
Organization, differential expression and methylation of rDNA in artificial Solanum allopolyploids
25
Organization, differential expression and methylation of rDNA in artificial Solanum allopolyploids Nataliya Y. Komarova 1 , Thomas Grabe 1 , Dirk J. Huigen 2 , Vera Hemleben 1, * and Roman A. Volkov 1 1 Department of General Genetics, Center of Plant Molecular Biology (ZMBP), University of Tu ¨bingen, Auf der Morgenstelle 28, 72076 Tu ¨bingen, Germany (*author for correspondence; e-mail vera.hemleben@uni- tuebingen.de); 2 The Graduate School of Experimental Plant Sciences, Department of Plant Breeding, Wageningen Agricultural University, P.O. Box 386, 6700 AJ Wageningen, The Netherlands Received 16 July 2004; accepted in revised form 11 October 2004 Key words: developmental regulation, differential activation/repression, epigenetic silencing, nucleolus, subrepeats, uniparental expression Abstract Uniparental activity of ribosomal RNA genes (rDNA) in interspecific hybrids is known as nucleolar dominance (ND). To see if difference in rDNA intergenic spacers (IGS) might be correlated with ND, we have used artificial Solanum allopolyploids and back-crossed lines. Combining fluorescence in situ hybridization and quantification of the level of the rRNA precursor by real-time PCR, we demonstrated that an expression hierarchy exists: In leaves, roots, and petals of the respective allopolyploids, rDNA of S. lycopersicum (tomato) dominates over rDNA of S. tuberosum (potato), whereas rDNA of S. tuberosum dominates over that of the wild species S. bulbocastanum. Also in a monosomic addition line carrying only one NOR-bearing chromosome of tomato in a potato background the dominance effect was maintained. These results demonstrate that there is possible correlation between transcriptional dominance and number of conservative elements downstream of the transcription start in the Solanum rDNA. In anthers and callus tissues under-dominant rDNA was slightly (S. lycopersicum/S. tuberosum) or strongly (S. tuberosum/ S. bulbocastanum) expressed indicating developmental modulation of ND. In leaves and petals, repression of the respective parental rDNA correlated with cytosine methylation at certain sites conserved in the IGS, whereas activation of under-dominant rDNA in anthers and callus tissues was not accompanied by considerable changes of the methylation pattern. Introduction Interspecific hybridization and polyploidy (i.e. al- lopolyploidization) are widespread in plants (Grant, 1981; Levin, 2002). Genome rearrange- ments and alterations in gene expression were found in allopolyploids (Song et al., 1995; Wendel et al., 1995; Volkov et al., 1999; Lee and Chen, 2001; Osborn et al., 2003; Ridlle and Birchler, 2003). One of the first examples of interaction between parental genomes in hybrids is repre- sented by nucleolar dominance (ND; Honjo and Reeder, 1973). This fascinating epigenetic phe- nomenon was described originally by Navashin (1927, 1934), who observed that in interspecific hybrids of Crepis only chromosomes of one crossing partner carried secondary constrictions at metaphase. This chromosomal region was not lost in hybrids, but could be reactivated to produce normal nucleoli in hybrids with a different crossing partner. A relation between secondary constric- tions at metaphase and nucleoli formation during interphase was demonstrated (Heitz, 1931; McClintock, 1934). McClintock (1934) supposed Plant Molecular Biology 56: 439–463, 2004. Ó 2004 Kluwer Academic Publishers. Printed in the Netherlands. 439
-
Upload
uni-tuebingen -
Category
Documents
-
view
3 -
download
0
Transcript of Organization, differential expression and methylation of rDNA in artificial Solanum allopolyploids

Organization, differential expression and methylation of rDNAin artificial Solanum allopolyploids
Nataliya Y. Komarova1, Thomas Grabe1, Dirk J. Huigen2, Vera Hemleben1,* andRoman A. Volkov11Department of General Genetics, Center of Plant Molecular Biology (ZMBP), University of Tubingen, Aufder Morgenstelle 28, 72076 Tubingen, Germany (*author for correspondence; e-mail [email protected]); 2The Graduate School of Experimental Plant Sciences, Department of Plant Breeding,Wageningen Agricultural University, P.O. Box 386, 6700 AJ Wageningen, The Netherlands
Received 16 July 2004; accepted in revised form 11 October 2004
Key words: developmental regulation, differential activation/repression, epigenetic silencing, nucleolus,subrepeats, uniparental expression
Abstract
Uniparental activity of ribosomal RNA genes (rDNA) in interspecific hybrids is known as nucleolardominance (ND). To see if difference in rDNA intergenic spacers (IGS) might be correlated with ND, wehave used artificial Solanum allopolyploids and back-crossed lines. Combining fluorescence in situhybridization and quantification of the level of the rRNA precursor by real-time PCR, we demonstratedthat an expression hierarchy exists: In leaves, roots, and petals of the respective allopolyploids, rDNA ofS. lycopersicum (tomato) dominates over rDNA of S. tuberosum (potato), whereas rDNA of S. tuberosumdominates over that of the wild species S. bulbocastanum. Also in a monosomic addition line carrying onlyone NOR-bearing chromosome of tomato in a potato background the dominance effect was maintained.These results demonstrate that there is possible correlation between transcriptional dominance and numberof conservative elements downstream of the transcription start in the Solanum rDNA. In anthers and callustissues under-dominant rDNA was slightly (S. lycopersicum/S. tuberosum) or strongly (S. tuberosum/S. bulbocastanum) expressed indicating developmental modulation of ND. In leaves and petals, repressionof the respective parental rDNA correlated with cytosine methylation at certain sites conserved in the IGS,whereas activation of under-dominant rDNA in anthers and callus tissues was not accompanied byconsiderable changes of the methylation pattern.
Introduction
Interspecific hybridization and polyploidy (i.e. al-lopolyploidization) are widespread in plants(Grant, 1981; Levin, 2002). Genome rearrange-ments and alterations in gene expression werefound in allopolyploids (Song et al., 1995; Wendelet al., 1995; Volkov et al., 1999; Lee and Chen,2001; Osborn et al., 2003; Ridlle and Birchler,2003). One of the first examples of interactionbetween parental genomes in hybrids is repre-sented by nucleolar dominance (ND; Honjo and
Reeder, 1973). This fascinating epigenetic phe-nomenon was described originally by Navashin(1927, 1934), who observed that in interspecifichybrids of Crepis only chromosomes of onecrossing partner carried secondary constrictions atmetaphase. This chromosomal region was not lostin hybrids, but could be reactivated to producenormal nucleoli in hybrids with a different crossingpartner. A relation between secondary constric-tions at metaphase and nucleoli formation duringinterphase was demonstrated (Heitz, 1931;McClintock, 1934). McClintock (1934) supposed
Plant Molecular Biology 56: 439–463, 2004.� 2004 Kluwer Academic Publishers. Printed in the Netherlands.
439

that if the nucleolus organizer region (NOR) ofspecies A dominates over NOR of species B, and ifB dominates over C, then NOR of species Ashould dominate over C. The existence of suchhierarchy for Crepis species was later confirmedexperimentally (Wallace and Langbridge, 1971),and ND was demonstrated for many organisms(Reeder, 1985; Pikaard, 2000).
At the molecular level, NOR is composed oftandemly arranged repeated units of nuclear genescoding for 18S, 5.8S, and 25–28S rRNA (ribo-somal RNA genes, rDNA), which in higher plantsis transcribed by RNA polymerase I into a 35SrRNA precursor (pre-rRNA). Respectively, inhybrids demonstrating ND a differential tran-scription/silencing of parental rDNA could beanticipated as it was demonstrated for Xenopus(Honjo and Reeder, 1973). In order to explainND, several hypotheses were proposed. Accordingto the species-specific transcription factorhypothesis (Dover and Flavell, 1984; Reeder,1985), a molecular evolution of the rDNA pro-moter region should be accompanied by co-evo-lution of the respective transcription factors. Thisidea was confirmed by in vitro assays for humanand murine rDNA promoters (Grummt et al.,1982; Mishima et al., 1982). Similar specificity wasalso observed for plants: a tobacco rDNA pro-moter was not recognized in cell-free transcriptionextracts from bean (Fan et al., 1995) and the to-mato promoter remained inactive after transfec-tion into protoplasts of Arabidopsis thaliana(Doelling and Pikaard, 1996). However, thehypothesis cannot explain ND in hybrids of spe-cies which are sufficiently related to be crossable,because in this case the transcriptional machineryof one species can effectively use the promoter ofother species as shown applying protoplast trans-fection and in vitro transcription assays (Saez-Vasquez and Pikaard, 1997; Chen et al., 1998;Frieman et al., 1999).
Investigation of ND in allopolyploids of Triti-cum and related genera revealed that rDNA with alonger intergenic spacer (IGS) containing moresubrepeated elements upstream of transcriptioninitiation site (TIS) dominate over those withshorter IGS containing less upstream subrepeats(Gustafson et al., 1988; Sardana et al., 1993; Hou-chins et al., 1997). These data are in congruencewith the enhancer imbalance hypothesis originallyproposed for Xenopus, where upstream subrepeats
act as transcription enhancers (Busby and Reeder,1983; Moss, 1983; Pikaard and Reeder, 1988; Papeet al., 1989). Assuming that enhancers bind tran-scription factors present in limited amounts, NDcould be due to the competition for the factorsbetween the IGS of the respective parental species(Reeder, 1985; Flavell, 1986).
A correlation between DNA methylation andND was shown for Triticum–Secale (Flavell et al.,1988; Viera et al., 1990b; Sardana et al., 1993;Houchins et al., 1997) allopolyploids. Recently, itwas also demonstrated that in Arabidopsis differ-ential NOR silencing is connected with cytosinemethylation at the promoter of under-dominantrDNA and deacetylation/methylation of histoneH3, which results in chromatin condensation(Lawrence et al., 2004). However, differentialexpression of NORs not always correlate withrDNA methylation as it was shown for Brassica(Chen and Pikaard, 1997a) and barley (Papazovaet al., 2001). Furthermore, in contrast to Triticum,data obtained for allopolyploids of Brassica andArabidopsis (Chen and Pikaard, 1997a, b; Chenet al., 1998) were inconsistent with the enhancerimbalance hypothesis, because rDNA with longerspacers showed no dominance effect.
ND appears to be developmentally regulated:e.g. in natural allotetraploid Brassica napus dif-ferential expression of parental rDNA wasabolished in tissues originating from floral meri-stem (Chen and Pikaard, 1997b), but the exactdevelopmental cues were not identified.
Several cytological observations revealed thatat least in Triticeae chromosomal/genomic rear-rangements can modulate ND (Nicoloff, 1979;Schubert and Kunzel, 1990; Viera et al., 1990a;Neves et al., 1997; Georgiev et al., 2001; for reviewsee Pikaard, 2000; Volkov et al., 2004). Besides, forBrassica allopolyploids demonstrating ND (Chenand Pikaard, 1997b) no difference in transcriptionefficiency of the parental rDNA was found both inprotoplast transfection assays or in in vitro tran-scription systems (Saez-Vasquez and Pikaard,1997; Frieman et al., 1999), i.e. for appropriateregulation the rDNA promoter should be inte-grated into a chromosome.
Thus, comparison of the data obtained bycytological and molecular methods for the twobest studied model groups, e.g. Triticum–Aegilopsand Brassica–Arabidopsis, shows that the molecu-lar mechanisms responsible for ND remain not
440

fully understood. The relative importance of localstructural features of the rDNA, especially ofsubrepeated elements in the IGS, and the role ofthe chromosomal/genomic context remains a sub-ject of debate. In order to get further progress inour understanding of these questions, an approachshould be used, which allows evaluation of func-tional features of rDNA integrated into the chro-mosome. Particularly, somatic hybrids betweenspecies with well defined structural differences ofrDNA represent an attractive model system forfurther investigation of molecular mechanismscontrolling rRNA genes activity. Recently, wehave compared organization of the rDNA forseveral genera of the family Solanaceae (Borisjukand Hemleben, 1993; Borisjuk et al., 1997; Volkovet al., 1999, 2003a) and found that the molecularevolution of rDNA in this group occurred mainlyby amplification/deletion of subrepeats. To testwhether ND is connected with the repeated ele-ments in the IGS, we have selected as a modelsystem (i) synthetic allopolyploids and back-cross(BC) progeny between Solanum lycopersicum(LYC, tomato) and S. tuberosum (TUB, potato)(Jacobsen et al., 1995; Garriga-Caldere et al., 1997,1998) in comparison to (ii) allopolyploids betweenS. tuberosum and the tuber-bearing wild species S.bulbocastanum (BLB) also obtained by protoplastfusion (Schilde-Rentschler et al., 1993; Oberwalderet al., 1997).
Our results demonstrate that in the respectiveallopolyploids, BC lines and in a potato monoso-mic addition line carrying tomato chromosome 2rDNA of LYC dominates over rDNA of TUB inall tissues tested, whereas rDNA of TUB domi-nates over rDNA of BLB in leaves and petals, butnot in anthers and callus tissues. In contrast toTriticum–Aegilops and Brassica, ND correlateswith the number of the repeated elements presentin the IGS downstream but not upstream of TIS.A correlation between CG-methylation/de-meth-ylation and differential rDNA expression ofparental rDNA in the allopolyploids was found.
Materials and methods
Plant material and growth conditions
Allopolyploids of Solanum obtained by protoplastfusion and corresponding BC lines (Schilde-
Rentschler et al., 1993; Jacobsen et al., 1995; Gar-riga-Caldere et al., 1997, 1998; see Table 1) werepropagated in vitro onMSmedium (Murashige andSkoog, 1962). Shoots were transferred to soil andplants were grown in the greenhouse under light/dark conditions of 16/8 h at 24 �C. Callus tissueswere produced from leaves and cultivated in dark-ness at 21 �C using MS medium complementedwith 0.65 mg/l kinetin and 6 mg/l NAA.
Isolation of DNA, DNA blotting and restrictionmapping
Genomic DNA was isolated from different planttissues using DNeasy Plant kit (Qiagen, Valencia,CA). Restriction mapping of genomic rDNA andother standard procedures were carried outaccording to Sambrook et al. (1989). For DNAblot hybridization, RNA probes HP1 and HP2(Figure 1) were prepared; respective fragments ofrDNA were amplified by PCR from rDNA clonespNtm-1 (Y08427; Volkov et al., 1999) and pBER1(AF447407; Volkov et al., 2003a) using Hot-startTaq-polymerase (Qiagen) and sequence-specificprimer pairs (Pr15 + Pr16 and Pr17 + Pr18,respectively; Table 2). The T7 RNA polymerasepromoter fragment was added at the 50 end of theprimers Pr16 and Pr18. PCR products were puri-fied with Qiaquick PCR Purification kit (Qiagen)and used for generation of 32P-UTP labeled RNAprobes using Strip-EZ RNA kit (Ambion, Austin,TX).
The copy number of rDNA repeats in tomato,potato and LT-allopolyploids was determinedapplying dot-blot hybridization with LYC- andTUB-sense probes (Figure 2A; Table 2) specificfor rDNA of the parental species. The probes werelabeled with 32P using T4 polynucleotide kinase(Fermentas AB, Vilnus, Lithuania). For calibra-tion of signal intensity, subsequent dilutions ofPCR products containing the total IGS of LYC orTUB1 were applied on the same membrane. Toestimate the copy number of rDNA repeats in theparental forms (LYC, TUB1, TUB4 and BLB)dot-blot hybridization with 32P-labeled HP1 (Fig-ure 1) was performed. Additionally, to evaluatethe relative amounts of parental rDNA in LT- andTB-allopolyploids, the intensity of parental-spe-cific bands was quantified after DNA blottingand hybridization with HP1 and HP2. Forquantification, respective spots were cut from the
441

membranes and radioactivity was measured usingBeckman liquid scintillation counter LS 1801(Beckman, Orange County, CA). Taking into ac-count the length of PCR products used for cali-bration and the whole length of rDNA repeats indifferent Solanum species, the rDNA copy num-bers were calculated.
RNA isolation and one-step reverse transcription(RT)-PCR
Total cellular RNA was isolated from differentplant tissues using RNeasy kit (Qiagen). RNAquantity was measured using RiboGreen RNAQuantitation reagent (Molecular Probes, Leiden,
Netherlands), and its quality was checked byagarose-formaldehyde gel electrophoresis (Sam-brook et al., 1989). Only RNA without detectabledegradation of 25S rRNA was used. Total RNAwas treated with RNase-free DNase I (EpicentreTechnologies, Madison, WI) and checked for thepresence of genomic DNA using PCR. One-stepRT-PCR was performed with 50 ng of totalRNA per reaction applying one-step RT-PCR kit(Qiagen).
Cloning and sequencing of rDNA
Primers Pr1 + Pr2 (Table 2) deduced from thesequences of the 25S and 18S rRNA coding
Table 1. Plant material.
Species/hybrids Abbreviation Plant
accession no.
Ploidy level,
x = 12
Genome
composition
Source References
S. lycopersicum L.
(syn. Lycopersicon
esculentum Mill. ),
tomato.
LYC C31 2n = 2x 2LYC CPBR Jacobsen et al. (1995)
S. tuberosum L.,
potato
TUB1 87.1017-5 2n = 2x 2TUB CPBR Jacobsen et al. (1995)
TUB2 AM66-42 2n = 4x 4TUB CPBR Jacobsen et al. (1995)
TUB3 6704-1 2n = 4x 4TUB CPBR Garriga-Caldere et al.
(1997)
LYC + TUB1,
protoplast fusion
hybrid
LT-F1 C31-17-5 2n = 6x 2LYC + 4TUB CPBR Jacobsen et al. (1995)
LT-F1 · TUB2,
back-cross generation
LT-BC1A 93-6701 2n = 5x)3 1LYC + 4TUB CPBR Jacobsen et al. (1995)
LT-F1 · TUB3,
back-cross generation
LT-BC1B 94-2003 2n = 5x 1LYC + 4TUB CPBR Garriga-Caldere et al.
(1997)
LT-BC1B · TUB3,
monosomic addition
line
LT-BC2 95-2403-6 2n = 4x + 1 LChr2 + 4TUB CPBR Garriga-Caldere et al.
(1998),
D.J. Huigen (unpublished
results)
S. tuberosum L.,
potato
TUB4 B15 (H256/1) 2n = 2x 2TUB BLBP Schilde-Rentschler et al.
(1993)
S. bulbocastanum
Dun.
BLB BGRC N 08006 2n = 2x 2BLB GDC Schilde-Rentschler et al.
(1993)
BLB + TBR4,
protoplast fusion
hybrid
TB-1 Tu 7 2n = 4x 2TUB + 2BLB IPB Schilde-Rentschler et al.
(1993)
TB-2 Tu12 2n = 4x 2TUB + 2BLB IPB Schilde-Rentschler et al.
(1993)
CPBR, Center for Plant Breeding and Reproduction Research CPRO, Wageningen, The Netherlands; BLBP, Bayrische Landesanstaltfur Bodenkultur und Pflanzenbau, Freising, Germany; GDC, the German–Dutch Curatorium for Plant Genetic Resources,Braunschweig, Germany; IPB, Institute of Plant Biochemistry, University of Tubingen.
442

regions of tomato (Kiss et al., 1989a,b) and potato(Borisjuk et al., 1994) were used for PCR ampli-fication of the complete IGS of parental species(Table 1). For cloning, NotI restriction sites wereadded at the 50 ends of both primers. PCRamplification was performed with Pfu Turbo Hot-start DNA Polymerase (Stratagene, La Jolla, CA)applying the following program: initial DNAdenaturation at 95 �C, 2 min; 10 cycles at 95 �C,30 s; 65 �C, 1 min; 72 �C, 3.5 min; 20 cycles at95 �C, 30 s; 65 �C, 1 min; 72 �C, 3.5 min + 20 s/cycle; final extension 72 �C, 10 min. ObtainedPCR products were cloned and sequenced as de-scribed (Volkov et al., 2003a). Clones containingthe complete IGS of BLB and TUB4 previ-ously obtained and sequenced for the 50 externaltranscribed spacer (50 ETS) region (Volkov et al.,2003a), were additionally sequenced here forthe region upstream of TIS. The sequencesappeared in the GenBank/EMBL database underthe accession numbers AF464857–AF464860,AF464863–AF464866 and AY366528–AY366531.
The 30 end fragment of 25S rRNA codingregion was amplified with primers Pr19 + Pr20(Table 2) using genomic DNA and Hot-start Taq-polymerase (Qiagen) or total RNA from leavesand one-step RT-PCR kit (Qiagen). ObtainedPCR products were blunt-end cloned usingpGEM-T Vector System I (Promega, Madison,WI) and sequenced (accession numbers AY624484–AY624553).
Real-time PCR
For real-time RT-PCR quantification, a two-stepprotocol was applied as described recently(Volkov et al., 2003b) with minor modifications.Pre-rRNA transcripts were selectively convertedinto cDNA using 50 ng of total RNA and RNaseH-minus reverse transcriptase from ThermoScriptRT PCR System (Invitrogen, Carlsbad, CA) incombination with the primer Pr5 (Table 2), whichanneals to the highly conserved sequence at theborder of 50 ETS and 18S rRNA coding region(Figure 3A). The amount of pre-rRNA/cDNAdouble-stranded products obtained after reversetranscription was measured using PicoGreendsDNA Quantitation reagent (Molecular Probes).This method of template quantification improved
the reproducibility of data of subsequent real-time PCR.
The real-time PCR was performed in 50 ll ofreaction mixture composed of cDNA and mastermix (final concentrations: 1 u of Platinum Hot-Start Taq Polymerase [Invitrogen], 50 mM KCl,4 mM MgCl2, 20 mM Tris, pH 8.4, 400 lM eachdNTPs [Sigma, St. Louis, MO], 0.5 lM genespecific primers) using an iCycler iQ system (Bio-Rad, Hercules, CA). Amplification of PCRproducts was monitored via intercalation ofSYBR-Green (Molecular Probes; 1:250,000 dilu-tion of a 10,000· stock solution) using the fol-lowing program: initial polymerase activation:95 �C, 10 min; then 40 cycles at 94 �C, 20 s;67 �C, 55 s; 72 �C, 15 s.
Efficiency and specificity of real-time PCRquantification of pre-rRNA transcripts
Species-specific primer pairs for real-time PCR(Pr6 to Pr14; Table 2, Figure 3A) were designedusing Primer 3 Software (http://www.genome.wi.-mit.edu/cgi-bin/ primer/primer3.cgi) and experi-mentally checked for efficiency and specificity.Firstly, the whole IGS of parental species wasamplified by PCR and obtained products wereused as templates in further tests. Amplificationefficiency was validated by comparison of theo-retically expected and experimentally determinedthreshold values in dilution series of the wholeIGS PCR products using 10, 1, 0.1 or 0.01 ng perreaction. Accordingly, PCR conditions wereoptimized for high amplification efficiency(>95%) for all primer pairs used. To provespecificity of PCR, species-specific primers weretested in combinations with homologous (the samespecies) or heterologous (other species) templates.No amplification was observed in heterologouscombinations. The identity of the PCR productswas additionally confirmed by direct DNAsequencing.
In real-time PCR experiments, two pairs ofprimers specific for rDNA of the parentalspecies (Pr9 + Pr10 and Pr9 + Pr11 for LYC;Pr6 + Pr7 and Pr6 + Pr8 for TUB; Pr12 +Pr13 and Pr12 + Pr14 for BLB) were applied inparallel (see Figure 3A). Two concentrations ofcDNA (1 and 0.1 ng) were routinely measured,and duplicate samples were run for each con-centration. All experiments were repeated at least
443

two times for cDNA prepared for two batches ofplants. Using standardized conditions, deviationsof threshold values were less than 1.0 cycle forindependent cDNA preparations and less than0.5 cycle for replicates of the same cDNA. Rel-ative abundance of parental-specific pre-rRNAwas presented in % of total content of pre-RNAin the allopolyploids. The same results wereobtained with the two different primers pairs (seeabove). Changes in the relative concentrations ofPCR products/steady-state RNA levels werechecked for statistical significance according tot-test (Engel, 1997).
Methylation assay
In order to compare the level of cytosine meth-ylation in the rDNA of parental species and inthe corresponding allopolyploids, CG-methyla-tion-sensitive restriction endonucleases wereused. Firstly, genomic DNA was treated withCG-methylation-insensitive restriction endonuc-leases EcoRI + VspI, or EcoRI + SspI, whichcompletely cut rDNA producing species-specificpatterns of restriction fragments (Figures 5A
and 6A). It was reported (McClelland et al.,1994) that the digestion at EcoRI recognition site(GAATTC) can be blocked by multiple over-lapping CG-methylation, i.e. only in the contextm5CGAATTm5CG. However, later experimentsperformed by the supplier of the enzyme(Fermentas AB) used in our experiments havenot confirmed this observation, demonstratingthat even this sequence can be cut. In any case,EcoRI sites present in the rDNA of Solanum arenot flanked by CG-dinucleotides at both ends. Inaccordance, we never observed incompletedigestion of rDNA by application of EcoRI.
After first digestion genomic DNA was addi-tionally cut with one of two CG-methylation-sen-sitive enzymes, AatII or SalI. According to theinformation of the supplier (Fermentas AB), AatIIcuts only fully unmethylated recognition site, butneither GACGTm5C nor GAm5CGTC; SalI doesnot cut GTm5CGAC but cuts GTCGAm5C. Afterelectrophoretic separation in agarose gels andDNA blotting, membranes were hybridized with32P-labeled RNA probes, HP1 or HP2 (Figures 5and 6) to visualize the IGS region upstream ordownstream of TIS, respectively. To evaluate theintensity of bands observed after exposition of
Table 2. PCR primers and FISH probes.
Name Sequence
Pr1 50CATAGCGGCCGCAGACGACTTAAATACGCGAC30
Pr2 50CATAGCGGCCGCATGGCTTAATCTTTGAGACAA30
Pr3 50GGGIAGAGGTGTTGGAGGCAGA30
Pr4 50CCCAAGGGGGTTGAGAATTTAATCAC30
Pr5 50GCAGGATCAACCAGGTAGCATTC30
Pr6 50GGGCGAGTGGCAAGTTTGATG30
Pr7 50CGCCCAAGGGGGTTGAGAATTTA30
Pr8 50CAACCTAACCACGCAAGCCGA30
Pr9 50GGGCGAGTGGCGAGTTCTAGAG30
Pr10 50GAAGACCAAGGTTCACACAGGGGA30
Pr11 50CCAACCTAACCACACATGCCCAA30
Pr12 50TGTGGAAACGGCAGTGCTTTCG30
Pr13 50TGAAGTTCCTGCACGGGCACC30
Pr14 50GGGTTAGGACGTCGCTGCTCG30
Pr15 50TTCACCAAGTGTTGGATTGTT30
Pr16 50TAATACGACTCACTATAGGGAGGGCTGAATCTCAGTGGA30
Pr17 50GTATCCTGTGCTGCATACCTAAT30
Pr18 50TAATACGACTCACTATAGGGAGGCGTCTACCGAATCCGAGAAT30
LYC-sense probe 50CTAGAGCTCCTGTTACGGCTAACTCTAGG30
LYC anti-sense probe 50CCTAGAGTTAGCCGTAACAGGAGCTCTAG30
TUB-sense probe 50CATTTTTCTCGACCGGGCTAGACGCGTTT30
TUB anti-sense probe 50AAACGCGTCTAGCCCGGTCGAGAAAAATG30
444

membranes with X-ray film, two approaches wereused: (i) autoradiographs were quantified usingBio1D Image Analysis Software (Vilber Lourmat,Marne-La-Valee, France), or (ii) individual spotswere cut out from the membranes and radioac-tivity was measured using Beckman liquid scintil-lation counter LS 1801.
Fluorescence in situ hybridization (FISH)
FISH was modified from Schwarzacher andHeslop-Harrison (2000). Root tips, approximately2 cm in length, were harvested from plants culti-vated in vitro. Metaphase accumulation was per-formed by incubation of the root tips in 2 mMhydroxyquinoline for 2 h at room temperature andthen for 2 h at 4 �C. After fixation in etha-nol:acetic acid (3:1 [v/v]) root tips were stored at)20 �C until further treatment.
Fixed roots were washed in SCB (10 mM so-dium-citrate buffer, pH 4.8), digested in enzymesolution containing 90 u/ml cellulase (Sigma,C1184) and 13.5 u/ml pectinase (Sigma, P4716)dissolved in SCB for 1 h at 37 �C in a humidchamber, and washed in SCB for 15 min. Eachroot tip was cut into small fragments (1–3 mm),and incubated with 20 ll 50% (v/v) acetic acid for3 min. The root tips were covered with cover-slips,squashed, immersed in liquid nitrogen, and thencover-slips were removed.
In order to discriminate between the rDNA andpre-rRNA of LYC and TUB in the allopolyploidgenomes, species-specific sense and anti-sense oli-gonucleotide probes were designed according tothe 50 ETS sequence (Figure 2A; Table 2). Species-specificity of the hybridization probes was checkedby FISH performed with root tips of the parentalspecies applying the combination of both sense oranti-sense probes for rDNA and pre-rRNAdetection, respectively. In the majority of the FISHexperiments, LYC- and TUB-specific probes la-beled with rhodamin (red signal) or fluorescein(green signals), respectively, were used. Addition-ally, in control experiments LYC probes labeledwith fluorescein and TUB probes labeled withrhodamin were tested.
To detect rDNA, total cellular RNA was re-moved from slides by RNase A (100 lg/ml)treatment for 1 h at 37 �C. Proteins were re-moved by pepsin treatment (5 lg/ml) for 15 minat 37 �C. Slides were rinsed in 2· SSC (1· SSC is
0.15 M NaCl and 0.015 M sodium citrate), threetimes for 5 min, fixed in 4% paraformaldehydefor 10 min at room temperature, and then rinsedin 2· SSC two times for 5 min. To denatureDNA, slides were incubated for 2 min at 75 �C inthe solution containing 70% formamide, 2· SSC,dehydrated by submerging in 70%, 90%, and100% (v/v) ice cooled ethanol and air dried. Forhybridization, 30 ll of DNA-detection solution(5· SSPE, 5· Denhardt’s solution, 0.5% SDS,50 ng/ll DNA of E. coli) containing 2 pmol ofboth sense probes was added to each slide, cov-ered with a cover-slip and incubated overnight at37 �C.
Detection of pre-rRNA in interphase nucleiwas performed analogously, but RNAse treatmentand DNA denaturation were omitted. Forhybridization, 30 ll of RNA-detection solution(20% formamide, 600 mM NaCl, 50 mM Tris-HCl, pH 7.5, 4.5 mM Na pyrophosphate, 0.2%polyvinylpirrolidone 40, 0.2% Ficoll 400, 5 mMEDTA, 10% Dextran sulfate) containing 2 pmolof both anti-sense probes was used for one slide.
After hybridization slides were washed with 6·SSC three times for 30 min, then incubated for2 min at 63 �C, and finally washed with 4· SSC,0.2% Tween 20 for 5 min. To eliminate unspecificbackground, slides were additionally incubatedwith 5% BSA for 30 min at 37 �C and washedwith 4�SSC, 0.2% Tween 20 for 5 min at roomtemperature.
Chromosomes were stained with 100 ll DAPI(1 lg/ml) per slide for 10 min, and the prepara-tions were analyzed with a Zeiss epifluorescencemicroscope (Oberkochen, Germany) with theappropriate filters and photographed on KodakElite Chrom 400 ASA film. Color figures andoverlays were prepared from digitized images ofthe film negatives by using Adobe PhotoShopsoftware (Mountain View, CA).
Results
Organization of rDNA intergenic spacerin Solanum spec.
In order to characterize the organization of rDNAof the parental species and to design species-spe-cific probes for detection of the parental rDNAand the respective transcription products in the
445

allopolyploids studied (Table 1), restriction map-ping of genomic DNA and cloning/sequencing ofthe IGS region (Figure 1) was performed.
In the genome of LYC, line C31 (the tomatoparent of the LYC + TUB somatic allopolyploidLT-F1 and the back-cross line LT-BC1A; seeTable 1), only one IGS length class was found,which contains two arrays of repeated elementsupstream and downstream of TIS (Figure 1). TwoIGS clones of LYC, completely sequenced here,were identical in length (3258 bp) and sequence(99.9% similarity). A 86-bp motif present as aunique sequence in the IGS of LYC downstreamof the 25S rDNA appears to be amplified in theIGS of the other species studied (see below) pro-ducing type I subrepeats. In addition to the 54-bpsubrepeats (type IIa) upstream of TIS already de-scribed for the IGS of LYC (Perry and Palukaitis,1990), we have detected two copies of type IIbsubrepeats, which represent a duplication of thelast 20 bp of type IIa subrepeats already describedfor the IGS of potato (Borisjuk and Hemleben,1993). Compared with potato, the IGS of LYCpossesses less subrepeats upstream of TIS (Fig-ure 1), but this length difference is compensated bya six times repeated 141-bp element (type III)present downstream of TIS (Borisjuk and Hemle-ben, 1993). Each of the type III subrepeats con-tains a 40-bp conservative element (CE), which ishighly similar to those of the distantly relatedNicotiana (Borisjuk et al., 1997) and Atropa (I.I.Panchuk, N. Borisjuk, V. Hemleben, and R.A.Volkov, unpublished results) species.
Four lines of potato, which were used for theproduction of the allopolyploids and back-crossedlines, were characterized. One major IGS lengthvariant (3.0 kb) was detected in the rDNA ofTUB1 (line 87.1017/5, the potato partner of theLT-F1 line; see Table 1). In addition, minor IGSvariants were found, which show some length andsequence heterogeneity upstream of TIS, e.g. aSspI recognition site was detected only in the IGSof approximately 10–15% of rDNA repeats ofTUB1 (see Figure 1). Several IGS clones of TUB1were isolated, two of which were completely se-quenced (Figure 1). They appeared to be identicalin length and sequence (99.8% similarity). TheIGS of potato lines AM66-42 (TUB2, the crossingpartner for LT-F1 to obtain LT-BC1A) and 6704(TUB3, the crossing partner for LT-F1 to obtainLT-BC1B and LT-BC2; Table 1), were character-
ized only by restriction mapping; one length vari-ant (3.2 kb) was found in both lines. For thepotato dihaploid breeding line H256/1 (TUB4),which was used to produce TUB + BLB somatichybrids (TB-1 and TB-2; Table 1), two IGS vari-ants (3.1 and 3.3 kb) were detected (Figure 1).Interestingly, the 3.1-kb variant lacks the EcoRIsite present in the 50 ETS of the other Solanumspecies. Two clones representing both IGS variantsof TUB4 were completely sequenced (Figure 1),and a comparison showed that the IGS clones ofTUB1 and TUB4 are very similar in the 50 ETS(97.5–99.8% similarity) and differ mainly innumber and alteration of upstream subrepeats.The 141-bp type III element, which is six timesrepeated in the IGS of LYC, occurs only once inpotato (TUB1 and TUB4); in addition, it containsinternal duplications around CE (Figure 1), and asecond copy of CE (CE2) is present in the 50 ETS,as we have recently demonstrated for the majorityof South American tuber-bearing potatoes(Volkov et al., 2003a).
In the genome of BLB, the fusion partner forTB-1 and TB-2 allopolyploids (Table 1), two IGSvariants (3.0 and 3.4 kb) were detected by restric-tion mapping. Two IGS clones sequenced representboth IGS variants; they demonstrate a high se-quence similarity (97%) and differ mainly in thenumber of the subrepeated elements upstream ofTIS. The overall structural organization of IGSof BLB is analogous to TUB (Figure 1). Subre-peats of the same structural types I, IIa, and IIb,and an unique AT-rich region are present up-stream of TIS. In contrast to LYC, the 141-bptype III element containing CE occurs only oncein the 50 ETS of BLB, and, in contrast to TUB,no duplication of CE is present. Additionally,three to four IGS clones were partially sequencedfor each of the species demonstrating more than99% sequence similarity to the respective com-pletely sequenced clones.
Comparison of the 50 ETS sequences showedthat different regions evolve with different rates(Volkov et al., 2003a). Two major parts can bedistinguished: a variable region (VR), differing inlength, and a conservative region, approximately560 bp long (CR, see Figure 1). In this conserva-tive region, the level of similarity among the spe-cies studied ranges from 87% to 92%, i.e. even inthe comparatively slowly evolving region sufficientdifferences are present, which can be used to
446

design species-specific PCR primers and probes forfluorescent in situ hybridization (FISH).
Presence of parental rDNA in Solanumallopolyploids and back-cross lines
The parental rDNA in the allopolyploids wasmonitored either by FISH, and/or by DNA blothybridization, and by PCR using genomic DNA astemplate.
In order to discriminate LYC and TUBrDNA, corresponding species-specific probes lo-cated in the 50 ETS were designed for FISH(Figure 2A). Applying a mixture of two sense(RNA-like) probes, four green TUB-specific andtwo red LYC-specific signals were detected inroot tip cells of LT-F1 (Figure 2D), confirmingthat the LT-F1 line originated from the fusion ofone diploid tomato and of two diploid potatoprotoplasts (Jacobsen et al., 1995). In LT-BC1A,obtained by back-crossing of LT-F1 with thetetraploid TUB2, one LYC-specific and fourTUB-specific signals were found using the senseprobes (Figure 2E), confirming the pentaploidconstitution of this line (Jacobsen et al., 1995).Additionally, our results show that an extra-longchromosome observed in the LT-F1 andLT-BC1A (Jacobsen et al., 1995) does not con-tain rDNA sequences.
For discrimination of TUB and BLB rDNA inTB-1 and TB-2, we could not apply FISH, becauseit was not possible to design parental-specifichybridization probes due to high similarity of therespective 50 ETS. However, it was possible to useDNA blot hybridization and PCR.
For DNA blot hybridization, using doubledigestion with EcoRI and SspI, RFLP patternscharacteristic for each of the parental plants(LYC, TUB1-4, BLB) were obtained, and therespective allopolyploids/BC-lines showed thecombination of the parental patterns (not shown).
For PCR, the primers Pr3 and Pr4 were used(Table 2) annealing to the sequence motifs con-served in Solanum rDNA, which allowed ampli-fication of the 50 ETS region downstream of TISof all fusion partners (Figure 3A); the corre-sponding PCR products showed species-specificlengths and could be easily discriminated (Fig-ure 3B). As expected, only one intensive PCRproduct was obtained for LYC (1118 bp) and
for BLB (411 bp). For the 50 ETS of TUB1 andTUB4 two PCR products were obtained: a ma-jor 472-bp product, which is in agreement withrestriction mapping and sequencing data, and aminor 398-bp product, which according to thesequencing data (Volkov et al., 2003a) probablyrepresent a pseudogene. Therefore, the methodsused revealed that the fragment patterns/hybrid-ization signals observed for LT- and TB-allopo-lyploids/BC-lines exhibited a combination of thecorresponding patterns of the parental species,demonstrating the presence of the parentalrDNA arrays in the hybrid genomes. The geno-mic constitution of LT-F1 and LT-BC1A wasanalyzed applying genomic in situ hybridization(GISH) and RFLP (Jacobsen et al., 1995;Garriga-Caldere et al., 1997). Four completechromosome sets of potato and two of tomatoincluding a novel extra-long chromosome werefound in LT-F1. In LT-BC1A nine tomatochromosomes including the extra-long one weredetected by GISH. According to RFLP analysisthe nine chromosomes represented six (1, 3, 6, 8,9, and 10) of the 12 chromosomes of the haploidcomplement of tomato, suggesting that in LT-BC1A some of the tomato chromosomes may bepresent as two copies (Jacobsen et al., 1995).However, only one RFLP marker was used perchromosome. Our data obtained by differentmethods obviously showed that one copy ofchromosome 2 carrying the rDNA (Gebhardtet al., 1991) is also present in the LT-BC1A.
In order to determine the copy number ofrDNA repeats in LYC, TUB1, LT-F1 and LT-BC1A, we carried out dot-blot hybridization withparental-specific probes (Table 2: LYC and TUB,sense). Also, to estimate the copy number ofrDNA repeats in the parental species (LYC,TUB1, TUB4, and BLB) the HP1 probe (see Fig-ure 1) was used for dot-blot hybridization. Addi-tionally, to evaluate the relative amounts ofparental rDNA in the allopolyploids, we havequantified the intensity of parental-specific bandsafter DNA blotting and hybridization. Takingtogether, the results showed (Table 3) that rDNArepeats amount to approximately 7.5% of thegenome of LYC, to 5% in the genomes of TUB1and TUB4, and to 5.5% in BLB, which reasonablycorresponds to the estimations recently performedfor different species and varieties of Solanum(Schweizer et al., 1993).
447

The rDNA repeat copy numbers expected fromthe genome constitution of the LT-F1 and LT-BC1A were mainly in good agreement with theexperimental data (Table 3), demonstrating thatno significant amplification/deletion of parentalrDNA took place in the allopolyploids. No evi-dence for amplification/deletion of parental rDNAwas found in TB-1 and TB-2; this is in agreementwith previous analyses (Stelzer, 1996).
Differential expression of parental rDNA
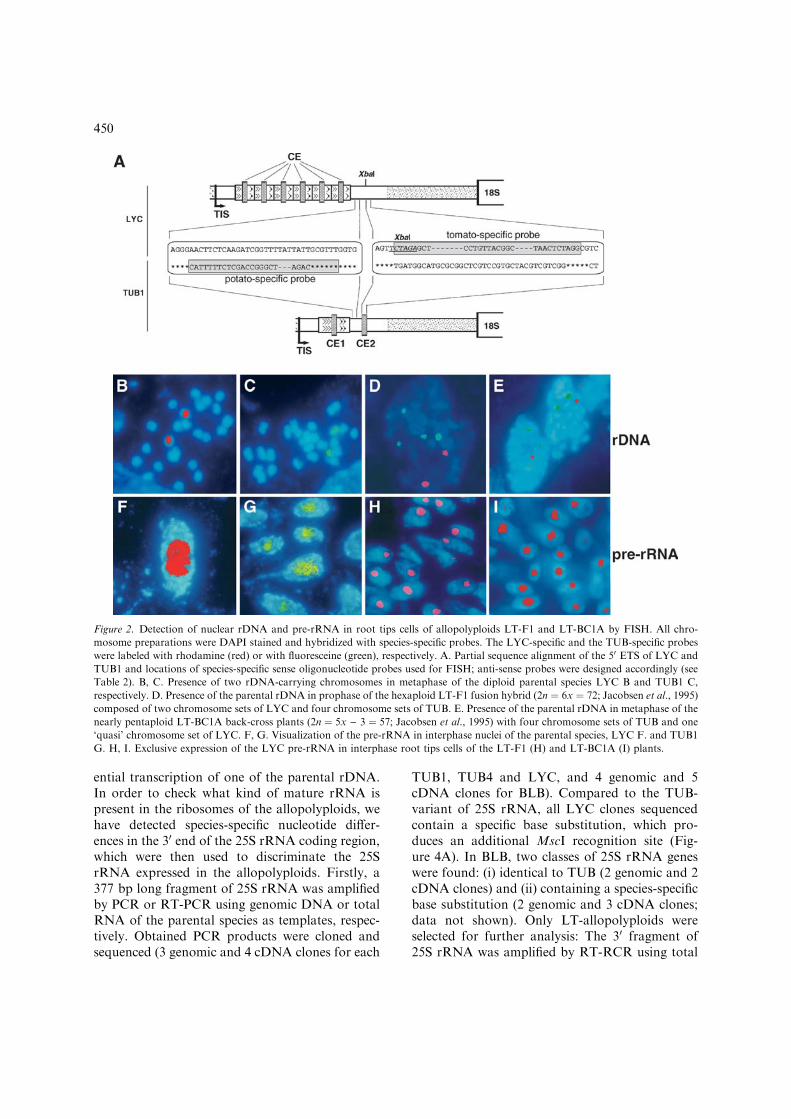
Firstly, FISH studies were applied to character-ize the parental rDNA expression in root tips ofLT-F1 and LT-BC1A. A mixture of anti-senseprobes designed for the VR of the 50 ETS(Figure 2A) and specifically labeled for tomato(red) and potato (green) was used for differentialpainting of parental pre-rRNA in interphasenuclei. Actually, in both, LT-F1 and LT-BC1A,only red (tomato) signals filling the nucleoluswere distinguished, demonstrating preferentialexpression of LYC pre-rRNA in root tip cells(Figures 2H and I).
In the LT-allopolyploids the character of theparental rDNA expression was also tested apply-ing competitive one-step RT-PCR with totalrRNA isolated from leaves as template and prim-ers Pr3 and Pr4 (Figure 3B). Identical sets offragments were amplified for LYC and LT-F1,demonstrating that rDNA of tomato was exclu-sively expressed, whereas potato rDNA appearedto be completely inactive. In leaves of LT-BC1A, aweak TUB-specific PCR product was detected inaddition to the prominent tomato-specific product(Figure 3B), showing at least partial transcrip-tional activity of potato rDNA in the back-crossedplants.
In TB-1 and TB-2 allopolyploids, differentialactivity of parental rDNA was also studiedapplying one-step RT-PCR with the primersPr3 and Pr4. Using total RNA isolated fromleaves, exclusively a TUB-specific product wasamplified (see Figure 3B), demonstrating tran-scriptional dominance of TUB4 rDNA over BLBrDNA.
Interestingly, only one RT-PCR product (472bp) was obtained with total RNA of TUB1 andTUB4 as templates, although two fragments (398and 472 bp) were found using genomic DNA for
PCR (see Figure 3B). This observation agrees withour sequencing data that the shorter minor prod-uct represents transcriptionally silenced rDNArepeats.
Quantification of parental pre-rRNA levelsby real-time RT-PCR
Bearing in mind that in allopolyploids of Brassicaone of the parental rDNA silenced in vegetativetissues can be reactivated in floral organs (Chenand Pikaard, 1997b), we investigated tissue-speci-ficity of ND in Solanum allopolyploids. In order toprecisely evaluate the relative abundance of pre-rRNA transcribed from the parental rDNA indifferent tissues of the hybrids, we applied the real-time RT-PCR quantification using RNA/cDNAprepared from different tissues in combinationwith species-specific primers (Pr6 to Pr14;Figure 3C, Table 2).
In LT-F1 the rDNA of tomato delivered 100%of the total pre-rRNA in leaves and petals, andabout 99% in anthers and callus (Figure 3C). Sim-ilarly in LT-BC1A, the transcripts of LYC rDNArepresent the main fraction of the pre-rRNA pool,whereas the relative amount of the potatopre-rRNA was about 1% in leaves and petals andincreased to approximately 3–5% in callus and an-thers (Figure 3C), indicating a weak de-repressionof potato rDNA. We have also quantified the levelsof parental pre-rRNA in leaves of anotherback-cross line, LT-BC1B, which was indepen-dently obtained from LT-F1 by crossing with adifferent tetraploid breeding line of potato, TUB3(Table 1). In leaves of this line LYC pre-rRNAamounts to 97% similar as in LT-BC1A(Figure 3C).
In addition, we studied a monosomic additionline LT-BC2, which was produced by crossing ofLT-BC1B with TUB3. This line possesses onlyone tomato chromosome 2 carrying the LYCrDNA, and four complete sets of potato chro-mosomes (D.J. Huigen, unpublished results; seeTable 1). Actually, real-time RT-PCR quantifi-cation showed that in leaves of LT-BC2 pre-rRNA of LYC amounts to 95% (Figure 3C),indicating that a single tomato chromosome 2 isstill sufficient to provide preferential expressionof tomato rDNA and genes located at any othertomato chromosomes are obviously not neces-sary.
448

449
- type IIa and IIb subrepeats;
- type III subrepeat;
- type I subrepeat;
- type III subrepeat with internal duplications around CE
0.5 kb
LYC
25S 18S
TIS
CE
18S
TIS
CE 1 CE 2
18S
TIS
CE 1 CE 2
18S
TIS
CE 1 CE 2
18S
TIS
CE
TUB1
TUB4
BLB
Pr 2
Pr 1
VR CR
18S
TIS
CE
EcoRI
EcoRI
EcoRI
EcoRI
EcoRI
EcoRI
EcoRI
SspI
Ssp *I
SspI
SspI
VspI
VspI
VspI
VspI
VspI
VspIVspI
HP1
BamHI
BamHI
BamHI
BamHI
BamHI
BamHI
XbaI
XbaI
XbaI
25S
EcoRI
25S
EcoRI
25S
EcoRI
25S
EcoRI
25S
EcoRI
25S
EcoRI
HP2
Figure 1. Organization of the IGS in the rDNA of Solanum lycopersicum (LYC), S. tuberosum (TUB1 and TUB4), and S. bulbo- castanum (BLB). TIS, transcription initiation site; TTS, transcription termination site; NTS, non-transcribed spacer region; ETS, external transcribed spacer; ITS, internal transcribed spacer; VR and CR, variable and conservative regions in the 5´ ETS; CE, conserved element. Position of restriction endonucleases recognition sites (BamHI, EcoRI, SspI, VspI, XbaI) and hybridization probes 1 and 2 (HP1, HP2), used for mapping of genomic rDNA, and location of primers Pr1 and Pr2, applied for PCR amplification of the IGS are indicated. SspI restriction site present only in the minor fraction of rDNA of TUB1 is marked by an asterisk.
In TB-1 and TB-2, the rDNA of TUB4 deliv- ered up to 94–98% of the total pre-rRNA in leaves and petals confirming dominance of TUB over BLB. Remarkably, approximately equal amounts of both parental pre-rRNA were found in anthers and callus (Figure 3C), demonstrating de-repres-sion of under-dominant rDNA of BLB.
Evaluation of parental-specific 25S rRNA in allopolyploids The different abundance of parental pre-RNA found in allopolyploids could reflect a differential processing possibly connected with different length of the 5´ ETS (see Figure 1), but not the prefer-
ential transcription of one of the parental rDNA.In order to check what kind of mature rRNA ispresent in the ribosomes of the allopolyploids, wehave detected species-specific nucleotide differ-ences in the 30 end of the 25S rRNA coding region,which were then used to discriminate the 25SrRNA expressed in the allopolyploids. Firstly, a377 bp long fragment of 25S rRNA was amplifiedby PCR or RT-PCR using genomic DNA or totalRNA of the parental species as templates, respec-tively. Obtained PCR products were cloned andsequenced (3 genomic and 4 cDNA clones for each
TUB1, TUB4 and LYC, and 4 genomic and 5cDNA clones for BLB). Compared to the TUB-variant of 25S rRNA, all LYC clones sequencedcontain a specific base substitution, which pro-duces an additional MscI recognition site (Fig-ure 4A). In BLB, two classes of 25S rRNA geneswere found: (i) identical to TUB (2 genomic and 2cDNA clones) and (ii) containing a species-specificbase substitution (2 genomic and 3 cDNA clones;data not shown). Only LT-allopolyploids wereselected for further analysis: The 30 fragment of25S rRNA was amplified by RT-RCR using total
Figure 2. Detection of nuclear rDNA and pre-rRNA in root tips cells of allopolyploids LT-F1 and LT-BC1A by FISH. All chro-
mosome preparations were DAPI stained and hybridized with species-specific probes. The LYC-specific and the TUB-specific probes
were labeled with rhodamine (red) or with fluoresceine (green), respectively. A. Partial sequence alignment of the 50 ETS of LYC and
TUB1 and locations of species-specific sense oligonucleotide probes used for FISH; anti-sense probes were designed accordingly (see
Table 2). B, C. Presence of two rDNA-carrying chromosomes in metaphase of the diploid parental species LYC B and TUB1 C,
respectively. D. Presence of the parental rDNA in prophase of the hexaploid LT-F1 fusion hybrid (2n ¼ 6x ¼ 72; Jacobsen et al., 1995)
composed of two chromosome sets of LYC and four chromosome sets of TUB. E. Presence of the parental rDNA in metaphase of the
nearly pentaploid LT-BC1A back-cross plants (2n ¼ 5x ) 3 ¼ 57; Jacobsen et al., 1995) with four chromosome sets of TUB and one
‘quasi’ chromosome set of LYC. F, G. Visualization of the pre-rRNA in interphase nuclei of the parental species, LYC F. and TUB1
G. H, I. Exclusive expression of the LYC pre-rRNA in interphase root tips cells of the LT-F1 (H) and LT-BC1A (I) plants.
450

Figure 3. Differential expression of parental rDNA in different tissues of allopolyploids of Solanum. (A) Organization of 50 ETS in
parental species and location of primers used for PCR amplification. (B) Presence and expression of parental rDNA in leaves of the
allopolyploids. Genomic DNA and total RNA were isolated from leaves of parental species and allopolyploids and used as templates
for PCR (DNA) or RT-PCR (RNA) using primers Pr3 + Pr4. Lengths of the PCR products are shown. Lanes M represent the size
marker (100 bp Gene Ruler DNA Ladder Mix; Fermentas AB). (C) Pre-rRNA level for parental rDNA in different tissues of
allopolyploids of Solanum. Total cellular RNA was isolated from different tissues of the parental species and allopolyploids and the 50
ETS part of pre-rRNA was converted into cDNA using reverse transcriptase and primer Pr5. Equal amounts of cDNA were subjected
to real-time PCR quantification with primers specific for rDNA of the parental species. Relative abundance of parental-specific
pre-rRNA is presented in % of total content of pre-RNA in different tissues of allopolyploids. Bars show means ± SD (n ¼ 4–6).
451

RNA as template prepared from leaves and an-thers of LT-F1 and LT-BC1A, respectively, andthen the PCR products were cloned. For each ofthese RNAs, 10 clones were randomly selected andsequenced. Among these 40 clones only two clonesobtained from anthers of LT-BC1A were identifiedas TUB-specific, whereas the remaining 38 cloneswere LYC-specific. Also, we applied digestion ofRT-PCR products with MscI for discrimination ofparental 25S rRNA expressed in LT-allopolyp-loids (Figure 4B): Prominently, LYC-specificproducts were obtained; a weak TUB-specificproduct was found for anthers of LT-BC1A andleaves of LT-BC1B and LT-BC2, agreeing wellwith the real-time RT-PCR quantification of pre-RNA levels (see Figure 3). An additional weakproduct of reduced length (about 300 bp) couldrepresent a LYC-derived non-transcribed rDNA-related pseudogene located at a chromosome dif-ferent from number 2 because this product couldbe observed neither by RT-PCR nor by PCR usinggenomic DNA as template in LT-BC2 line(Figure 4B).
Hence, the dominance of the LYC pre-rRNAobserved in LT-allopolyploids remained unaffectedat the level of 25S rRNA, i.e. no differentialprocessing takes place.
Methylation of rDNA
Methylation of cytosine residues has been dis-cussed to be involved in regulation of rDNAexpression (Flavell et al., 1988; Pikaard and
Reeder, 1988; Torres-Ruiz and Hemleben, 1994).In order to compare the level of cytosine methyl-ation in the rDNA of the parental species andcorresponding allopolyploids, several combina-tions of CG-methylation-sensitive (AatII, SalI)and CG-methylation-insensitive (EcoRI, VspI andSspI) restriction endonucleases, selected accordingto the presence of their recognition sites in therDNA (Figures 5A and 6A), were applied. Firstly,genomic DNA was treated by CG-methylation-insensitive enzymes, which completely digest theDNA. After that the probes were additionallytreated with one of the CG-methylation-sensitiveenzymes, which selectively cut only unmethylatedDNA. In order to selectively visualize restrictionfragments of the IGS regions upstream or down-stream of TIS two different labeled probes, HP1and HP2, were used for DNA blot hybridization(Figures 5A and 6A). Therewith, species-specificpatterns of restriction fragments were obtainedand used for discrimination of methylated andde-methylated parental rDNA.
Autoradiographs showed that the methylatedrDNA fraction provided the most intensive signalsin all species/allopolyploids studied (Figures 5Band 6B); de-methylated rDNA appeared as severaladditional bands of reduced size. Among theparental species the rDNA of tomato is highlymethylated compared to the potato species (TUB,BLB) as confirmed by subsequent quantification ofthe autoradiographs.
To estimate the degree of methylationof parental rDNA in the allopolyploids, we
Table 3. Ribosomal DNA copy numbers in parental diploids and allopolyploids of Solanum.
Parental species Genome size, in pg
per 1C
Content of
rDNA in %
Copy number
of repeated
rDNA units
per 1C
Allopolyploids b rDNA copy number in
allopolyploids/cell
Experimentally
determined a
Used for
calculations
Theoretically
expected
Experimentally
determined
S. lycpopersicum,
LYC
1.0–2.5 1.75 7.5 ± 1.0 13 000 LT-F1 26 000 (LYC)
+ 30 000 (TUB)
26 000 (LYC)
+ 26000 (TUB)
S. tuberosum,
TUB1
0.9–2.1 1.5 5.0 ± 1.0 7500 LT-BC1A 13 000 (LYC)
+ 30 000 (TUB)
16 000 (LYC)
+ 28 000 (TUB)
S. tuberosum,
TUB4
0.9–2.1 1.5 5.0±1.0 7500 TB-1 and TB-2 15 000 (TUB4)
+ 16 400 (BLB)
15 000 (TUB4)
+ 16 000 (BLB)
S. bulbocastanum,
BLB
Nd c 1.5 5.5 ± 1.0 8200
a Bennett and Smith (1976) and Arumuganathan and Earle (1991).b For genome composition see Table 1.c Not determined.
452

compared the relative intensities of species-specificmethylated and respective de-methylated frag-ments, especially, the intensity of the shortestde-methylated restriction products, which canbe easily distinguished on autoradiographs(Figure 5B). In leaves of LT-allopolyploids com-
pared with the parental species the intensity ofTUB-specific de-methylated products appeared tobe markedly weaker, upstream (AatII: 735 bp,SalI: 1105 bp) and downstream (AatII: 609 bp,SalI: 716 bp) of TIS. In contrast, no changes inmethylation patterns of the LYC-rDNA were ob-
Figure 4. Detection of parental-specific mature 25S rRNA in Solanum allopolyploids. (A) Partial sequence alignment of 25S rRNA
coding region of LYC, TUB1, TUB3 and BLB. The 377 bp long 30 fragment of 25S rRNA coding region was amplified by PCR/RT-
PCR, cloned and sequenced. Position of primers (Pr19 and Pr20) used for amplification and localization of LYC-specific MscI
recognition site (underlined) are shown. (B) Presence of 25S rRNA genes of LYC and TUB and their expression in leaves and anthers
of LT-F1, LT-BC1A , LT-BC1B and LT-BC2. The 30 fragment of 25S rRNA coding region was amplified by PCR (DNA) or RT-PCR
(RNA), obtained PCR products were digested with MscI and analyzed in agarose gel. For comparison, non-digested PCR products of
LYC (LYC/nd) and a mixture of digested PCR products of LYC and TUB in molar ratio 10 to 1 (LYC:TUB, 10:1) were loaded on the
same gel.
453

served for the region upstream of TIS. Also, forthe region downstream of TIS intensity of theLYC-specific de-methylated fragments (AatII:501 bp, SalI: 824 bp) remained either unaffected inLT-F1 or only slightly reduced in LT-BC1A, al-though the intensity of the LYC-specific methy-lated fragments (1633 and 1744 bp for the regionsupstream and downstream of TIS, respectively)
decreased because LT-F1 and especially LT-BC1Aplants contain less LYC-rDNA per cell (seeTable 3). Therefore, the data show that in theLT-allopolyploids compared to the parental spe-cies a slight de-methylation of LYC rDNA andsignificant hypermethylation of TUB rDNAoccurred. These observations were confirmed byquantification of the intensity of the respective
Figure 5. Cytosine methylation in rDNA of LYC, TUB1, and LT-allopolyploids. (A) Schematic presentation of the IGS region of
LYC and TUB1; the positions of restriction endonucleases recognition sites for EcoRI (E), VspI (V), AatII (A) and SalI (S) as
predicted from sequences of the IGS clones and locations of hybridization probes (HP1 and HP2) used for rDNA mapping are shown.
(B) rDNA methylation assay: Genomic DNA isolated from different tissues of parental species and allopolyploids were firstly double-
digested with CG-methylation-insensitive EcoRI and VspI, and after that treated with an excess of CG-methylation-sensitive AatII or
SalI (upper and lower panels, respectively). DNA fragments obtained were separated by agarose gel electrophoresis followed by DNA
blotting and hybridization with probe HP1 or HP2, respectively. Arrows indicate unmethylated products, demonstrating obvious
changes in intensity.
454
bands (not shown). Only minimal variations in themethylation pattern were found between differenttissues.
To evaluate the methylation degree of theparental rDNA in the TB-allopolyploids, we ap-plied digestion of genomic DNA with EcoRI and
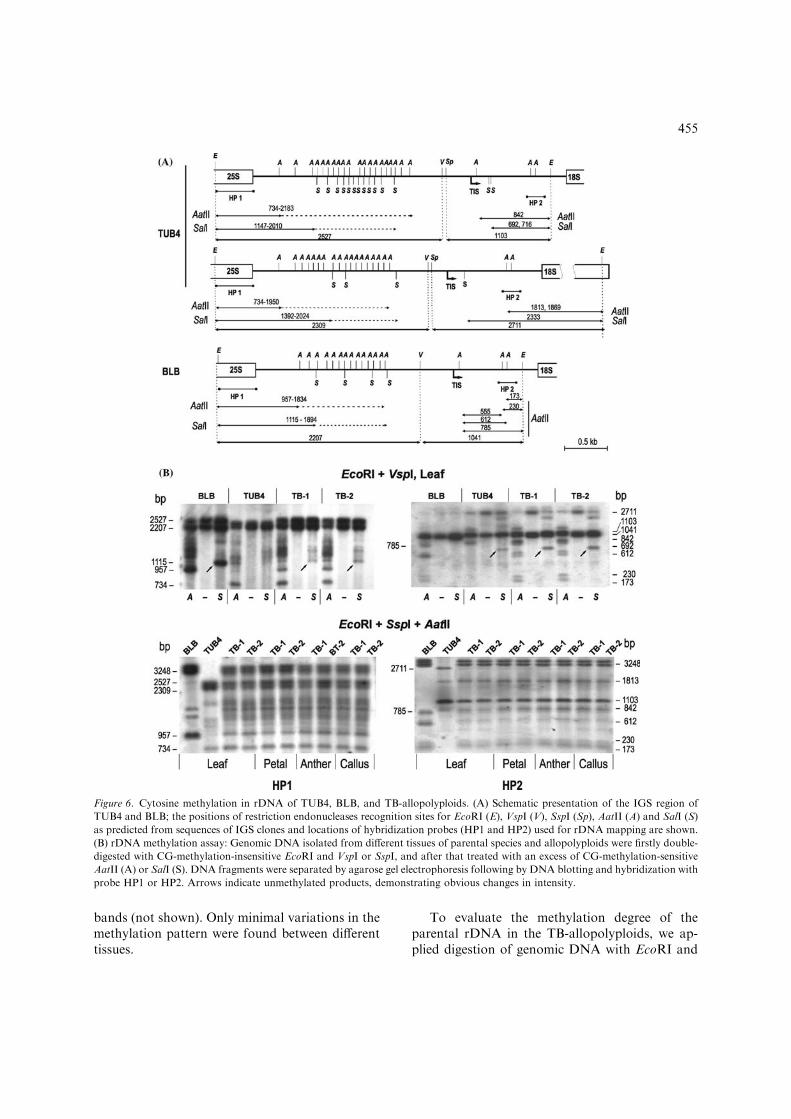
Figure 6. Cytosine methylation in rDNA of TUB4, BLB, and TB-allopolyploids. (A) Schematic presentation of the IGS region of
TUB4 and BLB; the positions of restriction endonucleases recognition sites for EcoRI (E), VspI (V), SspI (Sp), AatII (A) and SalI (S)
as predicted from sequences of IGS clones and locations of hybridization probes (HP1 and HP2) used for rDNA mapping are shown.
(B) rDNA methylation assay: Genomic DNA isolated from different tissues of parental species and allopolyploids were firstly double-
digested with CG-methylation-insensitive EcoRI and VspI or SspI, and after that treated with an excess of CG-methylation-sensitive
AatII (A) or SalI (S). DNA fragments were separated by agarose gel electrophoresis following by DNA blotting and hybridization with
probe HP1 or HP2. Arrows indicate unmethylated products, demonstrating obvious changes in intensity.
455

VspI with/without addition of AatII or SalI,respectively. Data obtained (Figure 6B) showedthat for TB-1 and TB-2 the relative intensity ofTUB-specific de-methylated restriction fragmentsremained nearly unchanged for the region up-stream of TIS and slightly increased for the regiondownstream of TIS (SalI: 692 bp), indicating aweak de-methylation of transcriptionally domi-nant TUB rDNA. In contrast, the under-dominantrDNA of BLB endured additional methylationupstream of TIS (SalI: 1115 bp). In order to betterdiscriminate methylated rDNA fragments specificfor each of the parental species, we used also an-other combination of restriction enzymes: EcoRIand SspI with addition of AatII (Figure 6) or SalI(data not shown), demonstrating that the methyl-ation patterns are very similar in different tissuesof TB-allopolyploids.
Discussion
Organization of rDNA in parental species
In the genomes of the parental Solanum speciesused for production of artificial allopolyploids andBC lines studied here only one or two variants ofthe rDNA are present as shown by restrictionmapping and further confirmed by cloning/sequencing (Figure 1). The obtained clones for theIGS region should represent ‘normal’, functionallyactive genes, because they contain highly conservedTIS and adjacent fragments of 18S and 25S rRNAcoding regions. Individual rDNA repeats appear tobe nearly identical in the same genome, e.g. twocompletely sequenced LYC IGS clones differ onlyby two nucleotides and show only three basesdifference to the earlier sequenced IGS of tomato,cv. Rutgers (Schmidt-Puchta et al., 1989; Perryand Palukaitis, 1990). Even different length vari-ants present in TUB3 and BLB reveal a highsequence similarity, from 97% to 100% in dif-ferent regions of the IGS, which is probably aresult of intensive intragenomic homogenizationof rDNA repeats.
Using species-specific oligonucleodide probeshomologous to the 50 ETS for FISH, we showedthat only one rDNA locus per chromosomal set ispresent in tomato and potato, which is in agreementwith the RFLP-mapping and cytological observa-tions that in tomato, potato and diploid tuber-
bearing Solanum species 35S rDNA is present as asingle locus at the short arm of chromosome 2 (Yehand Peloquin, 1965; Vallejos et al., 1986; Ganalet al., 1988; Visser et al., 1988; Gebhardt et al., 1991;Tanksley et al., 1992). Particularly for tomato, agenomic rDNA clone of pea (Vallejos et al., 1986)was used as a probe for in situ hybridization.However, in addition to an intensive signal at theshort arm of chromosome 2 four minor loci werefound in meiosis of tomato, if the complete tomatorDNA repeat was used for in situ hybridization (Xuand Earle, 1996). The authors speculate that the‘failure of the earlier workers to detect the minorrDNA sites may be due to the absence of the minorsite sequences in the heterologous probe from pea’(Xu and Earle, 1996). Therefore, it looks likelythat the minor rDNA sites do not contain rRNAcoding sequences, but perhaps only amplifiedfragments homologous to spacer regions, i.e. theycould represent rDNA related orphans/pseudog-enes already detected in several plants includingSolanaceae (Stupar et al., 2002; Volkov et al.,2003a; Lim et al., 2004). These orphans/pseud-ogenes can be either distributed throughout thegenome or clustered as tandem repeats. They areusually non-transcribed and demonstrate an in-creased rate of molecular evolution, but stillpossess apparent homology to functional genesand could be detected by hybridization. In ourexperiments, in order to exclude an influence ofthe putative pseudogenes we have used specificprobes/primers designed from sequences of func-tional rRNA repeats.
Thus, the available data show that one or twovariants of complete rDNA repeats are present asa single locus at chromosome 2 of the parentalSolanum species. At sequence level, a very highintragenomic homogeneity of rDNA repeats wasdemonstrated. On the other hand, comparativestudies of the rDNA of the Solanum species re-vealed prominent structural differences in numberof subrepeated elements upstream and down-stream of TIS. Therefore, the species represent anappropriate model to test the idea that structuralfeatures of the IGS could determine ND ininterspecific hybrids. In spite of high relatednessof the species studied, sufficient sequence vari-ability was found to design species-specificprobes/primers for detection and discriminationof parental rDNA and pre-rRNA in the respec-tive allopolyploids.
456

Organization of IGS and differential expressionof parental rDNA in Solanum allopolyploids
In different tissues of tomato/potato hexaploidLT-F1 and in derived BC lines the rDNA of LYCstrongly dominates over rDNA of TUB, providingfrom 95% to 100% of rRNA. Also, in leaves andpetals of symmetric TB-allotetraploids (Schilde-Rentschler et al., 1993; Stelzer, 1996), rDNA ofTUB dominates over BLB, providing 94–98% ofpre-rRNA. Estimation of rDNA repeat copynumbers showed that both LT-F1 and LT-BC1Apossess more rDNA repeats of TUB than of LYC.Equal amount of rDNA repeats from bothparental species, TUB3 and BLB, were detected inTB-1 and TB-2 (Table 2). Therefore, the differen-tial expression of rDNA appears to be indepen-dent on the amount of parental rDNA repeatspresent in the allopolyploids.
It was demonstrated that in natural and artifi-cial allopolyploids structural rearrangements ofrDNA might occur (Borisjuk et al., 1988; Clusteret al., 1996; Volkov et al., 1999), which couldinfluence transcriptional activity of rDNA. How-ever, in the allopolyploids/BC-lines studied onlyparental rDNA variants were detected, i.e. there isno evidence for recombination events.
Could some specific structural features of theIGS determine the differential expression ofparental rDNA? All species studied here possess asimilar overall organization of the IGS (Figure 1).The major differences are (i) the number of sub-repeats upstream and downstream of TIS, and (ii)the length of the AT-rich region directly upstreamof TIS. Regarding the presumptive enhancerfunction proposed for upstream subrepeats(Flavell, 1986), it could be anticipated that in LT-allopolyploids rDNA of potato, which containstwo times more upstream subrepeats, could dom-inate over rDNA of tomato. However, our data donot confirm this; in contrast a strong dominance ofLYC rDNA was demonstrated.
A possible function of upstream subrepeatsremains a subject of debates. Originally, an abilityto enhance transcription from the rDNA promoterwas found for 60- and 81-bp upstream subrepeats(Labhart and Reeder, 1984) and also demon-strated for ‘region 0 and region 1’ upstream sub-repeats (Mougey et al., 1996) of Xenopus.However, Caudy and Pikaard (2002) showed laterthat ‘region 0 and region 1 repeats do not display
significant enhancer activity’. In plants, a tran-scription enhancer function of upstream subre-peats was proposed for Triticeae (Flavell, 1986;Sardana et al., 1993), but in Arabidopsis no effectof upstream subrepeats on the RNA polymerase Ipromoter activity was found both in vitro (Doel-ling and Pikaard, 1995) and in vivo (Wanzenbocket al., 1997). Surprisingly, the upstream Sal-repeats from the IGS of A. thaliana can enhancethe transcription of protein coding genes by RNApolymerase II (Schlogelhofer et al., 2002) and cansubstitute for Xenopus repeats to enhance RNApolymerase I transcription in injected frog oocytes(Doelling et al., 1993). Besides, in natural andartificial allopolyploids between Brassica oleracea,B. nigra, and B. rapa no correlation between NDand the size of the IGS or the number of subre-peats were found (Chen and Pikaard, 1997a,b).For Brassica it is difficult to discuss presumptiveenhancer function of upstream subrepeats, becausein the species studied these elements have differentsize and belong to different structural classes(Bennett and Smith, 1991; Tremousaygue et al.,1992; Da Rocha and Bertrand, 1995; Bhatia et al.,1996). In contrast to Brassica, all Solanum speciesstudied here possess upstream subrepeats of thesame structural types differing only by a few basesubstitutions. Accordingly, our experiments clear-ly demonstrate that the number of upstream sub-repeats in the IGS do not determine preferentialexpression of rDNA in Solanum allopolyploids.Taking into account that AT-rich sequences mayfunction as enhancers of transcription (Borisjuket al., 2000; Volkov et al., 2004), it could be sup-posed that the length of the AT-rich region up-stream of TIS modulates differential transcriptionof rDNA. However, we observed that BLB rDNA,containing the longest AT-rich region, is com-pletely silenced in leaves and petals of TB-1 andTB-2.
Surprisingly, our experiments suggest a corre-lation between ND and the number of CE down-stream of TIS. For Brassicaceae, a protein factorNF-D was described, which binds both DNA andRNA at the pre-rRNA processing site in the 50
ETS (Saez-Vasquez et al., 2004). It was also pro-posed that the NF-D could interact with rDNAtranscription. In Solanum lycopersicon, the pre-rRNA processing signal is located 250 bp down-stream of the last downstream repeated element(Perry and Palukaitis, 1990), and no apparent se-
457

quence similarity was found between the NF-Dbinding site and corresponding 50 ETS region ofSolanum (not shown). In Vigna radiata, down-stream subrepeats present in the IGS can stimulatetranscription from the rDNA promoter as shownapplying both in vitro transcription with isolatednuclei or transient expression assays in protoplasts(Zentgraf and Hemleben, 1994). Therefore, weassume that in the IGS of Solanum the number ofdownstream subrepeats or perhaps only CE canalso stimulate transcription. An obvious conser-vation of CE in distantly related Solanaceae(Borisjuk et al., 1997; Volkov et al., 2003a) sug-gests that this sequence motif could be a recogni-tion site for DNA-interacting proteins involved inmodulation of rDNA transcription. Proteins,possibly involved in enhancement of rDNA tran-scription via interaction with downstream subre-peats and also with the rDNA promoter region,were described for Cucumis sativus (Zentgraf andHemleben, 1992; 1994); proteins specifically inter-acting with downstream subrepeats in the IGS ofSolanum are currently characterized in our lab(N.Y. Komarova, V. Hemleben, and R.A. Volkov,unpublished).
In addition to the enhancer imbalancehypothesis, a species-specific transcription factorhypothesis was proposed to explain ND (for re-view see Pikaard, 2000). However, this hypothesiscannot explain ND observed by crossing of closelyrelated species, such as that of Solanum used in ourexperiments. The closer relationship of tomato(LYC) to the tuber-bearing potatoes (TUB, BLB)was demonstrated using chloroplast DNA(Spooner et al., 1993), 5S rDNA (Volkov et al.,2001), and 50 ETS region (Volkov et al., 2003a;N.Y. Komarova, V. Hemleben and R.A. Volkov,unpublished results) as molecular markers. AllSolanum species studied here possess the samebasal chromosomal number (x ¼ 12); besides, highco-linearity of chromosomes of tomato and potatowas demonstrated (Gebhardt et al., 1991; Tanksleyet al., 1992).
Pre-rRNA maturation
Which molecular mechanism provides differentialaccumulation of parental pre-rRNA in interspe-cific hybrids? Available data show that ND is aphenomenon regulated at the level of transcription(Chen and Pikaard, 1997a). However, it was never
directly shown that differential expression ofrDNA remains unaffected post-transcriptionally.Here, we have quantified for the Solanum allopo-lyploids/BC-lines either the unprocessed pre-rRNA (real-time PCR; Figure 3C), simultaneouslyunprocessed and partially processed transcripts(FISH and competitive PCR, Figures 2 and 3B),and completely processed 25S rRNA (competitivePCR; Figure 4). Actually, also the 25S rRNA,representing one of the rRNA components of themature 60S ribosome subunits, was mostly sup-plied by the respective dominant parental rDNA,indicating that differential expression of parentalrDNA is also reflected in the mature rRNA.
Methylation of parental rDNA
Similar to other eukaryots, plants often containmuch more rDNA repeats then used for transcrip-tion. Respectively, only a fraction of rDNA istranscriptionally active and exists in open chro-matin state (for reviews see Hemleben et al., 1988;Grummt and Pikaard, 2003; Volkov et al., 2004).At the other hand, transcriptional silencing ofrDNAoften correlates withmethylation of cytosine(Flavell et al., 1988; Sardana et al., 1993; Torres-Ruiz and Hemleben, 1994; Chen and Pikaard,1997a,b;Houchins et al., 1997; Santoro et al., 2002),although several exceptions were also described(Macleod andBird, 1982; Chen and Pikaard, 1997a,b; Papazova et al., 2001). Recently, it was shownthat concerted changes in cytosine methylation andspecific histone modifications dictate the ‘on andoff’ states of rRNA genes in Arabidopsis. The sameregulation mechanism is used for gene dosagecontrol and ND (Lawrence et al., 2004).
In plants, CG and CNG motives are the maintargets of cytosine methylation (Gruenbaum et al.,1981). Also, density of methylation vs. methyla-tion at specific sites should be discriminated. Forinstance, it has been shown that methylation of asingle HpaII site is crucial for regulation of tran-scription of mouse rDNA in vivo and in vitro(Santoro et al., 2002). In our experiments we usedtwo methylation sensitive restriction enzymes,AatII and SalI, both of which are blocked by CG-methylation. Methylation at AatII sites, GAC-GTC, was of special interest, because only thismotif is highly conserved in the upstream sub-repeated region of the IGS of distantly related
458

Solanaceae, e.g. Solanum, Nicotiana and Atropa(R.A. Volkov, unpublished). At the other hand,the AatII recognition sequence represents a C-box,a binding site for bZIP proteins, one of the largestand most conserved group of eukaryotic tran-scription factors/repressors (Izawa et al., 1993;Niu et al., 1999).
Our data show that rDNA of LYC is nearlycompletely methylated at the AatII recognitionsites upstream of TIS and highly methylated atAatII and SalI sites downstream of TIS. Con-versely, the IGS of TUB and BLB are lessmethylated, especially upstream of TIS. In LT-al-lopolyploids the silenced TUB1 rDNA enduredadditional methylation, particularly upstream ofTIS, whereas the dominant LYC rDNA under-went a slight de-methylation. Also, in TB-1 andTB-2 the under-dominant BLB rDNA becameadditionally methylated at upstream subrepeatsand the dominant TUB4 rDNA seems to beslightly de-methylated. Thus, in leaves and petalsof Solanum allopolyploids a correlation betweenthe relative methylation status at AatII and SalIsites and preferential expression of parental rDNAwas found. The data give the impression that inSolanum the hypermethylated rRNA genes repre-sent inactive rDNA repeats as it was demonstratedfor Arabidopsis (Lawrence et al., 2004) and thatND is a phenomenon regulating via differentialsilencing rather than via activation of parentalrDNA resembling the mechanism described formammals (Santoro et al., 2002). In the IGS ofSolanum allopolyploids additional methylationwas better pronounced for the upstream regioncontaining putative enhancer subrepeats. There-fore, it could be speculated that the methylationstatus at certain sites but not the number of up-stream subrepeats may be important for differen-tial expression of rDNA.
Tissue-specificity of differential expressionof rDNA and developmental regulation
A strong dominance of tomato rDNA was foundin all organs of LT-F1 and LT-BC1A. In anthersand callus tissues of LT-BC1A a weak de-repres-sion of potato rDNA was observed. In leaves andpetals of TB-allopolyploids, rDNA of TUB dom-inates over BLB. However, in contrast to LT-al-lopolyploids, uniparental expression of rDNA wasnot found in anthers and callus tissues; similar
results were obtained for two allopolyploid lines,TB-1 and TB-2. In Brassica napus, a natural allo-tetraploid originating from diploid B. rapa andB. oleracea, rDNA transcripts of B. rapa weredetected at similar levels in all organs tested,whereas transcripts of B. oleracea were found onlyin organs delivered from the floral meristem (floralbuds, sepals, petals, anthers, and siliques) atapproximately 30–50% of B. rapa rDNA tran-script level (Chen and Pikaard, 1997b). The au-thors proposed that ND is a developmentallyregulated phenomenon and that at the floralmeristem transition stage under-dominant rDNAcan be re-activated. Our data generally confirmthis point of view and show in addition that acti-vation of under-dominant rDNA during floraldevelopment occurs in Solanum allopolyploids at alater stage, because, in contrast to B. napus, pref-erential expression of the parental rDNA remainsunaffected in petals. Moreover, activation of un-der-dominant rDNA of TUB or BLB, respectively,was found in callus demonstrating that ND is aphenomenon connected with tissue differentiation.In general, activation of under-dominant rDNAcan be well (TB-allopolyploids, B. napus) orweakly (LT-allopolyploids) manifested.
Remarkably, in different tissues of Solanumallopolyploids studied only minimal alterations inrDNA methylation were observed (Figures 5 and6), although in anthers and callus tissues of TB-1and TB-2 an obvious activation of BLB rDNAtakes place. Hence, the differential activation/silencing of rDNA in anthers and callus tissues isnot connected with changes of cytosine methyla-tion at AatII/SalI sites in the IGS. Perhapsselective methylation at other sites could play arole.
Genome constitution and differential expression
Several observations demonstrated that ND is aphenomenon regulated at the chromosomal/genomic level: e.g. changes of chromosomallocalization of NOR or presence/absence of somechromosomal arms can result in alteration of ND(Nicoloff, 1979; Schubert and Kunzel, 1990; Vieraet al., 1990a,b; Neves et al., 1997; Georgiev et al.,2001), indicating that factors like the positionwithin the three-dimensional space of the nucleus(Csink and Henikoff, 1996) and activity of un-linked genes should be taking into consideration.
459

In contrast, a very strong dominance of LYC-rDNA observed in our studies in LT-F1, contain-ing two complete tomato and four potatochromosome sets, was only slightly reduced in theback-cross lines LT-BC1A/B, carrying four potatoand one incomplete tomato chromosome sets, andeven in the monosomic addition line LT-BC2,which possess only one tomato chromosome 2 atthe back-ground of four potato genomes. Thesilencing of TUB-rDNA cannot be a specific fea-ture of a particular potato variety, because severalpotato lines were used for the production of the BCplants studied. Therefore, dominance of LYC-rDNA seems to be independent of the chromo-some/genomic context, i.e. tomato chromosome 2alone is able to grant transcriptional dominance ofLYC rDNA. Although at the moment we cannotexclude that an unknown gene located at thechromosome 2 can be crucial for the ND, theavailable data allow the assumption that localfeatures of rDNA, especially the number and/ormethylation status of subrepeated elements in theIGS, are the main factors responsible for differen-tial expression of rDNA in Solanum allopolyploids.
Acknowledgements
This work was supported by the Deutsche Fors-chungsgemeinschaft (DFG) and the DeutscheAkademische Austauschdienst (DAAD). We aregrateful to Prof. Dr. E. Jacobsen and Dr. H.J. deJong (Wageningen University, The Netherlands)for providing S. lycopersicum + S. tuberosumallopolyploids and parental forms and Dr.L. Schilde-Rentschler and Prof. Dr. H. Ninne-mann (University of Tubingen, Germany) forproviding S. tuberosum + S. bulbocastanumallopolyploids and parental lines.
References
Arumuganathan, K. and Earle, E.D. 1991. Nuclear DNAcontent of some important plant species. Plant Mol. Biol.Rep. 9: 208–218.
Bennett, R.I. and Smith, A.G. 1976. Nuclear DNA amounts inangiosperms. Phils. Trans. R. Soc. London B274: 227–274.
Bennett, R.I. and Smith, A.G. 1991. The complete nucleotidesequence of the intergenic spacer region of an rDNA operonfrom Brassica oleracea and its comparison with othercrucifers. Plant Mol. Biol. 16: 1095–1098.
Bhatia, S., Singth Negi, M. and Lakshmikumaran, M. 1996.Structural analysis of the rDNA intergenic spacer of Brassicanigra: evolutionary divergence of the spacers of the threediploid Brassica species. J. Mol. Evol. 43: 460–468.
Borisjuk, N., Borisjuk, L., Komarnytsky, S., Timeva, S.,Hemleben, V., Gleba, Y. and Raskin, I. 2000. Tobaccoribosomal DNA spacer element stimulates amplification andexpression of heterologous genes. Nat. Biotechnol. 18: 1303–1306.
Borisjuk, N., Borisjuk, L., Petjuch, G. and Hemleben, V. 1994.Comparison of nuclear ribosomal RNA genes amongSolanum species and other Solanaceae. Genome 37: 271–279.
Borisjuk, N.V., Davidjuk, Y.M., Kostishin, S.S.,Miroshnichenko, G.P., Velasco, R., Hemleben, V. andVolkov, R.A. 1997. Structural analysis of rDNA in thegenus Nicotiana. Plant Mol. Biol. 35: 655–660.
Borisjuk. N. and Hemleben, V. 1993. Nucleotide sequence ofpotato rDNA intergenic spacer. Plant Mol. Biol. 21:381–384.
Borisjuk, N.V., Momot, V.P. and Gleba Y. 1988. Novel class ofrDNA repeat units in somatic hybrids between Nicotianaand Atropa. Theor. Appl. Genet. 76: 108–112.
Busby, S.J. and Reeder, R.H. 1983. Spacer sequences regulatetranscription of ribosomal gene plasmids injected intoXenopus embryos. Cell 34: 989–996.
Caudy, A.A. and Pikaard, C.S. 2002. Xenopus ribosomal RNAgene intergenic spacer elements conferring transcriptionalenhancement and nucleolar dominance-like competition inoocytes. J. Biol. Chem. 277: 31577–31584.
Chen, Z.J., Comai, L. and Pikaard, C.S. 1998. Gene dosage andstochastic effects determine the severiti and direction ofuniparental ribosomal RNA gene silencing (nucleolar dom-inance) in Arabidopsis allopolyploids. Proc. Natl. Acad. Sci.USA 95: 14891–14896.
Chen, Z.J. and Pikaard, C.S. 1997a. Epigenetic silencing ofRNA polymerase I transcription: a role for DNA methyl-ation and histone modification in nucleolar dominance.Genes Dev. 11: 2124–2136.
Chen, Z.J. and Pikaard, C.S. 1997b. Transcriptional analysis ofnucleolar dominance in polyploid plants: biased expression/silencing of progenitor rRNA genes is developmentallyregulated in Brassica. Proc. Natl. Acad. Sci. USA 94:3442–2447.
Cluster, P.D., Calderini, O., Puppilli, F., Crea, F., Damiani, F.and Arcioni, S. 1996. The fate of ribosomal genes in threeinterspecific somatic hybrids of Medicago sativa; threedifferent outcomes including the rapid amplification of newspacer-length variants. Theor. Appl. Genet. 93: 801–808.
Csink, A.K. and Henikoff, S. 1996. Genetic modification ofheterochromatic association and nuclear organization inDrosophila. Nature 381: 529–531.
Da Rocha, P.S.C.F. and Bertrand, H. 1995. Structure andcomparative analysis of the rDNA intergenic spacer ofBrassica rapa: implications for the function and evolution ofthe Cruciferae spacer. Eur. J. Biochem. 229: 550–557.
Doelling, J.H., Gaudino, R.J. and Pikaard, C.S. 1993. Func-tional analysis of Arabidopsis thaliana rRNA gene andspacer promoters in vivo and by transient expression. Proc.Natl. Acad. Sci. USA 90: 7528–7532.
Doelling, J.H. and Pikaard, C.S. 1995. The minimal ribosomalRNA gene promoter of Arabidopsis thaliana includes acritical element at the transcription initiation site. Plant J. 8:683–692.
460

Doelling, J.H. and Pikaard, C.S. 1996. Species-specificity ofrRNA gene transcription in plants manifested as a switch inpolymerase-specificity. Nucleic Acids Res. 24: 4725–4732.
Dover, G.A. and Flavell, R.V. 1984. Molecular co-evolution:rDNA divergence and the maintenance of function. Cell 38:622–623.
Engel, J. 1997. Signifikante Schule der schlichten Statistik.Filander Verlag GmbH, Furth, Germany.
Fan, H., Kimitaka, Y., Miyanishi, M., Sugita, M. andSugiura, M. 1995. In vitro transcription of plant RNApolymerase I-dependent rRNA genes is species-specific.Plant J 8: 295–298.
Flavell, R.B. 1986. The structure and control of expression ofribosomal RNA genes. Ox. Surv. Plant Mol. Cell Biol. 3:252–274.
Flavell, R.B., O’Dell, M. and Thompson, W.F. 1988.Regulation of cytosine methylation in ribosomal DNAand nucleolus organizer expression in wheat. J. Mol. Biol.204: 523–534.
Frieman, M., Chen, Z.J., Saez-Vasquez, J., Shen, L.A. andPikaard, C.S. 1999. RNA polymerase I transcription in aBrassica interspecific hybrid and its progenitors: tests oftranscription factor involvement in nucleolar dominance.Genetics 152: 451–460.
Ganal, M.W., Lapitan, N.L.V. and Tanksley, S.D. 1988. Amolecular and cytogenetic survey of major repeated DNAsequences in tomato (Lycopersicon esculentum). Mol. Gen.Genet. 213: 262–268.
Garriga-Caldere, F., Huigen, D.J., Angrisano, A., Jacobsen,E. and Ramanna, M.S. 1998. Transmission of alientomato chromosomes from BC1 to BC2 progenies derivedfrom backcrossing potato(+)tomato fusion hybrids topotato: the selection of single additions for seven differenttomato chromosomes. Theor. Appl. Genet. 96: 155–163.
Garriga-Caldere, F., Huigen, D.J., Filotico, F., Jacobsen, E.and Ramanna, M.S. 1997. Identification of alien chromo-somes through GISH and RLFP analysis and the potentialfor the establishment of potato lines with monosomicadditions of tomato chromosomes. Genome 40: 666–673.
Gebhardt, C., Ritter, E., Barone, A., Debener, T., Walkeme-ier, B., Schachtschabel, U., Kaufmann, H., Thomson,R.D., Bonierbale, M.W., Ganal, M.W., Tanksley, S.D.and Salamini, F. 1991. RFLP maps of potato and theiralignment with the homologous tomato genome. Theor.Appl. Genet. 83: 49–57.
Georgiev, S., Papazova, N. and Gecheff, K. 2001. Transcrip-tional activity of an inversion split NOR in barley(Hordeum vulgare L.). Chromosome Res. 9: 507–514.
Grant, V. 1981. Plant Speciation, 2nd edn. ColumbiaUniversity Press, New York.
Gruenbaum, Y., Naveh-Many, T., Cedar, H. and Razin, A.1981. Sequence specificity of methylation in higher plantDNA. Nature 292: 860–862.
Grummt, I. and Pikaard, C.S. 2003. Epigenetic mechanismscontrolling RNA polymerase I transcription. Nat. Rev.Mol. Cell Biol. 4: 641–649.
Grummt, I., Roth, E. and Paule, M.R. 1982. rRNAtranscription in vitro is species-specific. Nature 296:173–174.
Gustafson, J.P., Dera, A.R. and Petrovic, S. 1988. Expressionof modified rye ribosomal RNA genes in wheat. Proc.Natl. Acad. Sci. USA 85: 3943–3945.
Heitz, E. 1931. Nucleolen und Chromosomen in der GattungVicia. Planta 15: 495–505.
Hemleben, V., Ganal, M., Gerstner, J., Schiebel, K., Torres,R.A. 1988. Organization and length heterogeneity of plantribosomal RNA genes. In: G. Kahl (Ed.), The Architec-ture of Eukaryotic Gene. VHC. Weinheim, pp. 371–384.
Honjo, T. and Reeder, R.H. 1973. Preferential transcriptionof Xenopus laevis ribosomal RNA in interspecific hybridsbetween Xenopus laevis and Xenopus mulleri. J. Mol. Biol.80: 217–228.
Houchins, K., O’Dell, M., Flavell, R.B. and Gustafson, J.P.1997. Cytosine methylation and nucleolar dominance incereal hybrids. Mol. Gen. Genet. 55: 294–301.
Izawa, T., Foster, R. and Chue, N.-H. 1993. Plant bZIP proteinDNA binding specificity. J. Mol. Biol. 230: 1131–1144.
Jacobsen, E., De Jong, J.H., Kamstra; S.A., Van Den Berg,P.M.M.M. and Ramanna, M.S. 1995. Genomic in situhybridization (GISH) and RFLP analysis for the identifi-cation of alien chromosomes in the backcross progenyof potato(+)tomato fusion hybrids. Heredity 74: 250–257.
Kiss, T., Kis, M. and Solymosy, F. 1989a. Nucleotide sequenceof a 25S rRNA gene from tomato. Nucleic Acids Res. 17: 796.
Kiss, T., Szkukalek, A. and Solymosy, F. 1989b. Nucleotidesequence of a 17S (18S) rRNA gene from tomato. NucleicAcids Res. 17: 2127.
Labhart, P. and Reeder, R.H. 1984. Enhancer-like properties ofthe 60/81 bp elements in the ribosomal gene spacer ofXenopus laevis. Cell 37: 285–289.
Lawrence, R.J., Earley, K., Pontes, O., Silva, M., Chen, Z.J.,Neves, N., Viegas, W. and Pikaard, C.S. 2004. A concertedDNA methylation/histone methylation switch regulatesrRNA gene dosage control and nucleolar dominance. Mol.Cell 13: 599–609.
Lee, H.S. and Chen, Z.J. 2001. Protein-coding genes areepigenetically regulated in Arabidopsis allopolyploids. Proc.Natl. Acad. Sci. 98: 6753–6758.
Levin, D.A. 2002. The Role of Chromosomal Change in PlantEvolution. Oxford University Press, Oxford, UK.
Lim, K.Y., Skalicka, K., Koukalova, B., Volkov, R.A.,Matyasek, R., Hemleben, V., Leitch, A.R. and Kovarik,A. 2004. Dynamic changes in the distribution of a satellitehomologous to intergenic 26-18S rDNA spacer in theevolution of Nicotiana. Genetics 166: 1935–1946.
Macleod, D. and Bird, A. 1982. DNAase I sensitivity andmethylation of active versus inactive rRNA genes in xenopusspecies hybrids. Cell 29: 211–218.
McClintock, B. 1934. The relationship of a particular chromo-somal element to the development of the nucleoli in Zeamays. Z. Zellforsch. Mikrosk. Anat. 21: 294–328.
Mishima, Y., Financsek, I., Kominami, R. and Muramatsu, M.1982. Fractionation and reconstitution of factors requiredfor accurate transcription of mammalian ribosomal genes:identification of species-dependent initiation factor. NucleicAcids Res. 10: 6659–6670.
Moss, T. 1983. A transcription function for the repetitiveribosomal spacer in Xenopus laevis. Nature 302: 223–228.
Mougey, E.B., Pape, L.K. and Sollner-Webb, B. 1996. J.Virtually the entire Xenopus laevis rDNA multikilobaseintergenic spacer serves to stimulate polymerase I transcrip-tion. Biol. Chem. 271: 27138–27145.
Murashige, T. and Skoog, F. 1962. A revised medium for rapidgrowth and bioassays with tobacco tissue cultures. Physiol.Plant. 15: 437–497.
Navashin, M. 1927. Changes in number and form of chromo-somes as a result of hybridization. Z. Zellforsch. Mikrosk.Anat. 6: 195–223.
461

Navashin, M. 1934. Chromosomal alterations caused byhybridization and their bearing upon certain general geneticproblems. Cytologia 5: 169–203.
Neves, N., Silva, M., Heslop-Harrison, J.S. and Viegas, W.1997. Nucleolar dominance in triticales: control by unlinkedgenes. Chromosome Res. 5: 125–131.
Nicoloff, H. 1979. Nucleolar dominance is observed in barleytranslocation lines with specifically reconstructed SATchromosomes. Theor. Appl. Genet. 55: 247–251.
Niu, X.P., Renshaw-Gegg, L., Miller, L. and Guiltinan, M.J.1999. Bipartite determinants of DNA-binding specificity ofplant basic leucine zipper proteins. Plant Mol. Biol. 41:1–13.
Oberwalder, B., Rouss, B., Schilde-Rentschler, L., Hemleben,V. and Ninnemann, H. 1997. Asymmetric fusion betweenwild and cultivated species of potato (Solanum spp.) –detection of asymmetric hybrids and genome elimination.Theor. Appl. Genet. 94: 1104–1112.
Osborn, T.C., Pires, J.C., Birchler, J.A., Auger, D.L., Chen,Z.J., Lee, H.S., Comai, L., Madlung, A., Doerge, R.W.,Colot, V. and Martienssen, R.A. 2003. Understandingmechanisms of novel gene expression in polyploids. TrendsGenet. 19: 141–147.
Papazova, N., Hvarleva, T., Atanassov, A. and Gecheff, K.2001. The role of cytosine methylation for rRNA geneexpression in reconstructed karyotypes of barley. Biotechnol.Equipment 15: 35–44.
Pape, L.K., Windile, J.J., Mougey, E.B. and Sollner-Webb, B.1989. The Xenopus ribosomal DNA 60- and 81-base-pairrepeats are position-dependent enhancers that function atthe establishment of the preinitiation complex: analysis invivo and in an enhancer-responsive in vitro system. Mol. CellBiol. 9: 5093–5104.
Perry, K.L. and Palukaitis, P. 1990. Transcription of tomatoribosomal DNA and the organization of the intergenicspacer. Mol. Gen. Genet. 221: 102–112.
Pikaard, C.S. 2000. Nucleolar dominance: uniparental genesilencing on a multi-megabase scale in genetic hybrids. PlantMol. Biol. 43: 163–177.
Pikaard, C.S. and Reeder, R.H. 1988. Sequence elementsessential for function of the Xenopus laevis ribosomal DNAenhancers. Mol. Cell Biol. 8: 4282–4288.
Reeder, R.H. 1985. Mechanisms of nucleolar dominance inanimals and plants. J. Cell Biol. 101: 2013–2016.
Riddle, N.C. and Birchler, J.A. 2003. Effects of reuniteddiverged regulatory hierarchies in allopolyploids and specieshybrids. Trends Genet. 19: 597–600.
Saez-Vasquez, J., Caparros-Ruiz, D., Barneche, F. andEcheverria, M. 2004. A plant snoRNP complex containingsnoRNAs, fibrillarin, and nucleolin-like proteins is compe-tent for both rRNA gene binding and pre-rRNA processingin vitro. Mol. Cell. Biol. 24: 7284–7297.
Saez-Vasquez, J. and Pikaard, C.S. 1997. Extensive purificationof a putative RNA polymerase I holoenzyme from plantsthat accurately initiates rRNA gene transcription in vitro.Proc. Natl. Acad. Sci. USA 94: 11869–11874.
Sambrook, J., Fritsch, E. and Maniatis, T. 1989. MolecularCloning: A Laboratory Manual, 2nd edn. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
Santoro, R., Li, J. and Grummt, I. 2002. The nucleolarremodeling complex NoRC mediates heterochromatin for-mation and silencing of ribosomal gene transcription.Nature Genet. 32: 393–396.
Sardana, R., O’Dell, A. and Flavell, R. 1993. Correlationbetween the size of the intergenic regulatory region, thestatus of cytosine methylation of rRNA genes and nucleolarexpression in wheat. Mol. Gen. Genet. 236: 155–162.
Schilde-Rentschler, L., Ruoss, B. and Ninnemann, H. 1993.Availability of new genetic resources from wild species forpotato breeding. In: EAPR 12th Conference Trienniale,Paris, pp. 195–196.
Schlogelhofer, P., Nizhynska, V., Feik, N., Chambon, C.,Potuschak, T., Wanzenbock, E.M., Schweizer, D. andBachmair, A. 2002. The upstream Sal repeat-containingsegment of Arabidopsis thaliana ribosomal DNA intergenicregion (IGR) enhances the activity of adjacent protein-coding genes. Plant Mol. Biol. 49: 655–667.
Schmidt-Puchta, W., Gunther, I. and Sanger, H.L. 1989.Nucleotide sequence of the intergenic spacer (IGS) of thetomato ribosomal DNA. Plant Mol. Biol. 13: 251–253.
Schubert, I. and Kunzel, C. 1990. Position-dependent NORactivity in barley. Chromosoma 99: 352–359.
Schwarzacher, T. and Heslop-Harrison, P. 2000. PracticalIn Situ Hybridization. The Bath Press, Bath, UK.
Schweizer, G., Borisjuk, N., Borisjuk, L., Stadler, M., Stelzer,T., Schilde, L. and Hemleben, V. 1993. Molecular analysis ofhighly repeated genome fractions in Solanum and their use asmarkers for the characterization of species and cultivar.Theor. Appl. Genet. 85: 801–808.
Stelzer, T. 1996. Vorkommen repetitiver DNA-Elemente unddetaillierte Genomanalyse von somatischen Hybriden ausintra- und interspezifischen, symmetrischen Fusionen derGattung Solanum. Ph.D. Dissertation, Verlag Ulrich E.Grauer, Sturttgart, Germany.
Song, K., Lu, P., Tang, K. and Osborn, T.C. 1995. Rapidgenome change in synthetic polyploids of Brassica and itsimplications for polyploid evolution. Proc. Natl. Acad. Sci.92: 7719–7723.
Spooner, D.M., Anderson, G.J. and Jansen, R.K. 1993. Chloro-plast DNA evidence for the interrelationships of tomatoes,potatoes, and pepinos (Solanaceae). Am. J. Bot. 80: 676–688.
Stupar, R.M., Song, J., Tek, A.L., Cheng, Z., Dong, F. andJiang, J. 2002. Highly condensed potato pericentromericheterochromatin contains rDNA-related tandem repeats.Genetics 162: 1435–1444.
Tanksley, S.D., Ganal, M.W., Prince, J.P., De Vicente, M.C.,Bonierbale, M.W., Broun, P., Fulton, T.M., Giovannoni,J.J., Grandillo, S., Martin, G.B., Messeguer, R., Miller, J.C.,Miller, L., Paterson, A.H., Pineda, O., Roder, M.S., Wing,R.A., Wu, W. and Young, N.D. 1992. High densitymolecular linkage maps of the tomato and potato genomes.Genetics 132: 1141–1160.
Torres-Ruiz, R.A. and Hemleben, V. 1994. Pattern and degreeof methylation in ribosomal RNA genes of Cucurbita pepo L.Plant Mol. Biol. 26: 1167–1179.
Tremousaygue, D., Laudie, M., Grellet, F. and Delseny, M.1992. The Brassica oleracea rDNA spacer revisited. PlantMol. Biol. 18: 1013–1018.
Vallejos, C.E., Tanksley, S.D. and Bernatzky, R. 1986. Local-ization in the tomato genome of DNA restriction fragmentscontaining sequences homologous to the rRNA (45S), hemajor chlorophyll a/b binging polypeptide and the ribuloso-bisphosphate carboxylase genes. Genetics 112: 93–105.
Viera, A., Mello-Sampayo, T. and Viegas, W.S. 1990a. Geneticcontrol of 1R nucleolus organizer region in the presence ofwheat genome. Genome 33: 713–718.
462

Viera, A., Morais, L., Barao, A., Mello-Sampayo, T. andViegas, W.S. 1990b. 1R chromosome nucleolus organizerregion activation by 5-azacytidine in wheat · rye hybrids.Genome 33: 707–712.
Visser, R., Hoekstra, R., Van der Leji, F., Witholt, B. andFeenstra,W. 1988. In situ hybridization to somatic metaphasechromosomes of potato. Theor. Appl. Genet. 76: 420–424.
Volkov, R.A., Borisjuk, N.V., Panchuk, I.I., Schweizer, D. andHemleben, V. 1999. Elimination and rearrangement ofparental rDNA in allotetraploid Nicotiana tabacum. Mol.Biol. Evol. 16: 311–320.
Volkov, R.A., Zanke, C., Panchuk, I.I. and Hemleben, V. 2001.Molecular evolution of 5S rDNA of Solanum species (sect.Petota): application for molecular phylogeny and breeding.Theor. Appl. Genet. 103: 1273–1282.
Volkov. R.A., Komarova, N.Y., Panchuk, I.I. and Hemleben,V. 2003a. Molecular evolution of rDNA external transcribedspacer and phylogeny of sect. Petota (genus Solanum) Mol.Phylogenet. Evol. 29: 187–202.
Volkov, R.A., Panchuk, I.I. and Schoffl, F. 2003b. Heat-stress-dependency and developmental modulation of gene expres-sion: the potential of house-keeping genes as internalstandards in mRNA expression profiling using real-timeRT-PCR. J. Exp. Bot. 391: 2343–2349.
Volkov, R.A., Medina, F.J., Zentgraf, U. and Hemleben V.2004. Molecular cell biology: Organization and molecularevolution of rDNA, nucleolar dominance, and nucleolusstructure. In: Progress in Botany 65. Springer-Verlag,Berlin–Heidelberg, pp. 106–146.
Wallace, H. and Langbridge, W.H.R. 1971. Differential amp-hiplasty and the control of ribosomal RNA synthesis.Heredity 27: 1–13.
Wanzenbock, E.M., Schofer, C, Schweizer, D. and Bachmair,A. 1997. Ribosomal transcription units integrated viaT-DNA transformation associate with the nucleolus anddo not require upstream repeat sequences for activity inArabidopsis thaliana. Plant J. 11: 1007–1016.
Wendel, J.F., Schnabel, A. and Seelanan, T. 1995. Bidirectionalinterlocus concerted evolution following allopolyploidspeciation in cotton (Gossypium). Proc. Natl. Acad. Sci. 92:280–284.
Xu, J. and Earle, E.D. 1996. High resolution physical mappingof 45S (5.8S, 18S and 25S) rDNA gene loci in the tomatogenome using a combination of karyotyping and FISH ofpachytene chromosomes. Chromosoma 104: 545–550.
Yeh, B.P. and Peloquin, S.J. 1965. The nucleolus associatedchromosome of Solanum species and hybrids. Am. J. Bot. 52:626.
Zentgraf, U. and Hemleben, V. 1992. Complex formation ofnuclear proteins with the spacer of Cucumis sativus ribo-somal DNA. Nucl. Acids Res. 20: 3685–3691.
Zentgraf, U. and Hemleben, V. 1994. Structural organizationand regulation of transcription by RNA polymerase I ofplant nuclear ribosomal RNA genes. In: L. Nover (Ed.),Plant Promoters and Transcription Factors. Results andProblems in Cell Differentiation 20, Springer-Verlag, Berlin–Heidelberg, pp. 3–24.
463




















