
Patterns of adult abundance in Chthamalus stellatus (Poli) and C. montagui Southward (Crustacea:...
15
Patterns of adult abundance in Chthamalus stellatus (Poli) and C. montagui Southward (Crustacea: Cirripedia) emerge during late recruitment A.M. Power a, * , J. Delany b , D. McGrath c , A.A. Myers d , R.M. O’Riordan d a Department of Zoology, Room 226 Martin Ryan Institute, National University of Ireland, Galway, Ireland b Dove Marine Laboratory, University of Newcastle, Cullercoats, North Shields, Tyne and Wear NE30 4PZ, UK c Department of Life Sciences, Galway-Mayo Institute of Technology, Galway, Ireland d Department of Zoology, Ecology and Plant Science, University College Cork, Lee Maltings, Prospect Row, Cork City, Ireland Received 15 September 2004; received in revised form 18 October 2005; accepted 15 November 2005 Abstract The aim of this study was to investigate when adult distribution patterns are established in the barnacles Chthamalus stellatus and C. montagui . Adult dzonesT were identified by analysing field counts of both species at mid and upper shore heights. Monthly collections of cyprids, b 1 month old metamorphs and recruits (all metamorphosed individuals older than approximately 1 month) were made for C. stellatus and C. montagui in natural barnacle beds at six shores in SW Ireland. This was carried out over one year in 1996/1997, using a hierarchical sampling design. Abundance of total recruits (0–3 months old) was compared between adult zones after the main settlement season had ended. In addition, scales of variability in 0–3 month recruitment into adult zones were compared between the species at two scales: shores (1000s of metres) and sites within shores (10s of metres). Older recruits of each species, up to 11 months of age, were also compared between adult zones. The majority of settlement (measured as attached cyprids) occurred between August and October 1996. In October, there was no effect of adult zone on the abundance of total (0–3 month) recruitment up to that point in either species. Despite this homogeneity in recruitment between adult zones, significant spatial variation was found in 0–3 month recruits of both species at both of the scales examined. In C. stellatus the amount of variation associated with the larger scale (shore) was more than twice that of sites or of the residual variation (replicates within sites). 0–3 month recruitment in C. montagui was also most variable at the scale of shores but the residual variability (between replicates within site) was of similar magnitude to that of shores. Variability in 0–3 month C. montagui recruitment was relatively low at the scale of sites. There was a small but consistent input of recruits to adult zones over 9 months of the year, complicating the assessment of when adult patterns were set-up in these species. By June 1997, characteristic patterns of adult dominance had been established at all shores. Settlement had completely ceased by this time and individual barnacles were potentially 11 months old. Neither settlement nor early recruitment are significant in determining adult zonation patterns in these species. Instead, differential mortality patterns in individuals up to the age of 11 months are implicated in determining patterns of distribution of both species. D 2005 Elsevier B.V. All rights reserved. Keywords: Barnacles; Chthamalus ; Recruitment; Regulation; Spatial variation 0022-0981/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.jembe.2005.11.012 * Corresponding author. Tel.: +353 91 493015; fax: +353 91 494526. E-mail address: [email protected] (A.M. Power). Journal of Experimental Marine Biology and Ecology 332 (2006) 151 – 165 www.elsevier.com/locate/jembe
Transcript of Patterns of adult abundance in Chthamalus stellatus (Poli) and C. montagui Southward (Crustacea:...

www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolog
Patterns of adult abundance in Chthamalus stellatus (Poli) and
C. montagui Southward (Crustacea: Cirripedia) emerge during
late recruitment
A.M. Power a,*, J. Delany b, D. McGrath c, A.A. Myers d, R.M. O’Riordan d
a Department of Zoology, Room 226 Martin Ryan Institute, National University of Ireland, Galway, Irelandb Dove Marine Laboratory, University of Newcastle, Cullercoats, North Shields, Tyne and Wear NE30 4PZ, UK
c Department of Life Sciences, Galway-Mayo Institute of Technology, Galway, Irelandd Department of Zoology, Ecology and Plant Science, University College Cork, Lee Maltings, Prospect Row, Cork City, Ireland
Received 15 September 2004; received in revised form 18 October 2005; accepted 15 November 2005
Abstract
The aim of this study was to investigate when adult distribution patterns are established in the barnacles Chthamalus stellatus
and C. montagui. Adult dzonesT were identified by analysing field counts of both species at mid and upper shore heights. Monthly
collections of cyprids, b1 month old metamorphs and recruits (all metamorphosed individuals older than approximately 1 month)
were made for C. stellatus and C. montagui in natural barnacle beds at six shores in SW Ireland. This was carried out over one year
in 1996/1997, using a hierarchical sampling design. Abundance of total recruits (0–3 months old) was compared between adult
zones after the main settlement season had ended. In addition, scales of variability in 0–3 month recruitment into adult zones were
compared between the species at two scales: shores (1000s of metres) and sites within shores (10s of metres). Older recruits of each
species, up to 11 months of age, were also compared between adult zones.
The majority of settlement (measured as attached cyprids) occurred between August and October 1996. In October, there was
no effect of adult zone on the abundance of total (0–3 month) recruitment up to that point in either species. Despite this
homogeneity in recruitment between adult zones, significant spatial variation was found in 0–3 month recruits of both species at
both of the scales examined. In C. stellatus the amount of variation associated with the larger scale (shore) was more than twice that
of sites or of the residual variation (replicates within sites). 0–3 month recruitment in C. montagui was also most variable at the
scale of shores but the residual variability (between replicates within site) was of similar magnitude to that of shores. Variability in
0–3 month C. montagui recruitment was relatively low at the scale of sites.
There was a small but consistent input of recruits to adult zones over 9 months of the year, complicating the assessment of when
adult patterns were set-up in these species. By June 1997, characteristic patterns of adult dominance had been established at all
shores. Settlement had completely ceased by this time and individual barnacles were potentially 11 months old. Neither settlement
nor early recruitment are significant in determining adult zonation patterns in these species. Instead, differential mortality patterns
in individuals up to the age of 11 months are implicated in determining patterns of distribution of both species.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Barnacles; Chthamalus; Recruitment; Regulation; Spatial variation
0022-0981/$ - s
doi:10.1016/j.jem
* Correspondi
E-mail addr
y and Ecology 332 (2006) 151–165
ee front matter D 2005 Elsevier B.V. All rights reserved.
be.2005.11.012
ng author. Tel.: +353 91 493015; fax: +353 91 494526.
ess: [email protected] (A.M. Power).

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165152
1. Introduction
Identifying patterns of distribution of species and the
factors underlying these patterns is a fundamental goal
in ecology. Barnacles have open populations, i.e., local
recruitment is potentially uncoupled from local repro-
duction by a dispersal phase (Roughgarden et al., 1985;
Hughes, 1990). Larval recruitment in open populations
is analogous to birth in closed populations (Caley et al.,
1996), therefore attempts to understand how adult dis-
tribution patterns arise in barnacles have concentrated
on determining whether adult patterns were set-up at
larval settlement (i.e., dbirthT) or instead, at a later stagein benthic life due to post-settlement mortality (or
ddeathT) (see Connell, 1985; Caley et al., 1996 for
reviews).
Investigations into whether small or variable settle-
ment can regulate species’ distribution and abundance
are carried out by seeking evidence of density-depen-
dence between the abundance of settlement and subse-
quent adult densities (see Caley et al., 1996). Settlement
regulation has been predicted from a theoretical model
for small settlement densities (Roughgarden et al.,
1985) and subsequently shown in field studies (e.g.,
Connell, 1985; Gaines and Roughgarden, 1985; Ken-
dall and Bedford, 1987; Minchinton and Scheibling,
1991; Carroll, 1996; Jeffery, 2003). However, contrary
to predictions, settlement regulation has also been
shown at large settlement densities (Raimondi, 1990;
Menge, 2000). In addition, mortality factors such as
predation were important in determining density of
adults even when settlement densities were low (Car-
roll, 1996; Menge, 2000). Where intertidal vertical
zonation of adults of species exist, another approach
has been adopted. In this case, processes that contribute
to the establishment of vertical zonation patterns have
been examined by comparing the vertical limits of
settlement and recruitment, with the upper and lower
vertical limits of adults (e.g., Hatton, 1938; Connell,
1961a,b; Denley and Underwood, 1979; Strathmann
and Branscomb, 1979; Wethey, 1983; Raimondi,
1988). In some cases, both approaches have been si-
multaneously examined, by seeking evidence of densi-
ty-dependent relationships between different life stages
at several shore heights (e.g., Minchinton and Schei-
bling, 1991; Carroll, 1996; Menge, 2000).
All open populations are driven by recruitment in the
sense that recruits are required to have arrived and
survived at some time in the past to set the initial condi-
tion (i.e., density of recruits), on which post-settlement
factors may act to produce adult populations (Caffey,
1985; Carroll, 1996). Nevertheless considerable evi-
dence has accumulated to indicate that mortality due to
predation, competition, disturbance, or a combination of
these three is important in structuring barnacle popula-
tions (Hatton, 1938; Connell, 1961a,b; Moyse and
Knight-Jones, 1967; Lubchenco and Menge, 1978;
Wethey, 1983, 1984; Menge, 1991, 2000; Lively et al.,
1993; Carroll, 1996; Pineda et al., 2002). However, not
all of these studies examined the relative contribution
of settlement and early post-settlement mortality (e.g.,
Menge, 1991, 2000; Lively et al., 1993). Connell
(1985) defined settlement as the point when an indi-
vidual first makes permanent contact with the substra-
tum. This has rarely been observed directly in the
field; more usually, sampling occurs at an unknown
time after settlement. Strictly, all studies where indi-
viduals have survived a period of time between set-
tlement and census are more correctly defined as
recruitment studies and not settlement studies (for
further discussion see Connell, 1985; Underwood
and Denley, 1984).
In a review, Underwood and Denley (1984) pro-
posed that settlement patterns could be used to frame
an alternative hypothesis to explain regulation of popu-
lations. The importance of events at settlement such as
larval supply or arrival at adult zones (Gaines and
Roughgarden, 1985; Gaines et al., 1985; Gaines and
Bertness, 1993; Minchinton and Scheibling, 1991),
larval distribution in the water column (Grosberg,
1982), larval settlement rate (Denley and Underwood,
1979; Bushek, 1988; Raimondi, 1990; Sutherland,
1990), larval quality (Jarrett and Pechenik, 1997;
Miron et al., 1999) or settlement cues (Raimondi,
1988) have been shown (see review by Connell, 1985
and discussion by Underwood and Denley, 1984; Un-
derwood and Fairweather, 1989).
Dense populations of the barnacles Chthamalus stel-
latus and C. montagui occur in SW Ireland, which
offers an opportunity to examine simultaneously how
distribution and abundance of both species are estab-
lished. Populations of these species approach the north-
ern edges of their ranges in SW Ireland and although
they can overlap vertically over shore heights and
horizontally over gradients of exposure, zones of nu-
merical dominance of each species do occur. C. mon-
tagui usually dominates the upper barnacle zone and
can be significantly more abundant there than it is on
the midshore, where C. stellatus may be dominant
(Power, 2000; Delany et al., 2003). C. stellatus-domi-
nated zones tend to be found on more wave-exposed
shores (2–3 Ballantine scale (Ballantine, 1961)) while
C. montagui-dominated zones tend to be at less wave-
exposed locations (3–6 Ballantine scale) (Power, 2000).

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 153
It has been postulated that population distribution
and abundance of these two species may be established
at settlement: larger cyprid size and a greater dispersal
of larval phases in C. stellatus relative to C. montagui
may favour its increased settlement on offshore islands,
while less distantly-occurring larvae and larval reten-
tion in more sheltered conditions may favour C. mon-
tagui (Crisp et al., 1981; Burrows, 1988; Burrows et al.,
1999; Ross et al., 2003). However, Power et al. (1999a)
and Delany et al. (2003) have shown in SW Ireland that
neither attached cyprid abundance (dsettlementT), normetamorphs (b1 month old) favoured conspecific
adult zones compared to zones dominated by adults
of the other species (although this contrasts with a
recent report from Plymouth, S England (Jenkins,
2005)). In addition to no distributional differences be-
tween the species at settlement/metamorph stages, there
was significantly higher post-settlement mortality in the
dwrongT adult zone in both species between 6 and 12
months after settling on the shore (Burrows, 1988;
Delany et al., 2003). This poses the question: when
do zonation patterns emerge in these barnacle species?
In order to examine at what stage adult zonation
patterns were established, the present study examined
recruitment variation between adult zones, separately in
C. stellatus and C. montagui. Total recruitment was
compared after the main settlement season had ended
in different adult zones for each species (i.e., the anal-
ysis compared the sum of cyprids plus metamorphosed
individuals plus recruits to a maximum age of 3 months
old). All sampling was carried out in natural barnacle
beds and spatial variation between adult zones was
examined at two scales: shores (1000s of metres) and
sites within shores (10s of metres). A similar compar-
ison, this time omitting sites, was carried out for older
recruits (to a maximum possible age of 11 months old).
Previous studies have noted that total abundance of
C. stellatus cyprids was significantly higher than that of
C. montagui cyprids, despite the fact that metamorphs
of the two species did not significantly differ (Power et
al., 1999a; Power, 2000; Delany et al., 2003). This will
not affect the interpretation of recruitment into adult
zones in the present study because total recruitment was
examined separately in each species. However if, as has
been suggested, a different proportion between settlers
and metamorphs exists in the two species and this is
due to a different rate of metamorphosis (Power et al.,
1999a), it has important consequences for comparing
settlement in the species since the attached cyprid as a
settlement unit would not be directly equivalent be-
tween the two. On the other hand, if the species differ
in their proportion between cyprids and metamorphs
because of different early post-settlement survival, this
is also a potentially important source of variation to
measure. We examined the difference between the spe-
cies in cyprids (expressed as a proportion of cyprids
plus metamorphs) to see whether these proportions
differed consistently, or whether the pattern was due
to disproportionate effects at particular adult barnacle
zones, shores or dates.
2. Methods
2.1. Study sites and sampling
Zones of adult dominance were established by sam-
pling counts of each species as adults in 15 independent
replicate (10�10) cm quadrats at six shores in Co.
Cork, SW Ireland. The shores were: Bullens Bay East
(BBE), Bullens Bay West (BBW), Old Head of Kinsale
(OHK), Garrettstown (GARR), Nohoval (NOH) and
Castlepark (CPK) (for further details see Power, 2000;
Delany et al., 2003). Comparisons of species abundance
were made between mid (1.5–2.4 m above CD) and
upper (3.75–4.65 m above CD) shore levels at all
shores except for one shore, CPK, where the upper
level was 2.85–3.75 m above CD due to the barnacle
zone occurring over a reduced vertical extent at that
shore.
Settlement and recruitment of C. stellatus and C.
montagui were assessed using a hierarchical experi-
mental design within zones where either C. stellatus
or C. montagui were more abundant as adults. We
wished to test the hypothesis that recruitment in a single
species was different between adult zones therefore we
chose randomly-selected representative replicate shores
within each zone. Zones were thus kept independent by
being sampled on different shores. Sampling was car-
ried out from July 1996 to June 1997 within the C.
stellatus (=mid shore) zone at three of the six shores
(BBE, NOH and OHK) and similarly within the C.
montagui (=upper shore) zone at three remaining
shores (GARR, BBW and CPK). Shores were located
within 30 km of each other. Three sites were sampled
per shore, these sites were defined as areas of (2�2) m
within the designated zone and were haphazardly se-
lected on each visit to be at least 5 m but no greater than
30 m apart. Sampling was carried out monthly on each
shore within 5 days either side of the lowest spring tide.
On each sampling occasion, all cyprids and metamor-
phosed individuals within six replicate (10�10) cm
quadrats per site in natural barnacle beds were removed
and placed in 70% ethanol. Replicate quadrats were
stratified to exclude pools and crevices.

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165154
In addition to sampling recruitment at independent
adult zones (at different shores -see above), we exam-
ined recruitment of each species between adult zones
within shores. Two shores were monitored each day at
the beginning of the Chthamalus settlement season in
1998. Once cyprid settlement was observed, two
patches were sampled in each adult zone on each
shore between 28 July and 13 August. Patches were
approximately 5 m wide and were sampled in the mid
and upper shore zones, corresponding to zones of
adult abundance in C. stellatus and C. montagui. In
each patch 30 replicate collections were made of
cyprids and metamorphs in (10�10) cm quadrats.
During sampling occasions with heavy settlement rep-
licate size and number was reduced to 18 (5�5) cm
quadrats.
2.2. Identification of cyprids, metamorphs and recruits
Identification of cyprid to species was possible fol-
lowing measurement of each individual’s carapace
length (see O’Riordan et al., 1999; Power et al.,
1999b). Metamorphs were identified according to Bur-
rows (1988) and determined to be b1 month-old based
on criteria of size, colour and degree of calcification
(Power et al., 1999a).
The beginning of the settlement season in 1998 was
observed to within a day of its initiation at the study
shores. Therefore for the experiment comparing settle-
ment/recruitment between adult zones within shores,
the age of metamorphs could be assumed to be b17
days old. Note that this was slightly younger than
potential maximum age of metamorphs in the spatial
variability study, in which metamorphs were aged to b1
month old.
In analyses of individualsN1 month old and b11
months old, maximum recruit age was determined as
the time elapsed between the initiation of settlement in
1996 and the sample collection date for each of the four
dates analysed.
2.3. Statistical analyses of age classes
2.3.1. Adult zones
We tested the hypothesis that adult species abun-
dance differed between mid shore zones, where C.
stellatus was the more abundant barnacle species and
upper shore zones, where C. montagui was more abun-
dant. A three-factor analysis of variance (ANOVA) was
carried out: both zone and species were fixed, orthog-
onal factors with two levels each; three random shores
were nested within each zone.
2.3.2. Variation in 0–3 month recruits between adult
zones and at spatial scales
Total recruitment was compared in each species
separately between two adult zones after the main set-
tlement season had ended. This was carried out at two
spatial scales. Because all age groups were removed and
identified this single point estimate of recruitment
includes cyprids, metamorphs and recruits of up to a
maximum age of 3 months old. Sites were haphazardly
selected on each date, so to include this spatial scale in
ANOVA, analysis was restricted to a single point after
the main settlement season had occurred in both species
(October). Zone had two levels and was considered as a
fixed, orthogonal factor. Shore had three levels, was
nested in zone and was random. Site was nested in the
shore�zone interaction, was random with three levels,
there were six replicate quadrats sampled within each
site. Spatial scales of recruitment were examined sepa-
rately in each species so that the magnitude of random
effects of scale could be examined for each species
separately in the model. From the mean squares esti-
mates generated by ANOVA, we calculated the magni-
tude of effects according to the components of variation
associated with the random factors in the model (shores,
sites and residual) (Winer, 1971).
2.3.3. Abundance of cyprids+b17 day old metamorphs
between adult zones (within shores)
To examine whether abundance of settlement or
early recruitment (cyprids+b17 day old metamorphs)
varied between adult zones within shores we compared
mid (=C. stellatus zone, 1.5–2.4 m above CD) and
upper (=C. montagui zone, 3.75–4.65 m above CD)
for each species. Zone and species were considered to
be fixed orthogonal factors. Shores were random and
orthogonal and patches were random and nested within
shores. All four factors had two levels and nine inde-
pendent replicates were used for each species.
2.3.4. Recruit abundance (N1 month and b11 months
old) between adult zones
Patterns of species dominance in recruits between
adult zones were examined across the sampling dates
after the main settlement had occurred. Bad weather
made sampling impossible at all six shores in some
months. This missing data applied equally to both
species so, although it did not compromise the hypoth-
esis under investigation, this did have the effect of
making the design unbalanced. Therefore the analysis
was simplified to a comparison between zones (two
levels, fixed, orthogonal), shores (two levels, random,
nested in zone), species (two levels, fixed, orthogonal)

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 155
and dates (four levels, i.e., November, December, Jan-
uary and June; fixed and orthogonal). Nine independent
replicates were used for each species.
2.3.5. Cyprids as a proportion (of cyprids+b1 month
old metamorphs) between adult zones, shores and
sampling dates
We tested the hypothesis of a different proportion
between the two species of abundance in cyprids rela-
tive to b1 month old metamorphs. This comparison
was carried out across different adult zones and across
shores within zones over the main settlement season.
The dependent variable was a proportion between
cyprid abundance and metamorph abundance from
one month later. Since sites were haphazardly chosen
on each sampling occasion, we could not match sites
between successive visits so this factor is omitted from
the analysis. There were 4 factors in the ANOVA, zone
and species each had two levels and were orthogonal
and fixed. Date was also orthogonal and fixed and
included four levels representing four sampling occa-
sions when cyprids were sufficiently abundant, i.e.,
early August, late August, September and October.
Shore was nested in zone and was a random factor
with three levels. There were 18 replicate proportions.
Table 1
Analysis of variance of abundance in two barnacle species
Source of variation df
Zo 1
Sh (Zo) 4
Sp 1
Zo�Sp 1
Sp�Sh (Zo) 4
Residual 168
Transformation Ln (X +1)
Cochran’s C 0.190
P Significant ( P b0.05)
SNK multiple comparisons:
Zo within levels of Sp
C. montagui
C. stellatus
Sp within levels of Zo
Upper shore
Mid shore
Sp within (Sh�Zo)
Mid shore BBE
Mid shore OHK
Mid shore NOH
Upper shore GARR
Upper shore BBW
Upper shore CPK
Adult zones of dominance were compared between Chthamalus stellatus on
upper shore (3.75–4.65 m above CD). Sampling was carried out on six shores
of model terms: Zo=zone, Sh=shore, Sp=species. Abbreviations of shores
OHK=Old Head of Kinsale, NOH=Nohoval, BBE=Bullens Bay East.
For all analyses, homogeneity of variances were
checked using Cochran’s C and where variances were
found to be heterogenous, transformations were made.
In cases where transformations failed to homogenise
variances, we proceeded with the analysis on untrans-
formed data. Analysis of variance is robust to departures
from the assumption of homogeneity of variances if the
experiment is large and balanced (Underwood, 1997). In
addition, since violations of this assumption increase the
risk of Type I errors, non-significant results are accept-
able (Underwood, 1997). Multiple comparisons within
levels of significant factors performed according to
Student–Newman–Keuls (SNK) tests. Analysis of vari-
ance was conducted GMAV5 software for windows
(Institute of Marine Ecology, Sydney, Australia).
3. Results
3.1. Adult zones
Cochran’s C indicated significantly heterogeneous
variances between samples (P b0.05) which transfor-
mation failed to correct, but based on a large balanced
experimental design (in this case there were 15 repli-
cates) we proceeded with the analysis. C. stellatus were
MS F P
8.75 0.47 0.531
18.62 10.71 0.000
9.96 1.15 0.345
221.88 25.54 0.007
8.69 5.00 0.000
1.74
UpperNmid shore
Upperbmid shore
C. montagui NC. stellatus
C. montagui bC. stellatus
C. montagui bC. stellatus
C. montagui bC. stellatus
C. montagui bC. stellatus
C. montagui NC. stellatus
C. montagui NC. stellatus
C. montagui NC. stellatus
the mid shore (1.5–2.4 m above CD) and C. montagui zones on the
in SW Ireland, n =15. SNK=Student–Newman–Keuls. Abbreviations
: GARR=Garrettstown, BBW=Bullens Bay West, CPK=Castlepark,
C. stellatus zone C. montagui zone
0
50
100
150
200
250
S O N D J F M A M J
garr
bbw
cpk
0
50
100
150
200
250
S O N D J F M A M J
bbe
ohk
noh
Cyprids N= 4628
Metamorphs N= 14420
0
50
100
150
200
250
earlyAug
lateAug
earlyAug
lateAug
earlyAug
lateAug
earlyAug
lateAug
S O N D J F M A M J
bbe
ohk
noh
Cyprids N=4568
0
50
100
150
200
250
S O N D J F M A M J
garr
bbw
cpk
Metamorphs N= 5630M
ean
Abu
ndan
ce/d
m2
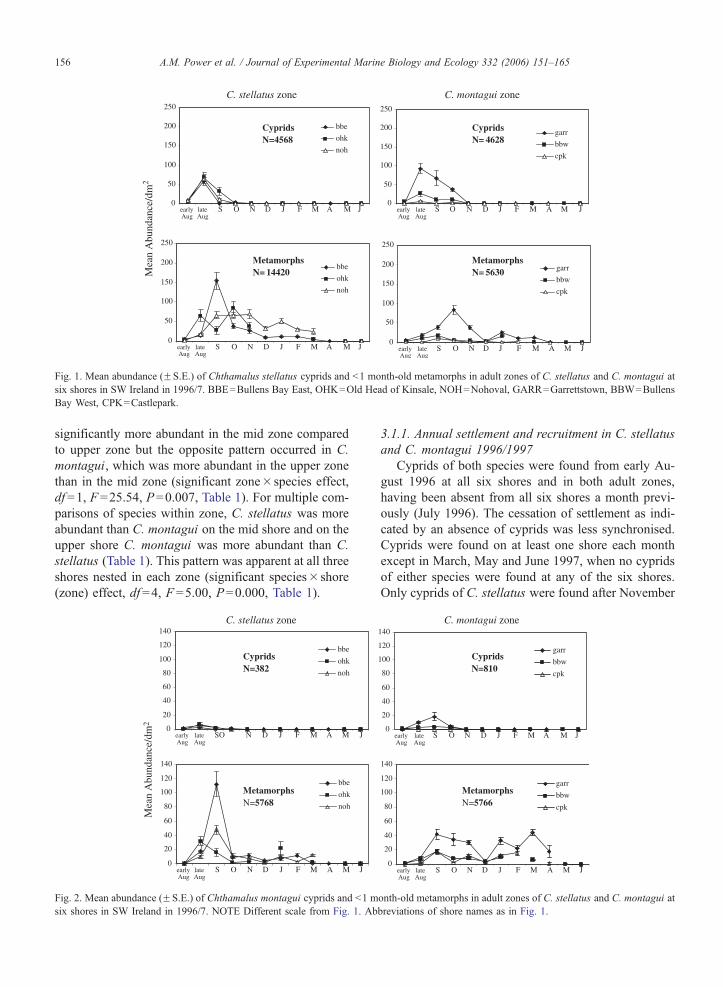
Fig. 1. Mean abundance (FS.E.) of Chthamalus stellatus cyprids and b1 month-old metamorphs in adult zones of C. stellatus and C. montagui at
six shores in SW Ireland in 1996/7. BBE=Bullens Bay East, OHK=Old Head of Kinsale, NOH=Nohoval, GARR=Garrettstown, BBW=Bullens
Bay West, CPK=Castlepark.
A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165156
significantly more abundant in the mid zone compared
to upper zone but the opposite pattern occurred in C.
montagui, which was more abundant in the upper zone
than in the mid zone (significant zone� species effect,
df =1, F =25.54, P=0.007, Table 1). For multiple com-
parisons of species within zone, C. stellatus was more
abundant than C. montagui on the mid shore and on the
upper shore C. montagui was more abundant than C.
stellatus (Table 1). This pattern was apparent at all three
shores nested in each zone (significant species� shore
(zone) effect, df =4, F =5.00, P=0.000, Table 1).
C. stellatus zone
0
20
40
60
80
100
120
140
earlyAug
lateAug
SO N D J F M A M J
bbe
ohk
noh
0
20
40
60
80
100
120
140
earlyAug
lateAug
S O N D J F M A M J
bbe
ohk
noh
Mea
n A
bund
ance
/dm
2
Cyprids N=382
Metamorphs N=5768
Fig. 2. Mean abundance (FS.E.) of Chthamalus montagui cyprids and b1 m
six shores in SW Ireland in 1996/7. NOTE Different scale from Fig. 1. Ab
3.1.1. Annual settlement and recruitment in C. stellatus
and C. montagui 1996/1997
Cyprids of both species were found from early Au-
gust 1996 at all six shores and in both adult zones,
having been absent from all six shores a month previ-
ously (July 1996). The cessation of settlement as indi-
cated by an absence of cyprids was less synchronised.
Cyprids were found on at least one shore each month
except in March, May and June 1997, when no cyprids
of either species were found at any of the six shores.
Only cyprids of C. stellatus were found after November
C. montagui zone
0
20
40
60
80
100
120
140
earlyAug
lateAug
S O N D J F M A M J
garr
bbw
cpk
0
20
40
60
80
100
120
140
earlyAug
lateAug
S O N D J F M A M J
garr
bbw
cpk
Cyprids N=810
Metamorphs N=5766
onth-old metamorphs in adult zones of C. stellatus and C. montagui at
breviations of shore names as in Fig. 1.
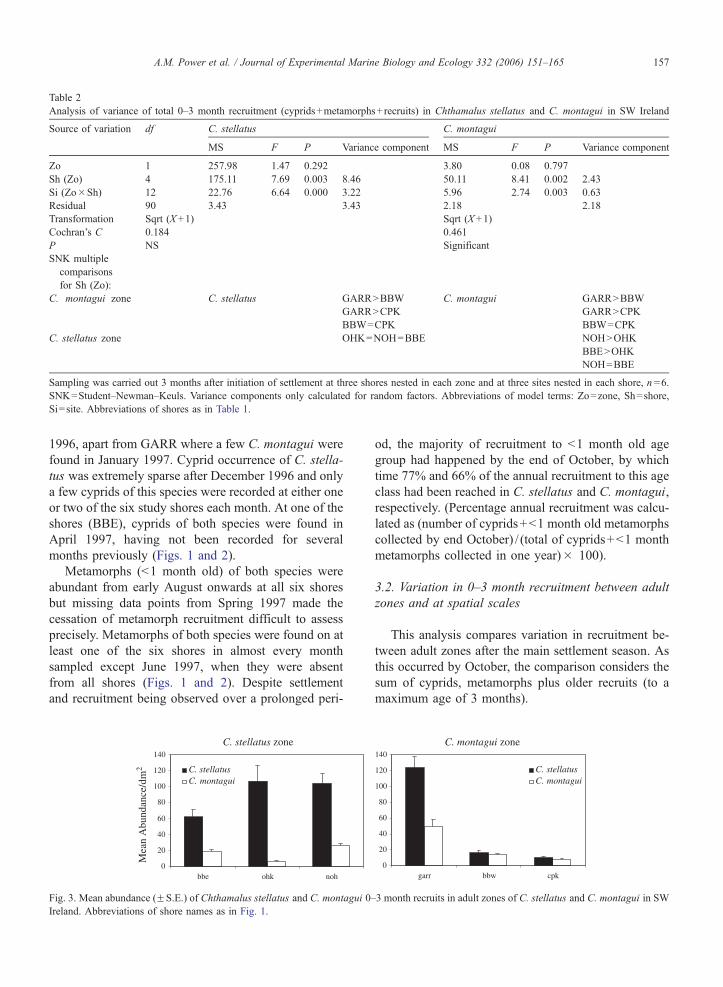
Table 2
Analysis of variance of total 0–3 month recruitment (cyprids+metamorphs+recruits) in Chthamalus stellatus and C. montagui in SW Ireland
Source of variation df C. stellatus C. montagui
MS F P Variance component MS F P Variance component
Zo 1 257.98 1.47 0.292 3.80 0.08 0.797
Sh (Zo) 4 175.11 7.69 0.003 8.46 50.11 8.41 0.002 2.43
Si (Zo�Sh) 12 22.76 6.64 0.000 3.22 5.96 2.74 0.003 0.63
Residual 90 3.43 3.43 2.18 2.18
Transformation Sqrt (X +1) Sqrt (X +1)
Cochran’s C 0.184 0.461
P NS Significant
SNK multiple
comparisons
for Sh (Zo):
C. montagui zone C. stellatus GARRNBBW C. montagui GARRNBBW
GARRNCPK GARRNCPK
BBW=CPK BBW=CPK
C. stellatus zone OHK=NOH=BBE NOHNOHK
BBENOHK
NOH=BBE
Sampling was carried out 3 months after initiation of settlement at three shores nested in each zone and at three sites nested in each shore, n =6.
SNK=Student–Newman–Keuls. Variance components only calculated for random factors. Abbreviations of model terms: Zo=zone, Sh=shore,
Si=site. Abbreviations of shores as in Table 1.
A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 157
1996, apart from GARR where a few C. montagui were
found in January 1997. Cyprid occurrence of C. stella-
tus was extremely sparse after December 1996 and only
a few cyprids of this species were recorded at either one
or two of the six study shores each month. At one of the
shores (BBE), cyprids of both species were found in
April 1997, having not been recorded for several
months previously (Figs. 1 and 2).
Metamorphs (b1 month old) of both species were
abundant from early August onwards at all six shores
but missing data points from Spring 1997 made the
cessation of metamorph recruitment difficult to assess
precisely. Metamorphs of both species were found on at
least one of the six shores in almost every month
sampled except June 1997, when they were absent
from all shores (Figs. 1 and 2). Despite settlement
and recruitment being observed over a prolonged peri-
C. stellatus zone
0
20
40
60
80
100
120
140
bbe ohk noh
C. stellatusC. montagui
Mea
n A
bund
ance
/dm
2
Fig. 3. Mean abundance (FS.E.) of Chthamalus stellatus and C. montagui 0–
Ireland. Abbreviations of shore names as in Fig. 1.
od, the majority of recruitment to b1 month old age
group had happened by the end of October, by which
time 77% and 66% of the annual recruitment to this age
class had been reached in C. stellatus and C. montagui,
respectively. (Percentage annual recruitment was calcu-
lated as (number of cyprids+b1 month old metamorphs
collected by end October) / (total of cyprids+b1 month
metamorphs collected in one year)� 100).
3.2. Variation in 0–3 month recruitment between adult
zones and at spatial scales
This analysis compares variation in recruitment be-
tween adult zones after the main settlement season. As
this occurred by October, the comparison considers the
sum of cyprids, metamorphs plus older recruits (to a
maximum age of 3 months).
C. montagui zone
0
20
40
60
80
100
120
140
garr bbw cpk
C. stellatusC. montagui
3 month recruits in adult zones of C. stellatus and C. montagui in SW
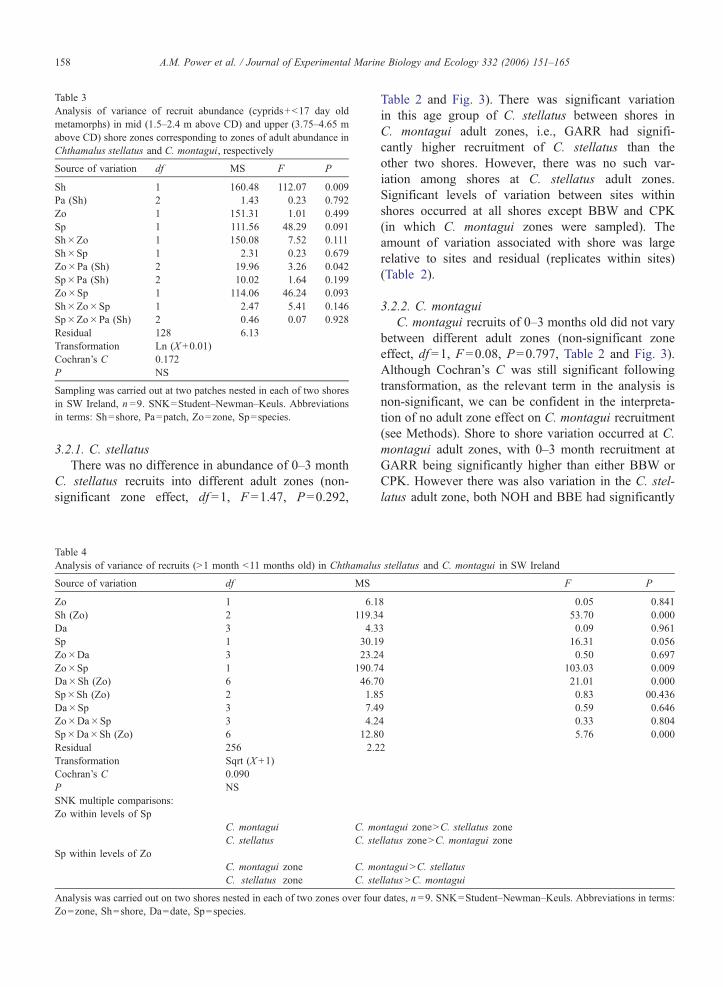
Table 3
Analysis of variance of recruit abundance (cyprids+b17 day old
metamorphs) in mid (1.5–2.4 m above CD) and upper (3.75–4.65 m
above CD) shore zones corresponding to zones of adult abundance in
Chthamalus stellatus and C. montagui, respectively
Source of variation df MS F P
Sh 1 160.48 112.07 0.009
Pa (Sh) 2 1.43 0.23 0.792
Zo 1 151.31 1.01 0.499
Sp 1 111.56 48.29 0.091
Sh�Zo 1 150.08 7.52 0.111
Sh�Sp 1 2.31 0.23 0.679
Zo�Pa (Sh) 2 19.96 3.26 0.042
Sp�Pa (Sh) 2 10.02 1.64 0.199
Zo�Sp 1 114.06 46.24 0.093
Sh�Zo�Sp 1 2.47 5.41 0.146
Sp�Zo�Pa (Sh) 2 0.46 0.07 0.928
Residual 128 6.13
Transformation Ln (X +0.01)
Cochran’s C 0.172
P NS
Sampling was carried out at two patches nested in each of two shores
in SW Ireland, n =9. SNK=Student–Newman–Keuls. Abbreviations
in terms: Sh=shore, Pa=patch, Zo=zone, Sp=species.
A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165158
3.2.1. C. stellatus
There was no difference in abundance of 0–3 month
C. stellatus recruits into different adult zones (non-
significant zone effect, df =1, F =1.47, P=0.292,
Table 4
Analysis of variance of recruits (N1 month b11 months old) in Chthamalu
Source of variation df MS
Zo 1 6.1
Sh (Zo) 2 119.3
Da 3 4.3
Sp 1 30.1
Zo�Da 3 23.2
Zo�Sp 1 190.7
Da�Sh (Zo) 6 46.7
Sp�Sh (Zo) 2 1.8
Da�Sp 3 7.4
Zo�Da�Sp 3 4.2
Sp�Da�Sh (Zo) 6 12.8
Residual 256 2.2
Transformation Sqrt (X +1)
Cochran’s C 0.090
P NS
SNK multiple comparisons:
Zo within levels of Sp
C. montagui C. mo
C. stellatus C. ste
Sp within levels of Zo
C. montagui zone C. mo
C. stellatus zone C. ste
Analysis was carried out on two shores nested in each of two zones over fou
Zo=zone, Sh=shore, Da=date, Sp=species.
Table 2 and Fig. 3). There was significant variation
in this age group of C. stellatus between shores in
C. montagui adult zones, i.e., GARR had signifi-
cantly higher recruitment of C. stellatus than the
other two shores. However, there was no such var-
iation among shores at C. stellatus adult zones.
Significant levels of variation between sites within
shores occurred at all shores except BBW and CPK
(in which C. montagui zones were sampled). The
amount of variation associated with shore was large
relative to sites and residual (replicates within sites)
(Table 2).
3.2.2. C. montagui
C. montagui recruits of 0–3 months old did not vary
between different adult zones (non-significant zone
effect, df =1, F =0.08, P=0.797, Table 2 and Fig. 3).
Although Cochran’s C was still significant following
transformation, as the relevant term in the analysis is
non-significant, we can be confident in the interpreta-
tion of no adult zone effect on C. montagui recruitment
(see Methods). Shore to shore variation occurred at C.
montagui adult zones, with 0–3 month recruitment at
GARR being significantly higher than either BBW or
CPK. However there was also variation in the C. stel-
latus adult zone, both NOH and BBE had significantly
s stellatus and C. montagui in SW Ireland
F P
8 0.05 0.841
4 53.70 0.000
3 0.09 0.961
9 16.31 0.056
4 0.50 0.697
4 103.03 0.009
0 21.01 0.000
5 0.83 00.436
9 0.59 0.646
4 0.33 0.804
0 5.76 0.000
2
ntagui zoneNC. stellatus zone
llatus zoneNC. montagui zone
ntagui NC. stellatus
llatus NC. montagui
r dates, n =9. SNK=Student–Newman–Keuls. Abbreviations in terms:
A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 159
more recruitment than OHK. Variation between sites
within shores only occurred at GARR. Shores and
replicates within site (residual variation) contributed
greatest to the variation in recruitment of 0–3 month
C. montagui and variation associated with sites was
much smaller (Table 2).
3.3. Abundance of cyprids+b17 day old metamorphs
between adult zones (within shores)
Recruitment (cyprids+b17 day metamorphs) of C.
montagui was extremely variable between shores and
patches. Abundance of C. montagui was low in both
zones except in patch 1 in the upper level at one of the
shores (BBW). However, there was no difference be-
tween adult zones in either species’ abundance (non-
significant zone� species interaction, df =1, F =46.24,
P=0.093, Table 3). This confirmed that recruitment to
the b17 day age group was not different between the
two adult zones at the same shore.
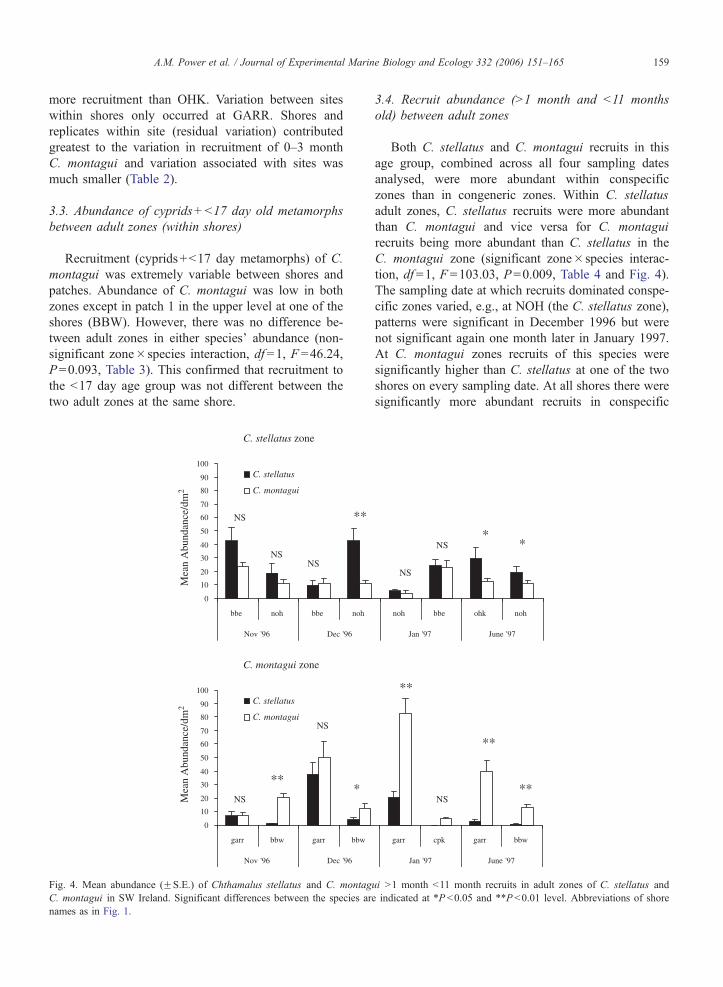
0
10
20
30
40
50
60
70
80
90
100
bbe noh bbe noh
Nov '96 Dec '96
C. stellatus
C. montagui
Mea
n A
bund
ance
/dm
2M
ean
Abu
ndan
ce/d
m2
C. montagui zone
C. stellatus zone
0
10
20
30
40
50
60
70
80
90
100
garr bbw garr bbw
Nov '96 Dec '96
C. stellatus
C. montagui
NS
NS NS
**
NS
**
NS
*
Fig. 4. Mean abundance (FS.E.) of Chthamalus stellatus and C. montag
C. montagui in SW Ireland. Significant differences between the species ar
names as in Fig. 1.
3.4. Recruit abundance (N1 month and b11 months
old) between adult zones
Both C. stellatus and C. montagui recruits in this
age group, combined across all four sampling dates
analysed, were more abundant within conspecific
zones than in congeneric zones. Within C. stellatus
adult zones, C. stellatus recruits were more abundant
than C. montagui and vice versa for C. montagui
recruits being more abundant than C. stellatus in the
C. montagui zone (significant zone� species interac-
tion, df =1, F =103.03, P=0.009, Table 4 and Fig. 4).
The sampling date at which recruits dominated conspe-
cific zones varied, e.g., at NOH (the C. stellatus zone),
patterns were significant in December 1996 but were
not significant again one month later in January 1997.
At C. montagui zones recruits of this species were
significantly higher than C. stellatus at one of the two
shores on every sampling date. At all shores there were
significantly more abundant recruits in conspecific
noh bbe ohk noh
Jan '97 June '97
garr cpk garr bbw
Jan '97 June '97
NS
NS * *
**
NS
**
**
ui N1 month b11 month recruits in adult zones of C. stellatus and
e indicated at *P b0.05 and **P b0.01 level. Abbreviations of shore

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165160
zones by June 1997, at which time recruits were N1
month old with a maximum potential age of 11 months.
3.5. Cyprids as a proportion (of cyprids+b1 month old
metamorphs) between adult zones, shores and sampling
dates
Proportions of cyprid to metamorph did not vary in
the two species according to whether we sampled in the
C. stellatus or C. montagui adult zone (non-significant
zone� species interaction, df =1, F =0.15, P=0.714,
Table 5). There was a consistently higher proportion
of cyprids relative to metamorphs in C. stellatus than in
C. montagui, irrespective of shore or zone sampled
(significant species� shore (zone) interaction, df =4,
F =3.29, P=0.011, Table 5). With respect to the four
dates analysed, C. stellatus always had a higher relative
abundance of cyprids compared to metamorphs than C.
montagui. This proportion was significantly higher in
C. stellatus than in C. montagui on 18 /24 sampling
occasions (significant species�date� shore (zone) in-
teraction, df =12, F =2.30, P=0.007, Table 5 and Fig.
Table 5
Analysis of variance of cyprids as a proportion (of cyprids+b1 month old m
Source of variation df M
Zo 1 76
Sh (Zo) 4 71
Da 3 66
Sp 1 36
Zo�Da 3 47
Zo�Sp 1 0.
Da�Sh (Zo) 12 26
Sp�Sh (Zo) 4 5.
Da�Sp 3 13
Zo�Da�Sp 3 15
Sp�Da�Sh (Zo) 12 4.
Residual 816 1.
Transformation ArcSin
Cochran’s C 0.049
P NS
SNK Multiple comparisons:
Sp within Sh (Zo)
C. montagui zone GA
BB
CP
C. stellatus zone BB
OH
NO
Sh within (Zo� Sp)
C. montagui zone C.
C.
C. stellatus zone C.
C.
Sampling was carried out at three shores nested in each adult zone over four
terms: Zo=zone, Sh=shore, Da=date, Sp=species. Abbreviations of shore
5). There was some variability between the shores in
the proportion of cyprids and metamorphs for both
species but only at the C. montagui adult zones. Pro-
portions were similar for both species at C. stellatus
zones.
4. Discussion
The regulation of population distribution and abun-
dance, through density-dependent relationships be-
tween settlers and adults, may hold true at some shore
heights but not at others (Minchinton and Scheibling,
1991; Menge, 2000). The present study examines ver-
tical distributions and confirms that factors acting on
recruitment and not on settlement were important in
explaining distributions of the barnacles C. stellatus
and C. montagui. A second major objective was to
determine when distinctive adult distributions were
established, by examining the spatial variation in re-
cruitment of each species at local scales in SW Ireland.
Related but smaller-scale studies have previously
shown that neither attached cyprids nor metamorphs
etamorphs) in Chthamalus stellatus and C. montagui in SW Ireland
S F P
.88 1.07 0.359
.65 40.14 0.000
.62 2.53 0.107
9.99 63.03 0.001
.92 1.82 0.197
91 0.15 0.714
.33 14.75 0.000
87 3.29 0.011
.68 3.33 0.057
.82 3.85 0.039
11 2.30 0.007
79
RR C. stellatus NC.montagui
W C. stellatus NC.montagui
K C. stellatus NC.montagui
E C. stellatus NC.montagui
K C. stellatus NC.montagui
H C. stellatus NC.montagui
montagui BBW=GARRNCPK
stellatus BBWNGARRNCPK
montagui NS
stellatus NS
dates, n =18. SNK=Student–Newman–Keuls. Abbreviations of model
s as in Table 1.
_________________________________
C. stellatus adult zone C. montagui adult zone
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
earlyAug
lateAug
Sept Oct earlyAug
lateAug
Sept OctearlyAug
lateAug
Sept Oct earlyAug
lateAug
Sept Oct earlyAug
lateAug
Sept OctearlyAug
lateAug
Sept Oct
bbe ohk noh garr bbw cpk
Mea
n P
ropo
rtio
n/dm
2
C. montagui
C. stellatus
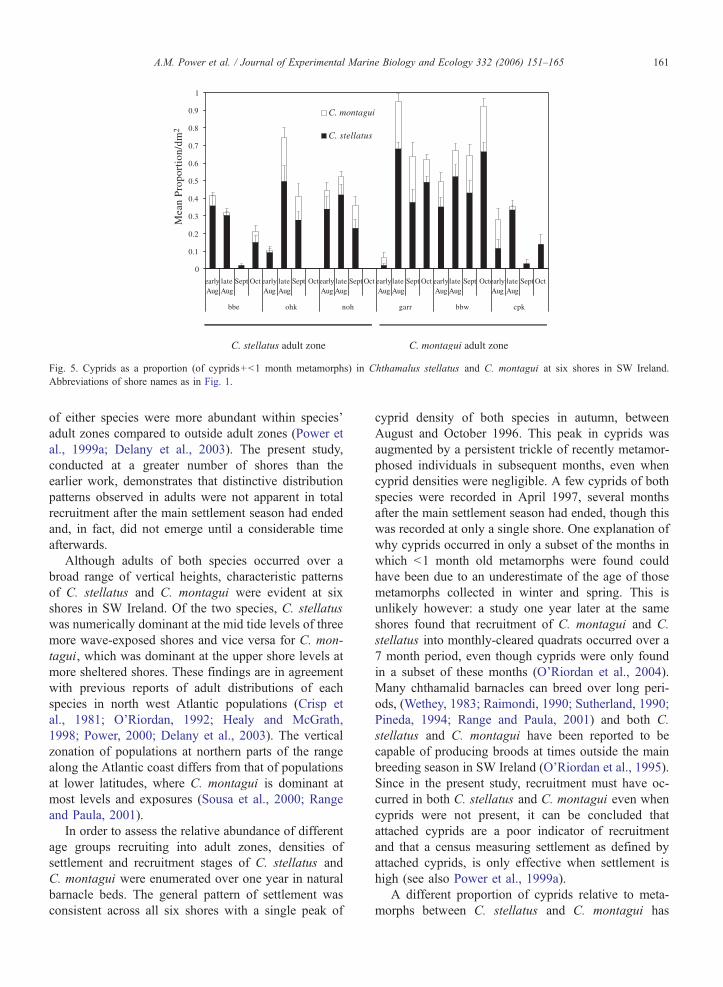
Fig. 5. Cyprids as a proportion (of cyprids+b1 month metamorphs) in Chthamalus stellatus and C. montagui at six shores in SW Ireland.
Abbreviations of shore names as in Fig. 1.
A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 161
of either species were more abundant within species’
adult zones compared to outside adult zones (Power et
al., 1999a; Delany et al., 2003). The present study,
conducted at a greater number of shores than the
earlier work, demonstrates that distinctive distribution
patterns observed in adults were not apparent in total
recruitment after the main settlement season had ended
and, in fact, did not emerge until a considerable time
afterwards.
Although adults of both species occurred over a
broad range of vertical heights, characteristic patterns
of C. stellatus and C. montagui were evident at six
shores in SW Ireland. Of the two species, C. stellatus
was numerically dominant at the mid tide levels of three
more wave-exposed shores and vice versa for C. mon-
tagui, which was dominant at the upper shore levels at
more sheltered shores. These findings are in agreement
with previous reports of adult distributions of each
species in north west Atlantic populations (Crisp et
al., 1981; O’Riordan, 1992; Healy and McGrath,
1998; Power, 2000; Delany et al., 2003). The vertical
zonation of populations at northern parts of the range
along the Atlantic coast differs from that of populations
at lower latitudes, where C. montagui is dominant at
most levels and exposures (Sousa et al., 2000; Range
and Paula, 2001).
In order to assess the relative abundance of different
age groups recruiting into adult zones, densities of
settlement and recruitment stages of C. stellatus and
C. montagui were enumerated over one year in natural
barnacle beds. The general pattern of settlement was
consistent across all six shores with a single peak of
cyprid density of both species in autumn, between
August and October 1996. This peak in cyprids was
augmented by a persistent trickle of recently metamor-
phosed individuals in subsequent months, even when
cyprid densities were negligible. A few cyprids of both
species were recorded in April 1997, several months
after the main settlement season had ended, though this
was recorded at only a single shore. One explanation of
why cyprids occurred in only a subset of the months in
which b1 month old metamorphs were found could
have been due to an underestimate of the age of those
metamorphs collected in winter and spring. This is
unlikely however: a study one year later at the same
shores found that recruitment of C. montagui and C.
stellatus into monthly-cleared quadrats occurred over a
7 month period, even though cyprids were only found
in a subset of these months (O’Riordan et al., 2004).
Many chthamalid barnacles can breed over long peri-
ods, (Wethey, 1983; Raimondi, 1990; Sutherland, 1990;
Pineda, 1994; Range and Paula, 2001) and both C.
stellatus and C. montagui have been reported to be
capable of producing broods at times outside the main
breeding season in SW Ireland (O’Riordan et al., 1995).
Since in the present study, recruitment must have oc-
curred in both C. stellatus and C. montagui even when
cyprids were not present, it can be concluded that
attached cyprids are a poor indicator of recruitment
and that a census measuring settlement as defined by
attached cyprids, is only effective when settlement is
high (see also Power et al., 1999a).
A different proportion of cyprids relative to meta-
morphs between C. stellatus and C. montagui has

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165162
already been shown by comparing total cyprids and
metamorphs collected over a year (Power et al.,
1999a; O’Riordan et al., 2004). This has been shown
in the present study to be a consistent pattern, there are
more cyprids relative to metamorphs in C. stellatus
than in C. montagui, irrespective of the adult zone,
shore or date sampled. This has previously been attrib-
uted to 1) a faster rate of metamorphosis in C. montagui
(see Cruz, 1999; Cruz et al., 2005), 2) temporal sepa-
ration of settlement of cyprids of the two species, e.g.,
during neap tides or at night time in C. montagui and
vice versa during spring tides or in day time in C.
stellatus (e.g., Raimondi, 1990 in C. anisopoma;
Cruz, 1999): the latter would have been overlooked in
the present study by sampling during daytime on spring
tides, see Power et al. (1999a) or 3) a higher metamor-
phic success in C. montagui or higher post-settlement
mortality in C. stellatus (e.g., Pineda, 1994 in Chtha-
malus spp.). If any of these three possibilities are true,
the measurement and interpretation of settlement rates
between the species are confounded since a cyprid is
not an equivalent unit between C. stellatus and C.
montagui (see Power, 2000 for further discussion).
Thus in the present study we have not compared the
species directly, instead we analysed settlement and
recruitment in each species separately (or summed
cyprids and metamorphs) so that potentially different
metamorphic rates will not affect the interpretation of
our results.
At a subset of the shores examined in the present
study, Delany et al. (2003) reported that adult zone had
no effect on total input of either cyprids or b1 month-
old metamorphs over the main settlement season be-
tween August and October 1996. By contrast, in Ply-
mouth dnewly settledT C. stellatus were reported to be
more abundant on the low shore at the most exposed
site, where adults of that species were also relatively
abundant (Burrows, 1988). A more recent study, also
from Plymouth, found that larval choice at settlement
was an important determinant of adult distributions
(Jenkins, 2005). In the present study, recruitment of
animals up to a maximum age of 3 months did not
vary between adult zones in either C. stellatus or C.
montagui. Comparisons between adult zones in the
present study did not depend on accurate ageing of
the individual barnacles because a comparison of re-
cruitment between species was made using a single
point estimate in October, after the majority of settle-
ment had taken place.
In both Delany et al. (2003) and the present study,
upper and mid shore zones were always sampled on
different shores, to maintain statistical independence of
adult zones. This is relevant because of evidence of
reduced recruitment at upper shore levels compared to
lower down on the shore in C. montagui (Kendall and
Bedford, 1987; Cruz, 1999). The opposite pattern has
been documented however, with fewer settlers below
compared to above mid-tide in C. stellatus (probably
actually C. montagui) (Connell, 1961a; Moyse and
Knight-Jones, 1967) and Chthamalus fissus (Menge,
1991). To examine how patterns of recruitment varied
vertically at the same shores, we carried out compar-
isons in 1998 between adult zones within shores for
cyprids and early metamorphs (in this case metamorphs
were b17 days old). Results of this experiment were
consistent with the hypothesis that adult zones were
unimportant in determining early species composition,
i.e., there was no difference in the abundance of either
species between mid and upper shore zones at the two
study shores. There was a large degree of variation in
settlers between shores and patches however, therefore
this result requires confirmation at more shores and
over a longer sampling period. Settlers/recruits of the
related species Chthamalus anisopoma (Raimondi,
1990) and Chthamalus spp. (Pineda, 1994) also did
not show any difference in abundance between shore
heights.
By contrast with early recruitment, a comparison of
predominantly older recruits up to a potential maximum
age of 11 months, did display the same distribution
patterns as adults. There was a significant effect of
adult zone for the entire group considered together
but significant effects were also seen on several dates
within the time period analysed. Particularly at upper-
shore C. montagui adult zones, a significant numerical
dominance of this species was observed 4, 5 or 6
months after the initiation of settlement. However this
pattern was not observed at all shores on any of those
dates and, once observed, was not necessarily main-
tained in subsequent months. Assessing the month
when zonation came about at individual shores was
complicated by the fact that only two of the three shores
in each zone were analysed for each of the sampling
dates (this was done to keep the experimental design
balanced by omitting shores for which there were miss-
ing data). By June 1997 when recruits were potentially
11 months old, all shores displayed significantly higher
recruit abundance within conspecific adult zones for
both species.
Variability in early life stages has been shown to be
very large in intertidal studies (e.g., Caffey, 1985;
Raimondi, 1990; Underwood and Chapman, 1996;
Benedetti-Cecchi et al., 2000; Jenkins et al., 2000;
Jeffery, 2003; O’Riordan et al., 2004). The spatial

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 163
scales at which variation occurs may indicate which
factors are important in the population dynamics of a
species. For example, if recruitment varies more at large
spatial scales, this is more likely to be due to larval
supply which also varies at large spatial scales, whereas
factors such as physical stress are more likely to cause
variability at smaller spatial scales (e.g., Caley et al.,
1996; Underwood and Chapman, 1996). There is evi-
dence to support these general predictions, for example,
at large spatial scales regulation of a species can occur
at settlement in some parts of the geographic range
(where input rates are low) but comes about due to
mortality at other parts of the range (where input is
higher) (Connell, 1985; Menge, 1991, 2000). On a
smaller scale within shores however, Denley and Un-
derwood (1979) found that although settlement was
important in regulating the upper and lower vertical
limits of barnacle species, post-settlement mortality
was more important in governing which species colo-
nised microhabitats within the barnacle zone (shade
patches). Specifying the spatial scale at which variation
has been examined is, at any rate, crucial for making
comparisons between studies and drawing general con-
clusions about the relative importance of various factors
(Underwood and Petraitis, 1993).
Despite homogeneous 0–3 month recruitment into
adult zones in the present study, significant variation
did occur in this age group for both species at all spatial
scales examined. In C. stellatus, more than twice as
much of this variation was attributed to the larger scale
of shore (i.e., 1000s of metres) than was attributed to
sites within shores (10s of metres) or residual error
(within 4 m2). This implies that factors at the scale of
shores were more important than those related to smal-
ler spatial scales (within shores) for 0–3 month C.
stellatus recruits. Similar recruits of C. montagui, var-
ied mainly at the scale of shores but residual variation
within sites (i.e., within 4 m2) was also high, indicating
that C. montagui displayed patchiness at both larger
and smaller spatial scales. Variation at the scale of
shores occurred in both species of recruits inside C.
montagui zones because of a consistently higher re-
cruitment of both species at GARR than at the other
two C. montagui zone shores. GARR, in common with
other C. montagui zones, was sampled at upper shore
levels, which are all subjected to fewer hours of tidal
inundation. Variation between GARR and the other
upper zones may have arisen due to the fact that of
the three shores where C. montagui zones were sam-
pled, this was the most exposed to wave action. There is
an increasing body of evidence to suggest that recruit-
ment is higher at sites with a greater exchange of water
(Hatton, 1938; Caffey, 1985; Raimondi, 1990; Bertness
et al., 1992; Burrows, 1988; Menge, 2000; Hancock
and Petraitis, 2001). At exposed shores where C. stel-
latus zones (mid-shore) were sampled, only C. monta-
gui recruit abundance varied between shores, as all
shores had similar abundances of C. stellatus.
Post-settlement mortality has long been described in
chthamalid barnacles, for example, classic studies into
the emergence of intertidal distributions of dC.stellatusT before the recognition of C. montagui as a
separate species, report that adult vertical patterns at
settlement were unimportant because spat were abun-
dant at middle (Hatton, 1938) and low (Connell,
1961a; Moyse and Knight-Jones, 1967) shore levels
but did not survive there. In the present study, vertical
zonation in both C. stellatus and C. montagui was
indistinct until a minimum of 5 months and a maxi-
mum of 11 months after the settlement season had
begun (when settlement rates were negligible). There-
fore zonation does not occur at settlement and mortal-
ity acting on populations several months after
settlement is more important in structuring vertical
zones in these barnacle species in SW Ireland. There
is evidence of significant differential mortality in indi-
viduals between 6–12 months of age outside conspe-
cific adult zones in C. stellatus and C. montagui
(Delany et al., 2003; Burrows, 1988). The process
which brings about differences in abundance at C.
stellatus and C. montagui zones has been suggested
to be associated with desiccation (Power et al., 2001).
C. stellatus was found in higher densities in wetter
areas than in adjacent dry areas on the upper shore.
Conversely, a higher abundance of C. montagui was
found in drier areas of the upper shore compared to
adjacent wet areas (Power et al., 2001). In that study
the pattern was significant in C. montagui (maximum
age of 5 months) before it was significant in C. stella-
tus (maximum age of 10 months). A laboratory inves-
tigation into species-specific mortality under different
submersion regimes has shown that C. montagui suf-
fered less mortality in the shortest immersion regime
but C. stellatus showed most mortality in the same
treatment (Burrows, 1988). Future studies which link
wetness of the habitat to differential survival of C.
stellatus and C. montagui would confirm this as a
causative factor in determining vertical patterns in
both of these barnacle species.
Acknowledgements
We are grateful to the European Community for
funding the work under the MAST-3 programme, con-

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165164
tract MAS3-CT95-0012. The manuscript was prepared
while AMP was in receipt of a post doctoral fellowship
(PDOC/01/006) funded by the Marine Institute and
National Development Plan (Ireland) under the Marine
RTDI Measure. Comments from an anonymous referee
and A.J. Underwood made a valuable contribution to
the manuscript. [AU]
References
Ballantine, W.J., 1961. A biologically defined exposure scale
for the comparative description of rocky shores. Field Stud.
1, 1–19.
Benedetti-Cecchi, L., Acunto, S., Bulleri, F., Cinelli, F., 2000. Pop-
ulation ecology of the barnacle Chthamalus stellatus in the north-
west Mediterranean. Mar. Ecol. Prog. Ser. 198, 157–170.
Bertness, M.D., Gaines, S.D., Stephens, E.G., Yund, P.O., 1992.
Components of recruitment in populations of the acorn barnacle
Semibalanus balanoides (Linnaeus). J. Exp. Mar. Biol. Ecol. 156,
199–215.
Burrows, M.T., 1988. The comparative biology of Chthamalus stel-
latus (Poli) and Chthamalus montagui Southward. PhD thesis,
University of Manchester.
Burrows, M.T., Hawkins, S.J., Southward, A.J., 1999. Larval devel-
opment of the intertidal barnacles Chthamalus stellatus and
Chthamalus montagui. J. Mar. Biol. Assoc. UK 79, 93–101.
Bushek, D., 1988. Settlement as a major determinant of intertidal
oyster and barnacle distributions along a horizontal gradient.
J. Exp. Mar. Biol. Ecol. 122, 1–18.
Caffey, H.M., 1985. Spatial and temporal variation in settlement and
recruitment of intertidal barnacles. Ecol. Monogr. 55, 313–332.
Caley, M.J., Carr, M.H., Hixon, M.A., Hughes, T.P., Jones, G.P.,
Menge, B.A., 1996. Recruitment and the local dynamics of
open marine populations. Ann. Rev. Ecolog. Syst. 27, 477–500.
Carroll, M., 1996. Barnacle population dynamics and recruitment
regulation in southcentral Alaska. J. Exp. Mar. Biol. Ecol. 199,
285–302.
Connell, J.H., 1961a. The influence of interspecific competition and
other factors on the distribution of the barnacle Chthamalus
stellatus. Ecology 42, 710–723.
Connell, J.H., 1961b. Effects of competition, predation by Thais
lapillus, and other factors on natural populations of the barnacle
Balanus balanoides. Ecol. Monogr. 31, 61–104.
Connell, J.H., 1985. The consequences of variation in initial settle-
ment vs. post-settlement mortality in rocky intertidal communi-
ties. J. Exp. Mar. Biol. Ecol. 93, 11–45.
Crisp, D.J., Southward, A.J., Southward, E.C., 1981. On the distri-
bution of the intertidal barnacles Chthamalus stellatus, Chthama-
lus montagui and Euraphia depressa. J. Mar. Biol. Assoc. UK 61,
359–380.
Cruz, T., 1999. Settlement patterns of Chthamalus spp. at Praia da
Oliveirinha (SW Portugal). Acta Oecol. 20, 285–287.
Cruz, T., Castro, J.J., Delany, J., McGrath, D., Myers, A.A., O’Rior-
dan, R.M., Power, A.-M., Rabaca, J., Hawkins, S.J., 2005. Tidal
rates of settlement of the intertidal barnacles Chthamalus stellatus
and Chthamalus montagui in western Europe: the influence of the
night/day cycle. J. Exp. Mar. Biol. Ecol. 318, 51–60.
Delany, J., Myers, A.A., McGrath, D., O’Riordan, R.M., Power,
A.M., 2003. Role of post-settlement mortality and dsupply-sideTecology in setting patterns of intertidal distribution in the chtha-
malid barnacles Chthamalus montagui and C. stellatus. Mar.
Ecol. Prog. Ser. 249, 207–214.
Denley, E.J., Underwood, A.J., 1979. Experiments in factors influ-
encing settlement, survival, and growth of two species of barna-
cles in New South Wales. J. Exp. Mar. Biol. Ecol. 36, 269–293.
Gaines, S.D., Bertness, M., 1993. The dynamics of juvenile dispersal:
why field ecologists must integrate. Ecology 74, 2430–2435.
Gaines, S., Roughgarden, J., 1985. Larval settlement rate: a leading
determinant of structure in an ecological community of the marine
intertidal zone. Proc. Nat. Acad. Sci. 82, 3707–3711.
Gaines, S., Brown, S., Roughgarden, J., 1985. Spatial variation in
larval concentrations as a cause of spatial variation in settlement
for the barnacle, Balanus glandula. Oecologia 67, 267–272.
Grosberg, R.K., 1982. Intertidal zonation of barnacles: the influence
of planktonic zonation of larvae on vertical distribution of adults.
Ecology 63, 894–899.
Hancock, K.M., Petraitis, P.S., 2001. Effects of herbivorous snails and
macroalgal canopy on recruitment and early survivorship of the
barnacle Semibalanus balanoides (L.). J. Exp. Mar. Biol. Ecol.
257, 205–218.
Hatton, H., 1938. Essais de bionomie explicative sur quelques
especes intercotidales d’algues et d’animaux. Ann. Inst. Ocean.
17, 241–348.
Healy, B., McGrath, D., 1998. Marine fauna of Co. Wexford, Ireland:
the fauna of rocky shores and sandy beaches. Ir. Fish. Invest.,
New Ser. 2, 1–71.
Hughes, T.P., 1990. Recruitment limitation, mortality, and population
regulation in open systems: a case study. Ecology 71, 12–20.
Jarrett, J.N., Pechenik, J.A., 1997. Temporal variation in cyprid
quality and juvenile growth capacity for an interidal barnacle.
Ecology 78, 1262–1265.
Jeffery, C.J., 2003. Determination of abundance and distribution of an
intertidal barnacle: settlement or post-settlement mortality? Mar.
Ecol. Prog. Ser. 246, 291–305.
Jenkins, S.R., 2005. Larval habitat selection, not larval supply, deter-
mines settlement patterns and adult distributions in two chthama-
lid barnacles. J. Anim. Ecol. 74, 893–904.
Jenkins, S.R., Aberg, P., Cervin, C., Coleman, R.A., Delany, J.,
Della-Santina, P., Hawkins, S.J., LaCroix, E., Myers, A.A.,
Lindegarth, M., Power, A.-M., Roberts, M.F., Hartnoll, R.G.,
2000. Spatial and temporal variation in settlement and recruit-
ment of the intertidal barnacle Semibalanus balanoides (Crusta-
cea: Cirripedia) over a European scale. J. Exp. Mar. Biol. Ecol.
243, 209–225.
Kendall, M.A., Bedford, M.L., 1987. Reproduction and recruitment of
the barnacle Chthamalus montagui at Aberystwyth (mid-Wales).
Mar. Ecol. Prog. Ser. 38, 305–308.
Lively, C.M., Raimondi, P.T., Delph, L.F., 1993. Intertidal community
structure: space–time interactions in the northern Gulf of Califor-
nia. Ecology 74, 162–173.
Lubchenco, J., Menge, B.A., 1978. Community development and
persistence in a low rocky intertidal zone. Ecol. Monogr. 59,
67–94.
Menge, B.A., 1991. Relative importance of recruitment and other
causes of variation in rocky intertidal community structure. J. Exp.
Mar. Biol. Ecol. 146, 69–100.
Menge, B.A., 2000. Recruitment vs. post recruitment processes as
determinants of barnacle population abundance. Ecol. Monogr.
70, 265–288.
Minchinton, T.E., Scheibling, R.E., 1991. The influence of larval
supply and settlement on the population structure of barnacles.
Ecology 72, 1867–1879.

A.M. Power et al. / Journal of Experimental Marine Biology and Ecology 332 (2006) 151–165 165
Miron, G., Boudreau, B., Bourget, E., 1999. Intertidal barnacle dis-
tribution: a case study using multiple working hypothesis. Mar.
Ecol. Prog. Ser. 189, 205–219.
Moyse, J., Knight-Jones, E.W., 1967. Biology of cirripede larvae.
Proceedings of the Symposium on Crustacea, Ernakulam, 1965,
Part 2, Marine Biological Society of India. Bangalore Press,
Bengal, pp. 595–611.
O’Riordan, R.M., 1992. Reproduction and recruitment of two inter-
tidal chthamalid barnacles. PhD thesis, National University of
Ireland, Cork.
O’Riordan, R.M., Myers, A.A., Cross, T.F., 1995. The reproductive
cycles of Chthamalus stellatus (Poli) and C. montagui Southward
in south-western Ireland. J. Exp. Mar. Biol. Ecol. 190, 17–38.
O’Riordan, R.M.,Myers, A.A.,McGrath, D., Delany, J., Power, A.-M.,
1999. The sizes at settlement in natural populations of the cyprids of
Chthamalus montagui and Chthamalus stellatus. J. Mar. Biol.
Assoc. UK 79, 365–366.
O’Riordan, R.M., Arenas, F., Arrontes, J., Castro, J.J., Cruz, T.,
Delany, J., Martınez, B., Fernandez, C., Hawkins, S.J.,
McGrath, D., Myers, A.A., Oliveros, J., Pannacciulli, F.G.,
Power, A.-M., Relini, G., Rico, J.M., Silva, T., 2004. Spatial
variation in the recruitment of the intertidal barnacles Chthama-
lus montagui Southward and Chthamalus stellatus (Poli) (Crus-
tacea: Cirripedia) over an European scale. J. Exp. Mar. Biol.
Ecol. 304, 243–264.
Pineda, J., 1994. Spatial and temporal patterns in barnacle settlement
along a southern California rocky shore. Mar. Ecol. Prog. Ser.
107, 125–138.
Pineda, J., Riebensahm, D., Medeiros-Bergen, D., 2002. Semibalanus
balanoides in winter and spring: larval concentration, settlement,
and substrate occupancy. Mar. Biol. 140, 789–800.
Power, A.M., 2000. The ecology of chthamalid barnacles: an evalu-
ation of settlement ands recruitment in regulating intertidal distri-
bution of Chthamalus stellatus and C. montagui in south-west
Ireland. PhD thesis, National University of Ireland, Cork.
Power, A.-M., Delany, J., Myers, A.A., O’Riordan, R.M., McGrath,
D., 1999. Prolonged settlement and prediction of recruitment of
two sympatric intertidal Chthamalus species from southwest Ire-
land. J. Mar. Biol. Assoc. UK 79, 941–943.
Power, A.-M., Piyapattanakorn, S., O’Riordan, R.M., Iyengar, A.,
Myers, A.A., Hawkins, S.J., Delany, J., McGrath, D., Maclean,
N., 1999. Verification of cyprid size as a tool in the identification
of two European species of Chthamalus barnacles using mtDNA
RFLP analysis. Mar. Ecol. Prog. Ser. 191, 251–256.
Power, A.-M., Myers, A.A., O’Riordan, R.M., McGrath, D., Delany,
J., 2001. An investigation into rock surface wetness as a parameter
contributing to the distribution of the intertidal barnacles, Chtha-
malus stellatus and C. montagui. Estuar. Coast. Shelf Sci. 52,
349–356.
Raimondi, P.T., 1988. Settlement cues and determination of the
vertical limit of an intertidal barnacle. Ecology 69, 400–407.
Raimondi, P.T., 1990. Patterns, mechanisms, consequences of vari-
ability in settlement and recruitment of an intertidal barnacle.
Ecol. Monogr. 60, 282–309.
Range, P., Paula, J., 2001. Distribution, abundance and recruitment of
Chthamalus (Crustacea: Cirripedia) populations along the central
coast of Portugal. J. Mar. Biol. Assoc. UK 81, 461–468.
Ross, P.M., Burrows, M.T., Hawkins, S.J., Southward, A.J., Ryan,
K.P., 2003. A key for the identification of the nauplii of common
barnacles of the British Isles, with emphasis on Chthamalus.
J. Crustac. Biol. 23, 328–340.
Roughgarden, J., Iwasa, Y., Baxter, C., 1985. Demographic theory for
an open marine population with space-limited recruitment. Ecol-
ogy 66, 54–67.
Sousa, E.B., Cruz, T., Castro, J.J., 2000. Distribution and abundance
of co-occurring chthamalid barnacles Chthamalus montagui and
Chthamalus stellatus (Crustacea, Cirripedia) on the southwest
coast of Portugal. Hydrobiologia 440, 339–345.
Strathmann, R.R., Branscomb, E.S., 1979. Adequacy of cues to
favourable sites used by settling larvae of two intertidal barna-
cles. In: Stancyk, S.E. (Ed.), Reproductive Ecology of Marine
Invertebrates. University of South Carolina Press, Columbia, S.C.,
pp. 77–89.
Sutherland, J.P., 1990. Recruitment regulates demographic variation
in a tropical intertidal barnacle. Ecology 71, 955–972.
Underwood, A.J., 1997. Experiments in Ecology, their Logical De-
sign and Interpretation Using Analysis of Variance. Cambridge
University Press, Cambridge.
Underwood, A.J., Chapman, M.G., 1996. Scales of spatial patterns of
distribution of intertidal invertebrates. Oecologia 107, 212–224.
Underwood, A.J., Denley, E.J., 1984. Paradigms, explanations,
and generalities in models for the structure of intertidal com-
munities on rocky shores. In: Strong, D.R., Simberloff, D.,
Abele, L., Thistle, A.B. (Eds.), Ecological Communities: Concep-
tual Issues and the Evidence. Princeton University Press, Prince-
ton, pp. 151-130.
Underwood, A.J., Fairweather, P.G., 1989. Supply-side ecology and
benthic marine assemblages. Tree 4, 16–20.
Underwood, A.J., Petraitis, P.S., 1993. Structure of intertidal
assemblages in different locations: how can local processes
be compared? In: Ricklefs, R., Schluter, D. (Eds.), Species Di-
versity in Ecological Communities. University of Chicago Press,
pp. 38–51.
Wethey, D.S., 1983. Geographic limits and local zonation: the barna-
cles Semibalanus (Balanus) and Chthamalus in New England.
Biol. Bull. 165, 330–341.
Wethey, D.S., 1984. Sun and shade mediate competition in the
barnacles Chthamalus and Semibalanus: a field experiment.
Biol. Bull. 167, 176–185.
Winer, B.J., 1971. Statistical Principles in Experimental Design.
McGraw-Hill, Tokyo.




















