
OSCAR Is a Receptor for Surfactant Protein D That Activates TNF-α Release from Human CCR2+...
15
of March 5, 2015. This information is current as Inflammatory Monocytes + Human CCR2 Release from α D That Activates TNF- OSCAR Is a Receptor for Surfactant Protein Colonna William Vermi, Karsten Skjødt, Erika Crouch and Marco Alex S. Holehouse, Derek E. Byers, Michael J. Holtzman, Alexander D. Barrow, Yaseelan Palarasah, Mattia Bugatti, ol.1402289 http://www.jimmunol.org/content/early/2015/02/24/jimmun published online 25 February 2015 J Immunol Material Supplementary 9.DCSupplemental.html http://www.jimmunol.org/content/suppl/2015/02/24/jimmunol.140228 Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2015 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology at Washington University School of Medicine Library on March 5, 2015 http://www.jimmunol.org/ Downloaded from at Washington University School of Medicine Library on March 5, 2015 http://www.jimmunol.org/ Downloaded from at Washington University School of Medicine Library on March 5, 2015 http://www.jimmunol.org/ Downloaded from at Washington University School of Medicine Library on March 5, 2015 http://www.jimmunol.org/ Downloaded from at Washington University School of Medicine Library on March 5, 2015 http://www.jimmunol.org/ Downloaded from at Washington University School of Medicine Library on March 5, 2015 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of OSCAR Is a Receptor for Surfactant Protein D That Activates TNF-α Release from Human CCR2+...

of March 5, 2015.This information is current as
Inflammatory Monocytes+Human CCR2 Release fromαD That Activates TNF-
OSCAR Is a Receptor for Surfactant Protein
ColonnaWilliam Vermi, Karsten Skjødt, Erika Crouch and MarcoAlex S. Holehouse, Derek E. Byers, Michael J. Holtzman, Alexander D. Barrow, Yaseelan Palarasah, Mattia Bugatti,
ol.1402289http://www.jimmunol.org/content/early/2015/02/24/jimmun
published online 25 February 2015J Immunol
MaterialSupplementary
9.DCSupplemental.htmlhttp://www.jimmunol.org/content/suppl/2015/02/24/jimmunol.140228
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2015 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
OSCAR Is a Receptor for Surfactant Protein D ThatActivates TNF-a Release from Human CCR2+ InflammatoryMonocytes
Alexander D. Barrow,* Yaseelan Palarasah,† Mattia Bugatti,‡ Alex S. Holehouse,x,{
Derek E. Byers,‖ Michael J. Holtzman,‖ William Vermi,*,‡ Karsten Skjødt,†
Erika Crouch,* and Marco Colonna*
Surfactant protein D (SP-D) is critical for maintenance of lung homeostasis and provides a first line of defense to pathogens at
mucosal surfaces. Polymorphisms in the SP-D–encoding gene SFTPD have been associated with chronic obstructive pulmonary
disease and ulcerative colitis. Identification of the immunoreceptors that bind SP-D is essential for understanding its contribution
to lung homeostasis and mucosal defense. We located a putative binding motif for the osteoclast-associated receptor (OSCAR)
within the SP-D collagenous domain. An OSCAR-Fc fusion protein specifically bound to the collagenous region of recombinant
SP-D and captured native SP-D from human bronchoalveolar lavage. OSCAR localized in an intracellular compartment of
alveolar macrophages together with SP-D. Moreover, we found OSCAR on the surface of interstitial lung and blood CCR2+
inflammatory monocytes, which secreted TNF-a when exposed to SP-D in an OSCAR-dependent fashion. OSCAR and SP-D did
not exclusively colocalize in lung, as they were also highly expressed in atherosclerotic plaques of human aorta, supporting a role
for this interaction in atherosclerosis. Our results identify the OSCAR:SP-D interaction as a potential therapeutic target in
chronic inflammatory diseases of the lung as well as other diseases involving tissue accumulation of SP-D, infiltration of inflam-
matory monocytes, and release of TNF-a. The Journal of Immunology, 2015, 194: 000–000.
Surfactant protein D (SP-D) is a member of the collagenouslectins (collectins), which provide a first line of humoralinnate immune defense (1–3). The collectin family also
includes, but is not limited to, mannan-binding lectin (MBL) andsurfactant protein A (SP-A). Collectins are soluble proteins thatare structurally characterized by an N-terminal collagenous re-gion, a flexible coiled coil neck region, and a C-terminal carbo-hydrate recognition domain (CRD), which binds various sugars ina calcium-dependent fashion (4). The hydrophobic N-terminalregion of the SP-D polypeptide encodes two cysteine (Cys) resi-
dues (Cys15 and Cys20). Multimeric SP-D dodecamers can beformed through N-terminal disulfide bonding of trimeric SP-Dmonomers. Within the collectin family, the formation of dodeca-mers is unique to SP-D, which can be observed as the charac-teristic cruciform structures by electron microscopy (5). Thecollectin family is expressed in different mucosae and plays animportant tissue-specific role in the innate immune response (4).SP-D is predominantly secreted by alveolar type II epithelial
cells, but is also produced outside of the lung, in the gastrointestinaland genital mucosae, salivary glands, prostate, kidney, pancreas,skin, and endothelial cells (6). SP-D can act as a pattern recog-nition receptor through binding of the CRD to evolutionary con-served glycolipids and glycoproteins associated with infectiousagents, such as LPS from certain bacterial species or viral enve-lope glycoproteins. SP-D can thus opsonize, neutralize, and ag-glutinate infectious microorganisms predisposing to eliminationby phagocytes. In the lung, SP-D also plays an important ho-meostatic role through CRD-dependent scavenging of surfactantphospholipids by alveolar macrophages (7, 8). SP-D–deficient(Sftpd2/2) mice developed accumulation of surfactant phospho-lipids in the lungs, as well as infiltration of monocytes and theproinflammatory activation of alveolar macrophages, leading tochronic inflammation, emphysema, and fibrosis (9, 10). Correctionof some, but not all, of the pulmonary abnormalities in Sftpd2/2
mice required the transgenic expression of SP-D with an intactcollagenous domain (11). SP-D–deficient children were suscepti-ble to more frequent pneumonias, and long-term outcome wasworse than SP-D–sufficient control children (12).Human SFTPD genotype can influence the assembly, concen-
tration, and biological function of SP-D in vivo (13). Interestingly,polymorphisms in SFTPD have been associated with susceptibil-ity to chronic and infectious lung diseases, such as chronic ob-structive pulmonary disease (COPD) (14, 15), emphysema (16),
*Department of Pathology and Immunology, Washington University School of Med-icine, St. Louis, MO 63110; †Department of Cancer and Inflammation, University ofSouthern Denmark, 5000 Odense, Denmark; ‡Department of Molecular and Trans-lational Medicine, Section of Pathology, School of Medicine, University of Brescia,Brescia 25123, Italy; xDivision of Biology and Biomedical Sciences, WashingtonUniversity, St. Louis, MO 63110; {Center for Biological Systems Engineering, Wash-ington University, St. Louis, MO 63130; and ‖Division of Pulmonary and CriticalCare Medicine, Department of Medicine, Washington University School of Medi-cine, St. Louis, MO 63110
ORCID: 0000-0002-4155-5729 (A.S.H.).
Received for publication September 10, 2014. Accepted for publication January 24,2015.
This work was supported by National Institutes of Health Grant 5R01HL097805(to M.C.) and a Marie Curie International Outgoing Fellowship (to A.D.B.).
Address correspondence and reprint requests to Prof. Marco Colonna at the currentaddress: BJC Institute of Health at Washington University, 425 South Euclid, CampusBox 8118, St. Louis, MO 63110. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BAL, bronchoalveolar lavage; COPD, chronicobstructive pulmonary disease; C1q, complement component 1q; CRD, carbohy-drate recognition domain; Cys, cysteine; ILT, Ig-like transcript; LAIR, leukocyte-associated Ig-like receptor; MBL, mannan-binding lectin; NCRD, neck CRD;OSCAR, osteoclast-associated receptor; SP-A, surfactant protein A; SP-D, surfac-tant protein D; TREM, triggering receptor expressed on myeloid cells.
Copyright� 2015 by The American Association of Immunologists, Inc. 0022-1767/15/$25.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1402289
Published February 25, 2015, doi:10.4049/jimmunol.1402289 at W
ashington University School of M
edicine Library on M
arch 5, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

pneumococcal lung disease (17), and tuberculosis (18), and mayeven influence clinical outcome following lung transplantation(19). Serum SP-D levels have been associated with lung functionor health status in patients with severe COPD (20). SFTPD ge-notype has also been associated with inflammatory bowel dis-eases, such as Crohn’s disease and ulcerative colitis (21). SP-D isalso produced by vascular endothelial cells and has been impli-cated in lipid homeostasis and vascular lipid deposition. Sftpd2/2
mice were protected from diet-induced atherosclerosis (22),whereas a polymorphism of SFTPD has been associated withcoronary artery disease (23). These data suggest an important rolefor SP-D in regulating pulmonary homeostasis in addition to rolesin the gut and vascular system, although the molecular basis forthis remains ill defined.Identification of the immunoreceptors that capture SP-D and
transduce intracellular signaling is therefore essential for under-standing how SP-D contributes to lung homeostasis and in-nate mucosal defense and SFTPD disease associations (3, 24).Osteoclast-associated receptor (OSCAR) is an activating receptorfor collagen expressed by osteoclasts that costimulates osteo-clastogenesis (25). OSCAR transmits intracellular signals throughthe associated adapter FcRg (26), which contains an ITAM thatrecruits the protein tyrosine kinase Syk. Although in mouse OS-CAR is exclusively expressed in osteoclasts, human OSCAR wasreported to be also expressed on monocytes, macrophages, neu-trophils, and dendritic cells (26). Human OSCAR was shown toenhance the proinflammatory response of monocytes, although themonocyte subset that specifically expressed OSCAR was not de-scribed (26, 27). The wider expression of OSCAR by humanmyeloid immune cells suggested to us that OSCAR might playa role in innate immunity in addition to the reported role inosteoclastogenesis. In this study, we identified SP-D as a candidateligand for OSCAR by a bioinformatics search for proteins en-coding OSCAR-binding motifs (25). We further demonstrated thatOSCAR and SP-D colocalized in alveolar macrophages and thatthe OSCAR:SP-D interaction triggered TNF-a production byinflammatory monocytes.
Materials and MethodsBioinformatics
To initially identify putative OSCAR-binding candidate proteins for furtherscrutiny, an exhaustive protein BLAST search was carried out using allpermutations of the 9-aa minimum OSCAR-binding motif and side chain–binding variants defined previously (25): G[A|P|G]PG[P|A][A|S]G[F|D|S|Y][A|R|P|Q]. In addition, a less stringent but longer 12-aa con-sensus OSCAR-binding sequence, derived from the same data and usedto generate the minimum OSCAR-binding sequence, was also used:GXPGPX9GFX9GXP (where X is any amino acid). BLAST searches weredone with an expected threshold of 20,000, a word size of 2, and using thePAM30 matrix, as recommended by National Center for BiotechnologyInformation when searching for short, nearly exact matches. Candidatemotifs in both collagen and noncollagen proteins were identified. Fornoncollagen proteins, putative motifs were selected in which proteinBLAST provided an alignment of 80% or better, although one or twosubstitutions between amino acids of similar physiochemical propertieswere treated as matching. As an additional crucial criterion, putative motifswere also required to lie within a collagenous domain, as had previously beenidentified either experimentally or through computational approaches. Forcollagens, an 8/9-aa sequence alignment with one of the minimum OSCAR-binding motif variant sequences was required for a motif to be recognized,although for the vast majority a perfect alignment was obtained. In the non-collagen proteins, the biological roles of each protein could be broadly clas-sified into receptor proteins, secreted proteins, or extracellular matrix proteins.
Plasmids
The human OSCAR-Fc construct has been described before (25). Theextracellular domain of human leukocyte-associated Ig-like receptor(LAIR)-1 was amplified from plasmid DNA containing the human LAIR-1
cDNA with Platinum Pfx DNA Polymerase (Invitrogen) using the fol-lowing forward, 59-CATCCTCGAGCAGGAGGAAGATCTGCCCAG-39and reverse, 59-GAATTCTAGAATGCTCAGCTTTCAGGCCTTG-39primers (XhoI and XbaI restriction sites underlined). XhoI- and XbaI-restricted PCR products were cloned into the Signal Ig plus vector,which encoded the LAIR-1 ectodomain in frame with human IgG1 Fc.Cloning and expression of Ig-like transcript (ILT)-1, -3, and -7 and trig-gering receptor expressed on myeloid cells (TREM)-1 and -2 Fc-fusionproteins were as described (28).
Cell culture, expression, and purification of recombinantFc-fusion proteins
The 293Twere transiently transfected with 48 mg plasmid DNA/10-cm dishusing Lipofectamine 2000 diluted in Optimem (both Invitrogen), accordingto the manufacturer’s instructions. Fc-fusion proteins were purified fromserum-free culture supernatants by protein A affinity chromatography.
Recombinant collectin and collectin-like molecules
The cloning and expression of recombinant full-length human SP-D and thefull-length and mutant isoforms of rat SP-D have been described in detailbefore (5, 29). Briefly, S15, 20 is a recombinant rat SP-D mutant witha substitution of serine for Cys15 and Cys20 that is assembled exclusivelyas SP-D trimers (29). Mini SP-D is a mutant form of rat SP-D that lackstwo internal exons (3, 4) encoding the SP-D collagenous domain. The neckCRD (NCRD) (trimeric neck + CRD) is a rat SP-D truncation mutantlacking the N-terminal peptide and collagen domains that retains lectinactivity (5). Recombinant human MBL was purchased from Sino Bio-logical (Beijing, China), and complement component 1q (C1q) purifiedfrom human serum was purchased from Complement Technology. Theendotoxin level in our stock (112 mg/ml) human SP-D reagent was de-termined at 120 EU/ml by Limulus amebocyte lysate assay (Lonza).
Immobilization of Abs, proteins, and peptides
Collagen I from rat tails and BSAwere purchased from Sigma-Aldrich. BSA(2%), collagens (2.5 mg/ml), recombinant SP-D, and isoforms (2.5 mg/ml)were immobilized onto either tissue culture or ELISA plates overnight in100 ml of 0.01 M acetic acid at 4˚C. Fc-fusion proteins (0.25 mg/ml) wereimmobilized onto plates in 100 ml coating buffer (15 mM Na2CO3, 35mM NaHCO3 [pH 9.6]) overnight at 4˚C. All peptides were certified LPSfree by Limulus amebocyte lysate assay (Lonza). All wells were washedthree times in TBS to remove excess proteins before blocking in 5% BSAin TBS for 1 h at 37˚C prior to performing binding assays.
Solid- and soluble-phase SP-D–binding assays
For solid-phase SP-D–binding assays, 5 mg/ml each Fc-fusion protein in100 ml TBS-binding buffer (10 mM Tris.HCl [pH 7.5], 150 mM NaCl,0.1% BSA, 0.05% Tween 20 6 5 mM CaCl2) was incubated at roomtemperature for 1 h in ELISA plates. Wells were then washed five times inTBS-binding buffer6 5 mM Ca2+ before incubating with 1:5000 goat anti-human Ig-HRP conjugate (Southern Biotech) for 1 h in TBS-binding buffer6 5 mM Ca2+. Wells were then washed an additional five times in TBS-binding buffer 6 5 mM Ca2+ before developing with O-phenylenediaminedihydrochloride, and the absorbance at 490 nM was recorded. For soluble-phase SP-D–binding assays, 2-fold dilutions of either recombinant humanSP-D dodecamer (starting at 125 ng/ml) or human bronchoalveolar lavage(BAL) (starting at 1/4 dilution) were made in TBS-binding buffer andincubated in Fc-fusion–coated wells for 3 h. Captured SP-D was detectedusing 2 mg/ml anti-human SP-D mAb (Hyb-246-04) for 1 h at roomtemperature, followed by rabbit anti-mouse IgG-HRP (Dako).
Generation and purification of mAbs
To generate blocking mAbs to the OSCAR ectodomain, NMRI mice wereimmunized s.c. three times with 20 mg human OSCAR-Fc. For generationof mAbs to the OSCAR cytoplasmic tail, mice were immunized witha synthetic peptide, representing the last 14 C-terminal residues (Cys-DWRSQNRAPAGIRP) of human OSCAR. The synthetic peptides werecoupled to diphtheria toxoid via the cysteine using amine-to-sulfhydrylcross-linker (Thermo Scientific). Three days prior to the fusion, the micereceived an i.v. injection with 50 mg immunogen administered togetherwith adrenalin. The fusion and selection were done essentially as described(30). The SP2/0-AG14 myeloma cell line was used as fusion partner.Positive clones for the OSCAR ectodomain were selected by differentialscreening of OSCAR-Fc versus an irrelevant Fc fusion by ELISA. Positiveclones for the OSCAR C-terminal cytoplasmic tail were identified byscreening to the peptide used for immunization, described above. Cloningwas performed by limited dilution. Single clones were grown in culture
2 OSCAR IS A NOVEL MONOCYTE SP-D RECEPTOR
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

flasks in RPMI 1640 plus 10% FCS, and mAbs were purified from culturesupernatant by protein A affinity chromatography.
Abs
Goat anti-human OSCAR polyclonal Ab was purchased from Santa CruzBiotechnologies (25). The SP-D mAb 245-01 was purchased from EnzoLife Sciences. The anti–SP-D mAb 246-04 was purchased from BioportoDiagnostics (Gentofte, Denmark). The mAb KP1, which recognizes humanCD68, was purchased from Serotec. Donkey anti-goat IgG Alexa 568and donkey anti-mouse IgG Alexa 488 were purchased from Invitrogen.Purified and PE-conjugated anti-human OSCAR mAb 11.1CN5 was pur-chased from Beckman Coulter. The HLA-DR-allophycocyanin, CD16-FITC, and CD56-PE-Cy7 anti-human Abs, used to define human mono-cytes, granulocytes, and NK cells, respectively, were purchased from BDPharmingen, and the CD14-Pacific Blue, CD45-AF700, and CCR2-PerCP-Cy5.5 anti-human Abs were all purchased from BioLegend. Mouse mAbto thyroid transcription factor 1, clone SPT24, was from NovocastraLaboratories. Mouse mAb to human CD163, which recognizes cells of themonocyte/macrophage lineage, was from Neomarkers. mAb clone 13-9-17was used in immunostainings for the OSCAR C terminus. Rabbit anti–SP-D clone H-120 was from Santa Cruz. Rabbit isotype control Ab was fromCell Signaling, and mouse IgG1 isotype control was from BD Biosciences.For immunohistochemistry, primary Abs were revealed with EnvisionDako Polymer HRP.
Microscopy
Paraffin-embedded human lung sections from COPD and normal controlsubjects were deparaffinized using citrisolve (Vector Laboratories). Sectionswere then rehydrated in a graded ethanol series before Ag retrieval byboiling in Ag retrieval solution (Vector Laboratories) in a microwave,according to the manufacturer’s instructions. Sections were blocked in fishgel (Sigma-Aldrich) for 1 h before incubating with primary Abs diluted infish gel overnight at 4˚C. Slides were then washed three times in largevolumes of PBS before incubating individually with secondary Abs, re-spectively. Tiramide enhancement of fluorescent immunostaining wasperformed after each individual secondary Ab step, according to themanufacturer’s instructions (Invitrogen). For confocal microscopy, lungsections were mounted with vectashield containing DAPI, which stainednuclei blue (Vector Laboratories), and analyzed on an Olympus FluoviewFV1000 confocal microscope. For light microscopy, sections stainedimmunohistochemically were photographed using the DP-70 Olympusdigital camera mounted on the Olympus BX60 microscope.
Human subjects and BAL isolation
Human clinical samples were obtained from lung transplant donors andrecipients, as described previously (31). In brief, lung explants were har-vested at the time of lung transplantation from COPD recipients (n = 2)with very severe disease (GOLD Stage 4). Control samples were obtainedfrom normal donors (n = 3) using lung explants that were otherwise notuseable for transplantation. BAL was performed under general anesthesiausing standard procedures: briefly, the bronchoscope was placed in theright middle lobe and the lavage was performed using 3 3 50 ml sterilesaline and aspirated under low pressure. The concentration of SP-D inBAL was determined at 3495 ng/ml, as previously described (32).
Flow cytometry
Cells were prepared from human lung explant tissue using protease di-gestion, as described previously (31). Single-cell suspensions were gen-erated from minced lung tissue that was subjected to collagenase (LiberaseBlendzyme III; Roche), hyaluronidase (Sigma-Aldrich), and DNase I(grade II; Roche) digestion for 45 min at 37˚C. For flow cytometry, frozenaliquots of cell preparations were thawed and washed in complete mediumbefore analysis. Following FcR blockade, cell suspensions were incubatedwith labeled Abs on ice prior to flow cytometric analysis. For peripheralblood leukocytes, granulocytes were defined by electronic gating as sidescatterhighCD45+CD562CD16+ cells and monocytes as side scatterlow
CD45+HLA-DR+CD562. Monocytes were further distinguished based onexpression of CD14, CD16, and CCR2 (27).
OSCAR internalization assays
Purified monocytes were left untreated (0 min) or treated with 2.5 mg/mlrecombinant human SP-D at 37˚C for 5, 15, 30, and 60 min, respectively.Monocytes were then placed on ice and washed twice with ice-cold PBSbefore fixing with 2% paraformaldehyde for 10 min on ice. After washingthree times in PBS, OSCAR internalization was determined by flowcytometry as a reduction in monocyte cell surface staining with non-blocking anti-OSCAR mAb, 11.1CN5.
Monocyte isolation
Human monocytes were isolated from 50 ml blood using the MonocyteNegative Isolation Kit (Invitrogen), according to the manufacturer’sinstructions.
Cytokine analysis
A total of 100,000 monocytes in flat-bottom 96-well plates was stimulatedwith 2.5 mg/ml recombinant human SP-D in the presence or absence of 5mg/ml purified mAbs. TNF-a in tissue culture supernatants was deter-mined using the BD CBA human inflammatory cytokine kit (BD Bio-sciences).
Statistical analysis
Statistical significance was determined using GraphPad Prism 6.0. Statis-tical differences were determined by two-tailed Student t test. A p value,0.05 was considered significant.
Study approval
All studies were reviewed and approved by institutional human and animalstudies committees, and all experiments were carried out according tonational and regional legislation and regulations. Informed consent wasobtained from all subjects included in this study.
ResultsOSCAR is a receptor for SP-D
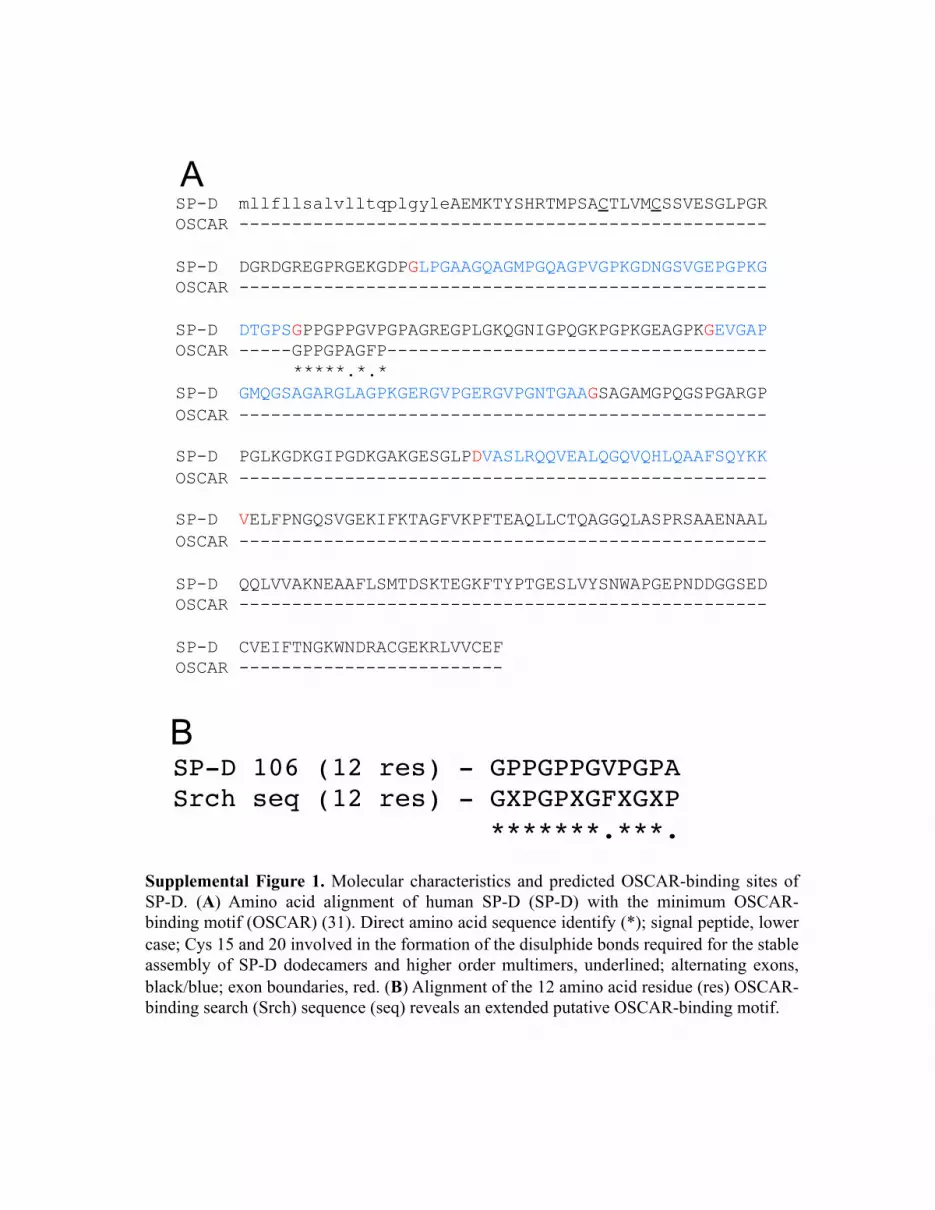
We hypothesized that OSCAR may interact with collagenousmolecules other than extracellular matrix collagens, which mayhave immunological roles. We conducted a bioinformatics searchusing two partially overlapping linear amino acid sequences (seeMaterials and Methods) derived from the previously definedtriple-helical OSCAR-binding motif (25). The search yielded 22noncollagen-based sequences (Table I) and 16 putative collagen-based OSCAR-binding motifs (Supplemental Table I). The col-lectins SP-A, SP-D, and MBL, and collectin-like molecules, suchas C1q (Table I), represented good candidates for interacting withOSCAR-expressing leukocytes because they play central roles intissue-specific immunity (1). Focusing on SP-D, the minimum9-aa OSCAR-binding motif used in the bioinformatics searchaligned with 77.8% direct amino acid sequence identity to the SP-D sequence 106–114 (GPPGPPGVP) (Supplemental Fig. 1A). The12-aa consensus OSCAR-binding motif used in the bioinformaticssearch aligned with 83.3% direct amino acid identity with theSP-D sequence 106–117 (GPPGPPGVPGPA) (Supplemental Fig.1B). The putative OSCAR-binding motifs are encoded by exon 3of human SFTPD, which encodes part of the SP-D collagenousdomain (Supplemental Fig. 1A). Because the 9- and 12-aa searchmotifs are largely overlapping, we conclude that each trimericsubunit of the SP-D monomer encodes one triple-helical OSCAR-binding sequence. Dodecameric SP-D, comprised of four trimericsubunits, would thus be expected to encode four triple-helicalOSCAR-binding motifs.We next determined whether OSCAR could bind to some of the
collectin or collectin-like molecules identified from the motifsearch (Table I). We assayed for binding of a human OSCAR Fc-fusion protein (OSCAR-Fc) to recombinant forms of SP-D, inaddition to C1q and MBL proteins (Fig. 1). OSCAR-Fc bound tofull-length dodecameric human and rat SP-D and trimeric subunitsof rat SP-D (Fig. 1A). OSCAR-Fc did not bind to Mini SP-D,a recombinant rat SP-D lacking the SP-D collagenous domainencoded by exons 2 and 3 of the SP-D gene (5), or a recombinantprotein encoding the NCRD region of rat SP-D (Fig. 1A).OSCAR-Fc binding was specific for SP-D because OSCAR-Fc didnot bind to either C1q or MBL (Fig. 1A), even though theseproteins were predicted to encode putative OSCAR-binding motifs(Table I). OSCAR-Fc binding to full-length dodecameric or tri-meric SP-D or collagen I was significantly increased by, but not
The Journal of Immunology 3
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

wholly dependent on, the presence of 5 mM Ca2+ (Fig. 1A). Be-cause collagen lacks lectin activity and neither recombinant lectin
domains (NCRD) nor Mini SP-D, which encodes an intact lectin
domain, bind to OSCAR (Fig. 1A), we conclude the small in-
crease in calcium-dependent binding of OSCAR-Fc to SP-D or
collagen must be mediated through OSCAR binding to collagen
domains and not the SP-D lectin domain. SP-D binding was not
observed for Fc-fusion proteins that encoded the extracellular
domains of human LAIR1 or ILT3, which are known to bind
collagen (33, 34), or TREM1, TREM2, ILT1, and ILT7 (Fig. 1A
and data not shown). Although SP-D was recently shown to bind
LAIR1 (35), we only detected SP-D binding to mouse LAIR1, but
not human LAIR1 (data not shown). This discrepancy may depend
on the sensitivity of our assay and the relative affinities of mouse
and human LAIR1 for SP-D. In conclusion, our results show that
OSCAR can bind specifically to the SP-D collagenous domain.Because SP-D is secreted as a soluble protein associated with
lung surfactant, we next determined whether OSCAR could bind to
soluble SP-D, as might reflect conditions in vivo. We incubateddilutions of human BAL (Fig. 1B) in wells coated with OSCAR-Fc
or LAIR1-Fc. The amount of protein captured by the immobilized
Fc-fusion proteins was then detected using the Hyb-246-04 mAb,
which is specific for the SP-D CRD (6). Dose-dependent binding
of a protein reactive with the Hyb-246-04 mAb was observed to
immobilized OSCAR-Fc, but not to LAIR1-Fc (Fig. 1B). These
results suggest OSCAR-Fc can capture native SP-D from human
BAL. However, because we have detected a putative OSCAR-binding
motif in SP-A (Table I) and SP-A has been shown to bind to Igs (36),
it may be possible that solid-phase OSCAR-Fc could also be cap-
turing SP-A from BAL. Captured SP-A may, in turn, bind to the Fc
portion of the secondary Abs used to detect bound SP-D. We there-
fore determined whether immobilized OSCAR-Fc could capture
soluble recombinant human SP-D dodecamer in the absence of SP-A.
Solid-phase OSCAR-Fc, but not LAIR1-Fc, captured soluble recom-
binant human SP-D dodecamer (Fig. 1C). These results show that
OSCAR can bind to soluble SP-D dodecamer.
Table I. Amino acid sequences and start positions of putative OSCAR-binding motifs within collagenous proteins (noncollagens)
Biological Class Protein Name Uniprot Sequence Position Notes
Extracellularmatrix andadhesion
EMILIN-1 Q9Y6C2 GPPGPPGLQGPP 820 Secreted adhesive protein, involved inelastic fiber anchoringGPPGPAGPPGSP 829
EMILIN-2 Q9BXX0 TVPGAEGFAGAP 876 Secreted adhesive protein, involved inelastic fiber anchoring/vessel
assemblyProtein sidekick-2 Q58EX2 GAPGPPGVP 1823 Adhesion protein (neuronal
development)Receptors Collectin-12 (scavenger
receptor with C-type lectintype I)
Q5KU26 GPPGPAGER 476 Pathogen recognition/phagocytosisGVPGPRGLPGLP 557
Macrophage receptor withcollagenous structure
(MARCO)
Q9UEW3 GPPGLAGFP 286 Pattern recognition receptor—bindsbacteria
Scavenger receptor class Amember 3 (SCARA3)
Q6AZY7 GAPGPPGPR 457 Scavenges oxidative speciesGPPGPPGSQ 595
Scavenger receptor class Amember 5 (SCARA5)
Q6ZMJ2 GPPGPKGDQ 306 Ferritin receptor—nontransferrin-dependent delivery of iron
Secreted C1q-related factor O75973 GPPGPPGDP 79 Implicated in excitatory synapseregulation on hippocampus neurons
Cell proliferation-inducingprotein 41
A1KY36 GLPGPPGFPGIG 523 —
Collagen and calcium-binding EGF domain-containing protein 1
Q6UXH8 GAPGPRGSPGPP 321 Angiogenic and lymphangioblasticgrowth during embryogenesis
Complement C1q-likeprotein 4
Q86Z23 GPPGPPGPRGPP 70 Implicated in excitatory synapseregulation on hippocampus neurons
Complement C1qsubcomponent subunit B
P02746 GGPGAPGAP 99 Part of the complement cascadeGAPGAPGPK 102
Complement C1qsubcomponent subunit C
P02747 GVPGPMGIPGEP 97 Part of the complement cascade
Complement C1q TNF-related protein 2
Q9BXJ5 GAPGPSGMMGRM 49 —GPPGRTGNRGKP 82
Ectodysplasin-A (EDA) Q92838 GPPGPPGPQGPP 189 Epithelial-mesenchymal signalingduring morphogenesis
Ficolin-2 Q15485 GPPGPNGAP 84 Liver-secreted, innate immunityprotein
Ficolin-3 O75636 GAPGPQGPP 57 May activate lectin complementpathway
Mannose-binding protein C P11226 GNPGPSGSPGPK 80 Innate immune calcium-dependentlectin
Surfactant protein A Q8IWL2 GPPGPMGPP 55 Collectin, found in the lungs, manyroles
Surfactant protein D P35247 GPPGPPGVP 106 Collectin, found in the lungs, manyrolesGPPGPPGVPGPA 106
Other Gliomedin Q6ZMI3 GPPGPPGPPGPP 236 Membrane protein involved information of the nodes of Ranvier
premRNA 39 end processingprotein WDR33
Q9C0J8 GPPGPRGMQGPP 742 Cleavage and polyadenylation of pre-mRNA 39 ends (nuclear protein)
4 OSCAR IS A NOVEL MONOCYTE SP-D RECEPTOR
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
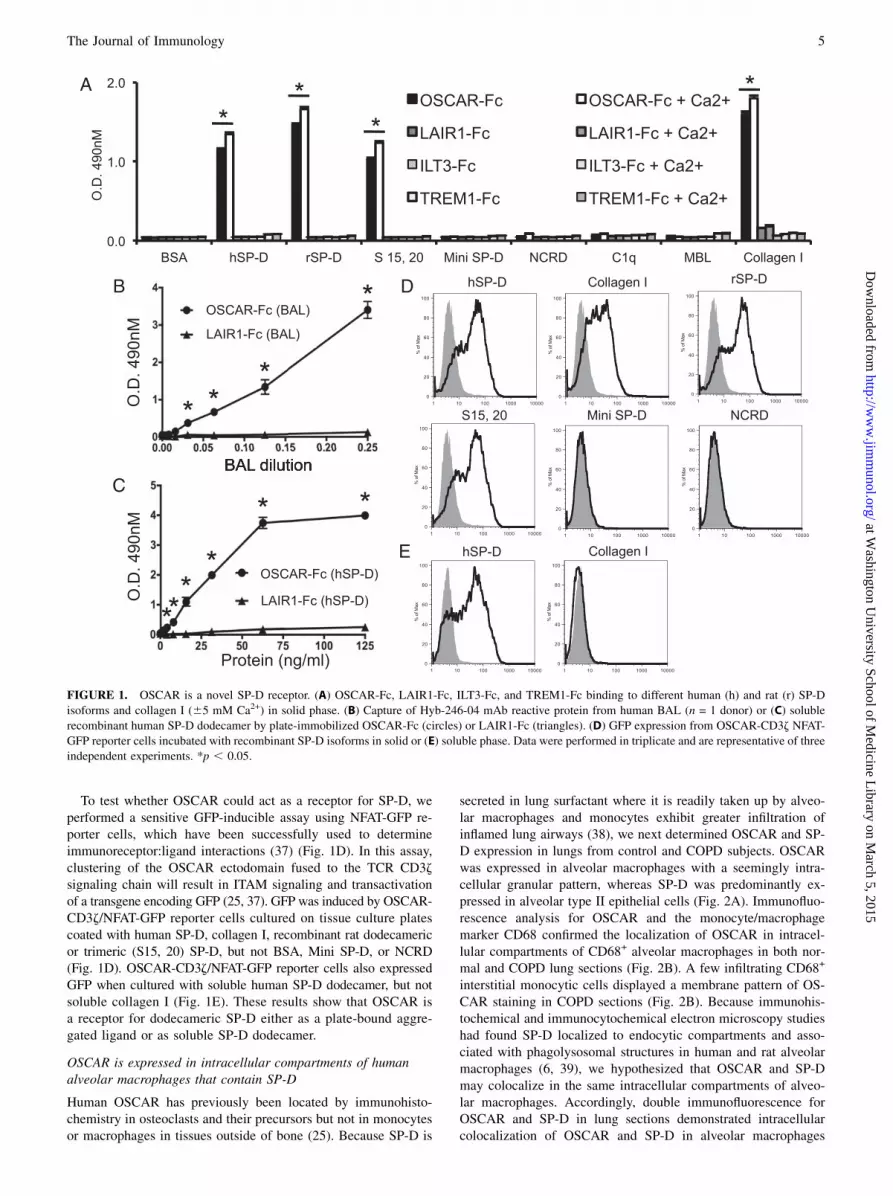
To test whether OSCAR could act as a receptor for SP-D, weperformed a sensitive GFP-inducible assay using NFAT-GFP re-porter cells, which have been successfully used to determineimmunoreceptor:ligand interactions (37) (Fig. 1D). In this assay,clustering of the OSCAR ectodomain fused to the TCR CD3zsignaling chain will result in ITAM signaling and transactivationof a transgene encoding GFP (25, 37). GFP was induced by OSCAR-CD3z/NFAT-GFP reporter cells cultured on tissue culture platescoated with human SP-D, collagen I, recombinant rat dodecamericor trimeric (S15, 20) SP-D, but not BSA, Mini SP-D, or NCRD(Fig. 1D). OSCAR-CD3z/NFAT-GFP reporter cells also expressedGFP when cultured with soluble human SP-D dodecamer, but notsoluble collagen I (Fig. 1E). These results show that OSCAR isa receptor for dodecameric SP-D either as a plate-bound aggre-gated ligand or as soluble SP-D dodecamer.
OSCAR is expressed in intracellular compartments of humanalveolar macrophages that contain SP-D
Human OSCAR has previously been located by immunohisto-chemistry in osteoclasts and their precursors but not in monocytesor macrophages in tissues outside of bone (25). Because SP-D is
secreted in lung surfactant where it is readily taken up by alveo-lar macrophages and monocytes exhibit greater infiltration ofinflamed lung airways (38), we next determined OSCAR and SP-D expression in lungs from control and COPD subjects. OSCARwas expressed in alveolar macrophages with a seemingly intra-cellular granular pattern, whereas SP-D was predominantly ex-pressed in alveolar type II epithelial cells (Fig. 2A). Immunofluo-rescence analysis for OSCAR and the monocyte/macrophagemarker CD68 confirmed the localization of OSCAR in intracel-lular compartments of CD68+ alveolar macrophages in both nor-mal and COPD lung sections (Fig. 2B). A few infiltrating CD68+
interstitial monocytic cells displayed a membrane pattern of OS-CAR staining in COPD sections (Fig. 2B). Because immunohis-tochemical and immunocytochemical electron microscopy studieshad found SP-D localized to endocytic compartments and asso-ciated with phagolysosomal structures in human and rat alveolarmacrophages (6, 39), we hypothesized that OSCAR and SP-Dmay colocalize in the same intracellular compartments of alveo-lar macrophages. Accordingly, double immunofluorescence forOSCAR and SP-D in lung sections demonstrated intracellularcolocalization of OSCAR and SP-D in alveolar macrophages
FIGURE 1. OSCAR is a novel SP-D receptor. (A) OSCAR-Fc, LAIR1-Fc, ILT3-Fc, and TREM1-Fc binding to different human (h) and rat (r) SP-D
isoforms and collagen I (65 mM Ca2+) in solid phase. (B) Capture of Hyb-246-04 mAb reactive protein from human BAL (n = 1 donor) or (C) soluble
recombinant human SP-D dodecamer by plate-immobilized OSCAR-Fc (circles) or LAIR1-Fc (triangles). (D) GFP expression from OSCAR-CD3z NFAT-
GFP reporter cells incubated with recombinant SP-D isoforms in solid or (E) soluble phase. Data were performed in triplicate and are representative of three
independent experiments. *p , 0.05.
The Journal of Immunology 5
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
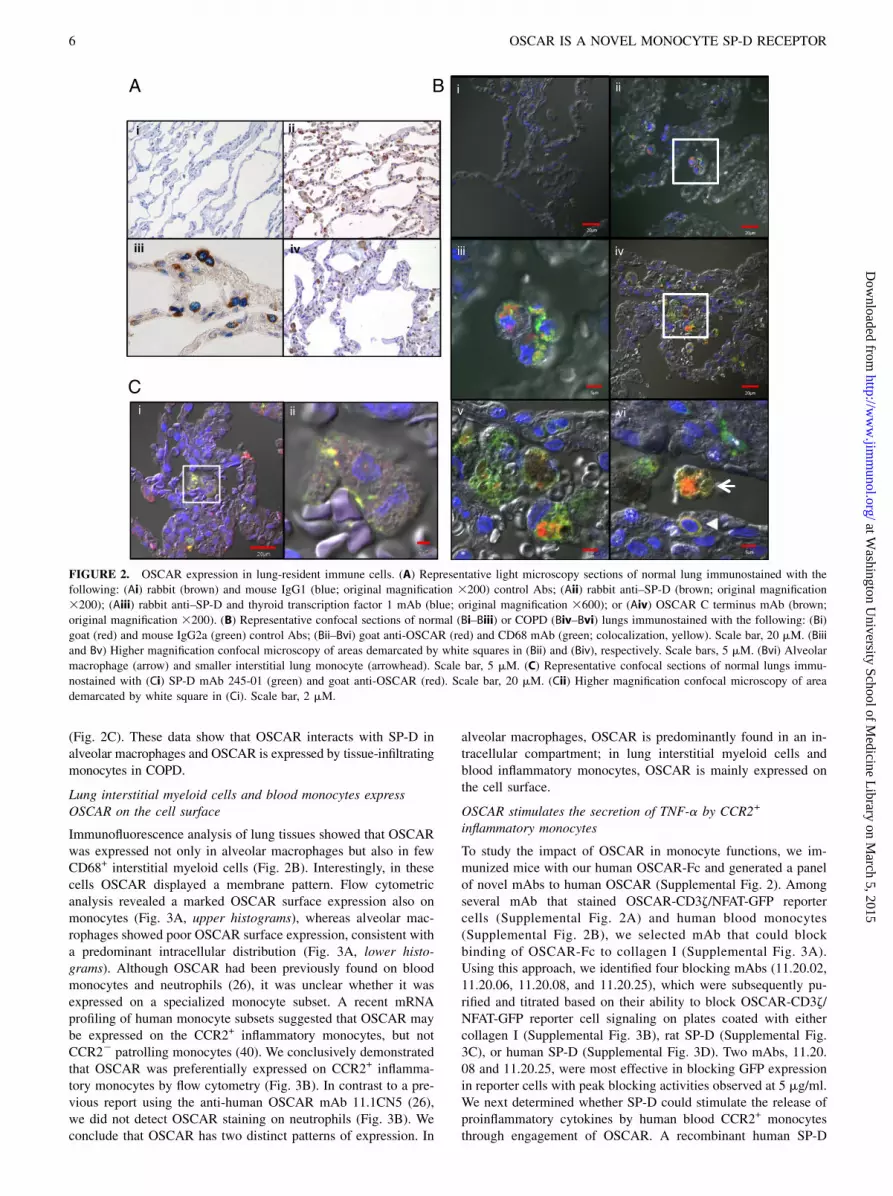
(Fig. 2C). These data show that OSCAR interacts with SP-D inalveolar macrophages and OSCAR is expressed by tissue-infiltratingmonocytes in COPD.
Lung interstitial myeloid cells and blood monocytes expressOSCAR on the cell surface
Immunofluorescence analysis of lung tissues showed that OSCARwas expressed not only in alveolar macrophages but also in fewCD68+ interstitial myeloid cells (Fig. 2B). Interestingly, in thesecells OSCAR displayed a membrane pattern. Flow cytometricanalysis revealed a marked OSCAR surface expression also onmonocytes (Fig. 3A, upper histograms), whereas alveolar mac-rophages showed poor OSCAR surface expression, consistent witha predominant intracellular distribution (Fig. 3A, lower histo-grams). Although OSCAR had been previously found on bloodmonocytes and neutrophils (26), it was unclear whether it wasexpressed on a specialized monocyte subset. A recent mRNAprofiling of human monocyte subsets suggested that OSCAR maybe expressed on the CCR2+ inflammatory monocytes, but notCCR22 patrolling monocytes (40). We conclusively demonstratedthat OSCAR was preferentially expressed on CCR2+ inflamma-tory monocytes by flow cytometry (Fig. 3B). In contrast to a pre-vious report using the anti-human OSCAR mAb 11.1CN5 (26),we did not detect OSCAR staining on neutrophils (Fig. 3B). Weconclude that OSCAR has two distinct patterns of expression. In
alveolar macrophages, OSCAR is predominantly found in an in-tracellular compartment; in lung interstitial myeloid cells andblood inflammatory monocytes, OSCAR is mainly expressed onthe cell surface.
OSCAR stimulates the secretion of TNF-a by CCR2+
inflammatory monocytes
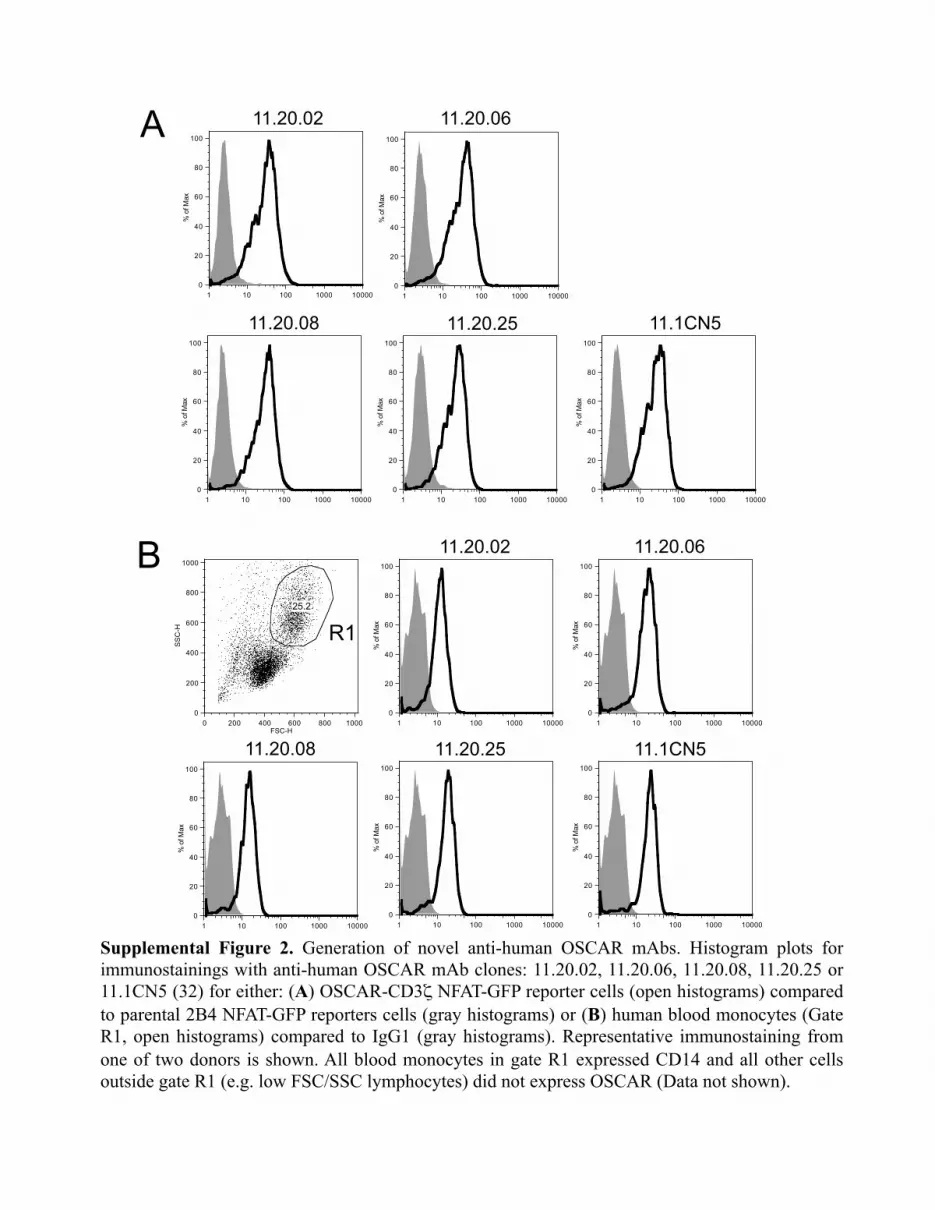
To study the impact of OSCAR in monocyte functions, we im-munized mice with our human OSCAR-Fc and generated a panelof novel mAbs to human OSCAR (Supplemental Fig. 2). Amongseveral mAb that stained OSCAR-CD3z/NFAT-GFP reportercells (Supplemental Fig. 2A) and human blood monocytes(Supplemental Fig. 2B), we selected mAb that could blockbinding of OSCAR-Fc to collagen I (Supplemental Fig. 3A).Using this approach, we identified four blocking mAbs (11.20.02,11.20.06, 11.20.08, and 11.20.25), which were subsequently pu-rified and titrated based on their ability to block OSCAR-CD3z/NFAT-GFP reporter cell signaling on plates coated with eithercollagen I (Supplemental Fig. 3B), rat SP-D (Supplemental Fig.3C), or human SP-D (Supplemental Fig. 3D). Two mAbs, 11.20.08 and 11.20.25, were most effective in blocking GFP expressionin reporter cells with peak blocking activities observed at 5 mg/ml.We next determined whether SP-D could stimulate the release ofproinflammatory cytokines by human blood CCR2+ monocytesthrough engagement of OSCAR. A recombinant human SP-D
FIGURE 2. OSCAR expression in lung-resident immune cells. (A) Representative light microscopy sections of normal lung immunostained with the
following: (Ai) rabbit (brown) and mouse IgG1 (blue; original magnification 3200) control Abs; (Aii) rabbit anti–SP-D (brown; original magnification
3200); (Aiii) rabbit anti–SP-D and thyroid transcription factor 1 mAb (blue; original magnification 3600); or (Aiv) OSCAR C terminus mAb (brown;
original magnification 3200). (B) Representative confocal sections of normal (Bi–Biii) or COPD (Biv–Bvi) lungs immunostained with the following: (Bi)
goat (red) and mouse IgG2a (green) control Abs; (Bii–Bvi) goat anti-OSCAR (red) and CD68 mAb (green; colocalization, yellow). Scale bar, 20 mM. (Biii
and Bv) Higher magnification confocal microscopy of areas demarcated by white squares in (Bii) and (Biv), respectively. Scale bars, 5 mM. (Bvi) Alveolar
macrophage (arrow) and smaller interstitial lung monocyte (arrowhead). Scale bar, 5 mM. (C) Representative confocal sections of normal lungs immu-
nostained with (Ci) SP-D mAb 245-01 (green) and goat anti-OSCAR (red). Scale bar, 20 mM. (Cii) Higher magnification confocal microscopy of area
demarcated by white square in (Ci). Scale bar, 2 mM.
6 OSCAR IS A NOVEL MONOCYTE SP-D RECEPTOR
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

dodecamer stimulated TNF-a release by blood CCR2+ monocytesfrom two different donors, which was blocked by the anti-humanOSCAR mAbs 11.20.08 and 11.20.25, respectively (Fig. 4A).OSCAR was not internalized by monocytes exposed to SP-D(Supplemental Fig. 3E). We conclude the SP-D/OSCAR interac-tion triggers a proinflammatory response in CCR2+ monocytes.
OSCAR is expressed in lipid-laden macrophages infiltratingatherosclerotic plaques
Although SP-D has been implicated in atherosclerosis (22, 23), themechanism is poorly understood. We hypothesized that SP-D mayengage OSCAR in the atherosclerotic plaque, promoting inflam-mation. To test this hypothesis, we investigated OSCAR expres-sion in atherosclerotic lesions of human aorta. OSCAR wasstrongly expressed in numerous lipid-laden macrophages infil-trating the tunica intima and in fewer macrophages penetrating thetunica media. SP-D was found in the vascular endothelium as wellas in many smooth muscular cells of the tunica media, consistentwith previous reports (22, 41) (Fig. 4B). These data suggest thatSP-D may contribute to atherosclerosis by binding OSCAR oninflammatory monocytes that infiltrate the atherosclerotic plaqueand stimulating the release of TNF-a, which will exacerbate thevascular lesion.
DiscussionIn humans and mice, OSCAR is expressed by osteoclasts and theirprecursors and can bind to collagens exposed on bone surfaces tocostimulate osteoclastogenesis (25). In humans, OSCAR is addi-
tionally expressed on monocytes, macrophages, and dendritic cells(26), suggesting that OSCAR may play a role in the immuneresponse. Motivated by this hypothesis, we designed a bio-informatics search for OSCAR-binding motifs in collagenousmolecules that might provide insight into OSCAR immunobiol-ogy. We identified putative OSCAR-binding motifs in the col-lectins, MBL, SP-A, and SP-D, and collectin-like molecules, suchas C1q, and ficolins, that all play important roles in innate im-munity (1). Of the putative ligands tested, we detected binding ofOSCAR-Fc to SP-D, but not to MBL or C1q. OSCAR may notbind to C1q because we only detected OSCAR-binding motifs inthe B and C chains of C1q and none in the A chain (Table I).Because OSCAR only binds to triple-helical motifs in collagenousmolecules and not to linear motifs (25), C1q therefore lacksa motif in the A chain to form a triple-helical sequence capable ofbinding OSCAR. Further structural and biophysical studies arerequired to determine the precise molecular binding specificitiesof OSCAR for collectin and collectin-like molecules.Our study demonstrates that human OSCAR is a novel myeloid
immunoreceptor for SP-D expressed by alveolar macrophages andinflammatory CCR2+ monocytes, which triggers the release ofTNF-a in inflammatory CCR2+ monocytes exposed to SP-D. Theidentification of SP-D receptors is essential to fully understand thefunction of SP-D in different tissues and organs in the context ofinfections and inflammation. Previous studies have shown that SP-D or the closely related collectin SP-A can interact with distinctimmunoreceptors, resulting in different functional outcomes.When not bound to a pathogen, SP-D and SP-A bind through their
FIGURE 3. Lung and blood monocytes ex-
press cell surface OSCAR. (A) Representative
histogram plots for lung cells triple stained with
mAbs to CD14, OSCAR, and HLA-DR (open
histograms) compared with isotype control mAb
(gray histograms) for low (Lo) forward scatter
(FSC)/side scatter (SSC) lung tissue monocytes
(gate R1, upper histogram panels) or high (Hi)
FSC/SSC alveolar macrophages (gate R2, lower
histogram panels). (B) Representative plots for
OSCAR immunostaining of peripheral blood
SSChighCD45+CD562CD16+ granulocytes (up-
per panels). Within the SSClowCD45+CD562
HLA-DR+ monocytic population, CCR2+CD14high
CD162 inflammatory monocytes express OSCAR
(bottom panels), but not CCR22CD14lowCD16high
noninflammatory monocytes (center panels).
Staining is representative of three different donors.
The Journal of Immunology 7
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

lectin domains to SIRP-a, delivering an inhibitory signal, whichprevents activation of mononuclear phagocytes and secretion ofinflammatory cytokines (24). In contrast, when microorganisms orcell debris are bound to the C-type lectin domain, SP-D and SP-Ainteract through their free collagen-like domain to CD91/calreticulin, promoting cell activation (24). SP-D also binds toCD14, interfering with the binding of LPS to the CD14-TLR4-MD2 complex, thereby reducing macrophage inflammatory re-sponses elicited by LPS (42, 43). Finally, a recent study showedthat the Ig-superfamily receptor LAIR1, which is known to bindcollagen, also binds to SP-D, delivering an inhibitory signal thatreduces inflammation (35). Because the OSCAR:SP-D interactionpromotes activation of inflammatory monocytes, we conclude thatthe ultimate functional effect of SP-D depends on the balancebetween inhibitory and activating receptors engaged at any onetime in a given tissue.Our study also demonstrates that, in alveolar macrophages,
OSCAR is predominantly found in an intracellular compartmentwhere it colocalizes with SP-D. This result suggests that OSCARmay capture SP-D in the alveolar space and drive SP-D inter-nalization into a phagolysosomal compartment in alveolar mac-rophages where SP-D and its cargo are processed and degraded.The internalization of SP-D may be crucial for the phagocytosisand killing of pathogens or the removal of necrotic and apoptoticcells in the lung as well as in various other mucosae and organs(2, 44). However, we did not detect any OSCAR internalizationin monocytes exposed to SP-D. It is possible that alveolar mac-rophages are capable of internalizing OSCAR:SP-D complexes,because macrophages are more phagocytic than monocytes. The
distribution of OSCAR between the cell membrane and theendosomal compartment in different myeloid cells might alsodepend on the relative abundance of SP-D to which these cells areexposed in their microenvironment. Alternatively, the differentsubcellular localizations of OSCAR in alveolar macrophages andmonocytes could be caused by the alternative splicing of OSCARtranscripts encoding OSCAR isoforms with different subcellulartrafficking properties (45). Future studies to determine the role, ifany, OSCAR isoforms may play in the cell biology and Ag pro-cessing of SP-D for alveolar macrophages and dendritic cells aretherefore merited.Previous studies have implicated SP-D in atherosclerosis and
coronary heart disease (22, 23, 41). We found that OSCAR ishighly expressed in CD163+ monocytes and macrophages infil-trating the atherosclerotic plaques in aorta. We envision that ex-posure of these OSCAR+ monocytes and macrophages to SP-Dproduced by endothelial cells in the tunica intima and by smoothmuscular cells in the tunica media may lead to the production ofTNF-a, exacerbating the degenerative process. Similar mecha-nisms may operate in lung tissue. For example, monocytes cantraffic into the lung during the steady state (46), but do not pref-erentially enter airway mucosal surfaces, unless there is a proin-flammatory or chemotactic stimulus (38). Therefore, CCR2+
monocytes that patrol lung interstitial tissues in the steady statemay not become exposed to SP-D secreted in airway lung sur-factant. In contrast, monocytes increasingly infiltrate the airwaysin response to a proinflammatory or chemotactic stimuli (38).Thus, upon tissue damage or the induction of proinflammatorystimuli in the lung airways, as might be operating in lung pa-
FIGURE 4. SP-D functionally interacts with OSCAR. (A) TNF-a release by human CCR2+ monocytes from two different donors treated with
soluble SP-D in the presence of either IgG1 or anti-OSCAR blocking mAbs, 11.20.08 or 11.20.25. (B) Representative serial sections of ath-
erosclerotic lesions (i–viii) from human aorta immunostained with the following: (i and v) rabbit anti–SP-D (brown; original magnification
3100); (ii and vi) anti-OSCAR C-terminal mAb 13-9-17 (brown; original magnification 3100); (iii and vii) high power views (original magni-
fication 3200) of areas defined by black rectangles in (ii) and (vi), representing intima and tunica media, respectively; (iv) OSCAR C-terminal
mAb 13-9-17 (brown) and anti-CD163 mAb (blue, original magnification 3600); (viii) rabbit (brown) and mouse IgG1 (blue) control Abs
(original magnification 3200).
8 OSCAR IS A NOVEL MONOCYTE SP-D RECEPTOR
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

thologies like COPD, CCR2+ monocytes will traffic from blood tothe damaged tissues, whereupon they may then become exposedto SP-D and release TNF-a. Thus, our characterization of theOSCAR:SP-D interaction and its functional impact in the secre-tion of proinflammatory cytokines by CCR2+ monocytes recruitedto the tissues may have important therapeutic implications notonly for inflammatory diseases of the lung and the mucosae butalso for atherosclerosis and its complications.
AcknowledgmentsWe thank Dr. Maria G. Quisgaard for providing BAL fluid and Loralyn
Benoit, Stoyan Ivanov, and Hani Suleiman for scientific discussions and
technical assistance.
DisclosuresA.D.B. is a named inventor on a patent application from the University
of Cambridge: Modulation of the activity and differentiation of cells
expressing the osteoclast-associated receptor. U.S. Patent Application
13122637.
References1. Holmskov, U., S. Thiel, and J. C. Jensenius. 2003. Collections and ficolins: hu-
moral lectins of the innate immune defense. Annu. Rev. Immunol. 21: 547–578.2. Wright, J. R. 2005. Immunoregulatory functions of surfactant proteins. Nat. Rev.
Immunol. 5: 58–68.3. Jakel, A., A. S. Qaseem, U. Kishore, and R. B. Sim. 2013. Ligands and receptors
of lung surfactant proteins SP-A and SP-D. Front. Biosci. (Landmark Ed.) 18:1129–1140.
4. Seaton, B. A., E. C. Crouch, F. X. McCormack, J. F. Head, K. L. Hartshorn, andR. Mendelsohn. 2010. Review: structural determinants of pattern recognition bylung collectins. Innate Immun. 16: 143–150.
5. White, M., P. Kingma, T. Tecle, N. Kacak, B. Linders, J. Heuser, E. Crouch, andK. Hartshorn. 2008. Multimerization of surfactant protein D, but not its collagendomain, is required for antiviral and opsonic activities related to influenza virus.J. Immunol. 181: 7936–7943.
6. Madsen, J., A. Kliem, I. Tornoe, K. Skjodt, C. Koch, and U. Holmskov. 2000.Localization of lung surfactant protein D on mucosal surfaces in human tissues.J. Immunol. 164: 5866–5870.
7. Ogasawara, Y., Y. Kuroki, and T. Akino. 1992. Pulmonary surfactant protein Dspecifically binds to phosphatidylinositol. J. Biol. Chem. 267: 21244–21249.
8. Persson, A. V., B. J. Gibbons, J. D. Shoemaker, M. A. Moxley, andW. J. Longmore. 1992. The major glycolipid recognized by SP-D in surfactant isphosphatidylinositol. Biochemistry 31: 12183–12189.
9. Wert, S. E., M. Yoshida, A. M. LeVine, M. Ikegami, T. Jones, G. F. Ross,J. H. Fisher, T. R. Korfhagen, and J. A. Whitsett. 2000. Increased metal-loproteinase activity, oxidant production, and emphysema in surfactant protein Dgene-inactivated mice. Proc. Natl. Acad. Sci. USA 97: 5972–5977.
10. Ikegami, M., C.-L. Na, T. R. Korfhagen, and J. A. Whitsett. 2005. Surfactantprotein D influences surfactant ultrastructure and uptake by alveolar type II cells.Am. J. Physiol. Lung Cell. Mol. Physiol. 288: L552–L561.
11. Kingma, P. S., L. Zhang, M. Ikegami, K. Hartshorn, F. X. McCormack, andJ. A. Whitsett. 2006. Correction of pulmonary abnormalities in Sftpd-/- micerequires the collagenous domain of surfactant protein D. J. Biol. Chem. 281:24496–24505.
12. Griese, M., M. Steinecker, S. Schumacher, A. Braun, P. Lohse, and S. Heinrich.2008. Children with absent surfactant protein D in bronchoalveolar lavage havemore frequently pneumonia. Pediatr. Allergy Immunol. 19: 639–647.
13. Leth-Larsen, R., P. Garred, H. Jensenius, J. Meschi, K. Hartshorn, J. Madsen,I. Tornoe, H. O. Madsen, G. Sørensen, E. Crouch, and U. Holmskov. 2005. Acommon polymorphism in the SFTPD gene influences assembly, function, andconcentration of surfactant protein D. J. Immunol. 174: 1532–1538.
14. van Diemen, C. C., D. S. Postma, Y. S. Aulchenko, P. J. L. M. Snijders,B. A. Oostra, C. M. van Duijn, and H. M. Boezen. 2010. Novel strategy toidentify genetic risk factors for COPD severity: a genetic isolate. Eur. Respir. J.35: 768–775.
15. Foreman, M. G., X. Kong, D. L. DeMeo, S. G. Pillai, C. P. Hersh, P. Bakke,A. Gulsvik, D. A. Lomas, A. A. Litonjua, S. D. Shapiro, et al. 2011. Poly-morphisms in surfactant protein-D are associated with chronic obstructive pul-monary disease. Am. J. Respir. Cell Mol. Biol. 44: 316–322.
16. Ishii, T., K. Hagiwara, K. Kamio, S. Ikeda, T. Arai, M. N. Mieno, T. Kumasaka,M. Muramatsu, M. Sawabe, A. Gemma, and K. Kida. 2012. Involvement ofsurfactant protein D in emphysema revealed by genetic association study. Eur. J.Hum. Genet. 20: 230–235.
17. Lingappa, J. R., L. Dumitrescu, S. M. Zimmer, R. Lynfield, J. M. McNicholl,N. E. Messonnier, C. G. Whitney, and D. C. Crawford. 2011. Identifying hostgenetic risk factors in the context of public health surveillance for invasivepneumococcal disease. PLoS One 6: e23413.
18. Silveyra, P., and J. Floros. 2012. Genetic variant associations of human SP-A andSP-D with acute and chronic lung injury. Front. Biosci. (Landmark Ed.) 17: 407–429.
19. Aramini, B., C. Kim, S. Diangelo, E. Petersen, D. J. Lederer, L. Shah,H. Robbins, J. Floros, S. M. Arcasoy, J. R. Sonett, and F. D’Ovidio. 2013. Donorsurfactant protein D (SP-D) polymorphisms are associated with lung transplantoutcome. Am. J. Transplant. 13: 2130–2136.
20. Kim, D. K., M. H. Cho, C. P. Hersh, D. A. Lomas, B. E. Miller, X. Kong,P. Bakke, A. Gulsvik, A. Agustı, E. Wouters, et al.; ECLIPSE, ICGN, andCOPDGene Investigators. 2012. Genome-wide association analysis of bloodbiomarkers in chronic obstructive pulmonary disease. Am. J. Respir. Crit. CareMed. 186: 1238–1247.
21. Tanaka, M., Y. Arimura, A. Goto, M. Hosokawa, K. Nagaishi, K. Yamashita,H. Yamamoto, T. Sonoda, M. Nomura, S. Motoya, et al. 2009. Genetic variantsin surfactant, pulmonary-associated protein D (SFTPD) and Japanese suscepti-bility to ulcerative colitis. Inflamm. Bowel Dis. 15: 918–925.
22. Sorensen, G. L., J. Madsen, K. Kejling, I. Tornoe, O. Nielsen, P. Townsend,F. Poulain, C. H. Nielsen, K. B. M. Reid, S. Hawgood, et al. 2006. Surfactantprotein D is proatherogenic in mice. Am. J. Physiol. Heart Circ. Physiol. 290:H2286–H2294.
23. Berg, K. K., H. O. Madsen, P. Garred, R. Wiseth, S. Gunnes, and V. Videm.2009. The additive contribution from inflammatory genetic markers on the se-verity of cardiovascular disease. Scand. J. Immunol. 69: 36–42.
24. Gardai, S. J., Y.-Q. Xiao, M. Dickinson, J. A. Nick, D. R. Voelker, K. E. Greene,and P. M. Henson. 2003. By binding SIRPalpha or calreticulin/CD91, lungcollectins act as dual function surveillance molecules to suppress or enhanceinflammation. Cell 115: 13–23.
25. Barrow, A. D., N. Raynal, T. L. Andersen, D. A. Slatter, D. Bihan, N. Pugh,M. Cella, T. Kim, J. Rho, T. Negishi-Koga, et al. 2011. OSCAR is a collagenreceptor that costimulates osteoclastogenesis in DAP12-deficient humans andmice. J. Clin. Invest. 121: 3505–3516.
26. Merck, E., C. Gaillard, M. Scuiller, P. Scapini, M. A. Cassatella, G. Trinchieri,and E. E. M. Bates. 2006. Ligation of the FcR gamma chain-associated humanosteoclast-associated receptor enhances the proinflammatory responses of humanmonocytes and neutrophils. J. Immunol. 176: 3149–3156.
27. Auffray, C., M. H. Sieweke, and F. Geissmann. 2009. Blood monocytes: de-velopment, heterogeneity, and relationship with dendritic cells. Annu. Rev.Immunol. 27: 669–692.
28. Bouchon, A., F. Facchetti, M. A. Weigand, and M. Colonna. 2001. TREM-1amplifies inflammation and is a crucial mediator of septic shock. Nature 410:1103–1107.
29. Brown-Augsburger, P., K. Hartshorn, D. Chang, K. Rust, C. Fliszar,H. G. Welgus, and E. C. Crouch. 1996. Site-directed mutagenesis of Cys-15 andCys-20 of pulmonary surfactant protein D: expression of a trimeric protein withaltered anti-viral properties. J. Biol. Chem. 271: 13724–13730.
30. Kohler, G., and C. Milstein. 1975. Continuous cultures of fused cells secretingantibody of predefined specificity. Nature 256: 495–497.
31. Byers, D. E., J. Alexander-Brett, A. C. Patel, E. Agapov, G. Dang-Vu, X. Jin,K. Wu, Y. You, Y. Alevy, J.-P. Girard, et al. 2013. Long-term IL-33-producingepithelial progenitor cells in chronic obstructive lung disease. J. Clin. Invest.123: 3967–3982.
32. Leth-Larsen, R., C. Nordenbaek, I. Tornoe, V. Moeller, A. Schlosser, C. Koch,B. Teisner, P. Junker, and U. Holmskov. 2003. Surfactant protein D (SP-D) serumlevels in patients with community-acquired pneumonia. Clin. Immunol. 108: 29–37.
33. Jiang, L., and A. N. Barclay. 2009. New assay to detect low-affinity interactionsand characterization of leukocyte receptors for collagen including leukocyte-associated Ig-like receptor-1 (LAIR-1). Eur. J. Immunol. 39: 1167–1175.
34. Meyaard, L. 2010. LAIR and collagens in immune regulation. Immunol. Lett.128: 26–28.
35. Olde Nordkamp, M. J. M., M. van Eijk, R. T. Urbanus, L. Bont, H. P. Haagsman,and L. Meyaard. 2014. Leukocyte-associated Ig-like receptor-1 is a novel in-hibitory receptor for surfactant protein D. J. Leukoc. Biol. 96: 105–111.
36. Lin, P. M., and J. R. Wright. 2006. Surfactant protein A binds to IgG and en-hances phagocytosis of IgG-opsonized erythrocytes. Am. J. Physiol. Lung Cell.Mol. Physiol. 291: L1199–L1206.
37. Arase, H., E. S. Mocarski, A. E. Campbell, A. B. Hill, and L. L. Lanier. 2002.Direct recognition of cytomegalovirus by activating and inhibitory NK cellreceptors. Science 296: 1323–1326.
38. Srivastava, M., S. Jung, J. Wilhelm, L. Fink, F. B€uhling, T. Welte, R. M. Bohle,W. Seeger, J. Lohmeyer, and U. A. Maus. 2005. The inflammatory versus con-stitutive trafficking of mononuclear phagocytes into the alveolar space of mice isassociated with drastic changes in their gene expression profiles. J. Immunol.175: 1884–1893.
39. Voorhout, W. F., T. Veenendaal, Y. Kuroki, Y. Ogasawara, L. M. van Golde, andH. J. Geuze. 1992. Immunocytochemical localization of surfactant protein D(SP-D) in type II cells, Clara cells, and alveolar macrophages of rat lung.J. Histochem. Cytochem. 40: 1589–1597.
40. Ingersoll, M. A., R. Spanbroek, C. Lottaz, E. L. Gautier, M. Frankenberger,R. Hoffmann, R. Lang, M. Haniffa, M. Collin, F. Tacke, et al. 2010. Comparisonof gene expression profiles between human and mouse monocyte subsets. Blood115: e10–e19.
41. Snyder, G. D., R. E. Oberley-Deegan, K. L. Goss, S. A. Romig-Martin,L. L. Stoll, J. M. Snyder, and N. L. Weintraub. 2008. Surfactant protein D isexpressed and modulates inflammatory responses in human coronary arterysmooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 294: H2053–H2059.
42. Sano, H., H. Chiba, D. Iwaki, H. Sohma, D. R. Voelker, and Y. Kuroki. 2000.Surfactant proteins A and D bind CD14 by different mechanisms. J. Biol. Chem.275: 22442–22451.
The Journal of Immunology 9
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

43. Yamada, C., H. Sano, T. Shimizu, H. Mitsuzawa, C. Nishitani, T. Himi, andY. Kuroki. 2006. Surfactant protein A directly interacts with TLR4 and MD-2and regulates inflammatory cellular response: importance of supratrimericoligomerization. J. Biol. Chem. 281: 21771–21780.
44. Clark, H., N. Palaniyar, P. Strong, J. Edmondson, S. Hawgood, and K. B. M. Reid.2002. Surfactant protein D reduces alveolar macrophage apoptosis in vivo.J. Immunol. 169: 2892–2899.
45. Kim, N., M. Takami, J. Rho, R. Josien, and Y. Choi. 2002. A novel member ofthe leukocyte receptor complex regulates osteoclast differentiation. J. Exp. Med.195: 201–209.
46. Jakubzick, C., E. L. Gautier, S. L. Gibbings, D. K. Sojka, A. Schlitzer,T. E. Johnson, S. Ivanov, Q. Duan, S. Bala, T. Condon, et al. 2013. Minimaldifferentiation of classical monocytes as they survey steady-state tissues andtransport antigen to lymph nodes. Immunity 39: 599–610.
10 OSCAR IS A NOVEL MONOCYTE SP-D RECEPTOR
at Washington U
niversity School of Medicine L
ibrary on March 5, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from
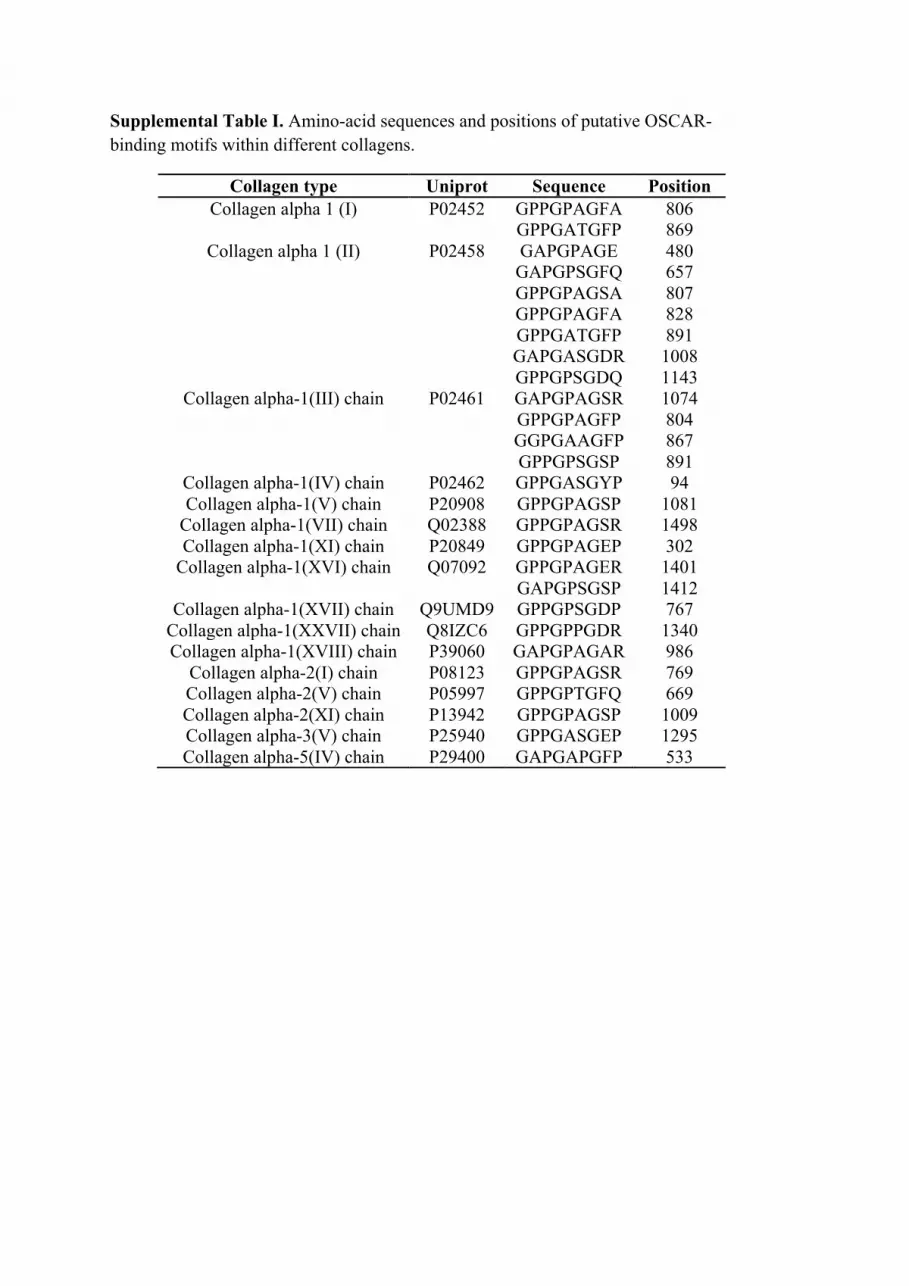
Supplemental Table I. Amino-acid sequences and positions of putative OSCAR-binding motifs within different collagens.
Collagen type Uniprot Sequence Position Collagen alpha 1 (I) P02452 GPPGPAGFA
GPPGATGFP 806 869
Collagen alpha 1 (II) P02458 GAPGPAGE GAPGPSGFQ GPPGPAGSA GPPGPAGFA GPPGATGFP GAPGASGDR GPPGPSGDQ
480 657 807 828 891 1008 1143
Collagen alpha-1(III) chain P02461 GAPGPAGSR GPPGPAGFP GGPGAAGFP GPPGPSGSP
1074 804 867 891
Collagen alpha-1(IV) chain P02462 GPPGASGYP 94 Collagen alpha-1(V) chain P20908 GPPGPAGSP 1081
Collagen alpha-1(VII) chain Q02388 GPPGPAGSR 1498 Collagen alpha-1(XI) chain P20849 GPPGPAGEP 302
Collagen alpha-1(XVI) chain Q07092 GPPGPAGER GAPGPSGSP
1401 1412
Collagen alpha-1(XVII) chain Q9UMD9 GPPGPSGDP 767 Collagen alpha-1(XXVII) chain Q8IZC6 GPPGPPGDR 1340 Collagen alpha-1(XVIII) chain P39060 GAPGPAGAR 986
Collagen alpha-2(I) chain P08123 GPPGPAGSR 769 Collagen alpha-2(V) chain P05997 GPPGPTGFQ 669 Collagen alpha-2(XI) chain P13942 GPPGPAGSP 1009 Collagen alpha-3(V) chain P25940 GPPGASGEP 1295 Collagen alpha-5(IV) chain P29400 GAPGAPGFP 533
SP-D mllfllsalvlltqplgyleAEMKTYSHRTMPSACTLVMCSSVESGLPGR OSCAR -------------------------------------------------- SP-D DGRDGREGPRGEKGDPGLPGAAGQAGMPGQAGPVGPKGDNGSVGEPGPKG OSCAR -------------------------------------------------- SP-D DTGPSGPPGPPGVPGPAGREGPLGKQGNIGPQGKPGPKGEAGPKGEVGAP OSCAR -----GPPGPAGFP------------------------------------
SP-D GMQGSAGARGLAGPKGERGVPGERGVPGNTGAAGSAGAMGPQGSPGARGP OSCAR -------------------------------------------------- SP-D PGLKGDKGIPGDKGAKGESGLPDVASLRQQVEALQGQVQHLQAAFSQYKK OSCAR -------------------------------------------------- SP-D VELFPNGQSVGEKIFKTAGFVKPFTEAQLLCTQAGGQLASPRSAAENAAL OSCAR -------------------------------------------------- SP-D QQLVVAKNEAAFLSMTDSKTEGKFTYPTGESLVYSNWAPGEPNDDGGSED OSCAR -------------------------------------------------- SP-D CVEIFTNGKWNDRACGEKRLVVCEF OSCAR -------------------------
*****.*.*
Supplemental Figure 1. Molecular characteristics and predicted OSCAR-binding sites of SP-D. (A) Amino acid alignment of human SP-D (SP-D) with the minimum OSCAR-binding motif (OSCAR) (31). Direct amino acid sequence identify (*); signal peptide, lower case; Cys 15 and 20 involved in the formation of the disulphide bonds required for the stable assembly of SP-D dodecamers and higher order multimers, underlined; alternating exons, black/blue; exon boundaries, red. (B) Alignment of the 12 amino acid residue (res) OSCAR-binding search (Srch) sequence (seq) reveals an extended putative OSCAR-binding motif.
A
SP-D 106 (12 res) - GPPGPPGVPGPA"Srch seq (12 res) - GXPGPXGFXGXP"
" " "*******.***. "
B
0 200 400 600 800 1000FSC-H
0
200
400
600
800
1000
SSC-H
25.2
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100%
of M
ax
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
1 10 100 1000 10000
0
20
40
60
80
100
% o
f Max
11.20.25 11.1CN5
11.20.02 11.20.06
11.20.08
A
B
11.20.25 11.1CN5
11.20.02 11.20.06
11.20.08
Supplemental Figure 2. Generation of novel anti-human OSCAR mAbs. Histogram plots for immunostainings with anti-human OSCAR mAb clones: 11.20.02, 11.20.06, 11.20.08, 11.20.25 or 11.1CN5 (32) for either: (A) OSCAR-CD3ζ NFAT-GFP reporter cells (open histograms) compared to parental 2B4 NFAT-GFP reporters cells (gray histograms) or (B) human blood monocytes (Gate R1, open histograms) compared to IgG1 (gray histograms). Representative immunostaining from one of two donors is shown. All blood monocytes in gate R1 expressed CD14 and all other cells outside gate R1 (e.g. low FSC/SSC lymphocytes) did not express OSCAR (Data not shown).
R1

Supplemental Figure 3. Screening for OSCAR blocking mAbs. (A) OSCAR-Fc binding to collagen I in the presence of tissue culture supernatants from different anti-human OSCAR hybridomas. Percentage (%) GFP expression from OSCAR-CD3ζ NFAT-GFP reporter cells cultured on tissue culture plates coated with: (B) collagen I; recombinant (C) rat or (D) human SP-D in the presence of different concentrations of purified mAbs: 11.20.02, 11.20.06, 11.20.08, 11.20.25, 11.1CN5 or IgG1 (0-5µg/ml, x-axis), respectively. (E) OSCAR cell surface expression on monocytes left untreated (blue, 0’) or treated with 2.5µg/ml human SP-D for 5 (green), 15 (orange), 30 (cyan) and 60 (pink) mins, respectively compared to IgG1 isotype control (red).
D C
A B
Sample %
Data.001 71
Data.010 55.9
Data.011 45.5
Data.012 45.2
Data.013 43.6
Data.014 51
100 101 102 103 104
FL1-H: Nowt
0
20
40
60
80
100
% o
f Max
60’ 30’ 15’ 5’ 0’ IgG1 isotype control
OSCAR
Sample %
Data.001 71
Data.010 55.9
Data.011 45.5
Data.012 45.2
Data.013 43.6
Data.014 51
100 101 102 103 104
FL1-H: Nowt
0
20
40
60
80
100
% o
f Max
11.1CN5 mAb
E




















