
Monsoon rain forest seedling dynamics, northern Australia: contrasts with regeneration in...
18
ORIGINAL ARTICLE Monsoon rain forest seedling dynamics, northern Australia: contrasts with regeneration in eucalypt-dominated savannas Jeremy Russell-Smith 1,2 * and Samantha A. Setterfield 1,3 1 Tropical Savannas Cooperative Research Centre, Charles Darwin University, Darwin, NT, 2 Bushfires Council of the Northern Territory, Winnellie, NT and 3 Charles Darwin University, Darwin, NT, Australia *Correspondence: Jeremy Russell-Smith, Tropical Savannas Cooperative Research Centre, Charles Darwin University, Darwin 0909, Northern Territory, Australia. E-mail: [email protected] ABSTRACT Aim To explore: (1) the relative influences of site conditions, especially moisture relations, on pathways and rates of monsoon rain forest seedling and sapling regeneration, especially of canopy dominants, in northern Australia; and (2) contrasts between regeneration syndromes of dominant woody taxa in savannas and monsoon rain forest. Location Four monsoon rain forest sites, representative of regional major habitat and vegetation types, in Kakadu National Park, northern Australia. Methods A decadal study involved: (1) initial assessment over 2.5 years to explore within-year variability in seed rain, dormant seed banks and seedling (< 50 cm height) dynamics; and (2) thereafter, monitoring of seedling and sapling (50 cm height to 5 cm d.b.h.) dynamics undertaken annually in the late dry season. On the basis of observations from this and other studies, regeneration syndromes of dominant monsoon rain forest taxa are contrasted with comparable information for dominant woody savanna taxa, Eucalyptus and Corymbia especially. Results Key observations from the monsoon rain forest regeneration dynamics study component are that: (1) peak seed rain inputs of rain forest taxa were observed in the wet season at perennially moist sites, whereas inputs at seasonally dry sites extended into, or peaked in, the dry season; (2) dormant soil seed banks of woody rain forest taxa were dominated by pioneer taxa, especially figs; (3) longevity of dormant seed banks of woody monsoon rain forest taxa, including figs, was expended within 3 years; (4) seedling recruitment of monsoon rain forest woody taxa was derived mostly from wet season seed rain with limited inputs from soil seed banks; (5) at all sites rain forest seedling mortality occurred mostly in the dry season; (6) rain forest seedling and sapling densities were consistently greater at moist sites; (7) recruitment from clonal reproduction was negligible, even following unplanned low intensity fires. Main conclusions By comparison with dominant savanna eucalypts, dominant monsoon rain forest taxa recruit substantially greater stocks of seedlings, but exhibit slower aerial growth and development of resprouting capacity in early years, lack lignotubers in mesic species, and lack capacity for clonal reproduction. The reliance on sexual as opposed to vegetative reproduction places monsoon rain forest taxa at significant disadvantage, especially slower growing species on seasonally dry sites, given annual–biennial fires in many north Australian savannas. Keywords Australia, clonal reproduction, disturbance, Kakadu National Park, monsoon forest, savanna, seed bank, seed rain, tropical rain forest. Journal of Biogeography (J. Biogeogr.) (2006) 33, 1597–1614 ª 2006 The Authors www.blackwellpublishing.com/jbi 1597 Journal compilation ª 2006 Blackwell Publishing Ltd doi:10.1111/j.1365-2699.2006.01527.x
Transcript of Monsoon rain forest seedling dynamics, northern Australia: contrasts with regeneration in...

ORIGINALARTICLE
Monsoon rain forest seedling dynamics,northern Australia: contrasts withregeneration in eucalypt-dominatedsavannas
Jeremy Russell-Smith1,2* and Samantha A. Setterfield1,3
1Tropical Savannas Cooperative Research
Centre, Charles Darwin University, Darwin,
NT, 2Bushfires Council of the Northern
Territory, Winnellie, NT and 3Charles Darwin
University, Darwin, NT, Australia
*Correspondence: Jeremy Russell-Smith,
Tropical Savannas Cooperative Research Centre,
Charles Darwin University, Darwin 0909,
Northern Territory, Australia.
E-mail: [email protected]
ABSTRACT
Aim To explore: (1) the relative influences of site conditions, especially moisture
relations, on pathways and rates of monsoon rain forest seedling and sapling
regeneration, especially of canopy dominants, in northern Australia; and (2)
contrasts between regeneration syndromes of dominant woody taxa in savannas
and monsoon rain forest.
Location Four monsoon rain forest sites, representative of regional major habitat
and vegetation types, in Kakadu National Park, northern Australia.
Methods A decadal study involved: (1) initial assessment over 2.5 years to
explore within-year variability in seed rain, dormant seed banks and seedling
(< 50 cm height) dynamics; and (2) thereafter, monitoring of seedling and
sapling (50 cm height to 5 cm d.b.h.) dynamics undertaken annually in the late
dry season. On the basis of observations from this and other studies, regeneration
syndromes of dominant monsoon rain forest taxa are contrasted with comparable
information for dominant woody savanna taxa, Eucalyptus and Corymbia
especially.
Results Key observations from the monsoon rain forest regeneration dynamics
study component are that: (1) peak seed rain inputs of rain forest taxa were
observed in the wet season at perennially moist sites, whereas inputs at seasonally
dry sites extended into, or peaked in, the dry season; (2) dormant soil seed banks
of woody rain forest taxa were dominated by pioneer taxa, especially figs; (3)
longevity of dormant seed banks of woody monsoon rain forest taxa, including
figs, was expended within 3 years; (4) seedling recruitment of monsoon rain
forest woody taxa was derived mostly from wet season seed rain with limited
inputs from soil seed banks; (5) at all sites rain forest seedling mortality occurred
mostly in the dry season; (6) rain forest seedling and sapling densities were
consistently greater at moist sites; (7) recruitment from clonal reproduction was
negligible, even following unplanned low intensity fires.
Main conclusions By comparison with dominant savanna eucalypts, dominant
monsoon rain forest taxa recruit substantially greater stocks of seedlings, but
exhibit slower aerial growth and development of resprouting capacity in early
years, lack lignotubers in mesic species, and lack capacity for clonal reproduction.
The reliance on sexual as opposed to vegetative reproduction places monsoon
rain forest taxa at significant disadvantage, especially slower growing species on
seasonally dry sites, given annual–biennial fires in many north Australian
savannas.
Keywords
Australia, clonal reproduction, disturbance, Kakadu National Park, monsoon
forest, savanna, seed bank, seed rain, tropical rain forest.
Journal of Biogeography (J. Biogeogr.) (2006) 33, 1597–1614
ª 2006 The Authors www.blackwellpublishing.com/jbi 1597Journal compilation ª 2006 Blackwell Publishing Ltd doi:10.1111/j.1365-2699.2006.01527.x

INTRODUCTION
Monsoon rain forest occurs across northern Australia
typically as small (< 5 ha), floristically simple island-like
patches (mostly < 100 species per patch), in a vast expanse
of fire-prone eucalypt-dominated savanna (Russell-Smith,
1991). Patches occur on a variety of substrates and
perennially moist to seasonally arid site conditions, defined
by a number of floristic and structural types which describe
generally a subset of more complex and extensive rain forest
vegetation growing in lowland, climatically more equable
regions of north-eastern Australia (Webb & Tracey, 1981;
Russell-Smith, 1991; Bowman, 1999). Adult populations of
monsoon rain forest taxa in any one patch are mostly small
(< 50 individuals: Russell-Smith & Lee, 1992), with evidence
for populations of some species occurring as widely-
dispersed, genetically connected metapopulations (Shapcott,
1998a,b, 2000).
Fire frequencies over extensive areas of coastal to sub-coastal
northern Australia are often annual to biennial, are mostly
anthropogenic in origin, and occur mostly late in the seven-
month dry season under severe fire-weather conditions
(Williams et al., 2002; Russell-Smith et al., 2003a). Many
monsoon rain forest patches are afforded some level of fire
protection, occurring either at perennially moist sites (e.g.
springs) or, in seasonally dry situations, in rugged, rocky
terrain. Additionally, internal patch ground fuels are typically
compact and non-grassy, and may be relatively moist by
comparison with those in adjoining seasonal savannas
(Bowman & Wilson, 1988). Under fire regimes characterized
by frequent late dry season fires, critical issues at patch margins
include the capacity of component woody species to regenerate
(from seed, resprouting), and attendant rates of regeneration
(Russell-Smith & Stanton, 2002).
By contrast with tropical forest systems in perhumid
climates, it remains the case that very limited work has been
reported on the regeneration pathways and dynamics of
forest assemblages in the markedly seasonal tropics (Murphy
& Lugo, 1986; Gerhardt & Hytteborn, 1992). Available
observations suggest that: (1) fruiting phenological patterns
exhibit substantial variability between different study sites
(Bach, 2002); (2) seed banks are dominated by early
successional species, as per rain forests occurring under
more perhumid conditions (e.g. Garwood, 1989); (3)
seedling recruitment occurs mostly in association with
moist, rainy periods, and survivorship is affected markedly
by seasonal moisture deficit conditions (Lieberman & Li,
1992); (4) many woody species exhibit the capacity to
resprout following stem death (Murphy & Lugo, 1986;
Bowman, 1991a; Russell-Smith & Stanton, 2002); and (5)
clonal reproduction is likely to be more prevalent at
seasonally dry sites (Ewel, 1977; Lieberman & Li, 1992;
Russell-Smith, 1996).
Various of the above observations accord with similar
regeneration patterns observed for woody taxa in savanna
assemblages, for example: enhanced seedling recruitment
associated with consistently moist periods (Hoffmann, 1996;
Setterfield & Williams, 1996); substantial capacity of stems
to resprout following fire (Bond & van Wilgen, 1996;
Williams et al., 2002); and enhanced clonal reproduction
following burning (Lacey, 1974; Hoffmann, 1998). As with
monsoon rain forest patch margins, woody structure devel-
opment in fire-prone savannas is strongly dependent on
rates of stem growth and the frequency and intensity of fire
(Kellman, 1984; Braithwaite & Estbergs, 1985; Higgins et al.,
2000).
This paper reports a comparative decadal assessment of
seedling and sapling regeneration dynamics, focusing especially
on canopy dominants, at four sites representative of contrast-
ing site conditions (e.g. substrate moisture status, soil type), to
address: (1) the relative significance of seed rain, soil seed
banks, and clonal reproduction for monsoon rain forest woody
species recruitment; (2) the relative influences of different site
conditions, especially moisture relations, on rates of monsoon
rain forest seedling and sapling regeneration; and (3) in the
Discussion section, contrasts between the regeneration syn-
dromes of dominant regional monsoon rain forest and
savanna taxa.
Study sites
Four monsoon rain forest sites, each occurring in Kakadu
National Park, northern Australia, were selected for detailed
study based on prior numerical classification of a comprehen-
sive north Australian data set (Russell-Smith, 1991). The upper
levels of that classification hierarchy describe four major types
occurring in higher rainfall coastal and sub-coastal areas: (1)
rain forests associated with sites of perennial moisture, either
with neutral to basic, fine-textured soils (Groups 1, 2); or (2)
acidic, sandy soils (Groups 3–7); and (3) rain forests associated
with seasonally dry substrates, either in rugged sandstone
terrain (Group 8); or (4) occurring on a wide variety of other
substrate types (e.g. coastal dunes, granite or basalt outcrops,
lateritic landforms: Group 9). An analagous upper-level
classificatory hierarchy was reproduced from floristic
10 · 10 m quadrat data (n ¼ 303) for 33 northern Australian
monsoon rain forest patches, derived for the purposes of
characterizing habitat conditions of rain forest sapling banks
(Russell-Smith, 1996).
For the purposes of this study only one replicate of each of
the four floristic types was selected since: (1) previous
replicated studies (all one-off surveys, or of relatively short-
term duration) had already established major patterning in
forest typology (Russell-Smith, 1991), dormant seed banks
(Russell-Smith & Lucas, 1994), sapling banks (Russell-Smith,
1996), seed rain phenology (Bach, 2002); (2) the prime focus
of this study was to document the recruitment dynamics of
canopy dominants characteristic of the respective forest types
over a decadal period; (3) the four selected sites included
diagnostic dominant tree species (Russell-Smith, 1991) char-
acteristic of each of the respective four forest types (Table 1);
and (4) the logistic realities of sampling isolated replicate
J. Russell-Smith and S. A. Setterfield
1598 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
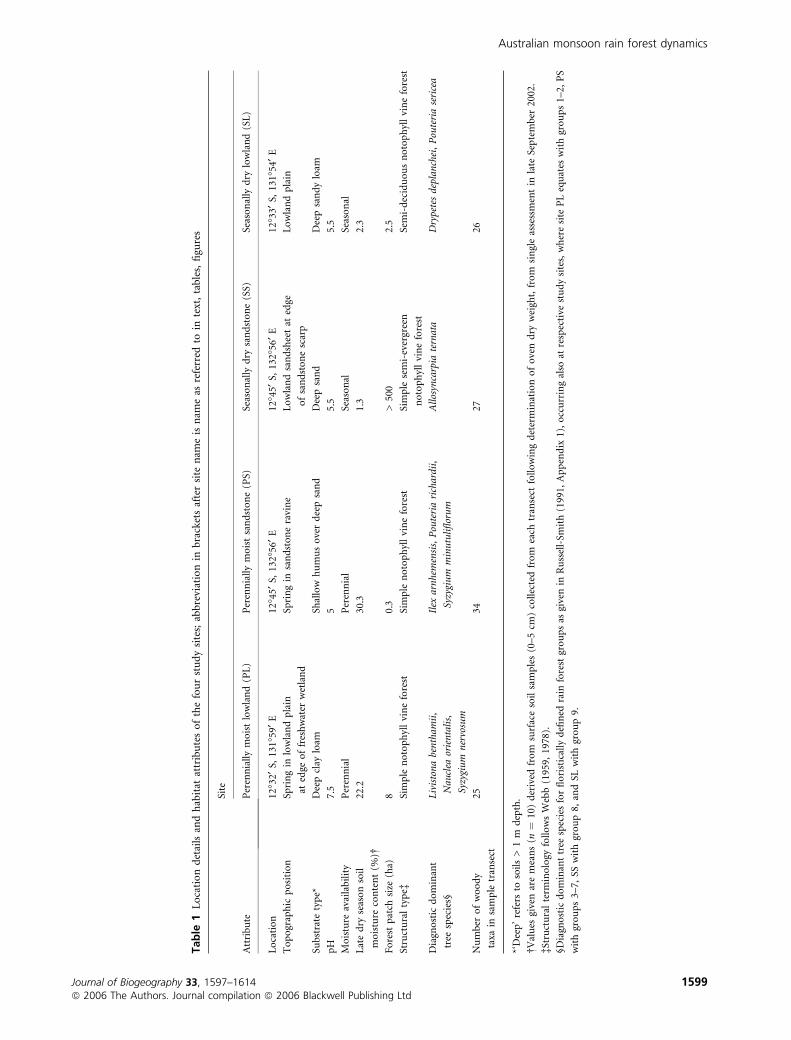
Tab
le1
Lo
cati
on
det
ails
and
hab
itat
attr
ibu
tes
of
the
fou
rst
ud
ysi
tes;
abb
revi
atio
nin
bra
cket
saf
ter
site
nam
eis
nam
eas
refe
rred
toin
text
,ta
ble
s,fi
gure
s
Att
rib
ute
Site
Per
enn
iall
ym
ois
tlo
wla
nd
(PL
)P
eren
nia
lly
mo
ist
san
dst
on
e(P
S)Se
aso
nal
lyd
rysa
nd
sto
ne
(SS)
Seas
on
ally
dry
low
lan
d(S
L)
Lo
cati
on
12�3
2¢S,
131�
59¢E
12�4
5¢S,
132�
56¢E
12�4
5¢S,
132�
56¢E
12�3
3¢S,
131�
54¢E
To
po
grap
hic
po
siti
on
Spri
ng
inlo
wla
nd
pla
in
ated
geo
ffr
esh
wat
erw
etla
nd
Spri
ng
insa
nd
sto
ne
ravi
ne
Lo
wla
nd
san
dsh
eet
ated
ge
of
san
dst
on
esc
arp
Lo
wla
nd
pla
in
Sub
stra
tety
pe*
Dee
pcl
aylo
amSh
allo
wh
um
us
ove
rd
eep
san
dD
eep
san
dD
eep
san
dy
loam
pH
7.5
55.
55.
5
Mo
istu
reav
aila
bil
ity
Per
enn
ial
Per
enn
ial
Seas
on
alSe
aso
nal
Lat
ed
ryse
aso
nso
il
mo
istu
reco
nte
nt
(%)�
22.2
30.3
1.3
2.3
Fo
rest
pat
chsi
ze(h
a)8
0.3
>50
02.
5
Stru
ctu
ral
typ
e�Si
mp
len
oto
ph
yll
vin
efo
rest
Sim
ple
no
top
hyl
lvi
ne
fore
stSi
mp
lese
mi-
ever
gree
n
no
top
hyl
lvi
ne
fore
st
Sem
i-d
ecid
uo
us
no
top
hyl
lvi
ne
fore
st
Dia
gno
stic
do
min
ant
tree
spec
ies§
Liv
isto
na
ben
tham
ii,
Nau
clea
orie
nta
lis,
Syzy
giu
mn
ervo
sum
Ilex
arn
hem
ensi
s,P
oute
ria
rich
ard
ii,
Syzy
giu
mm
inu
tuli
flor
um
All
osyn
carp
iate
rnat
aD
rype
tes
dep
lan
chei
,P
oute
ria
seri
cea
Nu
mb
ero
fw
oo
dy
taxa
insa
mp
letr
anse
ct
2534
2726
*‘D
eep
’re
fers
toso
ils
>1
md
epth
.
�Val
ues
give
nar
em
ean
s(n¼
10)
der
ived
fro
msu
rfac
eso
ilsa
mp
les
(0–
5cm
)co
llec
ted
fro
mea
chtr
anse
ctfo
llo
win
gd
eter
min
atio
no
fo
ven
dry
wei
ght,
fro
msi
ngl
eas
sess
men
tin
late
Sep
tem
ber
2002
.
�Str
uct
ura
lte
rmin
olo
gyfo
llo
ws
Web
b(1
959,
1978
).
§Dia
gno
stic
do
min
ant
tree
spec
ies
for
flo
rist
ical
lyd
efin
edra
info
rest
gro
up
sas
give
nin
Ru
ssel
l-Sm
ith
(199
1,A
pp
end
ix1)
,o
ccu
rrin
gal
soat
resp
ecti
vest
ud
ysi
tes,
wh
ere
site
PL
equ
ates
wit
hgr
ou
ps
1–2,
PS
wit
hgr
ou
ps
3–7,
SSw
ith
gro
up
8,an
dSL
wit
hgr
ou
p9.
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1599ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
patches mitigated against being able to undertake a thorough
longitudinal (temporal) investigation across many sites.
Site names (and abbreviations as used throughout the
paper), locations, habitat conditions, and diagnostic species are
outlined in Table 1. Two sites were located at small evergreen
rain forest patches associated with perennial springs: site PL on
pH-neutral deep clay loam at the edge of a lowland freshwater
wetland, and PS on acid humic soils over deep sand in rugged
sandstone terrain. The other sites were associated with well-
drained, seasonally dry, acidic substrates: SS comprising
extensive semi-evergreen forest on a deep sandsheet in rugged
sandstone, and SL comprising a small patch of semi-deciduous
forest on deep sandy loam in the gently undulating lowlands.
In other continental settings, monsoon rain forest equates with
structural types ranging from tropical semi-evergreen to
deciduous forests (Walter, 1984), monsoon forests (Whitmore,
1984), and tropical dry forests (Holdridge, 1967; Murphy &
Lugo, 1986).
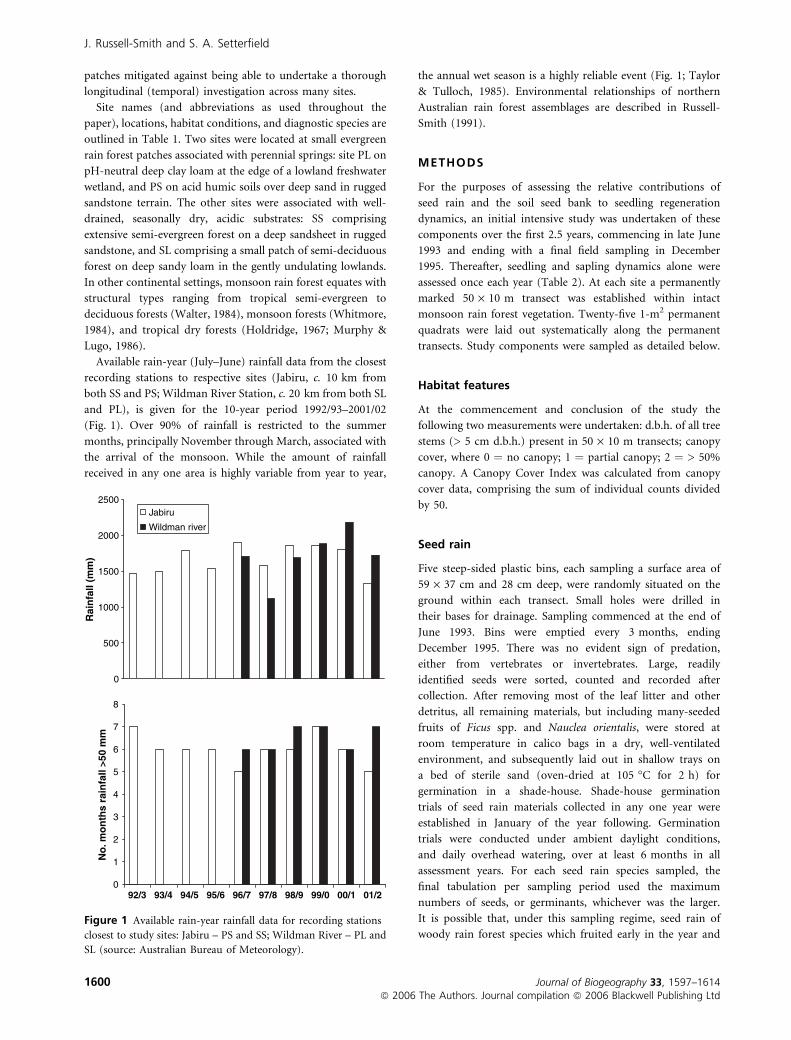
Available rain-year (July–June) rainfall data from the closest
recording stations to respective sites (Jabiru, c. 10 km from
both SS and PS; Wildman River Station, c. 20 km from both SL
and PL), is given for the 10-year period 1992/93–2001/02
(Fig. 1). Over 90% of rainfall is restricted to the summer
months, principally November through March, associated with
the arrival of the monsoon. While the amount of rainfall
received in any one area is highly variable from year to year,
the annual wet season is a highly reliable event (Fig. 1; Taylor
& Tulloch, 1985). Environmental relationships of northern
Australian rain forest assemblages are described in Russell-
Smith (1991).
METHODS
For the purposes of assessing the relative contributions of
seed rain and the soil seed bank to seedling regeneration
dynamics, an initial intensive study was undertaken of these
components over the first 2.5 years, commencing in late June
1993 and ending with a final field sampling in December
1995. Thereafter, seedling and sapling dynamics alone were
assessed once each year (Table 2). At each site a permanently
marked 50 · 10 m transect was established within intact
monsoon rain forest vegetation. Twenty-five 1-m2 permanent
quadrats were laid out systematically along the permanent
transects. Study components were sampled as detailed below.
Habitat features
At the commencement and conclusion of the study the
following two measurements were undertaken: d.b.h. of all tree
stems (> 5 cm d.b.h.) present in 50 · 10 m transects; canopy
cover, where 0 ¼ no canopy; 1 ¼ partial canopy; 2 ¼ > 50%
canopy. A Canopy Cover Index was calculated from canopy
cover data, comprising the sum of individual counts divided
by 50.
Seed rain
Five steep-sided plastic bins, each sampling a surface area of
59 · 37 cm and 28 cm deep, were randomly situated on the
ground within each transect. Small holes were drilled in
their bases for drainage. Sampling commenced at the end of
June 1993. Bins were emptied every 3 months, ending
December 1995. There was no evident sign of predation,
either from vertebrates or invertebrates. Large, readily
identified seeds were sorted, counted and recorded after
collection. After removing most of the leaf litter and other
detritus, all remaining materials, but including many-seeded
fruits of Ficus spp. and Nauclea orientalis, were stored at
room temperature in calico bags in a dry, well-ventilated
environment, and subsequently laid out in shallow trays on
a bed of sterile sand (oven-dried at 105 �C for 2 h) for
germination in a shade-house. Shade-house germination
trials of seed rain materials collected in any one year were
established in January of the year following. Germination
trials were conducted under ambient daylight conditions,
and daily overhead watering, over at least 6 months in all
assessment years. For each seed rain species sampled, the
final tabulation per sampling period used the maximum
numbers of seeds, or germinants, whichever was the larger.
It is possible that, under this sampling regime, seed rain of
woody rain forest species which fruited early in the year and
0
500
1000
1500
2000
2500
Rai
nfa
ll (m
m)
Jabiru
Wildman river
0
1
2
3
4
5
6
7
8
92/3 93/4 94/5 95/6 96/7 97/8 98/9 99/0 00/1 01/2
No
. mo
nth
s ra
infa
ll >5
0 m
m
Figure 1 Available rain-year rainfall data for recording stations
closest to study sites: Jabiru – PS and SS; Wildman River – PL and
SL (source: Australian Bureau of Meteorology).
J. Russell-Smith and S. A. Setterfield
1600 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

had inconspicuous, small and short-lived seeds may not
have been detected.
Soil seed bank
Soils were collected at the end of June in 1993, 1994 and 1995.
Twenty surface soil samples (0–5 cm) were collected randomly
within transects, bulked together, mixed thoroughly, split into
5–10 sub-samples per sampling period, stored at room
temperature in calico bags, and subsequently laid out in shallow
trays on a bed of sterile sand for germination in a shade-house as
per the seed rain samples. All soil sub-samples were weighed
prior to setting out in germination trays, and small sub-samples
taken for determination of oven dry weight. Germination trials
of 10 soil sub-samples collected in 1993, 1994, 1995 were
conducted in the following January to assess dormant seed
banks (sensu Russell-Smith & Lucas, 1994). Additionally, for
soil sub-samples collected in 1993, germination trials of 10 sub-
samples were undertaken immediately after collection for
assessment of post-wet season seed banks, and 5 sub-samples
were taken in January 1996 for assessment of seed bank
longevity. Germination trials were conducted over at least
6 months in all assessment years, under similar conditions as for
seed rain germination trials. Soil seed bank data are presented as
numbers of germinants per kg of oven dry weight soil.
Seedlings and clonal sprouts
Seedlings (and clonal sprouts < 50 cm tall), and saplings (50–
200 cm tall; 200 cm tall to 5 cm d.b.h.) were identified to
species and counted separately in each of the 25 1-m2
permanent quadrats. Individuals were not tagged but, where
required for determination of clonal reproduction, seedling-
sized clonal sprouts were identified by non-destructive exca-
vation. Sampling was undertaken first in late September 1993,
then in April/May and September/October (before the rains of
the ensuing wet season) in 1994 and 1995. Thereafter,
sampling was undertaken only in September/October (before
the rains) of each year until 2002, given that data from 1994–
95 indicated that, typically, minimum annual survivorship of
woody seedlings occurred at this time (see Results).
Functional groups
Data are presented for six functional groups: rain forest trees,
rain forest shrubs, savanna trees, savanna shrubs, graminoids,
and herbs. Rain forest woody taxa are those as listed by Liddle
et al. (1994). Savanna woody taxa include Callitris, Eucalyptus,
Corymbia, Melaleuca spp. and Acacia spp., with the exception
of the rain forest taxon A. auriculiformis. Nomenclature follows
Cowie & Albrecht (2005). Tree taxa are defined as those
typically attaining > 8 m height. Woody lianes/vines are
included here as trees or shrubs, depending on their stature.
Graminoids comprise Cyperaceae and Poaceae. Herbs are
defined as non-woody non-graminoids.
RESULTS
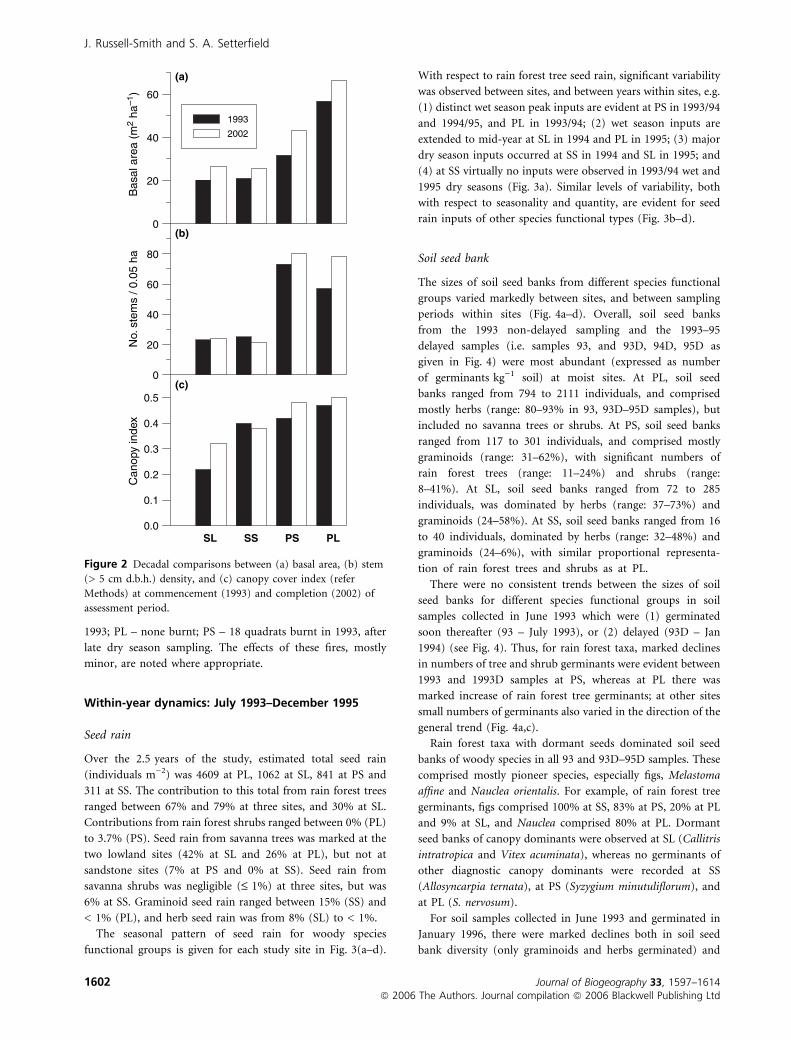
Basal area, stem density and canopy cover was greater at
perennially moist sites (PL and PS) than at seasonally dry sites
(Fig. 2). Over the 10-year assessment period, basal area
increased in all plots by between 17% (PL) and 36% (PS),
while stem density and canopy cover increased in three plots
but decreased marginally at SS (Fig. 2).
Over the period of assessment, low intensity, late dry season,
typically patchy fires were experienced at sites as follows: SS –
all 25 sampling quadrats burnt in 1994, four quadrats burnt in
1997, 10 quadrats burnt in 2001; SL – all 25 quadrats burnt in
Table 2 Sampling regime for different study components, where sampling periods given in quarters as follows: 1 ¼ January–March;
2 ¼ April–June; 3 ¼ July–September; 4 ¼ October–December
Study component
Sampling period
Within-year dynamics Decadal dynamics
1993 1994 1995 1996 1996 1997 1998 1999 2000 2001 2002
Quarter Quarter
2 3 4 1 2 3 4 1 2 3 4 1 2 3 3 3 3 3 3 3
Seed rain assessment
Sampling · · · · · · · · · ·Germination · · · · · ·
Soil seed bank assessments
Sampling · · ·Non-delayed germination · ·Delayed germination (6 months) · · · · · ·Delayed germination (3 years) · ·
Seedling, sapling counts · · · · · · · · · · · ·Tree stem, canopy cover, inventory · ·
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1601ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
1993; PL – none burnt; PS – 18 quadrats burnt in 1993, after
late dry season sampling. The effects of these fires, mostly
minor, are noted where appropriate.
Within-year dynamics: July 1993–December 1995
Seed rain
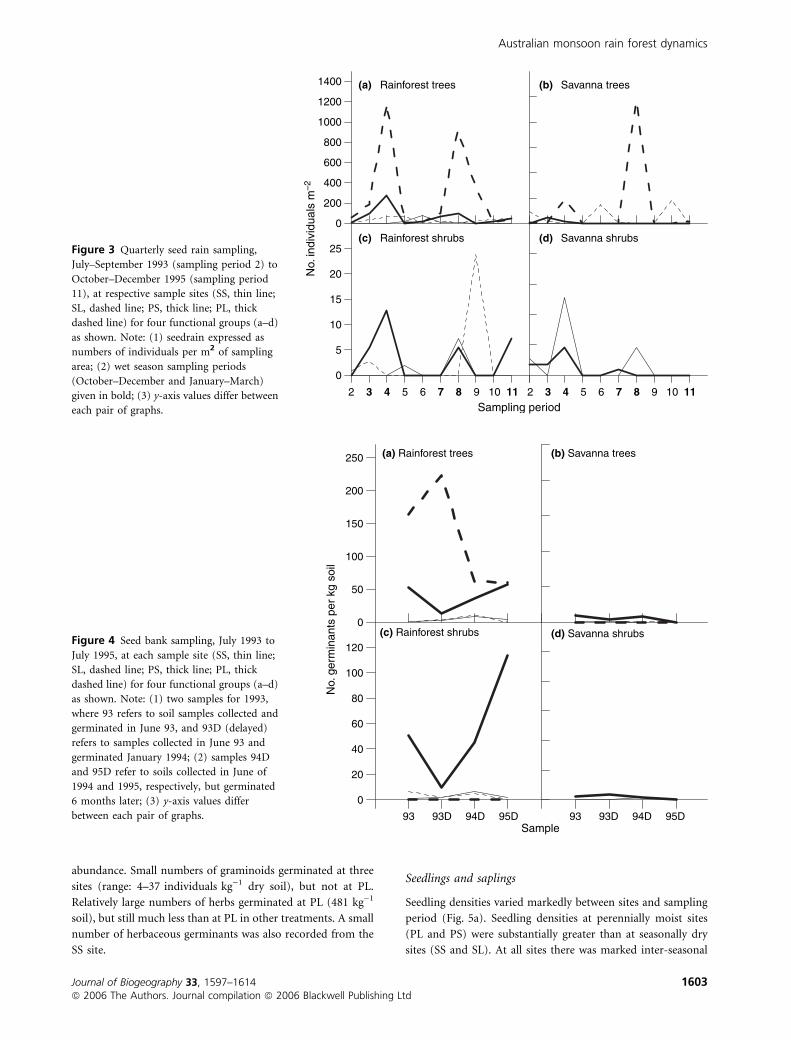
Over the 2.5 years of the study, estimated total seed rain
(individuals m)2) was 4609 at PL, 1062 at SL, 841 at PS and
311 at SS. The contribution to this total from rain forest trees
ranged between 67% and 79% at three sites, and 30% at SL.
Contributions from rain forest shrubs ranged between 0% (PL)
to 3.7% (PS). Seed rain from savanna trees was marked at the
two lowland sites (42% at SL and 26% at PL), but not at
sandstone sites (7% at PS and 0% at SS). Seed rain from
savanna shrubs was negligible (£ 1%) at three sites, but was
6% at SS. Graminoid seed rain ranged between 15% (SS) and
< 1% (PL), and herb seed rain was from 8% (SL) to < 1%.
The seasonal pattern of seed rain for woody species
functional groups is given for each study site in Fig. 3(a–d).
With respect to rain forest tree seed rain, significant variability
was observed between sites, and between years within sites, e.g.
(1) distinct wet season peak inputs are evident at PS in 1993/94
and 1994/95, and PL in 1993/94; (2) wet season inputs are
extended to mid-year at SL in 1994 and PL in 1995; (3) major
dry season inputs occurred at SS in 1994 and SL in 1995; and
(4) at SS virtually no inputs were observed in 1993/94 wet and
1995 dry seasons (Fig. 3a). Similar levels of variability, both
with respect to seasonality and quantity, are evident for seed
rain inputs of other species functional types (Fig. 3b–d).
Soil seed bank
The sizes of soil seed banks from different species functional
groups varied markedly between sites, and between sampling
periods within sites (Fig. 4a–d). Overall, soil seed banks
from the 1993 non-delayed sampling and the 1993–95
delayed samples (i.e. samples 93, and 93D, 94D, 95D as
given in Fig. 4) were most abundant (expressed as number
of germinants kg)1 soil) at moist sites. At PL, soil seed
banks ranged from 794 to 2111 individuals, and comprised
mostly herbs (range: 80–93% in 93, 93D–95D samples), but
included no savanna trees or shrubs. At PS, soil seed banks
ranged from 117 to 301 individuals, and comprised mostly
graminoids (range: 31–62%), with significant numbers of
rain forest trees (range: 11–24%) and shrubs (range:
8–41%). At SL, soil seed banks ranged from 72 to 285
individuals, was dominated by herbs (range: 37–73%) and
graminoids (24–58%). At SS, soil seed banks ranged from 16
to 40 individuals, dominated by herbs (range: 32–48%) and
graminoids (24–6%), with similar proportional representa-
tion of rain forest trees and shrubs as at PL.
There were no consistent trends between the sizes of soil
seed banks for different species functional groups in soil
samples collected in June 1993 which were (1) germinated
soon thereafter (93 – July 1993), or (2) delayed (93D – Jan
1994) (see Fig. 4). Thus, for rain forest taxa, marked declines
in numbers of tree and shrub germinants were evident between
1993 and 1993D samples at PS, whereas at PL there was
marked increase of rain forest tree germinants; at other sites
small numbers of germinants also varied in the direction of the
general trend (Fig. 4a,c).
Rain forest taxa with dormant seeds dominated soil seed
banks of woody species in all 93 and 93D–95D samples. These
comprised mostly pioneer species, especially figs, Melastoma
affine and Nauclea orientalis. For example, of rain forest tree
germinants, figs comprised 100% at SS, 83% at PS, 20% at PL
and 9% at SL, and Nauclea comprised 80% at PL. Dormant
seed banks of canopy dominants were observed at SL (Callitris
intratropica and Vitex acuminata), whereas no germinants of
other diagnostic canopy dominants were recorded at SS
(Allosyncarpia ternata), at PS (Syzygium minutuliflorum), and
at PL (S. nervosum).
For soil samples collected in June 1993 and germinated in
January 1996, there were marked declines both in soil seed
bank diversity (only graminoids and herbs germinated) and
0
20
40
60
Bas
al a
rea
(m2
ha–1
)
0
20
40
60
80
No.
ste
ms
/ 0.0
5 ha
0.0
0.1
0.2
0.3
0.4
0.5
Can
opy
inde
x1993
2002
(a)
(b)
(c)
SL SS PS PL
Figure 2 Decadal comparisons between (a) basal area, (b) stem
(> 5 cm d.b.h.) density, and (c) canopy cover index (refer
Methods) at commencement (1993) and completion (2002) of
assessment period.
J. Russell-Smith and S. A. Setterfield
1602 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
abundance. Small numbers of graminoids germinated at three
sites (range: 4–37 individuals kg)1 dry soil), but not at PL.
Relatively large numbers of herbs germinated at PL (481 kg)1
soil), but still much less than at PL in other treatments. A small
number of herbaceous germinants was also recorded from the
SS site.
Seedlings and saplings
Seedling densities varied markedly between sites and sampling
period (Fig. 5a). Seedling densities at perennially moist sites
(PL and PS) were substantially greater than at seasonally dry
sites (SS and SL). At all sites there was marked inter-seasonal
0
200
400
600
800
1000
1200
1400 Rainforest trees Savanna trees
0
5
10
15
20
25
No.
indi
vidu
als
m–2
(c) (d)
2 3 4 5 6 7 8 9 10 11 2 3 4 5 6 7 8 9 10 11Sampling period
(a)
Rainforest shrubs
(b)
Savanna shrubsFigure 3 Quarterly seed rain sampling,
July–September 1993 (sampling period 2) to
October–December 1995 (sampling period
11), at respective sample sites (SS, thin line;
SL, dashed line; PS, thick line; PL, thick
dashed line) for four functional groups (a–d)
as shown. Note: (1) seedrain expressed as
numbers of individuals per m2 of sampling
area; (2) wet season sampling periods
(October–December and January–March)
given in bold; (3) y-axis values differ between
each pair of graphs.
0
50
100
150
200
250 (a) Rainforest trees (b) Savanna trees
0
20
40
60
80
100
120
No.
ger
min
ants
per
kg
soil
(c) Rainforest shrubs (d) Savanna shrubs
Sample 93 93D 94D 95D 93 93D 94D 95D
Figure 4 Seed bank sampling, July 1993 to
July 1995, at each sample site (SS, thin line;
SL, dashed line; PS, thick line; PL, thick
dashed line) for four functional groups (a–d)
as shown. Note: (1) two samples for 1993,
where 93 refers to soil samples collected and
germinated in June 93, and 93D (delayed)
refers to samples collected in June 93 and
germinated January 1994; (2) samples 94D
and 95D refer to soils collected in June of
1994 and 1995, respectively, but germinated
6 months later; (3) y-axis values differ
between each pair of graphs.
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1603ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
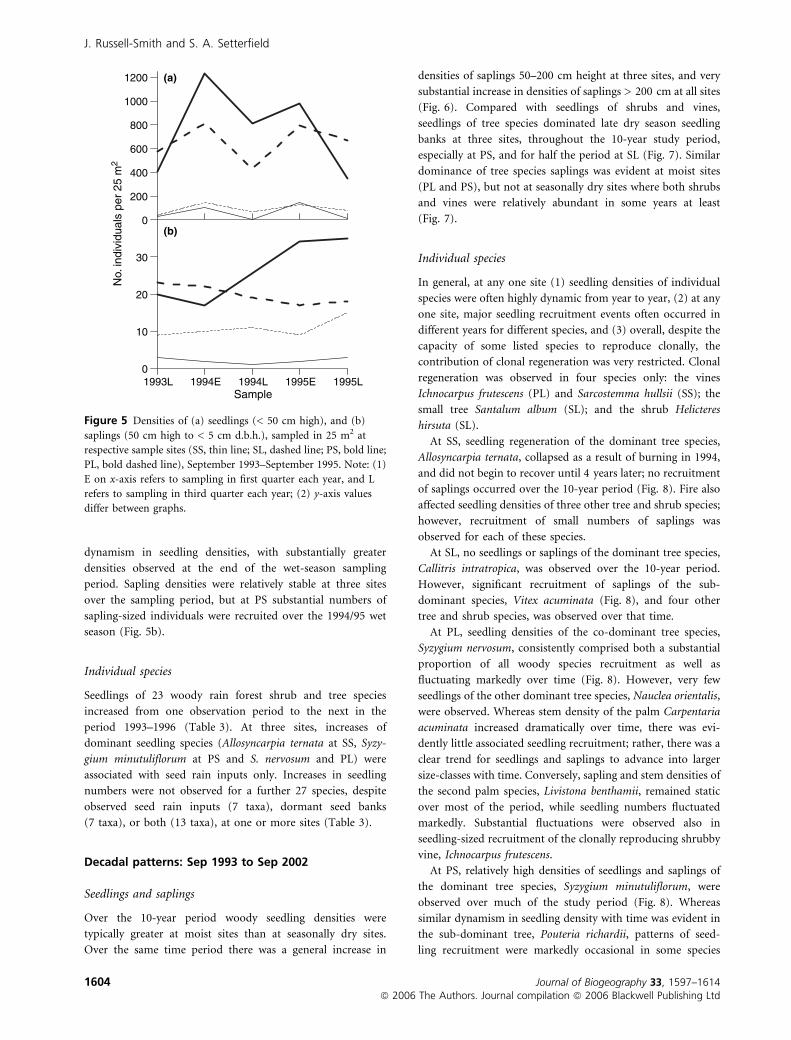
dynamism in seedling densities, with substantially greater
densities observed at the end of the wet-season sampling
period. Sapling densities were relatively stable at three sites
over the sampling period, but at PS substantial numbers of
sapling-sized individuals were recruited over the 1994/95 wet
season (Fig. 5b).
Individual species
Seedlings of 23 woody rain forest shrub and tree species
increased from one observation period to the next in the
period 1993–1996 (Table 3). At three sites, increases of
dominant seedling species (Allosyncarpia ternata at SS, Syzy-
gium minutuliflorum at PS and S. nervosum and PL) were
associated with seed rain inputs only. Increases in seedling
numbers were not observed for a further 27 species, despite
observed seed rain inputs (7 taxa), dormant seed banks
(7 taxa), or both (13 taxa), at one or more sites (Table 3).
Decadal patterns: Sep 1993 to Sep 2002
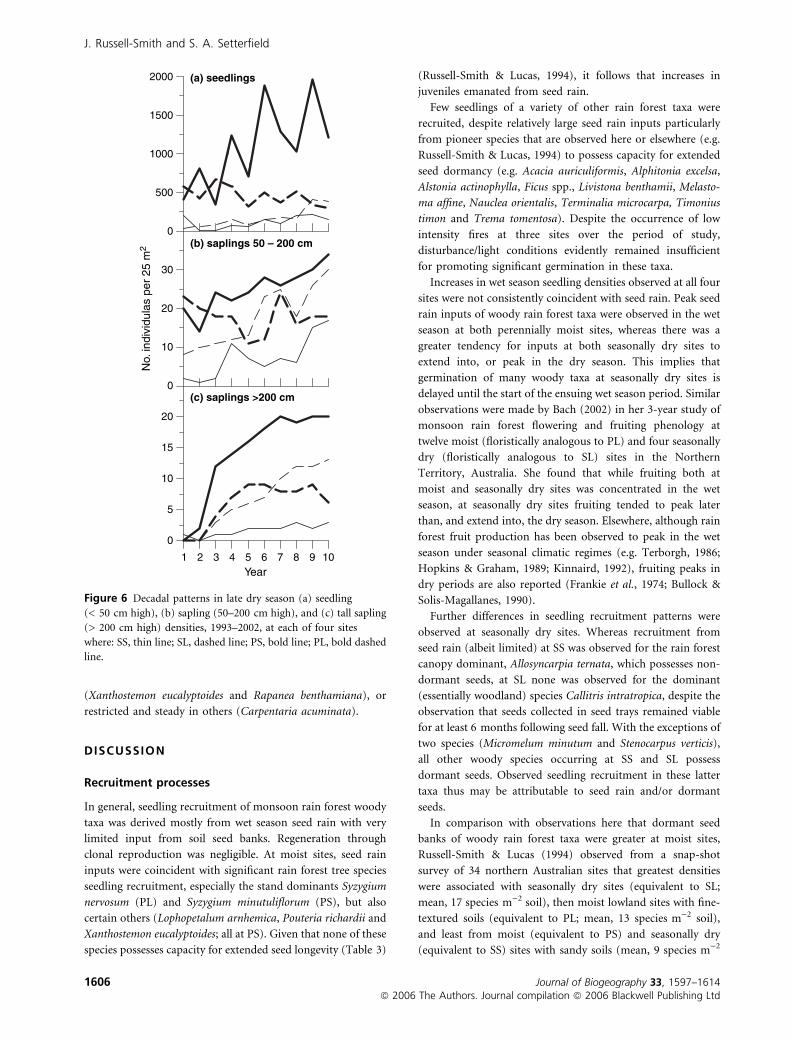
Seedlings and saplings
Over the 10-year period woody seedling densities were
typically greater at moist sites than at seasonally dry sites.
Over the same time period there was a general increase in
densities of saplings 50–200 cm height at three sites, and very
substantial increase in densities of saplings > 200 cm at all sites
(Fig. 6). Compared with seedlings of shrubs and vines,
seedlings of tree species dominated late dry season seedling
banks at three sites, throughout the 10-year study period,
especially at PS, and for half the period at SL (Fig. 7). Similar
dominance of tree species saplings was evident at moist sites
(PL and PS), but not at seasonally dry sites where both shrubs
and vines were relatively abundant in some years at least
(Fig. 7).
Individual species
In general, at any one site (1) seedling densities of individual
species were often highly dynamic from year to year, (2) at any
one site, major seedling recruitment events often occurred in
different years for different species, and (3) overall, despite the
capacity of some listed species to reproduce clonally, the
contribution of clonal regeneration was very restricted. Clonal
regeneration was observed in four species only: the vines
Ichnocarpus frutescens (PL) and Sarcostemma hullsii (SS); the
small tree Santalum album (SL); and the shrub Helicteres
hirsuta (SL).
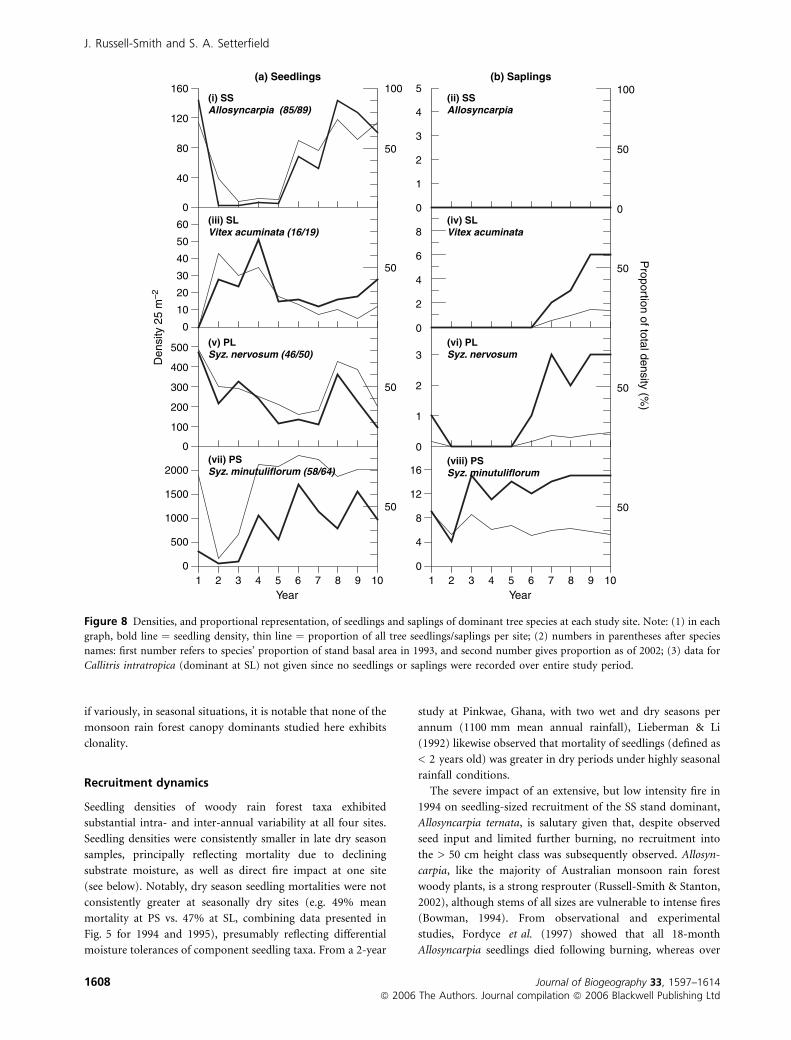
At SS, seedling regeneration of the dominant tree species,
Allosyncarpia ternata, collapsed as a result of burning in 1994,
and did not begin to recover until 4 years later; no recruitment
of saplings occurred over the 10-year period (Fig. 8). Fire also
affected seedling densities of three other tree and shrub species;
however, recruitment of small numbers of saplings was
observed for each of these species.
At SL, no seedlings or saplings of the dominant tree species,
Callitris intratropica, was observed over the 10-year period.
However, significant recruitment of saplings of the sub-
dominant species, Vitex acuminata (Fig. 8), and four other
tree and shrub species, was observed over that time.
At PL, seedling densities of the co-dominant tree species,
Syzygium nervosum, consistently comprised both a substantial
proportion of all woody species recruitment as well as
fluctuating markedly over time (Fig. 8). However, very few
seedlings of the other dominant tree species, Nauclea orientalis,
were observed. Whereas stem density of the palm Carpentaria
acuminata increased dramatically over time, there was evi-
dently little associated seedling recruitment; rather, there was a
clear trend for seedlings and saplings to advance into larger
size-classes with time. Conversely, sapling and stem densities of
the second palm species, Livistona benthamii, remained static
over most of the period, while seedling numbers fluctuated
markedly. Substantial fluctuations were observed also in
seedling-sized recruitment of the clonally reproducing shrubby
vine, Ichnocarpus frutescens.
At PS, relatively high densities of seedlings and saplings of
the dominant tree species, Syzygium minutuliflorum, were
observed over much of the study period (Fig. 8). Whereas
similar dynamism in seedling density with time was evident in
the sub-dominant tree, Pouteria richardii, patterns of seed-
ling recruitment were markedly occasional in some species
0
200
400
600
800
1000
1200
01993L 1994E 1994L 1995E 1995L
10
20
30
No.
indi
vidu
als
per
25 m
2
(b)
(a)
Sample
Figure 5 Densities of (a) seedlings (< 50 cm high), and (b)
saplings (50 cm high to < 5 cm d.b.h.), sampled in 25 m2 at
respective sample sites (SS, thin line; SL, dashed line; PS, bold line;
PL, bold dashed line), September 1993–September 1995. Note: (1)
E on x-axis refers to sampling in first quarter each year, and L
refers to sampling in third quarter each year; (2) y-axis values
differ between graphs.
J. Russell-Smith and S. A. Setterfield
1604 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Table 3 Responses of woody rain forest species at each study site, where: (a) seedlings (< 50 cm high) of listed taxa increased from one
observation period to the next over the period 1993–1996; and/or (b) seedrain and (c) seedbanks were observed in the period 1993–1995. After
species names # indicates taxa observed in this study, or elsewhere (Russell-Smith, 1996) to reproduce clonally; * indicates taxa observed
(Russell-Smith & Lucas, 1994) to possess dormant seedbanks. Family abbreviations given in parentheses, and given in full at end of table
Species
Seedlings Seedrain Seedbank
Site Site Site
PL PS SL SS PL PS SL SS PL PS SL SS
Acacia auriculiformis* (Mim.) + +
Aidia racemosa#* (Rub.) + + + +
Allosyncarpia ternata (Myrt.) + +
Alphitonia excelsa* (Rhamn.) + + +
Alstonia actinophylla* (Apoc.) +
Antidesma parvifolium# (Euph.) +
Breynia cernua#* (Euph.) +
Buchanania arborescens (Anac.) + +
Callitris intratropica (Cupr.) + +
Calophyllum sil (Clus.) + +
Canarium australianum* (Burs.) +
Canthium coprosmoides (Rub.) +
Carpentaria acuminata (Arec.) + + + +
Celtis philippensis (Ulm.) +
Choriceras tricorne# (Euph.) + +
Diospyros compacta# (Eben.) + +
Drypetes deplanchei# (Euph.) + + + +
Ficus leucotricha* (Mor.) + +
Ficus platypoda* (Mor.) + + +
Ficus racemosa* (Mor.) + + + +
Ficus scobina* (Mor.) + + + + + + +
Ficus virens* (Mor.) + + + + + + + +
Gmelina schlecteri (Verb.) +
Helicia australasica (Prot.) +
Helicteres hirsuta# (Sterc.) +
Horsfieldia australiana (Myrist.) + +
Ilex arnhemensis (Aquif.) +
Livistona benthamii (Arec.) + +
Lophopetalum arnhemica (Cel.) + +
Maranthes corymbosa* (Chrys.) +
Melastoma affine#* (Log.) + + + +
Melicope elleryana* (Rut.) + + + +
Nauclea orientalis* (Rub.) + + +
Notelaea microcarpa (Ole.) +
Polyalthia nitidissima (Ann.) + +
Pouteria richardii (Sap.) + +
Pouteria sericea* (Sap.) + + +
Rapanaea benthamiana (Myrs.) + +
Santalum album# (Lor.) +
Stenocarpus verticis (Prot.) + +
Strychnos lucida#* (Log.) + + + +
Syzygium minutuliflorum (Myrt.) + +
Syzygium nervosum (Myrt.) + +
Terminalia microcarpa* (Comb.) + + +
Timonius timon* (Rub.) + + +
Trema tomentosa* (Ulm.) + + + + + + +
Vavaea australiana (Sap.) + +
Vitex acuminata (Verb.) + + +
Vitex glabrata* (Verb.) + +
Xanthostemon eucalyptoides (Myrt.) + +
Abbreviations of family names: Anac. ¼ Anacardiaceae; Ann. ¼ Annonaceae; Apoc. ¼ Apocynaceae; Aquif. ¼ Aquifoliaceae; Arec. ¼ Arecaceae; Burs. ¼ Burseraceae;
Cel. ¼ Celastraceae; Chrys. ¼ Chrysobalanaceae; Clus. ¼ Clusiaceae; Comb. ¼ Combretaceae; Cupr. ¼ Cupressaceae; Eben. ¼ Ebenaceae; Euph. ¼ Euphorbiaceae;
Log. ¼ Loganiaceae; Lor. ¼ Loranthaceae; Mim. ¼ Mimosaceae; Mor. ¼ Moraceae; Myrist. ¼ Myristicaceae; Myrs. ¼ Myrsinaceae; Myrt. ¼ Myrtaceae;
Ole. ¼ Oleaceae; Prot. ¼ Proteaceae; Rhamn. ¼ Rhamnaceae; Rub. ¼ Rubiaceae; Rut. ¼ Rutaceae; Sap. ¼ Sapotaceae; Sterc. ¼ Sterculiaceae; Ulm. ¼ Ulmaceae;
Verb. ¼ Verbenaceae.
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1605ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
(Xanthostemon eucalyptoides and Rapanea benthamiana), or
restricted and steady in others (Carpentaria acuminata).
DISCUSSION
Recruitment processes
In general, seedling recruitment of monsoon rain forest woody
taxa was derived mostly from wet season seed rain with very
limited input from soil seed banks. Regeneration through
clonal reproduction was negligible. At moist sites, seed rain
inputs were coincident with significant rain forest tree species
seedling recruitment, especially the stand dominants Syzygium
nervosum (PL) and Syzygium minutuliflorum (PS), but also
certain others (Lophopetalum arnhemica, Pouteria richardii and
Xanthostemon eucalyptoides; all at PS). Given that none of these
species possesses capacity for extended seed longevity (Table 3)
(Russell-Smith & Lucas, 1994), it follows that increases in
juveniles emanated from seed rain.
Few seedlings of a variety of other rain forest taxa were
recruited, despite relatively large seed rain inputs particularly
from pioneer species that are observed here or elsewhere (e.g.
Russell-Smith & Lucas, 1994) to possess capacity for extended
seed dormancy (e.g. Acacia auriculiformis, Alphitonia excelsa,
Alstonia actinophylla, Ficus spp., Livistona benthamii, Melasto-
ma affine, Nauclea orientalis, Terminalia microcarpa, Timonius
timon and Trema tomentosa). Despite the occurrence of low
intensity fires at three sites over the period of study,
disturbance/light conditions evidently remained insufficient
for promoting significant germination in these taxa.
Increases in wet season seedling densities observed at all four
sites were not consistently coincident with seed rain. Peak seed
rain inputs of woody rain forest taxa were observed in the wet
season at both perennially moist sites, whereas there was a
greater tendency for inputs at both seasonally dry sites to
extend into, or peak in the dry season. This implies that
germination of many woody taxa at seasonally dry sites is
delayed until the start of the ensuing wet season period. Similar
observations were made by Bach (2002) in her 3-year study of
monsoon rain forest flowering and fruiting phenology at
twelve moist (floristically analogous to PL) and four seasonally
dry (floristically analogous to SL) sites in the Northern
Territory, Australia. She found that while fruiting both at
moist and seasonally dry sites was concentrated in the wet
season, at seasonally dry sites fruiting tended to peak later
than, and extend into, the dry season. Elsewhere, although rain
forest fruit production has been observed to peak in the wet
season under seasonal climatic regimes (e.g. Terborgh, 1986;
Hopkins & Graham, 1989; Kinnaird, 1992), fruiting peaks in
dry periods are also reported (Frankie et al., 1974; Bullock &
Solis-Magallanes, 1990).
Further differences in seedling recruitment patterns were
observed at seasonally dry sites. Whereas recruitment from
seed rain (albeit limited) at SS was observed for the rain forest
canopy dominant, Allosyncarpia ternata, which possesses non-
dormant seeds, at SL none was observed for the dominant
(essentially woodland) species Callitris intratropica, despite the
observation that seeds collected in seed trays remained viable
for at least 6 months following seed fall. With the exceptions of
two species (Micromelum minutum and Stenocarpus verticis),
all other woody species occurring at SS and SL possess
dormant seeds. Observed seedling recruitment in these latter
taxa thus may be attributable to seed rain and/or dormant
seeds.
In comparison with observations here that dormant seed
banks of woody rain forest taxa were greater at moist sites,
Russell-Smith & Lucas (1994) observed from a snap-shot
survey of 34 northern Australian sites that greatest densities
were associated with seasonally dry sites (equivalent to SL;
mean, 17 species m)2 soil), then moist lowland sites with fine-
textured soils (equivalent to PL; mean, 13 species m)2 soil),
and least from moist (equivalent to PS) and seasonally dry
(equivalent to SS) sites with sandy soils (mean, 9 species m)2
0
500
1000
1500
2000 (a) seedlings
0
10
20
30
No.
indi
vidu
las
per
25 m
2 (b) saplings 50 – 200 cm
1 2 3 4 5 6 7 8 9 10Year
0
5
10
15
20
(c) saplings >200 cm
Figure 6 Decadal patterns in late dry season (a) seedling
(< 50 cm high), (b) sapling (50–200 cm high), and (c) tall sapling
(> 200 cm high) densities, 1993–2002, at each of four sites
where: SS, thin line; SL, dashed line; PS, bold line; PL, bold dashed
line.
J. Russell-Smith and S. A. Setterfield
1606 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
soil). We note, however, that generalizations from one-off
studies may be fraught given the considerable temporal
variability in dormant seed banks evident at all four sites. As
in this study, dormant soil seed banks of woody rain forest taxa
were dominated by pioneers, especially figs. Similar observa-
tions concerning the dominance generally of pioneer taxa in
rain forest soil seed banks have been widely reported
(Garwood, 1989). Data presented here indicate that longevity
of dormant seed banks of woody monsoon rain forest taxa,
including figs, is expended under field conditions in less than
3 years.
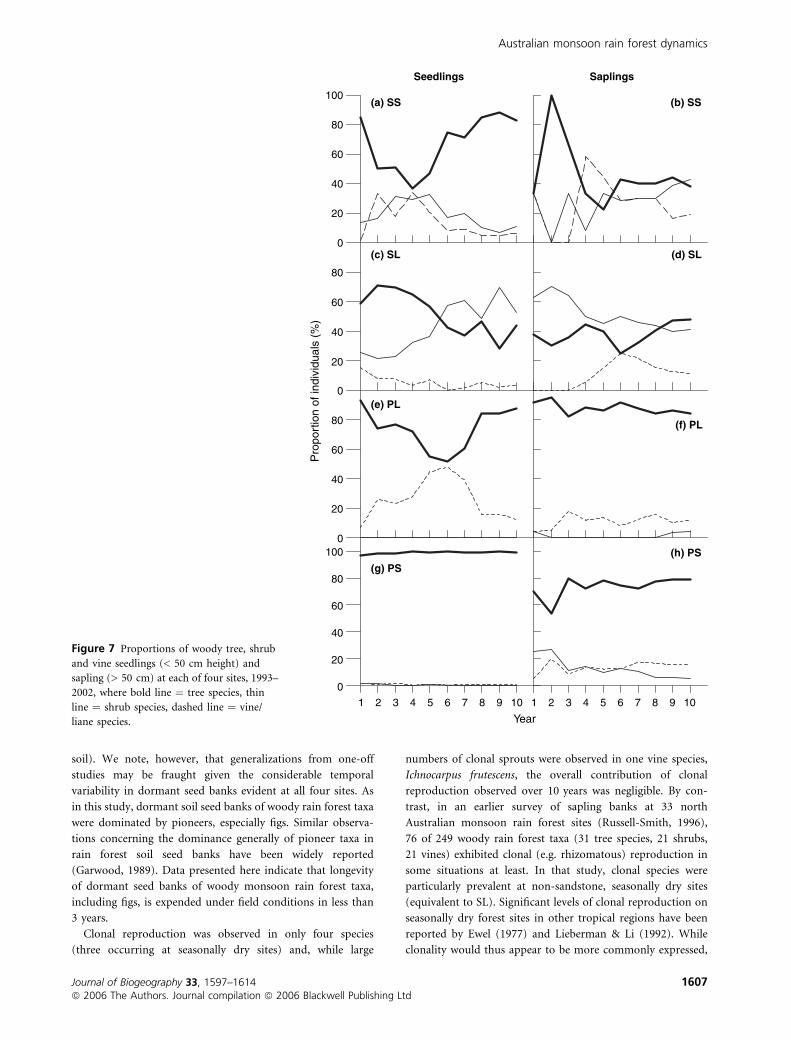
Clonal reproduction was observed in only four species
(three occurring at seasonally dry sites) and, while large
numbers of clonal sprouts were observed in one vine species,
Ichnocarpus frutescens, the overall contribution of clonal
reproduction observed over 10 years was negligible. By con-
trast, in an earlier survey of sapling banks at 33 north
Australian monsoon rain forest sites (Russell-Smith, 1996),
76 of 249 woody rain forest taxa (31 tree species, 21 shrubs,
21 vines) exhibited clonal (e.g. rhizomatous) reproduction in
some situations at least. In that study, clonal species were
particularly prevalent at non-sandstone, seasonally dry sites
(equivalent to SL). Significant levels of clonal reproduction on
seasonally dry forest sites in other tropical regions have been
reported by Ewel (1977) and Lieberman & Li (1992). While
clonality would thus appear to be more commonly expressed,
0
20
40
60
80
100(a) SS (b) SS
0
20
40
60
80
(c) SL (d) SL
0
20
40
60
80
Pro
port
ion
of in
divi
dual
s (%
)
(e) PL
(f) PL
1 2 3 4 5 6 7 8 9 10
Year
0
20
40
60
80
100
(g) PS
1 2 3 4 5 6 7 8 9 10
(h) PS
Seedlings Saplings
Figure 7 Proportions of woody tree, shrub
and vine seedlings (< 50 cm height) and
sapling (> 50 cm) at each of four sites, 1993–
2002, where bold line ¼ tree species, thin
line ¼ shrub species, dashed line ¼ vine/
liane species.
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1607ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
if variously, in seasonal situations, it is notable that none of the
monsoon rain forest canopy dominants studied here exhibits
clonality.
Recruitment dynamics
Seedling densities of woody rain forest taxa exhibited
substantial intra- and inter-annual variability at all four sites.
Seedling densities were consistently smaller in late dry season
samples, principally reflecting mortality due to declining
substrate moisture, as well as direct fire impact at one site
(see below). Notably, dry season seedling mortalities were not
consistently greater at seasonally dry sites (e.g. 49% mean
mortality at PS vs. 47% at SL, combining data presented in
Fig. 5 for 1994 and 1995), presumably reflecting differential
moisture tolerances of component seedling taxa. From a 2-year
study at Pinkwae, Ghana, with two wet and dry seasons per
annum (1100 mm mean annual rainfall), Lieberman & Li
(1992) likewise observed that mortality of seedlings (defined as
< 2 years old) was greater in dry periods under highly seasonal
rainfall conditions.
The severe impact of an extensive, but low intensity fire in
1994 on seedling-sized recruitment of the SS stand dominant,
Allosyncarpia ternata, is salutary given that, despite observed
seed input and limited further burning, no recruitment into
the > 50 cm height class was subsequently observed. Allosyn-
carpia, like the majority of Australian monsoon rain forest
woody plants, is a strong resprouter (Russell-Smith & Stanton,
2002), although stems of all sizes are vulnerable to intense fires
(Bowman, 1994). From observational and experimental
studies, Fordyce et al. (1997) showed that all 18-month
Allosyncarpia seedlings died following burning, whereas over
0
40
80
120
160
50
100(i) SSAllosyncarpia (85/89)
0
10
20
30
40
50
60
50
(iii) SLVitex acuminata (16/19)
0
2
4
6
8
50
(a) Seedlings (b) Saplings
(iv) SLVitex acuminata
0
100
200
300
400
500
Den
sity
25
m–2
50
(v) PLSyz. nervosum (46/50)
0
1
2
3
50
Proportion of total density (%
)
(vi) PLSyz. nervosum
1 2 3 4 5 6 7 8 9 10Year
0
500
1000
1500
2000
50
(vii) PSSyz. minutuliflorum (58/64)
1 2 3 4 5 6 7 8 9 10Year
0
4
8
12
16
50
(viii) PSSyz. minutuliflorum
0
1
2
3
4
5
0
50
100(ii) SSAllosyncarpia
Figure 8 Densities, and proportional representation, of seedlings and saplings of dominant tree species at each study site. Note: (1) in each
graph, bold line ¼ seedling density, thin line ¼ proportion of all tree seedlings/saplings per site; (2) numbers in parentheses after species
names: first number refers to species’ proportion of stand basal area in 1993, and second number gives proportion as of 2002; (3) data for
Callitris intratropica (dominant at SL) not given since no seedlings or saplings were recorded over entire study period.
J. Russell-Smith and S. A. Setterfield
1608 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

half of a 3-year-old seedling cohort (established initially under
nursery conditions) recovered from lignotuber resprouts.
Under the relatively benign fire disturbance, and consistent
rainfall seasonality conditions that existed over the decade of
study, densities of saplings increased overall at all sites. At
moist sites there was substantial recruitment also into the
> 5 cm d.b.h. stem class. Conversely, at seasonally dry sites,
only one individual was recruited into this class at SL, and four
stems were actually lost at SS. Such differential recruitment
rates highlight the relative vulnerability of monsoon rain
forests growing under seasonally dry conditions in the face of
contemporary north Australian fire regimes.
Comparisons with eucalypt-dominated savanna
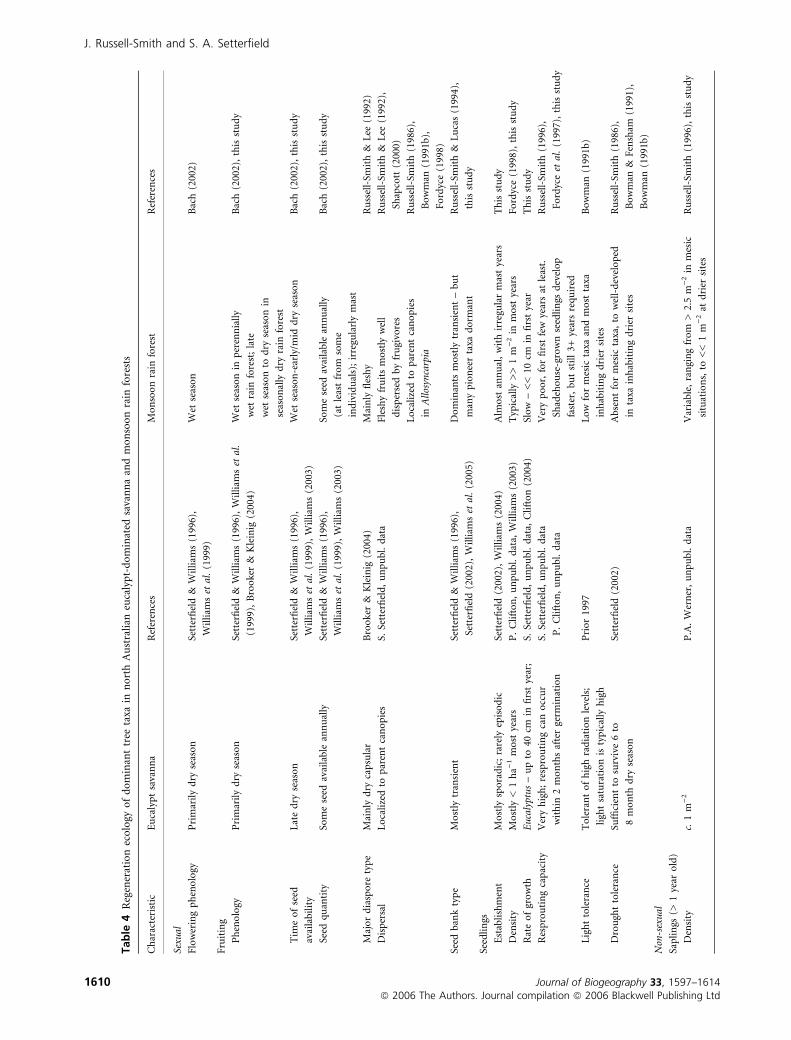
Available data concerning the regeneration ecologies/syn-
dromes of both monsoon rain forest dominants and dominant
northern Australian savanna eucalypts are summarized in
Table 4, derived from a variety of mostly recent studies. Major
contrasts are as follows:
Flowering and fruiting phenology
Flowering and fruiting phenology is strongly seasonal in both
monsoon rain forest and savanna dominants (Williams et al.,
1999; Bach, 2002), although there are significant differences in
timing of major phenological events. Flowering peaks before
the start of the wet season in rain forest at perennially moist
sites, and with the onset of the wet at seasonally dry sites.
Fruiting is concentrated in the wet season at moist sites and
peaks later at seasonally dry lowland sites (Bach, 2002; this
study). Fruiting in Allosyncarpia also occurs in the mid wet
season (Russell-Smith, 1986; Fordyce, 1998). By contrast,
flowering and fruit maturation of eucalypts occurs in the dry
season, with peaks in mid and late dry seasons (Williams et al.,
1999). Seed fall and resultant germination early in the wet
season facilitates establishment during the wet season before
rapid drying out of upper soil layers in the following dry
season (Setterfield & Williams, 1996; Williams, 2003). Newly
recruited seedlings of savanna dominants thus enjoy a longer
first wet season growth period than monsoon rain forest
counterparts at moist sites, and for Allosyncarpia at least in
seasonal forests.
However, this phenological pattern also results in eucalypt
flowering coinciding with current peak fire activity in northern
Australian savannas (Setterfield, 2002; Russell-Smith et al.,
2003a). Low intensity fires in the early dry season can cause a
marked reduction in ovule development and seed production
(Setterfield & Williams, 1996; Williams et al., 2003b). More
intense late dry season fires reduce floral and fruit reserves
across all savanna tree functional groups by more than 50%,
for 2–5 years (Williams et al., 2003b). Such fires also coincide
with early stages of floral primordial development in species
that flower in the early wet season (e.g. Terminalia ferdinan-
diana and Corymbia porrecta), or seed maturation in others
(e.g. Eucalyptus tetrodonta). Fires thus have a major impact on
savanna tree floral phenology, and contribute to the rarity of
sexual regeneration in dominant savanna woody species
(Setterfield, 2002; Williams, 2003). Nothing is known about
the effects of fires on monsoon rain forest reproductive
phenologies, although presumably such impacts would be
negative as for savanna dominants.
Seeds and seed banks
Despite such fire effects, adult eucalypts and other canopy
subdominants are capable of flowering and producing seed
in the season following fire (Setterfield, 2002; Williams et al.,
2003b), and providing annual input to the regeneration pool
even at frequently burnt sites (Williams, 1997; Setterfield,
2002). Dominant savanna species have short seed develop-
ment times (< 4 weeks in E. tetrodonta, E. miniata). The
dominant eucalypts are non-serotinous (Williams et al.,
1999), and seed banks are transient (Setterfield, 2002;
Williams et al., 2005). By contrast, most monsoon rain
forest canopy species have long fruit maturation periods
(Bach, 2002), longer fruiting periods (2–10 months; Bach,
2002), provide an important resource for regional frugivores
(Price et al., 1999; Palmer et al., 2000), and are widely
dispersed (Russell-Smith & Lee, 1992). While seed banks of
diagnostic dominant species are notably transient in three
regional monsoon rain forest types, those of many
co-occurring species exhibit extended longevity, particularly
pioneer taxa and those inhabiting seasonally dry sites.
Seedling density
As with savannas elsewhere, seedling regeneration occurs if
conducive environmental/moisture conditions coincide with
seed fall (Setterfield & Williams, 1996; Higgins et al., 2000).
Due to observed rarity of eucalypt savanna seedlings, however,
there have been few attempts to determine their density. After
8 years of monitoring the effects of fire on savanna at Cape
Cleveland, northern Queensland, Williams et al. (2003a) noted
only one recruitment event of the dominant overstorey tree,
Corymbia clarksoniana. A recent wet season survey for
Eucalyptus miniata seedlings within a 5-m radius of the
canopy of fecund adult trees (i.e. in the area of maximum
seedfall; S. Setterfield, unpubl. data) provided a density
estimate of 1.3 seedlings ha)1 (P. Clifton, unpubl. data). By
contrast, seedling densities of monsoon rain forest dominants
are typically substantially higher (c. 1 to >> 1 m)2), albeit
temporally highly variable, within close proximity to adults
(Russell-Smith, 1996; Fordyce, 1998; this study). Seedling
establishment of dominants is observed almost annually, with
very substantial recruitment in mast years (Fordyce, 1998; this
study).
Seedling growth rates
Vailable, if limited, observations indicate that seedling
growth rates of savanna dominants are substantially greater
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1609ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
Tab
le4
Reg
ener
atio
nec
olo
gyo
fd
om
inan
ttr
eeta
xain
no
rth
Au
stra
lian
euca
lyp
t-d
om
inat
edsa
van
na
and
mo
nso
on
rain
fore
sts
Ch
arac
teri
stic
Eu
caly
pt
sava
nn
aR
efer
ence
sM
on
soo
nra
info
rest
Ref
eren
ces
Sexu
al
Flo
wer
ing
ph
eno
logy
Pri
mar
ily
dry
seas
on
Sett
erfi
eld
&W
illi
ams
(199
6),
Wil
liam
set
al.
(199
9)
Wet
seas
on
Bac
h(2
002)
Fru
itin
g
Ph
eno
logy
Pri
mar
ily
dry
seas
on
Sett
erfi
eld
&W
illi
ams
(199
6),
Wil
liam
set
al.
(199
9),
Bro
oke
r&
Kle
inig
(200
4)
Wet
seas
on
inp
eren
nia
lly
wet
rain
fore
st;
late
wet
seas
on
tod
ryse
aso
nin
seas
on
ally
dry
rain
fore
st
Bac
h(2
002)
,th
isst
ud
y
Tim
eo
fse
ed
avai
lab
ilit
y
Lat
ed
ryse
aso
nSe
tter
fiel
d&
Wil
liam
s(1
996)
,
Wil
liam
set
al.
(199
9),
Wil
liam
s(2
003)
Wet
seas
on
-ear
ly/m
idd
ryse
aso
nB
ach
(200
2),
this
stu
dy
Seed
qu
anti
tySo
me
seed
avai
lab
lean
nu
ally
Sett
erfi
eld
&W
illi
ams
(199
6),
Wil
liam
set
al.
(199
9),
Wil
liam
s(2
003)
Som
ese
edav
aila
ble
ann
ual
ly
(at
leas
tfr
om
som
e
ind
ivid
ual
s);
irre
gula
rly
mas
t
Bac
h(2
002)
,th
isst
ud
y
Maj
or
dia
spo
rety
pe
Mai
nly
dry
cap
sula
rB
roo
ker
&K
lein
ig(2
004)
Mai
nly
fles
hy
Ru
ssel
l-Sm
ith
&L
ee(1
992)
Dis
per
sal
Lo
cali
zed
top
aren
tca
no
pie
sS.
Sett
erfi
eld
,u
np
ub
l.d
ata
Fle
shy
fru
its
mo
stly
wel
l
dis
per
sed
by
fru
givo
res
Lo
cali
zed
top
aren
tca
no
pie
s
inA
llos
ynca
rpia
Ru
ssel
l-Sm
ith
&L
ee(1
992)
,
Shap
cott
(200
0)
Ru
ssel
l-Sm
ith
(198
6),
Bo
wm
an(1
991b
),
Fo
rdyc
e(1
998)
Seed
ban
kty
pe
Mo
stly
tran
sien
tSe
tter
fiel
d&
Wil
liam
s(1
996)
,
Sett
erfi
eld
(200
2),
Wil
liam
set
al.
(200
5)
Do
min
ants
mo
stly
tran
sien
t–
bu
t
man
yp
ion
eer
taxa
do
rman
t
Ru
ssel
l-Sm
ith
&L
uca
s(1
994)
,
this
stu
dy
Seed
lin
gs
Est
abli
shm
ent
Mo
stly
spo
rad
ic;
rare
lyep
iso
dic
Sett
erfi
eld
(200
2),
Wil
liam
s(2
004)
Alm
ost
ann
ual
,w
ith
irre
gula
rm
ast
year
sT
his
stu
dy
Den
sity
Mo
stly
<1
ha)
1m
ost
year
sP
.C
lift
on
,u
np
ub
l.d
ata,
Wil
liam
s(2
003)
Typ
ical
ly>
>1
m)
2in
mo
stye
ars
Fo
rdyc
e(1
998)
,th
isst
ud
y
Rat
eo
fgr
ow
thE
uca
lypt
us
–u
pto
40cm
infi
rst
year
;S.
Sett
erfi
eld
,u
np
ub
l.d
ata,
Cli
fto
n(2
004)
Slo
w–
<<
10cm
infi
rst
year
Th
isst
ud
y
Res
pro
uti
ng
cap
acit
yV
ery
hig
h;
resp
rou
tin
gca
no
ccu
r
wit
hin
2m
on
ths
afte
rge
rmin
atio
n
S.Se
tter
fiel
d,
un
pu
bl.
dat
a
P.
Cli
fto
n,
un
pu
bl.
dat
a
Ver
yp
oo
r,fo
rfi
rst
few
year
sat
leas
t.
Shad
eho
use
-gro
wn
seed
lin
gsd
evel
op
fast
er,
bu
tst
ill
3+ye
ars
req
uir
ed
Ru
ssel
l-Sm
ith
(199
6),
Fo
rdyc
eet
al.
(199
7),
this
stu
dy
Lig
ht
tole
ran
ceT
ole
ran
to
fh
igh
rad
iati
on
leve
ls;
ligh
tsa
tura
tio
nis
typ
ical
lyh
igh
Pri
or
1997
Lo
wfo
rm
esic
taxa
and
mo
stta
xa
inh
abit
ing
dri
ersi
tes
Bo
wm
an(1
991b
)
Dro
ugh
tto
lera
nce
Suffi
cien
tto
surv
ive
6to
8m
on
thd
ryse
aso
n
Sett
erfi
eld
(200
2)A
bse
nt
for
mes
icta
xa,
tow
ell-
dev
elo
ped
inta
xain
hab
itin
gd
rier
site
s
Ru
ssel
l-Sm
ith
(198
6),
Bo
wm
an&
Fen
sham
(199
1),
Bo
wm
an(1
991b
)
Non
-sex
ual
Sap
lin
gs(>
1ye
aro
ld)
Den
sity
c.1
m)
2P
.A.
Wer
ner
,u
np
ub
l.d
ata
Var
iab
le,
ran
gin
gfr
om
>2.
5m
)2
inm
esic
situ
atio
ns,
to<
<1
m)
2at
dri
ersi
tes
Ru
ssel
l-Sm
ith
(199
6),
this
stu
dy
J. Russell-Smith and S. A. Setterfield
1610 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd
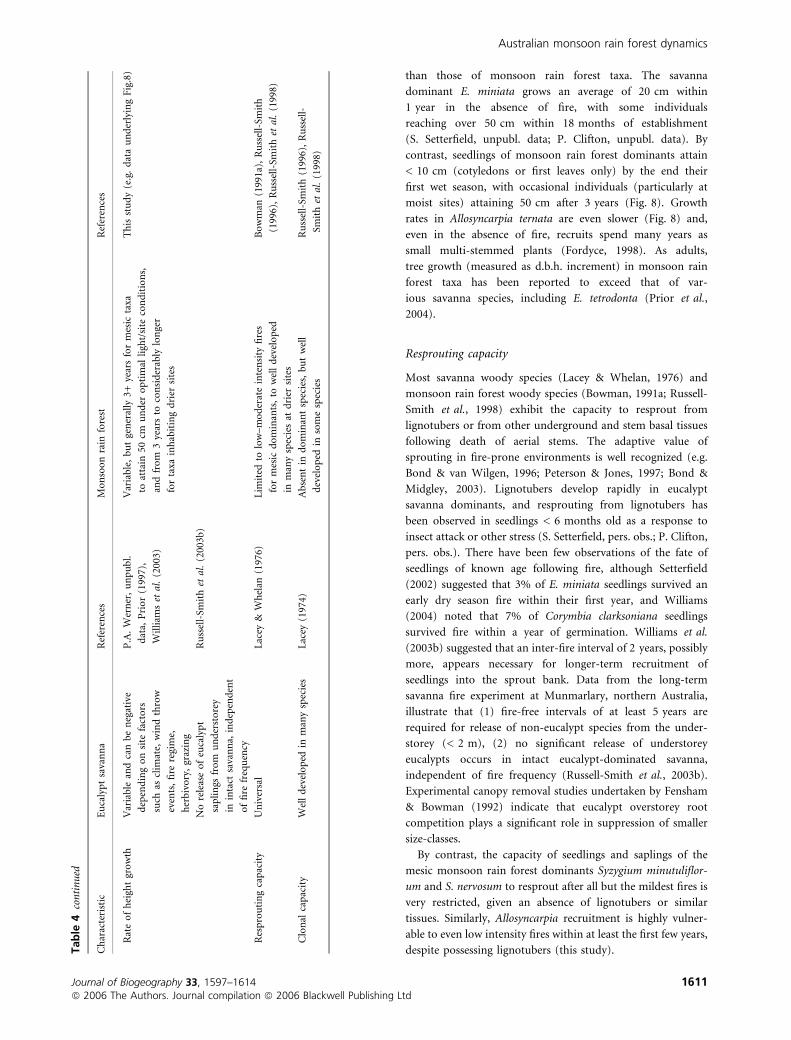
than those of monsoon rain forest taxa. The savanna
dominant E. miniata grows an average of 20 cm within
1 year in the absence of fire, with some individuals
reaching over 50 cm within 18 months of establishment
(S. Setterfield, unpubl. data; P. Clifton, unpubl. data). By
contrast, seedlings of monsoon rain forest dominants attain
< 10 cm (cotyledons or first leaves only) by the end their
first wet season, with occasional individuals (particularly at
moist sites) attaining 50 cm after 3 years (Fig. 8). Growth
rates in Allosyncarpia ternata are even slower (Fig. 8) and,
even in the absence of fire, recruits spend many years as
small multi-stemmed plants (Fordyce, 1998). As adults,
tree growth (measured as d.b.h. increment) in monsoon rain
forest taxa has been reported to exceed that of var-
ious savanna species, including E. tetrodonta (Prior et al.,
2004).
Resprouting capacity
Most savanna woody species (Lacey & Whelan, 1976) and
monsoon rain forest woody species (Bowman, 1991a; Russell-
Smith et al., 1998) exhibit the capacity to resprout from
lignotubers or from other underground and stem basal tissues
following death of aerial stems. The adaptive value of
sprouting in fire-prone environments is well recognized (e.g.
Bond & van Wilgen, 1996; Peterson & Jones, 1997; Bond &
Midgley, 2003). Lignotubers develop rapidly in eucalypt
savanna dominants, and resprouting from lignotubers has
been observed in seedlings < 6 months old as a response to
insect attack or other stress (S. Setterfield, pers. obs.; P. Clifton,
pers. obs.). There have been few observations of the fate of
seedlings of known age following fire, although Setterfield
(2002) suggested that 3% of E. miniata seedlings survived an
early dry season fire within their first year, and Williams
(2004) noted that 7% of Corymbia clarksoniana seedlings
survived fire within a year of germination. Williams et al.
(2003b) suggested that an inter-fire interval of 2 years, possibly
more, appears necessary for longer-term recruitment of
seedlings into the sprout bank. Data from the long-term
savanna fire experiment at Munmarlary, northern Australia,
illustrate that (1) fire-free intervals of at least 5 years are
required for release of non-eucalypt species from the under-
storey (< 2 m), (2) no significant release of understorey
eucalypts occurs in intact eucalypt-dominated savanna,
independent of fire frequency (Russell-Smith et al., 2003b).
Experimental canopy removal studies undertaken by Fensham
& Bowman (1992) indicate that eucalypt overstorey root
competition plays a significant role in suppression of smaller
size-classes.
By contrast, the capacity of seedlings and saplings of the
mesic monsoon rain forest dominants Syzygium minutuliflor-
um and S. nervosum to resprout after all but the mildest fires is
very restricted, given an absence of lignotubers or similar
tissues. Similarly, Allosyncarpia recruitment is highly vulner-
able to even low intensity fires within at least the first few years,
despite possessing lignotubers (this study).Tab
le4
con
tin
ued
Ch
arac
teri
stic
Eu
caly
pt
sava
nn
aR
efer
ence
sM
on
soo
nra
info
rest
Ref
eren
ces
Rat
eo
fh
eigh
tgr
ow
thV
aria
ble
and
can
be
neg
ativ
e
dep
end
ing
on
site
fact
ors
such
ascl
imat
e,w
ind
thro
w
even
ts,
fire
regi
me,
her
biv
ory
,gr
azin
g
No
rele
ase
of
euca
lyp
t
sap
lin
gsfr
om
un
der
sto
rey
inin
tact
sava
nn
a,in
dep
end
ent
of
fire
freq
uen
cy
P.A
.W
ern
er,
un
pu
bl.
dat
a,P
rio
r(1
997)
,
Wil
liam
set
al.
(200
3)
Ru
ssel
l-Sm
ith
etal
.(2
003b
)
Var
iab
le,
bu
tge
ner
ally
3+ye
ars
for
mes
icta
xa
toat
tain
50cm
un
der
op
tim
alli
ght/
site
con
dit
ion
s,
and
fro
m3
year
sto
con
sid
erab
lylo
nge
r
for
taxa
inh
abit
ing
dri
ersi
tes
Th
isst
ud
y(e
.g.
dat
au
nd
erly
ing
Fig
.8)
Res
pro
uti
ng
cap
acit
yU
niv
ersa
lL
acey
&W
hel
an(1
976)
Lim
ited
tolo
w–
mo
der
ate
inte
nsi
tyfi
res
for
mes
icd
om
inan
ts,
tow
ell
dev
elo
ped
inm
any
spec
ies
atd
rier
site
s
Bo
wm
an(1
991a
),R
uss
ell-
Smit
h
(199
6),
Ru
ssel
l-Sm
ith
etal
.(1
998)
Clo
nal
cap
acit
yW
ell
dev
elo
ped
inm
any
spec
ies
Lac
ey(1
974)
Ab
sen
tin
do
min
ant
spec
ies,
bu
tw
ell
dev
elo
ped
inso
me
spec
ies
Ru
ssel
l-Sm
ith
(199
6),
Ru
ssel
l-
Smit
het
al.
(199
8)
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1611ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Clonal capacity
Many Australian savanna species, including various eucalypt
dominants, reproduce clonally via sprouting from roots
(i.e. root suckers), rhizomes or stolons (Lacey, 1974; Lacey
& Whelan, 1976). Such vegetative regeneration offers obvious
advantages for recovery from, and recruitment of new shoots
following, burning – possibly at minimal developmental cost
(Peterson & Jones, 1997). Clonal eucalypts are found mainly
in tropical savannas, and apparently rarely elsewhere (Gill,
1997). Eucalyptus tetrodonta is the only eucalypt that can
produce aerial leafy shoots from the growth of root buds.
This trait is also exhibited by Erythrophleum chlorostachys,
another common savanna co-dominant. Superficially similar
reproduction is seen in the bloodwoods (Corymbia porrecta,
C. ptychocarpa and C. jacobsiana) but the aerial leafy shoots
develop from shallow rhizomes (Brooker & Kleinig, 2004). By
contrast, none of the monsoon rain forest dominants at the
four study sites possesses this regenerative capacity.
In summary, by comparison with dominant savanna
eucalypts, dominant monsoon rain forest taxa recruit substan-
tially greater stocks of seedlings, but exhibit slower aerial growth
and development of resprouting capacity in early years, lack
lignotubers in mesic species, and lack capacity for clonal
reproduction. Given annual–biennial fires in many north
Australian savannas, the study underscores the relative vulner-
ability of monsoon rain forest assemblages, especially those
comprising slower growing species on seasonally dry sites. As
noted by Hoffmann (1998) and Setterfield (2002), the reliance
on sexual as opposed to vegetative reproduction places such taxa
at significant disadvantage in fire-prone savanna environments.
ACKNOWLEDGEMENTS
Particular thanks are due to Diane Lucas for ongoing assistance
with field sampling, the staff of Kakadu National Park for
substantial logistical assistance, the traditional Aboriginal
owners of Kakadu National Park for permission to access
study sites, and J.S. Bach for inspiration. Professors Pat
Werner, Gordon Duff and two anonymous referees made
helpful comments on the text.
REFERENCES
Bach, C.S. (2002) Phenological patterns in monsoon rainfor-
ests in the Northern Territory, Australia. Austral Ecology, 27,
477–489.
Bond, W.J. & Midgley, J.J. (2003) The evolutionary ecology of
sprouting in woody plants. International Journal of Plant
Science, 164 (Suppl.): S103–S114.
Bond, W.J. & van Wilgen, B.W. (1996) Fire and plants.
Chapman & Hall, London, UK.
Bowman, D.M.J.S. (1991a) Recovery of some northern Aus-
tralian monsoon forest tree species following fire. Proceed-
ings of the Royal Society of Queensland, 101, 21–25.
Bowman, D.M.J.S. (1991b) Environmental determinants of
Allosyncarpia ternata forests that are endemic to western
Arnhem Land, northern Australia. Australian Journal of
Botany, 39, 575–589.
Bowman, D.M.J.S. (1994) Preliminary observations on the
mortality of Allosyncarpia ternata stems on the Arnhem
Land plateau, northern Australia. Australian Forestry, 57,
62–64.
Bowman, D.M.J.S. (1999) Australian rainforests: islands of green
in the land of fire. Cambridge University Press, Cambridge.
Bowman, D.M.J.S. & Fensham, R.J. (1991) Response of a
monsoon forest-savanna boundary to fire protection, Wei-
pa, northern Australia. Australian Journal of Ecology, 16,
111–118.
Bowman, D.M.J.S. & Wilson, B.A. (1988) Fuel characteristics
of coastal monsoon forests, Northern Territory, Australia.
Journal of Biogeography, 15, 807–817.
Braithwaite, R.W. & Estbergs, J.A. (1985) Fire patterns and
woody vegetation trends in the Alligator Rivers Region of
northern Australia. Ecology and management of the world’s
savannas (ed. by J.C. Tothill and J.J. Mott), pp. 359–364.
Australian Academy of Science, Canberra.
Brooker, M.I.H. & Kleinig, D.A. (2004) Field guide to eucalypts,
2nd edn. Bloomings Books, Melbourne.
Bullock, S.H. & Solis-Magallanes, J.A. (1990) Phenology of
canopy trees of a tropical deciduous forest in Mexico. Bio-
tropica, 22, 22–35.
Clifton, P.J. (2004) Effects of Andropogon gayanus on micro-
climate and woody seedling recruitment. Unpubl. Hons.
Thesis, Charles Darwin University, Darwin, Australia.
Cowie, I.D. & Albrecht, D.E. (2005) Checklist of the vascular
plants of the Northern Territory, Australia. Parks & Wildlife
Service of the Northern Territory, Darwin.
Ewel, J.J. (1977) Differences between wet and dry successional
tropical ecosystems. Geo-Eco-Trop, 1, 103–117.
Fensham, R.J. & Bowman, D.M.J.S. (1992) Stand structure and
the influence of overwood on regeneration in tropical eu-
calypt forest on Melville Island. Australian Journal of Botany,
40, 335–342.
Fordyce, I. (1998) Ecology, ecophysiology and biogeography of
the monsoon rainforest tree Allosyncarpia ternata S.T. Blake.
Unpublished PhD Thesis, Charles Darwin University, Darwin.
Fordyce, I.R., Eamus, D., Duff, G.A. & Williams, R.J. (1997)
The role of seedling age and size in the recovery of Allo-
syncarpia ternata following fire. Australian Journal of Ecol-
ogy, 22, 262–269.
Frankie, G.W., Baker, H.G. & Opler, P.H. (1974) Comparative
phenological studies of trees in tropical wet and dry forests in
the lowlands of Costa Rica. Journal of Ecology, 62, 881–913.
Garwood, N. (1989) Tropical soil seed banks: a review. Ecology
of soil seed banks (ed. by M.A. Leck, R.L. Simpson and V.T.
Parker), pp. 140–209. Academic Press, New York.
Gerhardt, K. & Hytteborn, H. (1992) Natural dynamics and
regeneration methods in tropical dry forests – an intro-
duction. Journal of Vegetation Science, 3, 361–364.
J. Russell-Smith and S. A. Setterfield
1612 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Gill, A.M. (1997) Eucalypts and fires: interdependent or
independent? In Eucalypt ecology: individuals to ecosystems
(ed. by J.E. Williams and J.C.Z. Woinarski), pp. 151–167.
Cambridge University Press, Cambridge.
Higgins, S.I., Bond, W.J. & Trollope, W.S.W. (2000) Fire,
resprouting and variability: a recipe for grass-tree coexistence
in savanna. Journal of Ecology, 88, 213–229.
Hoffmann, W.A. (1996) The effects of fire and cover on
seedling establishment in a neotropical savanna. Journal of
Ecology, 84, 383–393.
Hoffmann, W.A. (1998) Post-burn reproduction of woody
plants in a neotropical savanna: the relative importance of
sexual and vegetative reproduction. Journal of Applied
Ecology, 35, 422–433.
Holdridge, L.R. (1967) Life zone ecology. Tropical Science
Center, San Jose, Costa Rica.
Hopkins, M.S. & Graham, A.W. (1989) Community pheno-
logical patterns of a lowland tropical rainforest in north-
eastern Australia. Australian Journal of Ecology, 14, 399–413.
Kellman, M. (1984) Synergistic relationships between fire and
low soil fertility in Neotropical savannas: a hypothesis. Bi-
otropica, 16, 158–160.
Kinnaird, M.F. (1992) Phenology of flowering and fruiting of
an East African riverine forest ecosystem. Biotropica, 24,
187–194.
Lacey, C.J. (1974) Rhizomes in tropical eucalypts and their role
in recovery from fire damage. Australian Journal of Botany,
22, 29–38.
Lacey, C.J. & Whelan, P.I. (1976) Observations on the ecolo-
gical significance of vegetative reproduction in the Kather-
ine-Darwin region of the Northern Territory. Australian
Forestry, 39, 131–139.
Liddle, D.T., Russell-Smith, J., Brock, J., Leach, G.J. & Con-
nors, G.T. (1994) Atlas of vascular rain forest plants in the
Northern Territory, Australia. Flora of Australia, Supple-
mentary Series No. 3. Australian Government Publishing
Service, Canberra.
Lieberman, D. & Li, M. (1992) Seedling recruitment patterns
in a tropical dry forest in Ghana. Journal of Vegetation Sci-
ence, 3, 375–382.
Murphy, P.G. & Lugo, A.E. (1986) Ecology of tropical dry
forest. Annual Review of Ecology and Systematics, 17, 67–88.
Palmer, C., Price, O. & Bach, C. (2000) Foraging ecology of the
black flying fox (Pteropus alecto) in the seasonal tropics of
the Northern Territory, Australia. Wildlife Research, 27,
169–178.
Peterson, C.J. & Jones, R.H. (1997) Clonality in woody plants:
a review and comparison with clonal herbs. The ecology and
evolution of clonal plants (ed. by H. de Kroon and J. van
Groenendael), pp. 263–289. Backhuys Publishers, Leiden.
Price, O.F., Woinarski, J.C.Z. & Robinson, D. (1999) Very
large area requirements for frugivorous birds in monsoon
rainforests of the Northern Territory, Australia. Biological
Conservation, 91, 169–180.
Prior, L.D. (1997) Ecological physiology of Eucalyptus tet-
rodonta F. Muell and Terminalia ferdinandiana Excell sap-
lings in the Northern Territory. Unpublished PhD thesis,
Northern Territory University, Darwin.
Prior, L.D., Eamus, D. & Bowman, D.M.J.S. (2004) Tree
growth patterns in north Australian savanna habitats: sea-
sonal patterns and correlations with leaf attributes. Aus-
tralian Journal of Botany, 52, 303–314.
Russell-Smith, J. (1986) The forest in motion: exploratory
studies in western Arnhem Land, monsoonal northern
Australia. Unpublished PhD thesis, Australian National
University, Canberra.
Russell-Smith, J. (1991) Classification, species richness, and
environmental relations of monsoon rain forest in northern
Australia. Journal of Vegetation Science, 2, 259–278.
Russell-Smith, J. (1996) Regeneration of monsoon rainforest in
northern Australia: the sapling bank. Journal of Vegetation
Science, 7, 889–900.
Russell-Smith, J. & Lee, A.H. (1992) Plant populations of
monsoon rain forest in the Northern Territory, Australia.
Biotropica, 24, 471–487.
Russell-Smith, J. & Lucas, D.E. (1994) Regeneration of mon-
soon rain forest in northern Australia: the dormant seed
bank. Journal of Vegetation Science, 5, 161–168.
Russell-Smith, J. & Stanton, P. (2002) Fire regimes and fire
management of rainforest communities across northern
Australia. Flammable Australia: the fire regimes and biodi-
versity of a continent (ed. by R.A. Bradstock, J.E. Williams
and A.M. Gill), pp. 330–350. Cambridge University Press,
Cambridge, UK.
Russell-Smith, J., Ryan, P.G., Klessa, D., Waight, G. & Harwood,
R. (1998) Fire regimes, fire-sensitive vegetation, and fire
management of the sandstone Arnhem Plateau, monsoonal
northern Australia. Journal of Applied Ecology, 36, 829–846.
Russell-Smith, J., Yates, C.P., Edwards, A., Allan, G.E., Cook,
G.D., Cooke, P., Craig, R., Heath, B. & Smith, R. (2003a)
Contemporary fire regimes of northern Australia, 1997–
2001: change since Aboriginal occupancy, challenges for
sustainable management. International Journal of Wildland
Fire, 12, 283–297.
Russell-Smith, J., Whitehead, P.J., Cook, G.D. & Hoare, J.L.
(2003b) Response of Eucalyptus-dominated savanna to fre-
quent fires: lessons from Munmarlary, 1973–1996. Ecological
Monographs, 73, 349–375.
Setterfield, S.A. (2002) Seedling establishment in an Australian
tropical savanna: effects of seed supply, soil disturbance and
fire. Journal of Applied Ecology, 39, 949–959.
Setterfield, S.A. & Williams, R.J. (1996) Patterns of flowering
and seed production in Eucalyptus miniata and E. tetrodonta
in a tropical savanna woodland, northern Australia. Aus-
tralian Journal of Botany, 44, 107–122.
Shapcott, A. (1998a) The patterns of genetic diversity in Car-
pentaria acuminata (Arecaceae), and rainforest history in
northern Australia. Molecular Ecology, 7, 833–847.
Shapcott, A. (1998b) Vagile but inbred: patterns of inbreeeding
and the genetic structure within populations of the mon-
soon rain forest tree Syzygium nervosum (Myrtaceae) in
northern Australia. Journal of Tropical Ecology, 14, 595–614.
Australian monsoon rain forest dynamics
Journal of Biogeography 33, 1597–1614 1613ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd

Shapcott, A. (2000) Conservation and genetics in the frag-
mented monsoon rainforest in the Northern Territory,
Australia: a case study of three frugivore-dispersed species.
Australian Journal of Botany, 48, 397–407.
Taylor, J. & Tulloch, D. (1985) Rainfall in the wet-dry tropics:
extreme events at Darwin and similarities between years
during the period 1870–1973. Australian Journal of Ecology,
10, 281–295.
Terborgh, J. (1986) Community aspects of frugivory in tropical
forests. Frugivores and seed dispersal (ed. by A. Estrada and
T.H. Fleming), pp. 371–384. Dr W. Junk, Dordrecht.
Walter, H. (1984) Vegetation of the Earth, 3rd edn. Springer-
Verlag, Berlin.
Webb, L.J. (1959) A physiognomic classification of Australian
rainforests. Journal of Ecology, 47, 551–570.
Webb, L.J. (1978) A general classification of Australian rain-
forests. Australian Plants, 9, 349–363.
Webb, L.J. & Tracey, J.G. (1981) Australian rainforests:
patterns and change. Ecological biogeography of Australia
(ed. A. Keast), pp. 607–694. Dr Junk, The Hague.
Werner, P.A., Cowie, I.D. & Cusack, J.S. (2006) Juvenile tree
growth and demography in response to feral water buffalo in
savannas of northern Australia: an experimental field study
in Kakadu National Park. Australian Journal of Botany, 54,
283–296.
Whitmore, T.C. (1984) Tropical rain forests of the Far East, 2nd
edn. Clarendon Press, Oxford.
Williams, P.R. (2003) The timing of seed production in
eucalypt savanna of north-eastern Queensland. Queensland
Naturalist, 41, 31–37.
Williams, P.R. (2004) Post-fire survival within a year of ger-
mination by three perennial herbs and a eucalypt of north
Queensland. Queensland Naturalist, 42, 11–16.
Williams, P.R., Congdon, R.A., Grice, A.C. & Clarke, P.J.
(2003a) Effect of fire regime on plant abundance in a tro-
pical eucalypt savanna of north-eastern Australia. Austral
Ecology, 28, 327–338.
Williams, P.R., Congdon, R.A., Grice, A.C. & Clarke, P.J.
(2005) Germinable soil seed banks in a tropical savanna:
seasonal dynamics and effects of fire. Austral Ecology, 30, 79–
90.
Williams, R.J. (1997) Fire and floral phenology in a humid
tropical savanna at Kalpalga, Kakadu National Park. Bushfire
‘97: Proceedings of the Australasian Bushfire Conference (ed.
by B.J. McKaige, R.J. Williams and W.M. Waggitt), pp. 54–
59. CSIRO, Darwin.
Williams, R.J., Cook, G.D., Gill, A.M. & Moore, P.H.R. (1999)
Fire regime, fire intensity and tree survival in a tropical
savanna in northern Australia. Australian Journal of Ecology,
24, 50–59.
Williams, R.J., Griffiths, A.D. & Allan, G.E. (2002) Fire regimes
and biodiversity in the wet-dry tropical landscapes of
northern Australia. Flammable Australia: the fire regimes and
biodiversity of a continent (ed. by R.A. Bradstock, J.E. Wil-
liams and A.M. Gill), pp. 281–304. Cambridge University
Press, Cambridge, UK.
Williams, R.J., Muller, W.J., Wahren, C.-H., Setterfield, S.A. &
Cusack, J. (2003b) Vegetation. Fire in tropical savannas: the
Kapalga experiment (ed. by A.N. Andersen, G.D. Cook and
R.J. Williams), pp. 79–106. Springer-Verlag, New York.
BIOSKETCHES
Samantha Setterfield is a plant ecologist who has been
researching plant reproductive ecology in tropical savannas for
15 years. Her current research projects include studying the
impacts of introduced grasses on Australian savannas, and the
effect of fire on native savanna grass and trees.
Jeremy Russell-Smith is a consultant ecologist with over
20 years experience in northern Australia. He coordinates fire
research programs of the Bushfires Council of the Northern
Territory and the Tropical Savannas Cooperative Research
Centre and, as well, is involved with other natural resource
management projects in Southeast Asia. He has an abiding
interest in the ecology, biogeography and management of
monsoon rain forests.
Editor: Pauline Ladiges
J. Russell-Smith and S. A. Setterfield
1614 Journal of Biogeography 33, 1597–1614ª 2006 The Authors. Journal compilation ª 2006 Blackwell Publishing Ltd




















