
Microfossils from the Neoproterozoic Gangolihat Formation, Kumaun Lesser Himalaya: Their...
13
Microfossils from the Neoproterozoic Gangolihat Formation, Kumaun Lesser Himalaya: Their stratigraphic and evolutionary significance Meera Tiwari * , Indu Pant Wadia Institute of Himalayan Geology, 33, General Mahadev Singh Road, Dehradun, India article info Article history: Received 31 July 2007 Received in revised form 1 December 2008 Accepted 7 January 2009 Keywords: Neoproterozoic Gangolihat Dolomite Microbiota Lesser Himalaya India abstract A well-preserved microbial assemblage from the Gangolihat Dolomite, Kumaun Lesser Himalaya is described in the present paper. The present assemblage contains both filamentous (septate and non-sep- tate) and coccoidal microfossils besides acritarchs and sponge spicules. The assemblage includes 23 spe- cies belonging to 12 genera. Siphonophycus typicum is the most common species in the entire assemblage. The assemblage shows an abundance of filaments over coccoids and acritarchs. The assemblage includes Siphonophycus septatum, Siphonophycus. typicum, Siphonophycus robustum, Siphonophycus capitaneum, Siphonophycus kestron and Siphonophycus punctatum, Gunflintia minuta, Gunflintia grandis, Gunflintia barg- hoornii, Oscillatoriopsis media, Oscillatoriopsis obtusa, Oscillatoriopsis cuboides, Oscillatoriopsis sp., Nostoco- morpha, Chlorogloeaopsis contexta, Obruchevella parvissima, Sphaerophycus parvum, Myxococcoides minor, Leiosphaeridia crassa, Trachyhystrichosphaera vidalii, Trachyhystrichosphaera sp., Micrhystridium pallidum, Cymatiosphaera minuta and sponge spicules of monaxons and hexactinellid affinity. The assemblage sug- gests a Neoproterozoic age for Gangolihat Dolomite. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction In recent years, worldwide records of microscopic fossils of 2500–542 Ma has extremely enhanced our perception about early evolution of complex life. The discovery of the microbial life is doc- umented to a large extent by the assemblage preserved both in chert-phosphorite and shale and facies. A detailed study of shale and chert-phosphorite facies demonstrates that macroscopic fos- sils ranging from millimeter to decimeter are mostly preserved as carbonaceous compressions, while microscopic fossils ranging from micrometer to submillimeter are preserved as permineralized in chert and phosphate (Steiner, 1994; Zhang et al., 1998; Xiao et al., 2002). The chert biota is represented mostly as autochtho- nous, while the shale biota may be allochthonous too. These two preservational modes show altogether different sedimentary envi- ronments. The permineralized forms represent peritidal facies, whereas compressed forms represent shallow subtidal facies (Buick and Knoll, 1999). These different lithologies and mode of preservation are represented by cyanobacteria, multicellular eukaryotes such as algae and acritarchs and metazoans. Cyanobacteria, mostly represented by filaments and coccoids are the largest and most morphologically diverse group of photo- synthetic prokaryotes (Whitton, 1992). These are one of the main primary producers in past and modern marine environment and were also ecologically flexible, which facilitated tolerance to changing environments. Acritarcha (Evitt, 1963), known as a group of decay-resistant organic-walled microfossils, are among the first eukaryotes preserved in the fossil record of Proterozoic and Cam- brian (Zhang, 1986; Yan, 1991). These are variously interpreted as unicellular photosynthetic protists and some may also represent multicellular algae, fungi or even metazoans (Mendelson, 1987; Butterfield, 2004; Butterfield, 2005; Yin et al., 2007). Simple sphaeromorphs and morphologically complex acanthomorphs are the most common Proterozoic and Cambrian acritarchs. Their diversity was low in the Paleoproterozoic to Early Neoproterozoic and well-documented acritarch occurrences are confined to Neo- proterozoic (900–542 Ma) and younger age (Knoll, 1994; Schopf, 1992). In Neoproterozoic, their diversity increases during Late Riphean and Early Ediacaran followed by a decrease in diversity in Middle to Late Ediacaran and a subsequent increase throughout Cambrian (Knoll, 1984). In Cambrian, acritarch taxonomic diversity increased in step with animal radiation (Knoll, 1994; Vidal and Moczydlowska-Vidal, 1997). In India, Lesser Himalayan sequences show a clear change of diversity of acanthomorphic acritarchs from Neoproterozoic to Cambrian (Tiwari and Knoll, 1994; Tiwari, 1999; Tiwari and Pant, 2004). A variety of other eukaryotic phytoplank- tons also evolved during Proterozoic, which played a decisive role in forming their own environment. Recently, relicts of early multicellular life such as sponge spic- ules are found trapped within Proterozoic sediments which were earlier seen as a province of cyanobacteria only. The earliest 1367-9120/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.jseaes.2009.01.008 * Corresponding author. Address: 86 Janki Sadan, Vasant Vihar, Haldwani, India. Tel.: +91 0135 2525269; fax: +91 0135 2625212. E-mail addresses: [email protected], [email protected] (M. Tiwari). Journal of Asian Earth Sciences 35 (2009) 137–149 Contents lists available at ScienceDirect Journal of Asian Earth Sciences journal homepage: www.elsevier.com/locate/jaes
-
Upload
kunainital -
Category
Documents
-
view
0 -
download
0
Transcript of Microfossils from the Neoproterozoic Gangolihat Formation, Kumaun Lesser Himalaya: Their...

Journal of Asian Earth Sciences 35 (2009) 137–149
Contents lists available at ScienceDirect
Journal of Asian Earth Sciences
journal homepage: www.elsevier .com/ locate/ jaes
Microfossils from the Neoproterozoic Gangolihat Formation, Kumaun LesserHimalaya: Their stratigraphic and evolutionary significance
Meera Tiwari *, Indu PantWadia Institute of Himalayan Geology, 33, General Mahadev Singh Road, Dehradun, India
a r t i c l e i n f o a b s t r a c t
Article history:Received 31 July 2007Received in revised form 1 December 2008Accepted 7 January 2009
Keywords:NeoproterozoicGangolihat DolomiteMicrobiotaLesser HimalayaIndia
1367-9120/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.jseaes.2009.01.008
* Corresponding author. Address: 86 Janki Sadan, VTel.: +91 0135 2525269; fax: +91 0135 2625212.
E-mail addresses: meeratiwari2@rediffmail.(M. Tiwari).
A well-preserved microbial assemblage from the Gangolihat Dolomite, Kumaun Lesser Himalaya isdescribed in the present paper. The present assemblage contains both filamentous (septate and non-sep-tate) and coccoidal microfossils besides acritarchs and sponge spicules. The assemblage includes 23 spe-cies belonging to 12 genera. Siphonophycus typicum is the most common species in the entire assemblage.The assemblage shows an abundance of filaments over coccoids and acritarchs. The assemblage includesSiphonophycus septatum, Siphonophycus. typicum, Siphonophycus robustum, Siphonophycus capitaneum,Siphonophycus kestron and Siphonophycus punctatum, Gunflintia minuta, Gunflintia grandis, Gunflintia barg-hoornii, Oscillatoriopsis media, Oscillatoriopsis obtusa, Oscillatoriopsis cuboides, Oscillatoriopsis sp., Nostoco-morpha, Chlorogloeaopsis contexta, Obruchevella parvissima, Sphaerophycus parvum, Myxococcoides minor,Leiosphaeridia crassa, Trachyhystrichosphaera vidalii, Trachyhystrichosphaera sp., Micrhystridium pallidum,Cymatiosphaera minuta and sponge spicules of monaxons and hexactinellid affinity. The assemblage sug-gests a Neoproterozoic age for Gangolihat Dolomite.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
In recent years, worldwide records of microscopic fossils of2500–542 Ma has extremely enhanced our perception about earlyevolution of complex life. The discovery of the microbial life is doc-umented to a large extent by the assemblage preserved both inchert-phosphorite and shale and facies. A detailed study of shaleand chert-phosphorite facies demonstrates that macroscopic fos-sils ranging from millimeter to decimeter are mostly preservedas carbonaceous compressions, while microscopic fossils rangingfrom micrometer to submillimeter are preserved as permineralizedin chert and phosphate (Steiner, 1994; Zhang et al., 1998; Xiaoet al., 2002). The chert biota is represented mostly as autochtho-nous, while the shale biota may be allochthonous too. These twopreservational modes show altogether different sedimentary envi-ronments. The permineralized forms represent peritidal facies,whereas compressed forms represent shallow subtidal facies(Buick and Knoll, 1999). These different lithologies and mode ofpreservation are represented by cyanobacteria, multicellulareukaryotes such as algae and acritarchs and metazoans.
Cyanobacteria, mostly represented by filaments and coccoidsare the largest and most morphologically diverse group of photo-synthetic prokaryotes (Whitton, 1992). These are one of the main
ll rights reserved.
asant Vihar, Haldwani, India.
com, [email protected]
primary producers in past and modern marine environment andwere also ecologically flexible, which facilitated tolerance tochanging environments. Acritarcha (Evitt, 1963), known as a groupof decay-resistant organic-walled microfossils, are among the firsteukaryotes preserved in the fossil record of Proterozoic and Cam-brian (Zhang, 1986; Yan, 1991). These are variously interpretedas unicellular photosynthetic protists and some may also representmulticellular algae, fungi or even metazoans (Mendelson, 1987;Butterfield, 2004; Butterfield, 2005; Yin et al., 2007). Simplesphaeromorphs and morphologically complex acanthomorphs arethe most common Proterozoic and Cambrian acritarchs. Theirdiversity was low in the Paleoproterozoic to Early Neoproterozoicand well-documented acritarch occurrences are confined to Neo-proterozoic (900–542 Ma) and younger age (Knoll, 1994; Schopf,1992). In Neoproterozoic, their diversity increases during LateRiphean and Early Ediacaran followed by a decrease in diversityin Middle to Late Ediacaran and a subsequent increase throughoutCambrian (Knoll, 1984). In Cambrian, acritarch taxonomic diversityincreased in step with animal radiation (Knoll, 1994; Vidal andMoczydlowska-Vidal, 1997). In India, Lesser Himalayan sequencesshow a clear change of diversity of acanthomorphic acritarchs fromNeoproterozoic to Cambrian (Tiwari and Knoll, 1994; Tiwari, 1999;Tiwari and Pant, 2004). A variety of other eukaryotic phytoplank-tons also evolved during Proterozoic, which played a decisive rolein forming their own environment.
Recently, relicts of early multicellular life such as sponge spic-ules are found trapped within Proterozoic sediments which wereearlier seen as a province of cyanobacteria only. The earliest

138 M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149
sponges, which are one of the few animal groups involved in a bio-mineralization process, deposited hydrated silica in isolated skele-tal elements called spicules. These are preserved as either fusedspicules forming rigid skeleton or scattered spicules formingnon-rigid skeleton within the Mesohyl. The fossil record of non-ri-gid skeleton is meager because scattered spicules drop out of thedecomposing soft tissues after death of the animal and spreadwithin the sediments as isolated spicules. The spicular morphol-ogy, spicule size and their arrangement are the essential criterionin defining species and at times provide easy clues to differentiaterelated species. McCaffrey et al. (1994) reported the presence ofstem-group sponges in Neoproterozoic rocks, however, the fossilrecords of crown group sponges occur much later in the geologichistory. Earliest sponges of Early Ediacaran age (NeoproterozoicIII) have also been documented from phosphorite of Doushantuo,South China (Li et al., 1998), of Late Ediacaran age (NeoproterozoicIII) from the lowest skeletal fossil horizon in Mongolia (Brasieret al., 1997), and from the Neoproterozoic Ediacara fauna of southAustralia (Gehling and Rigby, 1996). In India also, Sponge spiculesof Early Ediacaran age and Neoproterozoic–Cambrian boundaryage are reported (Tiwari Meera, 1997, 1999; Tiwari et al., 2000).
In the Lesser Himalayan succession of India, the Krol–Tal se-quence, Deoban Formation, Vaishnodevi Limestone and Gangolihat
Fig. 1. Geological map of the a
Dolomite have proved to be significant as they have revealed well-preserved prokaryotic, eukaryotic and multicellular life forms (Sri-vastava and Kumar, 1997; Venkatachala and Kumar, 1998; Tiwariand Pant, 2004). These formations have created enormous curiosityamong paleobiologists for exploring the remains of multicellularlife in the Proterozoic carbonates. In the present paper, well-pre-served organic-walled microfossils, are reported from GangolihatDolomite of Kumaun Lesser Himalaya.
2. Geological setting
The Lesser Himalaya comprises of two major belts. The innerLesser Himalayan Belt including Jammu, Shali, Tejam and Buxa alsoextending into western Nepal and the outer Lesser Himalayan belt,the Krol Belt, extending from Solan to Nainital. The vast argillo-cal-careous succession exposed in the Inner Lesser Himalaya has beendescribed as Calc Zone of Badolisera and Calc Zone of Pithoragarh(Heim and Gansser, 1939). The succession comprises in the ascend-ing order the Rautgara–Gangolihat–Sor–Thalkedar Formations(Pant, 1985). It is overlain by a succession of quartzite and chloriteschist of Berinag Formation (Fig. 1). The Rautgara Formation con-sists of the oldest rocks, comprising quartzarenite, sublithareniteand slates (Fig. 9). This formation grades upward into Gangolihat
rea (after Valdiya, 1980).

Fig. 2. Stratigraphic column of the Jhiroli magnesite section showing position offossiliferous samples.
Table 1Generalized stratigraphic succession of the area based on Valdiya and Pant (1981).
Berinag formation Predominantly quartzarenite, locally conglomeratic with interbedded metavolcanics Locally mylonitizedschistose and sericitic quartzite
Berinag thrustMandhali formation Thalkedar
limestoneCherty dolomitic limestone with calcareous grey phyllites and sporadic stromatolites
Sor slate Olive green, brownish, grey and black slate with argillaceous and dolomitic limestoneGangolihat dolomite (Deobanformation)
Stromatolitic dolomite and lenticles of magnesite and TalcThinly bedded greyish limestone and marl
Rautgara formation Brown, dark, purple and olive green slate, quartzarenite, litharenite and sublithareniteBase not seen
M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149 139
Dolomite Formation. Valdiya (1980) has divided the carbonaterocks of the eastern Kumaun lesser Himalaya into two formations,the older Deoban Formation (= Gangolihat Dolomite) and youngerMandhali Formation containing Sor slate and Thalkedar LimestoneMembers. Gangolihat Dolomite is characterized by the thick dolo-mite, stromatolitic dolomites and magnesite developed around thetownship of Gangolihat in Pithoragarh district of Uttarakhandstate. In Bageshwar – Kapkot area, it is designated as Kapkot For-mation (Bhattacharya, 1974). The Sor slates consist of light greenand grey green slates, marls and thin limestone bands and theoverlying Thalkedar Limestone comprises of blue-grey bandedlimestone often with chert laminae, nodules and argillaceous lime-stone, alternating with calcareous grey phyllites. The calcareoussequence is overlain by predominantly arenaceous succession(quartzite and chlorite schist) of Berinag Formation. There is a pro-fuse development of stromatolites in the sequence. The general-ized stratigraphic succession based on Valdiya and Pant (1981) isgiven in Table 1.
3. Palaeobiological investigations
3.1. Materials and method
The main lithologies represented in Gangolihat Dolomite arecherty dolomite, stromatolitic dolomite, limestone, phyllite andshales. Microfossils were observed in thin sections only and arestudied under Olympus VANOX T microscope; some selected thinsections were also scanned under SEM. Microfossils present inchert are three dimensionally preserved and show more morpho-logical features for identification, whereas in shale diagenetic com-paction has flattened the fossils causing differences inidentification. All possible care was taken to identify the microfos-sils present in these two different facies.
3.2. Fossiliferous sections of the area
Carbonate stromatolites of the Proterozoic rarely exhibit micro-fossil remains of cyanobacteria, the dominant group of microor-ganisms, in majority of the shallow-marine stromatoliteoccurrences. Calcified cyanobacteria occur nearly continuouslyfrom the Cambrian to the Cretaceous, either forming skeletal stro-matolites, oncolites, or distinct microfossils such as Girvanella. Ger-not et al. (2001) believe that this appears enigmatic becausecyanobacterial filaments and cell remains have been preservedwhen early silicification occurred. Interestingly, cyanobacteria arefound preserved within the phosphatic part of stromatolite of Gan-golihat Dolomite especially in Jarmal and Bans sections. Distinc-tively diverse microbiota, especially in their contents ofoscillatoriacean trichomes, large unicellular eukaryotes and insome instances multicellular eukaryotic organisms, is found pre-served in chert nodules and bands within dolomite and shale ofGangolihat Dolomite. These organic-walled microfossils werestudied from samples collected from Gangolihat – Jarmalgaon sec-
tion (Fig. 3), Bageshwar – Kapkot section (Fig. 4), Pithoragarh –Chandak section (Fig. 5) and Jhiroli Magnesite section (Fig. 2).
3.2.1. Gangolihat – Jarmal road sectionIn the Gangolihat – Jarmal road section stromatolitic dolomite
and dolomite with chert bands and chert intraclasts are exposed(Fig. 3). The most conspicuous feature of this stromatolitic dolo-mite is the occurrence of collophane in the form of vein, streaksand stringers, emplaced generally along the interlaminal partingsand as non-uniform coatings on the exterior of the stromatolites.Thin section study of dolomite shows occurrence of dolomicrospa-rite with diagenetic veins of coarse quartz. Small circular and subrounded pelsparite of crystallized quartz with fine irregular trans-parent coatings are present throughout. The pellets are >0.5 mm indiameter and are cemented with sparite. Intraclasts are presentthroughout the thin section. The development of dismicrite is alsoobserved.

Fig. 3. Stratigraphic column of the section along Gangolihat –Jarmalgaon sectionshowing position of fossiliferous samples.
Fig. 4. Stratigraphic column of the section along Bageshwar –Kapkot sectionshowing position of fossiliferous samples.
140 M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149
The microbiota present within the pellets is represented byGunflintia barghoornii, Oscillatoriopsis media, Oscillatoriopsis cubo-ides, Oscillatoriopsis sp., Sphaerophycus parvum, Myxococcoides min-or, Leiosphaeridia crassa and some unidentified large sphericalbodies with characteristic outer rim. In the assemblage, G. barg-hoornii is characterized by multicellular, unbranched, curved orstraight, filaments possessing distinct septa (Fig. 6v). The diameterof the filament ranges from 20 to 21 lm and length is up to100 lm. Each septum is uniformly spaced and the cells are longerthan wide. G. barghoornii was first reported by Maithy (1975) fromthe Bushimay System, Zaire. The present form compares well withthe latter in having larger diameter range and a greater cell length.Trichomes identified as Oscillatoriopsis media are multicellular fila-ments with distinct cross wall, uniseriate, unbranched and withoutsheath (Fig. 6x, y, aa, dd). Cells are cylindrical with a diameterrange of 6.5–12.5 lm; cell length varies from 15.6 to 26.2 lm. To-tal length of the filament is �180 lm. Hormogones not observed.The present form compares very well with O. media reported fromthe Shorikha and Burovaya Formations, Siberian Platform and InfraKrol Formation (Sergeev, 2001; Mendelson and Schopf, 1982; Ti-wari and Pant, 2004) but differs from the latter in its larger celllength. Width of filaments is an important criterion for identifica-tion, whereas length may vary due to cell division. In the presentspecimens it seems that the filaments may have lost their crosswalls during diagenesis and preserved cells look longer. Therefore,the present forms are included within O. media. Filaments identi-fied as O. cuboides are multicellular, uniseriate and unbranchedand the cells of the filament are typically isodiametric, with a celldiameter of 10 lm and cell length of 10.5 lm. Total length of thefilament is �100 lm (Fig. 6u). These forms are closely comparablewith O. cuboides from the Lower Proterozoic Duck Creek DolomiteWestern Australia (Knoll et al., 1988), in its diameter range and iso-diametric nature of cells. Oscillatoriopsis sp. is characterized bymulticellular, unbranched filaments where the diameter of theindividual cell ranges from 12.9 to 13.5 lm and the cell length var-ies from 51 to 57 lm. Total length of the filament is �110 lm(Fig. 6r, s). The present forms are comparable to O. media (Mendel-son and Schopf, 1982) in its diameter range, but differ from the lat-ter in its length/width ratio, where the cells are 3–4 times longerthan wide. These forms are also comparable with Phormidella (Ven-katachala et al., 1990) in its diameter range which is 12.5–15.0 lm,but differ in cell length. Cell length of Phormidella is 25 lm, muchsmaller as compared to the present specimen, where it is 50–52 lm. Coccoidal colonies, consisting of numerous individuals,either paired or solitary are comparable to S. parvum (Fig. 8g).The paired cells are irregular, thin and smooth walled. Diameterof the cell varies from 5.2 to 10.1 lm. The size range is almostthe same as described from the Bitter Spring Formation (Schopf,1968) Infra Krol Formation and Deoban Formation of Lesser Hima-laya (Tiwari and Azmi, 1992; Shukla et al., 1987). Spheroidal cells,with a psilate surface feature, comprising few to many cells areidentified as M. minor (Fig. 8c, d, f). Cell diameter varies from 11to 37 lm. The size range of M. minor is larger than the form de-scribed by Schopf (1968) from the Bitter Spring Formation, Austra-lia. The present form compares well with the form present in theInfra Krol Formation and Semri Group (Venkatachala et al., 1990;Kumar and Srivastava, 1995). L. crassa possesses a thin vesicle wallwith a diameter range of 10–40.9 lm (Fig. 7d, e). Similar forms arereported from the Infra Krol Formation and Bylot Supergroup (Ti-wari and Pant, 2004; Hofmann and Jackson, 1994).
3.2.2. Bageshwar – Kapkot sectionThinly bedded limestone, massive bluish grey dolomite with
chert nodules and oolitic dolomite are present in Bageshwar – Kap-kot section (Fig. 4). The cherty band is �10 m thick where chertnodules range in size from 1 to 12 cm in diameter. Chert nodules

Fig. 5. Stratigraphic column of the section along Pithoragarh – Chandak section showing position of fossiliferous samples.
M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149 141
are lensoidal to ellipsoidal and are often arranged parallel to bed-ding. Chert nodules contain 5–10 lm sized pyrite crystals andblack organic matter giving black colour to these nodules. Often fil-aments and spherical bodies are found to be totally altered by pyr-ite giving bead-like appearance to them. The association of finegrained pyrite with organic matter is the most common type ofpreservation of organic structure in the algal chert.
The assemblage is characterized by Siphonophycus kestron, Gunf-lintia minuta, G. grandis, Nostocomorpha, Obruchevella parvissima,Micrhystridium pallidum and very small sponge spicules. Siphono-phycus kestron is characterized by non-septate filaments having across sectional diameter of 7–12microns (Fig. 6g). The present formis comparable to S. kestron described from the Draken Conglomer-ate Formation, Deoban Formation, Kheinjua Formation and Infrak-rol Formation (Knoll et al., 1991; Shukla et al., 1987; Kumar andSrivastava, 1995; Tiwari and Pant, 2004). Straight to curved, sep-tate and unbranched Filaments with a diameter of 1.1–1.9 lm,and a maximum length of �57 lm are identified as Gunflintia min-uta (Fig. 8r, s). Multicellular, unbranched fragments identified asGunflintia grandis are septate filaments, generally solitary andstraight (Fig. 8i, j, l, n, q). Diameter of the filaments ranges from2.1 to 4 lm and length ranges from 60 to 70 lm. These forms arecomparable with G. grandis from the Gunflint Iron Formation(Barghoorn and Tyler, 1965) and Deoban Formation (Kumar andSrivastava, 1992). Filaments identified as Nostocomorpha are unise-riate, unbranched, curved or straight in nature (Fig. 6t; 8. m). In thepresent assemblage these forms are preserved as chain like aggre-gates of spheroidal grains of iron minerals. Diameter of the fila-ment ranges from 3 to 8 lm and the maximum length of thefilament is �290 lm. These forms are comparable with forms fromthe Baffin Island Canada and Wernecke Mountains (Hofmann andJackson, 1994; Hofmann, 1984). Obruchevella parvissima is charac-terized by spirally coiled tubular filament with a coil diameter of8–12 lm and filament diameter of 2.5–3.5 lm (Fig. 8t, u). The
present specimen is comparable with O. parvissima from Meishu-cun Formation (Song, 1984) in its tube diameter, but differs in hav-ing smaller coil diameter. Micrhystridium pallidum is characterizedby small sub-circular vesicle of 8–10 micron diameter range, cov-ered with small 1–5 microm spines (Fig. 7j, k). Isolated hexactinel-lid and some polyactin spicules with a length of �10 lm anddiameter .5–1 lm are also preserved in the chert nodules. Raysof these spicules are needle like and are mostly transparent butsometimes black in colour (Fig. 8v, w, x). Such smaller forms arerare in the fossil records.
3.2.3. Pithoragarh – Chandak sectionIn Chandak area, Gangolihat Dolomite is further divided into
four members – the Chhera, Hiunpani, Chandak and Dhari, inascending order (Valdiya, 1980). Chhera member is mainly charac-terized by purple, violet and maroon slates and phyllites, interbed-ded with pink, white and light green marble (Chhera-Bans section).It is the lower most unit overlying Rautgara slates and quartzites. Itis in turn overlain by Hiunpani Member, which is characterized bywhite and light pink fine grained cherty dolomite alternating withchert lamina and intercalated with talcose phyllite (Hiunpani-Dha-ri section). Hiunpani Member is overlain by Chandak Memberwhich consist of detrital dolomite and dolomitic limestone show-ing development of bioherms of branching-columnar stromatoliteswhich are often phosphatic. In this area, lentiform magnesite is themost important unit. Dhari Member is the top most unit and ischaracterized by blue-grey limestone interbedded with calcareousshale or marl, slightly phosphatic. This unit is overlain by olivegreen, brown and grey Sor slate (Fig. 5). Sponge spicules are discov-ered from shale and pink limestone of the Chhera member exposedin the road section near village Chhera on Chandak-Chhera road(Tiwari et al., 2000).
Characteristically simple and small spicules, bearing six straightrays are present in this assemblage. Two size ranges of the spicules
142 M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149
are present; most of them are 8–10 lm in length with a diameterof �.5 lm and the others are 100–150 lm in length and �5 lm indiameter (Fig. 7l–o). Though spicular framework could not be builtdue to their scattered nature, the size range may be represented asdifferentiation of mega- and microscleres at an early stage ofsponge evolution. These spicules under SEM show clear hexactinel-lid morphology. According to Hooper (2000), the hexactinellidsponges consist of six-rayed siliceous spicules and axial canalwhich is always square in cross section.
3.2.4. Jhiroli Magnesite sectionJhiroli magnesite mine is situated �45 km from Almora, in the
eastern part of Kumaun Lesser Himalaya (Fig. 1). Fossiliferous sam-ples were collected from pit 1 of magnesite at Jhiroli (79�45007.5600:
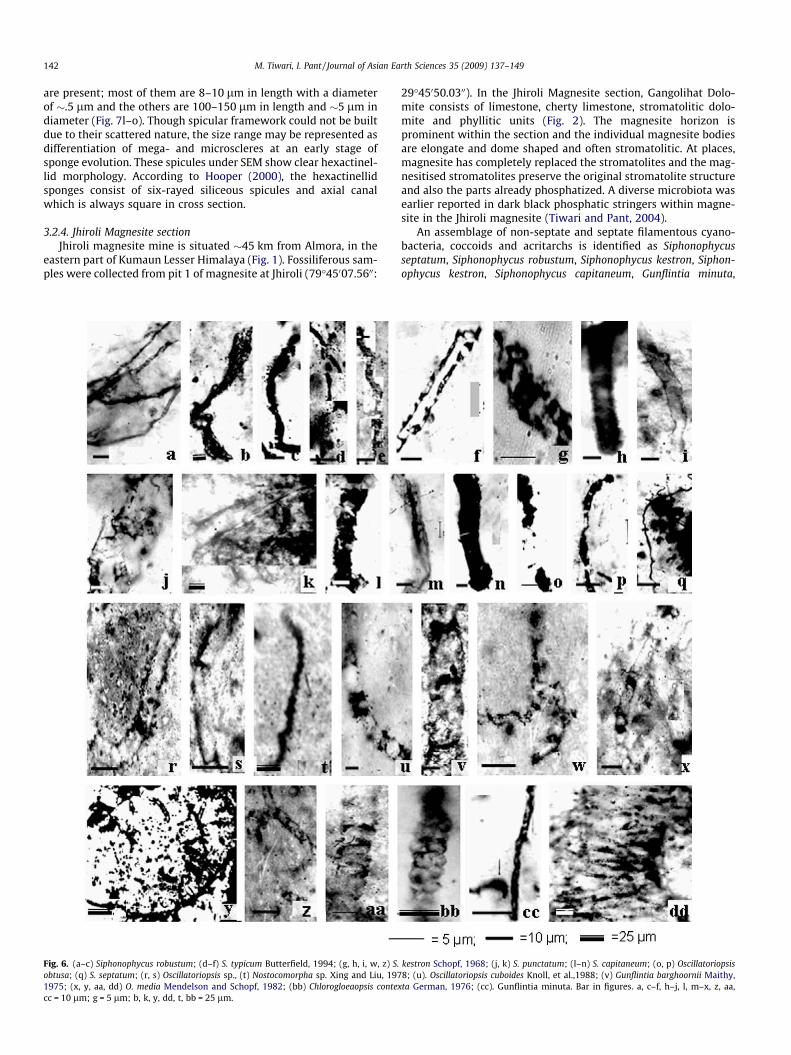
Fig. 6. (a–c) Siphonophycus robustum; (d–f) S. typicum Butterfield, 1994; (g, h, i, w, z) S.obtusa; (q) S. septatum; (r, s) Oscillatoriopsis sp., (t) Nostocomorpha sp. Xing and Liu, 1971975; (x, y, aa, dd) O. media Mendelson and Schopf, 1982; (bb) Chlorogloeaopsis contexcc = 10 lm; g = 5 lm; b, k, y, dd, t, bb = 25 lm.
29�45050.0300). In the Jhiroli Magnesite section, Gangolihat Dolo-mite consists of limestone, cherty limestone, stromatolitic dolo-mite and phyllitic units (Fig. 2). The magnesite horizon isprominent within the section and the individual magnesite bodiesare elongate and dome shaped and often stromatolitic. At places,magnesite has completely replaced the stromatolites and the mag-nesitised stromatolites preserve the original stromatolite structureand also the parts already phosphatized. A diverse microbiota wasearlier reported in dark black phosphatic stringers within magne-site in the Jhiroli magnesite (Tiwari and Pant, 2004).
An assemblage of non-septate and septate filamentous cyano-bacteria, coccoids and acritarchs is identified as Siphonophycusseptatum, Siphonophycus robustum, Siphonophycus kestron, Siphon-ophycus kestron, Siphonophycus capitaneum, Gunflintia minuta,
kestron Schopf, 1968; (j, k) S. punctatum; (l–n) S. capitaneum; (o, p) Oscillatoriopsis8; (u). Oscillatoriopsis cuboides Knoll, et al.,1988; (v) Gunflintia barghoornii Maithy,ta German, 1976; (cc). Gunflintia minuta. Bar in figures. a, c–f, h–j, l, m–x, z, aa,
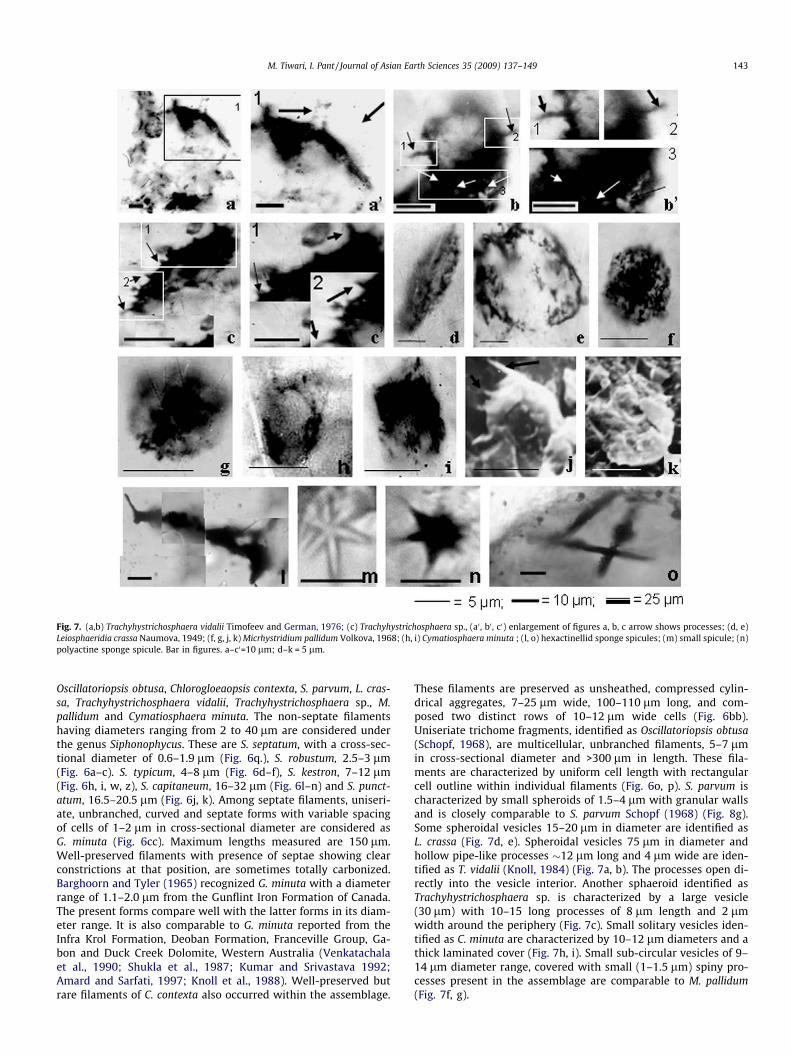
Fig. 7. (a,b) Trachyhystrichosphaera vidalii Timofeev and German, 1976; (c) Trachyhystrichosphaera sp., (a0 , b0 , c0) enlargement of figures a, b, c arrow shows processes; (d, e)Leiosphaeridia crassa Naumova, 1949; (f, g, j, k) Micrhystridium pallidum Volkova, 1968; (h, i) Cymatiosphaera minuta ; (l, o) hexactinellid sponge spicules; (m) small spicule; (n)polyactine sponge spicule. Bar in figures. a–c0=10 lm; d–k = 5 lm.
M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149 143
Oscillatoriopsis obtusa, Chlorogloeaopsis contexta, S. parvum, L. cras-sa, Trachyhystrichosphaera vidalii, Trachyhystrichosphaera sp., M.pallidum and Cymatiosphaera minuta. The non-septate filamentshaving diameters ranging from 2 to 40 lm are considered underthe genus Siphonophycus. These are S. septatum, with a cross-sec-tional diameter of 0.6–1.9 lm (Fig. 6q.), S. robustum, 2.5–3 lm(Fig. 6a–c). S. typicum, 4–8 lm (Fig. 6d–f), S. kestron, 7–12 lm(Fig. 6h, i, w, z), S. capitaneum, 16–32 lm (Fig. 6l–n) and S. punct-atum, 16.5–20.5 lm (Fig. 6j, k). Among septate filaments, uniseri-ate, unbranched, curved and septate forms with variable spacingof cells of 1–2 lm in cross-sectional diameter are considered asG. minuta (Fig. 6cc). Maximum lengths measured are 150 lm.Well-preserved filaments with presence of septae showing clearconstrictions at that position, are sometimes totally carbonized.Barghoorn and Tyler (1965) recognized G. minuta with a diameterrange of 1.1–2.0 lm from the Gunflint Iron Formation of Canada.The present forms compare well with the latter forms in its diam-eter range. It is also comparable to G. minuta reported from theInfra Krol Formation, Deoban Formation, Franceville Group, Ga-bon and Duck Creek Dolomite, Western Australia (Venkatachalaet al., 1990; Shukla et al., 1987; Kumar and Srivastava 1992;Amard and Sarfati, 1997; Knoll et al., 1988). Well-preserved butrare filaments of C. contexta also occurred within the assemblage.
These filaments are preserved as unsheathed, compressed cylin-drical aggregates, 7–25 lm wide, 100–110 lm long, and com-posed two distinct rows of 10–12 lm wide cells (Fig. 6bb).Uniseriate trichome fragments, identified as Oscillatoriopsis obtusa(Schopf, 1968), are multicellular, unbranched filaments, 5–7 lmin cross-sectional diameter and >300 lm in length. These fila-ments are characterized by uniform cell length with rectangularcell outline within individual filaments (Fig. 6o, p). S. parvum ischaracterized by small spheroids of 1.5–4 lm with granular wallsand is closely comparable to S. parvum Schopf (1968) (Fig. 8g).Some spheroidal vesicles 15–20 lm in diameter are identified asL. crassa (Fig. 7d, e). Spheroidal vesicles 75 lm in diameter andhollow pipe-like processes �12 lm long and 4 lm wide are iden-tified as T. vidalii (Knoll, 1984) (Fig. 7a, b). The processes open di-rectly into the vesicle interior. Another sphaeroid identified asTrachyhystrichosphaera sp. is characterized by a large vesicle(30 lm) with 10–15 long processes of 8 lm length and 2 lmwidth around the periphery (Fig. 7c). Small solitary vesicles iden-tified as C. minuta are characterized by 10–12 lm diameters and athick laminated cover (Fig. 7h, i). Small sub-circular vesicles of 9–14 lm diameter range, covered with small (1–1.5 lm) spiny pro-cesses present in the assemblage are comparable to M. pallidum(Fig. 7f, g).
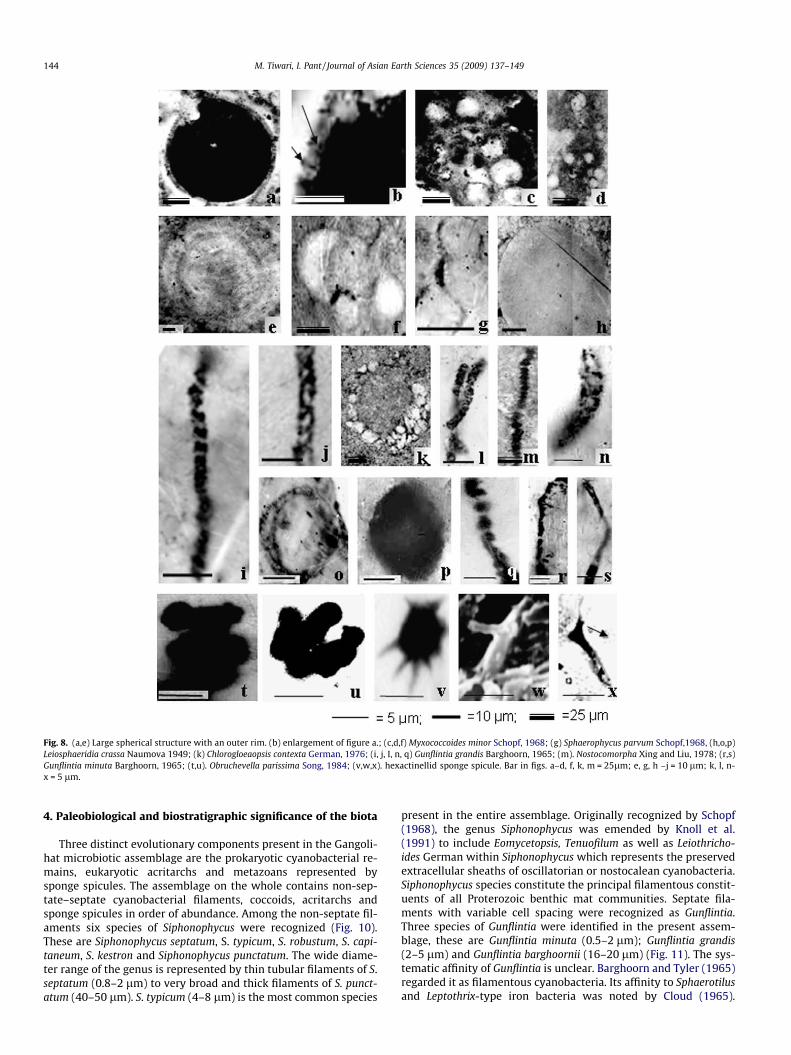
Fig. 8. (a,e) Large spherical structure with an outer rim. (b) enlargement of figure a.; (c,d,f) Myxococcoides minor Schopf, 1968; (g) Sphaerophycus parvum Schopf,1968, (h,o,p)Leiosphaeridia crassa Naumova 1949; (k) Chlorogloeaopsis contexta German, 1976; (i, j, I, n, q) Gunflintia grandis Barghoorn, 1965; (m). Nostocomorpha Xing and Liu, 1978; (r,s)Gunflintia minuta Barghoorn, 1965; (t,u). Obruchevella parissima Song, 1984; (v,w,x). hexactinellid sponge spicule. Bar in figs. a–d, f, k, m = 25lm; e, g, h –j = 10 lm; k, l, n-x = 5 lm.
144 M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149
4. Paleobiological and biostratigraphic significance of the biota
Three distinct evolutionary components present in the Gangoli-hat microbiotic assemblage are the prokaryotic cyanobacterial re-mains, eukaryotic acritarchs and metazoans represented bysponge spicules. The assemblage on the whole contains non-sep-tate–septate cyanobacterial filaments, coccoids, acritarchs andsponge spicules in order of abundance. Among the non-septate fil-aments six species of Siphonophycus were recognized (Fig. 10).These are Siphonophycus septatum, S. typicum, S. robustum, S. capi-taneum, S. kestron and Siphonophycus punctatum. The wide diame-ter range of the genus is represented by thin tubular filaments of S.septatum (0.8–2 lm) to very broad and thick filaments of S. punct-atum (40–50 lm). S. typicum (4–8 lm) is the most common species
present in the entire assemblage. Originally recognized by Schopf(1968), the genus Siphonophycus was emended by Knoll et al.(1991) to include Eomycetopsis, Tenuofilum as well as Leiothricho-ides German within Siphonophycus which represents the preservedextracellular sheaths of oscillatorian or nostocalean cyanobacteria.Siphonophycus species constitute the principal filamentous constit-uents of all Proterozoic benthic mat communities. Septate fila-ments with variable cell spacing were recognized as Gunflintia.Three species of Gunflintia were identified in the present assem-blage, these are Gunflintia minuta (0.5–2 lm); Gunflintia grandis(2–5 lm) and Gunflintia barghoornii (16–20 lm) (Fig. 11). The sys-tematic affinity of Gunflintia is unclear. Barghoorn and Tyler (1965)regarded it as filamentous cyanobacteria. Its affinity to Sphaerotilusand Leptothrix-type iron bacteria was noted by Cloud (1965).

Fig. 9. Generalized column of Gangolihat Dolomite.
M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149 145
Whereas, Golubic et al. (1995) showed differences in morphologi-cal characters between heterotrophic Sphaerotilus and chemo-trophic Leptothrix that may prove Gunflintia is the ancestor ofLeptothrix but not of Sphaerotilus. Another septate filament in theassemblage is dominated by genus Oscillatoriopsis. Four species ofOscillatoriopsis were recognized namely – Oscillatoriopsis obtusa,O. media, O. cuboides and O. sp. The genus Oscillatoriopsis waserected by Schopf (1968) to include the sheath-less trichomes ofOscillatoriacean cyanobacteria similar to modern genus Oscillatoriavoucher. Later, Mendelson and Schopf (1982) emended the genusand applied the generic name Oscillatoriopsis to all oscillatorianor Lyngbya-like trichomes, surrounded by a sheath less than1 lm thick. Subsequently, Butterfield et al. (1994) emended it toinclude all undifferentiated Oscillatoriopsis like trichomes withoutclearly developed extracellular sheaths, and differentiated specieson the basis of size range and cell shape (cf. Golubic and Focke,1978). Sergeev (2001), however, did not agree with this proposal.In his view, many morphologically distinguishable species in-cluded within this genus should be referred to other genera. Thegenus is widely distributed in Meso- to Neoproterozoic rocksworldwide. Septate filaments, characterized by Nostocomorphawith bead-like appearance, possessing a diameter of 3–8 lm, arepresent in large numbers mainly in the Utrora section. Comparablestructures are reported from various Proterozoic and Cambrianlocalities (Hofmann, 1984; Hofmann and Jackson, 1994). Charac-teristic but rare specimens of non-septate filaments, Chlorogloeaop-sis contexta are present only in the Jhiroli section. The genus wasrecognized from the Bushimay Group (1300–940 Ma) in Zaire(Maithy, 1975). German (1976) illustrated C. contextus with adiameter of 25 lm; the latter is more like C. zairensis. Hofmannand Jackson (1994), however, considered both to be conspecific
and attributed them to the genus C. contexta. Microfossils compa-rable to C. contexta are also reported from Vendian (Ediacaran) ofPodolia (German, 1985). Extraordinary small, coiled filaments rep-resented by Obruchevella parvissima are present in the assemblagealthough rarely. Obruchevella was originally proposed for tightlyand regularly coiled calcareous tubes of 6–18 lm filament diame-ter and 30–50 lm helix diameter, from Lower Cambrian carbon-ates of Siberia (Reitlinger 1948, 1959). They are generallyinterpreted as helical cyanobacteria, comparable to living Spirulinaor Phormidium species (Luchinina, 1975). O. parvissima is reportedfrom Early Cambrian Meishucun Stage of the Meishucun section ofChina and may represent a lineage that can be traced across theVendian–Cambrian boundary (Bengtson et al., 1992). Among theCoccoidal forms, S. parvum and M. minor are quite common inthe assemblage and are present in the Jhiroli and Jarmal sections.S. parvum has been reported from the Paleoproterozoic to Neopro-terozoic sequences throughout the world and Myxococcoides arethe common elements present throughout the Proterozoic.
Acritarchs in the assemblage are represented by simple spheri-cal and acanthomorphic forms identified as L. crassa, C. minuta, T.vidalii, Trachyhystrichosphaera sp. and M. pallidum. (Fig. 12). Amongthese, L. crassa shows an affinity with chlorophycean algae and isknown from upper Riphean to Lower Cambrian strata in Russiaand elsewhere (Yin and Guan 1999, and references therein). C. min-uta is observed only in the Jhiroli section and is an important partof the assemblage. Specimens of C. minuta are reported from Neo-proterozic-Cambrian boundary by Knoll and Swett (1987), Volkova(1969) and Baudet et al. (1989), but show a narrow size range.Small acanthomorphic acritarch M. pallidum is present in the Jhiroliand Utrora sections. M. pallidum was reported from the upper mostProterozoic and lower most Cambrian Formation of Central Mac-
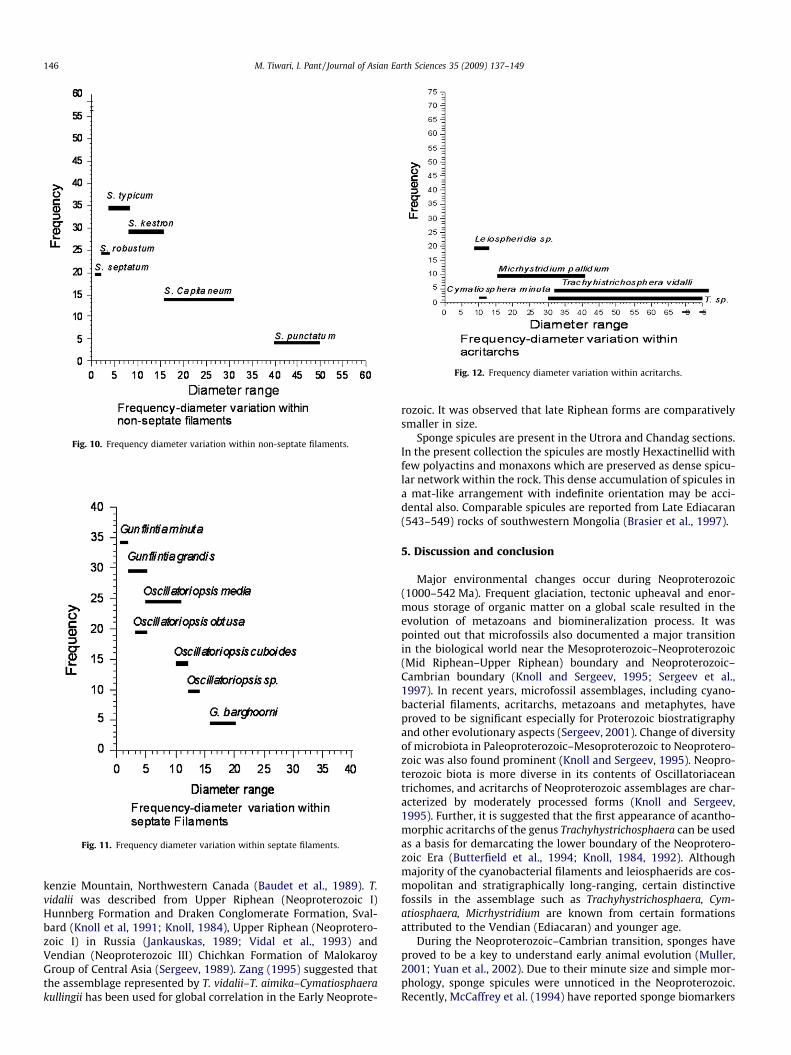
Fig. 10. Frequency diameter variation within non-septate filaments.
Fig. 11. Frequency diameter variation within septate filaments.
Fig. 12. Frequency diameter variation within acritarchs.
146 M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149
kenzie Mountain, Northwestern Canada (Baudet et al., 1989). T.vidalii was described from Upper Riphean (Neoproterozoic I)Hunnberg Formation and Draken Conglomerate Formation, Sval-bard (Knoll et al, 1991; Knoll, 1984), Upper Riphean (Neoprotero-zoic I) in Russia (Jankauskas, 1989; Vidal et al., 1993) andVendian (Neoproterozoic III) Chichkan Formation of MalokaroyGroup of Central Asia (Sergeev, 1989). Zang (1995) suggested thatthe assemblage represented by T. vidalii–T. aimika–Cymatiosphaerakullingii has been used for global correlation in the Early Neoprote-
rozoic. It was observed that late Riphean forms are comparativelysmaller in size.
Sponge spicules are present in the Utrora and Chandag sections.In the present collection the spicules are mostly Hexactinellid withfew polyactins and monaxons which are preserved as dense spicu-lar network within the rock. This dense accumulation of spicules ina mat-like arrangement with indefinite orientation may be acci-dental also. Comparable spicules are reported from Late Ediacaran(543–549) rocks of southwestern Mongolia (Brasier et al., 1997).
5. Discussion and conclusion
Major environmental changes occur during Neoproterozoic(1000–542 Ma). Frequent glaciation, tectonic upheaval and enor-mous storage of organic matter on a global scale resulted in theevolution of metazoans and biomineralization process. It waspointed out that microfossils also documented a major transitionin the biological world near the Mesoproterozoic–Neoproterozoic(Mid Riphean–Upper Riphean) boundary and Neoproterozoic–Cambrian boundary (Knoll and Sergeev, 1995; Sergeev et al.,1997). In recent years, microfossil assemblages, including cyano-bacterial filaments, acritarchs, metazoans and metaphytes, haveproved to be significant especially for Proterozoic biostratigraphyand other evolutionary aspects (Sergeev, 2001). Change of diversityof microbiota in Paleoproterozoic–Mesoproterozoic to Neoprotero-zoic was also found prominent (Knoll and Sergeev, 1995). Neopro-terozoic biota is more diverse in its contents of Oscillatoriaceantrichomes, and acritarchs of Neoproterozoic assemblages are char-acterized by moderately processed forms (Knoll and Sergeev,1995). Further, it is suggested that the first appearance of acantho-morphic acritarchs of the genus Trachyhystrichosphaera can be usedas a basis for demarcating the lower boundary of the Neoprotero-zoic Era (Butterfield et al., 1994; Knoll, 1984, 1992). Althoughmajority of the cyanobacterial filaments and leiosphaerids are cos-mopolitan and stratigraphically long-ranging, certain distinctivefossils in the assemblage such as Trachyhystrichosphaera, Cym-atiosphaera, Micrhystridium are known from certain formationsattributed to the Vendian (Ediacaran) and younger age.
During the Neoproterozoic–Cambrian transition, sponges haveproved to be a key to understand early animal evolution (Muller,2001; Yuan et al., 2002). Due to their minute size and simple mor-phology, sponge spicules were unnoticed in the Neoproterozoic.Recently, McCaffrey et al. (1994) have reported sponge biomarkers

M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149 147
in lower Neoproterozoic rocks and advocated the presence of stem-group sponges. Li et al. (1998) reported an assemblage of monaxonsponges with well preserved soft tissue from Doushantuo phos-phate deposit of south China, and considered them to be pre-Ediac-aran and Early Vendian (�580 Ma) in age. From southwesternMongolia Late Ediacaran (543–549 Ma) hexactinellid sponge spic-ules were reported (Brasier et al., 1997). These microfossils wereinterpreted variously as casts and moulds of cyanobacterial fila-ments, volcanic shards and twinned arsenopyrite crystals (Zhouet al., 1998). In the Neoproterozoic Ediacara fauna of south Austra-lia, some disc-like bodies interpreted as hexactinellid spicularimpression were documented (Gehling and Rigby, 1996) and werereinterpreted as stem-group sponge (Mehl, 1998), at the same timethe interpretation was found to be controversial (Li et al., 1998).During the Cambrian radiation, sponges evolved a variety of archi-tectural forms which are not observed in later periods. Hexactinellidsponge spicules are also reported from deposit of Mid Nemakit –Daldynian to Tommotian age (Rigby, 1987; Ding and Qian, 1988;Rozanov and Zhuravlev, 1992; Steiner et al., 1993; Brasier, 1992;Brasier et al., 1993; Qian, 1999) and Demosponges are recorded dur-ing the Atdabanian (Bengtson et al., 1990; Rozanov and Zhuravlev,1992; Zhang and Pratt, 1994; Reitner and Mehl, 1995). In India,sponge spicules of Cambrian affinity are reported from Lolab Forma-tion of northwestern Kashmir (Tiwari, 1997) and also from chert-phosphorite member of Tal Formation which is equivalent to Nem-akit – Daldynian to Early Tommotian age (Mazumdar and Banerjee,1998; Tiwari, 1999). It was observed that size range of isolatedhexactinellids present in Gangolihat Dolomite is much smaller (8–150 lm) than the reported occurrences of Cambrian sponge spic-ules. It may be due to originally smaller size of ancestral sponges.
6. Summary and conclusions
On the basis of lithological similarities and stromatolite biozo-nation (Kussiella kussiensis, Conophyton gargenicus, Conophytoncylindricus, Colonella columnaris, Plicatina antiqua, Baicalia bacalica,Minjaria uralica and Maslovia columnaris), it was established thatthe Deoban, Shali and Jammu (Great Limestone) Limestone beltsare correlatable and homotaxial with the Gangolihat Dolomiteand indicate an Early to Middle Riphean age, (Valdiya, 1969,1980; Tewari, 2003). Radiometrically, the age of the GangolihatDolomite (Deoban Formation) is poorly constrained due to incon-sistency of isotopic data, therefore no definite age could be estab-lished. Sarkar et al. (2000) obtained model Pb–Pb age of 1550–1700 Ma for galena hosted by silicified carbonate rocks presentin Deoban Formation exposed in Bageshwar, with a view that themodel ages of ore deposit are semiquantitative and a more accu-rate age of host rock need to be confirmed through more analysis.Earlier, Raha et al. (1978) had suggested 967 Ma for Jammu Lime-stone on the basis of Pb-isotopic age of galena. However, recently,an age of 839 ± 139 Ma was depicted on the basis of Re–Os iso-chron, from black shale of homotaxial Shali Formation exposed inHimachal Lesser Himalaya by Singh et al. (1999). They, however,feel that the uncertainties associated with the age are too largeto make a definite conclusion.
On the other hand, Deoban Formation has contributed signifi-cantly to the microbial record in recent years. A rich and diversifiedmicrobiota is described from Deoban Formation and the geologicalage was confined to Neoproterozoic. The presence of cyanobacte-ria, algae, acritarchs and possible nematodes suggests an EarlyVendian age for Deoban Formation (Srivastava and Kumar, 1997).Further, cyanobacterial filaments, sphaeromorphic acritarchs andvase-shaped microfossils suggest a Late Riphean-Vendian age forthe Great Limestone (Venkatachala and Kumar, 1998). Interest-ingly, on the other hand, Neoproterozoic–Cambrian age was sug-
gested on the basis of Paleobasidiospores in the eastern extensionof these carbonate rocks in Nepal (Brunel et al., 1985), and proto-conodonts from the Gangolihat Dolomite (Azmi and Paul, 2004).The discovery of distinct acritarchs and small sponge spicules sug-gests Neoproterozoic (Ediacaran) age for the Gangolihat Dolomite(Tiwari et al., 2000; Tiwari and Pant, 2004). However, the distinc-tive large acritarchs described from many formations of Early Ven-dian (Early Neoproterozoic III) age elsewhere e.g., the PertatatakaFormation of Australia (Zang and Walter, 1992), the DoushantuoFormation of China (Zhang et al., 1998), the Scotia Group of Spits-bergen (Knoll, 1992), and the Infrakrol Formation of India (Tiwariand Knoll, 1994) are missing in the Gangolihat Dolomite.
The fossil assemblage is dominated by long-ranging hypobra-dytelic cyanobacteria, the geological age of Gangolihat Dolomitemay be confined to the Terminal Neoproterozoic (Ediacaran) onthe basis of significant association of acritarchs and sponge spic-ules within the assemblage. Till date, such association of acritarchsand sponge spicules is unknown from Riphean age worldwide. Be-sides, presence of phosphorite and chert within the GangolihatDolomite thus provides an excellent source for understanding theearly evolution of animal life.
References
Amard, B., Sarfati Bertrand, J., 1997. Microfossils in 2000 Ma Old chertystromatolites of the Franceville Group, Gabon. Precambrian Research 81, 197–221.
Azmi, R.J., Paul, S.K., 2004. Dolomite of inner Kumaun lesser Himalaya: implicationon age and correlation. Current Science 86, 1653–1660.
Barghoorn, E.S., Tyler, S.A., 1965. Microorganisms from the Gunflint Chert. Science147, 563–577.
Baudet, D., Aitken, J.D., Vanguestaine, M., 1989. Palenology of uppermostProterozoic and lowermost Cambrian Formation, central MackenzieMountains, northwest Canada. Canadian Journal of Earth Science 26, 129–148.
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., Runnegar, B.N., 1990. EarlyCambrian fossils from South Australia. Memoir Association of AustralasianPalaeontology 9, 1–364.
Bhattacharya, A.R., 1974. Tectonic control of sedimentation in a part of northernKumaun Himalaya. Himalayan Geology 4, 348–360.
Brasier, M.D., 1992. Nutrient-enriched waters and the early skeletal fossil record.Geological Society London Journal 149, 621–630.
Brasier, M.D., Khomentovsky, V.V., Corfield, R.M., 1993. Stable isotopic calibration ofthe earliest skeletal fossil assemblage in eastern Siberia (Precambrian–Cambrian boundary). Terra Nova 5, 225–232.
Brasier, M.D., Green, O., Schields, G., 1997. Ediacaran sponge spicule clusters fromsouthwestern Mongolia and the origin of the Cambrian fauna. Geology 25, 303–306.
Brunel, M., Chaye, Albissin, d’, Locquin, M., 1985. The Cambrian age of magnesitefrom East Nepal as determined through the discovery of Palaeobasidiospores.Journal Geological Society of India 26, 255–260.
Buick, R., Knoll, A.K., 1999. Acritarchs and microfossils from the MesoproterozoicBangemall Group, northwestern Australia. Journal of Paleontology 73 (5), 744–764.
Butterfield, N.J., 2004. A vaucheriacean alga from the middle Neoproterozoic ofSpitsbergen: implications for the evolution of Proterozoic eukaryotes and theCambrian explosion. Paleobiology 30, 231–252.
Butterfield, N.J., 2005. Probable Proterozoic fungi. Paleobiology 31, 165–182.Butterfield, N.J., Knoll, A.H., Swett, K., 1994. Paleobiology of the Neoproterozoic
Svabergfjellet formation, Spitsbergen. Fossil and Strata 34, 1–84.Cloud, P., 1965. Significance of the Gunflint (Precambrian) microflora. Science 148,
27–35.Ding, W., Qian, Y., 1988. Late Sinian to early Cambrian small shelly fossils from
Yangjiping, Shimen, Hunan. Acta Micropalaeontologica Sinica 5, 39–55.Evitt, W.R., 1963. A discussion and proposals concerning fossil Dinoflagellates,
Hystrichospheres and Acritarchs (US). National Academy of SciencesProceedings 49, 158–164.
Gehling, J.G., Rigby, J.K., 1996. Long expected sponges from the NeoproterozoicEdiacara fauna of South Australia. Journal of Paleontology 70 (2), 185–195.
German, T.N. 1976. In: Timofeev, B.V., Hermann, T.N., Mikhailova, N.S. (Eds.).Microfossils from the Precambrian, Cambrian and Ordovician. Publ. Institute ofGeology Geochronology Academy of Science, USSR, p. 106 (in Russian).
German, T.N., 1985. In: Sokolov, B.S., Ivanovski, A.B. (Eds.), Vendian System, vol. 1.Nauka, Moscow, pp. 146–153.
Gernot et al., 2001. Photosynthesis-induced biofilm calcification and calciumconcentrations in Phanerozoic Oceans. Science 292, 1701–1704.
Golubic, S., Focke, J.P., 1978. Phormidium nendersonii Howe: identify andsignificance of a modern stromatolite building microorganism. Journal ofSedimentary Petrology 48, 751–764.

148 M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149
Golubic, S., Sergeev, V.N., Knoll, A.H., 1995. Mesoproterozoic archaellipsoides:akinites of heterocystous cyanobacteria. Lethaia 28, 285–298.
Heim, A., Gansser, A., 1939. Central Himalaya, Geological observation of the SwissExpedition in 1936. Mem. Soc. Helv. Sci. Nat. 73, 1–245.
Hofmann, H.J., 1984. Organic-walled microfossils from the latest Proterozoic andearliest Cambrian of the Wernecke Mountains, Yukon. Geological Survey ofCanada Paper 84-1B, 285–297.
Hofmann, H.J., Jackson, G.D., 1994. Shale facies microfossils from the ProterozoicBylot Supergroup, Baffin island, Canada, Paleontological Society Memoir.Journal of Paleontology 68 (Suppl.), 1–39.
Hooper, J.N.A., 2000. Sponguide: Guide to Sponge Collection and Identification. QldMuseum Australia. Available from: http://www.qmuseum.qld.gov.au/organisation/sections/SessileMarineInvertebrates/spong.pdf>.
Jankauskas, T.V., 1989. Precambrian Microfossils of the USSR. Nauka, Moscow. p.190.
Knoll, A.H., 1984. Microbiotas of the late Precambrian Hunnberg formation,Nordaustlandet, Svalbard. Journal of Paleontology 58, 131–162.
Knoll, A.H., 1992. Vendian microfossils in the metasedimentary chert of ScotiaGroup, Prins Karls Forland, Svalbard. Palaeontology 35, 751–774.
Knoll, A.H., 1994. Proterozoic and Early Cambrian protists: evidence for acceleratingevolutionary tempo. Proceeding National Academy of Science USA 41, 6743–6750.
Knoll, A.H., Sergeev, V.N., 1995. Taphonomic and evolutionary changes across theMesoproterozoic–Neoproterozoic transition News Jahrbuch Geol.Palaeontology Abh 195, 289–302.
Knoll, A.H., Swett, K., 1987. Micropaleontology across the Precambrian–Cambrianboundary in Spitsbergen. Journal of Palaeontology 61, 898–926.
Knoll, A.H., Strother, P.K., Rossi, S., 1988. Distribution and diagenesis of microfossilsfrom the lower Proterozoic Duck Creek Dolomite, Western Australia.Precambrian Research 38, 257–279.
Knoll, A.H., Swett, K., Mark, J., 1991. Palaeobiology of a Neoproterozoic tidal flat/lagoonal complex: the Draken Conglomerate Formation, Spitsbergan. Journal ofPalaeontology 65, 531–570.
Kumar, S., Srivastava, P., 1992. Middle to late Proterozoic microbiota from theDeoban Limestone, Garhwal Himalaya India. Precambrian Research 56, 291–318.
Kumar, S., Srivastava, P., 1995. Microfossils from the Kheinjua formation,Mesoproterozoic Semri group, Newari area Central India. PrecambrianResearch 74, 91–117.
Li, C.-W., Chen, J.Y., Hua, T.-E., 1998. Precambrian sponges with cellular structures.Science 279, 879–882.
Luchinina, V.A., 1975. Paleoalgologischeskaya kharakteristika kembriya Sibiriskoyplatformy (Yugo vostok) Palaeoalgological characteristic of the Cambrian of theSiberian Platform (Southeast). Nauka, Novosibirisk. p. 100.
Maithy, P.K., 1975. Microorganisms from the Bushimay System (Late Precambrian)of Kanshi, Zaire. The Palaeobotanists 22, 133–149.
Mazumdar, A., Banerjee, D.M., 1998. Siliceous sponge spicules in the Early CambrianChert-phosphorite Member of the Lower Tal Formation, Krol Belt, LesserHimalaya. Geology 26, 899–902.
McCaffrey, M.A., Moldowan, J.M., Lipton, P.A., Summons, R.E., Peters, K.E.,Jaganathan, A., Watt, D.S., 1994. Palaeoenvironmental implications of novelC30 steranes in Precambrian to Cenozoic age petroleum and bitumen.Geochimica Cosmochimica Acta 58, 529–532.
Mehl, D., 1998. Porifera and chancelloriidae from the Middle Cambrian of theGeorgina Basin, Australia. Palaeontology 41, 1153–1182.
Mendelson, C.V., 1987. Acritarchs. In: Lipps, J. (Ed.), Fossil Prokaryotes and Protists.University of Tennessee, Knoxville.
Mendelson, C.V., Schopf, J.W., 1982. Proterozoic microfossils from the SukhayaTunguska, Shorikha, and Yudoma Formations of the Siberian Platform, USSR.Journal of Paleontology 56, 42–83.
Muller, W.E.G., 2001. Review: how was metazoan threshold crossed? Thehypothetical Urmetazoa: Comparative Biochemistry Physiology, Part A.Molecular Integrated Physiology 129, 433–460.
Pant, C.C., 1985. Stromatolites from Middle Riphean Gangolihat Dolomites(Deoban), Berinag-Gangolihat area, Pithoragarh District Kumaun LesserHimalaya. Journal Geological Society of India 26, 1–9.
Qian, Y., 1999. Taxonomy and Biostratigraphy of Small Shelly Fossils in China.Science Press, Beijing, p. 247.
Raha, P.K., Chande, K.C., Balasubramanayam, S., 1978. Geochronology of the JammuLimestone, Udhampur District, Jammu, India. Journal of the Geological Societyof India 18, 93–95.
Reitlinger E.A., 1948. Kembriskei foraminiferi yakutii (Cambrian foraminifera ofYakutia). Bulletin of the Moscow Society of Naturalist, New Series 53, GeologySection, 23, 77–81.
Reitlinger E.A., 1959. Atlas Mikroskpichekikh Organicheskikh Ostatkov IProblematiki Drevnikh Tolshch Sibiri (Atlas of Microscopic Organic Remainsand Problematica of Ancient Deposits of Siberia). Trudy Geologicheskii Institute,Akademiya Nauk SSSR 25, 1–62 (translated by Associated Technical Services,Inc., New Jersey).
Reitner, J., Mehl, D., 1995. Early Paleozoic diversification of sponges: new data andevidence. Geology–Paleontology Mitt Innsbruck 20, 335–347.
Rigby, J.K., 1987. Early Cambrian sponges from Vermont and Pennsylvania, the onlyones described from North America. Journal of Paleontology 61, 451–461.
Rozanov, A.Y., Zhuravlev, A.Y., 1992. The Lower Cambrian fossil record of the SovietUnion. In: Lipps, J.H., Signor, P.W. (Eds.), Origin and Early Evolution of theMetazoa. Plenum Press, New York, pp. 205–282.
Sarkar, S.C., Chernyshev, I.V., Banerjee, H., 2000. Mid-proterozoic Pb–Pb ages forsome Himalayan base-metal deposits and comparison to deposits in Rajasthan,NW India. Precambrian Research 99, 171–178.
Schopf, J.W., 1968. Microflora of the Bitter Spring Formation, Late Precambrian,Central Australia. Journal of Paleontology 42, 651–688.
Schopf, J.W., 1992. Proterozoic prokaryotes; affinities, geological distribution andevolutionary trends. In: Schopf, J.W., Klein, C. (Eds.), The Proterozoic Biosphere:A Multidisciplinary Study. Cambridge University press, pp. 195–232.
Sergeev, V.N., 1989. Microfossils from transitional Precambrian–Phanerozoic strataof central Asia. Himalayan Geology 13, 269–278.
Sergeev, V.N., 2001. Paleobiology of the Neoproterozoic (Upper Riphean) Shorikhaand Burovaya Silicified Microbiotas, Turukhansk uplift, Siberia. Journal ofPaleontology 75, 427–448.
Sergeev, V.N., Knoll, A.H., Petrov, P.Yu., 1997. Paleobiology of the Mesoproterozoic–Neoproterozoic transition: the Sukhaya Tunguska formation, Turukhansk Uplift,Siberia. Precambrian Research 85, 201–239.
Shukla, M., Tewari, V.C., Yadav, V.K., 1987. Late Precambrian microfossils fromDeoban Limestone Formation, Lesser Himalaya, India. Palaeobotanist 35, 247–256.
Singh, S.K., Trivedi, J.R., Krishnaswami, S., 1999. Re-Os isotope systematics in blackshales from the Lesser Himalaya: Their chronology and role in the 187Os/188Osevolution of seawater. Geochimica et Cosmochimica Acta 63 (16), 2381–2392.
Song, X., 1984. Obruchevella from the early Cambrian Meishucun Stage of theMeishucun section, Jinning, Yunnan, China. Geological Magazine 121 (3), 179–183.
Srivastava, P., Kumar, S., 1997. Possible evidences of animal life in NeoproterozoicDeoban microfossil assemblage, Garhwal Lesser Himalaya, Uttar Pradesh.Current Science 72, 145–149.
Steiner, M., 1994. Die neoproterozoischen megaalgen sudchinas. Berlinergeowissenschaftliche Abhandlungen (e) 15, 1–146.
Steiner, M., Mehl, D., Reitner, J., Erdtmann, B.-D., 1993. Oldest entirely preservedsponges and other fossils from the lowermost Cambrian and a new faciesreconstruction of the Yangtze platform (China). Berliner Geowiss Abh. 9, 293–329.
Tewari, V.C., 2003. Correlation of Central Indian Vindhyans and Lesser Himalayanand Lesser Himalayan sequences, stratigraphy and Palaeobiology. GondwanaGeological Magazine 7, 383–394.
Tiwari, Meera, 1997. Nabaviella acanthomorpha n. Sp., a sponge spicule from thePrecambrian-Cambrian boundary interval in the Tethys sequence ofnorthwestern Kashmir. Journal of Geological Society of India 50, 655–658.
Tiwari, Meera, 1999. Organic-walled microfossils from the Chert-phosphoriteMember, Tal Formation, Precambrian-Cambrian Boundary, India. PrecambrianResearch 97, 99–113.
Tiwari, Meera, Azmi, R.J., 1992. Late proterozoic organic-walled microfossils fromthe Infrakrol Formation of Solan, Himachal Lesser Himalaya: an additional ageconstraint in the Krol Belt succession. Palaeobotanist 39, 387–394.
Tiwari, Meera, Knoll, A.H., 1994. Large acanthomorphic acritarch from the InfrakrolFormation of the Lesser Himalaya and their stratigraphic importance. Journal ofHimalayan Geology 5, 193–201.
Tiwari, Meera, Pant, C.C., 2004. Organic-walled microfossils from theNeoproterozoic black phosphatic stringers in the Gangolihat Dolomite, LesserHimalaya, India. Current Science 87, 1733–1738.
Tiwari, Meera, Pant, C.C., Tewari, V.C., 2000. Neoproterozoic sponge spicules andorganic-walled microfossils from the Gangolihat Dolomite, Lesser Himalaya,India. Current Science 79, 651–654.
Valdiya, K.S., 1969. Stromatolites of the Lesser Himalayan carbonate formations andthe Vindhyans. Journal of Geological Society of India 10, 1–25.
Valdiya, K.S., 1980. Geology of the Kumaun Lesser Himalaya: Wadia Institute ofHimalayan Geology. The Himachal Times Press, Dehradun. pp. 291.
Valdiya, K.S., Pant, C.C., 1981. On the existence of the Berinag Thrust, KumaunHimalaya. In: Sinha, A.K. (Eds.), Contemporary Geoscientific Researches inHimalaya, vol. 1, pp. 1–12.
Venkatachala, B.S., Kumar, Ashok, 1998. Fossil microbiota from the Vaishnodevilimestone, Himalayan foothills, and Jammu: age and Paleoenvironmentalimplications. Journal Geological Society of India 74 (1), 133–163.
Venkatachala, B.S., Shukla, M., Bansal, R. Acharyya, S.K., 1990. Upper proterozoicmicrofossils from the Infrakrol sediments, Nainital Synform, Kumaun Himalaya,India. In: Jain, K.P., Tiwari, R.S. (Eds.), Proc. Symp. Vistas in Indian Palaeobotany.Palaeobotanists 38, 29–38.
Vidal, G., Moczydlowska-Vidal, M., 1997. Biodiversity, speciation, and extinctiontrends of Proterozoic and Cambrian phytoplankton. Paleobiology 23 (2), 230–246.
Vidal, G., Moczydlowska, M., Rudavskaya, V.A., 1993. Biostratigraphical implicationsof a Chuaria-Tawuia assemblage and associated acritarchs from theNeoproterozoic of Yakutia. Palaeontology 36, 387–402.
Volkova, N.A., 1969. Acritarchs of the north west Russian platform. In: Rozanov,V.V., Missarzhevskii, N.A., Volkova, L.G., et al. (Eds.), Tommotian Stage and theCambrian Lower Boundary Problem. Nauka, Moscow, pp. 224–235.
Whitton, B.A., 1992. Diversity, ecology and taxonomy of the Cyanobacteria. In:Mann, N.H., Carr, N.G. (Eds.), Photosynthetic Prokaryotes, vols. 1–51. PlenumPress, New York.
Xiao, S., Yuan, X., Steiner, M., Knoll, A.H., 2002. Macroscopic carbonaceouscompressions in a terminal proterozoic shale; a systematic reassessment of aMiaohe biota, South China. Journal of Paleontology 76, 345–376.

M. Tiwari, I. Pant / Journal of Asian Earth Sciences 35 (2009) 137–149 149
Yan, Y., 1991. Shale- facies Microflora from the Changcheng System in Kuancheng,Hebei, and comparison with those of Neighboring areas. ActaMicropalaeontologica Sinica 12 (4), 349–373.
Yin, L., Guan, B., 1999. Organic walled microfossils of Neoproterozoic DongjiaFormation, Lushan County, Henan province, North China. Precambrian Research94, 121–137.
Yin, L., Zhu, M., Knoll, A.H., Yuan, X., Zhang, J., Hu, J., 2007. Doushantuo embryospreserved inside diapause eggs. Nature 446, 661–663.
Yuan, X., Xiao, S., Parsley, R. L., Zhou, C., Chen, Z., Hu, J., 2002. Towering sponges inan Early Cambrian Lagerstatte: disparity between non-bilaterian and bilateriansepifaunal tiers during the Neoproterozoic–Cambrian transition. Geology 30,363–366.
Zang, Wen-Long, 1995. Early Neoproterozoic sequence stratigraphy and acritarchsbiostratigraphy, Eastern Officer Basin, South Australia. Precambrian Research74, 119–175.
Zang, W., Walter, M.R., 1992. Late proterozoic and early Cambrian microfossils andbiostratigraphy, northern Anhui and Jiangsu, central eastern China.Precambrian Research 57, 243–323.
Zhang, Z., 1986. Clastic facies microfossils from the Chuanlinggou Formation(1800 Ma) near Jixian, North China. Geological Magazine 134 (2), 145–150.
Zhang, X., Pratt, B.R., 1994. New and extraordinary Early Cambrian sponge spiculeassemblage from China. Geology 22, 43–46.
Zhang, Y., Yin, L., Xiao, S., Knoll, A.H., 1998a. Permineralized fossils from theterminal Proterozoic Doushantuo Formation, South China. The PaleontologicalSociety Memoir 50, 52.
Zhang, Y., Yuan, X., Yin, L., 1998b. Interpreting Late Precambrian microfossils.Science 282, 1783.
Zhou, C., Yuan, X., Xue, Y., 1998. Sponge spicule-like pseudofossils from theNeoproterozoic Doushantuo formation in Weng’an, Guizhou, and China. ActaMicropalaeontologica Sinica 15, 380–384.




















