
Transport and metabolism of -lactate in Jerusalem artichoke mitochondria

REGULAR ARTICLE
Mycorrhizal colonization impacts on phenolic contentand antioxidant properties of artichoke leavesand flower heads two years after field transplant
Nello Ceccarelli & Maurizio Curadi &Luca Martelloni & Cristiana Sbrana &
Piero Picciarelli & Manuela Giovannetti
Received: 10 March 2010 /Accepted: 29 April 2010 /Published online: 18 May 2010# Springer Science+Business Media B.V. 2010
Abstract Greenhouse and field experiments werecarried out in order to investigate the influence ofmycorrhizal inoculation on total phenolic content(TPC) and antioxidant activity, expressed as antirad-ical power (ARP), of artichoke (Cynara cardunculusL. var. scolymus F.) leaves and flower heads extracts.The establishment of mycorrhizal symbiosis wasmonitored in pot and field grown plants, and thepersistence of the inoculated AMF in roots after2 years’ growth in the field was assessed by fungalITS sequencing. Both in the greenhouse and in thefield, marked increases in TPC and ARP were detectedin leaves and flower heads of artichoke plants inoculatedwith the AM fungal speciesGlomus intraradices, eitheralone or in mixture with Glomus mosseae. In the field,plants inoculated with Glomus mix showed flower
heads ARP content increases of 52.7 and 30.0% in thefirst and second year, respectively, compared withuninoculated plants. After 2 years’ growth in the fieldITS rDNA sequences clustering with those of G.mosseae and G. intraradices were retrieved only frominoculated plant roots. Our data show that mycorrhizalinoculation may represent an efficient and sustainablestrategy to improve productivity and enhance plantbiosynthesis of secondary metabolites with healthpromoting activities.
Keywords Artichoke (Cynara cardunculus L. var.scolymus) . Arbuscular mycorrhizal fungi (AMF) .
Glomus sp. . Flower heads . Phenolic content .
Antioxidant properties
Introduction
Arbuscular mycorrhizal (AM) symbioses are mutual-istic associations between soilborne fungi belongingto the phylum Glomeromycota and the roots of about80% of land plant species, including the mostimportant agricultural crops. AM fungi (AMF) playa key role in soil fertility and plant nutrition,enhancing the uptake and translocation of mineralnutrients—mainly P, N, S, K, Ca, Fe, Cu and Zn—from soil to host plants, by means of an extensivebelowground hyphal network, which spreads from
Plant Soil (2010) 335:311–323DOI 10.1007/s11104-010-0417-z
Responsible Editor: Angela Hodge.
N. Ceccarelli :M. Curadi : L. Martelloni : P. Picciarelli :M. Giovannetti (*)Department of Crop Plant Biology, University of Pisa,Viale delle Piagge 23,56124 Pisa, Italye-mail: [email protected]
C. SbranaInstitute of Biology and Agrobiotechnology, CNR,UOS Pisa,Via del Borghetto 80,56124 Pisa, Italy

colonised roots into the soil environment (Smith andRead 2008; Giovannetti and Avio 2002).
AMF colonization of root cortical cells producesseveral cytological and metabolic changes, such as amarked proliferation of plastids during arbusculeformation (Fester et al. 2001; Hans et al. 2004; Lohseet al. 2005). The activation of plastid biosyntheticpathways and Krebs cycle leads to increased meta-bolic activity and higher production of amino acids,fatty acids and apocarotenoids, linked to the forma-tion of symbiotic structures (Lohse et al. 2005). AMsymbiosis influences primary and secondary metabo-lism of host plants (Schliemann et al. 2008): itinduces important changes both in enzymatic activi-ties (i.e. superoxide dismutase and catalase) (Ruiz-Lozano et al. 1996; Marin et al. 2002) and inphysiological mechanisms leading to the accumula-tion of secondary metabolites, such as carotenoids andpolyphenols (Walter et al. 2000; Lambais et al. 2003;Fester et al. 2005; Marulanda et al. 2007; Toussaintet al. 2007). Some authors reported higher levels ofreactive oxygen species (ROS) in colonized roots, andsuggested that mycorrhizal plants respond to oxida-tive stresses by the accumulation of antioxidativeenzymes and carotenoids (Fester and Hause 2005).
Artichoke (Cynara cardunculus L. var. scolymusFiori) is a poliennal plant species (Asteraceae), nativeto the Mediterranean region and largely cultivated inItaly, which is the main world producer (FAOSTAT2007). Artichoke leaves are primarily used in thepharmaceutical industry, while immature flower headsrepresent a good dietary source of nutraceuticalcompounds, mostly polyphenolics with marked anti-oxidant activity (hydroxycinnamates and flavonoids)(Kraft 1997; Gebhardt 1997; Wang et al. 2003; Curadiet al. 2005). Artichoke leaves extracts possesshypocholesterolemic activity, mainly due to flavo-noids, such as luteolin that inhibits the HMG-CoA (3-hydroxy 3-methyl-glutaryl-CoA) reductase (Gebhardt1997). Antioxidant effects of artichoke extracts arelinked to free radicals scavenging activities ofmono- and di-caffeoylquinic acids (chlorogenicacid, cynarine and isomers); in particular, a stronginhibitory activity on LDL (low density lipoprotein)oxidation was reported for chlorogenic acid (Brownand Rice-Evans 1998). Owing to such biologicalactivities, artichoke extracts are supposed to exert aprotective action to prevent carcinogenesis andatherosclerosis (Sawa et al. 1999; Bundy et al.
2008). Accordingly, polyphenolics can be consideredimportant factors to improve the quality profile andthe nutritional and nutraceutical value of artichokeflower heads.
To investigate the effects of AM symbiosis onpolyphenolic production, we carried out microcosmand field experiments with artichoke plants inoculatedwith the AMF species Glomus mosseae and Glomusintraradices and with a combination of them. Here weassessed (a) total phenolic content (TPC) and antiox-idant activity, expressed as antiradical power (ARP)(Brand-Williams et al. 1995) of leaf extracts frommycorrhizal artichoke plantlets grown in microcosm;(b) TPC and ARP of flower heads extracts fromartichoke plants previously inoculated in microcosmand grown for 1 and 2 years in the field; (c) biomassand flower head production of artichoke plants, 1 and2 years after field transplant. We also monitored theestablishment of mycorrhizal symbiosis in microcosmand field grown plants, and assessed, by fungal ITSsequencing, the persistence of the inoculated AMF inroots after 2 years’ growth in the field.
Materials and methods
Chemicals and standards
5-O-caffeoyl-D-quinic acid (chlorogenic acid, CA),2,2-diphenyl-1-picrylhydrazyl (DPPH), Folin-Ciocalteu reagent and 1 ml semi-micro polystyrenecuvets were purchased from Sigma-Aldrich S.r.l.(Italy). Sodium carbonate (Na2CO3) was obtainedfrom J.T.Baker (Deventer, Holland). All the solventsused were double-distilled and HPLC grade.
Plant and fungal material
Eighty artichoke (C. cardunculus var. scolymus)offshoots were obtained from mother plants of thelate cv. Terom, cultivated in the field in the Depart-ment of Crop Plant Biology at S. Piero a Grado, Pisa,Italy. Young offshoots were excised on March 2005,and the leaves cut at about 30 cm of length. The AMfungal species used were G. mosseae (Nicol. andGerd.) Gerdemann and Trappe (isolate AZ 225C fromUSA, collector J.C. Stutz) and G. intraradicesSchenck and Smith (isolate IMA6 from France,collector V. Gianinazzi-Pearson). They were obtained
312 Plant Soil (2010) 335:311–323

from Helianthus annuus L. pot cultures maintained inthe collection of the Soil Microbiology Laboratory ofthe Department of Crop Plant Biology, University ofPisa, Italy.
Greenhouse experiment
This experiment was aimed at assessing TPC and ARPin leaves of artichoke plants inoculated with the twoAMfungal species and a mixture of them (Glomus mix).
Experimental setup Eighty microcosms were set up in7 l plastic pots containing a sandy soil which wassteam-sterilised (121°C for 40 min) to kill naturallyoccurring AMF. Each pot contained 1 artichokeoffshoot. Chemical and physical characteristics ofthe soil were as follows: pH (H2O), 8.1; clay; 4.9%,silt, 6.1%; sand, 88.9%; total N, 0.9 g kg−1; availableP (NaHCO3 soluble P, Olsen analysis) 4.15 mg kg−1,127 mg kg−1 available K. The experiment was acompletely randomized design with four inoculumtreatments (each fungal isolate, their mix and uninoc-ulated control), 20 replicates and 3 harvests. Inmycorrhizal treatments, each offshoot was inoculatedwith 50 g of crude inoculum (mycorrhizal roots andsoil containing spores and extraradical mycelium)originated from pot-cultures of each AM fungalspecies (G. mosseae and G. intraradices) or with50 g of a crude inoculum containing a mixture of bothendophytes (1:1, by volume) (Glomus mix). All thepots received 100 ml of a filtrate, obtained by sievinga mixture of the two inocula through a 50 μm porediameter sieve, to ensure a common microflora for alltreatments. Each pot was watered by a timer-controlled drip irrigation system. The experimentwas carried out under a PVC greenhouse withoutlateral walls.
Measurements Three randomly selected plants fromeach treatment were harvested after 30, 60 and 90 daysof growth. The following growth parameters wereassessed on each plant: fresh weight (FW) of thewhole plant, plant height (cm), root length (cm), rootFW, number of leaves, total leaf area (mm2), leaf dryweight (DW). DW was determined after drying at 70°Cfor 3 days. Percentage of AM colonized root length wasassessed on randomly collected 30 g of root samples,after root staining, by using the gridline intersect method(Giovannetti and Mosse 1980). Stained roots were
observed under a dissecting microscope and selectedcolonised root pieces were mounted on microscopeslides and observed under a Reichert-Jung (Austria)Polyvar light microscope to detect intraradical fungalstructures.
At the last harvest leaf samples were collected andstored at −20°C for TPC and ARP analyses of the leafextracts (see below).
Leaf extracts preparation Leaf samples (2.5 g FW)were collected from 6 plants per treatment, frozen inliquid nitrogen, hand crushed in a mortar, added with20 ml of 80%MeOH, and extracted at 4°C for 12 h on astirrer. After extraction, the plant material was centri-fuged at 4,000 rpm for 15 min, and the supernatantcollected and stored at 4°C in darkness. The extractionwas repeated three times, and the final volume of theextracts was 62.5 ml (=40 mg FW ml−1).
Field experiment
This experiment was aimed at assessing TPC and ARPof artichoke flower heads from 1-year and 2-year-fieldgrown plants, previously inoculated in pot.
Experimental setup At the end of the greenhouseexperiment, artichoke plants were trasplanted fromthe pots to the field. Chemical characteristics of thefield soil were as follows: pH (H2O), 7.5; NO3,332 mg kg−1; NH4, 9 mg kg−1; available P (P2O5),136 mg kg−1; exchangeable K (K2O), 487 mg kg−1.The field soil was a loam soil according to the USDAclassification (clay, 21.1%; silt, 40.2%; sand, 38.7%).The soil was harrowed before artichoke transplant ina single row (October 2005), with a distance ofabout 90 cm between plants. All the plants of asingle treatment were grouped together and followedby the plants of another treatment. Two g/plant of aternary fertilizer (NPK: 11-22-16) was distributedafter transplant. Weed control was carried outmanually. Offshoots were removed in spring andautumn.
Measurements Artichoke roots were sampled in Oc-tober 2007, after 2 years’ growth in the field, to assessmycorrhizal colonization. Duplicate samples (in the10–30 cm soil depth) from three different plant rootsystems for each treatment were collected by using asoil sampler (Eijkelkamp Agrisearch Equipment, NL).
Plant Soil (2010) 335:311–323 313

In order to determine total biomass production forthe different treatments, during the first and secondyear in the field, all offshoots produced were removedand fresh weights obtained. Main, secondary andtertiary flower heads produced by each treatment wereharvested at maturity, cleaned and fresh weightsobtained to assess total flower head production. Theflower heads were then stored at −20°C until analysedfor TPC and ARP. Taking into account that mostcommercial production of artichoke flower heads isrepresented by secondary heads, during the secondyear in the field TPC and ARP analyses were carriedout on the extracts obtained from such material. Threereplicates per treatment were analysed.
Molecular analysis of field root samples Samples ofrandomly collected fine roots were prepared from soilsampled from each plant treatment (12 samples, 1 mgeach). Roots were rinsed in sterile water and werepowdered by grinding in liquid nitrogen. DNAextraction was performed by using a DNeasy PlantMini Kit (Qiagen, USA) following manufacturer’srecommendation. To amplify ITS region the nestedPCR technique described by Renker et al. (2003) wasused. Briefly, the first PCR step was performed withthe primer pair SSU-Glom1 (ATT ACG TCC CTGCCC TTT GTA CA) and LSU-Glom1 (CTT CAATCG TTT CCC TTT CA), on a Eppendorf Master-cycler® personal thermal cycler (Eppendorf, Italy) ina total volume of 25 µl containing 0.125U of GoTaqFlexi DNA Polymerase (Promega, Italy), 1X Poly-merase reaction buffer (Promega), 0.2 mM (each)dNTPs, 1.5 mM MgCl2, 25 pmol of each of the twoprimers and 40–100 ng of the genomic DNA. Thethermal cycler was programmed as follows: a manual“hot start” at 94°C for 2 min, 16 touchdown cycles(30 s at 95°C, 30 s at 69 down to 55°C, 1 min at 72°C)and 15 fixed annealing temperature cycles (30 s at 95°C,30 s at 53°C, 1 min at 72°C), followed by a finalelongation of 10 min at 72°C. The product of first PCRstep was digested with the restriction enzyme AluI(5 µl product and 5U enzyme in a total volume of20 µl), purified with Montage PCR Centrifugal FilterDevices (Millipore, Italy) and diluted (1/10-100-500)to perform a nested PCR reaction. Dilutions (2 μl)were added as template, together with the ITS5/ITS4primer pair in the reaction mix described above. In thenested PCR the thermal cycler was programmed asfollows: initial denaturation at 95°C for 2 min, 7
touchdown cycles (45 s at 95°C, 45 s at 61 down to55°C, 1 min at 72°C) and 22 fixed annealingtemperature cycles (45 s at 95°C, 45 s at 55°C,1 min at 72°C), followed by a final elongation step of10 min at 72°C.
Amplified DNA fragments were purified by usingMontage PCR and quantified by a BioPhotometer(Eppendorf). Purified products were cloned intopGem®-T Easy vector according to the manufac-turer’s instructions (Promega). Putative positiveclones were screened by using standard SP6/T7amplifications, followed by a nested PCR usingITS1/ITS4, carried out as described above for ITS5/ITS4 PCR reaction. Amplification products (15clones from each root sample) were tested forrestriction fragment length polymorphism (RFLP)patterns by digesting the PCR products with HinfIand MboI (Takara, Wi, USA) restriction enzymes,according to manufacturer’s instructions, in order torecover sequence variability. Digested DNA waselectrophoresed through 2% MetaPhor agarose(BMA, ME USA) containing ethidium bromide(0.5 µg ml−1). A DNA Ladder (1 kb Plus, Invitrogen,Italy) was used as a molecular weight marker. DNAprofiles were acquired with the ImageMaster VDSsystem and analysed using ImageMaster 1D Elitesoftware (Amersham Bioscience, Italy). Twenty-sixclones containing recombinant plasmids with differentHinfI and MboI patterns were purified by Wizard®Plus SV Minipreps (Promega) and sequenced at BMRGenomics s.r.l. (Italy).
DNA sequences of the full ITS region weresubmitted to the European Molecular Biology Labo-ratory database under the accession numbers FN658690-92, FN 658987-91, FN 666277-82.
The glomeromycotan origin of the sequencesobtained in this study was verified by BLAST andby phylogenetic analysis of 5.8S-subunit sequences(Redecker et al. 1999; Hempel et al. 2007), whichwere aligned with those retrieved from NCBI data-base using the software ClustalW resulting in align-ments including 154 putatively homologous sites. Aphylogenetic tree was performed using Treecon forWindows version 1.3b, with the Neigbor-joining algo-rithm and setting the Kimura model to 2 (data notshown). The confidence of branching was assessedusing 1,000 bootstrap resamplings. Species separationfound in the 5.8S analysis was also confirmed by wholeITS regions alignments (data not shown).
314 Plant Soil (2010) 335:311–323

Determination of total phenolic compounds (TPC)
TPC was measured by means of the Folin-Ciocalteumethod previously described by Shadidi and Naczk(1995). Five g FW of leaves or edible parts (1 g fromeach of 5 artichoke heads collected from 5 differentplants) were extracted with 80% MeOH. The edibleparts were prepared by removing the outer fibrousbracts, then frozen with liquid nitrogen, homogenisedand extracted (3 times) with 80% MeOH for 12 h at4°C. The methanolic extracts were pooled and centri-fuged at 4,000g for 20 min. Raw extracts were storedin the dark at −20°C until analysis. Extract volume was125 ml (40 mg FW ml−1). Different diluitions of the80% MeOH extracts (0.25 ml) were mixed with0.25 ml of Folin-Ciocalteu reagent (diluted withdistilled H2O 1:1 v/v), 0.5 ml of a saturated Na2CO3
solution (20%) and 4 ml of H2O. The reaction mixtureswere left at room temperature for 30 min. Absorbanceat 725 nm (A725) was measured using a Hitachi U-3200spectrophotometer (Hitachi Ltd, Tokyo, Japan). Eachreplicate was analysed twice. Extract dilutions corre-sponded to 10, 5, 2.5, 1.25, 0.625, 0.250, 0.0625 mgFW. A calibration curve was obtained by mixingdifferent quantities of chlorogenic acid (CA) standard(0, 1, 5, 10, 15, 20, 25, 30, 35, 40, 50, 75 and 100 μg)with 0.25 ml of Folin-Ciocalteu reagent. TPC wasfinally expressed as mg CA equivalents g−1 FW.
Determination of antiradical power (ARP)
The antioxidant activity of the extracts expressed asantiradical power (ARP) was measured by means of the2,2-Diphenyl-1-pikrylhydrazyl (DPPH) test (Brand-Williams et al. 1995). In this assay the antioxidantactivity is monitored by measuring the reduction of thestable free radical DPPH by antioxidants or radicalspecies which results in loss of its absorbance at515 nm (A515). For DPPH analysis, a 153 μM stocksolution of DPPH in 80% MeOH was used. Analyseswere carried out using a Hitachi U-3200 spectropho-tometer. A standard curve was obtained measuring theA515 of 9 dilutions of DPPH (153, 125, 100, 80, 75, 60,50, 25, 10 μM). Analyses were performed by adding950 μl of a 153 μM DPPH solution to 50 μl of dilutedraw extract or methanolic CA standard solution in 1 mlpolystyrene cuvets. Extract dilutions were prepared bymixing 37.50, 31.75, 25.00, 18.75, 12.50, 6.25 μl ofeach extract with H2O up to a final volume of 50 μl.
To assess the ARP of the CA standard, the followingamounts were used: 0.5, 1, 2, 5, 10, 20, 30 μg. Valuesof A515 were read against a blank of 80% MeOHwithout DPPH. Each replicate sample per treatmentwas analysed twice. A515 of the samples was immedi-ately read after mixing DPPH with the raw extract(0 min = start reaction), then after 30, 60, 120, 180,240, 360, 480 min and 24 h, until reaction reached aplateau. After each reading, cuvets were stored inthe dark at room temperature. Different molarratios (moles of TPC, expressed as CA equiva-lents, per mole of DPPH) were tested. The exactinitial DPPH concentration (C0DPPH) of each samplewas calculated using the following linear regressionequation (C0DPPH ¼ 7:9910�5 � A515 þ 2:0610�7) asreported by Bondet et al. (1997). The percentage ofremaining DPPH was determined for each time intervaland the reaction kinetics for each molar ratio wereplotted (% of remaining DPPH against time). Based onthese reaction kinetics, the percentage of the remainingDPPH at the steady state was calculated and plottedagainst the molar ratios. The EC50 (molar ratiocorresponding to 50% of the remaining DPPH) wascalculated, and the antiradical power (ARP = 1/EC50)as defined by Brand-Williams et al. (1995) was thendetermined.
Statistical analysis
TPC and ARP data for every treatment are the mean of 3replicates (± SEM; n=3). TPC and ARP data weresubjected to one-way ANOVA analysis (P<0.05) and tounpaired t-test (interval of confidence = 95%) in order toassess the statistical significance of the within-treatmentand between-treatment differences. Statistical analysishas been performed using the GraphPad Prism 4software. Percentage root colonisation data were com-pared using one-way analysis of variance (ANOVA),with fungal treatment as factor, and submitted to arcsintransformations before analysis. Means and standarderrors reported in tables refer to untransformed data.
Results
Greenhouse experiment
Root colonization and plant growth parameters Rootcolonization showed significant differences among
Plant Soil (2010) 335:311–323 315
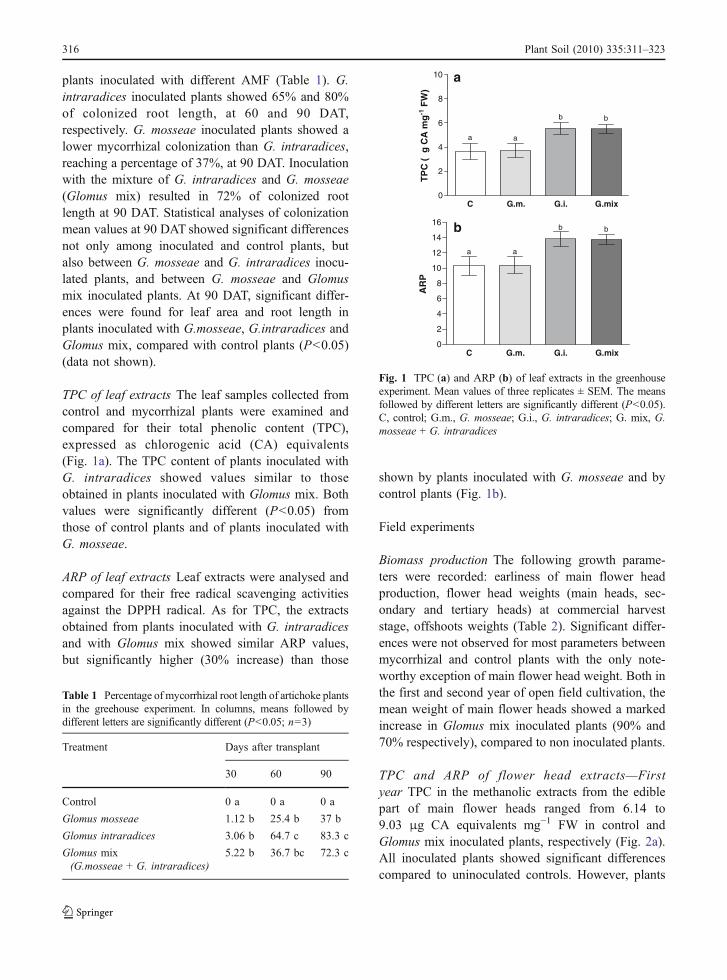
plants inoculated with different AMF (Table 1). G.intraradices inoculated plants showed 65% and 80%of colonized root length, at 60 and 90 DAT,respectively. G. mosseae inoculated plants showed alower mycorrhizal colonization than G. intraradices,reaching a percentage of 37%, at 90 DAT. Inoculationwith the mixture of G. intraradices and G. mosseae(Glomus mix) resulted in 72% of colonized rootlength at 90 DAT. Statistical analyses of colonizationmean values at 90 DAT showed significant differencesnot only among inoculated and control plants, butalso between G. mosseae and G. intraradices inocu-lated plants, and between G. mosseae and Glomusmix inoculated plants. At 90 DAT, significant differ-ences were found for leaf area and root length inplants inoculated with G.mosseae, G.intraradices andGlomus mix, compared with control plants (P<0.05)(data not shown).
TPC of leaf extracts The leaf samples collected fromcontrol and mycorrhizal plants were examined andcompared for their total phenolic content (TPC),expressed as chlorogenic acid (CA) equivalents(Fig. 1a). The TPC content of plants inoculated withG. intraradices showed values similar to thoseobtained in plants inoculated with Glomus mix. Bothvalues were significantly different (P<0.05) fromthose of control plants and of plants inoculated withG. mosseae.
ARP of leaf extracts Leaf extracts were analysed andcompared for their free radical scavenging activitiesagainst the DPPH radical. As for TPC, the extractsobtained from plants inoculated with G. intraradicesand with Glomus mix showed similar ARP values,but significantly higher (30% increase) than those
shown by plants inoculated with G. mosseae and bycontrol plants (Fig. 1b).
Field experiments
Biomass production The following growth parame-ters were recorded: earliness of main flower headproduction, flower head weights (main heads, sec-ondary and tertiary heads) at commercial harveststage, offshoots weights (Table 2). Significant differ-ences were not observed for most parameters betweenmycorrhizal and control plants with the only note-worthy exception of main flower head weight. Both inthe first and second year of open field cultivation, themean weight of main flower heads showed a markedincrease in Glomus mix inoculated plants (90% and70% respectively), compared to non inoculated plants.
TPC and ARP of flower head extracts—Firstyear TPC in the methanolic extracts from the ediblepart of main flower heads ranged from 6.14 to9.03 μg CA equivalents mg−1 FW in control andGlomus mix inoculated plants, respectively (Fig. 2a).All inoculated plants showed significant differencescompared to uninoculated controls. However, plants
Table 1 Percentage of mycorrhizal root length of artichoke plantsin the greehouse experiment. In columns, means followed bydifferent letters are significantly different (P<0.05; n=3)
Treatment Days after transplant
30 60 90
Control 0 a 0 a 0 a
Glomus mosseae 1.12 b 25.4 b 37 b
Glomus intraradices 3.06 b 64.7 c 83.3 c
Glomus mix(G.mosseae + G. intraradices)
5.22 b 36.7 bc 72.3 c
C G.m. G.i. G.mix0
2
4
6
8
10
a a
b b
TP
C (
µg C
A m
g-1
FW
)
a
C G.m. G.i. G.mix0
2
4
6
8
10
12
14
16
a a
b b
AR
P
b
Fig. 1 TPC (a) and ARP (b) of leaf extracts in the greenhouseexperiment. Mean values of three replicates ± SEM. The meansfollowed by different letters are significantly different (P<0.05).C, control; G.m., G. mosseae; G.i., G. intraradices; G. mix, G.mosseae + G. intraradices
316 Plant Soil (2010) 335:311–323
inoculated with Glomus mix or with G. intraradicesshowed a higher TPC value than plants inoculatedwith G. mosseae.
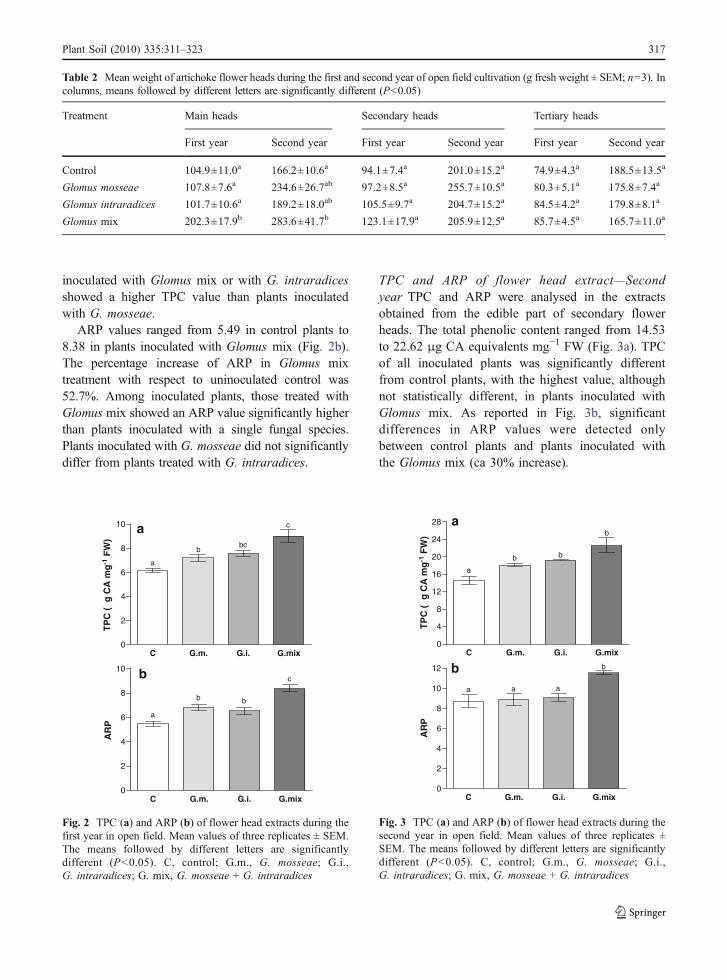
ARP values ranged from 5.49 in control plants to8.38 in plants inoculated with Glomus mix (Fig. 2b).The percentage increase of ARP in Glomus mixtreatment with respect to uninoculated control was52.7%. Among inoculated plants, those treated withGlomus mix showed an ARP value significantly higherthan plants inoculated with a single fungal species.Plants inoculated with G. mosseae did not significantlydiffer from plants treated with G. intraradices.
TPC and ARP of flower head extract—Secondyear TPC and ARP were analysed in the extractsobtained from the edible part of secondary flowerheads. The total phenolic content ranged from 14.53to 22.62 μg CA equivalents mg−1 FW (Fig. 3a). TPCof all inoculated plants was significantly differentfrom control plants, with the highest value, althoughnot statistically different, in plants inoculated withGlomus mix. As reported in Fig. 3b, significantdifferences in ARP values were detected onlybetween control plants and plants inoculated withthe Glomus mix (ca 30% increase).
Table 2 Mean weight of artichoke flower heads during the first and second year of open field cultivation (g fresh weight ± SEM; n=3). Incolumns, means followed by different letters are significantly different (P<0.05)
Treatment Main heads Secondary heads Tertiary heads
First year Second year First year Second year First year Second year
Control 104.9±11.0a 166.2±10.6a 94.1±7.4a 201.0±15.2a 74.9±4.3a 188.5±13.5a
Glomus mosseae 107.8±7.6a 234.6±26.7ab 97.2±8.5a 255.7±10.5a 80.3±5.1a 175.8±7.4a
Glomus intraradices 101.7±10.6a 189.2±18.0ab 105.5±9.7a 204.7±15.2a 84.5±4.2a 179.8±8.1a
Glomus mix 202.3±17.9b 283.6±41.7b 123.1±17.9a 205.9±12.5a 85.7±4.5a 165.7±11.0a
C G.m. G.i. G.mix0
4
8
12
16
20
24
28
a
b b
b
TP
C (µ
g C
A m
g-1
FW
)
a
C G.m. G.i. G.mix0
2
4
6
8
10
12
a a a
b
AR
P
b
Fig. 3 TPC (a) and ARP (b) of flower head extracts during thesecond year in open field. Mean values of three replicates ±SEM. The means followed by different letters are significantlydifferent (P<0.05). C, control; G.m., G. mosseae; G.i.,G. intraradices; G. mix, G. mosseae + G. intraradices
C G.m. G.i. G.mix0
2
4
6
8
10
a
bbc
c
TP
C (
µg C
A m
g-1
FW
)
a
C G.m. G.i. G.mix0
2
4
6
8
10
a
b b
cb
AR
P
Fig. 2 TPC (a) and ARP (b) of flower head extracts during thefirst year in open field. Mean values of three replicates ± SEM.The means followed by different letters are significantlydifferent (P<0.05). C, control; G.m., G. mosseae; G.i.,G. intraradices; G. mix, G. mosseae + G. intraradices
Plant Soil (2010) 335:311–323 317

Mycorrhizal root colonisation Root samples collectedin the field from AMF inoculated artichoke plantsshowed significantly higher mycorrhizal colonisation(30.4±5.7 for G. mosseae and 36.6±5.3% for G.intraradices) than uninoculated plants (16.8±2.0%)(P<0.05).
Molecular analysis of root samples The amplificationproducts obtained by using nested PCR protocol were550–650 bp, the expected size of full-length ITSregion. Combination of HinfI and MboI restrictionpatterns produced by cloned fragments allowed thegrouping of clones into 8 operative taxonomicalunits (OTUs), one of which (OTU 1) represented54.4% of clones (Table 3). Data showed that 73.1%,75.5% and 73.1% of clones obtained from roots ofplants treated with G. mosseae, G. intraradices andGlomus mix, respectively, produced sequences withhomology to those of the genus Glomus present inGenbank database, whereas only 5.5% of clonesobtained from uninoculated roots showed Glomussequences (Table 3).
Altogether we obtained 26 ITS sequences from the12 DNA extracts of artichoke roots, 14 correspondingto Glomeromycota (from OTU 1, 2, 5, 6) and 12 toother fungi.
Sequences obtained from clones belonging toOTU1 and OTU2 showed homology with sequencesattributed to “uncultured Glomus” after BLASTsearch, though the analysis of the 5.8S tree topologyshowed that sequences originating from OTU 2clustered within a clade containing many G. intra-radices database sequences, whereas sequences fromOTU1 did not cluster with known species sequences.Clones grouped into OTU 1 originated from roots ofall four treatments, whereas those belonging to OTU2 were isolated only from G. intraradices and Glomusmix inoculated plant roots. Sequences originating
from clones showing restriction patterns character-ising OTU 6, obtained from roots of G. mosseaeinoculated plants, clustered with database 5.8Ssequences of G. mosseae. Sequences from clonesgrouped into OTU 5, originating from roots ofuninoculated plants and of Glomus mix inoculatedplants, clustered with database 5.8S sequences ofGlomus caledonium-Glomus geosporum.
The whole ITS rDNA analysis confirmed similar-ities observed by 5.8S rDNA analysis.
Discussion
This is the first report of large increases of TPC andARP in both leaves and flower heads of artichokeplants inoculated with the AM fungal species G.intraradices and G. mosseae, assessed in greenhouseand in 2 years-field experiments.
Mycorrhizal colonisation and growth responses
G. intraradices showed significantly higher colonisa-tion, compared with G. mosseae (83% and 37%,respectively). Many plant species belonging to thefamily Asteraceae host AM fungal symbionts innature, and a wide variability in root colonisationhas been found among species in symbiosis with bothnative (Pawlowska et al. 1997; Bauer et al. 2003;Kumar et al. 2003; Apple et al. 2005) and exoticAMF (Ruiz-Lozano et al. 1996; Marin et al. 2002;Turnau and Mesjasz-Przybylowicz 2003; Araim et al.2009). G. mosseae colonised 68–70% of the rootlength in Lactuca sativa and a wild variety of Cynaracardunculus plants, respectively (Marin et al. 2002),whereas 10–100% of colonised root length wasobserved in Berkheya coddii and Echinacea pur-purea, respectively, in symbiosis with G. intraradices
Treatment OTU Analysed clones
1 2 3 4 5 6 7 8
Control 1 0 18 11 1 0 2 3 36
Glomus mosseae 36 0 6 8 0 2 0 0 52
Glomus intraradices 36 4 9 4 0 0 0 0 53
G.mosseae + G. intraradices 32 1 9 5 4 1 0 0 52
Analysed clones 105 5 42 28 5 3 2 3 193
Table 3 Distribution of thenumber of clones recoveredfrom Cynara scolymus rootsinto the different treatmentsand into OperativeTaxonomic Units (OTUs)after combinationof HinfI and MboI restrictionpatterns
318 Plant Soil (2010) 335:311–323

(Turnau and Mesjasz–Przybylowicz 2003; Araimet al. 2009). The consequences of multiple coloniza-tion of host plants by different AMF were previouslystudied (Lekberg et al. 2007; Maherali and Klironomos2007). G. intraradices BEG141 showed higher abilityto colonise lucerne roots than G. mosseae BEG 12(Alkan et al. 2006), despite G. intraradices displayedhigher infectivity, suggesting that symbiosis establish-ment may result not only from infectivity but also fromgrowth conditions and P availability in roots (Daft1983; Boddington and Dodd 2000; Alkan et al. 2006).
Mycorrhizal inoculation produced significantincreases of leaf area and root length of artichokeplantlets in greehouse. When transplanted in the field,plants inoculated with Glomus mix showed markedincreases of main flower head fresh weight both in thefirst and in second year (92.8% and 70.6%, respec-tively), in comparison with control plants and plantsinoculated with G. mosseae and G. intraradices. Alarge body of evidence has shown that plant growthresponses to mycorrhizal colonization depend both onplant and fungal species and on experimental andenvironmental conditions (Smith and Read 2008).Accordingly, cardoon, a variety of C. cardunculus,showed larger shoot dry weights (increases of 50%)and lower root:shoot ratio when inoculated with twoGlomus species, compared to non mycorrhizal plants(Marin et al. 2002). Increases in biomass productionwere previously reported in other species of theAsteraceae family. In L. sativa increases of leaf dryweight ranged from 95% to 191% in plants inoculatedwith G. mosseae and Glomus deserticola, respectively(Ruiz-Lozano et al. 1996).
In this work we analysed sequences of rDNA-ITS,widely used for AMF detection in roots (Renker et al.2003, 2005; Börstler et al. 2006; Sykorova et al.2007), obtained from artichoke roots after 2 yearsgrowth in the field: sequences clustering with those ofG. mosseae and G. intraradices were retrieved fromG. mosseae and G. intraradices inoculated plants,respectively, but not from uninoculated ones. Bothtypes of sequences occurred in Glomus mix inoculat-ed plant roots. Sequences clustering with G. geo-sporum/G. caledonium group were retrieved fromuninoculated plants and from G. mosseae inoculatedplants. These fungal species are frequently reported tooccur among AM fungal community in natural andagroecosystems (Landwehr et al. 2002; Galvan et al.2009).
Total phenolic content and antiradical power
In greenhouse-grown plants the enhanced phenoliccomplement of artichoke leaves obtained with my-corrhizal inoculation was affected by inoculumcomposition. TPC and ARP showed significantlyhigher values (50% and 33%, respectively) in theleaves of plants inoculated with Glomus mix, com-pared to control plants. This was apparently due to thepresence of G. intraradices in the fungal inoculum.All inoculated field grown plants showed higherphenolic compound content in the edible parts,compared with control plants. The highest valueswere observed in plants inoculated with both AMfungal species (Glomus mix). The higher TPC valuesfound in secondary flower heads (second year of fieldcultivation) in comparison with main flower heads(first year of field cultivation) are consistent withprevious data on CA content (Curadi et al. 2005).
Accumulation of secondary compounds, includingpolyphenols, in the roots of mycorrhizal plants wasreported by many authors (Morandi 1996; Fester et al.1999; Maier et al. 1999; Larose et al. 2002; Araimet al. 2009), while the influence of AM colonizationon metabolic activities in plant shoots was poorlyinvestigated (Devi and Reddy 2002). Some authorssuggested that accumulation of secondary compoundsin the shoots of AM plants may confer resistance tofungal pathogens (Toussaint 2007; Pozo and Azcon-Aguilar 2007; Herre et al. 2007). Enhanced phenoliccompounds content in mycorrhizal plants was found tobe correlated with reduced disease severity in Vicia fabaand tomato challenged with Botrytis fabae andPhytophthora nicotianae, respectively (Cordier et al.1996; Rabie 1998) and with reduction of Ralstoniasolanacearum population in tomato (Zhu and Yao 2004).
Evidences indicate that AM symbiosis induceschanges in host plant primary metabolism, includingphotosynthesis, and in water uptake and droughttolerance (Wright et al. 1998; Ruiz-Lozano 2003).Changes in secondary metabolism, such as phytohor-mone dynamics, structural modifications and activa-tion of defence mechanisms, were also found (Allenet al. 1982; Danneberg et al. 1992; Drüge andSchönbeck 1993; Medina et al. 2003; Strack et al.2003; Copetta et al. 2006). In mycorrhizal roots ofMedicago truncatula, activation of Krebs cycle and ofplastidial metabolism was detected, together withincreases in endogenous levels of fatty acids, carote-
Plant Soil (2010) 335:311–323 319

noids and aminoacids, such as tyrosine, which,together with phenylalanine, is the main precursor ofplant polyphenols in the phenylpropanoid metabolism(Lohse et al. 2005). Levels of transcripts encodingPAL showed transient increases in Oryza sativa plantscolonized by G. mosseae and in M. truncatulacolonised by G. versiforme (Harrison and Dixon1993; Blilou et al. 2000). In the latter plant species,increased chalcone synthase (CHS) transcripts accu-mulation was also found during colonization byGlomus versiforme (Harrison and Dixon 1993) andG. intraradices (Bonanomi et al. 2001). Furtherresearches should investigate whether mycorrhizalinoculation of artichoke plants affects the activity ofkey-enzymes of hydroxycinnamates biosynthesis,such as hydroxycinnamoyl-CoA:quinate hydroxycin-namoyltransferase (HQT) and hydroxycinnamoyl-CoA:shikimate/quinate hydroxycinnamoyltransferase(HCT), recently isolated in artichoke by Comino et al.(2007, 2009). Antioxidant activity of the edible partsof artichoke flower heads mirrored the pattern ofphenolics accumulation: ARP increases (52% and32% in the first and second year in the field,respectively) were higher in plants inoculated withboth Glomus species. So far, few studies investigatedAMF effects on the production of phenolic com-pounds in plants used for human nutrition, with theexception of some researches on medicinal andaromatic plants. Higher accumulation of antioxidantcompounds (rosmarinic acid and caffeic acid) wasreported in sweet basil shoots (Ocimum basilicum L.)inoculated with different Glomus species (Toussaintet al. 2007).
The larger increases in TPC and ARP valuesobtained with the mixed inoculum, regardless of theorigin of the extracts (leaves or flower heads) suggesta synergistic effect and may indicate a functionalcomplementarity between G. intraradices and G.mosseae, as suggested by Koide (2000). Suchphenomenon has been observed by other authors inM. truncatula L., Allium porrum L. and Medicagosativa L. plants simultaneously colonized by differentGlomus species (Jansa et al. 2008; Khan et al. 2008).Benefits of plants colonization by several AMFspecies were suggested to be due to successions ordifferential activity in time or to differences in AMFadaptation to environmental conditions (Daft andHogarth 1983; Pringle and Bever 2002; Sanders2004).
The general aim of the present study was todetermine whether different AM fungal species couldprovide an efficient and natural strategy to improvethe production and quality of artichoke plants, animportant Mediterranean crop. The results obtained inthe present 3 years’ study also underline that the effecton TPC and ARP values is strongly dependent on theAM fungal species used as inoculum, confirmingprevious reports on variable effects of different AMfungal isolates on host plant growth (Klironomos2003; Munkvold et al. 2004; Smith et al. 2004; Gaiet al. 2006).
Artichoke, a traditional component of the Mediter-ranean diet, may be regarded as a functional plantfood for its high content of secondary metaboliteswith antioxidant properties. The quality of plant foodis presently defined not only by its nutritionalfeatures, but also by its content in natural substancesthat prevent diseases induced by oxidative stress, suchas atherosclerosis and carcinogenesis. New varietiesof artichoke with higher polyphenolic content couldbe obtained by means of genetic engineering andbreeding programs (Niggeweg et al. 2004). However,more information is needed on plant genetic variabil-ity concerning polyphenolic contents as affected byplant development and environmental and agronomicconditions. To such aim, mycorrhizal inoculation withselected species of AMF represents a useful biotechno-logical application, affecting plant biosynthesis ofsecondary metabolites with health promoting activities.
Acknowledgements This research was supported by Minis-tero Italiano Università e Ricerca, PRIN project “ New methodsfor high quality artichoke production” N° 2005078347.
References
Alkan N, Gadkar V, Yarden O, Kapulnik Y (2006) Analysis ofquantitative interactions between two species of arbuscularmycorrhizal fungi, Glomus mosseae and G. intraradices,by real-time PCR. Appl Environ Microbiol 72:4192–4199
Allen MF, Moore TS Jr, Christensen M (1982) Phytohormonechanges in Bouteloua gracilis infected by vesicular-arbuscular mycorrhizae. II. Altered levels of gibberellin-like substances and abscisic acid in the host plant. Can JBot 60:468–471
Apple ME, Thee CI, Smith-Longozo VL, Cogar CR, Wells CE,Nowak RS (2005) Arbuscular mycorrhizal colonization ofLarrea tridentata and Ambrosia dumosa roots varies withprecipitation and season in the Mojave Desert. Symbiosis39:131–135
320 Plant Soil (2010) 335:311–323

Araim G, Saleem A, Arnason JT, Charest C (2009) Rootcolonization by an arbuscular mycorrhizal (AM) fungusincreases growth and secondary metabolism of purpleconeflower, Echinacea purpurea (L.) Moench. J Agr FoodChem 57:2255–2258
Bauer CR, Kellogg CH, Bridgham SD, Lamberti GA (2003)Mycorrhizal colonization across hydrologic gradients inrestored and reference freshwater wetlands. Wetlands23:961–968
Blilou I, Ocampo JA, Garcia GJ (2000) Induction of Ltp (lipidtransfer protein) and Pal (phenylalanine ammonia-lyase) geneexpression in rice roots colonized by the arbuscular mycor-rhizal fungus Glomus mosseae. J Exp Bot 51:1969–1977
Boddington CL, Dodd JC (2000) The effect of agriculturalpractices on the development of indigenous arbuscularmycorrhizal fungi. II. Studies in experimental microcosms.Plant Soil 218:145–157
Bonanomi A, Oetiker JH, Guggenheim R, Boller T, WiemkenA, Vogeli LR (2001) Arbuscular mycorrhiza in mini-mycorrhizotrons: first contact of Medicago truncatularoots with Glomus intraradices induces chalcone synthase.New Phytol 150:573–582
Bondet V, Brand-Williams W, Berset C (1997) Kinetics andmechanisms of antioxidant activity using the DPPH• freeradical method. Lebensm Wiss Technol 30:609–615
Börstler B, Renker C, Kahmen A, Buscot F (2006) Speciescomposition of arbuscular mycorrhizal fungi in twomountain meadows with differing management types andlevels of plant biodiversity. Biol Fert Soil 42:286–298
Brand-Williams W, Cuvelier ME, Berset C (1995) Use of a freeradical method to evaluate antioxidant activity. LebensmWiss Technol 28:25–30
Brown JE, Rice-Evans CA (1998) Luteolin-rich artichokeextracts protect low density lipoprotein from oxidation invitro. Free Radical Res 29:247–255
Bundy R, Walker AF, Middleton RW, Wallis C, Simpson HCR(2008) Artichoke leaf extract (Cynara scolymus L.)reduces plasma cholesterol in otherwise healthy hypocho-lesterolemic adults: a randomized, double blind placebocontrolled trial. Phytomedicine 15:668–675
Comino C, Lanteri S, Portis E, Acquadro A, Romani A, HehnA, Larbat R, Bourgaud F (2007) Isolation and functionalcharacterization of a cDNA coding a hydroxycinnamoyl-transferase involved in phenylpropanoid biosynthesis inCynara cardunculus L. BMC Plant Biol 7:14
Comino C, Hehn A, Moglia A, Menin B, Bourgaud F, LanteriS, Portis E (2009) The isolation and mapping of a novelhydroxycinnamoyltransferase in the globe artichokechlorogenic acid pathway. BMC Plant Biol 9:30
Copetta A, Lingua G, Berta G (2006) Effects of three AM fungion growth, distribution of glandular hairs, and essential oilproduction in Ocimum basilicum L. var. Genovese.Mycorrhiza 16:485–494
Cordier C, Gianinazzi S, Gianinazzi-Pearson V (1996) Coloni-sation patterns of root tissues by Phytophthora nicotianaevar parasitica related to reduced disease in mycorrhizaltomato. Plant Soil 185:223–232
Curadi M, Picciarelli P, Lorenzi R, Graifenberg A, Ceccarelli N(2005) Antioxidant activity and phenolic compounds inthe edible parts of early and late Italian artichoke (Cynarascolymus L.) varieties. Ital J Food Sci 17:33–43
Daft MJ (1983) The influence of mixed inocula on endomycor-rhizal development. Plant Soil 71:331–337
Daft MJ, Hogarth BG (1983) Competitive interactions amongstfour species of Glomus on maize and onion. T Br MycolSoc 80:339–345
Danneberg G, Latus C, Zimmer W, Hundeshagen B, Schneider-Poetsch H, Bothe H (1992) Influence of vesicular-arbuscular mycorrhiza on phytohormone balances inmaize (Zea mays L.). J Plant Physiol 141:33–39
Devi MC, Reddy MN (2002) Phenolic acid metabolism ofgroundnut (Arachis hypogaea L.) plants inoculated withVAM fungus and Rhizobium. Plant Growth Regul37:151–156
Drüge U, Schönbeck F (1993) Effect of vesicular-arbuscularmycorrhizal infection on transpiration, photosynthesis andgrowth of flax (Linum usitatissimum L.) in relation tocytokinin levels. J Plant Physiol 141:40–48
FAOSTAT (2007) http://faostat.fao.org/Fester T, Hause G (2005) Accumulation of reactive oxigen
species in arbuscular mycorrhyzal roots. Mycorrhiza15:373–379
Fester T, Maier W, Strack D (1999) Accumulation ofsecondary compounds in barley and wheat roots inresponse to inoculation with an arbuscular mycorrhizalfungus and co-inoculation with rhizosphere bacteria.Mycorrhiza 8:241–246
Fester T, Strack D, Hause G (2001) Reorganization of tobaccoroot plastids during arbuscule developement. Planta213:864–868
Fester T, Wray V, Nimtz M, Strack D (2005) Is stimulation ofcarotenoid biosynthesis in arbuscular mycorrhizal roots ageneral phenomenon? Phytochemistry 66:1781–1786
Gai JP, Feng G, Christie P, Li XL (2006) Screening ofarbuscular mycorrhizal fungi for symbiotic efficiency withsweet potato. J Plant Nutr 29:1085–1094
Galvan GA, Paradi I, Burger K, Baar J, Kuyper TW, ScholtenOE, Kik C (2009) Molecular diversity of arbuscularmycorrhizal fungi in onion roots from organic andconventional farming systems in the Netherlands. Mycor-rhiza 19:317–328
Gebhardt R (1997) Antioxidative and protective properties ofextracts from leaves of the artichoke (Cynara scolymus L.)against hydroperoxide-induced oxidative stress in culturedhepatocytes. Toxicol Appl Pharm 144:279–286
Giovannetti M, Avio L (2002) Biotechnology of arbuscularmycorrhizas. In: Khachatourians GG, Arora Dilip K (eds)Applied mycology and biotechnology, volume 2, agricultureand food production. Elsevier, Amsterdam, pp 275–310
Giovannetti M, Mosse B (1980) An evaluation of techniquesfor measuring vescicular arbuscular mycorrhizal infectionin roots. New Phytol 84:489–500
Hans J, Hause B, Strack D, Walter MH (2004) Cloning,characterization, and immunolocalization of a mycorrhiza-inducible 1-deoxy-D-xylulose-5-phosphate reductoisomer-ase in arbuscule-containing cells of maize. Plant Physiol134:614–624
Harrison MJ, Dixon RA (1993) Isoflavonoid accumulation andexpression of defense gene transcripts during the estab-lishment of vesicular-arbuscular mycorrhizal associationsin roots of Medicago truncatula. Mol Plant Microbe In6:643–654
Plant Soil (2010) 335:311–323 321

Hempel S, Renker C, Buscot F (2007) Differences in thespecies composition of arbuscular mycorrhizal fungi inspore, root and soil communities in a grassland ecosystem.Environ Microbiol 9:1930–1938
Herre EA, Mejia LC, Kyllo DA, Rojas E, Maynard Z, Butler A,Van Bael SA (2007) Ecological implications of anti-pathogen effects of tropical fungal endophytes andmycorrhizae. Ecology 88:550–558
Jansa J, Smith FA, Smith SE (2008) Are there benefits ofsimultaneous root colonization by different arbuscularmycorrhizal fungi? New Phytol 177:779–789
Khan IA, Ayub N, Mirza SN, Nizami SM, Azam M (2008)Synergistic effect of dual inoculation (Vesicular-Arbuscu-lar Mycorrhizae) on the growth and nutrients uptake ofMedicago sativa. Pakistan J Bot 40:939–945
Klironomos JN (2003) Variation in plant response to native andexotic arbuscular mycorrhizal fungi. Ecology 84:2292–2301
Koide RT (2000) Functional complementarity in the arbuscularmycorrhizal symbiosis. New Phytol 147:233–235
Kraft K (1997) Artichoke leaf extracts—recent findingsreflecting effects on lipid metabolism. Liver and gastroin-testinal tracts. Phytomedicine 4:369–378
Kumar A, Raghuwanshi R, Upadhyay RS (2003) Vesicular-arbuscular mycorrhizal association in naturally revegetatedcoal mine spoil. Trop Ecol 44:253–256
Lambais MR, Rios-Ruiz WF, Andrade RM (2003) Antioxidantresponses in bean (Phaseolus vulgaris) roots colonized byarbuscular mycorrhizal fungi. New Phytol 160:421–428
Landwehr M, Hildebrandt U, Wilde P, Nawrath K, Toth T, BiroB, Bothe H (2002) The arbuscular mycorrhizal fungusGlomus geosporum in European saline, sodic and gypsumsoils. Mycorrhiza 12:199–211
Larose G, Chenevert R, Moutoglis P, Gagne S, Piche Y,Vierheilig H (2002) Flavonoid levels in roots of Medicagosativa are modulated by the developmental stage of thesymbiosis and the root colonizing arbuscular mycorrhizalfungus. J Plant Physiol 159:1329–1339
Lekberg Y, Koide RT, Rohr JR, Aldrich-Wolfe L, Morton JB(2007) Role of niche restrictions and dispersal in thecomposition of arbuscular mycorrhizal fungal communi-ties. J Ecol 95:95–105
Lohse S, Schliemann W, Ammer C, Kopka J, Strack D, FesterT (2005) Organisation and metabolism of plastids andmitochondria in arbuscular mycorrhizal roots of Medicagotruncatula. Plant Physiol 139:329–340
Maherali H, Klironomos JN (2007) Influence of phylogeny onfungal community assembly and ecosystem functioning.Science 316:1746–1748
Maier W, Schmidt J, Wray V, Walter MH, Strack D (1999) Thearbuscular mycorrhizal fungus, Glomus intraradices,induces the accumulation of cyclohexenone derivativesin tobacco roots. Planta 207:620–623
Marin M, Ybarra M, Garcia-Ferriz F, Garcia-Ferriz L (2002)Effect of arbuscular mycorrhizal fungi and pesticides onCynara cardunculus growth. Agr Food Sci Finland11:245–251
Marulanda A, Porcel R, Barea JM, Azcon R (2007) Droughttolerance and antioxidant activities in lavander plantscolonized by native drought-tolerant of drught-sensitiveGlomus species. Microb Ecol 54:543–552
Medina MJH, Gagnon H, Pichq Y, Ocampo JA, Garrido JMG,Vierheilig H (2003) Root colonization by arbuscularmycorrhizal fungi is affected by the salicylic acid contentof the plant. Plant Sci 164:993–998
Morandi D (1996) Occurrence of phytoalexins and phenoliccompounds on endomycorrhizal interactions, and theirpotential role in biological control. Plant Soil 185:241–251
Munkvold L, Kjoller R, Vestberg M, Rosendahl S, Jakobsen I(2004) High functional diversity within species of arbus-cular mycorrhizal fungi. New Phytol 164:357–364
Niggeweg R, Michael A, Martin C (2004) Engineering plantswith increased levels of the antioxidant chlorogenic acid.Nature Biotechnol 22:746–754
Pawlowska TE, Blaszkowski B, Rühling Å (1997) Themycorrhizal status of plants colonizing a calamine spoilmound in southern Poland. Mycorrhiza 6:499–505
Pozo MJ, Azcon-Aguilar C (2007) Unraveling mycorrhiza-induced resistance. Curr Opin Plant Biol 10:393–398
Pringle A, Bever JD (2002) Divergent phenologies mayfacilitate the coexistence of arbuscular mycorrhizal fungiin a North Carolina grassland. Am J Bot 89:1439–1446
Rabie GH (1998) Induction of fungal disease resistance in Viciafaba by dual inoculation with Rhizobium leguminosarumand vesicular-arbuscular mycorrhizal fungi. Mycopatholo-gia 141:159–166
Redecker D, Hijri M, Dulieu H, Sanders IR (1999) Phyloge-netic analysis of a dataset of fungal 5.8S rDNA sequencesshows that highly divergent copies of internal transcribedspacers reported from Scutellospora castanea are ofascomycete origin. Fung Genet Biol 28:238–244
Renker C, Heinrichs J, Kaldorf M, Buscot F (2003) Combiningnested PCR and restriction digest of the internal tran-scribed spacer region to characterize arbuscular mycorrhi-zal fungi on roots from the field. Mycorrhiza 13:191–198
Renker C, Blanke V, Buscot F (2005) Diversity of arbuscularmycorrhizal fungi in grassland spontaneously developedon area polluted by a fertilizer plant. Environ Pollut135:255–266
Ruiz-Lozano JM (2003) Arbuscular mycorrhizal symbiosis andalleviation of osmotic stress. New perspectives formolecular studies. Mycorrhiza 13:309–317
Ruiz-Lozano JM, Azcon R, Palma JM (1996) Superoxidedismutase activity in arbuscular mycorrizal Lactucasativa plants subjected to drought stress. New Phytol 134:327–333
Sanders IR (2004) Intraspecific genetic variation in arbuscularmycorrhizal fungi and its consequences for molecularbiology, ecology, and development of inoculum. Can J Bot82:1057–1062
Sawa T, Nakao M, Akaike T, Ono K, Maeda H (1999)Alkylperoxyl radical-scavenging activity of various flavo-noids and other phenolic compounds: implications for theanti-tumor-promoter effect of vegetables. J Agr FoodChem 47:397–402
Schliemann W, Ammer C, Strack D (2008) Metabolic profilingof mycorrhizal roots of Medicago truncatula. Phytochem-istry 69:112–146
Shadidi F, Naczk M (1995) Methods of analysis and quantifi-cation of phenolic compounds. In: Shahidi F, Naczk M(eds) Food phenolics: sources. Chemistry, effects andapplications. Technomic, Lancaster, pp 287–293
322 Plant Soil (2010) 335:311–323

Smith SE, Read DJ (2008) Mycorrhizal symbiosis. AcademicPress, Cambridge, pp 1–787
Smith SE, Smith FA, Jakobsen I (2004) Functional diversity inarbuscular mycorrhizal (AM) symbioses: the contributionof the mycorrhizal P uptake pathway is not correlated withmycorrhizal responses in growth or total P uptake. NewPhytol 162:511–524
Strack D, Fester T, Hause B, Schliemann W, Walter M (2003)Arbuscular mycorrhiza: biological, chemical, and molec-ular aspects. J Chem Ecol 29:1955–1979
Sykorova Z, Ineichen K, Wiemken A, Redecker D (2007) Thecultivation bias: different communities of arbuscularmycorrhizal fungi detected in roots from the field, frombait plants transplanted to the field, and from a greenhousetrap experiment. Mycorrhiza 18:1–14
Toussaint JP (2007) Investigating physiological changes in theaerial parts of AM plants. What do we know and whereshould we be heading. Mycorrhiza 17:349–353
Toussaint JP, Smith FA, Smith SE (2007) Arbuscular mycor-rhizal fungi can induce the production of phytochemicals
in sweet basil irrespective of phosphorus nutrition.Mycorrhiza 17:291–297
Turnau K, Mesjasz-Przybylowicz J (2003) Arbuscular mycor-rhiza of Berkheya coddii and other Ni-hyperaccumulatingmembers of Asteraceae from ultramafic soils in SouthAfrica. Mycorrhiza 13:185–190
Walter MH, Fester T, Strack D (2000) Arbuscular mycorrhizalfungi induce the non-mevalonate methylerythriol phoshatepathway of isoprenoid biosynthesis correlated with accu-mulation of the “yellow pigment” and other apocarote-noids. Plant J 21:571–578
Wang M, Simon LE, Aviles IF, He K, Zheng Q, Tadmor Y (2003)Analysis of antioxidative phenolic compounds in artichoke(Cynara scolymus L.). J Agr Food Chem 51:601–608
Wright DP, Scholes JD, ReadDJ (1998) Effects of VAmycorrhizalcolonization on photosynthesis and biomass production ofTrifolium repens L. Plant Cell Environ 21:209–216
ZhuHH, YaoQ (2004) Localized and systemic increase of phenolsin tomato roots induced by Glomus versiforme inhibitsRalstonia solanacearum. J Phytopathol 152:537–542
Plant Soil (2010) 335:311–323 323
Copyright © 2022 FDOKUMEN




















