
Morphological variation of fruits and infrutescences inBiscutella L. (Cruciferae)
11
Feddes Repertorium 103 (1992) 5-6, 327-337 Berlin. August 1992 University of Reading, Department of Botany, School of Plant Sciences, Reading, Great Britain J. D. OLOWOKUDEJO Morphological variation of fruits and infrutescences in Biscirtellcl L. (Cruciferae) With one Figure, one Table and 3 Plates S u m ma ry The variation of fruits and infrutcscences in 51 taxa of the genus Biscutellu is reported. The basic structure of the fruit is similar in all species - a stalked strongly compressed indehiscent didymous silicula which breaks from the axis into two one-seeded loculi at mnturity. The peculiar and distinctive nature of the fruit constitutes the most important diagnostic feature for separating the genus from other genera. There is considerable variation in fruit size both within and between some species, e.g. the smallest fruits are found in B. microcurpu where they hardly exceed 2.5 mm x 5.0 mni while in B. rnegacarpucu they could be as big ;IS 11.0 mrn long and 18.5 mm wide. Prominent among species with significant intra-specific variation are B. lirsiranicu, B. setpercirens, B. glaciulis and B. crrlentina. The apex of the fruit is notched to varying extents except in B. uiiricirluttr where the margins run continuously with the style and the apex is therefore not notched. Style is persistent and contributes to the distinctive appearance of the fruit and the recognition of some species. Other variable characters include the pedicel. margin. valve shape and pubescence. The infrutescence varies from a simple raceme to a much-branched panicle. The flexuous branches of some taxa constitute an impor- tant feature for their preliminary identification. Some of these characters are of value in clarifying the taxonomic relationships among some component taxa. Introduction The genus Biscutelfa L. is widely distributed mainly in the Mediterranean region except for Z us am men fa ss un g Von 51 Taxa der Gattung Bisciitellu wird iiber die Variation der Fruchte und BlutenstSnde berichtet. Die Basisstruktur der Frucht ist bei alien Arten ihnlich: ein gestieltes, engzusammengeprentes unauf- geplatztes zweiteiliges Schotchen, das bei der Reife an der Achse in zwei einsamige Loculi aufbricht. Die eigenturnliche. abweichende Ausbildung der Frucht bildet das wiehtigste diagnostische Merkmal. das dic Gattung von den anderen Gattungen der Cruciferae trennt. In der FruchtgroBe selbst herrscht eine beacht- liche Spiinnweite sowohl innerhalb der Arten selbst als auch zwischen den rinzelnen Arten. So wurde die kleinste Frucht mit nur 2.5 x 5.0 mm bei B. micro- carpi gefunden, die griinte Frucht mit 11.0 x 18.5 mm lieferte B. macrocarpu. Die bedeutendsten unter den Arten mit kennzeichnender intraspezifischer Varia- tion sind B. hisiranica, B. senipercirens, B. yluciulis und B. calmrinn. Der Apex der Fruchte zeigt Ein- kerbungen unterschiedlichen AusmaDes auDer bei B. uuricirluta, bei der die Riinder gleichmiiRig iiber den Griffel verlaufen, weshalb der Apex nicht einge- tieft ist. Der Griffel steht aufrecht und bestimmt das unterschiedliche Aussehen der Fruchte sowie die Bestimmung einiger Arten. Andere variable Eigen- schaften betreffen den Pedicellus, die Nahte, die valvate Form und die Reife. Die Blutenstinde wech- seln zwischen einfacher Traubenform bis zu vielver- zweigten Blutenrispen. Die gebogenen Zweige einiger Taxa sind fur eine erste Bestimmung der Art von Bedeutung. Mehrere dieser angefuhrten Eigenschaf- ten sind fur die Klarung der taxonomischen Ver- wandtschaft wertvoll. one species complex, B. lnevigatn L., which extends into central Europe. The genus is tax- onomically difficult and there is little agreement between the various past taxonomic treatments
Transcript of Morphological variation of fruits and infrutescences inBiscutella L. (Cruciferae)

Feddes Repertorium 103 (1992) 5-6, 327-337 Berlin. August 1992
University of Reading, Department of Botany, School of Plant Sciences, Reading, Great Britain
J . D. OLOWOKUDEJO
Morphological variation of fruits and infrutescences in Biscirtellcl L. (Cruciferae)
With one Figure, one Table and 3 Plates
S u m m a r y
The variation of fruits and infrutcscences in 51 taxa of the genus Biscutellu is reported. The basic structure of the fruit is similar in all species - a stalked strongly compressed indehiscent didymous silicula which breaks from the axis into two one-seeded loculi at mnturity. The peculiar and distinctive nature of the fruit constitutes the most important diagnostic feature for separating the genus from other genera. There is considerable variation in fruit size both within and between some species, e.g. the smallest fruits are found i n B. microcurpu where they hardly exceed 2.5 mm x 5.0 mni while in B. rnegacarpucu they could be as big ;IS 11.0 mrn long and 18.5 mm wide. Prominent among species with significant intra-specific variation are B. lirsiranicu, B. setpercirens, B. glaciulis and B. crrlentina. The apex of the fruit is notched to varying extents except in B. uiiricirluttr where the margins run continuously with the style and the apex i s therefore not notched. Style is persistent and contributes to the distinctive appearance of the fruit and the recognition of some species. Other variable characters include the pedicel. margin. valve shape and pubescence. The infrutescence varies from a simple raceme to a much-branched panicle. The flexuous branches of some taxa constitute an impor- tant feature for their preliminary identification. Some of these characters are of value in clarifying the taxonomic relationships among some component taxa.
Introduction
The genus Biscutelfa L. is widely distributed mainly in the Mediterranean region except for
Z us a m m e n fa ss un g
Von 51 Taxa der Gattung Bisciitellu wird iiber die Variation der Fruchte und BlutenstSnde berichtet. Die Basisstruktur der Frucht ist bei alien Arten ihnlich: ein gestieltes, engzusammengeprentes unauf- geplatztes zweiteiliges Schotchen, das bei der Reife an der Achse in zwei einsamige Loculi aufbricht. Die eigenturnliche. abweichende Ausbildung der Frucht bildet das wiehtigste diagnostische Merkmal. das dic Gattung von den anderen Gattungen der Cruciferae trennt. In der FruchtgroBe selbst herrscht eine beacht- liche Spiinnweite sowohl innerhalb der Arten selbst als auch zwischen den rinzelnen Arten. So wurde die kleinste Frucht mit nur 2.5 x 5.0 mm bei B. micro- carpi gefunden, die griinte Frucht mit 11.0 x 18.5 m m lieferte B. macrocarpu. Die bedeutendsten unter den Arten mit kennzeichnender intraspezifischer Varia- tion sind B. hisiranica, B. senipercirens, B. yluciulis und B. calmrinn. Der Apex der Fruchte zeigt Ein- kerbungen unterschiedlichen AusmaDes auDer bei B. uuricirluta, bei der die Riinder gleichmiiRig iiber den Griffel verlaufen, weshalb der Apex nicht einge- tieft ist. Der Griffel steht aufrecht und bestimmt das unterschiedliche Aussehen der Fruchte sowie die Bestimmung einiger Arten. Andere variable Eigen- schaften betreffen den Pedicellus, die Nahte, die valvate Form und die Reife. Die Blutenstinde wech- seln zwischen einfacher Traubenform bis zu vielver- zweigten Blutenrispen. Die gebogenen Zweige einiger Taxa sind fur eine erste Bestimmung der Art von Bedeutung. Mehrere dieser angefuhrten Eigenschaf- ten sind fur die Klarung der taxonomischen Ver- wandtschaft wertvoll.
one species complex, B. lnevigatn L., which extends into central Europe. The genus is tax- onomically difficult and there is little agreement between the various past taxonomic treatments

328 Feddes Repert., Berlin 103 (1992) 5-6
as to the number of species recognised or the ranks of the recognised taxa (e.g. DE CANDOLLE
COSSON 1872; WILLKOMM & LANGE 1880; Rouv & FOUCAUD 1895; MALINOWSKI 1910; MA- CHATSCHKI-LAURICH 1926; GUINEA 1963; GUI- NEA & HEYWOOD 1964a, b; MAIRE 1967 among others). The major problem encountered in the genus is that of specific delimitation. Although the species usually have characteristic and re- cognizable appearance yet they are very difficult to describe precisely. The limits of variation of the species are not easy to clarify and define because of intermediate variants that frequently occur. There is also the presence of narrow endemics which may be morphologically close to other more widespread and better known species. Other problems of the genus have been highlighted in previous papers by OLOWOKUDE- JO (1985, 1986a, b).
In this investigation, the fruits and infrutes- cences of all species of Biscutella have been examined as part of a much wider programme of research into the systematics of the genus. Cruciferous fruits are extremely diverse and are relied upon to a large extent in the classification of the family at tribal, generic and specific levels.
Materials and methods
Plant material from three main sources were used for this investigation, and these include (a) herbarium collections (b) natural populations and (c) cultivated samples.
Herbarium specimens were either borrowed from or examined in the following institutions: B, BC, BCC, BCF, BM, G, GDA, GJO, GZU, HBG, K, LINN, LY, M A , MAF, RNG and W. Abbreviations follow HOLMCREN et al. (1981).
Field studies were conducted in central, eastern and southern Spain where 78 populations comprising 14 taxa were investigated and sampled in their natural habitats.
Fruits and seeds obtained from the field trips and those of known wild origin received from other collectors and botanic gardens were cultivated in the greenhouse and experimental plots of the School of Plant Sciences, University of Reading. The plants were raised to maturity and studied.
I8 1 1, I82 1 ; REICHENBACH 1 829 ; JORDAN 1864;
Assessmen t o f f r u i t c h a r a c t e r s
Fruits are selected from the lower half of the main infrutescence as these are the most typical. Fruits towards the top of the infrutescence are often smaller
and not fully developed. As much as possible. all measurements are made on mature fruits a t ap- proximately similar stage of development and in com- parable positions on different plants. At least five fruits per infrutescence were assessed while the total for each taxon ranges from 14 to 60.
The length ofeach fruit is measured from the base of the valve to the top while the width is measured a t the widest point. Pedicel length is measured from the junction with the stem to the base of the stipe. Width measurements are not taken for the pedicel but are referred to using relative terms such as stout or slender. The persistent style is measured from the tip of the stigma to the point of contact with the valves.
Descriptive statistics of means. standard devia- tions and errors are calculated for all variables.
M o r p h o l o g i c a l e x a m i n a t i o n All fruits are examined by means of a simple hand lens and dissecting microscope. The nature of the fruit apcx and margin are also determined.
The structure and branching patterns of the infrutescence are determined on fully grown and mature plants of each taxon.
Results The basic structure of the fruit in all species of the genus is similar: it is a stalked, strongly compressed indehiscent didymous silicula (or schizocarpoid) which breaks from the axis into two one-seeded loculi at maturity. The generic name, Biscutella, which means double shield, alludes to the peculiar and distinctive fruit resembling a pair of spectacles. The fruit is usually light to deep green when immature but turning dark green or light to dark brown or brownish black when fully developed. Some fruits of B. uariegata BOISS. et REUTER, B. mega- carpaeu BOSS et REUTER, B. foliosa MACH.- LAUR. and B. radicata COSSON are light to deep purple. The infrutescence may consist of a mixture of mature and immature fruits and flowers. Variation in some morphological cha- racters of the fruits is summarised in the Table. Representative samples of fruit types are illu- strated in Plates I - 111 while the infrutescence patterns are shown diagrammatically in the Figure.
F r u i t s ize
There is considerable intra- and inter-specific variation in fruit size within the genus (Table).
A - $ C i D 4
E
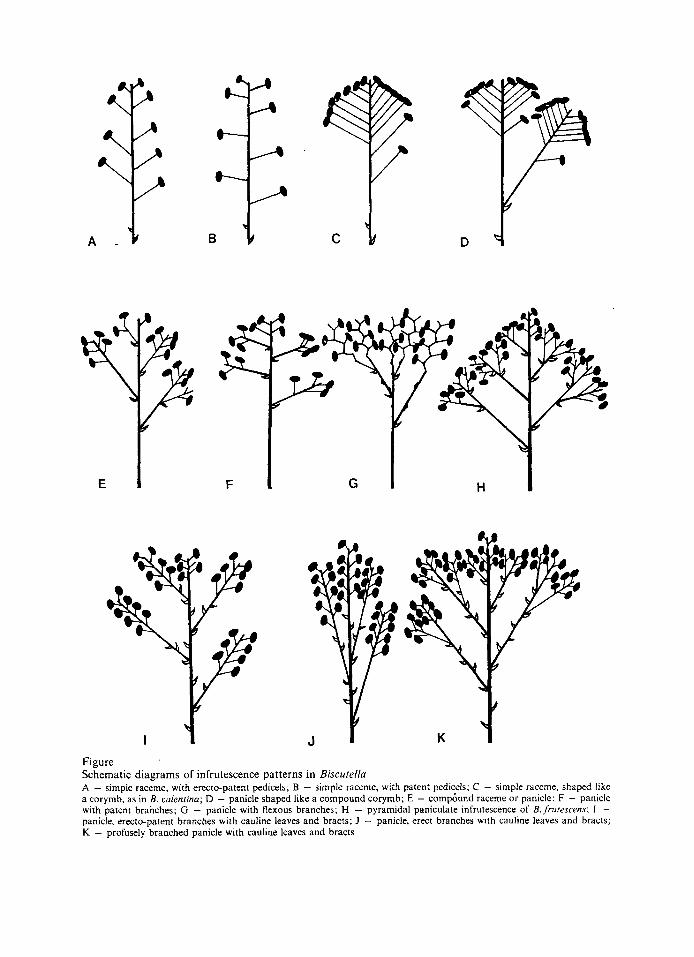
Figure Schematic diagrams of infrutescence patterns in Biscutella A - simple raceme, with erecto-patent pedicels; B - simple raceme, with patent pedicels; C - simple raceme, shaped like a corymb, as in B. ualentinu; D - panicle shaped like a compound corymb; E - compbund raceme or panicle: F - panicle wlth patent branches; G - panicle with flexous branches; H - pyramidal paniculate infrutescence of B. frutescms; I - panicle, erecto-patent branches with cauline leaves and bracts; J - panicle. erect branches with c a u l k leaves and bracts; K - profusely branched panicle with cauline leaves and bracts
330
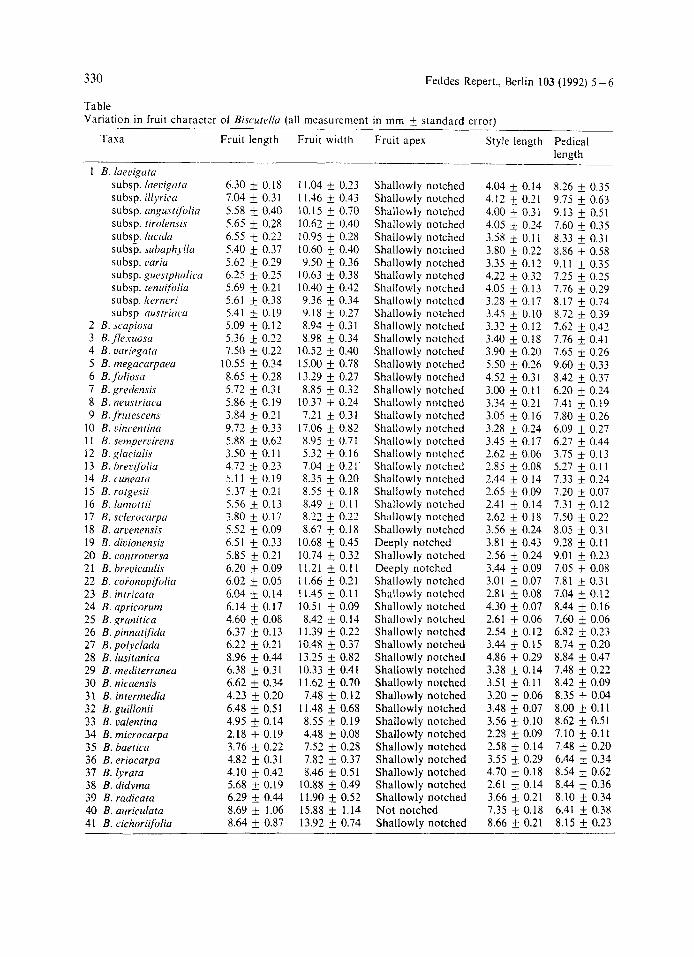
Table Variation in fruit character of Biscutrllu (all measurement in mm f standard error)
Feddes Repert., Berlin 103 (1992) 5 - 6
Taxa Fruit length Fruit width Fruit apex
1 B. ltrei ' I k ota subsp. luecigatu subsp. illjriccc subsp. ungust foliu subsp. tirolensis subsp. Iitciib subs;. suhaph~l la subsp. oariu subsp. gitrstplirrlic~ir subsp. tenrrifdia subsp. kerneri subsp. austriircu
2 B. scapiosu 3 B.flesuosa 4 B. vuriegatu 5 B. megacurpaeu 6 B. foliosu 7 B. greclensis 8 B. neusiriuc'ci 9 B.frirtescens
10 B. cincentina 1 I B. srmpercirens 12 B. gluciulis 13 B. hrecijoliu 14 B. c'uneuta 15 B. rutgeyii 16 B. lumottii 17 B. sclerociirpir 18 B. uraenensis 19 B. diuionensis 20 B. confrocersa 21 B. brevicairlis 22 B. coionopijoliu 23 B. intricata 24 B. apricorum 25 B. graniticu 26 B. pinnntijida 27 B. polyclada 28 B. lusitunicu 29 B. mediterranea 30 B. nicaensis 3 I B. intermedia 32 B. guillonii 33 B. valentina 34 B. microcarpa 35 B. baetica 36 B. eriocarpa 37 B. lyrata 38 B. didymu 39 B. radicata 40 B. auriculata 41 B. cichoriifolia
6.30 f 0.18 7.04 f 0.31 5.58 0.40 5.65 f 0.28 6.55 +. 0.22 5.40 k 0.37 5.62 f 0.29 6.25 & 0.25 5.69 f 0.21 5.61 f 0.38 5.41 f 0.19 5.09 f 0.12 5.36 & 0.22 7.50 f 0.22
10.55 k 0.34 8.65 f 0.28 5.72 k 0.31 5.86 k 0.19 3.84 0.21 9.73 f 0.33 5.88 f 0.62 3.50 f 0. I 1 4.72 & 0.23 5.1 I & 0.19 5.37 & 0.21 5.56 k 0.13 3.80 f 0.17 5.52 f 0.09 6.51 f 0.33 5.85 f 0.21 6.20 f 0.09 6.02 f 0.05 6.04 -t 0.14 6.14 & 0.17 4.60 f 0.08 6.37 0.13 6.22 f 0.21 8.96 f 0.44 6.38 f 0.31 6.62 f 0.34 4.23 f 0.20 6.48 f 0.51 4.95 0.14 2.18 + 0.19 3.76 f 0.22 4.82 f 0.31 4.10 f 0.42 5.68 f 0.19 6.29 f 0.44 8.69 f 1.06 8.64 f 0.87
1 1.04 f 0.23 1 1.46 & 0.43 10.15 f 0.70 10.62 f 0.40 10.95 f 0.28 10.60 f 0.40 9.50 f 0.36
10.63 f 0.38 10.40 f 0.42 9.36 f 0.34 9.18 f 0.27 8.94 f 0.31 8.98 f 0.34
10.52 f 0.40 15.00 f 0.78 13.29 f 0.27 8.85 f 0.32
10.37 f 0.24 7.21 f 0.31
17.06 f 0.82 8.95 f 0.71 5.32 f 0.16 7.04 f 0.21 8.35 f 0.20 8.55 f 0.18 8.49 0.1 1 8.21 f 0.22 8.67 f 0.18
10.68 k 0.45 10.74 f 0.32 11.21 f 0.11 11.66 f 0.21 11.45 f 0.1 I 10.51 f 0.09 8.42 f 0.14
11.39 k 0.22 10.48 f 0.37 13.25 f 0.82 10.33 Ifr 0.41 11.62 f 0.70 7.48 f 0.12
11.48 f 0.68 8.55 f 0.19 4.48 k 0.08 7.52 f 0.28 7.82 f 0.37 8.46 f 0.51
10.88 f 0.49 11.90 f 0.52 15.88 f 1.14 13.92 f 0.74
Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Deeply notched Shallowly notched Deeply notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Shallowly notched Not notched Shallowly notched
Style length
4.04 f 0.14 4.12 Ifr 0.21 4.00 f 0.31 4.05 i 0.24 3.58 f 0.1 I 3.80 f 0.22 3.35 f 0.12 4.22 k 0.32 4.05 & 0.13 3.28 k 0.17 3.45 & 0.10 3.32 f 0.12 3.40 f 0.18 3.90 f 0.20 5.50 f 0.26 4.52 +_ 0.31 3.00 f 0.1 1
3.05 f 0.16 3.28 & 0.24 3.45 f 0.17 2.62 i 0.06 2.85 f 0.08 2.44 f 0.14 2.65 f 0.09 2.41 f 0.14 2.62 f 0.18 3.56 f 0.24 3.81 f 0.43 2.56 f 0.24 3.44 f 0.09 3.01 2 0.07 2.81 f 0.08 4.30 f 0.07 2.61 f 0.06 2.54 f 0.12 3.44 f 0.15 4.86 f 0.29 3.38 k 0.14 3.51 f 0.11 3.20 0.06 3.48 f 0.07 3.56 f 0.10 2.28 f 0.09 2.58 f 0.14 3.55 f 0.29 4.70 f 0.18 2.61 f 0.14 3.66 0.21
8.66 0.21
3.34 2 0.21
7.35 * 0.18
Pedical length
8.26 f 0.35 9.75 f 0.63 9.13 f 0.51 7.60 f 0.35 8.33 f 0.31 8.86 f 0.58 9.11 f 0.35 7.25 f 0.25 7.76 0.39 8.17 f 0.74 8.72 f 0.39 7.62 f 0.42 7.76 f 0.41 7.65 f 0.26 9.60 f 0.33 8.42 f 0.37 6.20 f 0.24 7.41 & 0.19 7.80 f 0.26 6.09 f 0.27 6.27 f 0.44 3.75 f 0.13 5.27 f 0.1 1 7.33 f 0.24 7.20 f 0.07 7.31 f 0.12 7.50 f 0.22 8.05 & 0.31 9.18 f 0.1 1 9.01 f 0.23 7.05 f 0.08 7.81 f 0.31 7.04 f 0.12 8.44 f 0.16 7.60 f 0.06 6.82 f 0.23 8.74 f 0.20 8.84 f 0.47 7.48 f 0.22 8.42 f 0.09 8.35 f 0.04 8.00 f 0.11 8.62 f 0.51 7.10 f 0.11 7.48 f 0.20 6.44 f 0.34 8.54 f 0.62 8.44 f 0.36 8.10 f 0.34 6.41 f 0.38 8.15 Ifr 0.23

J. D. OLowoKuDEjo, Fruits and infrutescences in Biscufella 33 1
Prominent among species with significant intra-specific variation are B. lusitanica JORDAN in which a large-fruited variant measuring a- bout 9 - l l x 14- 17 mm has been described as a new variety (OLOWOKUDEJO 1986a) or even a species by GUINEA (1 963); B. semperuirens L. in which one taxon, var. tornentosa (LAG.) HEYWOOD has silicula measuring 7 - 8 x 12- 13 mm, while the fruits of remaining three yarieties are about 4-6 mm long and 6.5- 10.5 mm wide. The two varieties reco- gnised within B. glacialis (BOISS. & REUTEK) JORDAN are also partially distinguishable by their fruit size (OLOWOKUDEJO 1986b).
F r u i t a p e x a n d va lve
The apex of the fruit may or may not be notched. B. auricirlura L. (Plate IlA) is the only species without a notched apex. In the remai- ning taxa the apex is notched to varying extents. B. divionensis JORDAN, B. brevicaulis JORDAN and some specimens of B. intermetlia GOUAN have deeply notched apices while the remaining taxa usually possess shallowly notched apices (Table; Plate I A - F).
The shape of each of tfie two valves com- prising the silicula varies from oval to spherical. Taxa with deeply notched apices always have oval-shaped valves while those with shallow notching usually have spherical valves. Veins occur on the glabrous surface of the valves in some species but vary considerably in their degree of development and prominence. The venation pattern is conspicuously reticulate on the valves of B. laevigata L. subsp. lueviguta (Plate IC), B. laeuigata L. subsp. guestphalica MACH.-LAUR. (Plate I E), B. megacarpaea (Pla- te IF), and B. microcarpa (Plate 11 D). Embryo abortion in one of the two valves is a common phenomenon in some species e.g. B. microcarpa (Plate IlD).
The margins of the valve may be very narrow ( <0.3 mm) or fairly wide (>0.5 mm). The fruit margin is diaphanous in B. auriculata (Plate 11 A) and B. cichoriifolia (Plate I1 B) while it is membranous in the remaining species. B. auriculata is the only species of the genus in which the margins run continuously with the style (Plate IIA). The peripheral region of the valves is swollen and forms a continuous ridge in B. frutescens COSSON (Plate 1A). This feature
coupled with the flattened inner region of tfie valve makes this species very distinctive. The closest to this is found in B. microcarpa but the inner region of the valve are usually convex and not as flat.
S ty l e
The style is persistent and contributes to the distinctive appearance of the fruit. At the distal end is the capitate or conical stigma which may be bilobed and wider than the style e.g. B. mega- carparu (Plate ID), B. irz[crrncdia (Plate IB), B. auriculata (Plate I1 A), B. cichoriifolia (Pla- te I1 B) and B. microcarpa (Plate I1 D). The stig- ma may also be much reduced and therefore smaller in diameter than the top of the style e.g. B. frutescens (Plate IA), B. laeviguta subsp. guestphalica (Plate I E ) and B. ual~nrina (Pla- te MA). The style length varies little within and between the species. B. auriculuta and B. cicho- riifolia have the longest styles which are about 7.35 mm and 8.66 mm respectively. Within the remaining taxa the style length varies from 2.28 mrn in B. microcarpu to 5.50 mm in B. me- gacarpuea (Table). The style is either cylindrical or quadrangular in cross section.
Pedice l
The fruit pedicel is variable in length and thickness. The shortest pedicels are found in B. gfacialis where they hardly exceed 4 - 5 mm in length while in B. cichoriifolia they may be as long as 8 -9 mm (Table). The pedicels of B. auricirlata (Plate IIA) and B. cichoriifolia (Plate IIB) are usually more robust and stout than those of the remaining species which are generally slender (Plate I). The disposition of pedicels on the infrutescence is also variable; patent, erecto-patent and erect pedicels may be found within the genus (Plates 11, Ill).
F r u i t i n d u m e n t u m
Pedicels and valves may be sparsely or densely hairy or totally glabrous. There is considerable variation in pubescence both within and be- tween taxa. The density of hairs on the pedicel is usually affected by age. A densely hairy pedicel at the early fruiting stage may become sparsely hairy or completely glaborous when the fruit reaches full maturity. In some populations of

332 Feddes Repert., Berlin 103 (1992) 5 - 6
B. lamotti JORDAN have long, slender and flexuous infrutescence branches (Fig. 1 G). Al- though this character is not diagnostic of these taxa, but it is usually confined to them. Mo- reover, B. frutescens is well characterised by its pyramidal paniculate infrutescence (Fig. 1 H). Cauline leaves or bracts usually occur at the base of infrutescence branches as shown in Fig. 1 E - 1 H, but in B. cichoriifolia (Fig. 1 I) and B. auriculata (Fig. 1 J , K) additional cau- line leaves or bracts are found on the pe- duncles.
B. valentina the young pedicels are usually whi- tish tomentose (Plate IIIA).
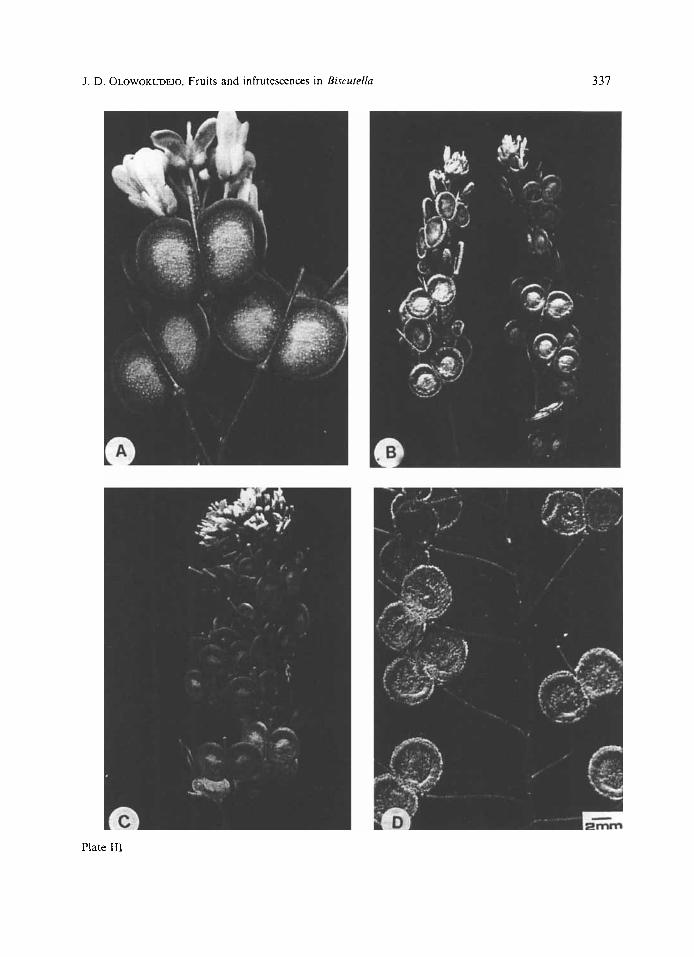
Simple, uniseriate trichomes which are va- riously shaped are found on the valves of some species. Some of these are shown in Plates 1-111. This feature is extremely variable both within and between taxa. For example, glabrous and hairy fruits are found within several taxa e.g. B. megacarpaea with hairy (Plate ID), and glabrous (Plate I F ) fruits, B. intermedia (Plate IB); B. auriculata (Plate IIC, E), B. didyma (Plate 111B) and B. lyrata (Plate IIIC, D). This hairs may be clavate, pilose, spinose or papillose. A mixture of any two or more of these hairs may be found on the same fruit surface, as has been reported elsewhere (OLO- WOKUDEJQ 1985). Pilose hairs are usually confi- ned to the edges of the valves in B. didyma (Plate 111B) while in B. lyrata the hairs are concen- trated on the central and peripheral regions of valves (Plate IIID). However, the intra- specific variability of the hairs makes them unreliable for purposes of identification and classification.
I n f r u t e s c e n c e
This is the fruit-bearing portion of the shoot, derived directly from the inflorescence. I t is usually terminal and indeterminate. The first fruit to develop is usually the lowest and maturation of the others takes place acrope- tally. The various patterns of arrangement of the fruits are shown diagrammatically in Fig. 1. The infrutescence may be a simple raceme with either erecto-patent pedicels (Fig. 1 A) or patent pedicels (Fig. 1B). In some populations of B. valentina, the infrutescence may appear like a simple corymb (Fig. 1 C) or a compound one (Fig. 1 D) because of the varying length of the pedicels. The oldest and lowermost flowers usually abort resulting in a compact terminal infrutescence which may even resemble a capi- tulum.
In most taxa, the infrutescence is a panicle (Fig. 1 E - 1 K) which may differ in some other features from one species to the other. For example the infrutescences of B. sempervirens var. tomentosa usually have distinctly patent branches which are restricted to the top of the main axis (Fig. 1F). Several specimens of B. sclerocarpa REVEL, B. polyclada JORDAN and
Discussion
The preceeding observations show that the basic architecture of Biscutella fruits is remarka- bly similar in all species of the genus. The peculiar and distinctive fruits thus constitute the most important diagnostic feature for sepa- rating the genus from other genera of the family. This is in contrast to the general situation in the Cruciferae which has been described as one of the most difficult families in terms of the delimitation of genera (ROLLIN 1938,1959; DA- VIS & HEYWOOD 1963; AL-SHEHBAZ 1973). Di- thyrea is the only genus of the family with a fruit type approaching that of Biscutella but this genus is confined to the New World and also differs from Biscutella in a number of characters which have been elucidated by PAY-
Although the taxonomic utility of fruit and infrutescence characters within the genus is obscured by the uniformity of certain features in many taxa, yet some species possess several distinctive characteristics that are systemati- cally significant. The extremely small silicula of B. microcarpa and the exceptionally large fruits of B. megacarpaea constitute important dis- tinguishing features. The infrutescence and fruits of B. frutescens are remarkable in the genus for their unique pyramidal shape and swollen margins respectively. This correlates with the distinct anatomical structure of the petiole (OLOWOKUDEJO 1987). The flexuous infrutescence branches of B. sclerocarpa, B. po- lyclada and B.lamottii can be used for the preliminary identification of these taxa. Other features such as apex, margin and style show taxonomically valuable variation. The deeply notched apices of B. divionensis, and B. brevi-
SON (1918).

J . D. OLOWOKUDEJO. Fruits and infrutescences in Biscurella 333
caulis separate these species from the rest. B. auriculata is easily recognisable as the only species without a notched apex and with mar- gins that are excurrent with the style.
Moreover, the fruit characters are of great assistance in clarifying the relationships of the component species. For instance, a diagnostic combination of stout pedicels, diaphanous mar- gins, relatively long styles and leafy infrutes- cences,- reveals the close morphological simi- larity between B. auriculata and B. cirhoriifolia. These features, among others, also distinguish these species from other taxa within the genus and support their elevation into a subgeneric rank as was proposed by OLOWOKUDEJO (1986a). The observed pattern of fruit varia- tion in B. coronopifolia, B. granitica, B. intri- cafa, B. polyclada and B. pinnat f ida does not support the continued separation of these taxa (OLOWOKUDEJO 1986b). The random varia- tion of fruit features in the various subspecies of B. laevigata complex shows that the diploid and tetraploid taxa cannot be separated by fruit size as earlier noted by OLOWOKUDEJO & HEY- WOOD (1984). Some species, e.g. B. vafentina and B. sempervirens, which have been shown to be widely distributed and highly variable mor- phologically (OLOWOKUDEJO 1 9 8 6 ~ d), usu- ally exhibit wide variations in fruit charac- teristics.
The presence or absence, and type of indu- mentum on the fruit valve have formed the basis for the recognition of many infraspecific taxa in B. auriculata, B. didyma and B. lyrata (MARE 1967; VAN OOSTSTROOM 1970; RAFFAELLI 1985, 1990). Previous investigation of the fruit surface (OLOWOKUDEJO 1985) has revealed the inconsi- stency of the micromorphological features and the variable nature of the indumentum which make them unreliable as taxonomic markers. ROLLINS (1958) has also demonstrated that the presence or absence of hairs in the fruit of Dithyrea, a related genus, is a simply inherited characteristic under a single gene control and therefore of no taxonomic significance.
A c k n o w l e d g e m e n t s
The author wishes to thank the Head of Botany Department, University of Reading, for permission to use the laboratory and greenhouse facilities and the Directors and Curators of herbaria consulted for kindly placing their material at his disposal.
’
References
AL-SHEHBAZ, I. A,, 1973, Biosystemaiics of the genus Thelypodium (Cruciferae). Contr. Gray Herb.,
COSSON, E. S. C., 1872, Bull. SOC. Bot. France, 19.
DAVIS, P. H.; HEYWOOD, V. H., 1963, Principles of Angiosperm taxonomy. Edinburgh.
DE CANDOLLE, A. P., 1811, Ann. Mus. Nat. Hist. Paris, 18, 295-301.
DE CANDOLLE, A. P., 1821, Regni vegetabilis systema Naturale, 2, 407-417.
GUINEA, E., 1963, El genero Biscuiella. Anal. Inst. Bot. Cavanille. 21, 387-405.
GUINEA, E.; HEYWOOD, V. H., 1964, Biscztrella. In: Flora Europaea, Vol. I./ed. by T. G. TUTIN et al. Cambridge. 325 - 330.
HOLMGREN, P. K.; KEUKEN, W.; SCHOFIELD, E. K., 1981, Index herbariorum. Part I . The herbaria of the World. ed. 7. Regnum veg., 106.
JORDAN, A., 1864, Diagnoses d’especes nouvelles ou meconnues, Vol. 1. Paris. Leipzig. 292 - 329.
MACHATSCHKI-LAURICH, B., 1926, Die Arten der Gattung Biscutella L. sect. Thlaspidiurn (MED.) DC. Bot. Arch., 13, 1 - 115.
MAIRE, R., 1967, Flora de I’Afrique du Nord, Vol. 13,
MALINOWSKI, E., 1910, Monographie du genre Biscu- rella L. I. Classification et distribution gPogra- phique. Bull. Internat. Acad. Sci. Cracovie. CI.
OLOWOKUDEJO, J . D., 1985, Scanning electron mic- roscopy of fruits in the genus Biscure/la (Crucife- rae). Phytornorphology, 35, 273 - 286.
OLOWOKUDEJO, J . D., I986a, The infrageneric classi- fication of Biscutella (Cruciferae). Brittonia, 38,
OLOWOKUDEJO, J. D., 1986b, New nomenclatural combinations and three new varieties in the genus Biscutel/a (Cruciferae). Feddes Repert., 97,
OLOWOKUDEJO, J. D., 1986c, Variation and tax- onomy of Biscutella valenrina in Spain. Can. J. Bot., 64, 2965 -2972.
OLOWOKUDUO, J. D., 1986d, Population variation in Biscutella sempervirens (Cruciferae) in Spain. Willdenowia, 16, 95- 102.
OLOWOKUDEJO, J. D., 1987, Taxonomic value of pe- tiole anatomy in the genus Biscutella L. (Cru- ciferae). Bull. Jard. Bot. Natl. Belg., 57, 307 to 320.
OLOWOKUDEJO, J. D.; HEYWOOD, V. H., 1984, Cyto- taxonomy and breeding system of the genus Biscutella (Cruciferae). Plant. Syst. & Evol., 145,
PAYSON, E. B., 1918, Notes on certain Cruciferae.
204, 3- 148.
222 - 223.
126-147.
Math.. 1910, I I 1 - 139.
86 - 88.
565 - 570.
291 - 309.
Ann. Mo. Bot. Card., 5, 143-151.

Feddes Repert., Berlin 103 (1992) 5-6
ROY. G.; FOUCAUD, J., 1895. Flore de France. Bd. 2. Paris et Rochefort. 104- 115.
VAN OOSTSTROOM, S. J., 1970. Adventive Biscirtella’s. Gorteria, 5 , 44-46.
WILLKOMM, H . M.; LANCE, J . M. C.. 1880. Prodro- mus florat: hispanicae. Bd. 3. Stuttgart. 760 to 765.
RAFFAELLI, M.. 1985, Taxonomic rearrangement of Biscutella lyraia (Cruciferae). Taxon, 34,695 - 697.
RAFFAELLI, M.. 1991. Biscurellu. 1. ser. LJi-arae MAL- IN. (Cruciferae) in Italia. Discussione sui carattcri morfologici e tassonomia. Webbia, 45. 1-30,
REICHENBACH. H. G. L.. 1829, lconographia botani- ca seu plantae criticae. Vol. VII. Lipsiae.
ROLLINS. R. C.. 1938, Smelocskia and Polyctenium.
ROLLINS, R. C., 1958, The genetic evolution of a taxonomic character in Ditliyrea (Cruciferae). Rhodora, 60, 145- 152.
ROLLINS, R. C., 1959, The genus Syntlzlipsis (Cruci- ferae). Rhodora, 61, 253 -264.
Rhodora. 40. 294- 305. Address of the author: Dr. J . DELE OLOWOKUDEJO, University of Reading, Department of Botany, School of Plant Sciences, Reading RG6 IAS, Great Britain
Manuscript received: November, 4th, 1991.
Explanafions to Plates I - 111
P l a t e I Variation of fruits in Bisciriella: (A) B.fruiescen7, silicula with distinctly swollcn margin. (B) B. inierniediu, glabrous and pubescent fruits with shallowly and deeply notched apices. C B. laevigura subsp. luecigutu, glabrous valves with a network of veins. (D) B. megucarpueu, big. densely pubescent fruits. (E) B. Iireviguru subsp. guestphalica. glabrous fruit with prominent reticulate venation. (F) B. megucarpaea, glabrous fruit with reticulate venation and well-developed membranous margin
P la t e I 1 Variation of fruits and infrutescences in Biscutella: (A) B. auriculara, densely pubescent fruit with wide diaphanous margin and stout pedicel. (B) B. cichoriifolia, pubescent fruit with wide diaphanous margin and stout pedicel. (C) B. atrriculuta, portion of infrutescence with patent and erecto-patent, pubescent fruits. (D) B. microcarpa, portion of infrutescence with patent fruits and aborted valves. (E) B. auriculata, portion of infrutescence with patent and erecto-patent glabrous fruits
P l a t e 111 Variation of infrutescences in Biscutellu: (A) B. valmiinu, fruits, with pubescent valves and pedicel and terminal flowers. (B) B. didyma, two infrutescence branches with pubescent and glabrous fruits. (C) B. lyratu, glabrous fruit and terminal flowers. (D) B. lyrara, densely pubescent fruits with patent and erectopatent dispositions

J. D. OLOWOKUDEJO, Fruits and infrutescences in Biscutella 335
Plate I

336 Feddes Repert., Berlin 103 (1 992) 5 - 6
Plate I1
J. D. OLOWOKUDEJO, Fruits and infrutescences in Biscziiella 337
Plate 111




















