
Patterns of interspecific associations of stem gallers on willows: Patterns of stem gallers on...
11
© 2003 Blackwell Publishing Ltd. http://www.blackwellpublishing.com/journals/ddi 443 BIODIVERSITY RESEARCH Diversity and Distributions (2003) 9, 443–453 Blackwell Publishing Ltd. Patterns of interspecific associations of stem gallers on willows JENS-PETER KOPELKE 1 *, JENS AMENDT 2 and KARSTEN SCHÖNROGGE 3 1 Forschungsinstitut Senckenberg, Senckenberganlage 25, D-60325 Frankfurt, Germany (correspondence address), 2 Johann Wolfgang Goethe Universität, Zentrum für Rechtsmedizin, Kennedyallee 104, D-60596 Frankfurt, Germany and 3 Centre of Ecology and Hydrology, CEH Dorset, Winfrith Technology Centre, Winfrith Newburgh, Dorchester, Dorset, DT2 8ZD, U.K. Abstract. The pattern of interspecific associa- tions of three stem-galling sawfly species (Euura atra, E. elaeagnos, E. purpureae) and three stem- galling gallmidge species ( Rabdophaga sp. 3–5) was investigated on five willow taxa (Salix alba, S. fragilis, S. × rubens, S. elaeagnos, S. purpurea) at five natural sites in Central Europe. The willow species harboured specific species associations of two stem gallers, each pair consisting of one Euura and one Rabdophaga species. The stem gallers were patchily distributed and their densi- ties varied significantly among willow host plant species, host plant individuals, and host plant sexes. Four of the six species showed a significant increase in galling rate with shoot length. The other two species were the sawfly and cecidomyiid pair that induce galls on S. purpurea. The pre- ference of stem gallers to longer shoots was gen- erally not related to higher larval performance in terms of survival. Only one species, Rabdophaga sp. 5, was found to be more abundant on male plants. The correlation of densities of the species pairs of stem gallers was independent of willow sexes. Species pairs of stem gallers co-occurring on the same willow species tended to attack different shoots within the same host plant in- dividual. When species pairs co-occurred on shoots they were usually found in similar densities as when occurring alone on shoots. The stem-galling sawflies usually formed galls at the basal part of a shoot, whereas the gallmidge R. sp. 5 (R. sp. 3 and R. sp. 4 showed no clear tendency) preferred the middle or distal part of a shoot. This is inter- preted with differences of their phenology and oviposition period. Key words. Cecidomyiidae, Distribution, Euura spp., interspecific association, plant vigour hypo- thesis, Rabdophaga spp., Salix spp., stem galler, Tenthredinidae. INTRODUCTION Gall-forming sawflies and gallmidges on willows (Salix spp.) are highly specialized herbivores, inducing their galls in young, meristematic tissue of different plant organs (Price & Craig, 1984; Clancy et al., 1986; Clancy & Price, 1989; Harris, 1994; Kopelke, 1998, 1999). The stem-galling midges of the genus Rabdophaga (Cecidomyiidae) and the sawfly genus Euura (Tenthredinidae) exhibit distinct host plant specificity by attacking exclusively a single willow species except for E. atra and Rabdophaga sp. 4 which attack three willow species (Kopelke, 1990, 1998, 1999; Kopelke & Amendt, 2002). Gall formation represents arguably the most intimate form of plant–herbivore interaction. In many galls, including sawfly and gall midge galls, the gall former not only affects the develop- mental pattern of a whole or part of a plant organ, but also controls, to at least some extent, the physiology of the plant tissue surrounding the * Corresponding author. E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Patterns of interspecific associations of stem gallers on willows: Patterns of stem gallers on...

© 2003 Blackwell Publishing Ltd. http://www.blackwellpublishing.com/journals/ddi 443
BIODIVERSITY RESEARCH
Diversity and Distributions (2003) 9, 443–453
Blackwell Publishing Ltd.
Patterns of interspecific associations of stem gallers on willowsJENS-PETER KOPELKE1*, JENS AMENDT 2 and KARSTEN SCHÖNROGGE3 1Forschungsinstitut Senckenberg, Senckenberganlage 25, D-60325 Frankfurt, Germany (correspondence address), 2Johann Wolfgang Goethe Universität, Zentrum für Rechtsmedizin, Kennedyallee 104, D-60596 Frankfurt, Germany and 3Centre of Ecology and Hydrology, CEH Dorset, Winfrith Technology Centre, Winfrith Newburgh, Dorchester, Dorset, DT2 8ZD, U.K.
Abstract. The pattern of interspecific associa-tions of three stem-galling sawfly species (Euuraatra, E. elaeagnos, E. purpureae) and three stem-galling gallmidge species (Rabdophaga sp. 3–5)was investigated on five willow taxa (Salix alba,S. fragilis, S. × rubens, S. elaeagnos, S. purpurea)at five natural sites in Central Europe. The willowspecies harboured specific species associationsof two stem gallers, each pair consisting of oneEuura and one Rabdophaga species. The stemgallers were patchily distributed and their densi-ties varied significantly among willow host plantspecies, host plant individuals, and host plantsexes. Four of the six species showed a significantincrease in galling rate with shoot length. Theother two species were the sawfly and cecidomyiidpair that induce galls on S. purpurea. The pre-ference of stem gallers to longer shoots was gen-erally not related to higher larval performance interms of survival. Only one species, Rabdophaga
sp. 5, was found to be more abundant on maleplants. The correlation of densities of the speciespairs of stem gallers was independent of willowsexes. Species pairs of stem gallers co-occurringon the same willow species tended to attackdifferent shoots within the same host plant in-dividual. When species pairs co-occurred on shootsthey were usually found in similar densities aswhen occurring alone on shoots. The stem-gallingsawflies usually formed galls at the basal part ofa shoot, whereas the gallmidge R. sp. 5 (R. sp. 3and R. sp. 4 showed no clear tendency) preferredthe middle or distal part of a shoot. This is inter-preted with differences of their phenology andoviposition period.
Key words. Cecidomyiidae, Distribution, Euuraspp., interspecific association, plant vigour hypo-thesis, Rabdophaga spp., Salix spp., stem galler,Tenthredinidae.
INTRODUCTION
Gall-forming sawflies and gallmidges on willows(Salix spp.) are highly specialized herbivores,inducing their galls in young, meristematic tissueof different plant organs (Price & Craig, 1984;Clancy et al., 1986; Clancy & Price, 1989; Harris,1994; Kopelke, 1998, 1999). The stem-gallingmidges of the genus Rabdophaga (Cecidomyiidae)and the sawfly genus Euura (Tenthredinidae)
exhibit distinct host plant specificity by attackingexclusively a single willow species except forE. atra and Rabdophaga sp. 4 which attack threewillow species (Kopelke, 1990, 1998, 1999; Kopelke& Amendt, 2002).
Gall formation represents arguably the mostintimate form of plant–herbivore interaction. Inmany galls, including sawfly and gall midge galls,the gall former not only affects the develop-mental pattern of a whole or part of a plant organ,but also controls, to at least some extent, thephysiology of the plant tissue surrounding the* Corresponding author. E-mail: [email protected]

444 J.-P. Kopelke, J. Amendt and K. Schönrogge
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
feeding larva or adult (Harris, 1994; Higton &Mabberly, 1994; Leitch, 1994; Schönrogge et al.,2000; Stone et al., 2002). Thus, many factors thatmight affect the distribution of free-feeding insectlarvae, such as the distribution of secondarychemistry within or between host plants, environ-mental and weather-related factors, are likely toaffect galling herbivores less in their feedingstages. However, genetic variation, hybridization,and possibly plant volatile emissions as an indi-cator of the former, can affect the ovipositionbehaviour of the females and influence gall dis-tributions in this way.
One reoccurring determinant of abundancepatterns among gallers has been proposed inthe ‘plant-vigour hypothesis’ (Price et al., 1987a;Price, 1991). This suggests that host plant organs(i.e. active meristems) that show the most vigor-ous growth are likely to provide the best habitatfor gallers (Price & Ohgushi, 1995; Price et al.,1997; Fritz et al., 2000). It suggests that vigor-ously growing plant organs are best supplied with,for instance, water and nutrients and are thus bestsuited for the fast growing gall tissues and theirinhabitants. Physiologically there is no directevidence that galls will benefit from a high levelof supply to the surrounding tissue, although gallsinduced by insects from different taxonomicgroups have been shown to be physiological sinks(Shorthouse & Rohfritsch, 1992; Bagatto et al., 1991,1997; but see Hartley & Lawton, 1992). However,the implication of the plant-vigour hypothesis,that gallers should prefer vigorously growing plantorgans and for stem gallers to prefer the relativelyrarer long shoots over shorter ones, has been foundto be true for a number of different and taxo-nomically diverse galling insects (Tenthredinidae— Roininen & Danell, 1997; Adelgidae — McKinnonet al., 1999; Cynipidae — Pieres & Price, 2000;Cecidomyiidae — Cornelissen & Fernandes, 2001;Psyllidae — Faria & Fernandes, 2001 — but alsosee de Bruyn, 1995). While the evidence for theplant-vigour hypothesis has become more general,it is not clear what gives rise to this pattern. Itis possible that eggs laid on vigorous shoots aremore likely to develop into galls but fail to do soon shorter shoots. Alternatively, females mightselect vigorous shoots at the time of oviposition,which would presumably mean that they couldidentify vigour in active meristems, since theshoots would not have grown at this time.
Another nonexclusive hypothesis addressingherbivore abundance distributions states thatherbivore attack patterns are determined by thesex of the host plant. In some willow species,sex-related differences in both growth (Danellet al., 1991; Elmqvist et al., 1991) and defence (Priceet al., 1989; Hjältén, 1992), have been observed.Some studies have also documented intersexualvariation in herbivory on dioecious plants. The mostcommon pattern reported is for male plants tosupport higher densities of herbivores (Boecklenet al., 1990, 1994; Hjältén, 1992; Fritz, 1995).
Both the plant-vigour hypothesis and the sex-bias hypothesis refer to relationships betweenherbivores and host plants. Little attention hasbeen given to whether variation in plant charactersmight affect interspecific competition betweenherbivorous insects. Different species of herbi-vores might respond in different ways to variationamong host plants, leading to positive or negativecorrelations in species abundances, respectively(Fritz et al., 1986, 1987b; Boecklen & Price, 1989;Fritz & Price, 1990; Fritz, 1990; Price et al., 1999).This could result in different assortments ofspecies among modules within plants or differentphenotypes. Interspecific competition has beena major paradigm in community ecology, butsedentary insects, such as gall formers, stemborers, and leaf miners have shown little evidenceof interspecific competition at naturally occurringdensities (Fritz et al., 1987a).
In this study we investigate patterns of stemgaller attack in five natural willow stands alongthe floodplains of the Rhine River in CentralEurope. Of the three Euura species, E. atra occurson Salix alba, S. fragilis and the hybrid S. × rubens,while E. elaeagnos is specific to S. elaeagnos,and E. purpureae is specific to S. purpurea.Intriguingly, the three Rhabdophaga speciesmirror the host specificity of the sawfliesexactly (R. sp. 3 on S. purpurea, R. sp. 4 onS. alba, S. fragilis, and S. × rubens, R. sp. 5 onS. elaeagnos) so that although the species co-occurlocally, there are three distinct species pairs oneach host plant species each of one stem-gallingsawfly and one stem-galling Rhabdophaga species.We establish the distribution of abundance, sur-vival, and parasitoid attack patterns within theguild of six stem-galling species in the light of theplant-vigour hypothesis, while also testing fordifferences in galling rates on male and female

Patterns of stem gallers on willows 445
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
plants. We investigate interrelations between thepartners in the three species pairs of Euura andRhabdophaga that share the same host plant,where we test whether or not the gall formers ofan associated species pair attack shoots of thesame length or the same parts (basal, median,distal) within a shoot (i.e. niche separation).
METHODS
Stem-galling species
The gall-forming sawflies and gallmidges of thisstudy (Euura spp., Rabdophaga spp.) developand pupate in closed stem galls. The sawfliesinitiate gall formation by the female’s injection ofstimulating substances ( Mccalla et al., 1962; Meyer,1987; Kopelke, 1998, 1999), while gallmidgesinduce gall formations by the larvae’s secretionof cecidogenous fluids while feeding on theplant tissue (Meyer, 1987; Harris, 1994). Afteremerging in spring the sawflies are immediatelyready for mating and oviposition. In contrast, thegallmidges require some days for eggs to mature.Both the stem-galling Euura and Rabdophagaspecies produce one generation each year.
Stem gallers were studied on the willow speciesSalix alba, S. fragilis, S. × rubens (= alba × fragilis),S. elaeagnos, and S. purpurea from several naturalsites. Six stem-galling species have been studied(E. atra, E. elaeagnos, E. purpureae, Rabdophagasp. 3–5), most of which induce moderatelyspindle-shaped galls, except for E. elaeagnos, whichforms conspicuous spindle-shaped galls. In thecase of the stem-galling Rabdophaga spp. thetaxonomical status of the species has not yetbeen clarified (Kopelke & Amendt, 2002). There aresome indications that sibling species on differentwillow species may be involved; in the presentstudy they are designated as Rabdophaga sp. 3–5.
Sampling of stem gallers
The studies were conducted in 1999 at fivelocations along the river Rhine (Central Europe:Germany and Switzerland) (Fig. 1). The com-position, age, and abundances of the Salixspecies varied greatly between the stands. A total of635 individual willow trees were mapped in spring,checking the sexes when the catkins occurred. Onlyin a few species were the sex ratios balanced.
Most of the species at all sites were dominated byfemale trees ( Kopelke & Amendt, 2002). Accord-ing to the age pattern of the host plant commu-nities there was not much difference between thesites at the lower (site no. 1 and 2) and upperRhine (site no. 3 and 4), the communities beingcomposed especially of older plants of S. caprea,S. elaeagnos, S. fragilis, S. purpurea, S. × rubens,and S. viminalis.
To determine plant vigour the length of 4912shoots from 82 willow individuals of different ageand sex (sites combined, alba: 18, elaeagnos: 32,fragilis: 5, purpurea: 12, × rubens: 15) were meas-ured, and grouped into 6 shoot length classes(5–10, 11–20, 21–30, 31–40, 41–60, 61-infinitymm). By the end of growing season at least 50shoots (= the current year’s growth of a stemwith leaves) were collected haphazardly from allaspects of each plant. The shoots were keptrefrigerated in plastic bags to preserve the stageof development until dissection. All shoots weresplit longitudinally to check the status of the
Fig. 1 Map of the sites with willow species studied,adjacent to the Rhine River in Central Europe.

446 J.-P. Kopelke, J. Amendt and K. Schönrogge
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
herbivores (alive, dead, parasitized) and theposition of the galls (basal third, median, or distalthird) were recorded.
Statistics
We used two different types of general linearmodels (GLM) both of which were nested to theappropriate level. To assess galling rates withshoot length we used a square-root–square-roottransformation for the count data and a nested where, for E. atra and R. sp. 4, shootlength was nested within tree within sex (of thewillow trees) within Salix species and withinlocality. Significances at all levels were assessedby calculating F-ratios from the appropriatemean-squares and degrees of freedom. Since theother 4 species were specific to only one Salixspecies those analyses had one nesting level less.
Survival and parasitoid attack were analysedas nested analyses of deviance using GLMs withbinomial errors. Significance was assessed onthe deletion of terms using χ2 or in some casesF-ratios to correct for overdispersion.
Finally, correlations of galling rates among thepaired species were analysed as nested analysesof deviance using GLMs with poisson errors.Significance was assessed as described above. Wealso assessed the likelihood of occurrence of onespecies in relation to the other as a binary (pres-ence/absence) variable again using GLMs withbinomial errors.
For all models the standardized residuals andfitted values of the minimal adequate modelswere inspected for independence and normality(Crawley, 2002). Please note that the figures showoverall relationships (not nested), while all thetest statistics were carried out nested.
RESULTS
Plant vigour and gall densities
Of the six gall-inducing species, four showed asignificant higher galling rate on longer shootsthan on short ones. The four species includeE. atra and R. sp. 4 on the three willow speciesS. alba, S. fragilis and S. rubens, and E. elaeagnosand R. sp. 5 on S. elaeagnos. The two speciesthat showed no increase in galling rates withshoot length were the cecidomyiid R. sp. 3 and
E. purpureae, both on S. purpurea (Table 1, Fig. 2).Both species also have the lowest overall gallingrates (E. purpureae 0.046 ± 0.008 galls per shootand R. sp. 3 0.042 ± 0.009 galls per shoot) whichis about half the galling rate of the next lowestone of E. elaeagnos (0.083 ± 0.008) and about aquarter compared to E. atra (0.211 ± 0.014) andR. sp. 4 and R. sp. 5 (0.154 ± 0.009 and 0.139 ±0.013 galls per shoot).
Gall densities in all six species were analysednested within shoot length within trees withinmale/female willows and within localities. For thetwo species that lacked a relationship with shootlength it was still evident that galling rates differedsignificantly among individual trees (Table 1).
For one of the Rhabdophaga species, R. sp. 5,galling rates on male plants were significantlyhigher than on female plants. At location three(see Fig. 1) the mean number of galls per shooton male willows was 0.375 ± 0.07, which wastwice that on the female plants with 0.16 ± 0.084.At location four the difference was threefold(male: 0.644 ± 0.136 and female: 0.20 ± 0.046),while at location five galling rates were aboutthe same and very low (male: 0.009 ± 0.005 andfemale: 0.009 ± 0.005).
In contrast to the other four species, E. atraand R. sp. 4 induce galls on three different spe-cies of willow. There is, however, no indicationthat galling rates of either species differ amongthe willow species (Table 1, Fig. 2) nor in factamong the different localities that were sampled.
Plant vigour, survival and parasitoid attack
Gall former survival and parasitoid attack rateswere analysed as logistic regression with shootlength nested within the other variables as before.Of the six species, only R. sp. 5 on S. elaeagnosshowed a significant negative correlation betweenshoot length and survival, i.e. the proportion ofgalls that would produce an adult gall former(χ2 = 27.99, d.f. = 10, P < 0.01, Fig. 3).
Parasitoid attack (parasitoid species com-bined), as one source of mortality, was also sig-nificantly correlated with shoot length for onlyone of the six species, E. purpureae ( χ2 = 16.91,d.f. = 7, P < 0.05). Here parasitoid attack rateswere lower on the longer shoots (Fig. 4). Therelationship between parasitoid attack rates andshoot length for at least one of the cecidomyiidae,

Patterns of stem gallers on willows 447
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
R. sp. 4, was also close to significance (P = 0.06),and again the trend was negative with lowerattack rates on longer shoots.
Interspecific patterns of galling rates
To investigate possible relationships of theabundance among co-occurring partners in eachof the species pairs we looked at both the gallabundance of one partner in relation to theabundance of the other (nested GLM with poissonerrors) and the likelihood of one partner occur-ring on the same shoot as the other using pres-ence/absence data rather than gall counts (nestedGLM with binomial errors). All six models wereagain nested within all the levels described beforeand are shown in Table 1.
The relationships between the likelihood ofoccurrences were consistent with those for abun-dances (E. atra — R. sp. 4: χ2 = 322.49, d.f. = 33,P < 0.001; E. elaeagnos — R. sp. 5: χ2 = 155.93,d.f. = 16, P < 0.001, and E. purpureae — R. sp. 3:χ2 = 47.38, d.f. = 7, P < 0.01). Within all threespecies pairs there are significant negative rela-tionships between the abundances of the Cecid-omyiids and the sawflies (E. atra — R. sp. 4:χ2 = 156.41, d.f. = 35, P < 0.001; E. elaeagnos —R. sp. 5: χ2 = 77.97, d.f. = 18, P < 0.001, andE. purpureae — R. sp. 3: χ2 = 22.48, d.f. = 8, P <0.01). (Fig. 5) illustrates the overall relationshipswithout nesting and shows that data points(= shoots) where both species of a pair co-occurare comparatively rare and at high density of oneof the partners virtually absent.
Table 1 Parameters used in nested s for assessing galling rates with shoot length are: shoot lengthnested within tree within sex (of the willow trees) within Salix species (only Euura atra and Rabdophaga sp.4) and within locality
Species Parameter d.f. F P<
E. atra Location 3,1 3.9 NSSalix within location 1,4 0.4 NSSex within Salix within location 4,29 1.74 NSTree within Sex within Salix within Location 29,141 1.41 NSLength (cat) within Tree within Sex within Salix within Location 141,2125 3.1 0.001
R. sp. 4 Location 3,1 10.85 NSSalix within location 1,4 0.073 NSSex within Salix within location 4,29 0.072 NSTree within Sex within Salix within Location 29,141 4.09 0.001Length (cat) within Tree within Sex within Salix within Location 141,2125 1.7 0.001
E. elaeagnos Location 2,3 2.45 NSSex within Location 3,25 0.66 NSTree within Sex within Location 25,128 4.13 0.001Length (cat) within Tree within Sex within Location 128,1620 1.24 0.001
R. sp. 5 Location 2,3 3.19 NSSex within Location 3,25 9.37 0.001Tree within Sex within Location 25,128 2.04 0.05Length (cat) within Tree within Sex within Location 128,1620 1.83 0.001
E. purpuerae Location 1,1 3.1 NSSex within Location 1,9 0.24 NSTree within Sex within Location 9,55 4.45 0.05Length (cat) within Tree within Sex within Location 55,762 0.98 NS
R. sp. 3 Location 1,1 5.2 NSSex within Location 1,9 0.43 NSTree within Sex within Location 9,55 2.93 0.05Length (cat) within Tree within Sex within Location 55,762 1.35 NS
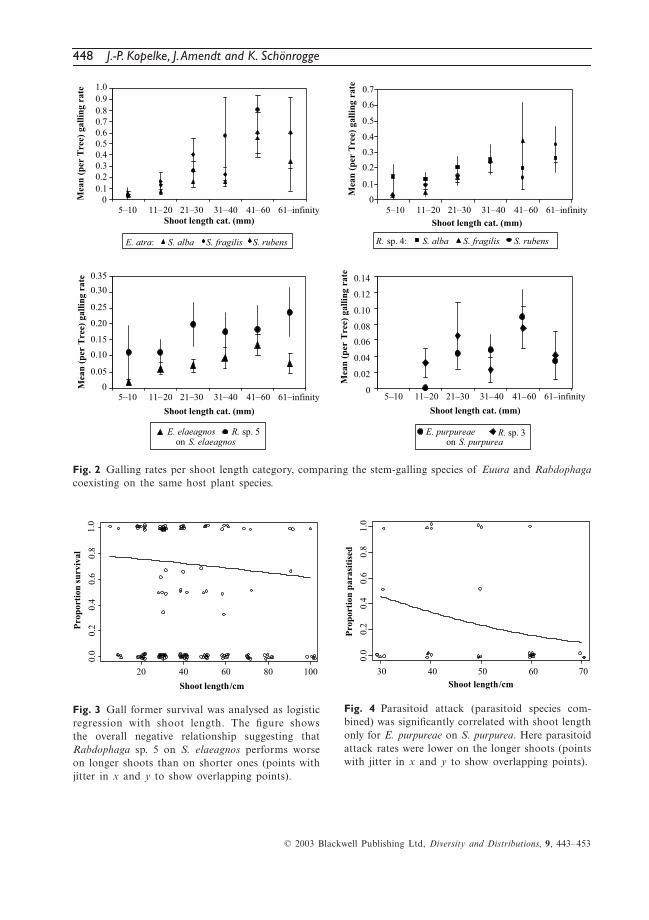
448 J.-P. Kopelke, J. Amendt and K. Schönrogge
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
Fig. 3 Gall former survival was analysed as logisticregression with shoot length. The figure showsthe overall negative relationship suggesting thatRabdophaga sp. 5 on S. elaeagnos performs worseon longer shoots than on shorter ones (points withjitter in x and y to show overlapping points).
Fig. 2 Galling rates per shoot length category, comparing the stem-galling species of Euura and Rabdophagacoexisting on the same host plant species.
Fig. 4 Parasitoid attack (parasitoid species com-bined) was significantly correlated with shoot lengthonly for E. purpureae on S. purpurea. Here parasitoidattack rates were lower on the longer shoots (pointswith jitter in x and y to show overlapping points).
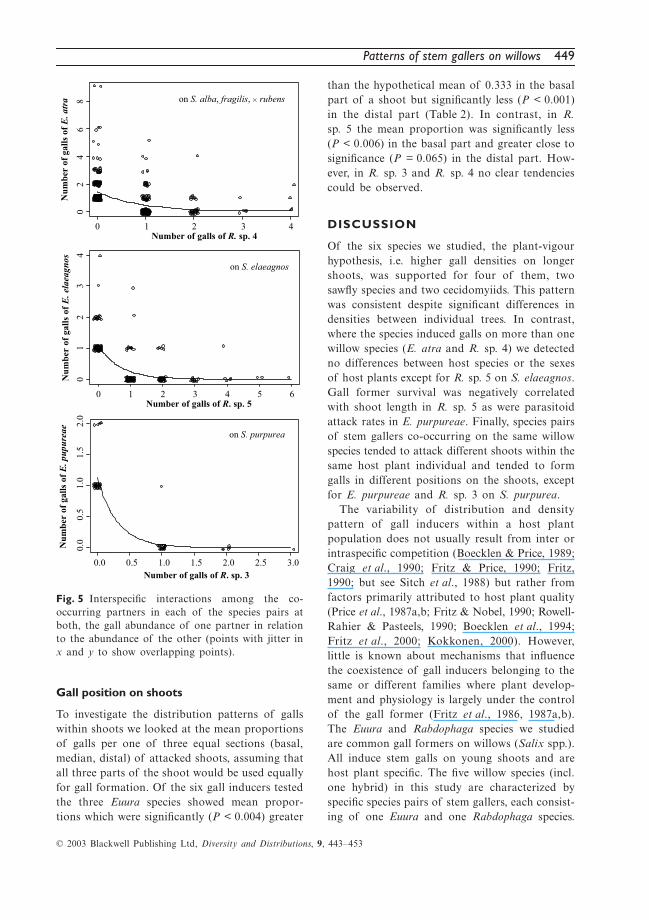
Patterns of stem gallers on willows 449
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
Gall position on shoots
To investigate the distribution patterns of gallswithin shoots we looked at the mean proportionsof galls per one of three equal sections (basal,median, distal) of attacked shoots, assuming thatall three parts of the shoot would be used equallyfor gall formation. Of the six gall inducers testedthe three Euura species showed mean propor-tions which were significantly (P < 0.004) greater
than the hypothetical mean of 0.333 in the basalpart of a shoot but significantly less (P < 0.001)in the distal part (Table 2). In contrast, in R.sp. 5 the mean proportion was significantly less(P < 0.006) in the basal part and greater close tosignificance (P = 0.065) in the distal part. How-ever, in R. sp. 3 and R. sp. 4 no clear tendenciescould be observed.
DISCUSSION
Of the six species we studied, the plant-vigourhypothesis, i.e. higher gall densities on longershoots, was supported for four of them, twosawfly species and two cecidomyiids. This patternwas consistent despite significant differences indensities between individual trees. In contrast,where the species induced galls on more than onewillow species (E. atra and R. sp. 4) we detectedno differences between host species or the sexesof host plants except for R. sp. 5 on S. elaeagnos.Gall former survival was negatively correlatedwith shoot length in R. sp. 5 as were parasitoidattack rates in E. purpureae. Finally, species pairsof stem gallers co-occurring on the same willowspecies tended to attack different shoots within thesame host plant individual and tended to formgalls in different positions on the shoots, exceptfor E. purpureae and R. sp. 3 on S. purpurea.
The variability of distribution and densitypattern of gall inducers within a host plantpopulation does not usually result from inter orintraspecific competition (Boecklen & Price, 1989;Craig et al., 1990; Fritz & Price, 1990; Fritz,1990; but see Sitch et al., 1988) but rather fromfactors primarily attributed to host plant quality(Price et al., 1987a,b; Fritz & Nobel, 1990; Rowell-Rahier & Pasteels, 1990; Boecklen et al., 1994;Fritz et al., 2000; Kokkonen, 2000). However,little is known about mechanisms that influencethe coexistence of gall inducers belonging to thesame or different families where plant develop-ment and physiology is largely under the controlof the gall former (Fritz et al., 1986, 1987a,b).The Euura and Rabdophaga species we studiedare common gall formers on willows (Salix spp.).All induce stem galls on young shoots and arehost plant specific. The five willow species (incl.one hybrid) in this study are characterized byspecific species pairs of stem gallers, each consist-ing of one Euura and one Rabdophaga species.
Fig. 5 Interspecific interactions among the co-occurring partners in each of the species pairs atboth, the gall abundance of one partner in relationto the abundance of the other (points with jitter inx and y to show overlapping points).
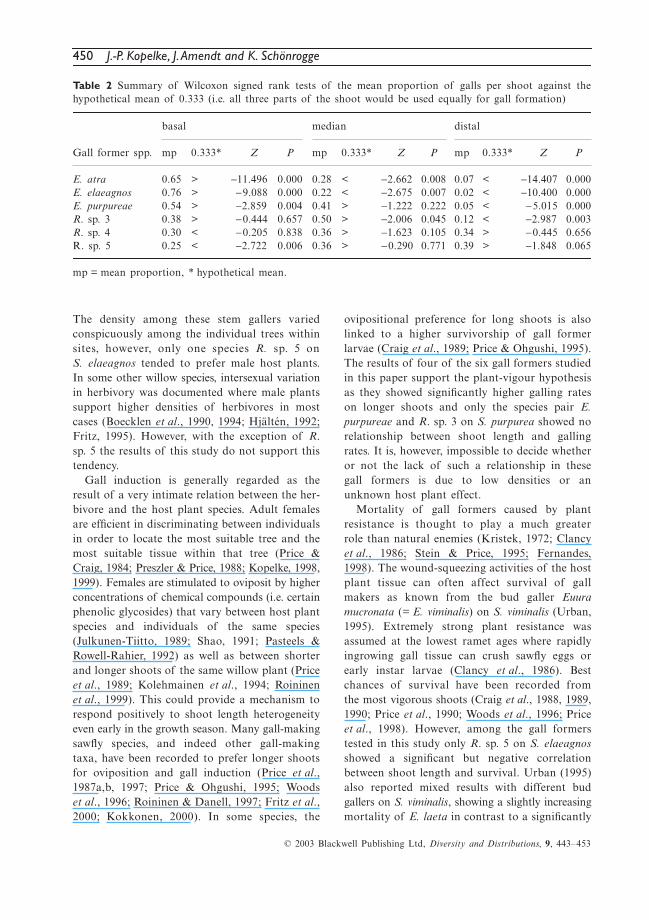
450 J.-P. Kopelke, J. Amendt and K. Schönrogge
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
The density among these stem gallers variedconspicuously among the individual trees withinsites, however, only one species R. sp. 5 onS. elaeagnos tended to prefer male host plants.In some other willow species, intersexual variationin herbivory was documented where male plantssupport higher densities of herbivores in mostcases (Boecklen et al., 1990, 1994; Hjältén, 1992;Fritz, 1995). However, with the exception of R.sp. 5 the results of this study do not support thistendency.
Gall induction is generally regarded as theresult of a very intimate relation between the her-bivore and the host plant species. Adult femalesare efficient in discriminating between individualsin order to locate the most suitable tree and themost suitable tissue within that tree (Price &Craig, 1984; Preszler & Price, 1988; Kopelke, 1998,1999). Females are stimulated to oviposit by higherconcentrations of chemical compounds (i.e. certainphenolic glycosides) that vary between host plantspecies and individuals of the same species(Julkunen-Tiitto, 1989; Shao, 1991; Pasteels &Rowell-Rahier, 1992) as well as between shorterand longer shoots of the same willow plant (Priceet al., 1989; Kolehmainen et al., 1994; Roininenet al., 1999). This could provide a mechanism torespond positively to shoot length heterogeneityeven early in the growth season. Many gall-makingsawfly species, and indeed other gall-makingtaxa, have been recorded to prefer longer shootsfor oviposition and gall induction (Price et al.,1987a,b, 1997; Price & Ohgushi, 1995; Woodset al., 1996; Roininen & Danell, 1997; Fritz et al.,2000; Kokkonen, 2000). In some species, the
ovipositional preference for long shoots is alsolinked to a higher survivorship of gall formerlarvae (Craig et al., 1989; Price & Ohgushi, 1995).The results of four of the six gall formers studiedin this paper support the plant-vigour hypothesisas they showed significantly higher galling rateson longer shoots and only the species pair E.purpureae and R. sp. 3 on S. purpurea showed norelationship between shoot length and gallingrates. It is, however, impossible to decide whetheror not the lack of such a relationship in thesegall formers is due to low densities or anunknown host plant effect.
Mortality of gall formers caused by plantresistance is thought to play a much greaterrole than natural enemies (Kristek, 1972; Clancyet al., 1986; Stein & Price, 1995; Fernandes,1998). The wound-squeezing activities of the hostplant tissue can often affect survival of gallmakers as known from the bud galler Euuramucronata (= E. viminalis) on S. viminalis (Urban,1995). Extremely strong plant resistance wasassumed at the lowest ramet ages where rapidlyingrowing gall tissue can crush sawfly eggs orearly instar larvae (Clancy et al., 1986). Bestchances of survival have been recorded fromthe most vigorous shoots (Craig et al., 1988, 1989,1990; Price et al., 1990; Woods et al., 1996; Priceet al., 1998). However, among the gall formerstested in this study only R. sp. 5 on S. elaeagnosshowed a significant but negative correlationbetween shoot length and survival. Urban (1995)also reported mixed results with different budgallers on S. viminalis, showing a slightly increasingmortality of E. laeta in contrast to a significantly
Table 2 Summary of Wilcoxon signed rank tests of the mean proportion of galls per shoot against thehypothetical mean of 0.333 (i.e. all three parts of the shoot would be used equally for gall formation)
Gall former spp.
basal median distal
mp 0.333* Z P mp 0.333* Z P mp 0.333* Z P
E. atra 0.65 > −11.496 0.000 0.28 < −2.662 0.008 0.07 < −14.407 0.000E. elaeagnos 0.76 > −9.088 0.000 0.22 < −2.675 0.007 0.02 < −10.400 0.000E. purpureae 0.54 > −2.859 0.004 0.41 > −1.222 0.222 0.05 < −5.015 0.000R. sp. 3 0.38 > −0.444 0.657 0.50 > −2.006 0.045 0.12 < −2.987 0.003R. sp. 4 0.30 < −0.205 0.838 0.36 > −1.623 0.105 0.34 > −0.445 0.656R. sp. 5 0.25 < −2.722 0.006 0.36 > −0.290 0.771 0.39 > −1.848 0.065
mp = mean proportion, * hypothetical mean.

Patterns of stem gallers on willows 451
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
decreasing mortality of E. mucronata (= E. vimi-nalis) with increasing shoot length. There isprobably no general pattern that may explain thepattern of survival of gall formers within a pop-ulation of variable shoot length.
Parasitoid attack, as another source of mortal-ity, was correlated with shoot length only inE. purpureae, showing lower parasitoid attack rateson longer shoots. Thus, either parasitoid attack isnot or inverse density dependent, or the galls ona shoot are not regarded as a patch by parasitoidfemales. Considering that the oviposition sites,i.e. buds, on a long shoot are more spaced outthan on short ones what is recognized as a patchmight differ between short and long shoots, mak-ing shoot as a unit inappropriate to assess spatialdensity dependence.
Within all three species pairs tested in thisstudy, significant negative relationships betweenthe abundances of the cecidomyiids and the saw-flies have been found. It stands to argue whetherthese relationships are suggestive of directcompetition, i.e. adult females that fight for oviposi-tion sites, or pre-emption and avoidance. E. atraand R. sp. 4 attacked 6% of the shortest and 43%of the longest shoots while E. elaeagnos andR. sp. 5 attacked 6% of the shortest and 21% ofthe longest shoots. All four species showed posi-tive relationships between galling rates and shootlength suggesting that at these gall densities thegall former — host plant relationship is still over-riding any possible changes of gall distributionsdue to interspecific interactions. We also showed thatinterspecific competition between co-occurringspecies pairs was not only minimized by attack-ing different shoots within the same willowindividual but by attacking different parts of ashoot. Euura spp. favoured especially the basalparts of a shoot, whereas at least one of the Rab-dophaga spp. preferred the median and distalparts. This could be due to the different pheno-logies of sawflies and gallmidges. The stem-gallingEuura species emerge earlier in spring than thestem-galling Rabdophaga species, and the femalesare able to oviposit immediately after hatching(Kopelke, 1998). The gallmidges emerge later andfemales need some time for eggs to maturate.Both gall-forming genera are likely to prefer theapical meristem for oviposition. As the shootcontinuous to grow on an autumn shoot the ear-lier Euura galls would be located in the basal,
while the later Rhabdophaga galls would be in themedian and distal sections.
Patchy distribution patterns as well as largevariation of gall former densities among a willowpopulation indicate different attractiveness of thehost plants. Among the factors influencing thedistribution and density of gall formers plant vig-our and possibly associated physiological factorsand/or the impact of natural enemies is likely tobe important. Although they utilise the samehost plant resource (individual, shoot) inter-specific competition in this system of co-occurringstem gallers is unlikely due to phenological dif-ferences that translate into spatial separation onindividual shoots.
ACKNOWLEDGMENTS
This study was supported by the national sciencefoundation (DFG, J.-P.K. & J.A.) and the Natu-ral Environment Research Council (GR3/12847),UK (K.S.).
REFERENCES
Bagatto, G. & Shorthouse, J.D. (1997) Accumula-tion of mineral nutrients by the galler Hemadasnubilipennis (Hymenoptera: Pteromalidae) and itsparasitoids on Lowbush Blueberry: Implicationsof feeding behaviour. In: Ecology and Evolution ofPlant-Feeding Insects in Natural and Man-MadeEnvironments (ed. A. Raman), pp. 159–168. Inter-national Scientific Publications, New Delhi.
Bagatto, G., Zmijowskyj, T.J. & Shorthouse, J.D.(1991) Galls induced by Diplolepis spinosa influ-ence distribution of mineral nutrients in the shrubRose. Hortscience, 20, 1283–1284.
Boecklen, W.J., Mopper, S. & Price, P. (1994)Sex-biased herbivory in arroyo willow: are theregeneral patterns among herbivores? Oikos, 71,267–272.
Boecklen, W.J. & Price, P.W. (1989) Size and shapeof sawfly assamblages on arroyo willow. Ecology,70, 1463–1471.
Boecklen, W.J., Price, P.W. & Mopper, S. (1990) Sexand drugs and herbivores: Sex-biased herbivory inArroyo willow (Salix lasiolepis). Ecology, 71, 581–588.
de Bruyn, L. (1995) Plant stress and larval perform-ance of a dipterous gall former. Oecologia, 101,461–466.
Clancy, K.M. & Price, P.W. (1989) Effect of plant re-sistance, competition, and enemies on a leaf-gallingsawfly (Hymenoptera: Tenthredinidae). Environ-mental Entomology, 18, 284–290.

452 J.-P. Kopelke, J. Amendt and K. Schönrogge
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
Clancy, K.M., Price, P.W. & Craig, T.P. (1986) Lifehistory and natural enemies of an undescribedsawfly near Pontania pacifica (Hymenoptera:Tenthredinidae) that forms leaf galls on arroyowillow, Salix lasiolepis. Annals of the EntomologicalSociety of America, 79, 884–892.
Cornelissen, T.G. & Fernandes, G.W. (2001) Patternsof attack by herbivores on the tropical shrubBauhinia brevipes (Leguminosae): Vigour orchance? European Journal of Entomology, 98, 37–40.
Craig, T.P., Itami, J.K. & Price, P.W. (1989) A strongrelationship between oviposition preference andlarval performance in a shoot-galling sawfly.Ecology, 70, 1691–1699.
Craig, T.P., Itami, J.K. & Price, P.W. (1990)Intraspecific competition and facilitation by ashoot-galling sawfly. Journal of Animal Ecology,59, 147–159.
Craig, T.P., Price, P.W., Clancy, K.M., Waring, G.L.& Sacchi, C.F. (1988) Forces preventing coevolu-tion in the three-trophic-level system: willow, agall-forming herbivore, and parasitoid. In: Chem-ical Mediation of Coevolution (ed. K.C. Spencer),pp. 57–80. Academic Press, New York.
Crawley, M.J. (2002) Statistical Computing. Wiley &Sons., Chichester, UK.
Danell, K., Hjälten, J., Ericson, L. & Elmqvist, T.(1991) Vole feeding on male and female willowshoots along a gradient of plant productivity.Oikos, 62, 145–152.
Elmqvist, T., Cates, R.G., Harper, J.K. & Gardfjell, H.A.(1991) Flowering in males and females of aUtah willow, Salix rigida and effects on growth,tannins, phenolic glycosides and sugars. Oikos, 61,65–72.
Faria, M.L. & Fernandes, G.W. (2001) Vigour of adioecious shrub and attack by a galling herbivore.Ecological Entomology, 26, 37–45.
Fernandes, G.W. (1998) Hypersensitivity as a phe-notypic basis of plant induced resistance against agalling insect (Diptera: Cecidomyiidae). Environ-mental Entomology, 27, 260–267.
Fritz, R.S. (1990) Variable competition betweeninsect herbivores on genetically variable host plants.Ecology, 71, 2008–2011.
Fritz, R.S. (1995) Direct and indirect effects of plantgenetic variation on enemy impact. EcologicalEntomology, 20, 18–26.
Fritz, R.S., Crabb, B.A. & Hochwender, C.G. (2000)Preference and performance of a gall-inducingsawfly: a test of the plant-vigor hypothesis. Oikos,89, 555–563.
Fritz, R.S., Gaud, W.S., Sacchi, C.F. & Price, P.W.(1987a) Variation in herbivore density among hostplants and its consequences for community struc-ture. Oecologia, 72, 577–588.
Fritz, R.S., Gaud, W.S., Sacchi, C.F. & Price, P.W.(1987b) Patterns of intra- and interspecific
association of gall-forming sawflies in relation toshoot size on their willow host plant. Oecologia,73, 159–169.
Fritz, R.S. & Nobel, J. (1990) Host plant variationin mortality of the leaf-folding sawfly on thearroyo willow. Ecological Entomology, 15, 25–35.
Fritz, R.S. & Price, P.W. (1990) A field test of inter-specific competition on oviposition of gall-formingsawflies on willow. Ecology, 71, 99–106.
Fritz, R.S., Sacchi, C.F. & Price, P.W. (1986) Com-petition versus host plant phenotype in speciescomposition: willow sawflies. Ecology, 67, 1608–1618.
Harris, K.M. (1994) Gall midges (Cecidomyiidae):classification and biology. In: Plant Galls (ed.M.A.J. Williams), pp. 201–211, Systematics Asso-ciation, Special, Vol. 49 Clarendon Press, Oxford.
Hartley, S.E. & Lawton, J.H. (1992) Host plant manip-ulation by gall insects: a test of the nutritionhypothesis. Journal of Animal Ecology, 61, 113–119.
Higton, R.N. & Mabberly, D.J. (1994) A willow gall fromthe galler’s point of view. In: Plant Galls — Organisms,Interactions, Populations (ed. M.A.J. Williams),pp. 301–312. Clarendon Press, Oxford.
Hjältén, J. (1992) Plant sex and hare feeding prefer-ences. Oecologia, 89, 253–256.
Julkunen-Tiitto, R. (1989) Phenolic constituents ofSalix: a chemotaxonomic survey of further Fin-nish species. Phytochemistry, 28, 2115–2125.
Kokkonen, K. (2000) Mixed significance of plantvigor: two species of galling Pontania in a hybrid-izing willow complex. Oikos, 90, 97–106.
Kolehmainen, J., Roininen, H., Julkunen-Tiitto, R.& Tahvanainen, J. (1994) Importance of phenolicglucosides in host selection of shoot gallingsawfly. Journal of Chemical Ecology, 20, 2455–2466.
Kopelke, J.-P. (1990) Wirtsspezifität als Differen-zierungskriterium bei gallenbildenden Blattwespe-narten der Gattung Pontania O. Costa (Hymenoptera:Tenthredinidae: Nematinae). Mitteilungen der Deut-schen Gesellschaft für Allgemeine und AngewandteEntomologie, 7, 527–534.
Kopelke, J.-P. (1998) Eiablage-Strategien bei gal-lenbildenden Arten der Blattwespen-GattungenPontania, Euura und Phyllocolpa (Hymenoptera:Tenthredinidae: Nematinae). Entomologia Genera-lis, 22(3/4), 251–275.
Kopelke, J.-P. (1999) Gallenerzeugende BlattwespenEuropas — Taxonomische Grundlagen, Biologieund ökologie (Tenthredinidae: Nematinae: Euura,Phyllocolpa, Pontania). Courier ForschungsinstitutSenckenberg, 212, 1–183.
Kopelke, J.-P. & Amendt, J. (2002) Species associa-tions of gall formers on willows (Salix spp.) of thefloodplains of the Rhine River (Tenthredinidae:Euura, Phyllocolpa, Pontania; Cecidomyiidae:Dasineura, Iteomyia, Rabdophaga). EntomologiaGeneralis, 26(3), 173–193.
Kristek, J. (1972) Die Blattwespen Euura laetaund Euura mucronata in den Korbweidenanlagen

Patterns of stem gallers on willows 453
© 2003 Blackwell Publishing Ltd, Diversity and Distributions, 9, 443–453
Mährens. Acta Scientiarum Naturalium AcademiaeScientiarum Bohemoslovacae Brno, 6, 1–96.
Leitch, I.J. (1994) Induction and development of thebean gall caused by Pontania proxima. In: PlantGalls: Organisms, Interactions, Populations (ed.M.A.J. Williams), pp. 283–300. Oxford SciencePublications, Oxford.
McCalla, D.R., Genthe, M.K. & Hovanitz, W.(1962) Chemical nature of an insect gall growthfactor. Plant Physiology, 37, 98–103.
McKinnon, M.L., Quiring, D.T. & Bauce, E. (1999)Influence of tree growth rate, shoot size and foliarchemistry on the abundance and performance ofa galling adelgid. Functional Ecology, 13, 859–867.
Meyer, J. (1987) Plant Galls and Gall Inducers.Gebrüder Bornträger Verlag, 291 S, Berlin, Stuttgart.
Pasteels, J.M. & Rowell-Rahier, M. (1992) Thechemical ecology of herbivory on willows. Pro-ceedings of the Royal Society of Edinburgh, 98B,63–73.
Pieres, C.S.S. & Price, P.W. (2000) Patterns ofhost plant growth and attack and establishmentof gall-inducing wasp (Hymenoptera: Cynipidae).Environmental Entomology, 29, 49–54.
Preszler, R.W. & Price, P.W. (1988) Host quality andsawfly populations: a new approach to life tableanalysis. Ecology, 69, 2012–2020; New York.
Price, P.W. (1991) The plant vigor hypothesis andherbivore attack. Oikos, 62, 244–251.
Price, P.W., Cobb, N., Craig, T.P., Fernandes, G.W.,Itami, J.K., Mopper, S. & Preszler, R.W. (1990)Insect herbivore population dynamics on treesand shrubs: New approaches relevant to latentand eruptive species and life table development.In: Insect–Plant Interactions (ed. E.E. Bernays),Vol. 2, pp. 1–38. CRC Press, Boca Raton, Florida.
Price, P.W. & Craig, T.P. (1984) Life history, pheno-logy, and survivorship on a stem-galling sawfly,Euura lasiolepis (Hymenoptera: Tenthredinidae),on the arroyo willow, Salix lasiolepis, in northernArizona. Annals of the Entomolecular Society ofAmerica, 77, 712–719.
Price, P.W., Craig, T.P. & Hunter, M. (1998) Populationecology of a gall-inducing sawfly, Euura lasiolepis,and relatives. In: Insect Populations in Theoryand in Practice (eds J.P. Dempster & I.F.G. McLean),pp. 323–340, Kluwer. Academic Publishers.
Price, P.W. & Ohgushi, T. (1995) Preference andperformance linkage in a Phyllocolpa sawfly on thewillow, Salix miyabeana, on Hokkaido. Researcheson Population Ecology, 37(1), 23–28.
Price, P.W., Roininen, H. & Ohgushi, T. (1999)Comparative plant–herbivore interactions involv-ing willows and three gall-inducing sawfly speciesin the genus Pontania (Hymenoptera: Tenthredini-dae). Ecoscience, 6, 41–50.
Price, P.W., Roininen, H. & Tahvanainen, J. (1987a)Plant age and attack by the bud galler, Euuramucronata. Oecologia, 73, 334–337.
Price, P.W., Roininen, H. & Tahvanainen, J. (1987b)Why does the bud-galling sawfly, Euura mucro-nata, attack long shoots? Oecologia, 74, 1–6.
Price, P.W., Roininen, H. & Tahvanainen, J. (1997)Willow tree shoot module length and the attackand survival pattern of a shoot-galling sawfly,Euura atra (Hymenoptera: Tenthredinidae). Ento-mologia Fennica, 8, 113–119.
Price, P.W., Waring, G.L., Julkunen-Titto, R.,Tahvanainen, J., Mooney, H. & Craig, T.P. (1989)The carbon-nutrient balance hypothesis in withinspecies phytochemical variation of Salix lasiolepis.Journal of Chemical Ecology, 15, 1117–1131.
Roininen, H. & Danell, K. (1997) Mortality factorsand resource use of the bud-galling sawfly, Euuramucronata (Hartig), on willows (Salix spp.) inarctic Eurasia. Polar Biology, 18, 325–330.
Roininen, H., Price, P.W., Julkunen-Tiitto, R.,Tahvanainen, J. & Ikonen, A. (1999) Ovipositionstimulant for a gall-inducing sawfly, Euura lasi-olepis, on willow is a phenolic glucoside. Journalof Chemical Ecology, 25, 943–953.
Rowell-Rahier, M. & Pasteels, J.M. (1990) Phenolg-lycosides and interactions at three trophic levels:Salicaceae — herbivores — predatores. In: Insect–Plant Interactions (ed. E.A. Bernays), 2, pp. 75–94.CRC Press, Boca Raton.
Schönrogge, K., Harper, L.J. & Lichtenstein, C.P.(2000) The protein content of tissues in cynipidgalls (Hymenoptera: Cynipidae): Similarities betweencynipid galls and seeds. Plant Cell and Environ-ment, 23, 215–222.
Shao, Y. (1991) Phytochemischer Atlas der SchweizerWeiden. Dissertation ETH Zürich, no. 9532. 199 S.
Shorthouse, J.D. & Rohfritsch, O. (1992) Biology ofInsect-Induced Galls. Oxford University Press NewYork.
Sitch, T.A., Grewcock, D.A. & Gilbert, F.S. (1988)Factors affecting components of the fitness in agall-making wasp (Cynips divisa Hartig). Oecolo-gia, 76, 371–375.
Stein, S.T. & Price, P.W. (1995) Relative effects ofplant resistance and natural enemies by plantdevelopmental age on sawfly (Hymenoptera:Tenthredinidae) preference and performance.Environmental Entomology, 24, 909–916.
Stone, G.N., Schönrogge, K., Atkinson, R.,Bellido, D. & Pujade-Villar, J. (2002) The populationbiology of oak gall wasps (Hymenoptera: Cynipidae).Annual Review of Entomology, 47, 633–668.
Urban, J. (1995) Mortalität der Blattwespen-ArtenEuura laeta und E. mucronata infolge physiologischerUrsachen und durch Abwehr-Überwallungswachstumder Wirtspflanzen-Gewebe (Symphyta: Tenthredi-nidae). Entomologia Generalis, 20(1/2), 103–121.
Woods, J.O., Carr, T.G., Price, P.W., Stevens, L.E. &Cobb, N.S. (1996) Growth of coyote willow andthe attack and survival of a mid-rib galling sawfly,Euura sp. Oecologia, 108, 714–722.




















