
Motor Patterns in Humans Walking With Bent Posture Versus ...
13
Interactions Between Posture and Locomotion: Motor Patterns in Humans Walking With Bent Posture Versus Erect Posture R. GRASSO, M. ZAGO, AND F. LACQUANITI Human Physiology Section of the Scientific Institute Santa Lucia and the University of Rome “Tor Vergata”, 00179 Rome, Italy Grasso, R., M. Zago, and F. Lacquaniti. Interactions between posture and locomotion: motor patterns in humans walking with bent posture versus erect posture. J. Neurophysiol. 83: 288 –300, 2000. Human erect locomotion is unique among living primates. Evolution selected specific biomechanical features that make human locomotion mechanically efficient. These features are matched by the motor patterns generated in the CNS. What happens when humans walk with bent postures? Are normal motor patterns of erect locomotion main- tained or completely reorganized? Five healthy volunteers walked straight and forward at different speeds in three different postures (regular, knee-flexed, and knee- and trunk-flexed) while their motion, ground reaction forces, and electromyographic (EMG) activity were recorded. The three postures imply large differences in the position of the center of body mass relative to the body segments. The elevation angles of the trunk, pelvis, and lower limb segments relative to the vertical in the sagittal plane, the ground reaction forces and the rectified EMGs were analyzed over the gait cycle. The waveforms of the elevation angles along the gait cycle remained essentially un- changed irrespective of the adopted postures. The first two harmonics of these kinematic waveforms explain .95% of their variance. The phase shift but not the amplitude ratio between the first harmonic of the elevation angle waveforms of adjacent pairs was affected system- atically by changes in posture. Thigh, shank, and foot angles covaried close to a plane in all conditions, but the plane orientation was systematically different in bent versus erect locomotion. This was explained by the changes in the temporal coupling among the three segments. For walking speeds .1ms 21 , the plane orientation of bent locomotion indicates a much lower mechanical efficiency relative to erect locomotion. Ground reaction forces differed prominently in bent versus erect posture displaying characteristics intermediate between those typical of walking and those of running. Mean EMG activity was greater in bent postures for all recorded muscles independent of the functional role. The waveforms of the muscle activities and muscle synergies also were affected by the adopted posture. We conclude that maintaining bent postures does not interfere either with the generation of segmental kinematic waveforms or with the planar constraint of intersegmental covariation. These characteristics are maintained at the expense of adjustments in kinetic parameters, mus- cle synergies and the temporal coupling among the oscillating body segments. We argue that an integrated control of gait and posture is made possible because these two motor functions share some common principles of spatial organization. INTRODUCTION The issue of the relationship between posture and locomo- tion is of great theoretical and experimental relevance (see Burleigh et al. 1994; Lacquaniti et al. 1997; Massion 1992; Mori 1987; Winter 1991; Zernicke and Smith 1996). Neuro- physiological studies indicate that the control of posture and locomotion are interdependent at many different levels of the CNS, from the motor cortex to the basal ganglia, the brain stem, and the spinal cord. Thus basal ganglia are connected reciprocally with ponto-mesecephalic structures (the pedunco- lopontine nucleus) belonging to the mesencephalic locomotor region (an area known to modulate spinal locomotion oscilla- tors) (Shik and Orlovsky 1976), on the one side, and with the axial motor cortex, on the other side. These structures partic- ipate in the preparation and initiation of locomotion by pro- viding the appropriate spatial frameworks necessary to incor- porate postural adjustments in the locomotion process (Garcia- Rill 1986; Grasso et al. 1999). Also when specific areas in the hypothalamus or the brain stem are stimulated, changes in posture are triggered just before step initiation (Mori 1987; Mori et al. 1978, 1983, 1989). Interactions between pathways controlling posture and gait exist even at the level of spinal premotor interneurons (Jankowska and Edgley 1993). In sum, it has been suggested that the “activation of setting mecha- nisms in the level of postural muscle tone and that of the spinal stepping generator are not separate phenomena” (Mori 1987). If the interrelationship between posture and locomotion is well recognized, the implications vis-a `-vis the issue of the motor pattern generation are not well understood (see, how- ever, Zernicke and Smith 1996). In particular, it is not clear whether or not the setting of different task-dependent postural configurations of the body should affect the waveforms of the basic motor patterns that are rhythmically output during walk- ing. This question hinges on the more general problem of the nature of the control waveforms output by central locomotor networks (see Lacquaniti et al. 1999; Zernicke and Smith 1996). It is generally thought that the multisegment motion of mammals locomotion is controlled by a network of coupled oscillators (central pattern generators or CPGs) (see Grillner 1981; Pearson 1993; Rossignol 1996). Although it often is assumed that CPGs control patterns of muscle activity, an equally plausible hypothesis holds that they control patterns of limb segment motion instead (Bianchi et al. 1998b; Borghese et al. 1996; Grasso et al. 1998; Shen and Poppele 1995). This hypothesis is based on the following observations (for a re- view, see Lacquaniti et al. 1999). In walking the waveforms of limb segment angular motion are much simpler and more consistent than the corresponding waveforms of muscle activ- ity, both in man (Borghese et al. 1996; Grasso et al. 1998) and The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 288 0022-3077/00 $5.00 Copyright © 2000 The American Physiological Society
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Motor Patterns in Humans Walking With Bent Posture Versus ...

Interactions Between Posture and Locomotion: Motor Patterns inHumans Walking With Bent Posture Versus Erect Posture
R. GRASSO, M. ZAGO, AND F. LACQUANITIHuman Physiology Section of the Scientific Institute Santa Lucia and the University of Rome “Tor Vergata”,00179 Rome, Italy
Grasso, R., M. Zago, and F. Lacquaniti. Interactions betweenposture and locomotion: motor patterns in humans walking with bentposture versus erect posture.J. Neurophysiol.83: 288–300, 2000.Human erect locomotion is unique among living primates. Evolutionselected specific biomechanical features that make human locomotionmechanically efficient. These features are matched by the motorpatterns generated in the CNS. What happens when humans walk withbent postures? Are normal motor patterns of erect locomotion main-tained or completely reorganized? Five healthy volunteers walkedstraight and forward at different speeds in three different postures(regular, knee-flexed, and knee- and trunk-flexed) while their motion,ground reaction forces, and electromyographic (EMG) activity wererecorded. The three postures imply large differences in the position ofthe center of body mass relative to the body segments. The elevationangles of the trunk, pelvis, and lower limb segments relative to thevertical in the sagittal plane, the ground reaction forces and therectified EMGs were analyzed over the gait cycle. The waveforms ofthe elevation angles along the gait cycle remained essentially un-changed irrespective of the adopted postures. The first two harmonicsof these kinematic waveforms explain.95% of their variance. Thephase shift but not the amplitude ratio between the first harmonic ofthe elevation angle waveforms of adjacent pairs was affected system-atically by changes in posture. Thigh, shank, and foot angles covariedclose to a plane in all conditions, but the plane orientation wassystematically different in bent versus erect locomotion. This wasexplained by the changes in the temporal coupling among the threesegments. For walking speeds.1 m s21, the plane orientation of bentlocomotion indicates a much lower mechanical efficiency relative toerect locomotion. Ground reaction forces differed prominently in bentversus erect posture displaying characteristics intermediate betweenthose typical of walking and those of running. Mean EMG activitywas greater in bent postures for all recorded muscles independent ofthe functional role. The waveforms of the muscle activities andmuscle synergies also were affected by the adopted posture. Weconclude that maintaining bent postures does not interfere either withthe generation of segmental kinematic waveforms or with the planarconstraint of intersegmental covariation. These characteristics aremaintained at the expense of adjustments in kinetic parameters, mus-cle synergies and the temporal coupling among the oscillating bodysegments. We argue that an integrated control of gait and posture ismade possible because these two motor functions share some commonprinciples of spatial organization.
I N T R O D U C T I O N
The issue of the relationship between posture and locomo-tion is of great theoretical and experimental relevance (see
Burleigh et al. 1994; Lacquaniti et al. 1997; Massion 1992;Mori 1987; Winter 1991; Zernicke and Smith 1996). Neuro-physiological studies indicate that the control of posture andlocomotion are interdependent at many different levels of theCNS, from the motor cortex to the basal ganglia, the brainstem, and the spinal cord. Thus basal ganglia are connectedreciprocally with ponto-mesecephalic structures (the pedunco-lopontine nucleus) belonging to the mesencephalic locomotorregion (an area known to modulate spinal locomotion oscilla-tors) (Shik and Orlovsky 1976), on the one side, and with theaxial motor cortex, on the other side. These structures partic-ipate in the preparation and initiation of locomotion by pro-viding the appropriate spatial frameworks necessary to incor-porate postural adjustments in the locomotion process (Garcia-Rill 1986; Grasso et al. 1999). Also when specific areas in thehypothalamus or the brain stem are stimulated, changes inposture are triggered just before step initiation (Mori 1987;Mori et al. 1978, 1983, 1989). Interactions between pathwayscontrolling posture and gait exist even at the level of spinalpremotor interneurons (Jankowska and Edgley 1993). In sum,it has been suggested that the “activation of setting mecha-nisms in the level of postural muscle tone and that of the spinalstepping generator are not separate phenomena” (Mori 1987).
If the interrelationship between posture and locomotion iswell recognized, the implications vis-a`-vis the issue of themotor pattern generation are not well understood (see, how-ever, Zernicke and Smith 1996). In particular, it is not clearwhether or not the setting of different task-dependent posturalconfigurations of the body should affect the waveforms of thebasic motor patterns that are rhythmically output during walk-ing. This question hinges on the more general problem of thenature of the control waveforms output by central locomotornetworks (see Lacquaniti et al. 1999; Zernicke and Smith1996).
It is generally thought that the multisegment motion ofmammals locomotion is controlled by a network of coupledoscillators (central pattern generators or CPGs) (see Grillner1981; Pearson 1993; Rossignol 1996). Although it often isassumed that CPGs control patterns of muscle activity, anequally plausible hypothesis holds that they control patterns oflimb segment motion instead (Bianchi et al. 1998b; Borgheseet al. 1996; Grasso et al. 1998; Shen and Poppele 1995). Thishypothesis is based on the following observations (for a re-view, see Lacquaniti et al. 1999). In walking the waveforms oflimb segment angular motion are much simpler and moreconsistent than the corresponding waveforms of muscle activ-ity, both in man (Borghese et al. 1996; Grasso et al. 1998) and
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
288 0022-3077/00 $5.00 Copyright © 2000 The American Physiological Society

cat (Shen and Poppele 1995). Moreover the temporal changesof the elevation angles of the lower limb segments do notevolve independently of each other, but they are tightly cou-pled together (Bianchi et al. 1998b; Borghese et al. 1996).When the elevation angles are plotted one versus the others,they describe a regular loop constrained close to a plane,common to both the stance and swing phase. The specificorientation of the plane of angular covariation reflects thephase relationships between the elevation angles of the seg-ments and therefore the timing of the intersegmental coordi-nation. The phase delay shifts systematically with increasingspeed both in man (Bianchi et al. 1998b) and cat (Shen andPoppele 1995). Because in man this phase-shift is correlatedwith the net mechanical power output over a gait cycle (Bian-chi et al. 1998b), we hypothesized that the control of kinematicphase can be used by the nervous system for limiting theoverall energy expenditure with increasing speed (Bianchi etal. 1998a). Finally, we observed that a reversal of the directionof walking from forward to backward involves the same wave-forms (time-reversed) of the elevation angles as in forwardgait, with a simple reversal of the delay in the phase couplingbetween limb segments, at the expense of a reorganization ofthe patterns of muscle activity (Grasso et al. 1998).
How does a change in walking posture affect these locomo-tor patterns? Although we normally walk with an erect posture,we can as easily walk stooped (as it happens in a low tunnel).Here we compared normal erect walking (regular, RE) withtwo different styles of bent walking, namely knee-flexed walk-ing (KF) and knee-flexed plus trunk-flexed walking (KT). REwalking is a unique feature of human locomotion (nonhumanprimates normally walk with a bent posture). Its evolutionaryhistory indicates highly specific adaptations of the skeletal andmuscular apparatus (Crompton et al. 1998; Spoor et al. 1994).Also, erect posture is mechanically efficient in humans becausethe center of body mass vaults over the supporting limb like aninverted pendulum, thereby limiting energy expenditure bymeans of an exchange of the forward kinetic energy with thegravitational potential energy (Cavagna et al. 1977). Maintain-ing KF and KT posture while walking may interfere with thependulum mechanism. McMahon et al. (1987) showed that insubjects running knee-flexed, reaction moments acting on theknee are increased and the effective vertical spring stiffness ofthe legs is decreased relative to normal running. We introducedcondition KT in addition to KF to assess the specific role oftrunk orientation in the generation of locomotor patterns. Infact it has been proposed that the trunk may act as a referencein the control of posture and movement (Darling and Miller1995; Massion et al. 1997; Mouchnino et al. 1993). This rolethen could be disrupted when the trunk is flexed.
In looking for the effects of changes of walking posture onthe locomotor patterns, we keep in mind that there exist inev-itable mechanical consequences of a bent posture. In particular,KF and KT walking must involve an offset in the mean valueof several kinematic and kinetic variables as compared with REwalking. For instance, because the legs are flexed, the meanelevation angles of the limb segments in the former tasks willbe generally different from those measured in the latter task.Also the mean level of muscle activity will be increasedbecause of the reduced mechanical advantage of bent limbs(Biewener 1990). With regard to the issue of locomotor patterngeneration, however, the key point concerns the waveform of
the kinetic and kinematic signals rather than their mean value.According to our previous hypothesis, locomotion is controlledin a kinematic space (see Lacquaniti et al. 1999). If so, onewould expect that the changes in posture should leave thekinematic waveforms essentially unchanged. The interrelation-ship between posture and locomotion should manifest itselfinstead on the phase delay between the motion of different limbsegments; that is, the control parameter that is set centrallyaccording to the specific task demands. (As we noted in thepreceding text, the phase delay varies as a function of changesin walking speed or reversal of walking direction.) An addi-tional implication of our hypothesis is that the conservation ofkinematic templates across changes in body posture can occuronly at the expense of a reorganization of muscle patterns andsynergies.
M E T H O D S
General procedures have been previously described (Bianchi et al.1998b; Borghese et al. 1996). Kinematic data were obtained by meansof the ELITE system (Ferrigno et al. 1990). Four 100-Hz TV cameraswere spaced on the recording side of the walkway to enhance spatialaccuracy. After three-dimensional (3D) calibration, the SD spatialaccuracy of the system was better than 1.5 mm (Bianchi et al. 1998b).The position of selected points on the side of the dominant lower limbwas recorded by attaching the infrared reflective markers to the skinoverlying the following landmarks: gleno-humeral joint (GH), ante-rior superior iliac spine (ASIS), posterior superior iliac spine (PSIS),greater trochanter (GT), lateral femur epicondyle (LE), lateral malle-olus (LM), and fifth metatarso-phalangeal joint (VM). VM markerwas placed on the shoe after verifying the correspondence on the barefoot. ASIS and PSIS coordinates were averaged to obtain ilium (IL)position. The body was modeled as an interconnected chain of rigidsegments: GH-IL for the trunk, IL-GT for the pelvis, GT-LE for thethigh, LE-LM for the shank, and LM-VM for the foot. The limb axiswas defined as GT-LM.
Ground reaction forces were recorded at 500 Hz by means of amulticomponent force platform (0.63 0.4 m, Kistler 9281B), placedapproximately at the center of the walkway. EMG activity was re-corded by means of surface electrodes from the gluteus maximus(GM), biceps femoris (long head) (BF), rectus femoris (RF), vastuslateralis (VL), lateral gastrocnemius (GCL), and tibialis anterior (TA).EMG signals were preamplified (1003) at the recording site, digi-tized, and transmitted to the remote amplifier via 15-m optic fibers.These signals were band-pass filtered (10-Hz high-pass and 200-Hzlow-pass, 4-pole Bessel filters), and sampled at 500 Hz. Sampling ofkinematic, force, and EMG data were synchronized.
Protocol
Experiments were approved by the Ethics Committee of SantaLucia Institute and conformed with the Declaration of Helsinki on theuse of human subjects in research. Five healthy volunteers (2 females,3 males, 21–36 yr age range) participated after giving informedconsent. Before the recording session, the dominant lower limb ofeach subject was determined according to standard criteria (Vanden-Abeele 1980). All subjects proved to be right-dominant. Before theexperiment, we asked our subjects to adopt static KF and KT postureson the force platform and computed the instantaneous position of thecenter of pressure of the net reaction forces during 20-s records(which were repeated 2 times). Fifteen seconds of steady state datawere averaged to obtain the mean anteroposterior displacement of thecenter of pressure relative to RE condition, which was 0.386 0.48and 0.436 0.66 cm (mean6 SD) forward for KF and KT, respec-tively. This small displacement is not statistically significant.
289POSTURAL GEOMETRY AND GAIT COORDINATION

Subjects then were instructed to walk (with their shoes on) with thearms folded on the chest, at a freely chosen, roughly constant speedwithin the ELITE sampling volume. They were encouraged to varythe speed across trials. Because no additional constraints were used,the speed range overlapped but did not coincide exactly in differentexperiments. To avoid modifications of the natural step length, sub-jects were asked not to pay attention whether or not they stepped ontothe force platform. Walks were performed in one of the three differentpostures (RE, KF, and KT), randomly alternated from trial to trial.
Data analysis
Three-dimensional kinematic data were filtered with an optimallow-pass FIR filter with automatic bandwidth selection (D’Amico andFerrigno 1990). The angle of elevation of theith segment in thesagittal plane with respect to the vertical was computed asai 5arctan[(xd 2 xp)/(yp 2 yd)], subscripts p and d denoting proximal anddistal endpoints of the segment, respectively, andx and y the hori-zontal and vertical coordinates in the sagittal plane, respectively.Elevation angles are positive in the forward direction relative to thevertical (i.e., when the distal marker falls anterior to the proximal).Gait cycle (T) was defined as the time interval between two successivemaxima in the time series of the limb axis elevation, step length (S) asthe linear translation of GT marker duringT, and average speed asV 5 S/T. Stance phase was defined as the interval during which thevertical reaction force exceeded 7% of body weight. The maximumelevation of the limb axis slightly precedes the heel touch-down(Borghese et al. 1996). Different trials from each subject were en-semble-averaged after time-interpolation of the kinematic data over Tto fit a normalized 1,000-points time base. Postural geometry wasmeasured by taking the mean, maximum and minimum value of theelevation angle of each segment over T. Thex, y coordinates of thelocation of the center of body mass (CM) are computed as the meanof x, y coordinates of the respective centers of mass of seven bodysegments: the H.A.T. plus the thigh, shank and foot of the right andleft limbs. H.A.T. is comprised of head, folded arms, trunk, and pelvisand is assumed to be one rigid link (corresponding to GH-GT seg-ment) (see Winter 1991). The center of mass of each of these seg-ments was derived from anthropometric measurements taken on thesubject and the geometric models based on the gamma-scannermethod (Zatsiorsky et al. 1990). Motion of the side of the bodycontralateral to the recording one was estimated by time-shifting therecorded data by T/2, under the assumption of symmetrical gait(Bianchi et al. 1998b).
EMGs were rectified numerically and low-pass filtered (in bothtime directions to avoid tail and phase distortions) by means of aButterworth filter, with cutoff at 25 Hz. Cross-correlation functions(CCF) between pairs of muscles were computed as previously de-scribed (Grasso et al. 1998).
Intersegmental coordination among the lower limb segments (thigh,shank, and foot) was evaluated starting from elevation angle wave-forms both in the time domain (1) and in position space (2).
1) Because of the periodic structure of the elevation angle wave-forms over the gait cycle (Bianchi et al. 1998b; Borghese et al. 1996),we decomposed such waveforms into their Fourier series components.The series was truncated at the 10th harmonic which had a frequencycorresponding to the highest low-pass cutoff frequency in the dataseries.
The amplitude transfer ratio (Gpd) and phase shift (Fpd) betweenthe corresponding Fourier harmonics from the elevation angles ofadjacent limb segmentsp and d were computed, respectively, asAk(d)/Ak(p) and asQk(d) 2 Qk(p) whereAk andQk are the amplitudeand phase respectively of thekth order Fourier series component.
2) The intersegmental coordination was evaluated in position spaceas previously described (Borghese et al. 1996). Briefly, the changes ofthe elevation angles of the thigh, shank, and foot covary linearlythroughout the gait cycle. The thigh-shank-foot 3D loops describe
paths that can be fitted by a plane that is computed by means oforthogonal regression (Bianchi et al. 1998b; Borghese et al. 1996). Ineach trial, we computed the covariance matrixR of the ensemble oftime-varying elevation angles over the gait cycle, after subtraction oftheir respective mean value. Eigenvaluesl and eigenvectorsu arecomputed by factoring the covariance matrixR from the set oforiginal signals by using a singular value decomposition algorithmsuch that
R 5 ULUT
whereU andL are the eigenvector and eigenvalue matrices, respec-tively, and T denotes matrix transpose. The sum of the eigenvalues isequal to the sum of the variances of the original signal waveforms.The three eigenvectorsu1 2 u3 of R, rank-ordered on the basis of thecorresponding eigenvalues, correspond to the orthogonal directions ofmaximum and minimum variance in the sample scatter. The first twoeigenvectorsu1 2 u2 lie on the best-fitting plane of angular covaria-tion. The third eigenvector (u3) is the normal to the plane and definesthe plane orientation in the position-space of the elevation angles. Foreach eigenvector, the parametersuit, uis, and uif correspond to thedirection cosines with the positive semi-axis of the thigh, shank andfoot angular coordinates, respectively. The direction cosines of theplane normal provide a measure of the orientation of the plane. Inparticular,u3t denotes the direction cosine with respect to the thighaxis.
R E S U L T S
General gait parameters
Walking flexed was performed easily in both KF and KTconditions. The subjects had no difficulty in maintaining thatposture for the few seconds required to walk the 10-m path. Weanalyzed a total number of 463 gait cycles at speeds within therange 0.24 2.0 m s21. The range of speeds 0.54 1.5 m s21
was represented evenly across subjects and conditions.The well-known monotonic increase of step length with
walking speed was not significantly affected by postural ge-ometry. The slopes of the intraindividual regression lines were0.4860.11 m per 1 m s21 speed increment, 0.446 0.33, and0.336 0.38 for RE, KF, and KT, respectively (the interceptswere 0.7560.09 m, 0.846 0.41, 0.836 0.37). However thescatter was larger for KF and KT than for RE (correlationcoefficients were 0.976 0.02, 0.856 0.10, and 0.696 0.38,respectively). Repeated-measures ANOVA across the threeconditions yielded nonsignificant outcomes for the compari-sons between regression parameters from the three conditions.On average, the duration of the stance phase was;60% of thegait cycle in all conditions (61.96 3.9%, 62.16 2.7%, and62.86 2.6% for RE, KF, and KT, respectively) and decreasedwith increasing speed (on average by 6.5% of gait cycleduration per 1 m s21). The latter relationship was less consis-tent in KF and KT than in RE.
Geometric postural features
Figure 1 (top) shows stick-diagram series from one subjectwalking at approximately the same speed ('1 m s21) with thethree different postures. As implied by the task, postural ge-ometry differs drastically across the three conditions. In KFand KT, the lower limbs are flexed and the trunk is tilted, moreso in KT. The vertical oscillations of the trunk and pelvissegments are less pronounced in KF and KT than in RE. Theposition of the CM is displaced forward and downward in KF
290 R. GRASSO, M. ZAGO, AND F. LACQUANITI
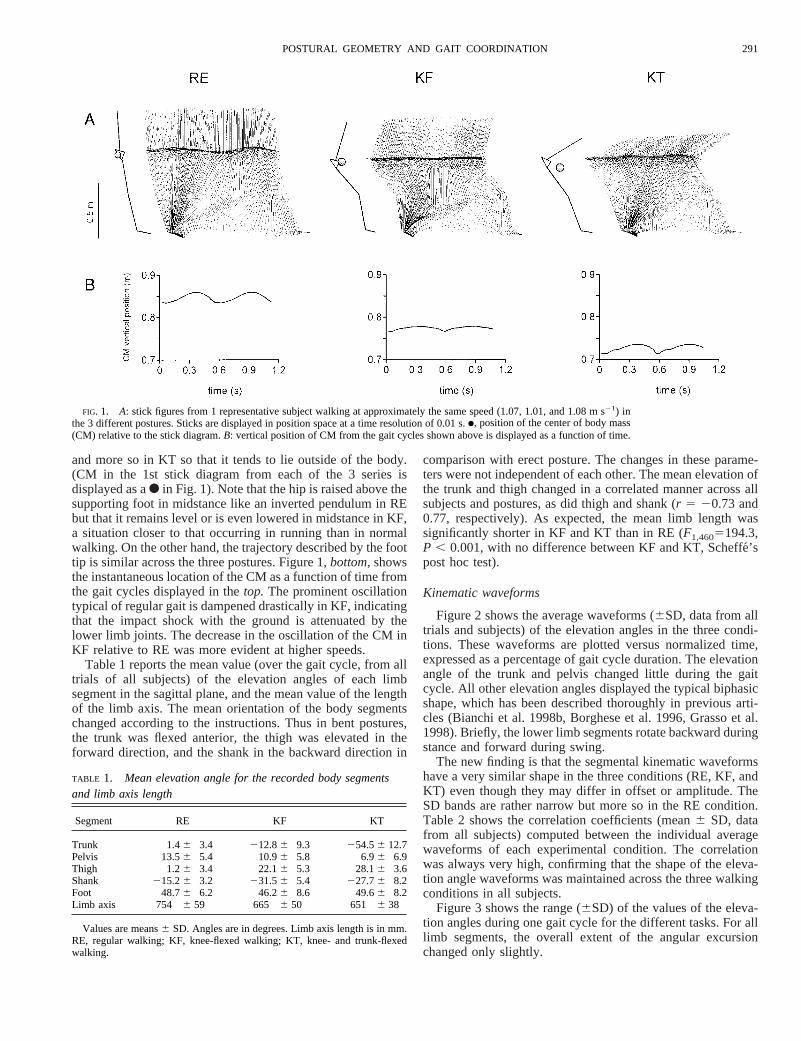
and more so in KT so that it tends to lie outside of the body.(CM in the 1st stick diagram from each of the 3 series isdisplayed as aF in Fig. 1). Note that the hip is raised above thesupporting foot in midstance like an inverted pendulum in REbut that it remains level or is even lowered in midstance in KF,a situation closer to that occurring in running than in normalwalking. On the other hand, the trajectory described by the foottip is similar across the three postures. Figure 1,bottom,showsthe instantaneous location of the CM as a function of time fromthe gait cycles displayed in thetop. The prominent oscillationtypical of regular gait is dampened drastically in KF, indicatingthat the impact shock with the ground is attenuated by thelower limb joints. The decrease in the oscillation of the CM inKF relative to RE was more evident at higher speeds.
Table 1 reports the mean value (over the gait cycle, from alltrials of all subjects) of the elevation angles of each limbsegment in the sagittal plane, and the mean value of the lengthof the limb axis. The mean orientation of the body segmentschanged according to the instructions. Thus in bent postures,the trunk was flexed anterior, the thigh was elevated in theforward direction, and the shank in the backward direction in
comparison with erect posture. The changes in these parame-ters were not independent of each other. The mean elevation ofthe trunk and thigh changed in a correlated manner across allsubjects and postures, as did thigh and shank (r 5 20.73 and0.77, respectively). As expected, the mean limb length wassignificantly shorter in KF and KT than in RE (F1,4605194.3,P , 0.001, with no difference between KF and KT, Scheffe´’spost hoc test).
Kinematic waveforms
Figure 2 shows the average waveforms (6SD, data from alltrials and subjects) of the elevation angles in the three condi-tions. These waveforms are plotted versus normalized time,expressed as a percentage of gait cycle duration. The elevationangle of the trunk and pelvis changed little during the gaitcycle. All other elevation angles displayed the typical biphasicshape, which has been described thoroughly in previous arti-cles (Bianchi et al. 1998b, Borghese et al. 1996, Grasso et al.1998). Briefly, the lower limb segments rotate backward duringstance and forward during swing.
The new finding is that the segmental kinematic waveformshave a very similar shape in the three conditions (RE, KF, andKT) even though they may differ in offset or amplitude. TheSD bands are rather narrow but more so in the RE condition.Table 2 shows the correlation coefficients (mean6 SD, datafrom all subjects) computed between the individual averagewaveforms of each experimental condition. The correlationwas always very high, confirming that the shape of the eleva-tion angle waveforms was maintained across the three walkingconditions in all subjects.
Figure 3 shows the range (6SD) of the values of the eleva-tion angles during one gait cycle for the different tasks. For alllimb segments, the overall extent of the angular excursionchanged only slightly.
FIG. 1. A: stick figures from 1 representative subject walking at approximately the same speed (1.07, 1.01, and 1.08 m s21) inthe 3 different postures. Sticks are displayed in position space at a time resolution of 0.01 s.●, position of the center of body mass(CM) relative to the stick diagram.B: vertical position of CM from the gait cycles shown above is displayed as a function of time.
TABLE 1. Mean elevation angle for the recorded body segmentsand limb axis length
Segment RE KF KT
Trunk 1.46 3.4 212.86 9.3 254.56 12.7Pelvis 13.56 5.4 10.96 5.8 6.96 6.9Thigh 1.26 3.4 22.16 5.3 28.16 3.6Shank 215.26 3.2 231.56 5.4 227.76 8.2Foot 48.76 6.2 46.26 8.6 49.66 8.2Limb axis 754 6 59 665 6 50 651 6 38
Values are means6 SD. Angles are in degrees. Limb axis length is in mm.RE, regular walking; KF, knee-flexed walking; KT, knee- and trunk-flexedwalking.
291POSTURAL GEOMETRY AND GAIT COORDINATION
Intersegmental coordination—time domain
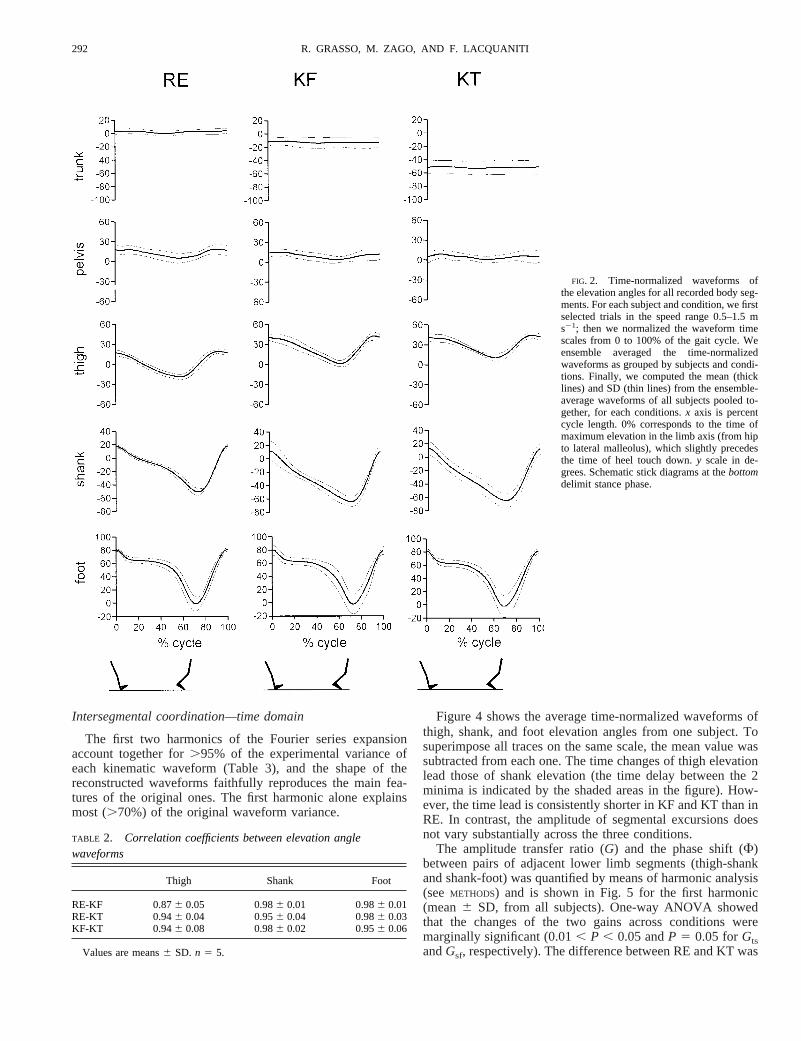
The first two harmonics of the Fourier series expansionaccount together for.95% of the experimental variance ofeach kinematic waveform (Table 3), and the shape of thereconstructed waveforms faithfully reproduces the main fea-tures of the original ones. The first harmonic alone explainsmost (.70%) of the original waveform variance.
Figure 4 shows the average time-normalized waveforms ofthigh, shank, and foot elevation angles from one subject. Tosuperimpose all traces on the same scale, the mean value wassubtracted from each one. The time changes of thigh elevationlead those of shank elevation (the time delay between the 2minima is indicated by the shaded areas in the figure). How-ever, the time lead is consistently shorter in KF and KT than inRE. In contrast, the amplitude of segmental excursions doesnot vary substantially across the three conditions.
The amplitude transfer ratio (G) and the phase shift (F)between pairs of adjacent lower limb segments (thigh-shankand shank-foot) was quantified by means of harmonic analysis(seeMETHODS) and is shown in Fig. 5 for the first harmonic(mean6 SD, from all subjects). One-way ANOVA showedthat the changes of the two gains across conditions weremarginally significant (0.01, P , 0.05 andP 5 0.05 forGtsandGsf, respectively). The difference between RE and KT was
FIG. 2. Time-normalized waveforms ofthe elevation angles for all recorded body seg-ments. For each subject and condition, we firstselected trials in the speed range 0.5–1.5 ms21; then we normalized the waveform timescales from 0 to 100% of the gait cycle. Weensemble averaged the time-normalizedwaveforms as grouped by subjects and condi-tions. Finally, we computed the mean (thicklines) and SD (thin lines) from the ensemble-average waveforms of all subjects pooled to-gether, for each conditions.x axis is percentcycle length. 0% corresponds to the time ofmaximum elevation in the limb axis (from hipto lateral malleolus), which slightly precedesthe time of heel touch down.y scale in de-grees. Schematic stick diagrams at thebottomdelimit stance phase.
TABLE 2. Correlation coefficients between elevation anglewaveforms
Thigh Shank Foot
RE-KF 0.876 0.05 0.986 0.01 0.986 0.01RE-KT 0.946 0.04 0.956 0.04 0.986 0.03KF-KT 0.946 0.08 0.986 0.02 0.956 0.06
Values are means6 SD. n 5 5.
292 R. GRASSO, M. ZAGO, AND F. LACQUANITI
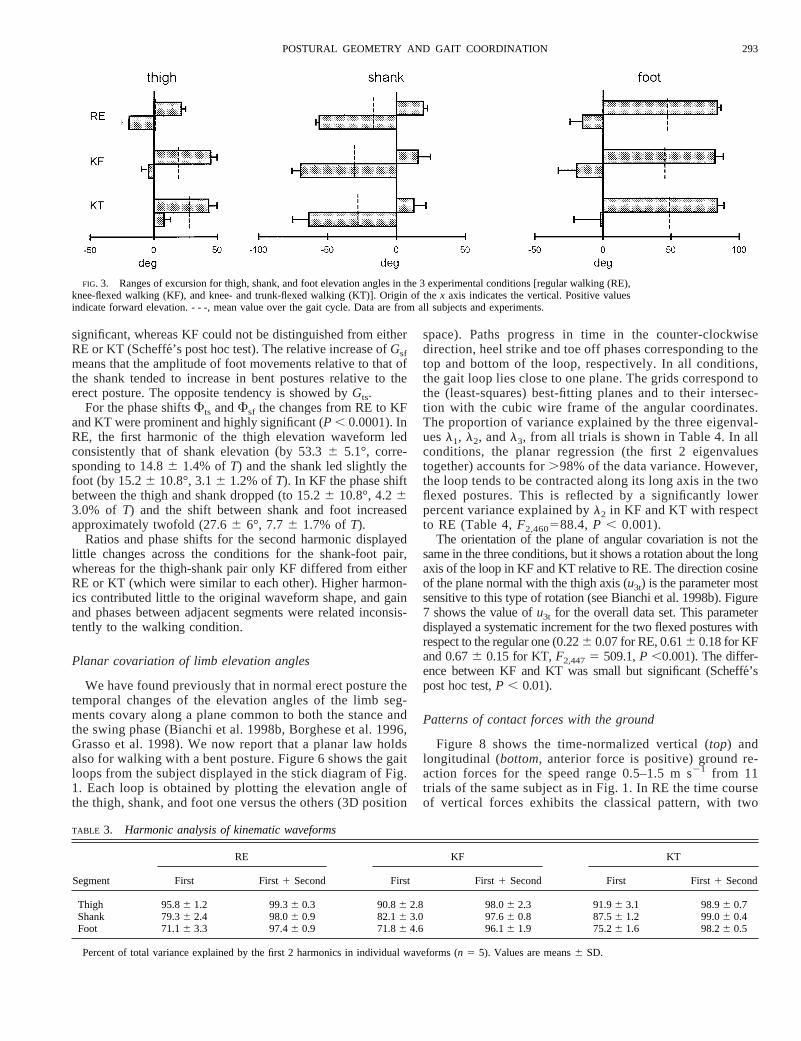
significant, whereas KF could not be distinguished from eitherRE or KT (Scheffe´’s post hoc test). The relative increase ofGsfmeans that the amplitude of foot movements relative to that ofthe shank tended to increase in bent postures relative to theerect posture. The opposite tendency is showed byGts.
For the phase shiftsFts andFsf the changes from RE to KFand KT were prominent and highly significant (P , 0.0001). InRE, the first harmonic of the thigh elevation waveform ledconsistently that of shank elevation (by 53.36 5.1°, corre-sponding to 14.86 1.4% ofT) and the shank led slightly thefoot (by 15.26 10.8°, 3.16 1.2% ofT). In KF the phase shiftbetween the thigh and shank dropped (to 15.26 10.8°, 4.263.0% of T) and the shift between shank and foot increasedapproximately twofold (27.66 6°, 7.76 1.7% ofT).
Ratios and phase shifts for the second harmonic displayedlittle changes across the conditions for the shank-foot pair,whereas for the thigh-shank pair only KF differed from eitherRE or KT (which were similar to each other). Higher harmon-ics contributed little to the original waveform shape, and gainand phases between adjacent segments were related inconsis-tently to the walking condition.
Planar covariation of limb elevation angles
We have found previously that in normal erect posture thetemporal changes of the elevation angles of the limb seg-ments covary along a plane common to both the stance andthe swing phase (Bianchi et al. 1998b, Borghese et al. 1996,Grasso et al. 1998). We now report that a planar law holdsalso for walking with a bent posture. Figure 6 shows the gaitloops from the subject displayed in the stick diagram of Fig.1. Each loop is obtained by plotting the elevation angle ofthe thigh, shank, and foot one versus the others (3D position
space). Paths progress in time in the counter-clockwisedirection, heel strike and toe off phases corresponding to thetop and bottom of the loop, respectively. In all conditions,the gait loop lies close to one plane. The grids correspond tothe (least-squares) best-fitting planes and to their intersec-tion with the cubic wire frame of the angular coordinates.The proportion of variance explained by the three eigenval-uesl1, l2, andl3, from all trials is shown in Table 4. In allconditions, the planar regression (the first 2 eigenvaluestogether) accounts for.98% of the data variance. However,the loop tends to be contracted along its long axis in the twoflexed postures. This is reflected by a significantly lowerpercent variance explained byl2 in KF and KT with respectto RE (Table 4,F2,460588.4, P , 0.001).
The orientation of the plane of angular covariation is not thesame in the three conditions, but it shows a rotation about the longaxis of the loop in KF and KT relative to RE. The direction cosineof the plane normal with the thigh axis (u3t) is the parameter mostsensitive to this type of rotation (see Bianchi et al. 1998b). Figure7 shows the value ofu3t for the overall data set. This parameterdisplayed a systematic increment for the two flexed postures withrespect to the regular one (0.226 0.07 for RE, 0.616 0.18 for KFand 0.676 0.15 for KT,F2,4475 509.1,P ,0.001). The differ-ence between KF and KT was small but significant (Scheffe´’spost hoc test,P , 0.01).
Patterns of contact forces with the ground
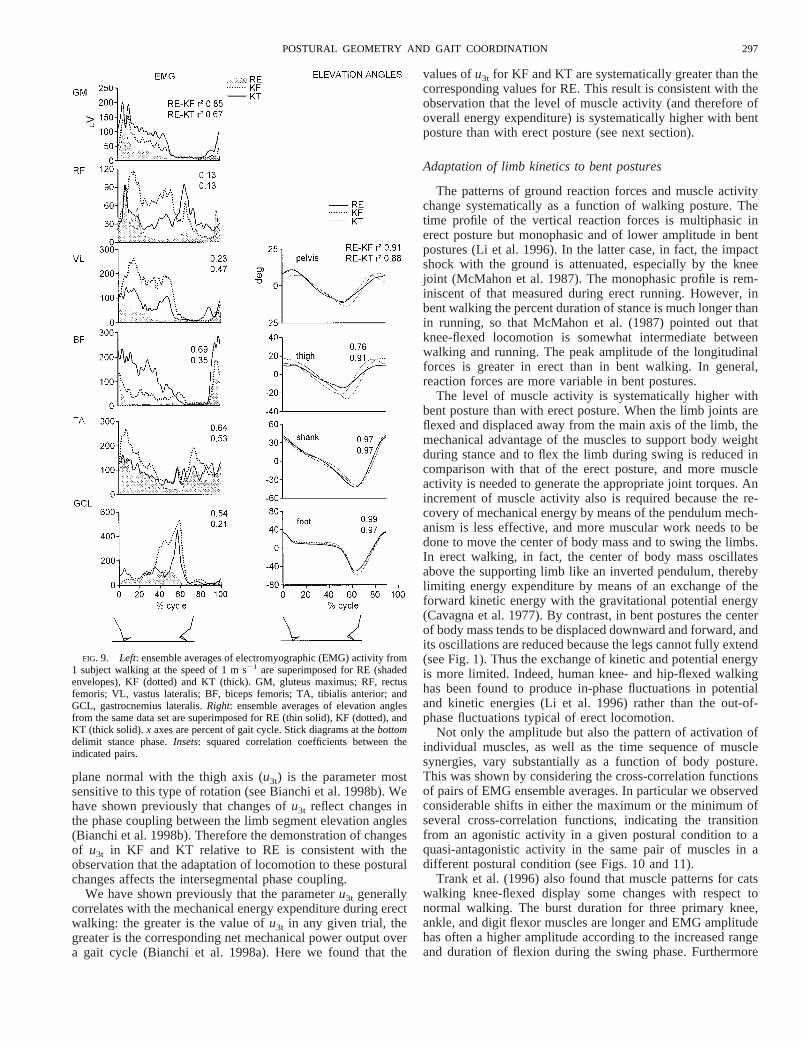
Figure 8 shows the time-normalized vertical (top) andlongitudinal (bottom, anterior force is positive) ground re-action forces for the speed range 0.5–1.5 m s21 from 11trials of the same subject as in Fig. 1. In RE the time courseof vertical forces exhibits the classical pattern, with two
FIG. 3. Ranges of excursion for thigh, shank, and foot elevation angles in the 3 experimental conditions [regular walking (RE),knee-flexed walking (KF), and knee- and trunk-flexed walking (KT)]. Origin of thex axis indicates the vertical. Positive valuesindicate forward elevation. - - -, mean value over the gait cycle. Data are from all subjects and experiments.
TABLE 3. Harmonic analysis of kinematic waveforms
Segment
RE KF KT
First First1 Second First First1 Second First First1 Second
Thigh 95.86 1.2 99.36 0.3 90.86 2.8 98.06 2.3 91.96 3.1 98.96 0.7Shank 79.36 2.4 98.06 0.9 82.16 3.0 97.66 0.8 87.56 1.2 99.06 0.4Foot 71.16 3.3 97.46 0.9 71.86 4.6 96.16 1.9 75.26 1.6 98.26 0.5
Percent of total variance explained by the first 2 harmonics in individual waveforms (n 5 5). Values are means6 SD.
293POSTURAL GEOMETRY AND GAIT COORDINATION
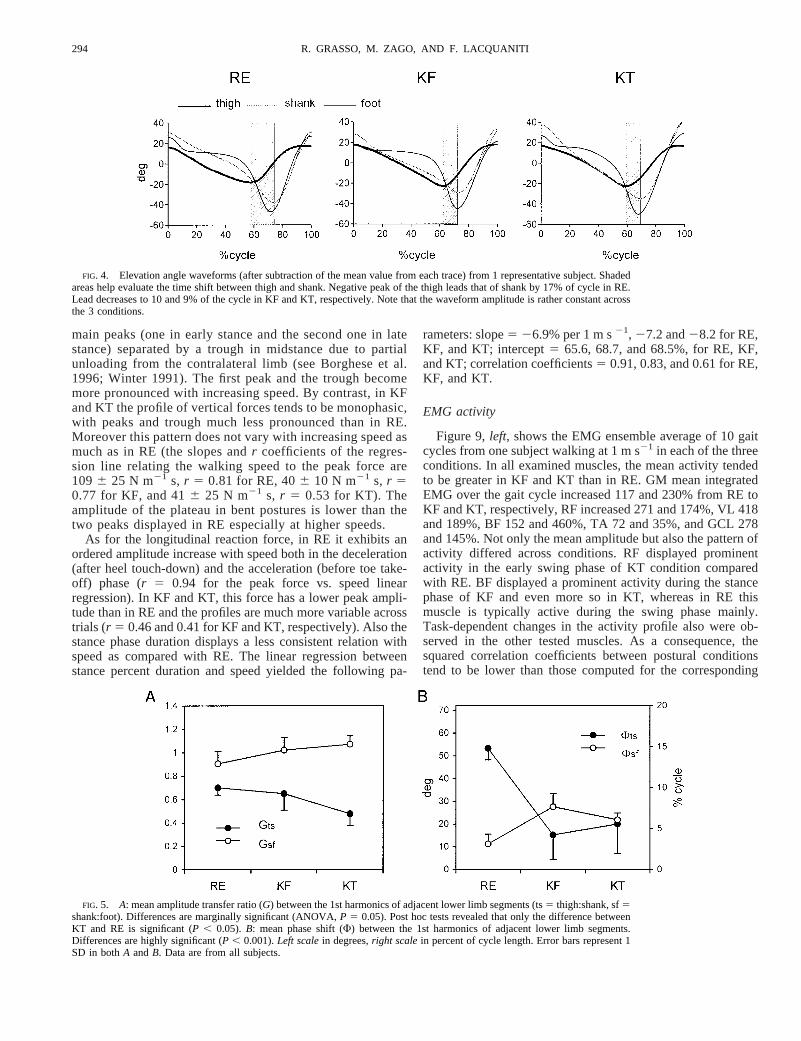
main peaks (one in early stance and the second one in latestance) separated by a trough in midstance due to partialunloading from the contralateral limb (see Borghese et al.1996; Winter 1991). The first peak and the trough becomemore pronounced with increasing speed. By contrast, in KFand KT the profile of vertical forces tends to be monophasic,with peaks and trough much less pronounced than in RE.Moreover this pattern does not vary with increasing speed asmuch as in RE (the slopes andr coefficients of the regres-sion line relating the walking speed to the peak force are1096 25 N m21 s, r 5 0.81 for RE, 406 10 N m21 s, r 50.77 for KF, and 416 25 N m21 s, r 5 0.53 for KT). Theamplitude of the plateau in bent postures is lower than thetwo peaks displayed in RE especially at higher speeds.
As for the longitudinal reaction force, in RE it exhibits anordered amplitude increase with speed both in the deceleration(after heel touch-down) and the acceleration (before toe take-off) phase (r 5 0.94 for the peak force vs. speed linearregression). In KF and KT, this force has a lower peak ampli-tude than in RE and the profiles are much more variable acrosstrials (r 5 0.46 and 0.41 for KF and KT, respectively). Also thestance phase duration displays a less consistent relation withspeed as compared with RE. The linear regression betweenstance percent duration and speed yielded the following pa-
rameters: slope5 26.9% per 1 m s21, 27.2 and28.2 for RE,KF, and KT; intercept5 65.6, 68.7, and 68.5%, for RE, KF,and KT; correlation coefficients5 0.91, 0.83, and 0.61 for RE,KF, and KT.
EMG activity
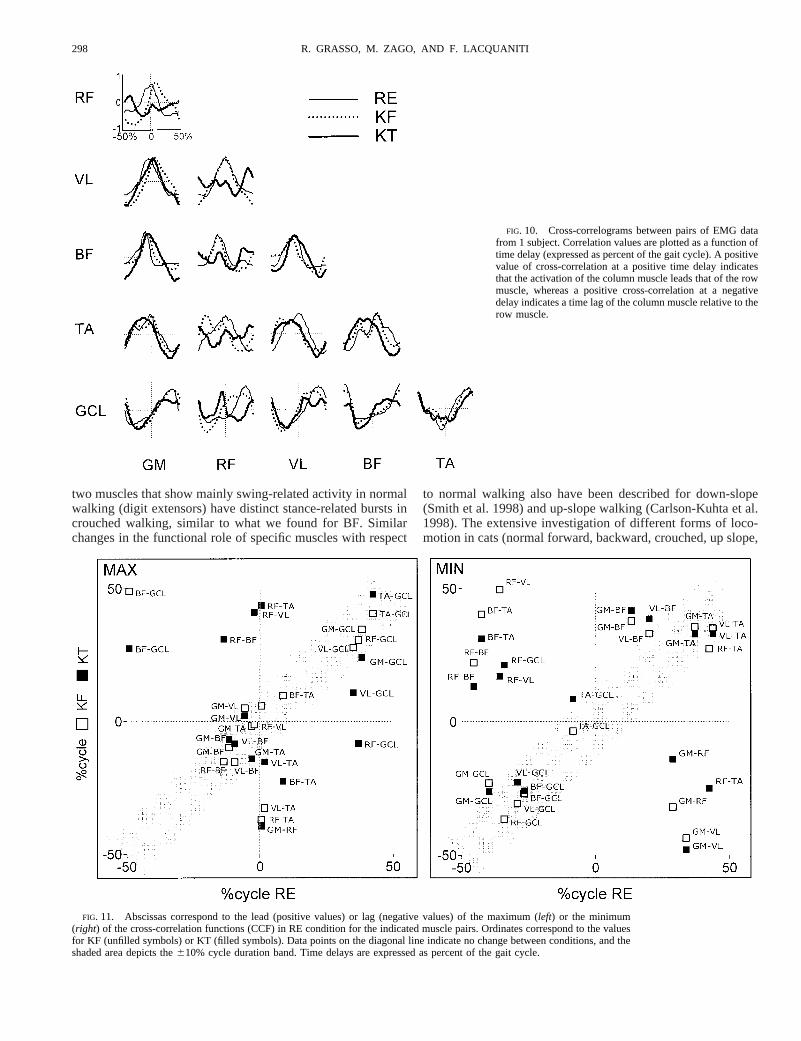
Figure 9,left, shows the EMG ensemble average of 10 gaitcycles from one subject walking at 1 m s21 in each of the threeconditions. In all examined muscles, the mean activity tendedto be greater in KF and KT than in RE. GM mean integratedEMG over the gait cycle increased 117 and 230% from RE toKF and KT, respectively, RF increased 271 and 174%, VL 418and 189%, BF 152 and 460%, TA 72 and 35%, and GCL 278and 145%. Not only the mean amplitude but also the pattern ofactivity differed across conditions. RF displayed prominentactivity in the early swing phase of KT condition comparedwith RE. BF displayed a prominent activity during the stancephase of KF and even more so in KT, whereas in RE thismuscle is typically active during the swing phase mainly.Task-dependent changes in the activity profile also were ob-served in the other tested muscles. As a consequence, thesquared correlation coefficients between postural conditionstend to be lower than those computed for the corresponding
FIG. 4. Elevation angle waveforms (after subtraction of the mean value from each trace) from 1 representative subject. Shadedareas help evaluate the time shift between thigh and shank. Negative peak of the thigh leads that of shank by 17% of cycle in RE.Lead decreases to 10 and 9% of the cycle in KF and KT, respectively. Note that the waveform amplitude is rather constant acrossthe 3 conditions.
FIG. 5. A: mean amplitude transfer ratio (G) between the 1st harmonics of adjacent lower limb segments (ts5 thigh:shank, sf5shank:foot). Differences are marginally significant (ANOVA,P 5 0.05). Post hoc tests revealed that only the difference betweenKT and RE is significant (P , 0.05). B: mean phase shift (F) between the 1st harmonics of adjacent lower limb segments.Differences are highly significant (P , 0.001).Left scalein degrees,right scalein percent of cycle length. Error bars represent 1SD in bothA andB. Data are from all subjects.
294 R. GRASSO, M. ZAGO, AND F. LACQUANITI

changes in the elevation angles (right). This was true for allsubjects.
Not only the patterns of activation of individual muscles butalso the time sequence of activation of different muscles, ormuscle synergies, varied substantially as a function of bodyposture. To quantify this kind of synergies, we computed theCCF between pairs of EMG ensemble averages. The CCFs forthe three tasks are superimposed in Fig. 10. The patterns ofmuscle synergies differ across conditions as indicated by thedifferent shape of the CCF. For instance, in RE and KF thepeak in the pair RF-VL occurs at;0% of the gait cycle,indicating agonistic activity. By contrast, in KT the peak oc-curs at;40% lead, indicating quasi-antagonistic activity inthese two muscles. Most other pairings of muscles exhibitsignificant time shifts in either the maximum or the minimumof the CCF in KF and KT conditions relative to the REcondition. This is shown by the scatter plot of these values inFig. 11. Several data points (13/30 and 18/30 for the maximumand minimum, respectively) fall outside the610% cycle du-ration band (shaded area).
D I S C U S S I O N
Kinematic waveforms
The purpose of the present study was to verify how walkingwith a bent posture affects the generation of gait motor pat-terns. In bent postures (KF and KT), the lower limbs and thetrunk are flexed as compared with the erect posture (RE). Inspite of the drastic changes in postural geometry, the oscilla-tions of each limb segment relative to the vertical are verysimilar across different postures.
The adaptation of the intersegmental coordination to thedifferent postures is achieved by changing the phase delaysbetween the motion of different limb segments. Thus the phaselead of thigh oscillations relative to shank oscillations is sys-tematically shorter with bent postures than with an erect pos-ture, whereas the phase lead of the shank relative to the foot
oscillations is longer in the former than in the latter. As a result,there is an approximately constant phase de´calage (of;20°corresponding to'5% of the gait cycle) between adjacent limbsegments in the proximal-to-distal direction in bent walking.The behavior of a flexed lower limb then can be equated to thatof a rolling wheel with the pivot corresponding to the kneejoint.
Somewhat similar results have been reported previously inthe comparison of level walking versus crouched walking incats (Trank et al. 1996). (The latter is an attitude typical ofstalking behavior in this species.) The profiles of hindlimbkinematics were rather similar between the two forms of loco-motion either for the hip, knee, ankle, or the metatarso-pha-langeal joint angles (see Fig. 3 of Trank et al. 1996) in spite ofprominent changes in their mean values and ranges of excur-sion. The timing of motion reversals (from flexion to exten-sion) differed at some joints.
Therefore postural adaptation acts on the same variable,intersegmental kinematic phase, which has been shown previ-ously to be centrally modified as a function of changes inwalking speed (Bianchi et al. 1998b) or walking direction(Grasso et al. 1998). Intersegmental phase plays a role ofglobal control variable similar to that previously advocated forthe network of coupled oscillators involved in the generation oflocomotion (see Grillner 1981; Pearson 1993; Rossignol 1996).According to the kinematic view we have exposed in theintroduction, each unit oscillator would directly control a limbsegment, alternately generating forward and backward oscilla-tions of the segment. Intersegmental coordination would beachieved by coupling unit oscillators with a variable phase.Variable coupling could result, for instance, by changing thesynaptic strength (or polarity) of the relative spinal connec-tions. Supraspinal centers may drive or modulate functionalsets of coordinating spinal interneurons to generate differentwalking modes. We shall take up the issue of the mechanismsof postural modulation of the coordinating network in a sub-sequent section.
Planar covariation of elevation angles
The changes in the elevation angles of the thigh, shank, andfoot covary along a plane common to both the stance and theswing phase. More than 98% of the data variance is explainedby the planar regression in all postural conditions. However,the orientation of the plane differs across conditions: the planeis rotated about the long axis of the loop with bent postures ascompared with the erect posture. The direction cosine of the
TABLE 4. Percent variance explained by the eigenvalues of theelevation angle covariance matrix
RE KF KT
l1 87.26 2.8 90.06 3.7 91.56 2.3l2 11.66 2.7 8.46 3.9 7.16 2.3l3 1.26 0.4 1.66 0.6 1.46 0.5
Values are means6 SD.
FIG. 6. Mean gait loops from 1 subject acrossthe 3 experimental conditions. A loop represents1 gait cycle and is obtained by plotting theaverage thigh waveform vs. the shank and footwaveforms (after mean values subtraction).Paths progress in time in the counterclockwisedirection, heel contact and toe-off phases corre-sponding approximately to the top and bottom ofthe loop, respectively. Interpolation planes resultfrom orthogonal planar regression fitting. Notethat the loop lies close to the plane in all condi-tions but it displays different orientations in KFand KT vs. RE.
295POSTURAL GEOMETRY AND GAIT COORDINATION
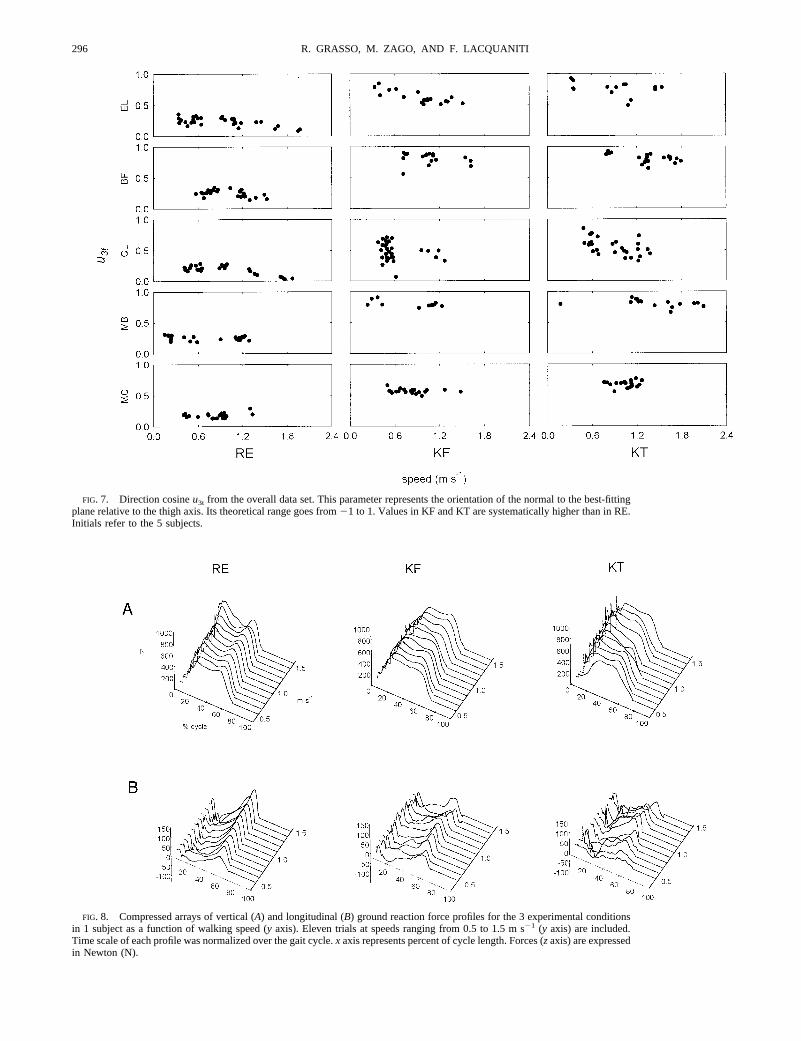
FIG. 7. Direction cosineu3t from the overall data set. This parameter represents the orientation of the normal to the best-fittingplane relative to the thigh axis. Its theoretical range goes from21 to 1. Values in KF and KT are systematically higher than in RE.Initials refer to the 5 subjects.
FIG. 8. Compressed arrays of vertical (A) and longitudinal (B) ground reaction force profiles for the 3 experimental conditionsin 1 subject as a function of walking speed (y axis). Eleven trials at speeds ranging from 0.5 to 1.5 m s21 (y axis) are included.Time scale of each profile was normalized over the gait cycle.x axis represents percent of cycle length. Forces (zaxis) are expressedin Newton (N).
296 R. GRASSO, M. ZAGO, AND F. LACQUANITI
plane normal with the thigh axis (u3t) is the parameter mostsensitive to this type of rotation (see Bianchi et al. 1998b). Wehave shown previously that changes ofu3t reflect changes inthe phase coupling between the limb segment elevation angles(Bianchi et al. 1998b). Therefore the demonstration of changesof u3t in KF and KT relative to RE is consistent with theobservation that the adaptation of locomotion to these posturalchanges affects the intersegmental phase coupling.
We have shown previously that the parameteru3t generallycorrelates with the mechanical energy expenditure during erectwalking: the greater is the value ofu3t in any given trial, thegreater is the corresponding net mechanical power output overa gait cycle (Bianchi et al. 1998a). Here we found that the
values ofu3t for KF and KT are systematically greater than thecorresponding values for RE. This result is consistent with theobservation that the level of muscle activity (and therefore ofoverall energy expenditure) is systematically higher with bentposture than with erect posture (see next section).
Adaptation of limb kinetics to bent postures
The patterns of ground reaction forces and muscle activitychange systematically as a function of walking posture. Thetime profile of the vertical reaction forces is multiphasic inerect posture but monophasic and of lower amplitude in bentpostures (Li et al. 1996). In the latter case, in fact, the impactshock with the ground is attenuated, especially by the kneejoint (McMahon et al. 1987). The monophasic profile is rem-iniscent of that measured during erect running. However, inbent walking the percent duration of stance is much longer thanin running, so that McMahon et al. (1987) pointed out thatknee-flexed locomotion is somewhat intermediate betweenwalking and running. The peak amplitude of the longitudinalforces is greater in erect than in bent walking. In general,reaction forces are more variable in bent postures.
The level of muscle activity is systematically higher withbent posture than with erect posture. When the limb joints areflexed and displaced away from the main axis of the limb, themechanical advantage of the muscles to support body weightduring stance and to flex the limb during swing is reduced incomparison with that of the erect posture, and more muscleactivity is needed to generate the appropriate joint torques. Anincrement of muscle activity also is required because the re-covery of mechanical energy by means of the pendulum mech-anism is less effective, and more muscular work needs to bedone to move the center of body mass and to swing the limbs.In erect walking, in fact, the center of body mass oscillatesabove the supporting limb like an inverted pendulum, therebylimiting energy expenditure by means of an exchange of theforward kinetic energy with the gravitational potential energy(Cavagna et al. 1977). By contrast, in bent postures the centerof body mass tends to be displaced downward and forward, andits oscillations are reduced because the legs cannot fully extend(see Fig. 1). Thus the exchange of kinetic and potential energyis more limited. Indeed, human knee- and hip-flexed walkinghas been found to produce in-phase fluctuations in potentialand kinetic energies (Li et al. 1996) rather than the out-of-phase fluctuations typical of erect locomotion.
Not only the amplitude but also the pattern of activation ofindividual muscles, as well as the time sequence of musclesynergies, vary substantially as a function of body posture.This was shown by considering the cross-correlation functionsof pairs of EMG ensemble averages. In particular we observedconsiderable shifts in either the maximum or the minimum ofseveral cross-correlation functions, indicating the transitionfrom an agonistic activity in a given postural condition to aquasi-antagonistic activity in the same pair of muscles in adifferent postural condition (see Figs. 10 and 11).
Trank et al. (1996) also found that muscle patterns for catswalking knee-flexed display some changes with respect tonormal walking. The burst duration for three primary knee,ankle, and digit flexor muscles are longer and EMG amplitudehas often a higher amplitude according to the increased rangeand duration of flexion during the swing phase. Furthermore
FIG. 9. Left: ensemble averages of electromyographic (EMG) activity from1 subject walking at the speed of 1 m s21 are superimposed for RE (shadedenvelopes), KF (dotted) and KT (thick). GM, gluteus maximus; RF, rectusfemoris; VL, vastus lateralis; BF, biceps femoris; TA, tibialis anterior; andGCL, gastrocnemius lateralis.Right: ensemble averages of elevation anglesfrom the same data set are superimposed for RE (thin solid), KF (dotted), andKT (thick solid).x axes are percent of gait cycle. Stick diagrams at thebottomdelimit stance phase.Insets: squared correlation coefficients between theindicated pairs.
297POSTURAL GEOMETRY AND GAIT COORDINATION
two muscles that show mainly swing-related activity in normalwalking (digit extensors) have distinct stance-related bursts incrouched walking, similar to what we found for BF. Similarchanges in the functional role of specific muscles with respect
to normal walking also have been described for down-slope(Smith et al. 1998) and up-slope walking (Carlson-Kuhta et al.1998). The extensive investigation of different forms of loco-motion in cats (normal forward, backward, crouched, up slope,
FIG. 10. Cross-correlograms between pairs of EMG datafrom 1 subject. Correlation values are plotted as a function oftime delay (expressed as percent of the gait cycle). A positivevalue of cross-correlation at a positive time delay indicatesthat the activation of the column muscle leads that of the rowmuscle, whereas a positive cross-correlation at a negativedelay indicates a time lag of the column muscle relative to therow muscle.
FIG. 11. Abscissas correspond to the lead (positive values) or lag (negative values) of the maximum (left) or the minimum(right) of the cross-correlation functions (CCF) in RE condition for the indicated muscle pairs. Ordinates correspond to the valuesfor KF (unfilled symbols) or KT (filled symbols). Data points on the diagonal line indicate no change between conditions, and theshaded area depicts the610% cycle duration band. Time delays are expressed as percent of the gait cycle.
298 R. GRASSO, M. ZAGO, AND F. LACQUANITI

and down slope) (reviewed in Zernicke and Smith 1996) hasmade clear that the coupling between muscle groups is animportant functional variable and may be modified centrallydepending on the behavioral context. Even robust stance-sus-taining synergies, which rely on the excitatory coupling be-tween hip and knee extensor units, may be switched off inparticular cases (i.e., in down-slope walking). To explain thesetransitions to different synergies without dropping the schemaof an array of joint-related CPG units (Grillner 1981), Zernickeand Smith (1996; see also Prochazka 1996) proposed that therule for setting interunit connections may be issued by su-praspinal control following conditional statements related tothe behavioral goal.
Our view is that, in response to the kinematic referencesignals, the appropriate muscle synergies are determined in aflexible manner to fit the current mechanical constraints. In-deed, the relationship between muscle activity and whole bodymechanics seems too variable and complex to allow a directcontrol of the latter without an intermediate kinematic repre-sentation. Neurophysiological data indicate that intermediatekinematic representations may be found in both premotor(Bizzi et al. 1991) and proprioceptive sensory areas of thespinal cord (Bosco et al. 1996) as well as in somatosensory andmotor cortices (Helms Tillery et al. 1996; Lacquaniti et al.1995; Scott and Kalaska 1997).
Integrated control of gait and posture
Postural adaptation conserves the general kinematic wave-forms with an appropriately tuned intersegmental phase andallocates specific patterns of muscle activity as a function ofthe required kinematic coordination. Integrated control of gaitand posture is made possible because these two motor func-tions share some common organizational principles (Lacqua-niti et al. 1997; Massion 1992). First, the frame of reference forthe kinematic coordination for both postural responses andlocomotion seems to be anchored to the vertical. Second, acontrol of the position of the center of body mass for static ordynamic equilibrium is involved in both gait (Cavagna et al.1977) and posture (Massion 1992). Also the planar law ofintersegmental kinematic coordination applies to both tasks.The planar law involved in postural responses has been re-ported in previous work (Lacquaniti and Maioli 1994a,b). Thechanges in the geometric configuration of the forelimbs andhindlimbs in cats pitched by variable tilts of the support plat-form lie close to one plane. This planar covariation is notaffected by adding loads that shift the animal’s center of mass.Intersegmental coordination of lower limbs and trunk also hasbeen described for human postural responses evoked by per-turbations of the support platform (Nashner and McCollum1985) or by axial bending movements (Massion 1992). It is notsurprising to find that similar laws of intersegmental coordina-tion apply to the control of posture and locomotion. Locomo-tion must assure a forward progression compatible with dy-namic equilibrium, adapting to potentially destabilizing factors(e.g., changes in body posture or load, uneven terrain, obsta-cles, etc.) in an anticipatory fashion by means of coordinatedsynergies of upper limbs, trunk, and lower limbs (Dietz et al.1987; Hirschfeld and Forssberg 1991).
The concept of an integrated control of posture and locomo-tion also stems from neurophysiological data. The stimulation
of specific areas in the brain stem and hypothalamus in freelymoving cats causes the animal to adopt different locomotor andpostural styles (Mori et al. 1989). Flexed locomotion can beevoked in cats by stimulating the lateral hypothalamus (Mori etal. 1989). Also selective lesions in Deiters’ nucleus result in asignificant decrease in extensor muscle activity during loco-motion (Orlovsky 1972). Repetitive stimulation of medial lon-gitudinal fasciculus neurons may disrupt fictive locomotorrhythms (Floeter et al. 1993; Gossard et al. 1996). The linkbetween posture and gait control is also mediated throughafferent feedback. Thus Shik and Orlovsky (1976) proposedthat vestibulospinal and reticulospinal pathways carrying in-formation from both the sensory feedback and from the state ofthe spinal circuitry may operate during locomotion to “sculpt”the output patterns generated in the segmental CPGs (see alsoArmstrong 1988; Hasan and Stuart 1988).
A specific role in setting the spatial framework for thecontrol of the postural geometry of the trunk and the coordi-nation of lower limb segments is provided by the basal ganglia(Garcia-Rill 1986). A recent study addressed this issue inpatients with Parkinson’s disease (Grasso et al. 1999). Patientscould be switchedON by means of either a D1-D2 receptoragonistic drug (apomorphine) or by globus pallidum internum(GPi) electrical stimulation. It was found that the inclination ofthe trunk with respect to the vertical, the waveforms of theelevation angles, and the planar law of angular covariationchange all in parallel in the transition from theOFF to the ON
condition.
We thank D. Prissinotti for skillful technical help with the experiments.The financial support of Telethon-Italy (Grant 1159) is gratefully acknowl-
edged.Address for reprint requests: F. Lacquaniti, IRCCS Santa Lucia, via
Ardeatina 306, 00179 Rome, Italy.
Received 6 May 1999; accepted in final form 14 September 1999.
REFERENCES
ARMSTRONG, D. B. Review lecture: the supraspinal control of mammalianlocomotion.J. Physiol. (Lond.)405: 1–37, 1988.
BIANCHI, L., ANGELINI, D., AND LACQUANITI, F. Individual characteristics ofhuman walking mechanics.Pflugers Arch.436: 343–356, 1998a.
BIANCHI, L., ANGELINI, D., ORANI, G. P., AND LACQUANITI, F. Kinematiccoordination in human gait: relation to mechanical energy cost.J. Neuro-physiol.79: 2155–2170, 1998b.
BIEWENER, A. A. Biomechanics of mammalian terrestrial locomotion.Science250: 1097–1103, 1990.
BIZZI, E., MUSSA-IVALDI , F. A., AND GISZTER, S. Computations underlying theexecution of movement: a biological perspective.Science253: 287–291,1991.
BORGHESE, N. A., BIANCHI, L., AND LACQUANITI, F. Kinematic determinants ofhuman locomotion.J. Physiol. (Lond.)494: 863–879, 1996.
BOSCO, G., RANKIN , A., AND POPPELE, R. A representation of passive hindlimbpostures in cat spinocerebellar activity.J. Neurophysiol.76: 715–726, 1996.
BURLEIGH, A. L., HORAK, F. B., AND MALOUIN, F. Modification of posturalresponses and step initiation: evidence for goal-directed postural interac-tions.J. Neurophysiol.72: 2892–2902, 1994.
CARLSON-KUHTA, P., TRANK, V. T., AND SMITH, J. Forms of forward quadru-pedal locomotion. II. A comparison of posture, hindlimb kinematics, andmotor patterns for upslope and level walking.J. Neurophysiol.79: 1687–1701, 1998.
CAVAGNA , G., HEGLUND, N. C., AND TAYLOR, C. R. Mechanical work interrestrial locomotion: two basic mechanisms for minimizing energy expen-diture. Am. J. Physiol.233 (Regulatory Integrative Comp. Physiol.2):R243–R261, 1977.
CROMPTON, R. H., YU, L., WEIJIE, W., GUNTHER, M., AND SAVAGE, R. Themechanical effectiveness of erect and “bent-hip bent-knee” bipedal walkingin Australopithecus afarensis. J. Hum. Evol.35: 55–74, 1998.
299POSTURAL GEOMETRY AND GAIT COORDINATION

D’A MICO, M. AND FERRIGNO, G. Technique for the evaluation of derivativesfrom noisy biomechanical displacement data using a model-based band-width-selection procedure.Med. Biol. Eng. Comput.28: 407–415, 1990.
DARLING, W. G. AND MILLER, G. F. Perception of arm orientation in three-dimensional space.Exp. Brain Res.102: 495–502, 1995.
DIETZ, V., QUINTERN, J., AND SILLEM , M. Stumbling reactions in man: signif-icance of proprioceptive and pre-programmed mechanisms.J. Physiol.(Lond.)368: 149–163, 1987.
FERRIGNO, G., BORGHESE, N. A., AND PEDOTTI, A. Pattern recognition in 3Dautomatic human motion analysis.ISPRS J. Photogrammetry Remote Sens-ing 45: 227–246, 1990.
FLOETER, M. K., SHOLOMENKO, G. N., GOSSARD, J. P., AND BURKE, R. E.Disynaptic excitation from the medial longitudinal fasciculus to lumbosacralmotoneurons: modulation by repetitive activation, descending pathways,and locomotion.Exp. Brain Res.92: 407–419, 1993.
GARCIA-RILL , E. The basal ganglia and the locomotor regions.Brain Res. Rev.11: 47–63, 1986.
GOSSARD, J. P., FLOETER, M. K., DEGTYARENKO, A. M., SIMON, E. S., AND
BURKE, R. E. Disynaptic vestibulospinal and reticulospinal excitation in catlumbosacral motoneurons: modulation during fictive locomotion.Exp. BrainRes.109: 277–288, 1996.
GRASSO, R., BIANCHI, L., AND LACQUANITI, F. Motor patterns for human gait:backward versus forward locomotion.J. Neurophysiol.80: 1868–1886,1998.
GRASSO, R., PEPPE, A., STRATTA, F., ANGELINI, D., ZAGO, M., STANZIONE, P.,AND LACQUANITI, F. Basal ganglia and gait control: apomorphine adminis-tration and internal pallidum stimulation in Parkinson’s disease.Exp. BrainRes.126: 139–148, 1999.
GRILLNER, S. Control of locomotion in bipeds, tetrapods, and fish. In:Hand-book of Physiology. The Nervous System. Motor Control.Bethesda, MD:Am. Physiol. Soc., 1981, sect. 1, vol. 2, part 1, p. 1179–1236.
HASAN, Z. AND STUART, D. G. Animal solutions to problems of movementcontrol: the role of proprioceptors.Annu. Rev. Neurosci.11: 199–223, 1988.
HELMS TILLERY, S. I., SOECHTING, J. F., AND EBNER, T. J. Somatosensorycortical activity in relation to arm posture: nonuniform spatial tuning.J. Neurophysiol.76: 2423–2438, 1996.
HIRSCHFELD, H. AND FORSSBERG, H. Phase-dependent modulations of antici-patory postural activity during human locomotion.J. Neurophysiol.66:12–19, 1991.
JANKOWSKA, E. AND EDGLEY, S. Interactions between pathways controllingposture and gait at the level of spinal interneurones in the cat.Prog BrainRes.97: 161–171, 1993
LACQUANITI, F., GRASSO, R., AND ZAGO, M. Motor patterns for walking.NewsPhysiol. Sci.14: 168–174, 1999.
LACQUANITI, F., GUIGON, E., BIANCHI, L., FERRAINA, S., AND CAMINITI , R.Representing spatial information for limb movement: role of area 5 in themonkey.Cereb. Cortex5: 391–409, 1995.
LACQUANITI, F. AND MAIOLI , C. Coordinate transformations in the control ofcat posture.J. Neurophysiol.72: 1496–1515, 1994a.
LACQUANITI, F. AND MAIOLI , C. Independent control of limb position andcontact forces in cat posture.J. Neurophysiol.72: 1476–1495, 1994b.
LACQUANITI, F., MAIOLI , C., BORGHESE, N., AND BIANCHI, L. Posture andmovement: coordination and control.Arch. Ital. Biol. 135: 353–367, 1997.
LI, Y., CROMPTON, R. H., ALEXANDER, R. M., GUNTHER, M. M., AND WANG,W. J. Characteristics of ground reaction forces in normal and chimpanzee-like bipedal walking by humans.Folia Primatol. (Basel)66: 137–159, 1996.
MASSION, J. Movement, posture and equilibrium: interaction and coordination.Prog. Neurobiol.38: 35–56, 1992.
MASSION, J., POPOV, K., FABRE, J. C., RAGE, P.,AND GURFINKEL, V. Is the erectposture in microgravity based on the control of trunk orientation or center ofmass position?Exp. Brain Res.114: 384–389, 1997.
MCMAHON, T. A., VALIANT , G., AND FREDERICK, E. C. Groucho running.J. Appl. Physiol.62: 2326–2337, 1987.
MORI, S. Integration of posture and locomotion in acute decerebrate cats inawake, freely moving cats.Prog. Neurobiol.28: 161–195, 1987.
MORI, S., KAWAHARA , K., AND SAKAMOTO, T. Supraspinal aspects of locomo-tion in the mesencephalic cat.Symp. Soc. Exp. Biol.37: 445–468, 1983.
MORI, S., NISHIMURA, H., KURAKAMI , C., YAMAMURA , T., AND AOKI, M.Controlled locomotion in the mesencephalic cat: distribution of facilitatoryand inhibitory regions within pontine tegmentum.J Neurophysiol.41:1580–1591, 1978.
MORI, S., SAKAMOTO, T., OHTA, Y., TAKAKUSAKI , K., AND MATSUYAMA , K.Site-specific postural and locomotor changes evoked in awake, freely mov-ing intact cats by stimulating the brainstem.Brain Res.505: 66–74, 1989.
MOUCHNINO, L., AURENTY, R., MASSION, J., AND PEDOTTI, A. Is the trunk areference frame for calculating leg position?Neuroreport4: 125–127, 1993.
NASHNER, L. M. AND MCCOLLUM, G. The organization of human posturalmovements: a formal basis and experimental synthesis.Behav. Brain Sci.8:135–172, 1985.
ORLOVSKY, G. N. The effect of different descending systems on flexor andextensor activity during locomotion.Brain Res.40: 359–371, 1972.
PEARSON, K. G. Common principles of motor control in vertebrates andinvertebrates.Annu. Rev. Neurosci.16: 265–297, 1993.
PROCHAZKA, A. The fuzzy logic of visuomotor control.Can. J. Physiol.Pharmacol.74: 456–462, 1996.
ROSSIGNOL, S. Neural control of stereotypic limb movements. In:Handbook ofPhysiology. Exercise: Regulation and Integration of Multiple Systems.Washington, DC: Am. Physiol. Soc., 1996, sect. 12, p. 173–216.
SCOTT, S. H.AND KALASKA , J. F. Reaching movements with similar hand pathsbut different arm orientations. I. Activity of individual cells in motor cortex.J. Neurophysiol.77: 826–852, 1997.
SHEN, L. AND POPPELE, R. E. Kinematic analysis of cat hindlimb stepping.J. Neurophysiol.74: 2266–2280, 1995.
SHIK, M. L. AND ORLOVSKY, G. N. Neurophysiology of locomotor automatism.Physiol. Rev.56: 456–501, 1976.
SMITH, J. L., CARLSON-KUTHA, P., AND TRANK, T. V. Forms of forwardquadrupedal locomotion. III. A comparison of posture, hindlimb kinematics,and motor patterns for downslope and level walking.J. Neurophysiol.79:1702–1716, 1998.
SPOOR, F., WOOD, B., AND ZONNEVELD, F. Implications of early hominidlabyrinthine morphology for evolution of human bipedal locomotion.Na-ture 369: 645–648, 1994.
TRANK, T. V., CHEN C., AND SMITH, J. L. Forms of forward quadrupedallocomotion. I. A comparison of posture, hindlimb kinematics, and motorpatterns for normal and crouched walking.J. Neurophysiol.76: 2316–2326,1996.
VANDEN-ABEELE, J. Comments on the functional asymmetries of the lowerextremities.Cortex16: 325–329, 1980.
WINTER, D. A. The Biomechanics and Motor Control of Human Gait: Normal,Elderly and Pathological. Waterloo, Ontario: Waterloo Biomechanics Press,1991.
ZATSIORSKY, V., SELUYANOV, V., AND CHUGUNOVA, L. In vivo body segmentinertial parameters determination using a gamma-scanner method. In:Bio-mechanics of Human Movement: Applications in Rehabilitation, Sports andErgonomics,edited by N. Berme and A. Cappozzo. Worthington, OH:Bertec, 1990, p. 186–202.
ZERNICKE, R. F.AND SMITH, J. L. Biomechanical insights into neural control ofmovement. In:Handbook of Physiology. Exercise: Regulation and Integra-tion of Multiple Systems. Washington, DC: Am. Physiol. Soc., 1996, sect.12, p. 293–330.
300 R. GRASSO, M. ZAGO, AND F. LACQUANITI




















