
Nuclear Transfer of Synchronized African Wild Cat Somatic Cells into Enucleated Domestic Cat...
10
1032 BIOLOGY OF REPRODUCTION 69, 1032–1041 (2003) Published online before print 28 May 2003. DOI 10.1095/biolreprod.102.014449 Nuclear Transfer of Synchronized African Wild Cat Somatic Cells into Enucleated Domestic Cat Oocytes 1 Martha C. Go ´ mez, 2,3,4 Jill A. Jenkins, 5 Angelica Giraldo, 3,4 Rebecca F. Harris, 4 Amy King, 4 Betsy L. Dresser, 4,6 and Charles Earle Pope 4 Department of Animal Science, 3 Louisiana State University, Baton Rouge, Louisiana 70808 Audubon Center for Research of Endangered Species, 4 New Orleans, Louisiana 70131 National Wetlands Research Center, 5 U.S. Geological Survey, Lafayette, Louisiana 70506 Department of Biological Sciences, 6 University of New Orleans, New Orleans, Louisiana 70148 ABSTRACT The African wild cat is one of the smallest wild cats and its future is threatened by hybridization with domestic cats. Nucle- ar transfer, a valuable tool for retaining genetic variability, offers the possibility of species continuation rather than extinction. The aim of this study was to investigate the ability of somatic cell nuclei of the African wild cat (AWC) to dedifferentiate within domestic cat (DSH) cytoplasts and to support early development after nuclear transplantation. In experiment 1, distributions of AWC and DSH fibroblasts in each cell-cycle phase were assessed by flow cytometry using cells cultured to confluency and dis- aggregated with pronase, trypsin, or mechanical separation. Trypsin (89.0%) and pronase (93.0%) yielded higher propor- tions of AWC nuclei in the G 0 /G 1 phase than mechanical sepa- ration (82.0%). In contrast, mechanical separation yielded high- er percentages of DSH nuclei in the G 0 /G 1 phase (86.6%) than pronase (79.7%) or trypsin (74.2%) treatments. In both species, pronase induced less DNA damage than trypsin. In experiment 2, the effects of serum starvation, culture to confluency, and exposure to roscovitine on the distribution of AWC and DSH fibroblasts in various phases of the cell cycle were determined. Flow cytometry analyses revealed that the dynamics of the cell cycle varied as culture conditions were modified. Specifically, a higher percentage of AWC and DSH nuclei were in the G 0 /G 1 phase after cells were serum starved (83% vs. 96%) than were present in cycling cells (50% vs. 64%), after contact inhibition (61% vs. 88%), or after roscovitine (56% vs. 84%) treatment, respectively. In experiment 3, we evaluated the effects of cell synchronization and oocyte maturation (in vivo vs. in vitro) on the reconstruction and development of AWC-DSH- and DSH- DSH-cloned embryos. The method of cell synchronization did not affect the fusion and cleavage rate because only a slightly higher percentage of fused couplets cleaved when donor nuclei were synchronized by serum starvation (83.0%) than after ros- covitine (80.0%) or contact-inhibition (80.0%). The fusion ef- ficiency of in vivo and in vitro matured oocytes used as recipient cytoplasts of AWC donor nuclei (86.6% vs. 85.2%) was similar to the rates obtained with DSH donor nuclei, 83.7% vs. 73.0%, respectively. The only significant effect of source of donor nu- cleus (AWC vs. DSH) was on the rate of blastocyst formation in 1 Supported by the E.J. and Marjory Ourso Foundation and the John and Shirley Davies Foundation. 2 Correspondence: Martha C. Go ´mez, Audubon Center for Research of Endangered Species, 14001 River Road, New Orleans, LA, 70131. FAX: 504 3917707; e-mail: [email protected] Received: 9 December 2002. First decision: 9 January 2003. Accepted: 14 May 2003. Q 2003 by the Society for the Study of Reproduction, Inc. ISSN: 0006-3363. http://www.biolreprod.org vitro. A higher percentage of the embryos derived from AWC nuclei developed to the blastocyst stage than did embryos pro- duced from DSH nuclei, 24.2% vs. 3.3%, respectively (P , 0.05). In experiment 4, the effect of calcium in the fusion me- dium on induction of oocyte activation and development of AWC-DSH-cloned embryos was determined. The presence of calcium in the fusion medium induced a high incidence of cleav- age of DSH oocytes (54.3%), while oocyte cleavage frequency was much lower in the absence of calcium (16.6%). The pres- ence or absence of calcium in the fusion medium did not affect the fusion, cleavage, and blastocyst development of AWC-DSH- cloned embryos. In experiment 5, AWC-DSH-cloned embryos were transferred to the uteri of 11 synchronized domestic cat recipients on Day 6 or 7 after oocyte aspiration. Recipients were assessed by ultrasonography on Day 21 postovulation, but no pregnancies were observed. In the present study, after NT, AWC donor nuclei were able to dedifferentiate in DSH cytoplasts and support high rates of blastocyst development in vitro. Incom- plete reprogramming of the differentiated nucleus may be a ma- jor constraint to the in vivo developmental potential of the em- bryos. assisted reproductive technology, early development, gamete bi- ology INTRODUCTION Most of the 36 species of wild felids are classified as threatened, vulnerable or endangered [1]. As an example, the African wild cat (Felis silvestris libica) is one of the smallest wild cats, whose future is threatened by hybrid- ization with domestic cats (Felis silvestris catus). Much progress has been made toward applying assisted reproduc- tive technologies for aiding in the conservation of endan- gered felids. In particular, nuclear transfer (NT) is a valu- able tool for retaining genetic variability [2, 3] and offers the possibility of species continuation rather than extinc- tion. Interspecies NT into bovine, rabbit, and sheep cyto- plasts has been shown to support development of somatic cell nuclei from various mammalian species [4–8]. Accord- ingly, ooplasm of the domestic cat is a potential host for somatic cell nuclei from endangered felid species. During normal fertilization and embryo development, mitochondrial DNA (mtDNA) is maternally inherited [9]. Animals produced by NT are genetic duplicates of the do- nor cell nuclear genome. Because foreign mitochondria are introduced into the oocyte, some degree of heteroplasmy can occur after NT [10], which, according to speculation, may be detrimental. There are some species-specific epi- genetic factors present in the oocyte cytoplasm that may lead to nucleo:cytoplasmic incompatibilities either imme- diately after NT or at later stages of development [11].
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Nuclear Transfer of Synchronized African Wild Cat Somatic Cells into Enucleated Domestic Cat...

1032
BIOLOGY OF REPRODUCTION 69, 1032–1041 (2003)Published online before print 28 May 2003.DOI 10.1095/biolreprod.102.014449
Nuclear Transfer of Synchronized African Wild Cat Somatic Cells into EnucleatedDomestic Cat Oocytes1
Martha C. Gomez,2,3,4 Jill A. Jenkins,5 Angelica Giraldo,3,4 Rebecca F. Harris,4 Amy King,4
Betsy L. Dresser,4,6 and Charles Earle Pope4
Department of Animal Science,3 Louisiana State University, Baton Rouge, Louisiana 70808Audubon Center for Research of Endangered Species,4 New Orleans, Louisiana 70131National Wetlands Research Center,5 U.S. Geological Survey, Lafayette, Louisiana 70506Department of Biological Sciences,6 University of New Orleans, New Orleans, Louisiana 70148
ABSTRACT
The African wild cat is one of the smallest wild cats and itsfuture is threatened by hybridization with domestic cats. Nucle-ar transfer, a valuable tool for retaining genetic variability, offersthe possibility of species continuation rather than extinction.Theaim of this study was to investigate the ability of somatic cellnuclei of the African wild cat (AWC) to dedifferentiate withindomestic cat (DSH) cytoplasts and to support early developmentafter nuclear transplantation. In experiment 1, distributions ofAWC and DSH fibroblasts in each cell-cycle phase were assessedby flow cytometry using cells cultured to confluency and dis-aggregated with pronase, trypsin, or mechanical separation.Trypsin (89.0%) and pronase (93.0%) yielded higher propor-tions of AWC nuclei in the G0/G1 phase than mechanical sepa-ration (82.0%). In contrast, mechanical separation yielded high-er percentages of DSH nuclei in the G0/G1 phase (86.6%) thanpronase (79.7%) or trypsin (74.2%) treatments. In both species,pronase induced less DNA damage than trypsin. In experiment2, the effects of serum starvation, culture to confluency, andexposure to roscovitine on the distribution of AWC and DSHfibroblasts in various phases of the cell cycle were determined.Flow cytometry analyses revealed that the dynamics of the cellcycle varied as culture conditions were modified. Specifically, ahigher percentage of AWC and DSH nuclei were in the G0/G1
phase after cells were serum starved (83% vs. 96%) than werepresent in cycling cells (50% vs. 64%), after contact inhibition(61% vs. 88%), or after roscovitine (56% vs. 84%) treatment,respectively. In experiment 3, we evaluated the effects of cellsynchronization and oocyte maturation (in vivo vs. in vitro) onthe reconstruction and development of AWC-DSH- and DSH-DSH-cloned embryos. The method of cell synchronization didnot affect the fusion and cleavage rate because only a slightlyhigher percentage of fused couplets cleaved when donor nucleiwere synchronized by serum starvation (83.0%) than after ros-covitine (80.0%) or contact-inhibition (80.0%). The fusion ef-ficiency of in vivo and in vitro matured oocytes used as recipientcytoplasts of AWC donor nuclei (86.6% vs. 85.2%) was similarto the rates obtained with DSH donor nuclei, 83.7% vs. 73.0%,respectively. The only significant effect of source of donor nu-cleus (AWC vs. DSH) was on the rate of blastocyst formation in
1Supported by the E.J. and Marjory Ourso Foundation and the John andShirley Davies Foundation.2Correspondence: Martha C. Gomez, Audubon Center for Research ofEndangered Species, 14001 River Road, New Orleans, LA, 70131.FAX: 504 3917707; e-mail: [email protected]
Received: 9 December 2002.First decision: 9 January 2003.Accepted: 14 May 2003.Q 2003 by the Society for the Study of Reproduction, Inc.ISSN: 0006-3363. http://www.biolreprod.org
vitro. A higher percentage of the embryos derived from AWCnuclei developed to the blastocyst stage than did embryos pro-duced from DSH nuclei, 24.2% vs. 3.3%, respectively (P ,0.05). In experiment 4, the effect of calcium in the fusion me-dium on induction of oocyte activation and development ofAWC-DSH-cloned embryos was determined. The presence ofcalcium in the fusion medium induced a high incidence of cleav-age of DSH oocytes (54.3%), while oocyte cleavage frequencywas much lower in the absence of calcium (16.6%). The pres-ence or absence of calcium in the fusion medium did not affectthe fusion, cleavage, and blastocyst development of AWC-DSH-cloned embryos. In experiment 5, AWC-DSH-cloned embryoswere transferred to the uteri of 11 synchronized domestic catrecipients on Day 6 or 7 after oocyte aspiration. Recipients wereassessed by ultrasonography on Day 21 postovulation, but nopregnancies were observed. In the present study, after NT, AWCdonor nuclei were able to dedifferentiate in DSH cytoplasts andsupport high rates of blastocyst development in vitro. Incom-plete reprogramming of the differentiated nucleus may be a ma-jor constraint to the in vivo developmental potential of the em-bryos.
assisted reproductive technology, early development, gamete bi-ology
INTRODUCTION
Most of the 36 species of wild felids are classified asthreatened, vulnerable or endangered [1]. As an example,the African wild cat (Felis silvestris libica) is one of thesmallest wild cats, whose future is threatened by hybrid-ization with domestic cats (Felis silvestris catus). Muchprogress has been made toward applying assisted reproduc-tive technologies for aiding in the conservation of endan-gered felids. In particular, nuclear transfer (NT) is a valu-able tool for retaining genetic variability [2, 3] and offersthe possibility of species continuation rather than extinc-tion. Interspecies NT into bovine, rabbit, and sheep cyto-plasts has been shown to support development of somaticcell nuclei from various mammalian species [4–8]. Accord-ingly, ooplasm of the domestic cat is a potential host forsomatic cell nuclei from endangered felid species.
During normal fertilization and embryo development,mitochondrial DNA (mtDNA) is maternally inherited [9].Animals produced by NT are genetic duplicates of the do-nor cell nuclear genome. Because foreign mitochondria areintroduced into the oocyte, some degree of heteroplasmycan occur after NT [10], which, according to speculation,may be detrimental. There are some species-specific epi-genetic factors present in the oocyte cytoplasm that maylead to nucleo:cytoplasmic incompatibilities either imme-diately after NT or at later stages of development [11].

1033NUCLEAR TRANSFER INTO DOMESTIC CAT OOCYTES
These potential incompatibilities will affect, to some de-gree, the ultimate utility of NT technology. It has been dem-onstrated that mtDNA present in the oocyte cytoplasmplays an important role in nuclear cytoplasmic incompati-bilities [12] and affects embryo implantation [13]. Even so,offspring have been produced from embryos reconstructedby interspecies [5, 7, 8], cross-subspecies not expressingheteroplasmy [14], and cross-subspecies expressing heter-oplasmy [15] nuclear transfer. These results demonstratedthe possibility that, through nuclear transfer techniques, theooplasm of domestic species could be used to preserve ge-netic diversity of closely related endangered species. Re-cently, the feasibility of producing viable felid offspring bycloning has been demonstrated [16]. Although the ratio ofNT kittens produced to total embryos transferred was sim-ilar to that reported in other species, the importance of in-creasing the efficiency of felid NT is obvious.
The cell-cycle phase of the donor nucleus plays a majorrole in the effectiveness of the NT process and has multipleinfluences on embryo reconstruction [17]. In the first reporton production of a live offspring from somatic cells, donornuclei were in a quiescent stage (G0) [18], where the cellswere presumably arrested after having exited the normalcell cycle. Although, somatic cell cloning has beenachieved with donor cells in G1 and G2/M phase [19, 20],for NT, it is generally accepted that better efficiency is ob-tained when the donor nucleus is in the G1 or G0 phase. Toachieve cell-cycle synchronization at this desired phase,cultured cells may be subjected to a period of serum star-vation before NT. A disadvantage of this method is theincrease in DNA fragmentation that occurs after prolongedculture of donor cells [21], where the extent of the damageincreases with the duration of the starvation period [22].Alternatively, donor cells can be arrested in the G0/G1phase by contact inhibition [23] or by exposure to revers-ible cycle inhibitors such as roscovitine [24].
Fibroblasts and cumulus cells of the domestic cat havean inherently long G0/G1 phase [25]. Serum starvation hasbeen found to increase the percentage of both of these celltypes at the desired stage; however, a notable portion un-dergoes apoptosis [25]. Therefore, additional stimuli wouldincrease the percentage of properly synchronized cells. Fur-thermore, the dislodging process should be optimized toreduce effects on cell characteristics such as size and DNAintegrity because viable single cells with intact DNA areessential for successful nuclear transfer.
In the present study, we determined the ability of so-matic cell nuclei of the African wild cat to dedifferentiatein domestic cat cytoplasts and support early in vitro devel-opment after reconstruction. Accordingly, our objectiveswere (1) to compare the effect of different methods, en-zymatic and mechanical, for obtaining single viable cellsafter dissociation; (2) to determine the effect of differentcell-cycle inhibition treatments on the distribution of cellsin the various phases of the cell cycle; (3) to evaluate theeffect of different methods of cell-cycle synchronizationand type of oocyte maturation (in vivo vs. in vitro) on re-construction and in vitro development of domestic and Af-rican wild cat embryos derived by NT; (4) to determine ifthe presence of calcium in the fusion medium (a) inducesconcurrent oocyte activation and (b) affects in vitro devel-opment of African wild cat embryos derived by NT; and(5) to evaluate developmental competence after transferinto domestic cat recipients of NT embryos produced byfusion of African wild cat fibroblast cell nuclei with do-mestic cat cytoplasts.
MATERIALS AND METHODS
ChemicalsAll chemicals were obtained from Sigma Chemical Co. (St Louis, MO)
unless otherwise stated. Media were prepared weekly and filtered (0.22 m,#4192 Acrodisc; Pall Corp., Ann Arbor, MI) into sterile 15-ml tubes. Afterfiltration, bicarbonate-buffered culture media were gassed with 5% CO2 inair before storage at 48C and used within 7 days. Any use of trade, product,or firm names is for descriptive purposes only and does not imply en-dorsement by the U.S. government.
AnimalsAll animal procedures were approved by the Institutional Animal Care
and Use Committee of the Audubon Nature Institute for Research of En-dangered Species as required by the Health Research Extension Act of1985 (Public law 99-158). Domestic cat females were group housed in anenvironmentally controlled room with 14L:10D at 20–268C. The Africanwild cat male was housed in a separate room under the same conditions.The rooms were cleaned, and cats were fed (Science Diet, Hill Pet Nutri-tion, Topeka, KS) daily. Fresh water was available at all times.
Establishment and Culture of Donor FibroblastsCell lines were generated from skin tissue collected by biopsy from a
male African wild cat (AWC) and a female domestic shorthaired cat(DSH). The tissue was washed twice in Ca21 and Mg21-free DulbeccoPBS (PBS; Invitrogen, Grand Island, NY) and finely cut into 1-mm2 piec-es. Tissue pieces were plated in 75-cm2 tissue-culture flasks (Nunc, Den-mark) containing 7 ml of Dulbecco modified Eagles medium (DMEM)supplemented with 50 mg/ml gentamicin and 10% (v/v) fetal bovine serum(FBS; Hyclone, Logan, Utah), and cultured at 388C in 5% CO2/air. After7–10 days of incubation, monolayer outgrowths with fibroblastic-like mor-phology were disaggregated with either 2.5 mg/ml of pronase or 2.5 mg/ml of trypsin, then regrown and passaged 2–3 times before being resus-pended in DMEM with 10% FBS and 10% (w/v) dimethyl sulfoxide(DMSO) and cooled at 1.08C/min to 2808C (Mr. Frosty; Nalgene, Roch-ester, NY) before storage in liquid nitrogen.
Cell Treatments and Flow Cytometric AnalysisDistributions of AWC and DSH fibroblasts in the cell-cycle phases
were assessed by flow cytometric analysis between cells disaggregatedwith pronase, trypsin, or mechanical separation (experiment 1). Also, com-parisons of the cell-cycle distribution of AWC and DSH fibroblasts weremade between cycling cells, after serum starvation, after being cultured toconfluence, and after exposure to roscovitine (experiment 2). Flat-sidedtissue culture tubes (Nunc, Denmark) were plated with frozen/thawedAWC or DSH fibroblasts at a concentration of 1 3 105 cells.
Experiment 1. AWC and DSH cells were cultured until reaching 100%confluence (2–3 days) and were disaggregated with 2.5 mg/ml trypsin, 2.5mg/ml pronase, or by mechanical separation before fixation. Cell viabili-ties in each treatment were determined by staining with 0.4% trypan blue[26].
Experiment 2. AWC and DSH cells were cultured for 1 day and allo-cated to one of the following treatments in which culture continued until(1) confluency to 60%–70% (cycling cells); (2) confluency to 100%, fol-lowed by an additional 5 days in culture, during which time the culturemedium was replaced every other day (contact inhibition); (3) confluencyto 60%–80%, replacement of tissue culture medium with DMEM 1 0.5%FBS and 5 days additional culture (serum starved); and (4) confluency to100%, culture medium replaced with DMEM containing 15 mM roscovi-tine and culture for an additional 24 h (roscovitine). Cells from each treat-ment were disaggregated with 2.5 mg/ml pronase prior to fixation.
Cells from each treatment were pelleted by centrifugation, then washedonce with PBS, and stained with equal volumes of 0.112% sodium citratecontaining 50 mg/ml propidium iodide (PI), 100 mg/ml RNAse A, and0.1% (v/v) Triton X-100 for 20 min at 388C. Stained cells were filteredthrough a 30-mm nylon mesh (Small Parts, Miami Lakes, FL) and thedistribution of nuclei in the various phases of the cell cycle was analyzedwith a flow cytometer (FACScan; Becton Dickinson ImmunocytometrySystems, San Jose, CA [BDIS]). Nuclei were analyzed at a rate of 300per sec, and 5–10 K events collected in listmode files with FL2H linearset at 340, at 1024 channels, using linear size and scatter parameters anddoublet discrimination mode. Histograms, dot plots, and density plots weregenerated by using CellQuest software (BDIS). Percentages of cells oc-

1034 GOMEZ ET AL.
curring in the G0/G1, S, and G2/M phase of the cell cycle were calculatedthrough the ModFit LT (Verity Software House, Topsham, ME) and esti-mated with CellQuest software (BDIS).
Oocyte MaturationCat ovaries obtained after ovariohysterectomy from local veterinary
clinics were transported to the laboratory at ambient temperature in Hepesbuffered saline containing 50 mg/ml gentamicin. Within 2–6 h, the ovarieswere minced and cumulus-oocyte complexes (COCs) collected. Oocyteswith uniformly dark, finely granulated ooplasm surrounded by several lay-ers of compact cumulus cells [27] were cultured in modified TCM-199containing 1 IU/ml human chorionic gonadotropin (hCG), 0.5 IU/mlequine chorionic gonadotropin (eCG), 10 mg/ml epidermal growth factor(EGF), and 3 mg/ml BSA for 24 h in 5% CO2, 5% O2, and 90% N2 at388C [28].
The protocol for ovarian stimulation and laparoscopic oocyte recoveryto obtain in vivo-matured oocytes from donor females has been describedpreviously [29]. Briefly, domestic cats 1–7 yr of age were treated with atotal of 3–5 IU of porcine-follicle stimulating hormone (FSH; Sioux Bio-chemical, Sioux Center, IA) administered daily (s.c.) for 4 days, followedby 3 IU of porcine-luteinizing hormone (LH, Sioux Biochemical) on thefifth day. Twenty-four to 26 h after LH injection (i.m.), oocytes werecollected by laparoscopic aspiration of mature ovarian follicles into TLHepes medium supplemented with 10 IU/ml heparin (sodium salt, fromporcine intestinal mucosa; Steris Laboratories, Phoenix, AZ) and 50 mg/ml gentamicin.
Cumulus cells of in vivo- and in vitro-matured oocytes were removedby vortexing in 1 mg/ml of hyaluronidase for 5 min, followed by gentlemechanical pipetting. Denuded oocytes were placed in Tyrode solutioncontaining 1% MEM nonessential amino acids (NEAA), 3 mg/ml BSA(Fraction V, very low endotoxin; Serologicals, Norcross, GA) and supple-mented with 15 mM NaHCO3, 0.36 mM pyruvate, 2.2 mM calcium lac-tate, 1 mM glutamine, and 50 mg/ml gentamicin (IVC-1 medium) at 388Cin 5% CO2 until further use.
Nuclear Transfer and Embryo CultureAfrican wild cat and domestic cat fibroblasts were synchronized by
one of the following treatments: (1) contact inhibition, (2) serum starva-tion, or (3) roscovitine. A single cell, presumably synchronized in G0/G1,was introduced into the perivitelline space of each enucleated in vivo- orin vitro-matured oocyte to evaluate the effect of donor nucleus, cell syn-chronization, and oocyte maturation on the reconstruction and in vitrodevelopment of AWC-DSH- and DSH-DSH-cloned embryos (experiment3).
The NT procedure was performed by using a micromanipulator (ModelMMO-202D; Narishige Instrument, Tokyo, Japan) attached to an invertedmicroscope (Olympus IX-70, Olympus, Tokyo, Japan) equipped withHoffman modulation contrast optics and a temperature-controlled stage setat 378C (Olympus, Tokai Hit, Japan). Before enucleation, denuded M-IIoocytes were incubated for 15 min in Ca21- and Mg21-free modified Ty-rodes salt solution supplemented with 1% MEM nonessential amino acids,3 mg/ml BSA, 30 mM NaHCO3, 0.36 mM pyruvate, 1 mM glutamine,and 50 mg/ml gentamicin (ECM medium), 20 mg/ml Hoechst 33342 and20 mg/ml cytochalasin B (CCB). After incubation, oocytes were enucle-ated in ECM medium, in which NaHCO3 was reduced to 15 mM, and 15mM Hepes was added, along with 20 mg/ml of CCB and 2 mg/ml ofsucrose (330–340 mOsm, enucleation medium). The first polar body andapproximately 10% of the underlying cytoplasm were drawn into an enu-cleation pipette (outer diameter, 20 mM), with subsequent confirmation ofremoval of the metaphase spindle by epifluorescence microscopy. The timeof exposure to ultraviolet light was restricted to 1 sec.
A single AWC or DSH fibroblast cell (,20 mm in diameter), presum-ably synchronized in G0/G1 by one of the synchronization treatments, wasintroduced into the perivitelline space of the enucleated in vivo- or in vitro-matured oocyte. Fusion took place in fusion medium (0.3 M mannitol, 0.1mM Mg21, and 0.05 M Ca21) by placing each NT couplet between twostainless-steel electrodes attached to micromanipulators (LF-101; NepaGene, Tokyo, Japan). The distance between electrodes was 120 mm andmembrane fusion was induced by applying a 3-sec AC prepulse of 20V,1 MHz; followed by two 30-msec DC pulses of 240V/mm at intervals of0.5 sec. Pulses were delivered perpendicularly to the shared membranespace of the donor cell/cytoplasm. Following the fusion pulses, NT cou-plets were washed and cultured in IVC-1 medium, and after 30 min, fusionwas evaluated visually by confirming the presence or absence of the donorcell in the perivitelline space. To determine if the cell fusion method in-
duces concurrent oocyte activation, in vitro-matured DSH oocytes wereelectrically pulsed in fusion medium with the presence or absence of cal-cium by the same procedure as described above, and cultured for 2 daysto determine cleavage frequency. Also, AWC-DSH couplets reconstructedwith a single AWC fibroblast cell, presumably synchronized in G0/G1 byserum starvation treatment and introduced into the perivitelline space ofenucleated in vivo- or in vitro-matured oocytes, were electrically fused infusion medium with or without calcium (experiment 4).
Activation of the fused couplets (experiments 3 and 4) was performed2–3 h after fusion by placing the couplets between two electrodes in afusion chamber containing 3 ml of fusion medium and exposing them totwo 60-msec DC pulses of 120 V/mm. Then couplets were incubated in30-ml drops of IVC-1 medium supplemented with 10 mg/ml cycloheximideand 5 mg/ml CCB at 388C in 5% CO2 under mineral oil (4008; SageBioPharma, Bedminster, NJ) for 4 h. As controls for the activation pro-tocol, some denuded oocytes were parthenogenetically activated by thesame procedure as described for fused NT couplets.
Following activation, reconstructed couplets and parthenogeneticallyactivated oocytes were cultured in 500 ml of IVC-1 medium until Day 4,when reconstructed and parthenogenetic embryos were moved into 500 mlof Tyrode solution containing 1% NEAA, 2% MEM essential amino acids(EAA), 10% FBS, and supplemented as for IVC-1 medium (IVC-2 me-dium), and cultured in 5% CO2, 5% O2, 90% N2 at 388C up to the day ofembryo transfer or until Day 8.
Assessment of embryosThe number of cybrids and activated oocytes that cleaved was assessed
on Day 2 and on Day 4, further development (morula) was assessed vi-sually (light microscopy). The number of embryos developing to the blas-tocyst stage was determined on Days 7 and 8. Blastocyst cell numbers,both total and inner cell mass/trophectoderm cell (ICM/TPD) ratios, weredetermined by using a modification of a protocol previously described byWells [30]. Blastocysts were incubated for 1 h at 388C in 1 mg/ml ofHoechst 33342 stain solution. After incubation, blastocysts were exposedto 0.04% of Triton-X100 in PBS for 45–60 sec, and then cultured for 15min at 388C in 25 mg/ml of PI stain in water. Stained embryos were placedin a drop of mounting medium (glycerol; Hoechst 33342) on a clean glassmicroslide and cells were counted by epifluorescence microscopy [31].The ratio of ICM cells to TPD cells per embryo was determined by count-ing the number of blue (ICM) and red (TPD) cells stained by Hoechst andPI, respectively.
Embryo TransferExperiment 5. To evaluate the in vivo competence of the cloned em-
bryos, on Day 6 (n 5 3 recipients) and Day 7 or 8 (n 5 8 recipients),AWC-DSH-cloned embryos were transferred into the uteri of 11 gonad-otropin-treated (FSH/LH) domestic cat recipients on Day 6 or 7 after oo-cyte aspiration, as previously described [32]. For transfer, one uterine hornwas exteriorized through a 1.5-cm midventral incision and punctured witha sterile 16-gauge round-tipped, short bevel trocar. Embryos were aspiratedin ;50 ml of IVC-2 medium into a 14.5-cm tom-cat catheter (3.5 Fr.;Sherwood Medical, St Louis, MO), which was then threaded 4–5 cm intothe uterine lumen before depositing the embryos. To assess pregnancystatus, abdominal ultrasonography was done on recipient cats betweenDays 21 and 23 after ovulation (i.e., oocyte recovery).
Statistical AnalysesTwo and three-way ANOVA was used to analyze the data on cell-
cycle comparisons, one-way ANOVA to analyze the data on the numberof embryos that developed to premorula and morula stages, and three-wayANOVA to analyze the data for fusion, cleavage rate, and blastocyst de-velopment. Numbers of nuclei in blastocysts and in the ICM and TPDwere subjected to an arcsine transformation for each replication. The trans-formed values were analyzed by using one-way ANOVA on ranks. TheTukey multiple-comparison test was used to determine any differencesbetween two means after ANOVA. Values are reported as mean % 6standard error of the mean.
RESULTS
Experiment 1
Effect of disaggregation treatments on African wild catand domestic cat fibroblasts. The distribution of AWC nuclei

1035NUCLEAR TRANSFER INTO DOMESTIC CAT OOCYTES
TABLE 1. Percentages of African wild cat fibroblast cells in various phas-es of the cell cycle after culturing cells to confluence and cell separationby enzymatic or mechanical methods.
Dissagrega-tion method Na
Cell cycle phase (mean % 6 SEM)
G0/G1 S G2/M%
Aggregates
TrypsinPronaseMechanical
10 00010 00010 000
89 6 1b
93 6 0.4b
82 6 3c
4 6 0.22 6 0.30 6 0
7 6 1b
5 6 0.1b
18 6 3c
10b
2b
37c
a The total number of cells in each treatment was derived from threereplications.b,c Different superscripts within the same column indicate significant dif-ferences (P , 0.0001).
TABLE 2. Percentages of domestic cat fibroblast cells in various phasesof the cell cycle after culturing cells to confluence and cell separation byenzymatic or mechanical methods.
Dissagregationmethod Na
Cell cycle phase (mean % 6 SEM)
G0/G1 S G2/M
%Aggre-gates
TrypsinPronaseMechanical
11 33627 24741 931
74 6 3b
80 6 4c
87 6 2d
14 6 28 6 19 6 2
12 6 312 6 24 6 2
2b
5b
15c
a The total number of cells in each treatment was derived from at leastthree replications.b,c,d Different superscripts within the same column indicate significant dif-ferences (P , 0.0001).
FIG. 1. Typical histograms of DNA, ob-tained by using flow cytometry of domes-tic cat fibroblast cells cultured under a va-riety of cell-cycle inhibitors.
in the various phases of the cell cycle indicated that trypsinand pronase (89.0%, 93.0%) yielded higher proportions ofcells in the G0/G1 phase than did mechanical separation(82.0%; Table 1). The proportions were reversed for DSHnuclei, as the mechanical separation method yielded higherproportions of cells in the G0/G1 phase (87.0%) comparedwith that from trypsin (74.2%) and pronase (79.7%; Table2) treatment. However, in both AWC and DSH cells, cellaggregates were more prevalent after mechanical disaggre-gation (37.3%, 15.0%) than after trypsin (10.5%, 1.7%) orpronase (1.8%, 4.8%, respectively, P , 0.05; Tables 1–2)treatment, respectively. Cell-viability testing indicated thattrypsin and pronase treatments yielded higher percentages ofviable cells for AWC (85%–90%) and DSH (90%–95%)cells compared with mechanical separation (AWC 5 35%,DSH 5 50%; P , 0.05). Overall, these findings suggest thatpronase treatment yielded cells with higher viability and lesscellular and DNA damage (data not shown) and gave a high-er percentage of cells in the G0/G1 phase. Based on theseresults, pronase disaggregation was chosen as the preferredmethod for cell preparation prior to flow cytometric analysisor nuclear transfer in all further experiments.
Experiment 2
Effect of cell-cycle inhibitor treatments on the distribu-tion of cells in the various phases of the cell cycle. Flowcytometry analyses revealed that most AWC and DSH fi-broblast cells were in the G0/G1 phase, as shown by theprominent G0/G1 DNA peaks (Figs. 1, 2), and that the dy-namics of the cell cycle varied as culture conditions weremodified (Fig. 3). Higher percentages of AWC and DSHcells were in the G0/G1 phase after serum starvation com-pared with cycling cells, or after contact inhibition and ros-covitine treatments (P , 0.05; Figure 3). In contrast, cy-cling cultures contained higher percentages of cells in theS and G2/M phases than the other treatments. For both spe-cies, serum starvation resulted in lower percentages of cellsin the S and G2/M phases compared with contact inhibitionand roscovitine. Cycling AWC cells had a lower proportionof cells in the G0/G1 phase (50%) compared with cyclingDSH cells (64%; P , 0.05). Although AWC cells weresynchronized in the G0/G1 phase by all synchronizationtreatments, proportions of cells in G0/G1 were lower thanthe proportion of DSH cells synchronized in G0/G1 in eachtreatment (P , 0.05).

1036 GOMEZ ET AL.
FIG. 2. Typical histograms of DNA, ob-tained by using flow cytometry of Africanwild cat fibroblast cells cultured under avariety of cell-cycle inhibitors.
Experiment 3
Effect of donor nucleus, cell synchronization, and oo-cyte maturation on the reconstruction and development ofAWC-DSH- and DSH-DSH-cloned embryos. In this exper-iment, evaluations were made on the potential for in vitrodevelopment of NT embryos produced from in vitro- andin vivo-matured oocytes reconstructed with DSH or AWCcells from each cell-cycle synchronization treatment. Blas-tocyst development was influenced by the donor nucleusbut was not affected by the cell-cycle synchronizationmethod (Table 3). Higher percentages of blastocysts wereobserved (assessed as a percentage of cleaved couplets)when AWC donor cells were used compared with DSHcells (24.2% vs. 3.3%, respectively; P , 0.05; Table 3).The method of cell synchronization did not affect the fusionand cleavage rates. A slightly higher percentage of oocytescleaved when cells synchronized by serum starvation(83.0%) were used as the donor nucleus compared withroscovitine (80.0%) or contact-inhibition (80.0%), althoughthere were no statistical differences between the treatments.Despite the high percentage of embryos cleaving after NTin all treatments, 18% of AWC-DSH and 40% of DSH-DSH ‘presumptive’ embryos had undergone fragmentationand an intact donor nucleus was still present. The remainderof the ‘presumptive’ embryos did cleave (2–4 cells), withmost NT embryos arresting at the premorula stage (2–10cells) stage. There was not a statistically significant differ-ence between treatments. Indeed, only 14% of embryos re-constructed with DSH cells reached the morula/blastocyststage compared with 44% of embryos reconstructed withAWC cells (P , 0.05).
The fusion efficiency of in vivo- and in vitro-maturedoocytes used as recipient cytoplasts with AWC cells as do-
nor nuclei was not different (86.6% vs. 85.2%, respective-ly). Domestic cat cells (DSH) transferred into in vivo-ma-tured oocytes showed a similar fusion efficiency (83.7%)to AWC cells, but fusion efficiency of NT couplets usingin vitro-matured oocytes was slightly lower (78.0%), butnot statistically different (Table 3). Likewise, the cleavagerate of embryos reconstituted with DSH cells and in vitro-matured oocytes was numerically lower (73.0%) but notsignificantly different from the other groups.
The percentage of blastocysts developing after parthe-nogenetic activation of in vivo- and in vitro-matured oo-cytes was 23.3 6 2 and 27.4 6 5, respectively. The rate ofblastocyst development in embryos reconstructed withAWC cells and in vivo- (27.2 6 3) or in vitro- (23.1 63.2)matured oocytes was higher than that of embryos recon-structed with DSH cells and in vivo- (3.0 6 4) or in vitro-(2.3 6 3) matured oocytes (P , 0.05).
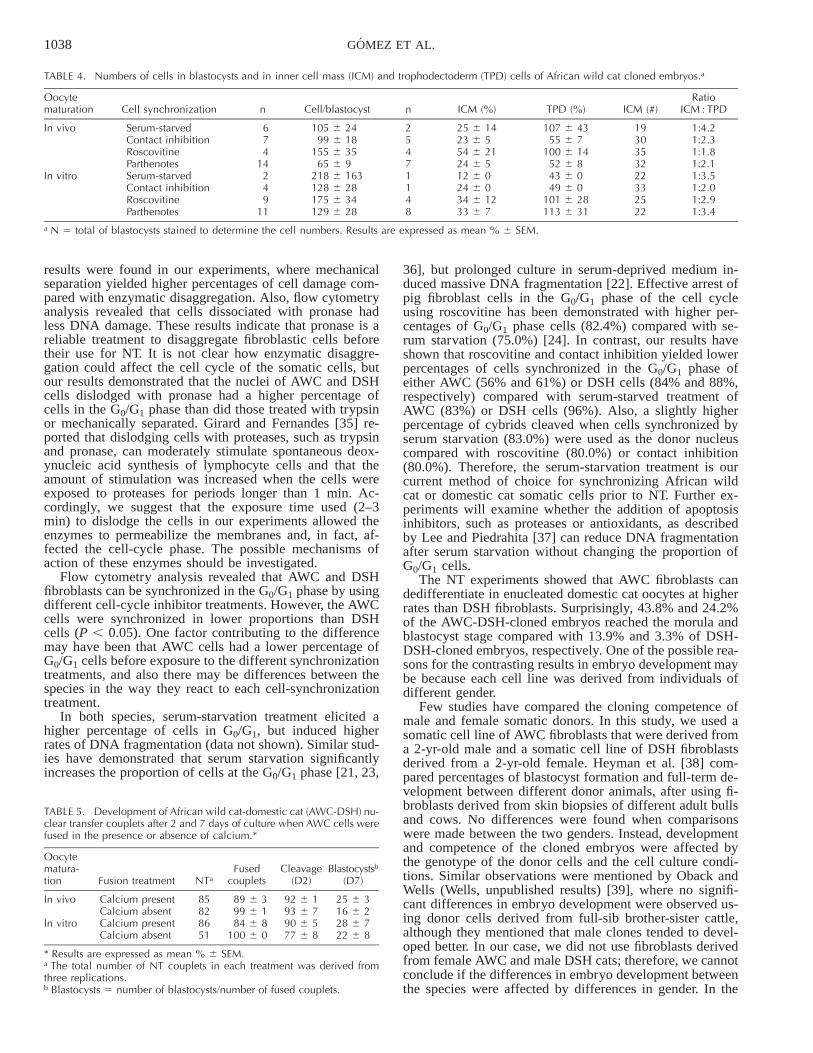
The total cell number in NT blastocysts was not affectedby the cell-cycle synchronization method or by type of oo-cyte,- and was similar to the cell number of the parthenotes(Table 4). The ICM cell numbers and the ratios of ICM/TPD cells were similar between treatments. We were notable to compare the total cell number of blastocysts derivedfrom the DSH cell line because of the low number of blas-tocysts obtained.
Experiment 4
Effect of calcium in the fusion medium on domestic catoocyte activation and development of AWC-DSH-clonedembryos. In this experiment, we evaluated the effect of cal-cium in the fusion medium on a) activation of domestic catoocytes and b) embryo development of AWC-DSH-clonedembryos.

1037NUCLEAR TRANSFER INTO DOMESTIC CAT OOCYTES
FIG. 3. Percentages of African wild catand domestic cat fibroblasts existing in thevarious phases of the cell cycle after treat-ment with different cell-cycle inhibitors.
TABLE 3. Effect of donor nucleus, cell synchronization, and oocyte maturation of the recipient cytoplast on in vitro development of DSH-DSH andAWC-DSH cloned embryos.a
Donornucleus
Oocytematuration
Donor nucleustreatment NT
Fusedcouplets Cleavage Premorulae Morulae Blastocysts
AWC In vivo Serum-starvedContact inhibitionRoscovitine
1435255
89 6 289 6 1081 6 8
89 6 382 6 980 6 9
85 6 869 6 962 6 22
38 6 6ab
35 6 7ab
47 6 17ab
28 6 3a
17 6 6a
33 6 14a
In vitro Serum-starvedContact inhibitionRoscovitine
787779
79 6 880 6 497 6 2
79 6 987 6 788 6 6
75 6 1978 6 483 6 9
46 6 13ab
42 6 3ab
51 6 9a
21 6 9a
28 6 4a
20 6 4a
DSH In vivo Serum-starvedContact inhibitionRoscovitine
354856
88 6 267 6 296 6 3
83 6 986 6 884 6 8
40 6 469 6 1976 6 12
4 6 2b
10 6 5ab
11 6 2ab
3 6 5b
2 6 1b
4 6 3b
In vitro Serum-starvedContact inhibitionRoscovitine
6010449
78 6 1070 6 586 6 7
79 6 774 6 567 6 17
44 6 339 6 1293 6 6
8 6 5b
9 6 3b
22 6 13ab
0 6 0b
3 6 2b
3 6 3b
a The total number of NT couplets in each treatment was derived from at least three replications. Results are expressed as mean % 6 SEM. Developmentrate of embryos at different stages, number of embryos/number of fused couplets.a,b Different superscripts within the same column indicate significant differences (P , 0.05). DSH, Domestic cat; AWC, African wild cat.
The presence of calcium in the fusion medium inducedcleavage in slightly more than half of the domestic cat oo-cytes (25/465 54.3%), while the percentage of cleaved oo-cytes in the absence of calcium was much lower (4/26516.6%; P , 0.05). Although the presence of calcium in thefusion medium induced activation of domestic cat oocytes,the fusion rate, cleavage rate, and blastocyst developmentof AWC-DSH-cloned embryos were not affected by thepresence of calcium in the fusion medium (Table 5).
Experiment 5
Embryo transfer of AWC-DSH-cloned embryos into do-mestic cat recipients. Derived AWC-DSH-cloned embryoswere transferred into 11 domestic cat recipients. Fifty-threemorulae (Day 6) were transferred to three of the recipientson Day 6 after oocyte recovery (mean 5 17.6), and 57blastocysts (Days 7 and 8), and 66 morulae (Day 7) weretransferred to eight of the recipients on Day 7 after oocyterecovery (mean 5 17.5 embryos). No pregnancies weredetected after abdominal ultrasonography on Days 21–23after oocyte recovery.
DISCUSSION
In the present study, we compared the ability of somaticcell nuclei of the African wild cat and the domestic cat todedifferentiate in domestic cat cytoplasts and to supportearly development after reconstruction. Specifically, we ex-amined the effect of type of oocyte maturation (in vivo vs.in vitro), different methods (mechanical and enzymatic) fordonor cell isolation, and cell synchronization treatments(serum starvation, contact inhibition, and roscovitine) onthe distribution of cells in the various phases of the cellcycle and the effect of calcium in the cell fusion mediumon embryo reconstruction after NT.
The cell dislodging process can affect cell viability andDNA integrity. Both mechanical and enzymatic disaggre-gation have been used to dislodge cells cultured in vitro.Mechanical separation re-suspends cells more quickly thanenzymatic digestion but may cause mechanical damage[33]. Enzymatic disaggregation using trypsin is the mostwidely used enzyme for tissue disaggregation, but can in-duce DNA damage [33], and is less effective at disaggre-gating fibroblastic cell lines than is pronase [34]. Similar
1038 GOMEZ ET AL.
TABLE 4. Numbers of cells in blastocysts and in inner cell mass (ICM) and trophodectoderm (TPD) cells of African wild cat cloned embryos.a
Oocytematuration Cell synchronization n Cell/blastocyst n ICM (%) TPD (%) ICM (#)
RatioICM : TPD
In vivo Serum-starvedContact inhibitionRoscovitineParthenotes
674
14
105 6 2499 6 18
155 6 3565 6 9
2547
25 6 1423 6 554 6 2124 6 5
107 6 4355 6 7
100 6 1452 6 8
19303532
1:4.21:2.31:1.81:2.1
In vitro Serum-starvedContact inhibitionRoscovitineParthenotes
249
11
218 6 163128 6 28175 6 34129 6 28
1148
12 6 024 6 034 6 1233 6 7
43 6 049 6 0
101 6 28113 6 31
22332522
1:3.51:2.01:2.91:3.4
a N 5 total of blastocysts stained to determine the cell numbers. Results are expressed as mean % 6 SEM.
TABLE 5. Development of African wild cat-domestic cat (AWC-DSH) nu-clear transfer couplets after 2 and 7 days of culture when AWC cells werefused in the presence or absence of calcium.*
Oocytematura-tion Fusion treatment NTa
Fusedcouplets
Cleavage(D2)
Blastocystsb
(D7)
In vivo Calcium presentCalcium absent
8582
89 6 399 6 1
92 6 193 6 7
25 6 316 6 2
In vitro Calcium presentCalcium absent
8651
84 6 8100 6 0
90 6 577 6 8
28 6 722 6 8
* Results are expressed as mean % 6 SEM.a The total number of NT couplets in each treatment was derived fromthree replications.b Blastocysts 5 number of blastocysts/number of fused couplets.
results were found in our experiments, where mechanicalseparation yielded higher percentages of cell damage com-pared with enzymatic disaggregation. Also, flow cytometryanalysis revealed that cells dissociated with pronase hadless DNA damage. These results indicate that pronase is areliable treatment to disaggregate fibroblastic cells beforetheir use for NT. It is not clear how enzymatic disaggre-gation could affect the cell cycle of the somatic cells, butour results demonstrated that the nuclei of AWC and DSHcells dislodged with pronase had a higher percentage ofcells in the G0/G1 phase than did those treated with trypsinor mechanically separated. Girard and Fernandes [35] re-ported that dislodging cells with proteases, such as trypsinand pronase, can moderately stimulate spontaneous deox-ynucleic acid synthesis of lymphocyte cells and that theamount of stimulation was increased when the cells wereexposed to proteases for periods longer than 1 min. Ac-cordingly, we suggest that the exposure time used (2–3min) to dislodge the cells in our experiments allowed theenzymes to permeabilize the membranes and, in fact, af-fected the cell-cycle phase. The possible mechanisms ofaction of these enzymes should be investigated.
Flow cytometry analysis revealed that AWC and DSHfibroblasts can be synchronized in the G0/G1 phase by usingdifferent cell-cycle inhibitor treatments. However, the AWCcells were synchronized in lower proportions than DSHcells (P , 0.05). One factor contributing to the differencemay have been that AWC cells had a lower percentage ofG0/G1 cells before exposure to the different synchronizationtreatments, and also there may be differences between thespecies in the way they react to each cell-synchronizationtreatment.
In both species, serum-starvation treatment elicited ahigher percentage of cells in G0/G1, but induced higherrates of DNA fragmentation (data not shown). Similar stud-ies have demonstrated that serum starvation significantlyincreases the proportion of cells at the G0/G1 phase [21, 23,
36], but prolonged culture in serum-deprived medium in-duced massive DNA fragmentation [22]. Effective arrest ofpig fibroblast cells in the G0/G1 phase of the cell cycleusing roscovitine has been demonstrated with higher per-centages of G0/G1 phase cells (82.4%) compared with se-rum starvation (75.0%) [24]. In contrast, our results haveshown that roscovitine and contact inhibition yielded lowerpercentages of cells synchronized in the G0/G1 phase ofeither AWC (56% and 61%) or DSH cells (84% and 88%,respectively) compared with serum-starved treatment ofAWC (83%) or DSH cells (96%). Also, a slightly higherpercentage of cybrids cleaved when cells synchronized byserum starvation (83.0%) were used as the donor nucleuscompared with roscovitine (80.0%) or contact inhibition(80.0%). Therefore, the serum-starvation treatment is ourcurrent method of choice for synchronizing African wildcat or domestic cat somatic cells prior to NT. Further ex-periments will examine whether the addition of apoptosisinhibitors, such as proteases or antioxidants, as describedby Lee and Piedrahita [37] can reduce DNA fragmentationafter serum starvation without changing the proportion ofG0/G1 cells.
The NT experiments showed that AWC fibroblasts candedifferentiate in enucleated domestic cat oocytes at higherrates than DSH fibroblasts. Surprisingly, 43.8% and 24.2%of the AWC-DSH-cloned embryos reached the morula andblastocyst stage compared with 13.9% and 3.3% of DSH-DSH-cloned embryos, respectively. One of the possible rea-sons for the contrasting results in embryo development maybe because each cell line was derived from individuals ofdifferent gender.
Few studies have compared the cloning competence ofmale and female somatic donors. In this study, we used asomatic cell line of AWC fibroblasts that were derived froma 2-yr-old male and a somatic cell line of DSH fibroblastsderived from a 2-yr-old female. Heyman et al. [38] com-pared percentages of blastocyst formation and full-term de-velopment between different donor animals, after using fi-broblasts derived from skin biopsies of different adult bullsand cows. No differences were found when comparisonswere made between the two genders. Instead, developmentand competence of the cloned embryos were affected bythe genotype of the donor cells and the cell culture condi-tions. Similar observations were mentioned by Oback andWells (Wells, unpublished results) [39], where no signifi-cant differences in embryo development were observed us-ing donor cells derived from full-sib brother-sister cattle,although they mentioned that male clones tended to devel-oped better. In our case, we did not use fibroblasts derivedfrom female AWC and male DSH cats; therefore, we cannotconclude if the differences in embryo development betweenthe species were affected by differences in gender. In the

1039NUCLEAR TRANSFER INTO DOMESTIC CAT OOCYTES
present study, because the type of recipient cytoplasts, thecell synchronization, and the nuclear transfer method wereessentially identical, the observed differences in develop-ment may be considered to be due in part to the origin ofthe donor nucleus. Underscoring this statement is that noconclusive reports have yet been published indicating thatmale somatic cells are influencing better embryo develop-ment after NT.
Significant effects have been reported on source of re-cipient cytoplast used for NT with lower rates of blastocystproduction and fetal survival from in vitro-matured cyto-plasts compared with in vivo-matured cytoplasts [40]. In-stead, in our NT experiments, the fusion efficiency, embryocleavage, blastocyst development, and total cell number inNT blastocysts were not affected by the source of oocytesused as recipient cytoplasts. We had previously demonstrat-ed in our laboratory a reduction in cleavage frequency andin vitro development of cat oocytes matured in vitro andfertilized by intracytoplasmic sperm injection comparedwith that of in vivo-matured oocytes [21]. However, mod-ification of the IVM medium by the addition of epidermalgrowth factor has improved both the number and quality ofembryos produced [28, 41]. We can suggest that, althoughthe IVM system is not fully optimized, improvements madein recent years to our IVM medium have improved thequality of in vitro-matured cytoplasts, making them com-parable with in vivo-matured cytoplasts in terms of embryodevelopment. The influence of source of recipient cyto-plasts needs consideration with regard to pregnancy rateand embryo survival following the transfer of NT-derivedembryos.
Direct comparisons of NT success between laboratoriesare not possible because of variations in the type of cells,embryo culture system, and NT procedures. The rates ofblastocyst development of domestic cat embryos recon-structed with cumulus cells (2.6%), granulose cells (3.0%),and domestic cat fetal fibroblasts (5.2%) [42, 43] were sim-ilar to the present results using DSH somatic cells (3.3%).A possible explanation for the early developmental failureof most of our reconstructed embryos may be inadequateor incomplete reprogramming of donor nuclei.
Kim et al. [44] suggest that failure of activated oocytesbefore NT to remodel somatic nuclei is caused by the ab-sence of cytoplasmic factors, such as maturation promotingfactor (MPF), that are needed to induce nuclear envelopebreakdown (NEBD). Because the electrical pulse necessaryto induce fusion of the donor cell to the recipient cytoplastduring NT can induce concurrent oocyte activation and atransient decrease of the maturation promoting factor activ-ity, chromatin remodeling may be affected. In fact, pig cy-brids that were fused in medium containing 0.1 mM ofcalcium and not receiving further chemical activation stim-ulus were activated and cleaved at higher rates (69%) thantheir counterparts fused under calcium-free conditions(10%) [45]. Similarly, in our study, a higher percentage ofoocytes cleaved after receiving electrical pulses in the pres-ence of calcium (54.3%) compared with those electricallypulsed in the absence of calcium (16.6%). These resultsindicate that domestic cat oocytes are susceptible to acti-vation during electrical fusion in the presence of calcium.However, the fusion of AWC-DSH couplets in the presenceor absence of calcium did not affect the fusion rates norembryo development to the blastocyst stage. Several stud-ies have shown that the level of active MPF declines sig-nificantly in oocytes within 2 h after electrical activationand remains depleted for at least 8–10 h, directly affecting
chromatin remodeling [46, 47]. We did not measure levelsof histone H1 kinase activity to determine the level of ac-tive MPF, but the proportions of couplets undergoing chro-matin remodeling after fusion in medium containing cal-cium is similar to those couplets fused in the absence ofcalcium. Therefore, we suggest that the levels of MPF afterfusion in the presence of calcium do not decline drasticallywithin the next 2–3 h, which was the delayed activationinterval we used before activating the reconstructed cou-plets. Because we do not know the extent to which thecalcium concentration (0.05 mM) in the fusion medium isaffecting the level of MPF activity during fusion, furtherstudies should be done to investigate the level of histoneH1 kinase activity.
In our NT experiments, we used oocytes of similar typeand source that were at the same meiotic stage (M-II, non-activated) when reconstructed. Only 18.0% of the AWC-DSH presumptive embryos were fragmented and were un-able to remodel the chromatin compared with 40.0% ofDSH-DSH presumptive embryos. From these results, it ap-pears that chromatin remodeling failure may be partiallydue to the recipient cytoplast, but there are likely otherfactors involved as well. Such unknown factors may havecontributed to the lower rate of chromatin remodeling inthe DSH somatic cell line relative to the remodeling ratein the AWC somatic cell line. Differences in the epigeneticstatus of the DSH somatic nuclei may be a cause of thelow rate of chromatin remodeling after NT [48]. A betterunderstanding of the molecular basis for epigenetic varia-tion within the donor cells will help to understand its influ-ence during cell reprogramming.
During the initial stages of development, embryos areunder the control of maternally derived proteins and tran-scripts accumulated in the oocyte. The timing of the tran-sition from maternal to embryonic control of development(MET) and the beginning of the transcription period is spe-cies specific [49]. For example, in the mouse, embryonicgenome activation and transcription begin at the 1-cellstage [50], whereas in domestic cats, the transition frommaternal to embryonic control of development and embry-onic transition occurs by the 5–8-cell stage [51]. Therefore,the possibility of a relationship between the high rate ofdevelopment failure of both DSH-DSH (40.0%) and AWC-DSH (37.5%) NT embryos at the 8–10-cell stage and thetransition from maternal to embryonic control of develop-ment cannot be overlooked. In spite of a similar rate ofearly development failure in embryos reconstructed fromeach of the two somatic cell lines, the embryos derivedfrom AWC somatic cells developed in vitro to the blasto-cyst stage (24.2%) at rates comparable with domestic catembryos produced by parthenogenetic activation of in vivo-(23.3%) and in vitro-matured (27.4%) oocytes or by intro-cytoplasmic sperm injection (ICSI) from in vivo- (29.5%)and in vitro-matured (19.0%) oocytes [29].
Although the AWC-DSH-cloned embryos developed tothe blastocyst stage in vitro, we were not able to producea pregnancy after embryo transfer. Interspecies and subspe-cies embryo transfer of nondomestic cat embryos into do-mestic cat recipients has resulted in the birth of Indian de-sert cat [52] and African wild cat kittens [53] and an earlypregnancy of interspecies panda-rabbit cloned embryos [6],demonstrating that the domestic cat is not only a model fordevelopment of in vitro assisted reproductive technologies,but it can also serve as a successful recipient of embryosfrom closely related small non-domestic cats. Therefore, wedo not think that species incompatibility contributed to the

1040 GOMEZ ET AL.
developmental failure of the AWC-DSH embryos trans-ferred into the domestic cat. More likely, an incompletereprogramming of the differentiated nucleus was a majorconstraint to the in vivo developmental potential of theseembryos [54]. Also, the influence of possible mitochondrialheteroplasmy and/or the deleterious effect of mtDNA of thederived AWC cloned embryos on preimplantation devel-opment should not be discounted. Nagao et al. [55] havedemonstrated that mouse embryos derived from nucleartransfer of interspecific species (M. musculus and M. spre-tus) had reduced development in vitro and decreased phys-ical performances after birth. Conversely, the birth of nor-mal Bos indicus calves derived from nuclear transfer of Bosindicus donor cells into Bos taurus oocyte recipients dem-onstrated that cross-subspecies nuclear transfer is a viableapproach to rescue closely related endangered species [14,15]. Meirelles et al. [14] demonstrated that donor cell-de-rived mtDNA (Bos indicus) was reduced during early em-bryo development and completely eliminated by the end ofthe gestation. However, Steinborn et al. [15] found coex-isting mtDNAs of both subspecies (heteroplasmy) inhealthy cloned Bos indicus calves. Both studies indicatedthat the presence of mtDNA derived from the recipient oo-cyte or mtDNAs derived from both recipient oocyte anddonor-cell (heteroplasmy) does not inhibit normal devel-opment. Although in our study we did not evaluate themitochondrial inheritance of the derived AWC cloned em-bryos, we suggest that it should be possible to establish apregnancy with cross-species- (DSH and AWC) derivedcloned embryos after transfer to surrogate domestic cats.Indeed, further research is needed to evaluate the precisemitochondrial inheritance pattern of AWC-cloned embryosderived by cross-subspecies nuclear transfer and its influ-ence on embryo implantation and survival rate.
In conclusion, the results of the current study indicatethat (1) African wild cat and domestic cat cells can be syn-chronized in the G0/G1 phase by different methods, (2) adistinct difference in in vitro developmental competence ofNT embryos produced from each of the two cat fibroblastcell lines, and (3) incomplete reprogramming of the differ-entiated nucleus may be a major constraint to the in vivodevelopmental potential of these embryos.
ACKNOWLEDGMENT
M.C.G. is grateful to Dr. Teija Peura for the training in somatic cellculture and nuclear transfer procedures. We are also most appreciative ofDr. Kazuchika Miyoshi for indicating how to use the electrofusion ma-chine, Jackie Coulon for coordinating and collecting ovaries from the vet-erinary clinics, Dr. Alex Cole and his veterinary staff for their help withthe surgery procedures, and Stella Sullivan and Kelly Trimble for theirexcellent cat care.
REFERENCES
1. Nowell P, Jackson P. Major issues in cat conservation. In: Status Sur-vey and Conservation Action Plan: Wild Cats. Gland, Switzerland:IUCN; 1996: 149–179.
2. Seidel GE Jr. Cloning, transgenesis, and genetic variance in Animals.Cloning Stem Cells 2001; 3:251–256.
3. Ryder OA. Cloning advances and challenges for conservation. TrendsBiotechnol 2002; 20:231–232.
4. Dominko T, Mitalipova M, Haley B, Beyhan Z, Memili E, McKusickB, First NL. Bovine oocyte cytoplasm supports development of em-bryos produced by nuclear transfer of somatic cell nuclei from variousmammalian species. Biol Reprod 1999; 60:1496–1502.
5. Lanza RP, Cibelli JB, Diaz F, Moraes CT, Farin PW, Farin CE, Ham-mer CJ, West MD, Damiani P. Cloning of an endangered species (Bosgaurus) using interspecies nuclear transfer. Cloning 2000; 2:79–90.
6. Chen DY, Wen DC, Zhang YP, Sun QY, Han ZM, Liu ZH, Shi P, Li
JS, Xiangyu JG, Lian L, Kou ZH, Wu YQ, Chen YC, Wang PY,Zhang HM. Interspecies implantation and mitochondria fate of panda-rabbit cloned embryos. Biol Reprod 2002; 67:637–642.
7. White KL, Bunch TD, Mitalipov S, Reed WA. Establishment of preg-nancy after the transfer of nuclear transfer embryos produced fromthe fusion of argali (Ovis ammon) nuclei into domestic Sheep (Ovisaries) enucleated oocytes. Cloning 1999; 1:47–54.
8. Loi P, Ptak G, Barboni B, Fulka J Jr, Cappai P, Clinton M. Geneticrescue of an endangered mammal by cross-species nuclear transferusing post-mortem somatic cells. Nature 2001; 19:962–964.
9. Cummins JM. Mitochondria: potential roles in embryogenesis and nu-cleocytoplasmic transfer. Hum Reprod 2001; 7:217–228.
10. Steinborn R, Schinogl P, Zakhartchenko V, Achmann R, SchernthanerW, Stojkpvic M, Wolf E, Muller M, Brem G. Mitochondrial DNAheteroplasmy in cloned cattle produced by fetal and adult cell cloning.Nat Genet 2000; 25:255–257.
11. Latham KE. Epigenetic modification and imprinting of the mamma-lian genome during development. Curr Top Dev Biol 1999; 43:1–49.
12. Yamazaki Y, Meirelles FV, Bordignon V, Garcia JM, Smith LC. Effectof mitochondrial DNA on developmental outcome of cytoplast-recon-structed mouse zygotes. Arq Fac Vet UFRGS 1999; 27:(abstract 299).
13. Nagao Y, Totsuka Y, Atomi Y, Kaneda H, Yonekawa H, Takahashi S,Imai H. Heterogeneous mitochondria DNA introduced by nucleartransfer influences the developmental ability of mouse embryos invitro. Theriogenology 1997; 47:(abstract 233).
14. Meirelles FV, Bordignon V, Watanabe Y, Watanabe M, Dayan A, LoboRB, Garcia JM, Smith LC. Complete replacement of the mitochondrialgenotype in a Bos indicus calf reconstructed by nuclear transfer to aBos taurus oocyte. Genetics 2001; 158:351–356.
15. Steinborn R, Schinogl P, Wells DN, Bergthaler A, Muller M, BremG. Coexistence of Bos Taurus and B. indicus mitochondrial DNAs innuclear transfer-derived somatic cattle clones. Genetics 2002; 162:823–829.
16. Shin T, Kraemer D, Pryor J, Liu L, Rugila J, Howe L, Buck S, Mur-phy K, Lyons L, Westhusin MA. Cat cloned by nuclear transplanta-tion. Nature 2002; 415(6874):859.
17. Campbell KH, Loi P, Otaegui Pj, Wilmut I. Cell cycle co-ordinationin embryo cloning by nuclear transfer. Rev Reprod 1996; 1:40–46.
18. Wilmut I, Schnieke AE, McWhir J, Kind AJ, Campbell KH. Viableoffspring derived from fetal and adult mammalian cells. Nature 1997;385:810–813.
19. Cibelli JB, Stice SL, Golueke PJ, Kane JJ, Jerry J, Blackwell C, Poncede Leon FA, Robl JM. Cloned transgenic calves produced from non-quiescent fetal fibroblasts. Science 1998; 280:1256–1258.
20. Ono Y, Shimozawa N, Ito M, Kono T. Cloned mice from fetal fibro-blast cells arrested at metaphase by a serial nuclear transfer. Biol Re-prod 2001; 64:44–50.
21. Kues WA, Anger M, Carnwath JW, Paul D, Motlik J, Niemman H.Cell cycle synchronization of porcine fetal fibroblasts: effects of serumdeprivation and reversible cell cycle inhibitors. Biol Reprod 2000; 62:412–419.
22. Peura TT. Serum starvation can cause excessive DNA damage insheep fetal fibroblasts. Theriogenology 2001; 55:(abstract 285).
23. Boquest AC, Billiy ND, Prather RS. Flow cytometric cell cycle anal-ysis of cultured porcine fetal fibroblast cells. Biol Reprod 1999; 60:1013–1019.
24. Arat S, Gibbons J, Rzucidlo SJ, Miyoshi K, Venable A, WalterburgR, Stice S. Bovine cloning using adult donor cells treated with ros-covitine. Biol Reprod 2001; 64(suppl):173 (abstract 171).
25. Bochenek M, Katska L, Kania G, Rynska B, Smorag Z, PienkowskiM. Cell cycle analysis of somatic cell lines in the cloning of thedomestic cat. Theriogenology 2001; 55:(abstract 256).
26. Shapiro H. Practical Flow Cytometry. New York: Wiley-Liss; 1993.27. Pope CE, McRae MA, Plair BL, Keller GL, Dresser BL. In vitro and
in vivo development of embryos produced by in vitro maturation andin vitro fertilization of cat oocytes. J Reprod Fertil 1997; 51:69–82.
28. Gomez MC, Pope CE, Davis AM, Harris RF, Dresser BL. Additionof epidermal growth factor (EGF) during in vitro maturation of do-mestic cat oocytes enhances fertilization frequency and blastocyst de-velopment in vitro. Theriogenology 2001; 5:(abstract 472).
29. Gomez MC, Pope CE, Harris R, Davis A, Mikota S, Dresser BL.Births of kittens produced by intracytoplasmic sperm injection of do-mestic cat oocytes matured in vitro. Reprod Fertil Dev 2000; 12:423–433.
30. Wells KD. Efficient stain for differential staining of trophectodermand inner cell mass cells. In: Proceedings of Genetically Engineering

1041NUCLEAR TRANSFER INTO DOMESTIC CAT OOCYTES
and Cloning Animals: Science, Society and Industry; 2000; CedarCity, UT. Abstract 12.
31. Harris RF, Gomez MC, Pope CE, Giraldo AM, King AL, Dresser BL.Differential staining of trophectoderm and inner cell mass cells of catin vitro derived blastocysts. Theriogenology 2003; 59(abstract 448).
32. Pope CE, Mcrae MA, Plair BL, Keller GL, Dresser BL. Successfulin vitro and in vivo development of in vitro fertilized two- to four-cell cat embryos following cryopreservation, culture and transfer. The-riogenology 1994; 42:513–25.
33. Freshney RI. Disaggregation of the tissue and primary culture. In:Freshney RI (eds.), Culture of Animal Cells: A Manual of Basic Tech-nique. New York: Wiley-Liss & Sons, Inc., Publisher; 1994:127–147.
34. Foley JH, Aftonomos B. The use of pronase in tissue culture: a com-parison with trypsin. J Cell Physiol 1970; 75:159–162.
35. Girard JP, Fernandes B. Studies on the mitogenic activity of trypsin,pronase and neuraminidase on human peripheral blood lymphocytes.Eur J Clin Invest 1976; 6:347–353.
36. Kasinathan P, Knott JG, Wang Z, Jerry DJ, Robl JM. Production ofcalves from G1 fibroblasts. Nature 2001; 19:1176–1178.
37. Lee CK, Piedrahita JH. Inhibition of apoptosis in serum starved por-cine embryonic fibroblasts. Mol Reprod Dev 2002; 62:106–112.
38. Heyman Y, Zhou QI, Lebourhis D, Chavatte-Palmer P, Renard JP,Vignon X. Novel approaches and hurdles to somatic cloning in cattle.Cloning Stem Cells 2002; 4:47–55.
39. Oback B, Wells D. Donor cells for nuclear cloning: many are called,but few are chosen. Cloning Stem Cells 2002; 4:147–168.
40. Wells DN, Misica PM, Day TA, Tervit HR. Production of clonedlambs from an established embryonic cell line: a comparison betweenin vivo- and in vitro-matured cytoplasts. Biol Reprod 1997; 57:385–393.
41. Gomez MC, Pope E, Harris R, Mikota S, Dresser B. Development ofin vitro matured, in vitro fertilized domestic cat embryos followingcryopreservation, culture and transfer. Theriogenology 2003; 60:239–251.
42. Kitiyanant Y, Saikhun J, Pavasuthipaisit K. Somatic nuclear transferin domestic cat oocytes treated with IGF-I for in vitro maturation.Theriogenology 2003; 59:1775–1786.
43. Skrzyszowska M, Katska L, Rynska B, Kania G, Smorag Z, Pien-kowski M. In vitro developmental competence of domestic cat em-
bryos after somatic cloning: a preliminary report. Theriogenology2002; 58:1615–1621.
44. Kim JM, Ogura A, Nagata M, Aoki F. Analysis of the mechanism forchromatin remodeling in embryos reconstructed by somatic nucleartransfer. Biol Reprod 2002; 67:760–766.
45. Boquest AC, Grupen CG, Harrison SJ, Mcllfatrick SM, Ashman RJ,d’Apice AJ, Nottle MB. Production of cloned pigs from cultured fetalfibroblast cells. Biol Reprod 2002; 66:1283–1287.
46. Grupen CG, Mau JC, McIlfatrick SM, Maddocks S, Nottle MB. Effectof 6-dimethylaminopurine on electrically activated in vitro maturedporcine oocytes. Mol Reprod Dev 2002; 62:387–396.
47. Campbell KHS, Ritchie WA, Wilmut I. Nuclear-cytoplasmic interac-tions during the first cell cycle of nuclear transfer reconstructed bovineembryos: implications for deoxyribonucleic acid replication and de-velopment. Biol Reprod 1993; 49:933–942.
48. Surani A. Reprogramming of genome function through epigenetic in-heritance. Nature 2001; 414:122–128.
49. Telford NA, Watson AJ, Schultz GA. Transition from the maternal toembryonic control in early mammalian development: a comparison ofseveral species. Mol Reprod Dev 1990; 26:90–100.
50. Bouniol C, Nguyen E, Devey P. Endogenous transcription occurs atthe 1-cell stage in the mouse embryos. Exp Cell Res 1995; 218:57–62.
51. Hoffert KA, Anderson GB, Wildt DE, Roth TL. Transition from ma-ternal to embryonic control of development in IVM/IVF domestic catembryos. Mol Reprod Dev 1997; 48:208–215.
52. Pope CE, Gelwicks EJ, Keller GL, Dresser BL. In vitro fertilizationin domestic and nondomestic cats including sequences of early nuclearevents, in vitro development cryopreservation and successful intra andinterspecies embryo transfer. J Reprod Fertil 1993; 47:189–201.
53. Pope CE, Gomez MC, Mikota SK, Dresser BL. Development of invitro produced African wildcat (Felis silvestris) embryos after cryo-preservation into domestic cat recipients. Biol Reprod 2000; 62(suppl1):321 (abstract 544).
54. Kikyo N, Wolffe AP. Reprogramming nuclei: insights from cloning,nuclear transfer and heterokaryons. J Cell Sci 2000; 113:11–20.
55. Nagao Y, Totsuka Y, Atomi Y, Kaneda H, Lindahl KF, Imai H, Yone-kawa H. Decreased physical performance of congenic mice with mis-match between the nuclear and the mitochondrial genome. Genes Gen-et Syst 1998; 73:21–27.




















