
Patterns of macroalgal diversity, community composition and long-term changes along the Swedish west...
20
Hydrobiologia 459: 83–102, 2001. © 2001 Kluwer Academic Publishers. Printed in the Netherlands. 83 Patterns of macroalgal diversity, community composition and long-term changes along the Swedish west coast M. Peders´ en 1 & P. Snoeijs 2∗ 1 Department of Botany, Stockholm University, SE-10691 Stockholm, Sweden 2 Department of Plant Ecology, Evolutionary Biology Centre, Uppsala University, Villavägen 14, SE-75236 Uppsala, Sweden Tel: +18-471-28-85. Fax: +18-553-419. E-mail: [email protected] ( ∗ Author for correspondence) Received 15 November 2000; in revised form 21 May 2001; accepted 1 June 2001 Key words: algae, ecology, depth distributions, diversity, long-term changes, Kattegat, Skagerrak Abstract This study analyses the complicated patterns of vertical distribution of the macroalgal vegetation in an area where brackish and marine waters meet and mix. Variables used to record vegetation characteristics are algal cover, spe- cies composition and diversity. The data set includes 64 diving profiles, all from sites exposed to wave action, along a ca. 260 km long coastline. The profiles belong to four categories: coastal sites in the Skagerrak (more marine), coastal sites in the Kattegat (more brackish), coastal sites in the Kattegat after a toxic phytoplankton bloom, and submerged offshore stone reefs in the Kattegat. The highest species diversity was found at the reefs, which are not affected directly by land runoff. At the reefs the 18 most common perennial species penetrate 2–11 m (on average 5.5 m) deeper than at the coastal sites. The virtual absence of sedimentation, and thus the availability of substratum, at the reefs may explain the differences so that the lower limit for the algae is determined by light penetration or by recruitment problems caused by strong currents at the reefs, whereas sedimentation limits the settlement of algae in coastal sites. Ordination analysis based on species composition reveals that the major environmental gradients structuring the algal vegetation in the Kattegat and the Skagerrak are salinity and water depth. The large data set of this study made it possible to quantify the downward dislocation of Atlantic intertidal species to the sublittoral along the Swedish west coast. For example, the mean upper limit of Corallina officinalis is 2 m in the Skagerrak but 12.5 m in the Kattegat and the mean occurrence interval of Fucus serratus is 0.9–2.7 m in the Skagerrak, but 1.1–6.3 m in the Kattegat. This downward dislocation is suggested to be the result of decreased competition when species successively disappear with lower salinity. Comparisons of the present study’s results with those of previous investigations show that eight common red algal species have moved upwards compared to the situation before the large-scale eutrophication started in the 1960s, e.g. Cystoclonium purpureum and Polysiphonia elongata by ca. 8 m, Phycodrys rubens and Delesseria sanguinea by ca. 5 m. A toxic phytoplankton bloom affected macroalgal community composition on the whole only slightly, but it had a negative effect on algal cover and species richness below a water depth of ca. 5 m, the algae were visibly damaged and the lower vegetation limit temporarily moved upwards. Introduction This paper presents the results from a large-scale field study of the macroalgal vegetation in the Skagerrak and Kattegat in 1988–1990. The study was carried out to be able to compare the littoral vegetation with earlier records to understand the effects of large-scale environmental changes in the ecosystem. It also docu- ments the short-term effects of a toxic phytoplankton bloom that struck the area in spring 1988. Only wave- exposed sites in open waters were included in the study because these will reflect large-scale changes and not local pollution. This study further makes com- parisons between the more marine Skagerrak and the
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Patterns of macroalgal diversity, community composition and long-term changes along the Swedish west...

Hydrobiologia 459: 83–102, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
83
Patterns of macroalgal diversity, community composition andlong-term changes along the Swedish west coast
M. Pedersen1 & P. Snoeijs2∗1Department of Botany, Stockholm University, SE-10691 Stockholm, Sweden2Department of Plant Ecology, Evolutionary Biology Centre, Uppsala University, Villavägen 14,SE-75236 Uppsala, SwedenTel: +18-471-28-85. Fax: +18-553-419. E-mail: [email protected] (∗Author for correspondence)
Received 15 November 2000; in revised form 21 May 2001; accepted 1 June 2001
Key words: algae, ecology, depth distributions, diversity, long-term changes, Kattegat, Skagerrak
Abstract
This study analyses the complicated patterns of vertical distribution of the macroalgal vegetation in an area wherebrackish and marine waters meet and mix. Variables used to record vegetation characteristics are algal cover, spe-cies composition and diversity. The data set includes 64 diving profiles, all from sites exposed to wave action, alonga ca. 260 km long coastline. The profiles belong to four categories: coastal sites in the Skagerrak (more marine),coastal sites in the Kattegat (more brackish), coastal sites in the Kattegat after a toxic phytoplankton bloom, andsubmerged offshore stone reefs in the Kattegat. The highest species diversity was found at the reefs, which are notaffected directly by land runoff. At the reefs the 18 most common perennial species penetrate 2–11 m (on average5.5 m) deeper than at the coastal sites. The virtual absence of sedimentation, and thus the availability of substratum,at the reefs may explain the differences so that the lower limit for the algae is determined by light penetration or byrecruitment problems caused by strong currents at the reefs, whereas sedimentation limits the settlement of algaein coastal sites. Ordination analysis based on species composition reveals that the major environmental gradientsstructuring the algal vegetation in the Kattegat and the Skagerrak are salinity and water depth. The large data setof this study made it possible to quantify the downward dislocation of Atlantic intertidal species to the sublittoralalong the Swedish west coast. For example, the mean upper limit of Corallina officinalis is 2 m in the Skagerrakbut 12.5 m in the Kattegat and the mean occurrence interval of Fucus serratus is 0.9–2.7 m in the Skagerrak, but1.1–6.3 m in the Kattegat. This downward dislocation is suggested to be the result of decreased competition whenspecies successively disappear with lower salinity. Comparisons of the present study’s results with those of previousinvestigations show that eight common red algal species have moved upwards compared to the situation before thelarge-scale eutrophication started in the 1960s, e.g. Cystoclonium purpureum and Polysiphonia elongata by ca.8 m, Phycodrys rubens and Delesseria sanguinea by ca. 5 m. A toxic phytoplankton bloom affected macroalgalcommunity composition on the whole only slightly, but it had a negative effect on algal cover and species richnessbelow a water depth of ca. 5 m, the algae were visibly damaged and the lower vegetation limit temporarily movedupwards.
Introduction
This paper presents the results from a large-scale fieldstudy of the macroalgal vegetation in the Skagerrakand Kattegat in 1988–1990. The study was carriedout to be able to compare the littoral vegetation withearlier records to understand the effects of large-scale
environmental changes in the ecosystem. It also docu-ments the short-term effects of a toxic phytoplanktonbloom that struck the area in spring 1988. Only wave-exposed sites in open waters were included in thestudy because these will reflect large-scale changesand not local pollution. This study further makes com-parisons between the more marine Skagerrak and the

84
more brackish Kattegat and includes the major sub-merged offshore stone reefs (‘fishing-banks’) in theKattegat. The latter are previously little known hotspots of algal diversity in the area. Our study willalso serve as a comprehensive baseline study for futureinvestigations of the macroalgal vegetation of the area.
Previous large-scale phycological studies in thisarea were predominantly floristically inclined (Kylin,1907, 1944, 1947, 1949; Rosenvinge, 1909–1931;Rosenvinge & Lund, 1941–1947; Lund 1950). Wærn(1952, 1965) described ecological communities andprocesses in a wider perspective, but unfortunatelyalso without a numerical basis for documenting thesecommunities and processes, except for single well-described diving profiles, some of which recently havebeen re-investigated (e.g. Johansson et al., 1998). Fur-ther recent investigations were also restricted to singleareas or localities, e.g. at Kullen in the southern Kat-tegat by von Wachenfeldt (1975) and Kornfeldt (1981,1984), and at Lysekil in the Skagerrak by Lundälv etal. (1986) and Svane & Gröndahl (1988). Contrarily,our study covers a 260 km long coastline and thus themajor part of the Skagerrak and Kattegat.
In the early days, algal collections were mainlymade by dredging (instead of diving), and such datamust be interpreted with caution because of interferingloose-lying algae and other anomalies, see the discus-sions in Michanek (1967), Nielsen (1991) and Nielsen& Dahl (1992a, b). Thus, no reliable data are availablefor direct comparative numerical vegetation analysesfor the Kattegat and Skagerrak area as a whole tocompare the situation before and after the extensiveeutrophication that started in the 1960s (Rosenberg etal., 1990; Richardsson & Heilman, 1995). However,in the present study, we were able to make comparis-ons for the depth distributions of a number of commonspecies for which the occurrence (but not abundance)with respect to water depth was well described byKylin in the 1940s.
The Skagerrak and Kattegat together form thenorthern part of the transitional zone between the mar-ine North Sea and the brackish Baltic Sea. The Balticcurrent (outflow from the Baltic Sea) flows at the sur-face and its vertical extension strongly depends onwind direction. A halocline separates this surface layerfrom the high-salinity North Sea water (32–34 psu)beneath it. In this highly dynamic area, it is impossibleto relate the distribution of macroalgae, which havebeen growing for months or years, to measured en-vironmental factors such as temperature, irradiance orsalinity. Constantly differing weather conditions affect
flow directions and mixing processes. There are, how-ever, well-defined differences in salinity between thetwo basins (Fonselius, 1995): surface-water salinitiesare 15–25 psu in the Kattegat and 20–30 psu in theSkagerrak, the halocline is found at 20–30 m depthin the southern Kattegat and at 10–20 m in the easternSkagerrak. There are probably also differences in sedi-mentation load between coastal sites (directly affectedby land-runoff) and offshore submerged stone reefs(not affected by direct land-runoff), simply because oftheir geographical position.
Minuscule tidal fluctuation (less than ca. 20 cm)is a typical feature at the rocky shores of the Swedishwest coast and water level is mainly regulated by at-mospheric pressure (Fonselius, 1995). This, togetherwith the salinity gradient, creates special conditionsthat greatly affect the structure and composition of thealgal vegetation. The typical Atlantic intertidal beltbecomes lost with the reduction and final disappear-ance of tidal movements along a gradient from thenortheastern Skagerrak to the southern Kattegat. Sim-ultaneously, the water becomes increasingly brackishalong this gradient. Many marine rocky-shore mac-roalgae that on the Atlantic and North Sea coasts arefound in intertidal and upper littoral zones occur atincreasingly greater water depth when following thesalinity gradient from the Skagerrak through the Kat-tegat, the Öresund and finally into the Baltic Sea.This phenomenon has been defined as the ‘downwardprocess’ (Wærn, 1952; 1965). It has been explainedby the absence of tides, avoidance of more variablesurface salinities and changes in competitive balancewhen marine species successively disappear along thesalinity gradient and euryhaline species (the intertidalspecies from the Atlantic coast) compete for sub-littoral space. The downward process is especiallyconspicuous for the large canopy- and belt-formingbrown algae such as Fucus and Laminaria species(Wærn, 1965; Snoeijs, 1999). An example of a speciesthat expands its depth range tremendously is Fucusvesiculosus, which is the only large perennial canopy-forming alga that penetrates into the brackish watersof the Baltic Sea as far as the Gulf of Bothnia. On theSwedish west coast it has a depth range of ca. 20 cmonly, but in the northern Baltic Sea proper it formsbelts between a depth of 1 and 10 m in the absence ofcompetitors. To be able to resolve the downward pro-cess, one has to disregard subordinate gradients suchas differences in exposure, aspects of inner and outerarchipelagos, bottom configurations and substrata andthe orientation of the rocks in relation to the light

85
(Wærn, 1965). In the present study, it was for the firsttime possible to quantify the downward process fromthe Skagerrak to the Kattegat for the species that arecommon in both areas, because of the large quantityof data we collected from exposed sites.
A larger number of previous studies has describedchanges in the macroalgal vegetation as a result of eu-trophication or decreased eutrophication of bays andfjords of the Kattegat and Skagerrak (e.g. Klavestad,1978; Wennberg, 1987; Bokn et al., 1992; Wal-lentinus, 1996). A common phenomenon of the de-cline or deterioration of algal communities in baysand fjords caused by eutrophication or other humanimpacts such as direct effects of municipal discharges(Norin & Wærn, 1973), ferry traffic (Rönnberg, 1981),pulp-mill effluents (Kautsky et al., 1988), coolingwater discharge (Snoeijs & Prentice, 1989; Snoeijs,1992), fish farming (Rönnberg et al., 1992), etc. is theloss of functional diversity. This does not necessar-ily include lower species diversity, because all largeleaf-like canopy-forming perennial algae can be lostwhereas the group small filamentous ephemeral algaebecomes richer in species (Snoeijs & Prentice, 1989).Such a loss of important functional algal groups mayaffect a whole ecosystem because many invertebratesand fish are, at least part of their lives, dependenton the perennial algal belts for substratum, food andshelter. An example of this is the decline and thedecreased depth penetration of the Fucus vesiculosusbelt all around the Baltic Sea coasts (Kangas et al.,1982; Kautsky et al., 1986; Vogt & Schramm, 1991;Schramm, 1996; Eriksson et al., 1998). Therefore,we have tried in this paper to summarise different as-pects of the algal vegetation in terms of functionaldiversity at exposed sites, thus reflecting the situ-ation in the open waters. The phylogenetic groups(red, brown and green algae) summarise much of thephysiological performances of the species. Algal func-tional types can roughly be summarised by size andshape classifications. Littler & Littler (1980) and Lit-tler et al. (1983) found that thin, rapidly growing,short-lived algae are characteristic for unstable envir-onments, whereas coarse, slower-growing, long-livedalgae are characteristic of stable (late-successional)environments. Functional diversity is here derived bya deductive approach classifying the different algalspecies into groups of functionally similar species(types) which are expected to be critical in determin-ing the operation (function) of the submerged littoralvegetation. In modelling terrestrial plant communities,functional types are seen as a necessary device for
reducing the complexity and often uncharted charac-teristics of species diversity in function and structure(Woodward & Cramer, 1996). One of the aims of thisstudy was to test if such an approach can be usefulfor an algal vegetation. In this paper we report on thedepth distributions of both species diversity and func-tional diversity of the vegetation according to salinity(Kattegat compared with Skagerrak), sedimentation(coastal sites compared with offshore submerged stonereefs in the Kattegat), and the occurrence of a short-term catastrophe (a toxic pelagic microalgal bloom inthe Kattegat).
Materials and methods
Fieldwork
The rocky-shore macroalgal vegetation in the Swedishpart of the Kattegat and the Skagerrak was investig-ated by SCUBA diving during four expeditions, 3–29September 1988, 4–24 June 1989, 3–14 September1989 and 7–27 June 1990, with the Uppsala-based re-search vessel ‘MS Sunbeam’. Two winter dives fromland in February 1990 were included as well. Al-together ca. 150 forty-five – sixty min dives wereperformed within this project with 4–6 divers particip-ating in each. In every dive, at least two divers werePhD-students in phycology or professional phycolo-gists (one for video-filming and one for making notesand sampling of algae), and the others were assistantsfor photographing and carrying sampling bags.
At each diving profile (5–10 m wide) the abund-ance of each encountered upright (non-crustose) spe-cies was recorded for every metre depth intervalbetween the upper and lower occurrence of the algalvegetation, except for the upper metre of the littoral,which was subdivided into 0–0.5 and 0.5–1 m depthintervals. For the estimation of species abundance,a 0–5 ordinal scale was used which is suitable forunderwater conditions when only limited time is avail-able for recording the vegetation: 0: not found, 1:one individual, 2: several individuals, 3: common, 4:abundant, 5: dominant). The recording of the vegeta-tion was achieved by (1) underwater notes on paperfor each depth interval, (2) spoken records duringvideo-filming for each depth interval, (3) sampling ofalgae for each depth interval, (4) photographic records.Special attention was given to detecting the lowest ho-rizontal depth occurrences of macroalgae in the lowerlittoral. The criteria we used for determining the low-est limit of an alga are those of professor Mats Wærn
86
(pers. comm.): the alga has to grow attached to ahorizontal substratum so that it will receive the max-imum possible light dose, there must be possibilities tosettle further down as well and algae growing on smallstones or loose shells should not be considered be-cause these may have fallen down from more shallowdepths or may have been transported by currents.
Identification of the algal samples, if necessaryusing light microscopy, and ‘ecological’ herbariumsheets on which all species from a certain depth aregathered on the same sheet were made by a crew ofscientists and their assistants on board of ‘MS Sun-beam’. The scientists had followed the underwaterwork on a TV-monitor and they had been able to in-struct the divers via a telephone on board. With thehelp of the algal samples brought up, the divers noteswere completed with rare and/or smaller species thatwere missing from the direct underwater observations.The video-films, photographic slides and herbariumsheets are kept at the Universities of Stockholm andUppsala under the name ‘WWF 1988–1990’ and aremeant to serve as a basis of reference for this andfuture studies.
Data compilation
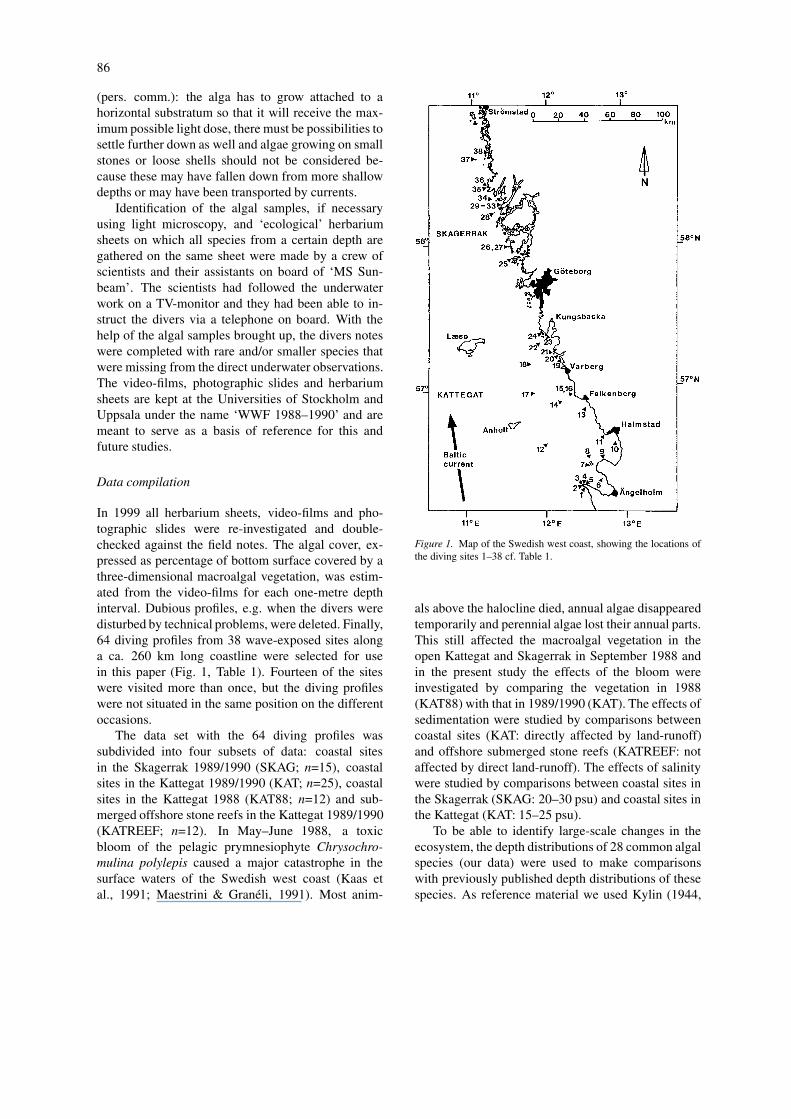
In 1999 all herbarium sheets, video-films and pho-tographic slides were re-investigated and double-checked against the field notes. The algal cover, ex-pressed as percentage of bottom surface covered by athree-dimensional macroalgal vegetation, was estim-ated from the video-films for each one-metre depthinterval. Dubious profiles, e.g. when the divers weredisturbed by technical problems, were deleted. Finally,64 diving profiles from 38 wave-exposed sites alonga ca. 260 km long coastline were selected for usein this paper (Fig. 1, Table 1). Fourteen of the siteswere visited more than once, but the diving profileswere not situated in the same position on the differentoccasions.
The data set with the 64 diving profiles wassubdivided into four subsets of data: coastal sitesin the Skagerrak 1989/1990 (SKAG; n=15), coastalsites in the Kattegat 1989/1990 (KAT; n=25), coastalsites in the Kattegat 1988 (KAT88; n=12) and sub-merged offshore stone reefs in the Kattegat 1989/1990(KATREEF; n=12). In May–June 1988, a toxicbloom of the pelagic prymnesiophyte Chrysochro-mulina polylepis caused a major catastrophe in thesurface waters of the Swedish west coast (Kaas etal., 1991; Maestrini & Granéli, 1991). Most anim-
Figure 1. Map of the Swedish west coast, showing the locations ofthe diving sites 1–38 cf. Table 1.
als above the halocline died, annual algae disappearedtemporarily and perennial algae lost their annual parts.This still affected the macroalgal vegetation in theopen Kattegat and Skagerrak in September 1988 andin the present study the effects of the bloom wereinvestigated by comparing the vegetation in 1988(KAT88) with that in 1989/1990 (KAT). The effects ofsedimentation were studied by comparisons betweencoastal sites (KAT: directly affected by land-runoff)and offshore submerged stone reefs (KATREEF: notaffected by direct land-runoff). The effects of salinitywere studied by comparisons between coastal sites inthe Skagerrak (SKAG: 20–30 psu) and coastal sites inthe Kattegat (KAT: 15–25 psu).
To be able to identify large-scale changes in theecosystem, the depth distributions of 28 common algalspecies (our data) were used to make comparisonswith previously published depth distributions of thesespecies. As reference material we used Kylin (1944,

87
Tabl
e1.
Sum
mar
yof
divi
ngsi
tes,
incl
udin
gda
tes
and
dept
hsof
inve
stig
atio
n
Are
aSi
tenr
Site
nam
ePo
sitio
nD
ate(
s)an
din
vest
igat
edde
pths
Kat
tega
tcoa
stal
site
s1
Ran
svik
56◦ 1
7′28′′
N,1
2◦28′ 5
4′′ E
8809
03:1
–13
m,9
0060
8:1–
15m
1988
,198
9,19
902
Åke
rsbe
rget
56◦ 1
7′53′′
N,1
2◦27′ 0
8′′ E
8909
13:0
–21
m3
Para
dish
amne
n56◦ 1
8′13′′
N,1
2◦27′ 0
7′′ E
8906
04:7
–21
m,8
9091
2:0-
21m
,900
203:
0–20
m,9
0060
7:0–
20m
4V
isitg
rotta
n56◦ 1
8′13′′
N,1
2◦27′ 5
7′′ E
8809
04:0
–3,8
–9an
d12
–13
m5
Abl
aham
n56◦ 1
8′08′′
N,1
2◦28′ 2
7′′ E
8809
05:0
–18
m,8
9060
5:0-
19m
,890
914:
0–18
m,9
0020
2:0–
13m
,900
607:
0–20
m6
Gry
tgru
nden
56◦ 2
0′46′′
N,1
2◦42′ 4
3′′ E
9006
09:1
–10
m7
Svar
tesk
är56◦ 2
6′11′′
N,1
2◦32′ 3
8′′ E
8809
06:0
–11
m,8
9060
8:0–
11m
9H
ovs
Hal
lar
56◦ 2
8′14′′
N,1
2◦42′ 2
4′′ E
8809
08:0
–13
m,8
9060
8:0–
12m
10På
arps
reve
t56◦ 3
7′06′′
N,1
2◦52′ 4
5′′ E
8809
09:6
–9m
,890
610:
5–8
m,8
9091
0:8–
10m
,900
614:
3–7
m11
Tylu
dden
56◦ 3
8′28′′
N,1
2◦44′ 2
9′′ E
8809
10:0
–5m
,890
610:
0–6
m,8
9090
5:0–
5m
,900
614:
0–5
m15
Glo
mm
aryg
gen
56◦ 5
5′49′′
N,1
2◦19′ 4
7′′ E
8809
13:5
–7m
16G
lom
men
sFy
r56◦ 5
5′51′′
N,1
2◦21′ 0
2′′ E
8809
14:0
–5m
19Ä
spev
ik57◦ 1
1′22′′
N,1
2◦11′ 0
4′′ E
8809
16:0
–5m
,900
616:
0–4
m20
Öst
raSa
ndan
57◦ 1
2′56′′
N,1
2◦09′ 2
3′′ E
8809
18:0
–5m
,890
615:
0–5
m,9
0061
6:0–
5m
21A
rvas
kär
57◦ 1
3′32′′
N,1
2◦05′ 0
6′′ E
9006
17:0
–18
m22
Lill
elan
d57◦ 1
8′23′′
N,1
1◦55′ 4
4′′ E
8906
15:0
.5–2
5m
23Ö
lmeu
dde
57◦ 2
1′28′′
N,1
2◦00′ 5
3′′ E
8906
18:0
–12
m24
Kyr
kefj
älls
und
57◦ 2
2′17′′
N,1
1◦56′ 2
9′′ E
8809
21:0
–5m
Kat
tega
tsto
nere
efs
8N
Wof
Hal
land
sV
äder
ö56◦ 2
8′08′′
N,1
2◦34′ 1
0′′ E
8909
11:1
0–18
m19
89,1
990
12St
ora
Mid
delg
rund
56◦ 3
2′54′′
N,1
2◦03′ 3
0′′ E
8906
11:8
–9an
d11
–12
m,9
0061
0:8–
12m
13K
nöla
grun
d56◦ 5
0′49′′
N,1
2◦29′ 1
1′′ E
9006
13:4
–11
m14
Mor
ups
Ban
k56◦ 5
2′36′′
N,1
2◦13′ 2
8′′ E
8906
12:1
2–26
m,8
9090
9:13
–26
m,9
0061
1:12
–25
m17
Lill
aM
idde
lgru
nd56◦ 5
5′00′′
N,1
1◦51′ 0
3′′ E
8909
08:7
–8an
d23
–24
m,9
0061
2:9–
10m
18Fl
aden
57◦ 0
8′48′′
N,1
1◦46′ 4
8′′ E
8906
14:9
–11
m,8
9090
3:9–
11,1
2–13
and
19–2
0m
,900
618:
8–20
m
Skag
erra
kco
asta
lsite
s25
Mar
stra
ndsö
n57◦ 5
3′28′′
N,1
1◦33′ 3
8′′ E
8906
17:0
–10
m19
89,1
990
26K
läde
shol
men
57◦ 5
6′41′′
N,1
1◦32′ 3
5′′ E
8906
18:8
–18
m27
Skap
holm
en57◦ 5
6′44′′
N,1
1◦32′ 0
7′′ E
8906
18:0
–18
m28
Bon
den
58◦ 1
2′35′′
N,1
1◦19′ 0
6′′ E
8906
20:0
–18
m29
Skäl
lhol
men
NW
58◦ 1
4′59′′
N,1
1◦22′ 5
4′′ E
9006
21:0
–10
m30
Skäl
lhol
men
S58◦ 1
4′54′′
N,1
1◦23′ 0
0′′ E
9006
21:0
–15
m31
Smed
jebr
otte
t58◦ 1
5′28′′
N,1
1◦22′ 0
0′′ E
9006
27:0
–23
m32
Har
pöbå
dar
58◦ 1
5′42′′
N,1
1◦22′ 0
5′′ E
8906
19:5
–23
m,9
0062
7:5–
24m
33Fl
atho
lmen
58◦ 1
5′43′′
N,1
1◦24′ 4
5′′ E
8906
19:0
–7m
34Sö
rgru
ndsb
erge
t58◦ 1
7′20′′
N,1
1◦11′ 0
1′′ E
9006
20:1
0–27
m35
Hål
lö58◦ 2
0′34′′
N,1
1◦12′ 4
3′′ E
8906
21:8
–22
m36
Bri
msk
är58◦ 2
1′02′′
N,1
1◦12′ 3
5′′ E
9006
26:0
–17
m37
Ärh
olm
en,V
äder
öarn
a58◦ 3
4′46′′
N,1
1◦04′ 4
3′′ E
8906
22:0
–24
m38
Val
önN
W58◦ 3
5′20′′
N,1
1◦14′ 2
8′′ E
8906
24:0
–17
m

88
Figure 2. Algal cover (a) and species richness (b) given as meansper water depth interval for each of the subsets of data: coastal sitesin the Kattegat 1989–1990 (KAT), coastal sites in the Kattegat 1988(KAT88), offshore submerged stone reefs in the Kattegat 1989–1990(KATREEF) and coastal sites in the Skagerrak 1989–1990 (SKAG).Error bars indicate standard error of the mean.
1947, 1949) who in these three books compiled hisknowledge of the macroalgae of the Swedish westcoast, which he gathered since he started his PhD-studies at Uppsala University in 1902 (PhD-thesis:Kylin, 1907). Kylin covered exactly the same area aswe did, but unfortunately he rarely mentioned differ-ences in depth distributions between the Kattegat andthe Skagerrak (only for 7 of the 28 species). There-fore, his upper depth limits are valid for the northernSkagerrak and his lower depth limits for the southernKattegat. Kylin’s investigations are based on dredging,and therefore, the lower limit is not completely trust-worthy. The second study we used to compare ourdepth distributions with is von Wachenfeldt (1975). Hemade a detailed study of the vegetation in the Öresundin 1963–1972, the narrow sound between Sweden andDenmark south of our study area, including ‘Kullen’
where our sites 1–5 also are situated. Therefore, hisupper depth limits are valid for the southern Kattegatand his lower depth limits for the southern Öresund.Von Wachenfeldt’s investigations are based on bothdredging and diving. The taxonomic nomenclatureused throughout this paper is according to the officialcheck-list of the Baltic Marine Biologists (Nielsen etal. 1995)
Data analysis
Detrended Correspondence Analysis (DCA), imple-mented with the programme CANOCO 4 (ter Braak& þmilauer, 1998), was carried out to summariseoverall patterns in the vegetation of the 64 diving pro-files. Community diversity was calculated as speciesrichness, Shannon index (log base=e) and, Pielou’sevenness measure (Diamond & Case, 1986). Fur-ther statistical tests, analysis of variance (ANOVA),unpaired t-test and Pearson’s product-moment correl-ation (Fowler et al., 1998), were performed with theMINITABTM statistical package.
The categories for describing algal functional di-versity were: (a) phylogenetic classification in red,brown and green algae, (b) classification into sizegroups of the maximum possible size for each species:<10 cm, 10–50 cm and >50 cm, and (c) classifica-tion into shape groups simplified after Littler & Littler(1980): filamentous (filaments <1 mm wide through-out the thallus), thread-like (threads >1 mm wide in atleast part of the thallus) and leaf-like (most of the thal-lus flat). In the calculations all depths down to 27 mwere included. If the lower limit of algal occurrencewas set by sedimentation (reported by the divers), thecover and species richness from this limit down to27 m were 0.
Results
Algal cover
The relationship between algal cover and water depthis shown in Figure 2a for each of the subsets ofdata. Correlation analyses showed significant neg-ative associations between water depth and meanalgal cover for SKAG (r=−0.86, p<0.001), KAT(r=−0.98, p<0.001), KAT88 (r=−0.96, p<0.001) andKATREEF (r=−0.95, p<0.001). Two-way ANOVAanalyses using subset of data and water depth as modelvariables showed that the algal cover was significantlydifferent between KAT and KAT88 (p<0.001), KAT

89
and KATREEF (p<0.001) and SKAG and KATREEF(p<0.001), but not between SKAG and KAT.
At coastal sites in the Skagerrak, the algal coverwas higher than at coastal sites in the Kattegat between1 and 8 m, but it was the other way around in the uppermetre, and between 12 and 19 (Fig. 2a). The latter waspartly caused by the much larger competition for spacebetween algae and marine sessile animals in the lowerlittoral of the Skagerrak. In the Kattegat, many of theseanimals do not occur because of the low salinity. An-other reason for the differences in depth distribution ofthe algal cover between the Skagerrak and the Kattegatis that the maximum development of the Laminariabelt (with high cover) is situated higher up in the Sk-agerrak (ca. 2–7 m) than in the Kattegat (ca. 7–12 m).The mean algal cover at the stone reefs in the Kattegatwas much higher than that at coastal sites of both theKattegat and Skagerrak at all investigated depths.
The mean algal cover in the Kattegat in 1988 wasclearly lower than in 1989/1990 below a depth of ca.5 metres. The lower cover in 1988 can be attributedto the toxic phytoplankton bloom in May–June 1988.In September 1988 the perennial macroalgae were stillvisibly damaged by the dose of microalgal toxin theyreceived in the spring of that year, although new thal-lus had begun to grow from most of them. In 1988 thevegetation penetrated down to 18 m at the most, butin 1989/1990 it was 25 m. Also, the upward shift ofthe lower vegetation limit in 1988 can be consideredan effect of the toxic bloom, which caused heavysedimentation when it died out, thus burying and suf-focating the lower macroalgal vegetation. The diversreported this phenomenon frequently in 1988.
Species richness
Shannon diversity was most correlated to the rich-ness component (n=748, r=0.92, p<0.001) and lessto the evenness component (n=748, r=0.51, p<0.001).All three diversity measures showed only weak cor-relations with algal cover (n=748; richness: r=0.36;Shannon r=0.43, evenness: r=0.25; all three p<0.001).The relationship between algal species richness andwater depth is shown in Fig. 2b for each of the subsetsof data. Correlation analyses showed significant neg-ative associations between water depth and mean algalspecies richness for SKAG (r=−0.96, p<0.001), KAT(r=−0.96, p<0.001), KAT88 (r=−0.92, p<0.001)and KATREEF (r=−0.94, p<0.001). Two-way AN-OVA analyses using subset of data and water depthas model variables showed that algal species rich-
ness was significantly different between KAT andKAT88 (p<0.001), KAT and SKAG (p<0.001) KATand KATREEF (p<0.001) and SKAG and KATREEF(p<0.001).
The mean algal species richness followed the se-quence: stone reefs in the Kattegat > coastal sitesin the Skagerrak > coastal sites in the Kattegat >
coastal sites in the Kattegat 1988. At the reefs speciesrichness showed a small peak between 18 and 22 mdepth under the halocline, a phenomenon that wasnot observed at the coastal sites. This peak is mainlycaused by an increase in red algae. At the reefs somespecies occurred which were not encountered at thecoastal sites in the Kattegat or Skagerrak in this in-vestigation: Chylocladia verticillata (red), Halarach-nion ligulatum (red), Lomentaria orcadensis (red),Plocamium cartilagineum (red), Sphondylothamnionmultifidum (red), Cutleria multifida (brown), and somethat were found at coastal sites in the Skagerrak but notin the Kattegat: Apoglossum ruscifolium (red), Bon-nemaissonia asparagoides (red), Pterosiphonia para-sitica (red). All of these are marine species that areless tolerant to lower salinity than the other algae en-countered in this study. Their occurrence at the reefs ismost likely a combination of absence of sedimentation(more rocky substratum available for the settlement ofmacroalgae) and higher salinity under the halocline.At coastal sites in the Kattegat species richness waslower in 1988 than in 1989/1990 below a depth of ca.5 m. This was related to the loss of some ephemeralspecies caused by the microalgal toxin; the perennialspecies were still there and had started to grow again.
Community analysis
The data set contained 103 species and 748 samples(=depth intervals) with altogether 8878 abundancerecords. The eigenvalues for the first four DCA ordina-tion axes were 0.53, 0.32, 0.24 and 0.23, respectively.Axis 1 explains most of the variation in the speciescomposition as can be concluded from its high eigen-value compared to that of axis 2. Axis 1 is stronglycorrelated with water depth (n=748, r=0.90, p<0.001;Fig. 3a). Axis 2 reflects the difference in the vegeta-tion between the more marine Skagerrak and the morebrackish Kattegat (Fig. 3b). This was tested by cor-relation analyses between axis 2 and distance (in km)from site 1 (r=0.56, p<0.001). For coastal sites in theKattegat the community composition in 1988 was notvery different from that in 1989/1990 as can be con-cluded from the mixed positions of the sample scores

90
for KAT88 and KAT. The sample scores for the stonereefs in the Kattegat (KATREEF) have a position inter-mediate SKAG and KAT/KAT88 on Axis 2, indicatingthat its community composition has elements of boththe Skagerrak and the Kattegat. No patterns in spe-cies composition related to time of year (September orJune) were discovered.
Four community variables were tested by correla-tion analysis on the first two DCA axes (all n= 748,p<0.001). Axis 1 was more correlated to algal cover(r=−0.44) than axis 2 (r=0.15). Axis 2 was morecorrelated to species richness and Shannon diversity(r=0.59, r=0.59) than axis 1 (r=−0.25, r=−0.32).Both axis were only weakly correlated to evenness(axis 1: r=−0.19; axis 2: r=0.16).
The species scores of the DCA ordination (Fig. 4a,b) reflect the same patterns as shown by the samplescores (Fig. 3). From left to right along Axis 1 thespecies follow a gradient from the upper littoral inthe far-left (e.g. Fucus vesiculosus, F. evanescens,F. spiralis, Pilayella littoralis, Porphyra purpurea,Urospora penicilliformis) to the lower littoral in thefar-right (e.g. Dilsea carnosa, Erythrodermis traillii,Lomentaria orcadensis , Odonthalia dentata, Pter-osiphonia parasitica). From the lower part of theordination to the upper part the species follow a gradi-ent from euryhaline species that occur more in theKattegat (e.g. Ceramium strictum, Cladophora spp.)to more marine species that occur more in the Skager-rak (e.g. Aglaothamnion hookeri, Chordaria flagel-liformis, Corallina officinalis, Desmarestia aculeata,Laminaria hyperborea, Lomentaria clavellosa). Redalgae dominate the lower half of the littoral zone (righthalf of the ordination) and brown and green algae aremore common in the upper half of the littoral zone,together with some red algae that are obligate in theupper littoral such as Porphyra spp. and Bangia at-ropurpurea (left half of the ordination). No patternrelated to the four subsets of data (areas) was foundfor the phylogenetic groups in the ordination analysis.Any patterns related to algal size and shape were alsomore complicated and could not be detected in the or-dination analysis. However, several patterns emergedwhen the relative proportions of groups of species(phylogenetic groups, size groups and shape groups)that make up the littoral macroalgal communities wereinvestigated.
Phylogenetic groups
The mean percentages of red, brown and green algalspecies in each depth interval are shown in Fig. 5a–c.Although species richness and the total depth penet-ration differed markedly between the four differentgroups of sites investigated (Fig. 2), the major trendsin the distributions of the phylogenetic groups wererather similar: the proportion of red algae increasedwith depth whereas that of green and brown algaedecreased. Red algal species were always more nu-merous than green and brown together, except in theupper metres of the littoral. The percentage brownalgae was relatively stable around 25% in the Kat-tegat between 1 and 17 m whereas in the Skagerrakthere was a successive drop from ca. 35% down toless than 5% between 1 and 17 m. This means that inthe more marine waters of the Skagerrak brown algaeare more confined to the upper littoral, whereas in theKattegat they are more evenly distributed throughoutthe littoral by the downward process. The proportionof green algal species was highest in the upper metreof the littoral everywhere and below this their contri-bution decreased at all coastal sites. The decrease wasfaster in the Skagerrak (from ca. 30 to <5% between0 and 6 m of depth) than in the Kattegat (from ca30 to <5% between 0 and 12 m of depth), indicat-ing that the more marine species that do not occur inthe Kattegat are found in the groups red and brownalgae. In the lower littoral, between a depth of 14and 24 m, the percentage green algae was highest (al-though only about 5–8%) at the stone reefs. This smallgroup of ‘deep-water’ green algae consisted of onlythree species, Cladophora rupestris, Chaetomorphamelagonium and Derbesia marina.
All three phylogenetic groups are represented withhigher diversity deeper down at the reefs than at thecoastal sites. At the reefs first brown algae disappearat 21 m, then green algae at 23 m, and finally redalgae at 26 m. This sequence was different to the or-der of disappearance with depth in the other subsetsof data where green algae always disappeared first in-stead of brown algae. This shows that species of allthree phylogenetic groups can penetrate deeper whensedimentation is less, but especially so the red algae.
Size groups
The mean percentages of the three size groups of algalspecies in each depth interval are shown in Figure 5d–f. The increasing trend of the medium-sized group

91
Figure 3. Plot of the first two axes of the DCA ordination, showing sample scores marked according to water depth (a) and subset of data (b).
10–50 cm, from ca 35% at the surface to 100% deep-est down, was similar for all four areas. The group>50 cm (only brown algae of the genera Ascophyllum,Chorda, Chordaria, Desmarestia, Fucus, Halidrys,Halosiphon and Laminaria) showed pronounced dif-ferences in relative proportions between the four areas.In the Skagerrak there was a rather constant decreasing
trend from ca 30% at a depth of 1 m down to 0% at22 m. In contrast, an increase from ca. 15% at thesurface to ca. 30% at a depth of 13 m was found inthe Kattegat. This once again shows that in the moremarine waters of the Skagerrak brown algae are moreconfined to the upper littoral, whereas in the Kattegatthey are more evenly distributed throughout the lit-

92
Figure 4. For legend see p. 93
toral by the downward process. It also shows that thismainly concerns the large brown algae, but it is prob-ably even valid for their obligate/typical brown algalepiphytes, such as Elachista fucicola on Fucus spp.and Sphacelaria cirrosa on Halidrys siliquosa.
The smallest size group, species <10 cm, showedmany different patterns. At the coastal sites in the Kat-tegat this group of small algae decreased steeply fromca. 50% at the water surface down to ca. 10% below adepth of 12 m. In 1988, this group was not represen-ted below 13 m, which was probably an effect of the

93
←−Figure 4. Plot of the first two axes of the DCA ordination (cf. Fig. 4), showing species scores marked according to phylogenetic group forthe 41 most abundant taxa (a) Brown algae: Chor fil = Chorda filum, Chor fla = Chordaria flagelliformis, Desm acu = Desmarestia aculeata,Desm vir = Desmarestia viridis, Ecto sil = Ectocarpus siliculosus, Fucu ser = Fucus serratus, Fucu ves = Fucus vesiculosus, Hali sil =Halidrys siliquosa, Lami dig = Laminaria digitata, Lami sac = Laminaria saccharina, Spha cir = Sphacelaria cirrosa, Green algae: Bryo plu= Bryopsis plumosa, Chae mel = Chaetomorpha melagonium, Clad fle = Cladophora flexuosa, Clad glo = Cladophora glomerata, Clad rup =Cladophora rupestris, Ente spp = Enteromorpha spp., Red algae: Ahnf pli = Ahnfeltia plicata, Bonn ham = Bonnemaisonia hamifera, Bron bys= Brongniartella byssoides, Call cor = Callithamnion corymbosum, Cera nod = Ceramium nodulosum, Cera str = Ceramium strictum, Chon cri =Chondrus crispus, Cocc tru = Coccotylus truncatus, Cora off = Corallina officinalis, Cyst pur = Cystoclonium purpureum, Dele san = Delesseriasanguinea, Dils car = Dilsea carnosa, Furc lum = Furcellaria lumbricalis, Lom cla = Lomentaria clavellosa, Mem ala = Membranoptera alata,Odon den = Odonthalia dentata, Phyc rub = Phycodrys rubens, Phyl pse = Phyllophora pseudoceranoides, Poly elo = Polysiphonia elongata,Poly fib = Polysiphonia fibrillosa, Poly fuc = Polysiphonia fucoides, Poly str = Polysiphonia stricta, Pter plu = Pterothamnion plumula, Rhodcon = Rhodomela confervoides and 42 common taxa (b) Brown algae: Asco nod = Ascophyllum nodosum, Aspe bul = Asperococcus bullosus,Dict foe = Dictyosiphon foeniculaceus, Ecto fas = Ectocarpus fasciculatus, Elac fuc = Elachista fucicola, Fucu eva = Fucus evanescens, Fucuspi = Fucus spiralis, Halo tom = Halosiphon tomentosus, Lami hyp = Laminaria hyperborea, Peta fas = Petalonia fascia, Pila lit = Pilayellalittoralis, Scyt lom = Scytosiphon lomentaria, Spha plu = Sphacelaria plumosa, Spon tom = Spongomorpha tomentosum, Green algae: Blidmin = Blidingia minima, Chae aer = Chaetomorpha aerea, Clad ser = Cladophora sericea, Codi fra = Codium fragile, Rhiz rip = Rhizocloniumriparium, Spon aer = Spongomorpha aeruginosa, Ulva lac = Ulva lactuca, Uros pen = Urospora penicilliformis. Red algae: Agla hoo =Aglaothamnion hookeri, Apog rus = Apoglossum ruscifolium, Bang atr = Bangia atropurpurea, Bonn asp = Bonnemaisonia asparagoides, Callcri = Callophyllis cristata, Cera arb = Ceramium arborescens, Dumo con = Dumontia confervoides, Lome orc = Lomentaria orcadensis, Palmpal = Palmaria palmata, Phyl tra = Phyllophora traillii, Plum plu = Plumaria plumosa, Poly bro = Polysiphonia brodiaei, Poly rot = Polyidesrotundus, Porp coc = Porphyropsis coccinea, Porp lin = Porphyra linearis, Porp pur = Porphyra purpurea, Porp umb = Porphyra umbilicalis,Pter par = Pterosiphonia parasitica, Ptil gun = Ptilota gunneri, Rhod pur = Rhodochorton purpureum. The species scores of the 20 leastabundant taxa are not shown.
Chrysochromulina bloom. In the Skagerrak and at thestone reefs, this size group was rather constant around30% in the upper 12 m. In the lower littoral, below ca.20 m this group often showed a maximum again whichcan be attributed to the occurrence of small more mar-ine species below the halocline. All size groups arerepresented with higher diversity deeper down at thereefs than at the coastal sites. First algae >50 cm dis-appear at 20 m, then algae <10 cm at 24 m, and finally,algae 10–50 cm at 26 m. This sequence was similarto the order of disappearance with depth in the othersubsets of data.
Shape groups
The mean percentages of the three shape groups ofalgal species in each depth interval are shown in Fig-ure 5g–i. The leaf-like species and the filamentousspecies show opposite trends. The mean relative pro-portions of leaf-like algae increased from ca 20–30%in the upper littoral to 100% in the lower littoral for allsubsets of data. The mean relative proportions of fila-mentous algae decreased from ca 50–60% in the upperlittoral to 0% in the lower littoral for all site groups.This implies that in exposed sites a leaf-like shape isan important trait for species in the lower littoral, anda thread-like or filamentous one in the upper littoral.In the coastal sites of the Kattegat a larger proportionof leaf-like species and smaller proportions of thread-like and filamentous species were found than in the
coastal sites of the Skagerrak. In the upper 1–4 m thereare more filamentous and less leaf-like species in theKattegat and below 6 m it is the other way around.This is not easy to explain, as it may be a combinedeffect of the loss of marine species in the Kattegat,the downward process or eutrophication in the Kat-tegat. The higher proportion of thread-like species atthe reefs (Fig. 5g) is probably an effect of the smallmarine species that are not found in coastal sites, suchasChylocladia verticillata, Lomentaria orcadensis andPlocamium cartilagineum.
All three shape groups are represented with higherdiversity deeper down at the reefs than at the coastalsites. Thread-like algae disappear at 22 m, filamentousalgae at 24 m, and finally, leaf-like algae at 26 m. Thissequence was similar to the order of disappearancewith depth in the other site groups. The alga with thelowest occurrence in the littoral in this study through-out was Phycodrys rubens at 27 m. Further low findswere Coccotylus truncatus at 26.5 m and Delesseriasanguinea at 26 m, followed by Dilsea carnosa andOdonthalia dentata at 24–25 m. These five species areall leaf-like red algae within the size group 10–50 cm.
Deepest occurrence of perennial species
Figure 6 shows the mean deepest occurrences of 18perennial algal species common at the stone reefs inthe Kattegat 1989/1990 (unaffected by land runoffand without visible sedimentation) and at the coastal

94
Figure 5. Depth distributions of the mean relative proportions (in%) of algal species richness within different phylogenetic groups (a–c), sizegroups (d–f) and shape groups (g–i) for all four subsets of data. Error bars indicate the standard error of the mean.

95
sites in the Kattegat 1989/1990 and the Skagerrak1989/1990 (with visible sedimentation). The deepestoccurrences of all 18 species were always deeper atthe reefs than at the coastal sites. The difference wasas much as 11.5 m for Polysiphonia elongata and 10 mfor Chaetomorpha melagonium when comparing thereefs with coastal sites in the Kattegat.
Quantification of the downward process
For 28 algal species the mean upper and lower limitsfor the species’ occurrences in the Skagerrak and Kat-tegat (excluding 1988) were compared (Fig. 7). These28 species belong to the 35 most abundant taxa in thedata set and most of them are perennials. The otherseven of the 35 most abundant taxa (mainly filament-ous ephemerals) had fragmented distributions in thelittoral and are not shown (Ceramium strictum, Clado-phora flexuosa, Cladophora glomerata,Ectocarpussiliculosus, Enteromorpha spp., Polysiphonia fibril-losa, Pterothamnion plumula). Except for the twoFucus species, all species in Figure 7a had signific-antly lower mean upper limits in the Kattegat (un-paired t-tests with significance level p<0.05). TheFucus species extended their depth distributions bylowering the lower depth limit by 4.5 (F. vesiculosus)and 3 m (F. serratus). This implies that a downwardprocess occurs for all 14 species. No significant down-ward process of the upper limit was found for the 14species in Figure 7b. Polysiphonia elongata was theonly species that had a significantly higher upper limitin the Kattegat than in the Skagerrak. The lower limitsof the 28 species were calculated only from the profilesthat were 18 m or deeper and also the stone reef pro-files were included. It is possible that the lower limitsin this investigation (except for the shallow ones of theFucus species and Ahnfeltia plicata) are mainly set bysedimentation (divers observations) in the Skagerrak,as opposed to the Kattegat, and thus they do not reflectthe depth distributions in the same way as the upperlimits do.
Comparison with previous records of depthdistributions
The depth distributions of the same 28 species asused for calculating the downward process were usedin comparisons with previously published algal depthdistributions. Several species showed in 1989–1990large deviations from their previous downward pro-cess, which is illustrated in Figure 7 by the upperdepth limits of Kylin (1944, 1947, 1949) and von
Figure 6. Mean maximum depth limits of 18 perennial macroalgalspecies in the coastal sites and stone reefs of the Kattegat and thecoastal sites of the Skagerrak. Error bars indicate the standard errorof the mean. Abbreviations cf. Figure 4.
Wachenfeldt (1975). Instead, some species showed aclear upward shift in the form of (1) higher upperlimit in 1989–1990 than in 1902–1944: Brongniar-tella byssoides, Cystoclonium purpureum, Delesseriasanguinea, Dilsea carnosa, Odonthalia dentata, Phy-codrys rubens, or (2) Absence of a previously cleardownward process: Bonnemaisonia hamifera, C. pur-pureum, D. sanguinea, P. rubens and Polysiphoniaelongata. Also Kylin mentioned for three of thesespecies that they had a downward process of their up-per vegetation limit, these are B. hamifera (0.5 m inthe Skagerrak – 10 m in the Kattegat), B. byssoides(3–5 m) and P. elongata (0.5–5 m). These eight spe-cies can all be considered opportunistic red algae,either ephemerals or perennials with annual parts. B.hamifera (= Trailliella intricata Batters) is a non-indigenous species from the Pacific Ocean that hasinvaded the area about 100 years ago (Kylin, 1916).Another result from the comparison is that the brownalga Halidrys siliquosa and its epiphyte Sphacelariacirrosa (previously known as S. bipinnata) had a muchdeeper distribution in both the Skagerrak and Kattegatthan previously reported by Kylin (1947), down to15–19 m instead of 3–5 m.
Discussion
The lower vegetation limit set by light
We have shown that macroalgal species diversity ismuch higher at the offshore stone reefs in the Kattegat

96
than at exposed coastal sites in either the Kattegat orthe Skagerrak. The stone reefs are unaffected by directland-runoff, being completely submerged under thewater surface and far away from land runoff from theSwedish and Danish coasts. The water currents thatpass the stone reefs are strong and below the halocline(at about 20 m depth) they contain marine North Seawater (>30 psu). This probably contributes to the highdiversity and good condition of the algae. Most likelythe main reason, however, is that the stone reefs havelittle sedimentation because of their remote positionand the continuous removal of particles by the strongcurrents. This was confirmed by visual observationsby our divers. High species diversity at Danish off-shore stone reefs has earlier been reported by Nielsen(1991) and Nielsen & Dahl (1992a, b). The only reefvisited both by us and Nielsen & Dahl (1992b) isStora Middelgrund. When comparing the species com-position found at 8–12 m, the observations are rathersimilar for the upright macroalgae in both studies, ex-cept that Nielsen & Dahl did not record Fucus serratusbut we recorded it in 1989 with a lower limit at ca.10 m depth. Nielsen & Dahl found 14 species at 25 mdepth, whereas we only found 1–3 species in three pro-files on stones lying in the gravel bottoms at MorupsBank at 25 m (all other reef profiles we visited wereless deep). The absolute lowest limit for the uprightalgal species at horizontal substrates at the reefs inour study was 26.5 m (Coccotylus truncatus). Stoneslocated further down were covered with crustose redalgae but no upright macroalgae occurred althoughplenty of substratum in the form of stones was present.
At 20 m depth we found 11 species at MorupsBank where the bottom consisted of stones and coarsegravel without much light reflection. At Fladen a whiteshell bottom with good light reflection at the samedepth supported 22 species of upright macroalgae.This suggests that the lower limit of algal vegetationis not regulated by light penetration through the wa-ter column only, but that reflection from the bottomcan contribute substantially to the light regime, af-fecting algal growth and diversity. At Morups Bank,the Coccotylus truncatus plants at 26.5 m depth weresmall (1–2 cm) and growing on a large horizontalstone. The algae at 24 m depth were surprisingly large(20–30 cm), probably because they have access to nu-trients continuously due to the strong currents. Despitethe large size and healthy appearance of the plants,the total cover of the vegetation was less than 1% at24 m, consisting of single individuals or small patcheswith large empty spaces in-between. There is no lack
of substratum, light and nutrients for more algae togrow here, but we suspect that here the reason forthe low total cover lies in recruitment problems. Thestrong currents are most likely involved in this by car-rying away fragments of thalli as well as spores andsporelings as they also do with sediment particles.
The lower vegetation limit set by sedimentation
In contrast to the offshore reefs, all the coastal siteswere visibly affected by sedimentation, even at placeswith deep profiles such as Paradishamn, Lilleland,Sörgrundsberget and Ärholmen. At none of the coastalsites the lower vegetation limit was clearly set bylight as described above for the coastal reefs. In-stead, the divers always noted that algal cover andabundance decreased in concert with increasing thick-ness of the sediment layer on algae and substratum.Sedimentation affects rocky-shore algae in two ways:by covering photosynthetic tissue, and by coveringthe substratum so that spores and sporelings of al-gae cannot settle. Algae with ‘tough’ thalli such asPhyllophora pseudoceranoides, Coccotylus truncatus,Polysiphonia elongata and Sphacelaria arctica seemto be most resistant to sedimentation (Eriksson et al.,1998; Johansson et al., 1998).
It may be argued that sedimentation is a naturalprocess at coastal sites close to land in contrast tooffshore areas. However, a general eutrophicationof the Kattegat and Skagerrak coastal areas throughland-runoff and air deposition has been documented(Rosenberg, 1990; Rosenberg et al., 1990), and sed-imentation has increased significantly. For example,Wassman (1990) calculated that the sedimentation ofparticular organic carbon out of the euphotic zone inthe southern Kattegat would have increased by 140–250% since 1950. Johannessen and Dahl (1996) com-piled oxygen concentration measurements from 31stations in the Skagerrak from 1927 to the 1990s andconcluded that the observed decreases in oxygen sincethe 1960s–1970s most likely are caused by increasedsedimentation of phytoplankton by increased nutrientload of the coastal waters. Probably also part of thesedimentation load on the rocky shores observed dur-ing the present study is caused by more than basic landrunoff, and increasing eutrophication will in the endturn rocky shores into soft bottoms. Besides sediment-ation, other factors related to land runoff (pollutants,freshwater discharges, etc.) may also have a negativeeffect on macroalgal species diversity at coastal sitescompared to the offshore reefs.

97
Figure 7. Comparisons of the mean depth ranges (grey bars) and the range between the lowest and highest occurrence throughout (error bars)of 28 macroalgal species in the Skagerrak (SKAG) and the Kattegat (KAT). To the left and right of each pair of grey bars (this study) the depthintervals reported by Kylin (1944, 1947, 1949) and von Wachenfeldt (1975), are given. (a) Species with significant differences (p<0.05) in theupper depth limits between the Skagerrak and the Kattegat, except for the two Fucus spp. which had no significant difference. (b) Species withno significant differences (p>0.05) in the upper depth limits between the Skagerrak and the Kattegat, except for Polysiphonia elongata whichhad a significant higher upper depth limit in the Kattegat. Abbreviations cf. Figure 4.
Functional diversity
All three classifications of algal functional diversityshowed strong correlations with depth: red algae,leaf-like shape and size group 10–50 cm increased,whereas brown and green algae, thin filamentous andthread-like shape, as well as the size groups <10 and>50 cm all decreased. Deviations from these generalpatterns could be explained by the occurrence of ahalocline (Skagerrak coast and Kattegat stone reefs)and changes in competitive balance by the downwardprocess. The vertical distribution of algae dependsgreatly on light penetration, and changes in the lightclimate can cause changes in the zonation of algal
phylogenetic groups. The depth distribution of algaedepends on light penetration and availability of sub-stratum. There is a classical correlation between thecolour of an alga and its depth range, although manyexceptions occur. This is because the algal pigmentsare designed to absorb specific wavelengths of thelight spectrum. All three algal groups have chlorophylla and are able to absorb blue and red light. Green algaehave a pigment composition close to that of terrestrialplants and they can most effectively use the red part ofthe light spectrum. As red light penetrates only a fewmeters into the water, green algae are mainly foundclose to the water surface but they can also be foundat greater depths in waters rich in blue light (Hillis-

98
Colinvaux, 1986), which is a possible explanation forthe deep penetration of Cladophora rupestris, Chaeto-morpha melagonium and Derbesia marina at the reefsin the present study. The photosynthetic efficiency isalso known to increase with depth in red alga richin phycoerythrin like Delesseria sanguinea (Lüning,1990). Algae also show different sensitivity to UV-Bradiation (Dring et al., 1996; Beardall et al., 1998)which can result in changes of functional diversitywith climatic change when species with UV toleranceor repair mechanisms are favoured.
The toxic phytoplankton bloom
The toxic Chrysochromulina bloom in 1988 affectedalgal species richness negatively. The perennial algaewere visibly damaged by the algal toxins, but newthallus had already grown out from most of them inSeptember 1988 and also annual algae had begun tocolonise again. Species composition was affected bythe bloom, but not dramatically and there were nomajor differences in the depth distributions of algalfunctional diversity. The most conspicuous effect ofthe bloom was the loss of biomass at all depths andthe upward shift of the lower vegetation limit by 7 m.The loss of biomass was especially pronounced froma depth of 7 m down to 12 m, which coincides withthe pycnocline in the southern Kattegat and here thetoxic effect was greatest and/or lasted longest (Lindahl& Rosenberg, 1989; Kaas et al., 1991). We suggestthat the upward shift of the lower vegetation limit wasmost likely caused indirectly by the toxic bloom aswell. Thick layers of sedimentation covered the mac-roalgae when the bloom died off and bottom anoxiawas generated even on the stone surfaces. This pro-cess was reinforced by large numbers of decomposinganimals that had suffered mass death from the toxin(C. Larsson, pers. comm.).
The downward process
We were able to quantify the downward process in theSkagerrak and the Kattegat for 14 species by com-paring means of the upper and lower depth limitsfor the two areas. Previously this event was knownfrom single depth records without a statistical basisand with sampling mainly based on dredgings (Kylin,1907, 1944–1949; Rosenvinge, 1909–1931; Rosen-vinge & Lund, 1941–1947; Lund, 1950; Wærn, 1965).We documented a downward process of the meanupper depth limit for Ahnfeltia plicata, Cladophorarupestris, Laminaria digitata, Laminaria saccharina
and, Rhodomela confervoides which would not havebeen found by merely comparing the highest recordsbetween the two areas (all were found in the upper0.5 m in both the Kattegat and the Skagerrak).
From our results, we can distinguish severaldifferent types of species responses that can beattributed to the downward process in the Skagerrakand Kattegat: (1) Some species increase theirvertical distribution along the Baltic gradient fromthe Skagerrak to the Kattegat mainly by lowering thelower limit. To this group belong Fucus vesiculosus,Fucus serratus, Ahnfeltia plicata, Rhodomela confer-voides andCladophora rupestris. These all grow inthe intertidal zone at the Atlantic coasts (Kornmann& Sahling, 1977; Russell, 1991; Bird & McLach-lan, 1992) and they penetrate relatively far into theBaltic Sea, to salinities less than 7 psu (Nielsen etal., 1995). They are probably highly competitive whensalinity decreases along the gradient. Two Laminariaspecies, L. digitata and L. saccharina, also belong tothis group, but their downward process started earlier(probably in the very north of the Skagerrak) as theirdepth intervals in both the Skagerrak and the Kattegatare much wider than on Atlantic coasts. Laminaria hy-perborea we found only in the Skagerrak and this spe-cies’ downward process started even earlier. (2) Somespecies have a decreased vertical distribution alongthe Baltic gradient from the Skagerrak to the Kattegatmainly by lowering their upper limit. To this group be-long Corallina officinalis, Desmarestia viridis, Dilseacarnosa,Halidrys siliquosa, Odonthalia dentata andSphacelaria cirrosa. At the Atlantic coasts these aresublittoral marine species, except that C. officinaliscan be found also in the lower intertidal zone (Korn-mann & Sahling, 1977; Bird & McLachlan, 1992).These species do not penetrate far into the Baltic Sea(Nielsen et al., 1995) and probably lose in competitionwith other algae when salinity decreases in the Kat-tegat. (3) Some species do not show major changesin their depth distributions along the Baltic gradientfrom the Skagerrak to the Kattegat. In this group wefind Ceramium nodulosum,Coccotylus truncatus, Fur-cellaria lumbricalis, Phyllophora pseudoceranoides,Polysiphonia fucoides, all of which penetrate far intothe Baltic Sea (Nielsen et al., 1995). For these species,the downward process at large has probably not yet be-gun in the open Kattegat. Furthermore, there are alsospecies that never take part in the downward processbut stick to their position in the upper littoral fringe,e.g. Fucus spiralis and Dictyosiphon foeniculaceus(Wærn, 1965).

99
The most important force driving the downwardprocess is probably the competition between algae thatare able to tolerate low salinity to different degrees.Most of the downward process on the Swedish westcoast takes place above the halocline and therefore, itis unlikely that the vertical distribution of the salinityof the water column is of large importance (except per-haps in the lower littoral). Furthermore, it continuesalso in the Baltic Sea where the halocline is situatedfar beneath the photic zone. The downward processtakes place also at the stone reefs, which excludesfreshwater runoff from land or a melting ice cover asmajor forces. Therefore, the most likely mechanism isthat some species are better competitors when salinitygets lower along the Swedish west coast southwardsand then along the Swedish east coast northwards,and they ‘press down’ the vertical distribution of otherspecies and successively also of each other. Low sa-linity may also decrease the success of fertilisation byreducing the mobility and longevity of gametes, andincreased polyspermy, which is lethal for most species(Serrão et al., 1999). Amongst the structurally import-ant intertidal species along the Baltic Sea gradient thefollowing sequence can be recognised: Laminaria hy-perborea, L. digitata, L. saccharina, Fucus serratus,F. vesiculosus. The first one disappears already in thenorthern Kattegat, the last one in the northern BalticSea (3–4‰). Species pressed down in their verticaldistribution by these five large brown-algal speciesare, e.g. Halidrys siliquosa, Sphacelaria cirrosa andDilsea carnosa.
An upward shift indicates eutrophication
When comparing today’s rocky-shore macroalgal ve-getation of the Skagerrak and Kattegat with studiesfrom around 1900, the vertical range of the total veget-ation seems to have moved upwards. If this is true, thereason would be a large-scale eutrophication resultingin increased production in the pelagic zone and sub-sequent decreased light availability for macroalgae inthe lowest littoral, in combination with increased sed-imentation from the pelagic zone. Rosenvinge (1909–1931) frequently recorded algal vegetation at 30–35 mdepth in the Skagerrak, Kattegat, around the Danishislands and at Davids Banke north of Bornholm (BalticSea). Single plants of Phycodrys rubens and Deles-seria sanguinea were even reported from 40 m depth,Coccotylus truncatus from 36 m, and Furcellarialumbricalis, Odonthalia dentata and Membranopteraalata from 30, 29 and 27 m, respectively. Our deepest
find throughout was 27 m (P. rubens in the Skager-rak). As Rosenvinge’s investigations were made bydredging, a direct comparison with our depths is im-possible, but if Rosenvinge’s observations are correct,the lower limit of algal occurrence has moved upwardsby more that 10 m in less than 100 years. Svane &Gröndahl (1988) relocated an exposed diving stationof Gislén (1929, 1930) outside the Gullmar fjord inthe Skagerrak and they concluded that the lower limitof ‘high algal cover’ had moved upwards from 15 to12 m.
There are many pitfalls in long-term comparativestudies of the algal littoral vegetation caused by dif-ferences in methods, sites of investigation, personsand scientific focus. However, because we used thedownward process as a standard, we could partly re-late to our own data and conclude that the absenceof this event for species that previously did have adocumented downward process can be considered aresult of a general eutrophication of the open watersalong the Swedish west coast. This has previouslybeen shown in the northern Baltic Sea where the Fucusvesiculosus belt has moved upwards by several meterssince the 1940s, which was explained by increasedsedimentation, overgrowth by epiphytic annual algaeand increased shading by plankton (Kautsky et al.,1986; Eriksson et al., 1998). We did not find evidencefor such an upward shift in the vertical distributions ofthe large brown algae in the Skagerrak and Kattegatarea.
We have shown that an upward shift of opportun-istic red algae from the lower littoral in the exposedopen waters of the Kattegat and Skagerrak was appar-ent in 1989–1990. It might be argued that these resultsstill can be a result of the toxic phytoplankton bloomin 1988. However, in 1997 Johansson et al. (1998)obtained similar results from their exactly re-locateddiving profiles in the open Skagerrak. They found sig-nificant increases in abundance since 1960–1961 forBonnemaisonia hamifera, Cystoclonium purpureum,Delesseria sanguinea and Phycodrys rubens at thecost of Chondrus crispus and Furcellaria lumbricalis,and they attributed these changes to a general large-scale eutrophication of the Skagerrak. Delesseria san-guinea and Phycodrys rubens are also known to in-crease with local eutrophication in shallow bays in thewestern Baltic Sea (e.g. Breuer & Schramm, 1988).Johansson et al. (1998) also found Brongniartellabyssoides, Cystoclonium purpureum, D. sanguineaand P. rubens much higher up in the littoral zone thanKylin (1944, 1947, 1949) reported, but P. elongata

100
was always found below 6 m, which is in accordancewith Kylins observations. The slow-growing peren-nial red alga Furcellaria lumbricalis was previouslya dominant belt-forming alga in the Skagerrak andKattegat area below the Fucus and Laminaria belts(Kylin, 1907; Gislén, 1929, 1930; von Wachenfeldt1975). Now it has been largely replaced by the spe-cies typical of the Delesseria belt, which used to besituated below the Furcellaria belt (Kornfeldt, 1979;Breuer & Schramm, 1988; Johansson et al., 1998;this study). The Delesseria belt has probably movedupwards because of less light penetration through in-creased plankton production and increased turbidityof the water column with eutrophication along witha greater supply of nutrients. Another reason for thelarge decrease of F. lumbricalis may have been thelarge-scale harvesting of this species for agar produc-tion through extensive trawling in the 1950s–1970sof its loose-lying forma aegagropila which may haveplayed an important role in the dispersal of the speciesin the past (Austin, 1959).
Note
Unfortunately, some of the stone reefs in the Kat-tegat are subject to exploitation of the high-qualitysand and gravel that have accumulated in crevicesbetween the accumulations of stones and boulders.During the present study, deep craters filled with de-composing macroalgae, dead animals and hydrogensulphide resulting from such activities were observedat Stora Middelgrund. Efforts are now being made togive some of the stone reefs in the Kattegat the statusof marine reserves to protect them from exploitation(Nordheim & Boedeker 1998).
Acknowledgements
This project was financed by research grants fromthe Swedish World Wildlife Fund (WWF) and theSwedish Council for Planning and Coordination ofResearch (FRN) to M. Pedersén. Further spon-sors of the project were the Kattegat School inHalmstad (photographic equipment), Disab AB andAdab Diving AB (diving equipment), Kodak AB (pho-tographic films), French Food AB (food) and FalconAB (beverages). We are grateful to M. Wærn andS. Pekkari (Dept. of Plant Ecology, Uppsala Univer-sity), Å. Forsberg (Dept. of Plant Physiology, Uppsala
University), A. Tolstoy (Swedish Threatened SpeciesUnit, Agricultural University, Uppsala) and L. Math-iesen (Dept. of Botany, Århus University, Denmark)for scientific assistance during the fieldwork on boardof ‘Sunbeam’. Project assistant was Staffan Söder-lund. Laboratory assistant K. Österlund took most ofthe photographs. Å. Forsberg kindly lent his divingboat to the project. The following divers have carriedout most of the underwater fieldwork: J.-E. Axén, P.Berggren, M. Björk, J. Collén, K. Engell-Sörensen,Å. Forsberg, P. Haecky, K. Haglund, M. Knapp, C.Larsson, I. Strömberg, A. Tolstoy and K. Österlund.Responsible skippers during the research cruises were:S.-R. Ekman, L. Granath, P. Hagman, P. Hallert, P.Jonsson, M. Knapp and C.G. Velin. All these peopleare gratefully acknowledged as well as numerous otherscientists, students and visitors who have assisted inthe fieldwork during one or a few days.
References
Austin, A. P., 1959. Observations on Furcellaria fastigiata (L.)Lam. forma aegagropila Reinke in Danish waters together witha note on other unattached algal forms. Hydrobiologia 14:255–277.
Beardall, J., S. Beer & J. A. Raven, 1998. Biodiversity of marineplants in an era of climatic change: some predictions based onphysiological performance. Bot. mar. 41: 113–123.
Bokn, T. L., S. N. Murray, F. E. Moy & J. B. Magnusson,1992. Changes of fucoid distributions and abundances in theinner Oslofjord, Norway: 1974–1980 versus 1988–1990. ActaPhytogeogr. Suec. 78: 117–124.
Bird, C. J. & J. L. McLachlan, 1992. Seaweed Flora of the Mari-times 1. Rhodophyta – The Red Algae. Biopress Ltd, Bristol.177 pp.
Breuer, G. & W. Schramm, 1988. Changes in macroalgal vegeta-tion of Kiel Bight (Western Baltic Sea) during the past 20 years.Kieler Meeresforsch. Sonderh. 6: 241–255.
Diamond, J. & T. J. Case, 1986. Community Ecology. Harper &Row, New York.
Dring, M. J., A. Wagner, J. Boeskov & K. Lüning, 1996. Sensitivityof intertidal and subtidal red algae to UVA and UVB radiation, asmonitored by chlorophyll fluorescence measurement: influenceof collection depth and season, and length of irradiation. Eur. J.Phycol. 31: 293–302.
Eriksson, B. K., G. Johansson & P. Snoeijs, 1998. Long-termchanges in the sublittoral zonation of brown algae in the southernBothnian Sea. Eur. J. Phycol. 33: 241–249.
Fonselius, S., 1995. Västerhavets och Östersjöns oceanografi.SMHI, Norrköping. 200 pp.
Fowler, J., L. Cohen, & P. Jarvis, 1998. Practical Statistics for FieldBiology, 2nd edn. J. Wiley. 259 pp.
Gislén, T., 1929. Epibioses of the Gullmar Fjord I. A study in marinesociology. Skr. Ser. Kungl. Svenska Vetensk. Akad. 3: 1–123.
Gislén, T., 1930. Epibioses of the Gullmar Fjord II. A study inmarine sociology. Skr. Ser. Kungl. Svenska Vetensk. Akad. 4:1–380.

101
Hillis-Colinvaux L., 1986. Deep water populations of Halimeda inthe economy of an atoll. Bull. Mar. Sci. 38: 155–169.
Johansson, G., B. K. Eriksson, M. Pedersén & P. Snoeijs, 1998.Long-term changes of macroalgal vegetation in the Skagerrakarea. Hydrobiologia 385: 121–138.
Johannessen, T. & E. Dahl, 1996. Declines in oxygen concentrationsalong the Norwegian Skagerrak coast, 1927–1993: A signal ofecosystem changes due to eutrophication? Limnol. Oceanogr. 41:766–778.
Kaas, H., J. Larsen, F. Møhlenberg, & K. Richardson, 1991. TheChrysochromulina polylepis bloom in the Kattegat (Scandinavia)May–June 1988. Distribution, primary production and nutrientdynamics in the late stage of the bloom. Mar. Ecol. Prog. Ser. 79:151–161.
Kangas, P., H. Autio, G. Hällfors, H. Luther, Å. Niemi & H. Sale-maa, 1982. A general model of the decline of Fucus vesiculosusat Tvärminne, south coast of Finland in 1977–1981. Acta Bot.Fenn. 118: 1–27.
Karlsson, J., M. Kuylenstierna & P. Åberg, 1992. Contribution to theseaweed flora of Sweden: New or otherwise interesting recordsfrom the Swedish west coast. Acta Phytogeogr. Suec. 78: 49–63.
Kautsky, H., U. Kautsky & S. Nellbring, 1988. Distribution of floraand fauna in an area receiving pulp mill effluents in the BalticSea. Ophelia 28: 139–155.
Kautsky, N., H. Kautsky, U. Kautsky & M. Wærn, 1986. Decreaseddepth penetration of Fucus vesiculosus (L.) since the 1940s in-dicates eutrophication of the Baltic Sea. Mar. Ecol. Prog. Ser. 28:1–8.
Klavestad, N., 1978. The marine algae of the polluted inner part ofthe Oslofjord. Bot. mar. 21: 71–97.
Kornfeldt, R. A., 1979. Makroalgers biomassa längs Halland-skusten. Sv. Bot. Tidskr. 73: 131–138.
Kornfeldt, R. A., 1981. The biomass of seaweeds at the Kullen,S Sweden. In Levring, T. (ed.), Proc Xth Int. Seaweed Symp.,Berlin: 687–692.
Kornfeldt, R.A., 1984. Variation in distribution and biomass ofmarine benthic algae of Kullen, S Sweden. Nord. J. Bot. 4:563–584.
Kornmann, P., P.-H. Sahling, 1983. Meeresalgen von Helgoland,2nd edn. Biologische Anstalt Helgoland, Hamburg. 289 pp.
Kylin, H., 1907. Studien über die Algenflora der schwedischenWestküste. K.W. Appelbergs Buchdruckerei, Uppsala. 288 pp.
Kylin, H., 1918. Über Spermothamnion roseolum (Ag.) Pringsh. undTrailliella intricata Batters. Bot. Notiser 1916: 83–92.
Kylin, H., 1944. Die Rhodophyceen der Schwedischen Westküste.Lunds Univ. Årsskr. N.F., Avd. 2, 40(2): 1–104.
Kylin, H., 1947. Die Phaeophyceen der Schwedishen Westküste.Lunds Univ. Årsskr. N.F., Avd. 2, 43(4): 1–99.
Kylin, H., 1949. Die Chlorophyceen der Schwedischen Westküste.Lunds Univ. Årsskr. N.F., Avd. 2, 45(4): 1–79
Lindahl, O. & R. Rosenberg, 1989. The Chrysochromulina polylepisalgal bloom along the Swedish west coast 1988. Swed. Environ.Protect. Agency Rep. 3602: 1–71.
Littler, M. M. & D. S. Littler, 1980. The evolution of thallus formand survival strategies in benthic marine macroalgae: field andlaboratory tests of a functional form model. Am. Nat. 116: 25–44.
Lund, S., 1950. The marine algae of Denmark. Contributions to theirnatural history, Phaeophyceae IV. Biol. Skr. 6(2): 1–80.
Lundälv, T., C.S. Larsson & A. Axelsson, 1986. Long-term trendsin algal-dominated rocky subtidal communities on the Swedishwest coast - a transitional system. Hydrobiologia 142: 81–95.
Lüning, K., 1990. Seaweeds – Their Environment, Biogeographyand Ecophysiology. J. Wiley & Sons, New York: 527 pp.
Maestrini, S. Y. & E. Granéli, 1991. Environmental conditions andecophysiological mechanisms which led to the 1988 Chryso-chromulina polylepis bloom: an hypothesis. Oceanol. Acta 14:397–413.
Michanek, G., 1967. Quantitative sampling of benthic organisms bydiving on the Swedish west coast. Helgoländer. wiss. Meeresun-ters. 15: 455–467.
Nielsen, R. & K. Dahl, 1992a. Marine algae south of the islandVejrø, the Samsø area, Denmark. Acta Phytogeogr. Suec. 78:111–116.
Nielsen, R. & K. Dahl, 1992b. Macroalgae at Briseis Flak, SchultzsGrund and Store Middelgrund, stone reefs in the southern andeastern part of Kattegat, Denmark. In Bjørnestad, E., L. Hager-man & K. Jensen (eds), Proc. 12th Balt. Mar. Biol. Symp., Olsen& Olsen, Fredensborg, p. 109–118.
Nielsen, R., 1991. Vegetation of Tønneberg Banke, a stone reef inthe northern Kattegat, Denmark. Int. J. mar. Biol. Oceanogr. 17(Suppl 1): 199–211.
Nielsen, R., A. Kristiansen, L. Mathiesen & H. Mathiesen, 1995.Distributional index of the benthic macroalgae of the Baltic Seaarea. Acta Bot. Fenn. 155: 1–51.
Nordheim, H. & D. Boedeker, 1998. Red list of marine and coastalbiotopes and biotope complexes of the Baltic Sea, Belt Seaand Kattegat. Baltic Sea Environ. Proc., Helsinki Commission.115 pp.
Norin, L.-L. & M. Wærn, 1973. The zone of algal low standing cropnear Stockholm. Oikos, suppl. 15: 179–184.
Richardson, K. & Heilmann J.P., 1995. Primary production in theKattegat: past and present. Ophelia 41: 317–328.
Rosenberg, R., 1990. Negative oxygen trends in Swedish coastalbottom waters. Mar. Pollut. Bull. 21: 335–339.
Rosenberg, R., R. Elmgren, S. Fleischer, P. Johnsson, G. Persson &H. Dahlin, 1990. Marine eutrophication case studies in Sweden.Ambio 19: 102-110.
Rosenvinge, L. K., 1909–1931. The marine algae of Denmark. Con-tributions to their natural history, Rhodophyceae I–IV. Kongel.Danske Vidensk. Selsk. Skr. 7: 1–630.
Rosenvinge, L. K. & S. Lund, 1941–1947. The marine algae ofDenmark. Contributions to their natural history, PhaeophyceaeI–III. Biol. Skr. 1(4): 1–79, 2(6): 1–59, 4(5): 1–99
Rönnberg, O., K. Ådjers, C. Ruokolahti & M. Bondestam, 1992.Effects of fish farming on growth, epiphytes and nutrient contentof Fucus vesiculosus L. in the Åland archipelago, northern BalticSea. Aquat. Bot. 42: 109–120.
Rönnberg, Ö., 1981. Traffic effects on rocky-shore algae in the Ar-chipelago Sea, SW Finland. Acta Acad. Aboensis, Ser. B, 41(3):1–86.
Russell, G., 1991. Vertical distribution. In Mathieson, A. C. & P. H.Nienhuis (eds), Intertidal and Littoral Ecosystems. Ecosystemsof the World 24. Elsevier, Amsterdam: 43–65.
Schramm, W., 1996. The Baltic Sea and its transition zones. InSchramm W. & P. H. Nienhuis (eds), Marine Benthic Vegetation.Recent Changes and the Effects of Eutrophication, EcologicalStudies, 123: 131–163, Springer.
Serrão, E. A., S. H. Brawley, J. Hedman, L. Kautsky & G.Samuelsson, 1999. Reproductive success of Fucus vesiculosus(Phaeophyceae) in the Baltic Sea. J. Phycol. 35: 254–269.
Snoeijs, P., 1992. Ecology and taxonomy of Enteromorpha speciesin the vicinity of the Forsmark nuclear power plant (BothnianSea). Acta Phytogeogr. Suec. 78: 11–23
Snoeijs, P., 1999. Marine and brackish waters. In: Rydin, H., P.Snoeijs & M. Diekmann, (eds), Swedish Plant Geography. ActaPhytogeogr. Suec. 84: 187–212.

102
Snoeijs, P. & I. C. Prentice, 1989. Effects of cooling water dischargeon the structure and dynamics of epilithic algal communities inthe northern Baltic. Hydrobiologia 184: 99–123.
Svane, I. & F. Gröndahl, 1988. Epibioses of the Gullmarfjord: Anunderwater stereophotographical transect analysis in comparisonwith the investigations of Gislén in 1926–1929. Ophelia 28: 95–110.
ter Braak, C. J. F. & P. þmilauer, 1998. CANOCO Reference Manualand User’s Guide to Canoco for Windows: Software for Canon-ical Community Ordination (version 4). Microcomputer Power,Ithaca, NY, USA. 352 pp.
Vogt, H. & W. Schramm, 1991. Conspicuous decline of Fucus inKiel Bay (Western Baltic): what are the causes? Mar. Ecol. Prog.Ser. 69: 189–194.
von Wachenfeldt, T., 1975. Marine benthic algae and the environ-ment in the Öresund, Parts I-III. PhD-thesis, Lund University.
Wærn, M., 1952. Rocky-shore algae in the Öregrund archipelago.Acta Phytogeogr. Suec. 30: 1–298.
Wærn, M., 1965. A vista of the marine vegetation. Acta Phytogeogr.Suec. 50: 15–27.
Wallentinus, I., 1996. The Northern Atlantic Coasts (The SwedishWest Coast, Norway and Iceland). In Schramm W. & P. H. Nien-huis (eds), Marine Benthic Vegetation. Recent Changes and theEeffects of Eutrophication, Ecological Studies. Springer: 123:165–184.
Wennberg, T., 1987. Long-term changes in the composition anddistribution of the macroalgal vegetation in the southern part ofLaholm Bay, south-west Sweden, during the last thirty years.Swed. Envir. Protect. Agency Rep. 3290: 1–47.
Wassman, P., 1990. Calculating the load of organic carbon to theaphotic zone in eutrophicated coastal waters. Mar. Pollut. Bull.21: 183–187.
Woodward, F. I. & W. Cramer, 1996. Plant functional types andclimatic changes: Introduction. J. Veget. Sc. 7: 306–308.




















