclass I molecule, differing from Gogo-B*0101 by 22 aa (Fig. 1 anddata not shown). Fifteen of those differences occur in the poly-morphic a1 and a2 domains that comprise the peptide bindingregion (PBR). Interestingly, Gogo-B01 is identical with HLA-B*2702 from residues 71 to 90 at the end of thea1 domain (Figs.
3336 GORILLA MHC MOLECULE BINDS PEPTIDES WITH ARGININE AT P2
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

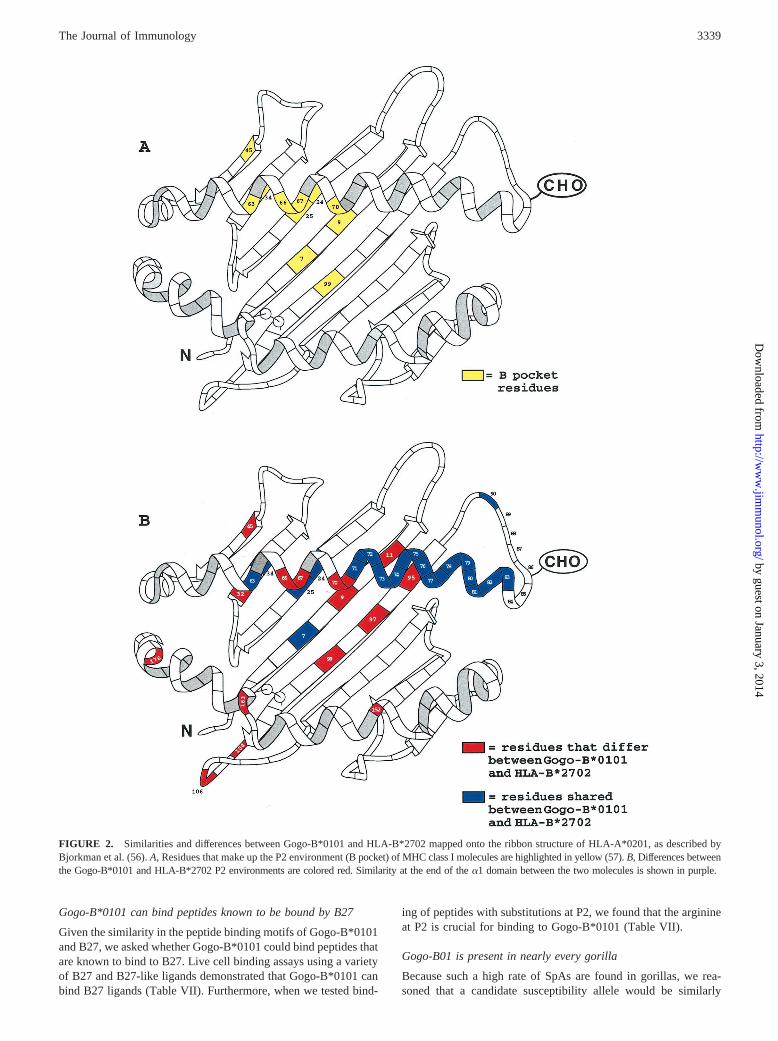
1 and 2). The amino acid sequence in this region is not unique toB27 and also is not conserved between all the subtypes of B27 (seeFig. 1, HLA-B*2705) and so may not be important in diseasepathogenesis. However, this region has been shown to influencepeptide binding specificity (46). Importantly, there are several dif-ferences between B27 and Gogo-B01 in the B pocket of the PBR(Figs. 1 and 2). In particular, the E45 and C67 residues of B27 arereplaced with M45 and S67 (Figs. 1 and 2).
Gorillas that develop SpAs express an MHC class I moleculethat binds peptides with arginine at position 2
We found that Gogo-B01 has only limited similarity to B27, pri-marily at the end of thea1 domain. Additionally, there are keydifferences between the B pocket residues of this molecule andB27. Moreover, previous studies have shown that substitution ofE45 for M45, the substitution present in Gogo-B01, greatly dimin-ished the ability of B27 to bind peptides with arginine at the sec-
ond amino acid of peptides bound by the MHC class I molecule(P2) (13, 14). However, examination of the amino acid residuesthat make up the P2 environment (47) in other HLA molecules thathave been shown to accommodate arginine at the second positionsuggested that Gogo-B*0101 should bind peptides with R2 at leastas well as HLA-Cw*0602 (Table III). Gogo-B01 is expressed inmost animals that develop SpAs. Therefore, we wished to deter-mine what peptides are bound by Gogo-B*0101.
When we defined the peptide binding motif of Gogo-B*0101 bypool sequencing of bound peptides, we found that arginine was thedominant anchor residue at position 2 (Tables IV and V). Sequenc-ing of individual ligands confirmed the presence of R at P2 inpeptides bound by Gogo-B*0101 (Table VI). Individual ligandsequences also revealed a strong preference for tryptophan (W) atthe C terminus, which was not apparent from the pool peptide data.This is consistent with previous work showing that HLA-B15 pep-tide ligands are preferentially anchored at their C termini while
Table I. Clinical signs in gorillas that developed SpAs
Name Sex Diagnosis Reference Description/Comment
Beta F AS 18 Stiffness, awkward, rigid gait; board-like rigidity of the dorsal spine andextreme bilateral contactures of the hips; chronic synovitis of thelarger peripheral joints; swelling and/or contractures involving theshoulders, elbows, wrist, knees, and ankles in a symmetric fashion;advanced sacroiliitis and lumber spondylitis
Kwashi(Beta’s maleoffspring)
M Inflammatorysynovitis
18 Swollen, painful right wrist; local inflammatory synovitis
Husani M ReA 19a Five months followingShigella flexnerienteritis at 6 mo of age,developed weight-bearing lameness of right leg; swelling of rightstifle and tarsus; arthrocentesis revealed acute to subacuteinflammatory joint disease; joint fluid cultures negative; symptomsresolved after 4 wk of treatment; swelling in stifle and lamenessrecurred within 3 days after medication was discontinued; right stiflewas then enlarged and firm; radiography showed increased soft tissueswelling; medication was changed, symptoms resolved and no furtheroccurrences were observed.
Holli F ReA 19a Six weeks following appearance of profuse watery diarrhea (S. flexneritype 2 isolated from feces), animal developed lameness of the rightleg and left arm; increasing severity of depression, lethargy, andlameness leading eventually to non-weight bearing on the left armand right leg; swelling of the left carpus and right tarsus withincreased heat and painful response to palpation of joints; bloodcount showed presence of leukocytosis with regenerative left shift;cytologic examination of the joint fluid showed neutrophilia butbacterial culture was negative; lameness resolved but intermittentjoint tenderness was observed for 3 mo. Nine to 10 mo later, mildlameness of left forelimb noted. No recurrences of lameness noted forthe next 2.5 years.
Harry M ReA D. Neiffer et al.,manuscript inpreparation
Febrile with loose watery stool at 6 and 11 mo of age; lameness withmalodorus, grayish stool at 15 mo of age (Shigellasp. type B—mostlikely flexneri isolated from feces). Placed on course of ciprofloxacin.One month later developed left leg lameness (foot held up with toesclenched); placed on anti-inflammatory; three further episodesbetween ages 21 and 39 mo characterized by lameness of legs and/orarms; on one occasion animal unwilling to move; pain elicited byextension of left elbow; episodes resolved with or without anti-inflammatory therapy; examination at 41 mo of age revealed effussionaround metacarpal-phalangeal joint of 2nd digit; radiographs revealedroughened femoral heads and acetabular margins, and bilateralmetaphyseal thickening of distal radii and ulnas with medialdeviation of radial diaphysis; at 48 mo of age severe left arm andright leg lameness with palpable hot swelling present over right stifle;arthrocentesis resulted in 12 ml viscous yellow fluid with 100%polymorphonuclear cells and no bacteria; blood work showed aleukocytosis with absolute neutrophilia. Radiographs revealedprogressive joint bone/cartilage changes and soft tissue swellingaround right stifle. Placed on long-term sulfasalazine therapy with noclinical signs of over 3 years.
a In Ref. 19, case 1 refers to Holli, case 2 refers to Husani.
3337The Journal of Immunology
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
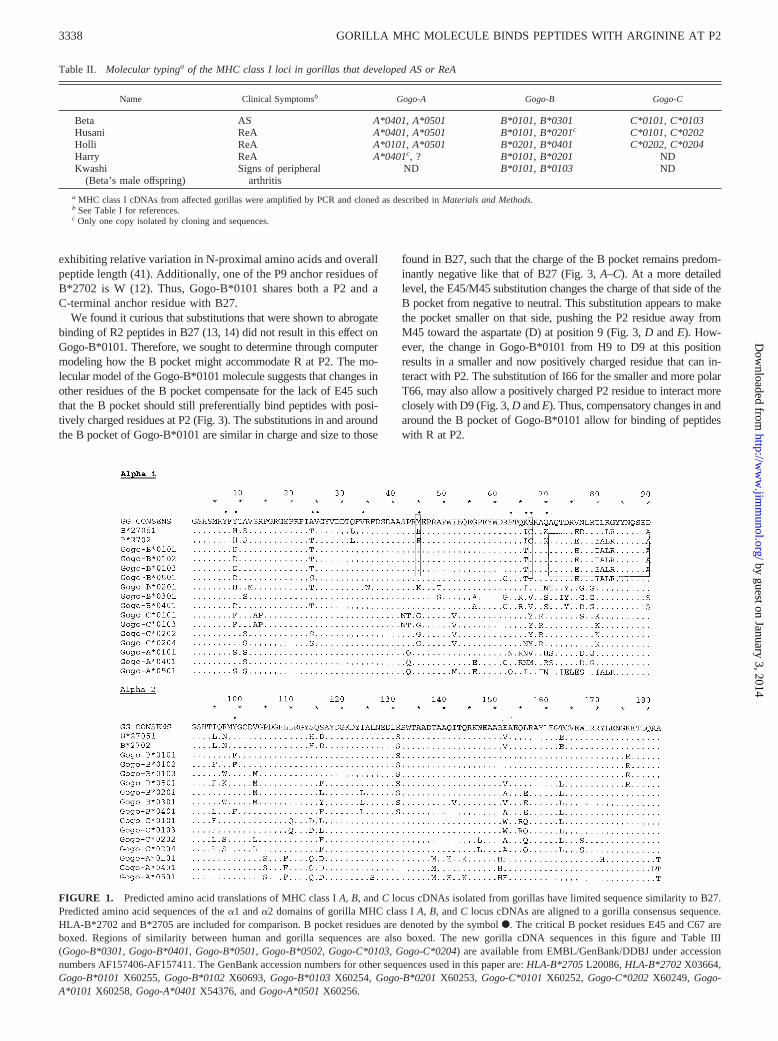
exhibiting relative variation in N-proximal amino acids and overallpeptide length (41). Additionally, one of the P9 anchor residues ofB*2702 is W (12). Thus, Gogo-B*0101 shares both a P2 and aC-terminal anchor residue with B27.
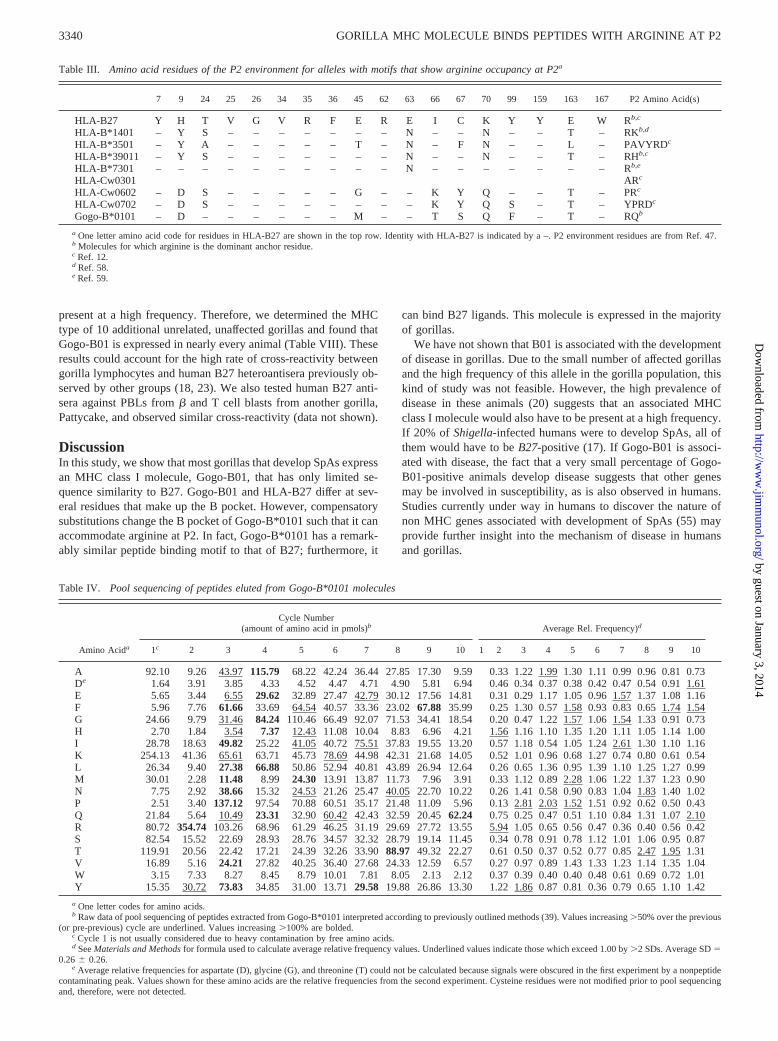
We found it curious that substitutions that were shown to abrogatebinding of R2 peptides in B27 (13, 14) did not result in this effect onGogo-B*0101. Therefore, we sought to determine through computermodeling how the B pocket might accommodate R at P2. The mo-lecular model of the Gogo-B*0101 molecule suggests that changes inother residues of the B pocket compensate for the lack of E45 suchthat the B pocket should still preferentially bind peptides with posi-tively charged residues at P2 (Fig. 3). The substitutions in and aroundthe B pocket of Gogo-B*0101 are similar in charge and size to those
found in B27, such that the charge of the B pocket remains predom-inantly negative like that of B27 (Fig. 3,A–C). At a more detailedlevel, the E45/M45 substitution changes the charge of that side of theB pocket from negative to neutral. This substitution appears to makethe pocket smaller on that side, pushing the P2 residue away fromM45 toward the aspartate (D) at position 9 (Fig. 3,D andE). How-ever, the change in Gogo-B*0101 from H9 to D9 at this positionresults in a smaller and now positively charged residue that can in-teract with P2. The substitution of I66 for the smaller and more polarT66, may also allow a positively charged P2 residue to interact moreclosely with D9 (Fig. 3,D andE). Thus, compensatory changes in andaround the B pocket of Gogo-B*0101 allow for binding of peptideswith R at P2.
FIGURE 1. Predicted amino acid translations of MHC class IA, B, andC locus cDNAs isolated from gorillas have limited sequence similarity to B27.Predicted amino acid sequences of thea1 anda2 domains of gorilla MHC class IA, B, andC locus cDNAs are aligned to a gorilla consensus sequence.HLA-B*2702 and B*2705 are included for comparison. B pocket residues are denoted by the symbolF. The critical B pocket residues E45 and C67 areboxed. Regions of similarity between human and gorilla sequences are also boxed. The new gorilla cDNA sequences in this figure and Table III(Gogo-B*0301, Gogo-B*0401, Gogo-B*0501, Gogo-B*0502, Gogo-C*0103, Gogo-C*0204) are available from EMBL/GenBank/DDBJ under accessionnumbers AF157406-AF157411. The GenBank accession numbers for other sequences used in this paper are:HLA-B*2705L20086,HLA-B*2702X03664,Gogo-B*0101X60255, Gogo-B*0102X60693, Gogo-B*0103X60254, Gogo-B*0201X60253, Gogo-C*0101X60252, Gogo-C*0202X60249, Gogo-A*0101 X60258,Gogo-A*0401X54376, andGogo-A*0501X60256.
Table II. Molecular typinga of the MHC class I loci in gorillas that developed AS or ReA
Name Clinical Symptomsb Gogo-A Gogo-B Gogo-C
Beta AS A*0401, A*0501 B*0101, B*0301 C*0101, C*0103Husani ReA A*0401, A*0501 B*0101, B*0201c C*0101, C*0202Holli ReA A*0101, A*0501 B*0201, B*0401 C*0202, C*0204Harry ReA A*0401c, ? B*0101, B*0201 NDKwashi
(Beta’s male offspring)Signs of peripheral
arthritisND B*0101, B*0103 ND
a MHC class I cDNAs from affected gorillas were amplified by PCR and cloned as described inMaterials and Methods.b See Table I for references.c Only one copy isolated by cloning and sequences.
3338 GORILLA MHC MOLECULE BINDS PEPTIDES WITH ARGININE AT P2
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
Gogo-B*0101 can bind peptides known to be bound by B27
Given the similarity in the peptide binding motifs of Gogo-B*0101and B27, we asked whether Gogo-B*0101 could bind peptides thatare known to bind to B27. Live cell binding assays using a varietyof B27 and B27-like ligands demonstrated that Gogo-B*0101 canbind B27 ligands (Table VII). Furthermore, when we tested bind-
ing of peptides with substitutions at P2, we found that the arginineat P2 is crucial for binding to Gogo-B*0101 (Table VII).
Gogo-B01 is present in nearly every gorilla
Because such a high rate of SpAs are found in gorillas, we rea-soned that a candidate susceptibility allele would be similarly
FIGURE 2. Similarities and differences between Gogo-B*0101 and HLA-B*2702 mapped onto the ribbon structure of HLA-A*0201, as described byBjorkman et al. (56).A, Residues that make up the P2 environment (Bpocket) of MHC class I molecules are highlighted in yellow (57).B, Differences betweenthe Gogo-B*0101 and HLA-B*2702 P2 environments are colored red. Similarity at the end of thea1 domain between the two molecules is shown in purple.
3339The Journal of Immunology
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
present at a high frequency. Therefore, we determined the MHCtype of 10 additional unrelated, unaffected gorillas and found thatGogo-B01 is expressed in nearly every animal (Table VIII). Theseresults could account for the high rate of cross-reactivity betweengorilla lymphocytes and human B27 heteroantisera previously ob-served by other groups (18, 23). We also tested human B27 anti-sera against PBLs fromb and T cell blasts from another gorilla,Pattycake, and observed similar cross-reactivity (data not shown).
DiscussionIn this study, we show that most gorillas that develop SpAs expressan MHC class I molecule, Gogo-B01, that has only limited se-quence similarity to B27. Gogo-B01 and HLA-B27 differ at sev-eral residues that make up the B pocket. However, compensatorysubstitutions change the B pocket of Gogo-B*0101 such that it canaccommodate arginine at P2. In fact, Gogo-B*0101 has a remark-ably similar peptide binding motif to that of B27; furthermore, it
can bind B27 ligands. This molecule is expressed in the majorityof gorillas.
We have not shown that B01 is associated with the developmentof disease in gorillas. Due to the small number of affected gorillasand the high frequency of this allele in the gorilla population, thiskind of study was not feasible. However, the high prevalence ofdisease in these animals (20) suggests that an associated MHCclass I molecule would also have to be present at a high frequency.If 20% of Shigella-infected humans were to develop SpAs, all ofthem would have to beB27-positive (17). If Gogo-B01 is associ-ated with disease, the fact that a very small percentage of Gogo-B01-positive animals develop disease suggests that other genesmay be involved in susceptibility, as is also observed in humans.Studies currently under way in humans to discover the nature ofnon MHC genes associated with development of SpAs (55) mayprovide further insight into the mechanism of disease in humansand gorillas.
Table III. Amino acid residues of the P2 environment for alleles with motifs that show arginine occupancy at P2a
7 9 24 25 26 34 35 36 45 62 63 66 67 70 99 159 163 167 P2 Amino Acid(s)
HLA-B27 Y H T V G V R F E R E I C K Y Y E W Rb,c
HLA-B*1401 – Y S – – – – – – – N – – N – – T – RKb,d
HLA-B*3501 – Y A – – – – – T – N – F N – – L – PAVYRDc
HLA-B*39011 – Y S – – – – – – – N – – N – – T – RHb,c
HLA-B*7301 – – – – – – – – – – N – – – – – – – Rb,e
HLA-Cw0301 ARc
HLA-Cw0602 – D S – – – – – G – – K Y Q – – T – PRc
HLA-Cw0702 – D S – – – – – – – – K Y Q S – T – YPRDc
Gogo-B*0101 – D – – – – – – M – – T S Q F – T – RQb
a One letter amino acid code for residues in HLA-B27 are shown in the top row. Identity with HLA-B27 is indicated by a –. P2 environment residues are from Ref. 47.b Molecules for which arginine is the dominant anchor residue.c Ref. 12.d Ref. 58.e Ref. 59.
Table IV. Pool sequencing of peptides eluted from Gogo-B*0101 molecules
Amino Acida
Cycle Number(amount of amino acid in pmols)b Average Rel. Frequency)d
1c 2 3 4 5 6 7 8 9 10 1 2 3 4 5 6 7 8 9 10
A 92.10 9.26 43.97115.79 68.22 42.24 36.44 27.85 17.30 9.59 0.33 1.22 1.991.30 1.11 0.99 0.96 0.81 0.73De 1.64 3.91 3.85 4.33 4.52 4.47 4.71 4.90 5.81 6.94 0.46 0.34 0.37 0.38 0.42 0.47 0.54 0.91 1.61E 5.65 3.44 6.55 29.62 32.89 27.47 42.7930.12 17.56 14.81 0.31 0.29 1.17 1.05 0.96 1.571.37 1.08 1.16F 5.96 7.76 61.66 33.69 64.54 40.57 33.36 23.0267.88 35.99 0.25 1.30 0.57 1.580.93 0.83 0.65 1.741.54G 24.66 9.79 31.46 84.24 110.46 66.49 92.07 71.53 34.41 18.54 0.20 0.47 1.22 1.571.06 1.54 1.33 0.91 0.73H 2.70 1.84 3.54 7.37 12.43 11.08 10.04 8.83 6.96 4.21 1.561.16 1.10 1.35 1.20 1.11 1.05 1.14 1.00I 28.78 18.63 49.82 25.22 41.05 40.72 75.51 37.83 19.55 13.20 0.57 1.18 0.54 1.05 1.24 2.611.30 1.10 1.16K 254.13 41.36 65.61 63.71 45.73 78.6944.98 42.31 21.68 14.05 0.52 1.01 0.96 0.68 1.27 0.74 0.80 0.61 0.54L 26.34 9.40 27.38 66.88 50.86 52.94 40.81 43.89 26.94 12.64 0.26 0.65 1.36 0.95 1.39 1.10 1.25 1.27 0.99M 30.01 2.28 11.48 8.99 24.30 13.91 13.87 11.73 7.96 3.91 0.33 1.12 0.89 2.281.06 1.22 1.37 1.23 0.90N 7.75 2.92 38.66 15.32 24.53 21.26 25.47 40.0522.70 10.22 0.26 1.41 0.58 0.90 0.83 1.04 1.831.40 1.02P 2.51 3.40 137.12 97.54 70.88 60.51 35.17 21.48 11.09 5.96 0.13 2.812.03 1.52 1.51 0.92 0.62 0.50 0.43Q 21.84 5.64 10.49 23.31 32.90 60.42 42.43 32.59 20.4562.24 0.75 0.25 0.47 0.51 1.10 0.84 1.31 1.07 2.10R 80.72 354.74 103.26 68.96 61.29 46.25 31.19 29.69 27.72 13.55 5.941.05 0.65 0.56 0.47 0.36 0.40 0.56 0.42S 82.54 15.52 22.69 28.93 28.76 34.57 32.32 28.79 19.14 11.45 0.34 0.78 0.91 0.78 1.12 1.01 1.06 0.95 0.87T 119.91 20.56 22.42 17.21 24.39 32.26 33.9088.97 49.32 22.27 0.61 0.50 0.37 0.52 0.77 0.85 2.471.95 1.31V 16.89 5.16 24.21 27.82 40.25 36.40 27.68 24.33 12.59 6.57 0.27 0.97 0.89 1.43 1.33 1.23 1.14 1.35 1.04W 3.15 7.33 8.27 8.45 8.79 10.01 7.81 8.05 2.13 2.12 0.37 0.39 0.40 0.40 0.48 0.61 0.69 0.72 1.01Y 15.35 30.72 73.83 34.85 31.00 13.7129.58 19.88 26.86 13.30 1.22 1.860.87 0.81 0.36 0.79 0.65 1.10 1.42
a One letter codes for amino acids.b Raw data of pool sequencing of peptides extracted from Gogo-B*0101 interpreted according to previously outlined methods (39). Values increasing.50% over the previous
(or pre-previous) cycle are underlined. Values increasing.100% are bolded.c Cycle 1 is not usually considered due to heavy contamination by free amino acids.d SeeMaterials and Methodsfor formula used to calculate average relative frequency values. Underlined values indicate those which exceed 1.00 by.2 SDs. Average SD5
0.266 0.26.e Average relative frequencies for aspartate (D), glycine (G), and threonine (T) could not be calculated because signals were obscured in the first experiment by a nonpeptide
contaminating peak. Values shown for these amino acids are the relative frequencies from the second experiment. Cysteine residues were not modified prior to pool sequencingand, therefore, were not detected.
3340 GORILLA MHC MOLECULE BINDS PEPTIDES WITH ARGININE AT P2
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

The question of whether HLA-B27 acts as an Ag-presentingmolecule in the mechanism of SpAs remains a central and, as yet,unanswered question. The peptide binding capabilities of the var-ious B27 subtypes either associated with disease susceptibility ornot associated with disease susceptibility have been described (Re-
viewed in Refs. 3 and 48). Although the results presented here donot show that Gogo-B01 is associated with susceptibility to dis-ease, they are consistent with recent data from transgenic rats (8)that suggest that the peptides bound by the B27 molecule play arole in the mechanism of disease.
Table V. Peptide binding motif of Gogo-B*0101
Residue
1 2 3 4 5 6 7 8 9 10
C Terminusa
7/8/9/10
Gogo-B*0101Dominantb R P E W
NFMAVG
Stronga L G M Y T F QI AY L
QH
Weaka Y H F Q I NE H K EQ IK N
HLA-B*2702c
Anchor residueR F
YILW
Other preferred residues K F G I I Y KL P K V L VXd K E Y V D
D V R T EE M D F RQ T HT ES Q
a C terminus preference was evident from sequence of individual ligands. See Table VI.b Determination of peptide binding motif according to previously established methods (39–41). Dominant:.3.5-fold increase over the previous cycle; anchor residue is
shown in bold and is applied if a position reveals a strong signal for only one residue as is the case with R at P2 (see Table IV); Strong:.2-fold increase over the previous cycle;Weak: 1.5- to 2-fold increase over the previous cycle.
c Motif for HLA-B*2702 is shown for comparison (12).d X indicates that several additional residues were detected at lower levels.
Table VI. Peptides bound by Gogo-B*0101
Source
Position
1 2 3 4 5 6 7 8 9 10
No match S D T C S M D T WHuman uridine nucleotide receptor, gi 1117913 P E D S S C R WNo match P V G P G S M P T WHumand-6 fatty acid desaturase, gi 4406528 I R F L E S H WHuman serine/threonine protein kinase pLK-1
506–514, gi423046A R L P Y L R T W
No exact match R P M A F Y S WNo exact match S D A Q P A L/I T S WHuman tubulina-4 chain, AA722099 S R C H L G AChromosome 7, HELA cDNA library
AA078075.1K R C S P Y
Similar but not exact match to serine threoninekinase
T R F E G Y F
No exact match W P G P A V S Z WHomo sapienshtgs, working draft AC007683.2 A S L G A K W
3341The Journal of Immunology
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

Interestingly, a strong C-terminal W residue was revealed in thesequence of individual ligands eluted from Gogo-B*0101. Thisobservation is consistent with results from peptides sequencedfrom HLA-B15 allotypes that showed that a S116 residue correlatedwith a strong C-terminal anchor residue (41, 49). Although the motifof HLA-B*2702 demonstrates a dominant W at P9 (11) and this isalso a preferred residue in the motif of HLA-B*2703 (50, 51), few
peptides or T cell epitopes identified to date contain this C-terminalresidue (12, 52). Further studies are necessary to elucidate a possiblerole in disease pathogenesis for peptides containing a C-terminal W.
The role of the free C67 in the B27 B pocket in disease mechanismhas been under investigation. Studies suggest that the free C67 can bemodified due to its chemical reactivity (53), and homocysteine-mod-ified B27 can be recognized by homocysteine-specific CTL in vivo
FIGURE 3. Molecular model of Gogo-B*0101compared with that of HLA-B27. InA–C, the top viewof the PBR is shown with negative charge in red, pos-itive charge in blue. InC, substitutions that result in adifference in charge or size are shown in pink, changesthat remain similar in charge and size are shown inlight blue. In D and E, the B pocket environment isshown with the peptide and it’s P2 side chain in pink.
Table VII. Capacity of Gogo-B*0101-positive cell lines to bind a panel of peptidesa
Peptide Origin Sequence
IC50 (nM)
ReferenceBeta721.221-
Gogo-B*0101
153.07 SIV (B27-like) IRYPKTFGW 34 54 60155.01 SIV (B27-like) IRFPKTFGW 12 30 60330-1 Human Fau protein (B27) RRFVNVVPTF 64 196 11330-3 HBNPBb-subunit 35–43 (B27) SRDKTIIMW 31 128 11405.1 Human actin (B27) KRGILTLKY 316 668 11405.2 Unknown (B27) GRFKLIVLY 1572 3091 11405.5 HIV env 584–592 (B27) ERYLKDQQL 7972 14579 61405.6 Flu Histone H3.3 (B27) RRYQKSTEL 26 53 105423.3 HFPS 191–199 (B27) KRYKSIVKY 36 100 115423.4 Human cytochrome C oxidase (B27) KRKKAYADF 3267 6115 11155.01 SIV (B27-like) IRFPKTFGW 12 30 60Chiron 50 SIV with substitution IKFPKTFGW 1836b 3274Chiron 51 SIV with substitution ILFPKTFGW 423 998Chiron 52 SIV with substitution IFFPKTFGW 2108 3174Chiron 53 SIV with substitution INFPKTFGW 17064 23443Chiron 54 SIV with substitution IDFPKTFGW 65757 107383
a Experiments performed with radiolabelled 153.07 IRYPKTFGW.b Numbers in bold indicate three times or greater decrease in binding capacity.
3342 GORILLA MHC MOLECULE BINDS PEPTIDES WITH ARGININE AT P2
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

(15). Furthermore, recent studies suggest that B27 forms nonconven-tional structures dependent on C67 (7). However, C67 does not appearto be required for SpA-like disease in transgenic rats because ratsmade transgenic with a S67 mutant of B27 developed a disease phe-notype similar to that of the homozygous wild-type B27 rats, althoughwith a lower prevalence of arthritis (54). Gogo-B01, expressed inmost gorillas that develop SpAs, also has a S67 in place of the C67.Thus, like transgenic rats, gorillas develop arthritis in the absence ofan MHC class I molecule that has a free cysteine. The recent obser-vation that B27 can form a C67-dependentb2m-free heavy chainhomodimer structure (7) has led to the proposition of an alternativetheory for the role of B27 in SpAs involving aberrant B27 heavy chaindimers (3).
In conclusion, here we present data showing the remarkablesimilarity between SpAs in gorillas andB27-associated SpAs inhumans. It is unclear from this study whether Gogo-B01 is asso-ciated with disease susceptibility in gorillas. However, given thesimilar peptide binding specificities of B27 and B01, the high per-centage of B01-positive animals in the gorilla population, and thehigh rate of disease in this species, it is tempting to speculate as tothe role in disease susceptibility of this molecule and the peptidesit binds.
AcknowledgmentsWe thank Kenneth W. Jackson of the William K. Warren Medical Re-search Institute, University of Oklahoma Health Sciences Center for as-sistance with the HPLC purification and Edman sequencing, and BobBecker of WRPRC for his invaluable assistance with computer graphics.We also thank Drs. Joel Taurog and Jose´ Lopez de Castro for helpfuldiscussions and critical review of initial versions of this manuscript.
References1. Benjamin, R., and P. Parham. 1990. Guilt by association: HLA-B27 and anky-
losing spondylitis.Immunol. Today 11:137.2. Burmester, G. R., A. Daser, T. Kabradt, A. Krause, N. A. Mitchison, J. Sieper,
and N. Wolf. 1995. Immunology of reactive arthritides.Annu. Rev. Immunol.13:229.
3. Allen, R. L., P. Bowness, and A. McMichael. 1999. The role of HLA-B27 inspondyloarthritis.Immunogenetics 50:220.
4. Khare, S. D., H. S. Luthra, and C. S. David. 1995. Spontaneous inflammatoryarthritis in HLA-B27 transgenic mice lackingb2-microglobulin: a model of hu-man spondyloarthropathies.J. Exp. Med. 182:1153.
5. Khare, S. D., J. Hansen, H. S. Luthra, and C. S. David. 1996. HLA-B27 heavychains contribute to spontaneous inflammatory disease in B27/humanb2-micro-glogulin (b2m) double transgenic mice with disrupted mouseb2m. J. Clin. Invest.98:2746.
6. Khare, S. D., M. J. Bull, J. Hanson, H. S. Luthra, and C. S. David. 1998. Spon-taneous inflammatory disease in HLA-B27 transgenic mice is independent ofMHC class II molecules: a direct role for B27 heavy chains and not B27-derivedpeptides.J. Immunol. 160:101.
7. Allen, R. L., C. A. O’Callaghan, A. J. McMichael, and P. Bowness. 1999. Cuttingedge: HLA-B27 can form a novelb2-microglobulin-free heavy chain homodimerstructure.J. Immunol. 162:5045.
8. Zhou, M., A. Sayad, W. A. Simmons, R. C. Jones, S. D. Maika, N. Satumtira,M. L. Dorris, S. J. Gaskell, R. S. Bordoli, R. B. Sartor, et al. 1998. The specificityof peptides bound to human histocompatibility leukocyte antigen (HLA)-B27influences the prevalence of arthritis in HLA-B27 transgenic rats.J. Exp. Med.188:877.
9. Guo, H. C., D. R. Madden, M. L. Silver, T. S. Jardetzky, J. C. Gorga,J. L. Strominger, and D. C. Wiley. 1993. Comparison of the P2 specificity pocketin three human histocompatibility antigens: HLA-A*6801, HLA-A*0201, andHLA-B*2705. Proc. Natl. Acad. Sci. USA 90:8053.
10. Jardetzky, T. S., W. S. Lane, R. A. Robinson, D. R. Madden, and D. C. Wiley.1991. Identification of self peptides bound to purified HLA-B27.Nature 353:326.
11. Rotzschke, O., K. Falk, S. Stevanovic, V. Gnau, G. N. Jung, and H.-G. Ram-mensee. 1994. Dominant aromatic/aliphatic C-terminal anchor in HLA-B*2702and B*2705 peptide motifs.Immunogenetics 39:74.
12. Rammensee, H. G., T. Friede, and S. Stevanoviic. 1995. MHC ligands and pep-tide motifs: first listing.Immunogenetics. 41:178.
13. Buxton, S. E., R. J. Benjamin, C. Clayberger, P. Parham, and A. M. Krensky.1992. Anchoring pockets in human histocompatibility complex leukocyte antigen(HLA) class I molecules: analysis of the conserved B (“45”) pocket of HLA-B27.J. Exp. Med. 175:809.
14. Rojo, S., F. Garcıa, J. A. Villadangos, and J. A. L. de Castro. 1993. Changes inthe repertoire of peptides bound to HLA-B27 subtypes and to site-specific mu-tants inside and outside pocket B.J. Exp. Med. 177:613.
15. Gao, X. M., P. Wordsworth, A. J. McMichael, M. M. Kyaw, M. Seifert, D. Rees,and G. Dougan. 1996. Homocysteine modification of HLA antigens and its im-munological consequences.Eur. J. Immunol. 26:1443.
16. Khan, M. A. 1992. An overview of clinical spectrum and heterogeneity of spon-dyloarthropathies.Rheum. Dis. Clin. North Am. 18:1.
17. Keat, A. 1983. Reiter’s syndrome and reactive arthritis in perspective.N. Engl.J. Med. 309:1606.
18. Adams, R. F., G. S. J. Flinn, and M. Douglas. 1987. Ankylosing spondylitis in anonhuman primate: a monkey tale.Arthritis Rheum. 30:956.
19. Raphael, B. L., P. P. Calle, N. Haramati, D. I. Watkins, M. D. Stetter, andR. A. Cook. 1995. Reactive arthritis subsequent toShigella flexnerienteritis intwo juvenile lowland gorillas (gorilla gorilla gorilla).J. Zoo Wildl. Med. 26:132.
20. Rothschild, B., and R. J. Woods. 1989. Spondyloarthropathy in gorillas.Semin.Arthritis Rheum. 18:267.
21. Ruvolo, M., T. R. Disotell, M. W. Allard, W. M. Brown, and R. L. Honeycutt.1991. Resolution of the African hominoid trichotomy by use of a mitochondrialgene sequence.Proc. Natl. Acad. Sci. USA 88:1570.
22. Sibley, C. G., and J. E. Ahlquist. 1984. The phylogeny of the hominoid primates,as indicated by DNA-DNA hybridization.J. Mol. Evol. 20:2.
23. Lawlor, D. A., E. Warren, P. Taylor, and P. Parham. 1991. Gorilla class I majorhistocompatibility complex alleles: comparison to human and chimpanzee classI. J. Exp. Med. 174:1491.
24. Watkins, D. I., Z. W. Chen, T. L. Garber, A. L. Hughes, and N. L. Letvin. 1991.Segmental exchange between MHC class I genes in a higher primate: recombi-nation in the gorilla between the ancestor of a human non-functional gene and anA locus gene.Immunogenetics 34:185.
25. Miller, G., and M. Lipman. 1973. Release of infectious Epstein-Barr virus bytransformed marmoset leukocytes.Proc. Natl. Acad. Sci. USA 70:190.
26. Lawlor, D. A., E. Warren, F. E. Ward, and P. Parham. 1990. Comparison of classI MHC alleles in humans and apes.Immunol. Rev. 113:147.
27. Klein, J., R. E. Bontrop, R. Dawkins, H. A. Erlich, U. B. Gyllensten, E. R. Heise,E. R. Jones, P. Parham, E. K. Wakeland, and D. I. Watkins. 1990. Nomenclaturefor the major histocompatibility complexes of different species: a proposal.Im-munogenetics 31:217.
28. Boyson, J. E., C. Shufflebotham, L. F. Cadavid, J. A. Urvater, L. A. Knapp,A. L. Hughes, and D. I. Watkins. 1996. The MHC class I genes of the rhesusmonkey: different evolutionary histories of MHC class I and II genes in primates.J. Immunol. 156:4656.
29. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman,J. A. Smith, and D. Struhl. 1992.Current Protocols in Molecular Biology. Vol.2. K. Janssen, ed., Wiley, New York.
30. Madden, D. R., J. C. Gorga, J. L. Strominger, and D. C. Wiley. 1991. Thestructure of HLA-B27 reveals nonamer self-peptides bound in an extended con-formation.Nature 353:321.
31. Jones, T. A., J. Y. Zou, S. W. Cowan, and M. Kjeldgaard. 1991. Improvedmethods for binding protein models in electron density maps and the location oferrors in these models.Acta Crystallogr. A47:110.
32. Ponder, J. W., and F. M. Richards. 1987. Tertiary templates for proteins: use ofpacking criteria in the enumeration of allowed sequences for different structuralclasses.J. Mol. Biol. 193:775.
33. X-PLOR. A System for X-ray Crystallography and NMR 3.1. Yale UniversityPress, New Haven, CT.
Table VIII. Eleven of 13 unrelated gorillas express Gogo-B01a,b
Name (ancestry) Gogo-B
Beta (wild born) B*0101, B*0301Husani (Pattycake3 Barney) B*0101, B*0201c
Hollid (Huerfanita3 Kongo) B*0201, B*0401Harry (Kweli 3 Babec) B*0101, B*0201Kwashi (Beta3 Carlos) B*0101, B*0103Choomba (wild born) B*0103, B*0501Katoomba (wild born) B*0101, B*0401Rann (wild born) B*0103, B*0401Shamba (wild born) B*0301, B*0401Bangae (wild born) B*0101, B*0103Calabare B*0102, B*0103Inaki (wild born) B*0101, B*0201Oko (wild born) B*0201f
Paki (wild born) B*0101, B*0502Ozoum (wild born) B*0101, or B*0103g
a MHC B locus alleles in bold indicates unrelated animal positive for aB01subtype.
b Gorillas were typed as in Table I.c Only one copy isolated by cloning and sequencing.d Holli is the half aunt of Husani.e Banga, Calabar, Inaki, and Oko were typed by Lawlor et al. (23).f Oko appears to be homozygous at theB locus.g Ozoum was typed indirectly by typing Rok, Banga’s offspring with Ozoum.
3343The Journal of Immunology
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

34. Shimizu, Y., and R. DeMars. 1989. Production of human cells expressing indi-vidual transferred HLA-A, -B, -C genes using an HLA-A, -B, -C null human cellline. J. Immunol. 142:3320.
35. Prilliman, K., M. Lindsey, Y. Zuo, K. Jackson, Y. Zhang, and W. Hildebrand.1997. Large-scale production of class I bound peptides: assigning a signature toHLA-B*1501. Immunogenetics 45:379.
36. Allen, T. M., J. Sidney, M.-F. del Guercio, R. L. Glickman, G. L. Lensmeyer,D. A. Wiebe, R. DeMars, C. D. Pauza, R. P. Johnson, A. Sette, and D. I. Watkins.1998. Characterization of the peptide-binding motif of a rhesus MHC class Imolecule (Mamu-A*01) that binds an immunodominant CTL epitope from SIV.J. Immunol. 160:6062.
37. Slingluff, C. L., A. L. Cox, R. A. Henderson, D. F. Hunt, and V. H. Engelhard.1993. Recognition of human melanoma cells hy HLA-A2.1-restricted cytotoxic Tlymphocytes is mediated by at least six shared peptide epitopes.J. Immunol.150:2955.
38. Kubo, R. T., A. Sette, H. M. Grey, E. Appella, K. Sakaguchi, N. Z. Zhu,D. Arnott, N. Sherman, J. Shabanowitz, H. Michel, et al. 1994. Definition ofspecific peptide motifs for four major HLA-A alleles.J. Immunol. 152:3913.
39. Falk, K., O. Rotzeschke, S. Stevanovic, G. N. Jung, and H.-G. Rammensee. 1991.Allele-specific motifs revealed by sequencing of self-peptides eluted from MHCmolecules.Nature 351:290.
40. Barber, L. D., B. Gillece-Castro, L. Percival, X. Li, C. Clayberger, andP. Parham. 1995. Overlap in the repertoires of peptides bound in vivo by a groupof related class I HLA-B allotypes.Curr. Biol. 5:179.
41. Prilliman, K. R., K. W. Jackson, M. Lindsey, J. Wang, D. Crawford, andW. H. Hildebrand. 1999. HLA-B15 peptide ligands are preferentially anchored attheir C termini.J. Immunol. 162:7277.
42. Mann, M., and M. Wilm. 1994. Error-tolerant identification of peptides in se-quence databases by peptide sequence tags.Anal. Chem. 66:4390.
43. Ruppert, J., J. Sidney, E. Celis, R. T. Kubo, H. M. Grey, and A. Sette. 1993.Prominent role of secondary anchor residues in peptide binding to HLA-A2.1molecules.Cell 74:929.
44. Sidney, J., S. Southwood, C. Oseroff, M. F. del Guercio, A. Sette, andH. M. Grey. 1998. Measurement of MHC/peptide interactions by gel filtration.Curr. Protocols Immunol. 18:3.1.
45. del Guercio, M. F., J. Sidney, G. Hermanson, C. Perez, H. M. Grey, R. T. Kubo,and A. Sette. 1995. Binding of a peptide antigen to multiple HLA alleles allowsdefinition of an A2-like supertype.J. Immunol. 154:685.
46. Barber, L. D., L. Percival, K. L. Arnett, J. E. Gumperz, L. Chen, and P. Parham.1997. Polymorphism in thea(1) helix of the Hla-B heavy chain can have anoverriding influence on peptide-binding specificity.J. Immunol. 158:1660.
47. Chelvanayagam, G. 1996. A roadmap for HLA-A, HLA-B, and HLA-C peptidebinding specificities.Immunogenetics 45:15.
48. Lopez de Castro, J. A. 1998. The pathogenetic role of HLA-B27 in chronicarthritis.Curr. Opin. Immunol. 10:59.
49. Prilliman, K. R., D. Crawford, H. D. Hickman, K. W. Jackson, J. Wang, andW. H. Hildebrand. 1999.a-2 domain polymorphism and HLA class I peptideloading.Tissue Antigens 54:450.
50. Griffin, T. A., J. Yuan, T. Friede, S. Stevanovic, K. Ariyoshi,S. L. Rowland-Jones, H. G. Rammensee, and R. A. Colbert. 1997. Naturallyoccurring A pocket polymorphism in HLA-B*2703 increases the dependence onan accessory anchor residue at P1 for optimal binding of nonamer peptides.J. Im-munol. 159:4887.
51. Boisgerault, F., V. Tieng, M. C. Stolzenberg, N. Dulphy, I. Khalil, R. Tamouza,D. Charron, and A. Toubert. 1996. Differences in endogenous peptides presentedby HLA-B*2705 and B*2703 allelic variants: implications for susceptibility tospondylarthropathies.J. Clin. Invest. 98:2764.
52. Rammensee, H., J. Bachmann, N. P. Emmerich, O. A. Bachor, and S. Stevanovic.1999. SYFPEITHI: database for MHC ligands and peptide motifs.Immunoge-netics 50:213.
53. Whelan, M. A., and J. R. Archer. 1993. Chemical reactivity of an HLA-B27 thiolgroup.Eur. J. Immunol. 23:3278.
54. Taurog, J. D., S. D. Maika, N. Satumtira, M. L. Dorris, I. L. McLean,H. Yanagisawa, A. Sayad, A. J. Stagg, G. M. Fox, A. Le O’Brien, et al. 1999.Inflammatory disease in HLA-B27 transgenic rats.Immunol. Rev. 169:209.
55. Brown, M., and P. Wordsworth. 1997. Predisposing factors to spondyloarthropa-thies.Curr. Opin. Rheumatol. 9:308.
56. Bjorkman, P. J., M. A. Saper, B. Samraoui, W. S. Bennett, J. L. Strominger, andD. C. Wiley. 1987. Structure of the human class I histocompatibility antigen,HLA-A2. Nature 329:506.
57. Saper, M. A., P. J. Bjorkman, and D. C. Wiley. 1991. Refined structure of thehuman histocompatibility antigen HLA-A2 at 2.6 Å resolution.J. Mol. Biol.219:277.
58. DiBrino, M., K. C. Parker, D. H. Margulies, J. Shiloach, R. V. Turner,W. E. Biddison, and J. E. Coligan. 1994. The HLA-B14 peptide binding site canaccommodate peptides with different combination of anchor residues.J. Biol.Chem. 269:32426.
59. Barber, L. D., L. Percival, and P. Parham. 1996. Characterization of the peptide-binding specificity of HLA-B*73.Tissue Antigens 47:472.
60. Evans, D. T., D. H. O’Connor, P. Jing, J. L. Dzuris, J. Sidney, J. da Silva,T. M. Allen, H. Horton, J. E. Venham, R. A. Rudersdorf, et al. 1999. Virus-specific cytotoxic T-lymphocyte responses select for amino-acid variation in sim-ian immunodeficiency virus Env and Nef.Nat. Med. 5:1270.
61. Shankar, P., J. A. Fabry, D. M. Fong, and J. Lieberman. 1996. Three regions ofHIV-1 gp160 contain clusters of immunodominant CTL epitopes.Immunol. Lett.52:23.
3344 GORILLA MHC MOLECULE BINDS PEPTIDES WITH ARGININE AT P2
by guest on January 3, 2014http://w
ww
.jimm
unol.org/D
ownloaded from




















