
Methylmercury production in the water column of an ultraoligotrophic lake of Northern Patagonia,...
8
Methylmercury production in the water column of an ultraoligotrophic lake of Northern Patagonia, Argentina Sergio Ribeiro Guevara a, * , Claudia P. Queimaliños b , María del Carmen Diéguez b , María Arribére a a Laboratorio de Análisis por Activación Neutrónica, Comisión Nacional de Energía Atómica, Centro Atómico Bariloche, Av. Bustillo km 9.5, 8400 Bariloche, Argentina b Laboratorio de Fotobiología, Centro Regional Universitario Bariloche, INIBIOMA, Universidad Nacional del Comahue-CONICET, Quintral 1250, 8400 Bariloche, Argentina article info Article history: Received 17 December 2007 Received in revised form 11 March 2008 Accepted 12 March 2008 Available online 28 April 2008 Keywords: Mercury Abiotic mercury methylation Phytoplankton 197 Hg radiotracer Freshwater Nahuel Huapi National Park abstract Methyl-mercury (CH 3 Hg + ) production was studied in freshwaters from lake Moreno, an ultraoligotrophic system belonging to Northern Patagonia. Hg 2+ labelled with high specific activity 197 Hg was spiked to water samples in concentrations of 10 ng l 1 , and incubated in laboratory for 3 d time trends under dif- ferent conditions. Experimental water was sampled daily to evaluate CH 3 197 Hg + production. Lake water used in the experiments was sampled just below the upper limit of the metalimnion (30 m depth), where maximum values of chlorophyll a have been measured previously. Sampling was performed in late autumn, when the plankton fraction <50 lm exhibited mercury concentrations up to 260 lgg 1 dry weight. The experiments analysed lake water filtered through 50, 20, and 0.2 lm (filter-sterilized) mesh nets. ASTM grade 1 water was also incubated for control. All the experiments were run in an environmen- tal chamber under controlled temperature and light regime. High Hg 2+ conversion to CH 3 Hg + , up to 50%, was measured in lake water, in a process stimulated by light. CH 3 Hg + production was two-fold higher after 3 d of incubation with illumination compared to total dark- ness. Sterile lake water showed conversions up to 30%, while the planktonic components seem to enhance the CH 3 Hg + production. Overall, our results provide evidence that lake Moreno waters favour CH 3 Hg + production in processes stimulated by light. Although biotic components certainly contribute to enhance mercury methylation, water chemistry plays a key role in this process. We hypothesize that dissolved organic matter, particularly its quality, could be decisive. Ó 2008 Elsevier Ltd. All rights reserved. 1. Introduction In natural aquatic ecosystems, inputs of mercury can be influ- enced by atmospheric deposition, geology, soil and vegetation type and extreme precipitation or catastrophic events (Leermakers et al., 1996; Fostier et al., 2000; Guimaraes et al., 2000; Ullrich et al., 2001). The pool of mercury is composed by elemental Hg, complexes of Hg 2+ and organic mercury forms, mainly methyl mer- cury (CH 3 Hg + ) which is a neurotoxic compound that can bioaccu- mulate through the food web (Boening, 2000). The distribution and transformation processes of mercury species are known to be controlled by several factors, including physical (i.e. tempera- ture and light), chemical, and biological (particularly microorgan- isms activity) (Sellers et al., 1996; Ullrich et al., 2001). Although there is substantial evidence regarding the fate of Hg in aquatic environments, the transformation and distribution mechanisms are still poorly understood (Ullrich et al., 2001). The westernmost area of North Patagonia (Argentina) is en- dowed with hundreds of large and deep ultraoligotrophic as well as smaller and shallower oligo to mesotrophic lakes. Although there is no relevant industrial or mining activity in the Nahuel Huapi National Park (Northwestern Patagonia), and no anthropo- genic mercury point sources have been identified, high mercury levels have been detected in different compartments of the aquatic ecosystems (Arribére et al., 2005; Ribeiro Guevara et al., 2005; Arri- bére et al., 2008). Mercury content in fish is high, up to 4 lgg 1 DW (dry weight) in muscle. While these values are not unusual in unpolluted lakes (Duffy et al., 1999; Ullrich et al., 2001), the fact that mercury bio-magnification was not observed in the surveyed lakes (Moreno and Nahuel Huapi) is atypical (Arri- bére et al., 2005). Mercury contents in sediments reached 7 lgg 1 (Ribeiro Guevara et al., 2005). The distinctive feature of these sys- tems, particularly of lake Moreno Oeste (West branch of lake Moreno), is that low Hg methylation was observed in the sedi- ments, the predominant source accepted for CH 3 Hg + production in most freshwater systems (Ullrich et al., 2001). A concurrent study being conducted by the authors on total mercury contents in different planktonic fractions of lake Moreno showed up decreasing concentrations with increasing organism size, reaching up to 260 lgg 1 DW in organisms ranging from 10 to 50 lm cell size, while the organisms >200 lm presented 0045-6535/$ - see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.chemosphere.2008.03.011 * Corresponding author. Tel.: +54 2944 445106; fax: +54 2944 445299. E-mail address: [email protected] (S. Ribeiro Guevara). Chemosphere 72 (2008) 578–585 Contents lists available at ScienceDirect Chemosphere journal homepage: www.elsevier.com/locate/chemosphere
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Methylmercury production in the water column of an ultraoligotrophic lake of Northern Patagonia,...

Chemosphere 72 (2008) 578–585
Contents lists available at ScienceDirect
Chemosphere
journal homepage: www.elsevier .com/locate /chemosphere
Methylmercury production in the water column of an ultraoligotrophic lakeof Northern Patagonia, Argentina
Sergio Ribeiro Guevara a,*, Claudia P. Queimaliños b, María del Carmen Diéguez b, María Arribére a
a Laboratorio de Análisis por Activación Neutrónica, Comisión Nacional de Energía Atómica, Centro Atómico Bariloche, Av. Bustillo km 9.5, 8400 Bariloche, Argentinab Laboratorio de Fotobiología, Centro Regional Universitario Bariloche, INIBIOMA, Universidad Nacional del Comahue-CONICET, Quintral 1250, 8400 Bariloche, Argentina
a r t i c l e i n f o
Article history:Received 17 December 2007Received in revised form 11 March 2008Accepted 12 March 2008Available online 28 April 2008
Keywords:MercuryAbiotic mercury methylationPhytoplankton197Hg radiotracerFreshwaterNahuel Huapi National Park
0045-6535/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.chemosphere.2008.03.011
* Corresponding author. Tel.: +54 2944 445106; faxE-mail address: [email protected] (S. Ribeiro
a b s t r a c t
Methyl-mercury (CH3Hg+) production was studied in freshwaters from lake Moreno, an ultraoligotrophicsystem belonging to Northern Patagonia. Hg2+ labelled with high specific activity 197Hg was spiked towater samples in concentrations of 10 ng l�1, and incubated in laboratory for 3 d time trends under dif-ferent conditions. Experimental water was sampled daily to evaluate CH3
197Hg+ production. Lake waterused in the experiments was sampled just below the upper limit of the metalimnion (�30 m depth),where maximum values of chlorophyll a have been measured previously. Sampling was performed in lateautumn, when the plankton fraction <50 lm exhibited mercury concentrations up to 260 lg g�1 dryweight. The experiments analysed lake water filtered through 50, 20, and 0.2 lm (filter-sterilized) meshnets. ASTM grade 1 water was also incubated for control. All the experiments were run in an environmen-tal chamber under controlled temperature and light regime.High Hg2+ conversion to CH3Hg+, up to 50%, was measured in lake water, in a process stimulated by light.CH3Hg+ production was two-fold higher after 3 d of incubation with illumination compared to total dark-ness. Sterile lake water showed conversions up to 30%, while the planktonic components seem toenhance the CH3Hg+ production. Overall, our results provide evidence that lake Moreno waters favourCH3Hg+ production in processes stimulated by light. Although biotic components certainly contributeto enhance mercury methylation, water chemistry plays a key role in this process. We hypothesize thatdissolved organic matter, particularly its quality, could be decisive.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
In natural aquatic ecosystems, inputs of mercury can be influ-enced by atmospheric deposition, geology, soil and vegetation typeand extreme precipitation or catastrophic events (Leermakerset al., 1996; Fostier et al., 2000; Guimaraes et al., 2000; Ullrichet al., 2001). The pool of mercury is composed by elemental Hg,complexes of Hg2+ and organic mercury forms, mainly methyl mer-cury (CH3Hg+) which is a neurotoxic compound that can bioaccu-mulate through the food web (Boening, 2000). The distributionand transformation processes of mercury species are known tobe controlled by several factors, including physical (i.e. tempera-ture and light), chemical, and biological (particularly microorgan-isms activity) (Sellers et al., 1996; Ullrich et al., 2001). Althoughthere is substantial evidence regarding the fate of Hg in aquaticenvironments, the transformation and distribution mechanismsare still poorly understood (Ullrich et al., 2001).
The westernmost area of North Patagonia (Argentina) is en-dowed with hundreds of large and deep ultraoligotrophic as well
ll rights reserved.
: +54 2944 445299.Guevara).
as smaller and shallower oligo to mesotrophic lakes. Althoughthere is no relevant industrial or mining activity in the NahuelHuapi National Park (Northwestern Patagonia), and no anthropo-genic mercury point sources have been identified, high mercurylevels have been detected in different compartments of the aquaticecosystems (Arribére et al., 2005; Ribeiro Guevara et al., 2005; Arri-bére et al., 2008). Mercury content in fish is high, up to4 lg g�1 DW (dry weight) in muscle. While these values are notunusual in unpolluted lakes (Duffy et al., 1999; Ullrich et al.,2001), the fact that mercury bio-magnification was not observedin the surveyed lakes (Moreno and Nahuel Huapi) is atypical (Arri-bére et al., 2005). Mercury contents in sediments reached 7 lg g�1
(Ribeiro Guevara et al., 2005). The distinctive feature of these sys-tems, particularly of lake Moreno Oeste (West branch of lakeMoreno), is that low Hg methylation was observed in the sedi-ments, the predominant source accepted for CH3Hg+ productionin most freshwater systems (Ullrich et al., 2001).
A concurrent study being conducted by the authors on totalmercury contents in different planktonic fractions of lake Morenoshowed up decreasing concentrations with increasing organismsize, reaching up to 260 lg g�1 DW in organisms ranging from10 to 50 lm cell size, while the organisms >200 lm presented

S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585 579
concentrations ranging from 0.2 to 2.4 lg g�1 DW (Arribére et al.,unpublished data). Total mercury concentrations in phyto andzooplankton of lake Moreno are extraordinarily high when evalu-ated in relation with values reported by other researchers. How-ever, mercury concentrations of the macro-zooplankton of lakeMoreno are close to those reported in the literature (Rask et al.,1994; Watras et al., 1998).
These previous results of mercury contents in plankton fromlake Moreno drove our attention to the role of the planktonic com-munity in mercury transformation processes, particularly in mer-cury methylation in the water column. Watras and Bloom (1994)found that planktonic production of CH3Hg+ was potentially a sig-nificant source in three lakes in Wisconsin, USA, but emphasizedthat direct estimates of methylation were needed to confirm theircalculations (Watras and Bloom, 1994). The evaluation of mercurymethylation in laboratory experiments by using labelled mercurycould represent a direct assessment of these processes in the watercolumn. More recently, Monperrus et al. (2007) found that CH3Hg+
formation is significantly influenced by both benthic and pelagicdynamics in a coastal lagoon. Mercury methylation processes havebeen studied in sediments, periphyton, and rooted macrophytesusing mercury labelled with a radioactive isotope (Guimaraeset al., 2000; Marvin-DiPasquale et al., 2003) or with stable isotopeenrichments (Mauro et al., 2002; Monperrus et al., 2007). The eval-uation of the mercury methylation potential is based on theamount of labelled Hg2+ converted to CH3Hg+. Furthermore, theknowledge and measurement of pre-existing native CH3Hg+ inthe environmental sample is unnecessary. These methods are nor-mally based on a simple and rapid CH3Hg+ separation technique forevaluation of the environmental factors influencing the CH3Hg+
production from the added Hg2+, by spiking the sample with la-belled Hg2+ and incubating it for a certain period under variousenvironmental conditions. Any labelled CH3Hg+ produced is thenseparated from the bulk, evaluating the activity of the product bygamma or beta counting when using a radiotracer. The specificactivity of the recovered CH3Hg+ after incubation is a measure ofthe methylation potential, as the result of all transformation pro-cesses (methylation, demethylation and reduction/oxidation pro-cesses) that occur simultaneously. Control samples are typically akey part of such studies, necessary to evaluate the carry-over ofunreacted Hg2+ into the organic extraction phase (Pérez Catánet al., 2007).
The study of mercury circulation and transformation processesin the water column using labelled mercury has been limited bythe impossibility of the existing techniques to simulate natural lev-els or real environmental contamination situations, since one ofthe factors affecting mercury transformation is its concentration(Zizek et al., 2008). In general, uncontaminated freshwaters con-tain less than 5 ng l�1 of total dissolved mercury (Ullrich et al.,2001). Natural concentrations in fresh and estuarine waters mayrange from 0.1 to 6 ng l�1 (Cossa et al., 1996; Leermakers et al.,1996; Fostier et al., 2000). In stream waters, dissolved mercurymay vary from 1 to 6 ng l�1 in a natural non-polluted area, whilein a deforested area of the same region it varies from 4 to 19 ng l�1
(Fostier et al., 2000). Up to 10–20 ng l�1 concentrations can befound in humic lakes or rivers rich in particulate mercury (Meili,1997). In the case of direct industrial contamination, the concen-trations in freshwater can range between 0.5 and 12 ng l�1 (Cossaet al., 1996). Mercury concentrations may range from 1 to 30 ng l�1
in rainwater (Leermakers et al., 1996; Fostier et al., 2000), and from9 to 60 ng l�1 in snow (Leermakers et al., 1996). Recent researchmeasured CH3Hg+ production from Hg2+ in marine and freshwatersystems using the short-lived radiotracer 197Hg (T1/2 = 64.14 h)produced from Hg isotopically enriched in 196Hg, with Hg2+ addi-tions of 10 and 12 ng l�1 (Ribeiro Guevara et al., 2007). The evalu-ation of CH3Hg+ production potentials in water samples using the
radiotracer 197Hg is the methodological approach selected to assessthe relevance of the water column of lake Moreno Oeste as aCH3Hg+ source to the trophic web.
2. Methods
2.1. Study site
Water samples were obtained from lake Moreno Oeste, the wes-tern branch of lake Moreno (41�50S; 71�330W, 758 m above see le-vel), a freshwater system included in the Nahuel Huapi basin(Nahuel Huapi National Park, Patagonia, Argentina). The samplingsite was located at the point of maximum depth of the lake, wherea previous study detected the highest mercury concentrations inplankton samples.
Nahuel Huapi National Park is the largest protected natural areaof Argentina, and comprises a drainage basin that includes threemajor river systems, 13 lakes of more than 10 km2, and severalhundred small lakes and ponds. All watersheds within the parkare glacial in origin and many of the lakes have been formed bythe damming effect of the frontal moraines left behind by retreat-ing glaciers. Within the Park’s limits there are pristine as well ashuman impacted areas, as San Carlos de Bariloche city and suburbswith a population of 100000 people. Its economy, as well as inother small towns and villages in the Park, is based on tourism.
Lake Moreno Oeste has a surface area of 6 km2 and a maximumdepth of 90 m. It is a warm monomictic system, stratifying fromlate spring to early autumn, with a marked thermocline around30 m depth (Queimaliños et al., 1999). It is an ultraoligotrophicsystem, with low dissolved organic carbon (DOC) concentrationsranging between 0.4 and 0.7 mg l�1. Chemical, physical, and limno-logical parameters of lake Moreno are shown in Table 1.
In this lake, chlorophyll a vertical distribution shows maximumvalues near the limit of the euphotic zone (1% of photosyntheticallyactive radiation, PAR, reaching up to 30 m), just below the upperlimit of the metalimnion (Queimaliños et al., 1999) or in hypolim-netic layers (Callieri et al., 2007). The autotrophic communities atthe chlorophyll a maximum are dominated by a mixotrophic ciliate(Ophrydium naumanni, 80–100 lm) and naked dinoflagellates(Gymnodinium paradoxum and Gymnodinium uberrimum, 30–40 lm) (Queimaliños et al., 1999, 2002).
Water samples for laboratory experiments were obtained inearly autumn (April 2007) at the end of the water stratification per-iod, when the highest concentrations of total Hg (260 lg g�1 DW)have been measured in the planktonic fraction <50 lm. The sampleswere obtained at 30 m depth, where peaks of chlorophyll a occur(Queimaliños et al., 1999). Water was collected at this depth priorto each experiment, using a messenger-activated 2 l Van Dorn bot-tle. Water temperature was measured within the sampler by meansof an attached thermometer. About 30 l of lake water were collectedfor each experimental set. Lake water was filtered in the fieldthrough a plankton net of 50 lm mesh to assure sample homogene-ity, and immediately transported to the laboratory in sterilizedplastic vessels, in darkness and thermally isolated.
2.2. Experimental
During three consecutive weeks in April 2007, we performedthree series of experiments to assess the potential of the water col-umn of lake Moreno Oeste for CH3Hg+ production. Basically, 3 dincubation time trends of lake water under different treatmentsafter amending Hg2+ labelled with high specific activity 197Hg,were daily evaluated for Hg2+ conversion to CH3Hg+. Three watertreatments were included in each experimental series, runningtriplicates for each case.

Table 1Lake Moreno characterization
Parameter Value
Water temperature (measured in situ after watersampling)
11–12 �C
Water conductivitya 37.1 lS cm�1
Water pHa 6.8Dissolved O2
b SaturationTotal phosphorus at sampling sitec 2–4 lg l�1
Total dissolved phosphorus at sampling sitec 1–2 lg l�1
Total nitrogena 50 lg l�1
Dissolved inorganic nitrogena 13 lg l�1
Dissolved organic carbon (DOC) at sampling sitec 0.4–0.7 mg l�1
Dissolved organic carbon (DOC)b 0.6 mg l�1
Na+a 1.2 mg l�1
K+a 0.3 mg l�1
Cl�a 1 mg l�1
Ca2+a 4.8 mg l�1
Mg2+a 1.4 mg l�1
SO2�4
a 1.3 mg l�1
HCO�3a 23 mg l�1
SiO2a 10 mg l�1
Chlorophyll a at sampling depth in sampling sitec 1 lg l�1
Chlorophyll ab 1 lg l�1
Phytoplankton abundance at sampling depth insampling site, in Aprilc
1000 cell ml�1
PAR illumination at sampling depth in sampling sited 10–30 lmol quanta m�2 s�1
Chemical, physical, and limnological parameters of lake Moreno and water samplesstudied.
a Díaz et al. (2007).b Souza et al. (2007).c Queimaliños et al. (1999).d Pérez et al. (2002).
580 S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585
The first experiment was designed to test the incidence of illu-mination on CH3Hg+ production assuming that light is one of themain abiotic factors influencing the CH3Hg+ concentration in thewater column directly, i.e. by photodegradation of CH3Hg+ (Sellerset al., 1996), and/or indirectly through its regulation of biotic vec-tors and photodegradation of dissolved organic matter (DOM),which may release methyl groups enhancing methylation reac-tions (Siciliano et al., 2005). Besides, light penetration in most An-dean lakes is a driving factor that strongly regulates thedistribution of aquatic organisms (Queimaliños et al., 1999; Pérezet al., 2002; Alonso et al., 2004). Lake water filtered through a50 lm mesh was incubated under two illumination conditions:complete darkness and a cycle of 12 h PAR (400–700 nm) + UV-A(320–400 nm):12 h dark. In order to assess CH3Hg+ production inlake water, ASTM grade 1 water was included as control, under alight regime of 12 h PAR:12 h dark. The second experiment aimedto determine the incidence of the biotic components of lakeMoreno Oeste waters in CH3Hg+ production. Two water treatmentswith the planktonic fractions <50 and <20 lm were set up. Theseplanktonic fractions were obtained by filtration of whole lakewater through 50 and 20 lm mesh nets, respectively. The formerincludes picoplankton (0.2–2 lm), constituted by bacteria and pic-ocyanobacteria; nanoplankton (mainly phytoflagellates between 2and 20 lm), and a fraction of microplankton (algae and ciliates be-tween 20 and 50 lm), while the latter comprises nanoplanktonicand picoplanktonic organisms. A third treatment in which all thebiological fractions comprised in the water were removed by filtra-tion (Millipore sterilized filters, 0.2 lm pore size), was set up todistinguish the direct incidence of the biotic components fromthe role of water chemistry. All treatments were run under an illu-mination regime of 12 h PAR:12 h dark. After the evaluation of thefirst and second experimental runs, a third experimental set wasdesigned to assess the role of lake water chemistry on CH3Hg+ pro-duction. Three dilution steps of filter-sterilised lake water withASTM grade 1 water, to 100%, 10%, and 1%, were studied. This
experiment was run in complete darkness. As the pH of ASTMgrade 1 water is around 5.2, and lake water has a pH of 6.5, thedilution range in our experimental treatments encompassed a pHgradient.
The experiments were performed in an environmental testchamber (Sanyo MLR-350), set at 12 �C, the temperature measuredin the lake at sampling. The water samples conditioned for eachexperiment were placed in the environmental chamber 12 h beforethe radiotracer amendment. The incubations were performed in 3 lglass flasks, except when treated with UV-A in which case 2.2 l UVtranslucent flasks (UVT, Plasmatic, Spain) were used. PAR wasprovided by fluorescent lamps (Sanyo Jet FL40SS-W/37) at55 lmol quanta m�2 s�1 and UV-A by Q-Panel 340 lamps, at0.70 W m�2. The samples incubated in darkness were placed intothe environmental chamber thoroughly wrapped with aluminiumfoil.
The CH3Hg+ production in the water samples was assessed byspiking Hg2+ labelled with high specific activity 197Hg, and measur-ing the radiotracer specific activity after CH3Hg+ extraction into anorganic phase. The ratio of the radiotracer specific activity in theCH3Hg+ extraction solution to the specific activity in the water isa measure of the CH3Hg+ production from the spiked Hg2+ duringeach incubation time. Sixty millilitres water samples were takenout daily from each incubation flask in glass vials, including time0 (water sampled immediately after labelled Hg2+ spiking) for con-trol, for CH3
197Hg+ extraction. Concurrently, 7 ml water sampleswere collected directly in a c-ray measurement vial to measurethe water 197Hg specific activity. The water 197Hg specific activityat time 0 was considered as the reference to compute CH3Hg+ pro-duction in each incubation flask. The 197Hg specific activity of totalHg in the water solution was measured for each incubation time toevaluate Hg losses from the aqueous system. Sample manipulationwas performed using sterilized elements and the working surfacewas properly cleaned using pure ethanol and a flame was kept dur-ing the extraction process.
The high specific activity 197Hg2+ radiotracer used in theseexperiments was produced by irradiation of 0.2 ml of a 2% HNO3
solution of Hg enriched to 51.6% in the 196Hg isotope (naturalabundance is 0.15%) in the nuclear research reactor RA-6, CentroAtómico Bariloche, Argentina (Ribeiro Guevara et al., 2007). Work-ing solutions were obtained by dilution in ASTM grade 1 water;0.1 ml of this solution was added to each incubation flask to startthe CH3Hg+ production time trend, homogenising immediatelythe solution with a sterilized glass bar. Hg2+ spikes were adjustedto obtain a final concentration ranging from 9 to 12 ng l�1
(Table 2).A solvent extraction method based on the use of KBr/H2SO4 was
employed to separate the labelled CH3Hg+ from the aqueous solu-tion to toluene. The extraction procedure and the method testingare described by Ribeiro Guevara et al. (2007). Other organic spe-cies are likely to be extracted in the toluene phase, but sinceCH3Hg+ is largely the predominant organic species in fresh watersystems (Meili, 1997; Ullrich et al., 2001) other contributions areassumed as negligible. X-ray and c-ray emissions associated withthe 197Hg decay were measured using coaxial and planar high-purity germanium detectors.
The bacterial activity was stopped for time 0 controls by addi-tion of extraction reagents immediately after tracer amendment(Pérez Catán et al., 2007). Sample collection for time 0 controlswas performed after 197Hg2+ tracer amendment and water homog-enisation, tasks that demanded about 1 min from tracer spikinguntil the extraction reagents were added to the water, and bioticor chemical CH3
197Hg+ production was hence stopped. Therefore,197Hg2+ recoveries on time 0 controls are associated to Hg2+ carryover but also to real biotic or chemical CH3
197Hg+ production dur-ing this period.

S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585 581
2.3. Statistical analyses
The incidence of the composition of lake Moreno water, and theeffects of dilution and radiation regime on the production CH3Hg+
were analysed using one-way repeated measures analysis of vari-ance. Whenever the effects of these factors were significant, posthoc contrasts were performed to compare treatments.
Assuming that all the experiments were set up in similar condi-tions, especially regarding to the chemical and biological featuresof lake water, we performed one comparison between treatmentsbelonging to different experiments. The assumption of biologicalhomogeneity was supported by studying the planktonic commu-nity structure from water samples obtained concurrently in eachsampling date following Utermöhl technique (Utermöhl, 1958).
3. Results
3.1. Considerations on the methodological approach
When spiking 197Hg2+ in the water under study, conversion toCH3
197Hg+ may occur during the incubation period, but other pro-cesses implying losses of labelled Hg from the aqueous systemsshould be taken into account for a correct evaluation of the meth-ylation process. Adsorption of 197Hg2+ onto the walls of the incuba-tion flasks (water pH is close to 7; Table 1), and 197Hg0 release fromthe aqueous solution in vapour form, are two potential pathwaysto push labelled Hg out of the system under study. The evolutionof the activity of total 197Hg radiotracer in water along the incuba-tion time trends (referred to the activity at time 0) showed randomvariations within ±10% in all cases. Fig. 1 shows the results ob-tained when incubating lake water sterilized and containing thetwo plankton fractions considered, under an illumination regimeof 12 h PAR:12 h dark. Similar patterns were obtained in the otherruns. Since no systematic decrease was observed during the 3 dincubation time trends, even though the variability is high, wecan assume that there were no relevant 197Hg losses from theaqueous solutions in any of the cases studied.
197Hg tracer recoveries on control water (ASTM grade 1 water)ranged from 0.5% to 2.9%, with similar values after 1–3 d incuba-tion periods, considering the uncertainties, but higher than time0 incubation (Table 2 and Fig. 2). Recoveries of time 0 controls oflake waters ranged from 0.4% to 1.2% (Table 2). Only the recoveryafter 3 d incubation time of control water resulted slightly higherthan the range of both kinds of controls. Although these valuesare higher than those observed in controls in a previous work inmarine and fresh waters (Ribeiro Guevara et al., 2007), our resultsof control water (ASTM grade 1) and time 0 incubations controlsare consistent. 197Hg tracer recoveries of controls are much lower
Table 2197Hg recoveries after CH3Hg+ extraction, referred to 197Hg2+ spiked (%)
Water 197Hg2+ spiked: final concentration (ng l�1
ASTM grade 1; 12:12 h PAR illumination 9.0Sterilized, 1% dilution, in darkness 12.2Sterilized, 10% dilution, in darkness 12.2
Sterilized, in darkness 12.2<50 lm, in darkness 11.9
Sterilized, 12:12 h PAR illumination 9.7<20 lm, 12:12 h PAR illumination 9.7<50 lm, 12:12 h PAR illumination 9.7<50 lm, 12:12 h UV-A + PAR illumination 9.0
Average values on triplicates. Water temperature during incubation: 11–12 �C.
than the recoveries in any case of incubated lake water, that rangedfrom 10% (sterilized lake water, 1 d incubation in darkness), to 52%(lake water <50 lm, 3 d incubation in 12 h PAR + UV-A:12 h dark)(Figs. 2–4 and Table 2). Therefore, actual Hg methylation was ob-tained in lake waters, while tracer recoveries on controls wouldbe associated mostly to Hg2+ carry over to the organic phase duringCH3Hg+ extraction (Ribeiro Guevara et al., 2007). Lake water di-luted to 1% in ASTM grade 1 water, incubated in dark condition,showed tracer recoveries ranging from 0.8% to 2.6%, decreasingto 1.2% after 3 d incubation. These values were similar to the con-trols’, considering uncertainties, while dilution to 10% showedslightly higher recoveries (Fig. 4 and Table 2). These results confirmthat the increase in 197Hg tracer recoveries after CH3Hg+ extractionin lake water represents Hg methylation during incubation associ-ated to water composition and experimental conditions.
The higher 197Hg tracer recoveries on time 0 controls can beassociated to an additional contribution to Hg2+ carry over of realbiotic and chemical CH3Hg+ production during the period elapsedfrom tracer spiking to the addition of CH3Hg+ extraction reagents,particularly regarding the high CH3Hg+ production rates that canbe extrapolated for short incubation times from the time trendstudied (Figs. 2–4). The differences of the 197Hg tracer recoveriesof time 0 controls of the different cases studied (from 0.4% to1.2%; Table 2) could, therefore, be associated to the variability ofthe period between spiking and addition of extraction reagents,and the incubation conditions, that showed to influence theCH3Hg+ production.
3.2. CH3Hg+ production
The first experimental run studied the 197Hg2+ conversion toCH3
197Hg+ in ASTM grade 1 water, and in lake water with plankton<50 lm in darkness and PAR + UV-A illumination. The time trend of197Hg recoveries increased notably over incubation time in lakewater (RM ANOVA, p < 0.0001), significantly higher than time 0controls. We found also significant differences between the threetreatments (p < 0.0001), with higher methylation in lake waterthan in ASTM grade 1 water. The CH3Hg+ production values rangedfrom 15.44 ± 0.77% to 23.5 ± 1.4% (after deduction of time 0 con-trols) for incubation in darkness, and from 27.3 ± 9.0% to50.8 ± 6.9% for water under PAR + UV-A (Fig. 2 and Table 2). The197Hg tracer recovery along the incubation time trend of lake waterin darkness showed a positive step after 1 d, but similar valuesafter 2 and 3 d of incubation (Fig. 2). This behaviour can be ex-plained either by saturation of the CH3Hg+ production reaction orby the incidence of demethylation processes. Formation of Hgcomplexes by high molecular weight DOC could also explain thesaturation tendency. CH3Hg+ production of lake water under
) Incubation time
0 (control) 1 d 2 d 3 d
Average SD Average SD Average SD Average SD
0.52 0.10 1.21 0.54 2.4 1.2 2.89 0.620.78 0.23 1.08 0.31 2.6 1.9 1.25 0.38– – 1.40 0.37 3.27 0.86 3.1 1.8
1.22 0.40 10.0 2.3 14.4 4.2 18.5 4.11.08 0.26 16.41 0.72 22.8 1.8 24.6 1.4
0.68 0.52 12.0 9.1 27 19 31 110.40 0.14 15.9 3.3 25.8 9.7 33 110.83 0.25 24.6 8.0 30 13 36 160.98 0.21 23 11 40.5 1.7 51.8 6.9
0 1 2 3
0.0
0.2
0.4
0.6
0.8
1.0
1.2
lake water sterilized lake water < 20 μm lake water < 50 μm
197 H
g ac
tivity
in w
ater
: rat
io to
tim
e 0
Incubation time (d)
Fig. 1. 197Hg activity in water sampled after each incubation period: ratio to themeasurement at time 0. Experimental run 2.
0 1 2 3
0
10
20
30
40
50
60
197 H
g re
cove
ry a
fter
CH
3Hg+ e
xtra
ctio
n (%
)
Incubation time (d)
ASTM grade 1 water Total plankton in dark Total plankton in 12-12 h light-dark
Fig. 2. Control water (ASTM grade 1) and lake water filtered <50 lm under twoillumination conditions. Experimental run 1. 197Hg measurements on toluene phaseafter CH3Hg+ extraction, relative to the specific activity of total 197Hg at time 0.Average values on triplicates (the uncertainty is the standard deviation of theaverage).
0 1 2 3
0
10
20
30
40
50
60
197 H
g re
cove
ry a
fter
CH
3Hg+ e
xtra
ctio
n (%
)
Incubation time (d)
lake water sterilized lake water < 20 μm lake water < 50 μm
Fig. 3. Lake water filtered <50, <20, and <0.2 lm (sterile). PAR illumination in12:12 h light:dark sequence. Experimental run 2. 197Hg measurements on toluenephase after CH3Hg+ extraction, relative to the specific activity of total 197Hg at time0. Average values on triplicates (the uncertainty is the standard deviation of theaverage).
582 S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585
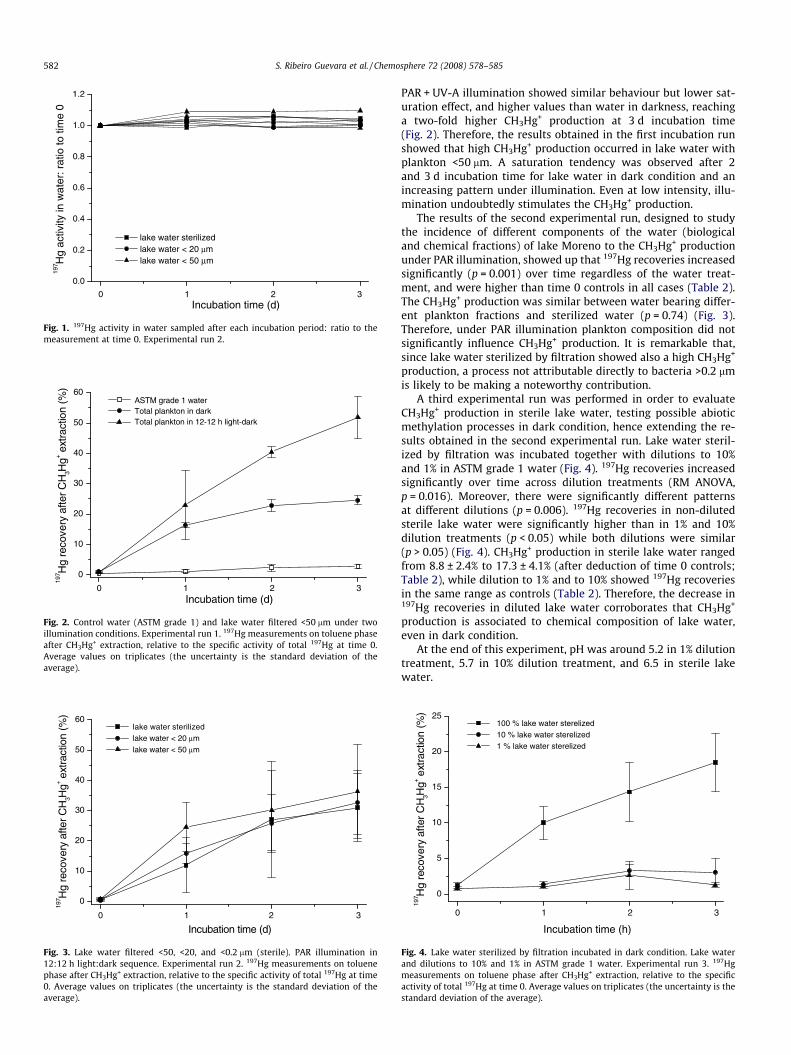
PAR + UV-A illumination showed similar behaviour but lower sat-uration effect, and higher values than water in darkness, reachinga two-fold higher CH3Hg+ production at 3 d incubation time(Fig. 2). Therefore, the results obtained in the first incubation runshowed that high CH3Hg+ production occurred in lake water withplankton <50 lm. A saturation tendency was observed after 2and 3 d incubation time for lake water in dark condition and anincreasing pattern under illumination. Even at low intensity, illu-mination undoubtedly stimulates the CH3Hg+ production.
The results of the second experimental run, designed to studythe incidence of different components of the water (biologicaland chemical fractions) of lake Moreno to the CH3Hg+ productionunder PAR illumination, showed up that 197Hg recoveries increasedsignificantly (p = 0.001) over time regardless of the water treat-ment, and were higher than time 0 controls in all cases (Table 2).The CH3Hg+ production was similar between water bearing differ-ent plankton fractions and sterilized water (p = 0.74) (Fig. 3).Therefore, under PAR illumination plankton composition did notsignificantly influence CH3Hg+ production. It is remarkable that,since lake water sterilized by filtration showed also a high CH3Hg+
production, a process not attributable directly to bacteria >0.2 lmis likely to be making a noteworthy contribution.
A third experimental run was performed in order to evaluateCH3Hg+ production in sterile lake water, testing possible abioticmethylation processes in dark condition, hence extending the re-sults obtained in the second experimental run. Lake water steril-ized by filtration was incubated together with dilutions to 10%and 1% in ASTM grade 1 water (Fig. 4). 197Hg recoveries increasedsignificantly over time across dilution treatments (RM ANOVA,p = 0.016). Moreover, there were significantly different patternsat different dilutions (p = 0.006). 197Hg recoveries in non-dilutedsterile lake water were significantly higher than in 1% and 10%dilution treatments (p < 0.05) while both dilutions were similar(p > 0.05) (Fig. 4). CH3Hg+ production in sterile lake water rangedfrom 8.8 ± 2.4% to 17.3 ± 4.1% (after deduction of time 0 controls;Table 2), while dilution to 1% and to 10% showed 197Hg recoveriesin the same range as controls (Table 2). Therefore, the decrease in197Hg recoveries in diluted lake water corroborates that CH3Hg+
production is associated to chemical composition of lake water,even in dark condition.
At the end of this experiment, pH was around 5.2 in 1% dilutiontreatment, 5.7 in 10% dilution treatment, and 6.5 in sterile lakewater.
0 1 2 3
0
5
10
15
20
25
197 H
g re
cove
ry a
fter
CH
3Hg+ e
xtra
ctio
n (%
)
Incubation time (h)
100 % lake water sterelized 10 % lake water sterelized 1 % lake water sterelized
Fig. 4. Lake water sterilized by filtration incubated in dark condition. Lake waterand dilutions to 10% and 1% in ASTM grade 1 water. Experimental run 3. 197Hgmeasurements on toluene phase after CH3Hg+ extraction, relative to the specificactivity of total 197Hg at time 0. Average values on triplicates (the uncertainty is thestandard deviation of the average).
0
0
10
20
30
197 H
g tr
acer
rec
over
y af
ter
CH
3Hg+ e
xtra
ctio
n (%
)
Incubation time (d)
lake water < 50 μm, in darkness
lake water sterile, in darkness
0 1 2 3
1 2 3
0
10
20
30
40
50
60 lake water < 50 μm in darkness
lake water < 50 μm PAR illumination
lake water < 50 μm UV-A + PAR illumination
a
b
Fig. 5. CH3Hg+ production in the incubation time trends: comparison between di-fferent experimental runs. (A) Incidence of the illumination condition in lake water<50 lm. (B) Direct incidence of plankton on CH3Hg+ production in dark condition.197Hg measurements on toluene phase after CH3Hg+ extraction, relative to the sp-ecific activity of total 197Hg at time 0. Average values on triplicates (the uncertaintyis the standard deviation of the average).
S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585 583
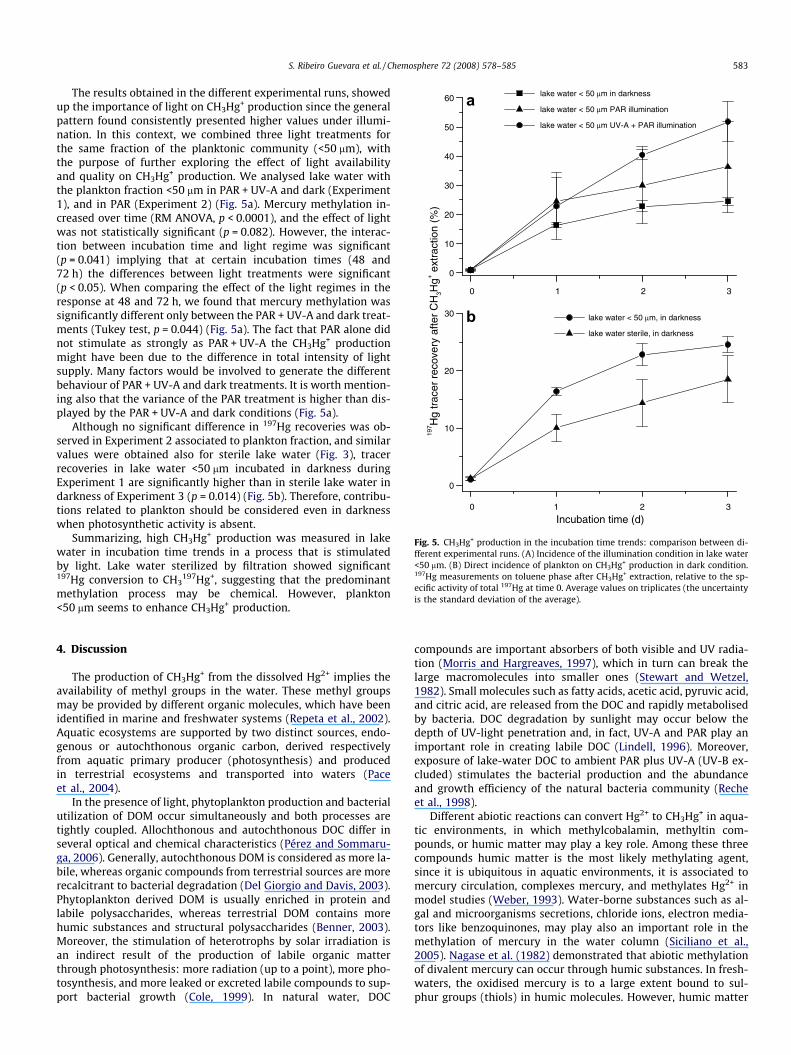
The results obtained in the different experimental runs, showedup the importance of light on CH3Hg+ production since the generalpattern found consistently presented higher values under illumi-nation. In this context, we combined three light treatments forthe same fraction of the planktonic community (<50 lm), withthe purpose of further exploring the effect of light availabilityand quality on CH3Hg+ production. We analysed lake water withthe plankton fraction <50 lm in PAR + UV-A and dark (Experiment1), and in PAR (Experiment 2) (Fig. 5a). Mercury methylation in-creased over time (RM ANOVA, p < 0.0001), and the effect of lightwas not statistically significant (p = 0.082). However, the interac-tion between incubation time and light regime was significant(p = 0.041) implying that at certain incubation times (48 and72 h) the differences between light treatments were significant(p < 0.05). When comparing the effect of the light regimes in theresponse at 48 and 72 h, we found that mercury methylation wassignificantly different only between the PAR + UV-A and dark treat-ments (Tukey test, p = 0.044) (Fig. 5a). The fact that PAR alone didnot stimulate as strongly as PAR + UV-A the CH3Hg+ productionmight have been due to the difference in total intensity of lightsupply. Many factors would be involved to generate the differentbehaviour of PAR + UV-A and dark treatments. It is worth mention-ing also that the variance of the PAR treatment is higher than dis-played by the PAR + UV-A and dark conditions (Fig. 5a).
Although no significant difference in 197Hg recoveries was ob-served in Experiment 2 associated to plankton fraction, and similarvalues were obtained also for sterile lake water (Fig. 3), tracerrecoveries in lake water <50 lm incubated in darkness duringExperiment 1 are significantly higher than in sterile lake water indarkness of Experiment 3 (p = 0.014) (Fig. 5b). Therefore, contribu-tions related to plankton should be considered even in darknesswhen photosynthetic activity is absent.
Summarizing, high CH3Hg+ production was measured in lakewater in incubation time trends in a process that is stimulatedby light. Lake water sterilized by filtration showed significant197Hg conversion to CH3
197Hg+, suggesting that the predominantmethylation process may be chemical. However, plankton<50 lm seems to enhance CH3Hg+ production.
4. Discussion
The production of CH3Hg+ from the dissolved Hg2+ implies theavailability of methyl groups in the water. These methyl groupsmay be provided by different organic molecules, which have beenidentified in marine and freshwater systems (Repeta et al., 2002).Aquatic ecosystems are supported by two distinct sources, endo-genous or autochthonous organic carbon, derived respectivelyfrom aquatic primary producer (photosynthesis) and producedin terrestrial ecosystems and transported into waters (Paceet al., 2004).
In the presence of light, phytoplankton production and bacterialutilization of DOM occur simultaneously and both processes aretightly coupled. Allochthonous and autochthonous DOC differ inseveral optical and chemical characteristics (Pérez and Sommaru-ga, 2006). Generally, autochthonous DOM is considered as more la-bile, whereas organic compounds from terrestrial sources are morerecalcitrant to bacterial degradation (Del Giorgio and Davis, 2003).Phytoplankton derived DOM is usually enriched in protein andlabile polysaccharides, whereas terrestrial DOM contains morehumic substances and structural polysaccharides (Benner, 2003).Moreover, the stimulation of heterotrophs by solar irradiation isan indirect result of the production of labile organic matterthrough photosynthesis: more radiation (up to a point), more pho-tosynthesis, and more leaked or excreted labile compounds to sup-port bacterial growth (Cole, 1999). In natural water, DOC
compounds are important absorbers of both visible and UV radia-tion (Morris and Hargreaves, 1997), which in turn can break thelarge macromolecules into smaller ones (Stewart and Wetzel,1982). Small molecules such as fatty acids, acetic acid, pyruvic acid,and citric acid, are released from the DOC and rapidly metabolisedby bacteria. DOC degradation by sunlight may occur below thedepth of UV-light penetration and, in fact, UV-A and PAR play animportant role in creating labile DOC (Lindell, 1996). Moreover,exposure of lake-water DOC to ambient PAR plus UV-A (UV-B ex-cluded) stimulates the bacterial production and the abundanceand growth efficiency of the natural bacteria community (Recheet al., 1998).
Different abiotic reactions can convert Hg2+ to CH3Hg+ in aqua-tic environments, in which methylcobalamin, methyltin com-pounds, or humic matter may play a key role. Among these threecompounds humic matter is the most likely methylating agent,since it is ubiquitous in aquatic environments, it is associated tomercury circulation, complexes mercury, and methylates Hg2+ inmodel studies (Weber, 1993). Water-borne substances such as al-gal and microorganisms secretions, chloride ions, electron media-tors like benzoquinones, may play also an important role in themethylation of mercury in the water column (Siciliano et al.,2005). Nagase et al. (1982) demonstrated that abiotic methylationof divalent mercury can occur through humic substances. In fresh-waters, the oxidised mercury is to a large extent bound to sul-phur groups (thiols) in humic molecules. However, humic matter

584 S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585
contains several different kinds of functional groups and, besidesthe co-ordination to sulphur, mercury more likely presents anadditional co-ordination to neighbouring carboxylic groups (Gård-feldt et al., 2003). In this sense, Weber (1993) emphasized that abi-otic methylation, which includes methylation by chemicalsreleased to the environment by biotic processes, may be a primarydriver of CH3Hg+ production.
The higher CH3Hg+ production observed in lake Moreno waterincluding plankton could be explained considering chemical pro-cesses stimulated by light and mediated by compounds associatedto plankton activity. Organisms belonging to the fraction <50 lmoccur with the highest abundances within the primary producersof lake Moreno (Queimaliños et al., 1999; Pérez et al., 2002), andtheir vital cycles and activity constitute a major source of theautochthonous DOM. Thus, many of these organic compoundsare likely to be in solution in lake Moreno waters and even in watersterilized by filtration, allowing hence CH3Hg+ production. Besides,enzymes or chemicals necessary for CH3Hg+ production may havebeen increased in the sterile water if substantial cell breakage orleakages are produced during the filtration step (Marvin-DiPasqu-ale, M., personal communication). In particular, Weber (1993) as-sessed that methyltin compounds and humic matter arepotential methyl donors for abiotic methylation of Hg2+ in theaquatic environment. Choi et al. (1994) studied Hg2+ methylationin cell extracts when evaluating the role of methylcobalamin inmethylation by Desulfovibrio desulfuricans LS, observing that theenzymatically catalyzed process was predominant over transmeth-ylation by free methylcobalamin. In our work, there is neither theattempt to evaluate the activity of D. desulfuricans LS, nor to assessthe methylcobalamin and methyltin compounds contents thatcould allow the direct comparison of the results.
In lake Moreno, we hypothesize that DOM, even in lower con-centrations than other freshwater systems studied, would playan important role in the methylation process, although this maybe more related with its quality rather than with its quantity. In-deed, Babiarz et al. (2003) stated that it is DOC quality over quan-tity what may account for CH3Hg+ production. Methylatingmolecules are possibly prone to light or bacterial degradationand thus, these processes may be recalled to explain the highmethylation values obtained in lake Moreno waters, even in ab-sence of organisms (sterile treatments). These methylating mole-cules in solution may provide the base line for methylation insterile water in darkness in our experiments. The stimulus pro-vided by PAR and UV-A for DOC degradation may have enhancedthe availability of methyl groups, even in sterile water. The factthat we found lower methylation values and a rapid saturation pat-tern in the dark treatment supports these ideas. In <50 lm treat-ments, increasing photosynthesis and consequently algalexcretion may have provided availability of substrate to bacterialproduction releasing methyl groups, which in turn were availablefor the methylation activity. Concomitantly, light-induced break-age of these organic exudates may also have supplied methyl radi-cals to the pool. The higher CH3Hg+ production in <50 lmtreatments with respect to sterile, both in darkness (Fig. 5b), couldbe explained by DOC bacterial degradation.
Summarizing, our results provide evidence that lake Morenowaters favour CH3Hg+ production, in processes stimulated bylight. Although biotic components certainly contribute to en-hance mercury methylation, water chemistry plays a key rolein this process. We hypothesize that DOM, particularly its quality,could be decisive. The high CH3Hg+ production observed in lakeMoreno waters is particularly significant since, from a bioaccu-mulation perspective, CH3Hg+ production in the water columncould have much higher impact than the sediments or otheraquatic compartments because of the direct linkage to pelagicfood webs.
Acknowledgements
The authors thank Milena Horvat, Mark Marvin-DiPasquale,Markus Meili and Jean Remy Guimaraes for their valuable com-ments. Acknowledgement is also due to Eduardo Scarnichia andthe reactor RA-6 operation staff for their assistance in the radio-tracer production and handling. This work was funded by ProjectPICT 13-13276, Agencia Nacional de Promoción Científica y Tec-nológica, Argentina.
References
Alonso, C., Rocco, V., Barriga, J.P., Battini, M.A., Zagarese, H., 2004. Surface avoidanceby freshwater zooplankton: field evidence on the role of ultraviolet radiation.Limnol. Oceanogr. 49, 225–232.
Arribére, M.A., Campbell, L.M., Kyser, K., Klassen, K., Ribeiro Guevara, S., Bubach,D.F., Vigliano, P., 2005. Aquatic food web transfer of metals in Nahuel HuapiNational Park, Argentina: expected and unique patterns. In: Proceedings of theSeventh Congress of the Society of Environmental Toxicology and Chemistry(SETAC) L.A., Santiago de Chile, Chile, October 16–20, 2005.
Arribére, M., Ribeiro Guevara, S., Bubach, D., Arcagni, M., Vigliano, P., 2008.Selenium and mercury in native and introduced fish species of PatagonianLakes, Argentina. Biol. Trace Elem. Res. doi:10.1007/s12011-007-8059-6.
Babiarz, C.L., Hurley, J.P., Krabbenhoft, D.P., Gilmour, C., Branfireun, B.A., 2003.Application of ultrafiltration and stable isotopic amendments to field studies ofmercury partitioning to filterable carbon in lake water and overland runoff. Sci.Total Environ. 304, 295–303.
Benner, R., 2003. Molecular indicators of bioavailability of dissolved organic matter.In: Findlay, S.E.G., Sinsabaugh, R.L. (Eds.), Aquatic Ecosystems: Interactivity ofDissolved Organic Matter. Academic Press, University of Toledo, OH, USA.
Boening, D.W., 2000. Ecological effects, transport, and fate of mercury: a generalreview. Chemosphere 40, 1335–1351.
Callieri, C., Modenutti, B., Queimaliños, C., Bertoni, R., Balseiro, E., 2007. Productionand biomass of picophytoplankton and larger autotrophs in Andeanultraoligotrophic lakes: differences in light harvesting efficiency in deeplayers. Aquat. Ecol. 41, 511–523.
Choi, S.C., Chase Jr, T., Bartha, R., 1994. Enzymatic catalysis of mercury methylationby Desulfovibrio desulfuricans LS. Appl. Environ. Microb. 60, 1342–1346.
Cole, J.J., 1999. Aquatic microbiology for ecosystem scientists: new and recycledparadigms in ecological microbiology. Ecosystems 2, 215–225.
Cossa, D., Coquery, M., Gobeil, C., Martin, M., 1996. Mercury fluxes at the oceanmargins. In: Baeyens, W., Ebinghaus, R., Vasiliev, O. (Eds.), Global and RegionalMercury Cycles: Sources, Fluxes and Mass Balances. NATO ASI Series 2.Environment, vol. 2. Kluwer Academic Publishers.
del Giorgio, P.A., Davis, J., 2003. Patterns in dissolved organic matter lability andconsumption across aquatic ecosystems. In: Findlay, D.L., Sinsabaugh, R.L.(Eds.), Aquatic Ecosystems: Interactivity of Dissolved Organic Matter. AcademicPress, University of Toledo, OH, USA.
Díaz, M., Pedrozo, F., Reynolds, C., Temporetti, P., 2007. Chemical composition andthe nitrogen-regulated trophic state of Patagonian lakes. Limnologica 37, 17–27.
Duffy, L.K., Scofield, E., Rodgers, T., Patton, M., Bowyer, R.T., 1999. Comparativebaseline levels of mercury, Hsp 70 and Hsp 60 in subsistence fish from theYukon-Kuskokwim delta region of Alaska. Comput. Biochem. Phys. C 124, 181–186.
Fostier, A.H., Forti, M.C., Guimaraes, J.R.D., Melfi, A.J., Boulet, R., Espirito Santo, C.M.,Krug, F.J., 2000. Mercury fluxes in a natural forested Amazonian catchment(Serra do Navio, Amapa State, Brazil). Sci. Total Environ. 260, 201–211.
Gårdfeldt, K., Munthe, J., Stromberg, D., Lindqvist, O., 2003. A kinetic study on theabiotic methylation of divalent mercury in the aqueous phase. Sci. TotalEnviron. 304, 127–136.
Guimaraes, J.R.D., Meili, M., Lars, D.H., de Castro e Silva, E., Roulet, M., NarvaezMauro, J.B., Alves de Lemos, R., 2000. Mercury net methylation in five tropicalflood plain regions of Brazil: high in the root zone of floating macrophyte matsbut low in surface sediments and flooded soils. Sci. Total Environ. 261, 99–107.
Leermakers, M., Meuleman, C., Baeyens, W., 1996. Mercury distribution and fluxesin Lake Baikal. In: Baeyens, W., Ebinghaus, R., Vasiliev, O. (Eds.), Global andRegional Mercury Cycles: Sources, Fluxes and Mass Balances. NATO ASI Series 2.Environment, vol. 2. Kluwer Academic Publishers.
Lindell, M., 1996. Effects of sunlight on organic matter and bacteria in lakes. Ph.D.Thesis, Lund University, Sweden.
Marvin-DiPasquale, M.C., Agee, J.L., Bouse, R.M., Jaffe, B.E., 2003. Microbial cycling ofin contaminated pelagic mercury and wetland sediments of San Pablo Bay,California. Environ. Geol. 43, 260–267.
Mauro, J.B.N., Guimaraes, J.R.D., Hintelmann, H., Watras, C.J., Haack, E.A., Coelho-Souza, S.A., 2002. Mercury methylation in macrophytes, periphyton, and water– comparative studies with stable and radio-mercury additions. Anal. Bioanal.Chem. 374, 983–989.
Meili, M., 1997. Mercury in lakes and rivers. In: Sigel, A., Sigel, H. (Eds.), Metal Ionsin Biological Systems, Mercury and its Effect on Environment and Biology, vol.34. Marcel Dekker Inc., New York, pp. 21–51 (Chapter 2).

S. Ribeiro Guevara et al. / Chemosphere 72 (2008) 578–585 585
Monperrus, M., Tessier, E., Point, D., Vidimova, K., Amouroux, D., Guyoneaud, R.,Leynaert, A., Grall, J., Chauvaud, L., Thouzeau, G., Donard, O.F.X., 2007. Thebiogeochemistry of mercury at the sediment water interface in the ThauLagoon. 2: Evaluation of mercury methylation potential in both surfacesediment and the water column. Estuar. Coast. Shelf Sci. 72, 485–496.
Morris, D.P., Hargreaves, B.R., 1997. The role of photochemical degradation ofdissolved organic carbon in regulating the UV transparency of three lakes on thePocono Plateau. Limnol. Oceanogr. 42, 239–249.
Nagase, H., Ose, O., Sato, T., Ishikawa, T., 1982. Methylation of mercury by humicsubstances in an aquatic environment. Sci. Total Environ. 25, 133–142.
Pace, M.L., Cole, J.J., Carpenter, S.R., Kitchell, J.F., Hodgson, J.R., Van de Bogert, M.C.,Bade, D.L., Kritzberg, E.S., Bastviken, D., 2004. Whole-lake carbon-13 additionsreveal terrestrial support of aquatic food webs. Nature 427, 240–243.
Pérez, G.L., Queimaliños, C.P., Modenutti, B.E., 2002. Light climate at the deepchlorophyll maxima in North Patagonian Andean lakes (Argentina). J. PlanktonRes. 24, 591–599.
Pérez, M.T., Sommaruga, R., 2006. Differential effect of algal- and soil-deriveddissolved organic matter on alpine lake bacterial community composition andactivity. Limnol. Oceanogr. 51, 2527–2537.
Pérez Catán, S., Ribeiro Guevara, S., Marvin-DiPasquale, M., Magnavacca, C., Cohen,I.M., Arribére, M., 2007. Methodological considerations regarding the use ofinorganic 197Hg(II) radiotracer to assess mercury methylation potential rates inlake sediment. Appl. Radiat. Isotopes 65, 987–994.
Queimaliños, C.P., Modenutti, B.E., Balseiro, G.E., 1999. Symbiotic association of theciliate Ophrydium naumanni with Chlorella causing a deep chlorophyll amaximum in an oligotrophic South Andes lake. J. Plankton Res. 21, 167–178.
Queimaliños, C.P., Pérez, G., Modenutti, B.E., 2002. Summer populationdevelopment and diurnal vertical distribution of dinoflagellates in anultraoligotrophic Andean lake (Patagonia, Argentina). Algol. Stud. 107, 117–129.
Rask, M., Metsala, R.T., Salonen, K., 1994. Mercury in the food chains of a smallpolyhumic forest lake in Southern Finland. In: Watras, C.J., Huckabee, J.W.(Eds.), Mercury Pollution – Integration and Synthesis. CRC Press Inc./LewisPublishers, Boca Raton.
Reche, I., Pace, M.L., Cole, J.J., 1998. Interactions of photobleaching and inorganicnutrients in determining bacterial growth on colored dissolved organic carbon.Microbial. Ecol. 36, 270–280.
Repeta, D.J., Quan, T.M., Aluwihare, L.I., Accardi, A., 2002. Chemical characterizationof high molecular weight dissolved organic matter in fresh and marine waters.Geochim. Cosmochim. Acta 66, 955–962.
Ribeiro Guevara, S., Rizzo, A., Sánchez, R., Arribére, M., 2005. Heavy metal inputs inNorthern Patagonia lakes from short sediment cores analysis. J. Radioanal. Nucl.Chem. 265, 481–493.
Ribeiro Guevara, S., Zizek, S., Repinc, U., Pérez Catán, S., Jacimovic, R., Horvat, M.,2007. Novel methodology for the study of mercury methylation and reductionin sediments and water using 197Hg radiotracer. Anal. Bioanal. Chem. 387,2185–2197.
Sellers, P., Kelly, C.A., Rudd, J.W.M., MacHutchon, A.R., 1996. Photodegradation ofmethyl mercury in lakes. Nature 380, 694–697.
Siciliano, S.D., ODriscoll, N.J., Tordon, R., Hill, J., Beauchamp, S., Lea, D.R., 2005.Abiotic production of methylmercury by solar radiation. Environ. Sci. Technol.39, 1071–1077.
Souza, M.S., Modenutti, B.E., Balseiro, E.G., 2007. Antioxidant defences in planktoniccrustaceans exposed to different underwater light irradiances in Andean lakes.Water Air Soil Pollut. 183, 49–57.
Stewart, A.J., Wetzel, R.G., 1982. Influence of dissolved humic materials on carbonassimilation and alkaline phosphatase activity in natural algal-bacterialassemblages. Freshwater Biol. 12, 369–380.
Ullrich, S.M., Tanton, T.W., Abdrashitova, S.A., 2001. Mercury in the aquaticenvironment: a review of factors affecting methylation. Crit. Rev. Environ. Sci.Tech. 31, 241–293.
Utermöhl, H., 1958. Zur vervollkommung der quantitativen phytoplankton-methodik. Mitt. Intemat. Verein. Limnol. 9, 1–38.
Watras, C.J., Bloom, N.S., 1994. The vertical distribution of mercury species inWisconsin lakes: accumulation in plankton layers. In: Watras, C.J., Huckabee,J.W. (Eds.), Mercury Pollution – Integration and Synthesis. CRC Press Inc./LewisPublishers, Boca Raton.
Watras, C.J., Backa, R.C., Halvorsena, S., Hudson, R.J.M., Morrisona, K.A., Wente, S.P.,1998. Bioaccumulation of mercury in pelagic freshwater food webs. Sci. TotalEnviron. 219, 183–208.
Weber, J.H., 1993. Review of possible paths for abiotic methylation of mercury (II) inthe aquatic environment. Chemosphere 26, 2063–2077.
Zizek, S., Ribeiro Guevara, S., Horvat, M., 2008. Validation of methodology fordetermination of the mercury methylation potential in sediments usingradiotracers. Anal. Bioanal. Chem. doi:10.1007/s00216-008-1968-1.




















