
Methane-derived carbonate build-ups and associated microbial communities at cold seeps on the lower...
14
Facies (2005) 51: 66–79 DOI 10.1007/s10347-005-0059-4 ORIGINAL ARTICLE Joachim Reitner · J¨ orn Peckmann · Andreas Reimer · Gabriela Schumann · Volker Thiel Methane-derived carbonate build-ups and associated microbial communities at cold seeps on the lower Crimean shelf (Black Sea) Received: 16 December 2004 / Accepted: 8 March 2005 / Published online: 2 July 2005 C Springer-Verlag 2005 Abstract In the euxinic waters of the NW’ Black Sea shelf, tower-like carbonate build-ups up to several metres in height grow at sites of cold methane seepage. These structures are part of an unique microbial ecosystem that shows a considerable biodiversity and a remarkable degree of organization. The accretion of the build-ups is promoted by the growth of centimetre-sized, methane-filled spheres constructed by calcifying microbial mats. Progressive min- eralization of these spheres involves the early precipitation of strongly luminescent high-Mg-calcite rich in iron sul- phides, and closely interfingered aragonite phases that fi- nally create the stable (mega-) thrombolithic fabric of the towers. Within the microbial mats, microorganisms occur in distinctive spatial arrangements. Major players among the microbial consortia are the archaea groups ANME-1 and ANME-2, Crenarchaeota, and sulphate-reducing bac- teria (SRB) of the Desulfosarcina/Desulfobacterium group. The intracellular precipitation of iron sulphides (greigite) by some of these bacteria, growing in close association with ANME-2, suggests iron cycling as an additional bio- geochemical pathway involved in the anaerobic oxidation of methane (AOM). Keywords Black Sea . Anaerobic oxidation of methane (AOM) . Magnetotactic bacteria . Carbonates . Cold seeps . FISH . Archaea J. Reitner () · A. Reimer · G. Schumann · V. Thiel Geowissenschaftliches Zentrum der Universit¨ at G ¨ ottingen GZG, Goldschmidtstraße 3, 37077 G¨ ottingen, Germany e-mail: [email protected] Tel.: +49-551-397950 Fax: +49-551-397918 J. Peckmann Forschungszentrum Ozeanr¨ ander, Universit¨ at Bremen, Postfach 330440, 28334 Bremen, Germany Introduction Most of the methane seeping upward in the marine sedi- mentary column is intercepted biologically by the anaero- bic oxidation of methane (AOM) and is thus prevented from reaching the atmosphere. There is now a consensus that AOM can be mediated by consortia of methane-oxidizing archaea and sulphate-reducing bacteria (SRB), although the precise pathway(s) and the roles of the respective mi- crobial components are not yet fully elucidated (Orphan et al. 2001; Valentine 2002 for a review; Nauhaus et al. 2004). AOM increases carbonate alkalinity according to the overall reaction: CH 4 + SO 2− 4 → HCO − 3 + HS − + H 2 O Therefore, calcium carbonate deposits are typically found as remnants of contemporary and ancient methane seepage (Goedert and Squires 1990; Peckmann et al. 1999; Greinert et al. 2001; Aloisi et al. 2002; Campbell et al. 2002; Lein et al. 2002; Peckmann and Thiel 2004). In the NW’ Black Sea, methane-seepage occurs in several areas along the shelf/slope SW of the Crimean Peninsula (Polikarpov et al. 1989; Egorov et al. 1998), and various morphologies of associated carbonates have been reported (Ivanov et al. 1991; Luth et al. 1999; Peckmann et al. 2001; Lein et al. 2002; Michaelis et al. 2002; Reitner et al. in press). These precipitates can roughly be divided into two groups: one forms below the sediment–water in- terface, as porous plates of carbonate-cemented sediments, irregular crusts, smooth-shaped lenticular concretions, flat pancake-shaped deposits, and large (metre-sized) tabular constructions. A different situation is observed below the chemocline, where the carbonates may grow free into the anoxic water column, forming tall build-ups, and in some places, impressive tower-like constructions that may reach several metres in height (Peckmann et al. 2001; Lein et al. 2002; Michaelis et al. 2002). Microbial mats are typically associated with the Black Sea seep carbonates and have been investigated by several studies. Laboratory experiments with mat samples from
-
Upload
uni-goettingen -
Category
Documents
-
view
0 -
download
0
Transcript of Methane-derived carbonate build-ups and associated microbial communities at cold seeps on the lower...

Facies (2005) 51: 66–79DOI 10.1007/s10347-005-0059-4
ORIGINAL ARTICLE
Joachim Reitner · Jorn Peckmann · Andreas Reimer ·Gabriela Schumann · Volker Thiel
Methane-derived carbonate build-ups and associated microbialcommunities at cold seeps on the lower Crimean shelf (Black Sea)
Received: 16 December 2004 / Accepted: 8 March 2005 / Published online: 2 July 2005C© Springer-Verlag 2005
Abstract In the euxinic waters of the NW’ Black Seashelf, tower-like carbonate build-ups up to several metresin height grow at sites of cold methane seepage. Thesestructures are part of an unique microbial ecosystem thatshows a considerable biodiversity and a remarkable degreeof organization. The accretion of the build-ups is promotedby the growth of centimetre-sized, methane-filled spheresconstructed by calcifying microbial mats. Progressive min-eralization of these spheres involves the early precipitationof strongly luminescent high-Mg-calcite rich in iron sul-phides, and closely interfingered aragonite phases that fi-nally create the stable (mega-) thrombolithic fabric of thetowers. Within the microbial mats, microorganisms occurin distinctive spatial arrangements. Major players amongthe microbial consortia are the archaea groups ANME-1and ANME-2, Crenarchaeota, and sulphate-reducing bac-teria (SRB) of the Desulfosarcina/Desulfobacterium group.The intracellular precipitation of iron sulphides (greigite)by some of these bacteria, growing in close associationwith ANME-2, suggests iron cycling as an additional bio-geochemical pathway involved in the anaerobic oxidationof methane (AOM).
Keywords Black Sea . Anaerobic oxidation of methane(AOM) . Magnetotactic bacteria . Carbonates . Coldseeps . FISH . Archaea
J. Reitner (�) · A. Reimer · G. Schumann · V. ThielGeowissenschaftliches Zentrum der Universitat Gottingen GZG,Goldschmidtstraße 3,37077 Gottingen, Germanye-mail: [email protected].: +49-551-397950Fax: +49-551-397918
J. PeckmannForschungszentrum Ozeanrander, Universitat Bremen,Postfach 330440,28334 Bremen, Germany
Introduction
Most of the methane seeping upward in the marine sedi-mentary column is intercepted biologically by the anaero-bic oxidation of methane (AOM) and is thus prevented fromreaching the atmosphere. There is now a consensus thatAOM can be mediated by consortia of methane-oxidizingarchaea and sulphate-reducing bacteria (SRB), althoughthe precise pathway(s) and the roles of the respective mi-crobial components are not yet fully elucidated (Orphanet al. 2001; Valentine 2002 for a review; Nauhaus et al.2004). AOM increases carbonate alkalinity according tothe overall reaction:
CH4 + SO2−4 → HCO−
3 + HS− + H2O
Therefore, calcium carbonate deposits are typically foundas remnants of contemporary and ancient methane seepage(Goedert and Squires 1990; Peckmann et al. 1999; Greinertet al. 2001; Aloisi et al. 2002; Campbell et al. 2002; Leinet al. 2002; Peckmann and Thiel 2004).
In the NW’ Black Sea, methane-seepage occurs inseveral areas along the shelf/slope SW of the CrimeanPeninsula (Polikarpov et al. 1989; Egorov et al. 1998), andvarious morphologies of associated carbonates have beenreported (Ivanov et al. 1991; Luth et al. 1999; Peckmannet al. 2001; Lein et al. 2002; Michaelis et al. 2002; Reitneret al. in press). These precipitates can roughly be dividedinto two groups: one forms below the sediment–water in-terface, as porous plates of carbonate-cemented sediments,irregular crusts, smooth-shaped lenticular concretions, flatpancake-shaped deposits, and large (metre-sized) tabularconstructions. A different situation is observed below thechemocline, where the carbonates may grow free into theanoxic water column, forming tall build-ups, and in someplaces, impressive tower-like constructions that may reachseveral metres in height (Peckmann et al. 2001; Lein et al.2002; Michaelis et al. 2002).
Microbial mats are typically associated with the BlackSea seep carbonates and have been investigated by severalstudies. Laboratory experiments with mat samples from

67
the anoxic zone build-ups have shown their capacityto perform methane oxidation and sulphate-reduction(Pimenov et al. 1997), which occur at a 1:1 stoichiometry(Michaelis et al. 2002). However, it has also been inferredfrom isotope data that bicarbonate may be used as anelectron acceptor by the anaerobic methane oxidizers (Leinet al. 2002). Recent work showed that AOM in anaerobicenrichment cultures from Black Sea sediments and watersamples was strongly stimulated by addition of acetate,methylamine and/or a mixture of acetate, formate, andlactate (Gal’chenko 2004). Although the exact metabolicpathways of AOM in the Black Sea environment yetremain unknown, an in situ transfer of methane carboninto the biomass of both, Archaea and SRB was inferredfrom organic matter and lipid biomarkers highly depletedin13C (Thiel et al. 2001; Michaelis et al. 2002; Blumenberget al. 2004; Gal’chenko 2004; Reitner et al. in press). Inaddition, Kruger et al. (2003) discovered a conspicuousnickel protein, similar to the terminal enzyme of archaealmethanogenesis, which is most likely involved in AOM.
Microscopy and culture independent molecular workidentified archaea of the ANME-1 (anaerobic methane ox-idizers) and ANME-2 groups, Crenarchaeota, and SRBof the Desulfosarcinales/Desulfobacterium (DSS) groupin mats associated with the carbonate build-ups (Pimenovet al. 1997; Michaelis et al. 2002; Tourova et al. 2002;Blumenberg et al. 2004). A mat sampled from an intrasedi-mentary carbonate concretion from the same area revealedANME-1 archaeal group that co-occurred with clusters ofSRB of the DSS group (Reitner et al. in press). The vibrio-like SRB were morphologically different from the coc-coid forms described by Michaelis et al. (2002), and theywere found to produce intracellular iron sulphide magne-tosomes, suggesting the involvement of iron cycling in themetabolism of the microbial consortia.
The range of patterns reported implicates considerableheterogeneity in the nature, metabolism, and spatial distri-bution of the methane-related microbiota. Research interesthas thus increasingly been concentrating on the differentmicrobial facies present in the Black Sea seep ecosystems.A recent study by Blumenberg et al. (2004) on the distribu-tions of lipid biomarkers gave important clues on differentmicrobial communities and their molecular and isotopicsignatures. Using materials from the same setting, we herepresent a survey on the principal petrological and biologicalfeatures using various microscopic techniques. The maingoal of this study is to establish (1) a better understandingof the complex petrology of methane-related carbonatesand the mechanisms of their formation, and (2) a compre-hensive visualization of the as-yet only rudimentarily un-derstood anatomy and heterogeneity of the seep-associatedmicrobial consortia.
Materials and methods
Samples were taken from the methane seep area onthe lower Crimean shelf explored during a German–Russian–Ukrainian joint expedition (GHOSTDABS) in
Fig. 1 Location of the methane seepage area (asterisk) on the lowerCrimean shelf explored during the German–Russian–Ukrainian jointexpedition in 2001. Samples were taken from the GHOSTDABS-field (44◦46.510′N, 31◦59.570′E) at about 230 m water depth
2001 (Fig. 1). For sampling, the manned submersibleJago was used from aboard the Russian R/V “ProfessorLogachev”. The samples described here were taken fromthe GHOSTDABS-field (44◦46.510′N, 31◦59.570′E) atabout 230 m water depth. Samples were fixed with 4%buffered formol, stored in 70% ethanol or in 50% phosphatebuffered saline (PBS). Samples for transmission electronmicroscopy (TEM), conventional scanning electron mi-croscopy (SEM), and field emission SEM (FE-SEM) werefixed with 4% buffered glutardialdehyde and post fixedwith 2% OsO4. To avoid drying artefacts, glutardialdehyde-fixed samples were dried with Peldri II (Pelco, USA) andhexamethyldisilazane (HMDS; Polysciences, USA), bothchemical alternatives to critical-point drying (Brown 1993).Thin-sections of microbial mats were cut using a Leicahardpart microtome. Image stacks with a Z-spacing of0.5 or 0.25 µm were obtained using a piezo-mover(Physik Instrumente GmbH & Co, Waldbronn) attachedto a Zeiss Axioplan microscope equipped with a Zeiss“Plan-Apochromat” 63×-objective (NA=1.4). Imageprocessing was carried out using the Metamorph Imagingsoftware (Universal Imaging Corporation, West Chester,PA) and the EPRTM deconvolution software (Scanalytics,Billerica, MA). Paraffin sections of decalcified sampleswere stained with various histochemicals, namely tolu-idine blue O (RNA, oligodeoxynucleotides, proteins,and glycosaminoglycans), 4′,6-diamidino-2-phenylindole(DAPI; double stranded DNA). For details of the stainingprocedures, see Romeis (1989), Manz et al. (2000), andHoffmann et al. (2003). Prior to fluorescence in situhybridization (FISH, explanation in Manz et al. 2000),the samples were stained with DAPI to control the cell

68
Fig. 2 Morphology of thecarbonate build-ups. A Drawingof a large, active tower. Gasemanates in continuous bubblestreams from openings in thespherical microbial mats at thetips of the structure (artwork:Conny Kaubisch, GZG). BMicrobial mat spheres growingat a methane outflow on a largecarbonate plate that has beenwinnowed by strong bottomcurrents. Note the fur-likeappearance of the black mats. CA juvenile build-up (ca. 25 cmhigh), consisting of agrape-shaped agglomeration ofbudding microbial mat spheres.Note the sediment cover of themats and the current-induced,funnel-like depression aroundthe base of the build-up. Thebuild-up grows on a bottomplate of carbonate-cementedsediment (see E). D A group ofgrowing, tower-like carbonatebuild-ups in an area of intensegas seepage (235 m waterdepth). E A subrecent towerdevoid of microbial mats. Notethe bottom plate ofcarbonate-cemented sedimentthat had rooted the build-up inthe sediment
numbers and vitality of the microbial communities. TheDAPI control-stain is important to avoid misinterpretationsand to recognize non-specific binding artefacts of theoligonucleotide probes. Each section was hybridized withtwo differently labelled oligonucleotide probes.
The probes were purchased 5′-labelled with the indo-carbocyanine dye Cy3 (Amersham Pharmacia Biotech)and Oregon Green (Molecular Probes) from Biome-tra (Gottingen, Germany), and Metabion (Planegg-Martinsried, Germany) were stored in TE buffer (10 mMTris/1 mM EDTA, pH 7.5) at −20◦C. In order to mini-mize mismatching of the oligonucleotide probes, severalsections of each mat sample were hybridized using variedbuffer stringencies (formamide concentrations) from 35 to
60% at set hybridization and washing temperatures. Whenusing bacterial together with archaeal probes, best resultswere obtained with a hybridization solution of 40% strin-gency. This solution was prepared by mixing 400 µl doublydistilled water with 400 µl formamide, 180 µl 5M NaCl,20 µl 1 M Tris (pH 8), and 1 µl 10% SDS. Fluorescentlylabelled oligonucleotides were added at a concentration of1 ng per µl of hybridization solution.
The following 16S rRNA oligonucleotide probes forfluorescence in situ hybridization (FISH) were applied:EUB338 (Amann et al. 1990a), ARCH915 (Amann et al.1990b), DSS658 (Manz et al. 1998), ANME-1 (Hinrichset al. 1999), EelMS932 (ANME-2, Boetius et al. 2000),Cren499 (Burggraf et al. 1994). Uncoated samples were

69
investigated by FE-SEM using a LEO 1530 Gemini in-strument at less than 1 kV. Energy dispersive X-ray spec-trometry (EDX) was performed on carbon-coated samplesusing the same instrument operated at 15 kV. In order toavoid dehydration artefacts and collapse of organic struc-tures, some samples were investigated fully hydrated usingan Oxford-Cryosystem attached to the FE-SEM. TEM in-vestigations on thin-sections were carried out with a ZeissEM 10 instrument at 60–80 kV. Carbonate samples forstable carbon isotope analysis were prepared using a hand-held microdrill. CO2 was generated by reaction with or-thophosphoric acid and analysed with a Finnigan MAT 252mass spectrometer at the University of Erlangen (Dr. M.Joachimski). The δ13C values are reported relative to the V-PDB standard (precision of values is ±0.05‰). Cathodo-luminescence investigations were carried out with a Citl8200 MK3A cold cathode mounted on a Zeiss Axiolabmicroscope. Micrographs were recorded at 20 kV voltageusing a cooled SPOT-CCD camera. The SPOT’s CCD chiphas a true resolution of 1315×1033 pixels. By exposingthe CCD three times (for red, green and blue, respectively),each colour was accurately recorded on every CCD pixel.Mapping of key elements (Mg, Sr, Mn) was carried outwith a JEOL JXA 8900RL electron microprobe housed inthe Center for Geosciences at the University of Gottingen(Department of Geochemistry, Dr. A. Kronz).
Results
Structure and formation of the carbonate towers
The metre-sized carbonate towers develop where gaseousmethane discharges into the water column (Fig. 2). Bottomwater currents frequently create a moat surrounding thebases of the build-ups (Fig. 2c). In places, the sedimentswere completely eroded from the roots of the towers, exca-vating large disc-shaped basal plates that anchor the towersin the sediment (Fig. 2e).
The initial growth of the towers starts with spherical,centimetre-sized microbial mat constructions displaying aunique architecture (Fig. 2b–c). The spheres are hollow,and are pervaded by gas. Openings control the gas pressure,releasing it either as a continuous bubble flow, or in moreor less regular intervals. The outer surface of the spheresis covered by a dark-grey to black-coloured microbial mat(referred to as ‘black mat’ hereafter). These few mm thickmats have greatly enlarged their surface by peculiar fur-like villi that trap sedimentary matter to a varying extent(Fig. 2c). Towards the interior of the spheres, the black matis underlain by a pink to orange-coloured layer of similarthickness (referred to as ‘pink mat’ in the following, seealso Fig. 3).
The spheres initially exhibit a soft, jelly-like consistency,and are easily compressible. The metabolism of the micro-bial community induces the precipitation of calcium car-bonate, a process that is likely stimulated by an increase incarbonate alkalinity upon sulphate-dependent AOM. Thecalcification process results in the formation of delicate
Fig. 3 A large tower retrieved from the GHOSTDABS field at 235 mwater depth (note researcher’s legs for scale). The cross sectionthrough the tilted tower shows a large-scale zonation of differentmicrobial mat types (see text)
walls that further stabilize the spheres. Small openings mayconnect neighbouring spheres, enabling gas supply and gasrelease (Fig. 4a).
The gradual lithification of the spheres involves a limita-tion of their maximum growth size to approximately 10 cm.Mature spheres generate buds from which juvenile spheresemerge. This results in the formation of grape-shaped ag-gregates (Figs. 2 and 4). Continuous budding and ongo-ing calcification finally create the tall tower-like build-ups.They typically show growing spheres at the tips, and a lessstructured surface with a massive, several centimetres thickcover of microbial mats on the flanks (Fig. 3). Here, as ob-served for the spheres, black mats are found at the outersea water contact, and are underlain by the pink mat.
As a result of complex calcification patterns, aragoniteand high-Mg-calcite phases coexist on a small spatial scaleand typically show a close interfingering, resulting in ahighly cavernous, not layered, and blotchy internal fab-ric (Fig. 4e). This fabric may, in places, trace the initialspherical constructions that once protruded into the watercolumn. Within the towers, no distinct central channel ex-ists. Instead, still abundant vugs and voids channel gas andfluids from the base towards the tips. These open spacesare lined by thin microbial mats differing from the blackand pink varieties by a green to brownish colour (Fig. 3,referred to as ‘green mat’ hereafter).
Early calcification within the black mat produces 100–200 µm-sized hemispherical to spherical crystal aggre-gates of fibrous high-Mg-calcite (Fig. 5b–c, e). Featuresunique to the high-Mg-calcite are a red-pink autofluores-cence (Figs. 5c, 6c), and a bright red-orange cathodolumi-nescence (Fig. 6b). The precipitation of the high-Mg-calcitestarts within accumulations of iron sulphides around mi-crobial colonies within the black mat. These iron sulphidesget enclosed in the high-Mg-calcite as strings and clustersof small framboidal aggregates (Fig. 4b–d). The calcifica-tion process first leads to the formation of high-Mg-calciteseams around the microbial colonies (Fig. 6b–c). This prob-ably causes the microbial activity to cease and is followedby permineralisation of the decaying colonies in the deeper

70
Fig. 4 Petrofabrics. AThin-section of microbial matspheres in an early stage ofmineralization. Delicate wallsconsist of iron sulphide andcarbonate precipitates. Arrowsdenote openings by which gas isreleased from, or exchangedbetween, individual spheres. BMagnification of A showingclose interfingering of carbonatephases. Iron sulphides (black)predominantly appear in earlyhigh-Mg-calcite. C Thin-sectionof a mature sphere showing anadvanced lithification stage.Further growth is inhibited dueto the development of asuccession of carbonatephases/cements. D Detail of C.Numbers denote (2) earlyhigh-Mg-calcite enriched in (1)iron sulphides, (3) earlyaragonite, (4) isopachousaragonite rim cement, (5) latemicritic calcite. E Cross-sectionthrough a tower-like build-up.Ongoing calcification creates amega-thrombolitic petrofabricthat may, in places, outline theoriginal spherical constructions
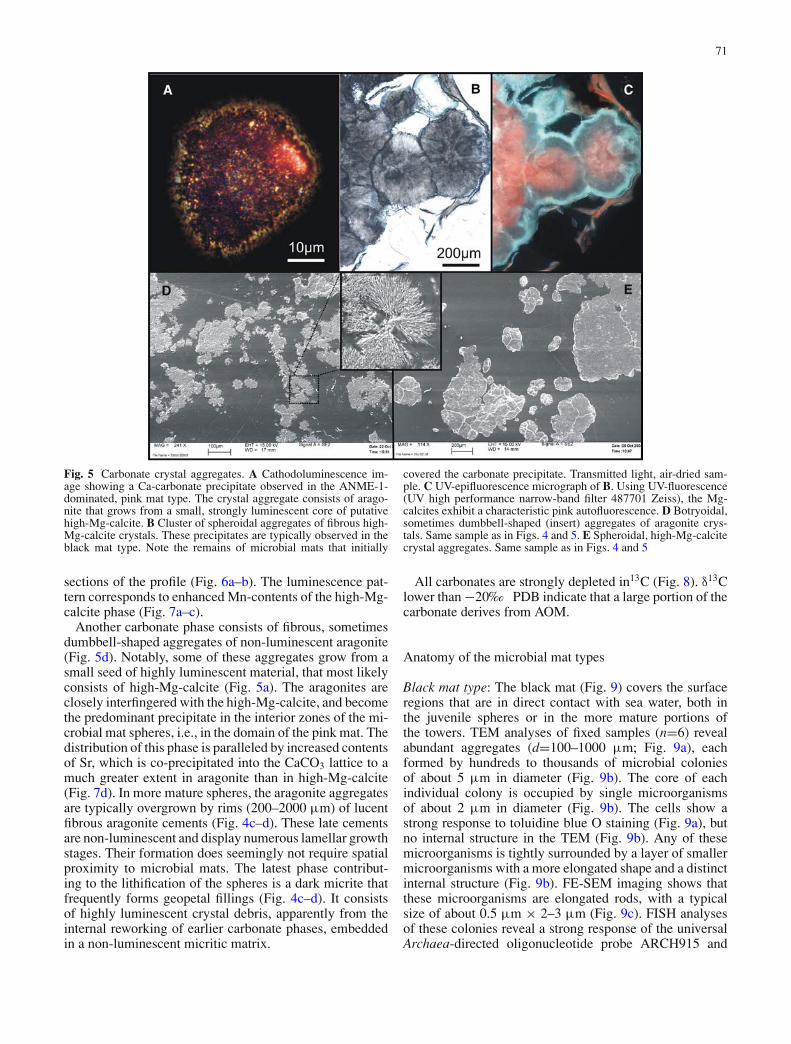
71
Fig. 5 Carbonate crystal aggregates. A Cathodoluminescence im-age showing a Ca-carbonate precipitate observed in the ANME-1-dominated, pink mat type. The crystal aggregate consists of arago-nite that grows from a small, strongly luminescent core of putativehigh-Mg-calcite. B Cluster of spheroidal aggregates of fibrous high-Mg-calcite crystals. These precipitates are typically observed in theblack mat type. Note the remains of microbial mats that initially
covered the carbonate precipitate. Transmitted light, air-dried sam-ple. C UV-epifluorescence micrograph of B. Using UV-fluorescence(UV high performance narrow-band filter 487701 Zeiss), the Mg-calcites exhibit a characteristic pink autofluorescence. D Botryoidal,sometimes dumbbell-shaped (insert) aggregates of aragonite crys-tals. Same sample as in Figs. 4 and 5. E Spheroidal, high-Mg-calcitecrystal aggregates. Same sample as in Figs. 4 and 5
sections of the profile (Fig. 6a–b). The luminescence pat-tern corresponds to enhanced Mn-contents of the high-Mg-calcite phase (Fig. 7a–c).
Another carbonate phase consists of fibrous, sometimesdumbbell-shaped aggregates of non-luminescent aragonite(Fig. 5d). Notably, some of these aggregates grow from asmall seed of highly luminescent material, that most likelyconsists of high-Mg-calcite (Fig. 5a). The aragonites areclosely interfingered with the high-Mg-calcite, and becomethe predominant precipitate in the interior zones of the mi-crobial mat spheres, i.e., in the domain of the pink mat. Thedistribution of this phase is paralleled by increased contentsof Sr, which is co-precipitated into the CaCO3 lattice to amuch greater extent in aragonite than in high-Mg-calcite(Fig. 7d). In more mature spheres, the aragonite aggregatesare typically overgrown by rims (200–2000 µm) of lucentfibrous aragonite cements (Fig. 4c–d). These late cementsare non-luminescent and display numerous lamellar growthstages. Their formation does seemingly not require spatialproximity to microbial mats. The latest phase contribut-ing to the lithification of the spheres is a dark micrite thatfrequently forms geopetal fillings (Fig. 4c–d). It consistsof highly luminescent crystal debris, apparently from theinternal reworking of earlier carbonate phases, embeddedin a non-luminescent micritic matrix.
All carbonates are strongly depleted in13C (Fig. 8). δ13Clower than −20‰ PDB indicate that a large portion of thecarbonate derives from AOM.
Anatomy of the microbial mat types
Black mat type: The black mat (Fig. 9) covers the surfaceregions that are in direct contact with sea water, both inthe juvenile spheres or in the more mature portions ofthe towers. TEM analyses of fixed samples (n=6) revealabundant aggregates (d=100–1000 µm; Fig. 9a), eachformed by hundreds to thousands of microbial coloniesof about 5 µm in diameter (Fig. 9b). The core of eachindividual colony is occupied by single microorganismsof about 2 µm in diameter (Fig. 9b). The cells show astrong response to toluidine blue O staining (Fig. 9a), butno internal structure in the TEM (Fig. 9b). Any of thesemicroorganisms is tightly surrounded by a layer of smallermicroorganisms with a more elongated shape and a distinctinternal structure (Fig. 9b). FE-SEM imaging shows thatthese microorganisms are elongated rods, with a typicalsize of about 0.5 µm × 2–3 µm (Fig. 9c). FISH analysesof these colonies reveal a strong response of the universalArchaea-directed oligonucleotide probe ARCH915 and

72
Fig. 6 Carbonate formation and fabrics. A Semi-thin-sectionthrough the wall of a microbial mat sphere at an early stage ofmineralization (compare Fig. 4a). The section encompasses the outerportions harbouring consortia of ANME-2 and magnetotactic SRB,and the inner regions with abundant ANME-1. Microbial biomass (in-cluding EPS) is stained blue. Mineralized portions appear dark. Notestrings of iron sulphide surrounding ANME-2-dominated colonies,and close interfingering of brownish high-Mg-calcite and (darker)aragonite phases at the transition towards the ANME-1-dominated in-terior facies. Transmitted light, toluidine blue O staining. B Cathodo-luminescence image showing a detail of A. This photomicrographdocuments the initial stages of seep carbonate formation, starting with
accumulations of iron sulphide (black) forming around the ANME-2-dominated colonies. The precipitation of strongly luminescent high-Mg-calcite emerges from these iron sulphide strings and enclosesthe colonies. This probably causes the microbial activity to cease,followed by permineralization of the decaying colonies in the deepersections of the profile. C UV-epifluorescence image showing a de-tail of B. Microbial biomass appears colourless. The high-Mg-calciteseams forming around the colonies of ANME-2 and magnetotacticSRB typically show a pink autofluorescence. This photomicrographillustrates the incorporation of strings of iron sulphide aggregatesinto the newly forming high-Mg-calcite
EelMS932 (ANME-2) for the microorganisms in the centreof the colonies, whereas the surrounding microorganismsare labelled by the bacterial probe EUB338 and theDSS658 probe targeting SRB of the DSS (Fig. 9d).
A remarkable feature of these SRB is that they containintracellular aggregates of crystallites (Fig. 9e, see alsoFig. 9b) consisting of iron sulphide (Fig. 9f). Iron sulphides,as strings or clusters of framboids, are also found enclosedin the high-Mg-calcite phase that is common within theblack mats (Figs. 4b and 6).
Pink mat type: Mats of a pink, sometimes more whitishor orange colour typically underlay the black mats. DAPI-staining and TEM reveal cylinder-shaped microorganismswith external sheaths as the most frequent members of themicrobial community (Fig. 10a–b). The cells show a dis-persed distribution rather than a clear tendency to aggregatein definite colonies. Single cells have a diameter of about0.5 µm, and are about 3 µm in length (Fig. 10c). Theyfrequently form segmented filaments in which the singlecells are separated by distinct invaginations. FISH analyses

73
Fig. 7 Interfingering ofcarbonate phases visualized bycathodoluminescence andelement mapping using WDX.Same sample as in Fig. 4a. ACathodoluminescence image ofthe high-Mg-calcite phases.Image mounted from 60 singlemicroscopic images. B WDXmapping of manganese.Increased concentrations (greencolours) coincide withhigh-Mg-calcite (luminescent).C WDX mapping ofmagnesium. Increasedconcentrations (green colours)coincide with high-Mg-calcite.D WDX mapping of strontium.Increased concentrations(green/yellow colours) coincidewith aragonite
identified these microorganisms as archaea of the ANME-1group (Fig. 10d).
A second major component of the microbial commu-nity are localized microbial clusters of some tenths of µmin diameter (Fig. 10a–b). FISH analyses revealed that thecolony-building organisms are SRB belonging to the DSS(Fig. 10c). These bacteria differ from those found in theblack mat in being rod-shaped (∼0.5 µm×1 µm) and non-magnetotactic.
The DSS clusters and ANME-1 show a much more ran-domized spatial arrangement as the distinctive consortiaobserved in the black mat, and there is only a slight, if any,tendency of ANME-1 archaeal cells to surround the DSSaggregates (Fig. 10a–b). It is interesting to see that emptyANME-1 sheaths make up a considerable portion of thepink mats (Fig. 10b–c) and were found as the major frac-tion by volume in some of the sections analysed (see alsoReitner et al. in press).
Green mat type: Microbes of the green mat (Fig. 11)inhabit the carbonate surfaces of cavities within the build-ups. Due to the irregular pore space, this environment can beconsidered as most heterogeneous with respect to the accessof sea water, gaseous methane and seepage fluids. Possi-bly as a consequence, the green mats exhibit a considerablygreater microbial diversity than the other mat types, and theresultant microheterogenity makes it difficult to highlightspecific features. Microscopy and FISH revealed that bothANME-1 and ANME-2 contribute to the archaeal popula-tion of the green mats (Fig. 11b, e). The mats also harboura third archaeal component represented by organisms la-belled by the Crenarchaeota-targeted oligonucleotide probe
Cren499 (Fig. 11f). Crenarchaeota occur sporadically in thegreen mat sections analysed, always in large (>100 µm)aggregates, and well separated from other consortia. How-ever, the morphology of these microorganisms and the de-tailed structure of their aggregates could not be resolvedwith confidence and need to be investigated in further de-tail. As in the pink mat, ANME-1 cells display a morerandomized distribution, whereas ANME-2 cells generallyoccur in colonies (10–200 µm in diameter) together withSRB belonging to the DSS (Fig. 11d). These SRB couldbe best characterized as elongated rods (0.5–1 µm × 2–3 µm) and are non-magnetotactic (Fig. 11b). They containabundant inclusions which may accumulate to as muchas ∼80% of the cell volume (Fig. 11b). These inclusionsare most reminiscent of the polyhydroxyalkanoates (PHA)storage inclusions found in many bacteria of different phy-logenetic affiliation (Schlegel 1985). Unlike in the blackmats, ANME-2 and bacterial cells are evenly distributedwithin the colonies and individual cells are not physicallyaggregated (Fig. 11a–c). Instead, each of these coloniesis surrounded by an approximately 200 nm thick, finelylayered organic envelope (Fig. 11a–b).
Discussion
Carbonate fabrics and mineralogy
From the data now available, it is evident that the Black Seaseep carbonates are true microbialites (sensu Burne andMoore 1987), where the chemical environment inducing

74
Fig. 8 Carbonate carbon and oxygen stable isotope data from severaltower-like build-ups (δ-values relative to the PDB standard). Datafrom a high-Mg-calcite intrasedimenatry concretion (Reitner et al.in press) are given for comparison. All samples were retrieved inthe area of the GHOSTDABS seep-field. Lowest δ13C-values wereobserved for an early, iron sulphide rich high-Mg-calcite phase froma microbial mat sphere
carbonate precipitation is created by methane-based mi-crobial metabolism. The not layered and somewhat chaoticpetrofabric of the carbonate towers can be characterizedas (mega-) thrombolitic (cf. Riding 2000). In some places,nevertheless, this fabric still outlines the contours of theinitial spherical constructions that once protruded into thewater column (Fig. 4e). Whereas thrombolites are knownfrom various settings including methane seeps (Riding2000; Shapiro and Fricke 2002) spherical forms similarto those observed in the Black Sea towers have not beenreported so far from other modern and fossil seep carbon-ates. If found preserved, these elements could be a usefulfacies marker for structures growing into an euxinic watercolumn.
The co-occurrence of high-Mg-calcite and aragonitephases has been recognized previously in Black Sea seepcarbonates (Peckmann et al. 2001; Lein et al. 2002). Thephenomenon is also known from other methane-derivedcarbonate crusts (e.g., from Mediterranean mud volcanoes;
Aloisi et al. 2002). It has been suggested that the precipi-tation of high-Mg-calcite versus aragonite may depend onthe availability of sulphate in the environment of crust for-mation (Peckmann et al. 2001; Aloisi et al. 2002). Indeed,earlier experimental work indicated that dissolved sulphateinhibits calcite precipitation (Walter 1986), although otherfactors such as pH, temperature, Mg2+/Ca2+ ratios andphosphate concentrations represent additional controls oncarbonate mineralogy (Walter 1986; Burton 1993). Highphosphate concentrations inhibit significantly the precipi-tation of aragonite. The role of phosphate during the forma-tion of polyphase carbonate minerals within the cold seepmicrobial mat is subject of further investigations.
The assumption of sulphate as one important control oncarbonate mineralogy offers a plausible explanation for thedistribution of Mg-calcite versus aragonite in the microbialmats studied. In this scenario, zones of Mg-calcite precipi-tation indicate environments where sulphate is rapidly con-verted into sulphide upon AOM. These conditions are foundwithin the black mat, where high abundances of ANME-2 and syntrophic iron sulphide-precipitating DSS are ob-served. Initial precipitation of aragonite, observed in theANME-1-dominated mat type, could be indicative of areasshowing lower rates of sulphate-dependent AOM, and thushigher steady state sulphate concentration. This model is infull agreement with recent laboratory experiments demon-strating a significantly (up to 20-fold) higher cell-specificAOM rate for ANME-2- compared to ANME-1-dominatedcommunities (Nauhaus et al. 2005). The results highlightthe potential of carbonate phases as possible tracers ofmetabolic activity in AOM environments, with possibleimplications for the interpretation of fossil methane-relatedcarbonates with their often complex carbonate mineralogyand growth patterns (cf., Peckmann and Thiel 2004).
The pronounced cathodoluminescence of the high-Mg-calcite can be regarded as a significant facies trait, becauseit typifies a primary process within a distinctive microbialand/or metabolic environment rather than a diagenetic pro-cess. Cathodoluminescence is commonly regarded as beinginduced by the enrichment of metal impurities in the min-eral lattice during diagenesis. Namely manganese is con-sidered as a principal activator of an orange luminescencein diagenetic carbonates (Pagel et al. 2000; Cazenave et al.2003). Indeed, the distribution of the luminescent high-Mg-calcite in the materials studied here is exactly paralleled byincreased Mn contents (Fig. 7b). The role of microbes inthe enrichment of Mn and other metals in individual seepcarbonate phases remains to be explored and is subject ofour ongoing studies.
The nature of the autofluorescence phenomenon ismore difficult to interpret. It can, for instance, be inducedby inclusions of organic molecules containing conju-gated systems. Possible autofluorescence inducers arechlorophyll (Podda et al. 2000) and, more likely in theenvironment studied, Ni-bearing porphinoids, which wererecently found in ANME-1-dominated microbial matsfrom the Black Sea towers (Kruger et al. 2003). Thesecompounds are closely related to nickel cofactor F430 ofmethyl-coenzyme M reductase, the terminal enzyme of

75
Fig. 9 Microbial inventory of the black mat. A Aggregates ofANME-2 archaeal group and associated SRB (DSS group) form-ing a large colony inside the black mat. The colony is lined by un-known filamentous microorganisms. Transmitted light, toluidine blueO staining. B Detail of A. TEM image showing individual aggregates,each with one ANME-2 archaeon in the centre of an envelope of vib-rioform DSS. Note the chains of intracellular greigite mineralisates.C Three-dimensional arrangement of ANME-2 and associated vibri-oform DSS. FE-SEM image of a native (unsputtered) Peldri II-driedsample. The network represents remains of extracellular polymericsubstances (EPS). D Fluorescence images of thin-sections treatedwith an Oregon green (green) labelled oligonucleotide probe spe-
cific for ANME-2 and red-fluorescent (Cy3) labelled oligonucleotideprobe specific for SRB of the Desulfosarcinales/Desulfobacterium(DSS 658) group. The abundance of DSS cells increases towards theouter margins of the colonies (see also A). E Longitudinal sectionof a vibrioform DSS specimen. Magnetosomes occur in a chain-likearrangement. A blotchy, non-uniform diffraction contrast is a typicalfeature of the greigite crystals. TEM micrograph. F Intracellular mag-netosomes visualized by FE-SEM. The ferrimagnetic greigite crystalsoccur as distinct white spherules. ‘x’ marks the beam-position for theEDX spectrum given in the insert. Copper peaks may result from theuse of Cu-grids, but could also be due to an incorporation of copperinto greigite magnetosomes (see Posfai et al. 1998)
methanogenesis, and were regarded as components ofarchaeal enzymes involved in AOM.
Microbiota
The irregular, thrombolithic structure of the towers bearsmacro- and microhabitats that substantially vary in their
shape and chemical environment (e.g., cryptic voids vs.surfaces at the sea water contact). Microbial metabolismand mineralization continuously modify these habitats intime and space, and the need for permanent adaptationsof the organisms involved offers an explanation for thevariability observed among the microbial consortia. Theintrinsic heterogeneity of the seep ecosystem hampers

76
Fig. 10 Microbial inventory of the pink mat. A Cylindrical ANME-1and associated microorganisms, most likely SRB of the DSS group.Note the randomized spatial arrangement with small, isolated SRB-aggregates in a matrix of ANME-1 cells. Semi-thin-section, trans-mitted light, toluidine blue ◦ staining. B TEM micrograph showingthe formation of small clusters by non-magnetic rod-shaped microor-ganisms, most likely SRB of the DSS group (black circle). Note thata large portion of the mat is made of empty ANME-1 sheaths. C
Three-dimensional arrangement of ANME-1 cells and their emptysheaths. FE-SEM image of a native (unsputtered) Peldri II-dried sam-ple. D Fluorescence image of thin-sections treated with an Oregongreen (green) labelled oligonucleotide probe specific for ANME-1and red-fluorescent (Cy3) labelled oligonucleotide probe specific forSRB of the DSS group. The probe response tracks, to some extent,the morphologies of cylindrical ANME-1 and rod-shaped DSS cells(see also B)
the correlation of observations made by independent stud-ies, but some general features can be summarized. Withregard to their morphological properties, the ANME-1archaea, most prominent in the pink mat, resemble ar-chaea reported from this seep area (Pimenov et al. 1997;Michaelis et al. 2002; Reitner et al. in press), and fromcold seeps in the Eel River Basin (Orphan et al. 2002).Putative ANME-1-dominated communities were first visu-alized in TEM sections of microbial mats associated withBlack Sea carbonate crusts by Pimenov et al. (1997). No-tably, these authors reported the absence of bacteria mor-phologically similar to SRB although sulphate-reductionwas indicated by radioisotopy. Michaelis et al. (2002), onthe other hand, identified consortia of ANME-1 and coc-coid DSS in a pink mat obtained from the same studyarea. The emerging picture is that these studies treatedsimilar material, as is here categorized as ‘pink mat’. Sub-stantial variation in the abundance of DSS occurs in theseANME-1-dominated mats, and large portions may evenbe devoid of associated bacteria. Monospecific ANME-1aggregates have also been found in sediments of the EelRiver Basin (Orphan et al. 2002). These observations sug-gest that ANME-1 does not necessarily depend on a closephysical association with a specific SRB partner for theirmetabolism.
The discrete consortia of ANME-2 and DSS observed inthe black mat are remarkably different from those reportedby earlier studies on the Black Sea mats. The DSS morpho-logically resemble forms associated with intrasedimentary
concretions from the same area (Reitner et al. in press).In concretion-associated mats, however, they formed denseaggregations that co-occurred in random association withANME-1. The size (∼30–80 nm i.d.) and arrangement ofthe iron sulphide precipitates inside the DSS cells are thesame as in the magnetosome chains formed by so-calledmagnetotactic bacteria (Posfai et al. 1998; Schuler 1999).Previous studies showed that the accumulation of intracel-lular iron sulphides characterizes anaerobic magnetotacticbacteria, with ferrimagnetic greigite (Fe3S4) formed as theprincipal mineral. The function of these precipitates is stillunder discussion. For motile magnetotactic bacteria, theuse of magnetic nanoparticles as a navigational device hasbeen proposed (Frankel et al. 1997), but this can hardly ac-count for the virtually immobile DSS living in the benthicmats studied here. Rather, the intracellular precipitation ofiron sulphides by some of these bacteria may infer iron cy-cling as an additional biogeochemical pathway associatedwith AOM (see also Reitner et al. in press).
The occurrence of Crenarchaeota in the green mat con-firms earlier work by Tourova et al. (2002), who identifiedgenes from two distinct phylogenetic crenarchaeotal lin-eages, in addition to members of the ANME-1 group in aBlack Sea seep-associated mat. Their occurrence in largeaggregates indicates that they are an integral component ofthe mat population rather than derived from the water col-umn via an input of planktonic Crenarchaeota. However,their physiology remains unknown and their morphologycould so far not definitely be unravelled.

77
Fig. 11 Microbial inventory of the green mat (Loga 01 38/12). AComplex arrangement of microbial colonies with various morpholo-gies forming different types of colonies within the green mat. Thecolonies in the left centre and in the left upper corner of the imageare occupied by consortia of ANME-2 and unknown microorganismshaving large, translucent inclusions in their cells (see also B). Semi-thin-section, transmitted light, toluidine blue O staining. B TEM mi-crograph showing a colony of ANME-2 cells and inclusion-bearingDSS (see also A and C). Note the different arrangement of ANME-2 and their bacterial partners, when compared to the ANME-2/DSSconsortia of the black mat (Fig. 3a–b). A wall-like structure separatesthe colonies from the surrounding environment. Bright rims liningthe ANME-2 cells are dehydration artefacts. C Fluorescence image(FISH) of a thin-section treated with ANME-2 targeted Oregon green(green) labelled oligonucleotide probe and a red-fluorescent (Cy3)labelled probe specific for SRB of the DSS group. The spherical
colonies exhibit a strong ANME-2 signal that partially superposesthe red DSS signal. D Fluorescence image (FISH) of a thin-sectiontreated with red-fluorescent (Cy3) labelled probe specific for SRB ofthe DSS group. These SRB are enriched within more or less sphericalcolonies (see A–C). The strong red signal of the DSS probe is linkedwith large numbers of ribosomes indicating strong metabolic activity.E Detail of A showing ANME-1 archaeal that are frequently observedinhabiting spaces in between the colonies. TEM micrograph. F Flu-orescence image of a thin-section treated with a general Archaea-targeted Oregon green (green) labelled oligonucleotide probe and ared-fluorescent (Cy3) labelled probe specific for the Crenarchaeota.The fluorescence signal traces, to some extent, the morphologies ofcylindrical ANME-1 and less shaped ANME-2 archaeal cells (seealso A). Note that the Crenarchaeota form a discrete colony withinthe complex fabric of the green mat

78
We have shown that the macroscopic appearances(colour) and modes of occurrence of the microbial matsare paralleled by specific biological and petrological fea-tures. From the study of numerous samples, we suggestthat these features represent key facets of the ecosystemassociated with the carbonate towers. However, we needto stress that the use of such categories necessarily im-plies major generalization and simplification. Deviationsand transitional varieties certainly exist, leaving many addi-tional scopes for future research. Likewise, our knowledgeabout the microheterogeneity of these mat systems is stilldeficient. In that respect, the data provided here should betaken as an initial approach rather than a conclusive docu-mentation of the outstanding methane-based environmentof the carbonate towers. The results support the idea thatsulphate-dependent AOM is the dominant biogeochemicalprocess that leads to the formation of the carbonate con-structions. However, it is evident that particular metabolicpathways occurring in AOM-environments are poorly un-derstood, and may be much more complex than anticipatedfrom the overall chemical reaction.
Conclusions
At places where gaseous methane discharges into theanoxic water column, formation of tower-like microbialitesis initiated by the growth of methane-filled calcifyingspheres constructed by microbial mats. Mineralization re-sulting in various calcite and aragonite phases, emergesfrom the microbial mats and proceeds towards the interiorof the spheres, finally creating an irregular ‘thrombolithic’fabric in the central portions of the towers. The biodiver-sity of the microbial mats associated with these build-upsis greater than previously recognized. Several mat varietiesoccur and harbour morphologically and phylogeneticallydistinct communities. Beside numerous yet uncharacter-ized microorganisms, all samples contain Archaea and SRBof the DSS, albeit in varying abundances and modes of oc-currence. ANME-1 generally show a randomized distribu-tion, and seemingly do not require close spatial proximityto a bacterial partner. ANME-2 depend on a tight physicalassociation with particular DSS, exhibiting a surprising de-gree of orderliness. The expression of intracellular greigitemagnetosomes in DSS associated with ANME-2 suggestsiron cycling as an additional process associated with AOM.Crenarchaeota may occur as an integral part of the micro-bial mat community, but their involvement in AOM remainsto be determined.
Acknowledgements This paper is dedicated to the memory of ErikFlugel. Always being open to new ideas, he promoted our workand stimulated our scientific development with great emphasis.He was more than a colleague. We thank the crew of the R/V‘Professor Logachev’ for excellent collaboration during the cruiseand Martin Ischebek for the on-board fixation of biological sam-ples. We are grateful to Joachim Hoefs, Bent T. Hansen, AndreasKronz (all GZG, Gottingen) and Michael Joachimski (Erlangen) foranalytical assistance. This study received financial support throughprogram GHOSTDABS (03G0559A) of the Bundesministerium furBildung und Forschung and the Deutsche Forschungsgemeinschaft
(GEOTECHNOLOGIEN). This is publication No. 11 of the researchprogram GHOSTDABS, and publication No. RCOM0298 of theDFG-Research Center for Ocean Margins
References
Aloisi G, Bouloubassi I, Heijs SK, Pancost RD, Pierre C, SinningheDamste JS, Gottschal JC, Forney LJ, Rouchy J-M (2002) CH4-consuming microorganisms and the formation of carbonatecrusts at cold seeps. Earth Planet Sci Lett 203:195–203
Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, StahlDA (1990a) Combination of 16S rRNA-targeted oligonu-cleotide probes with flow cytometry for analyzing mixedmicrobial populations. Appl Environ Microbiol 56:1919–1925
Amann RI, Krumholz L, Stahl DA (1990b) Fluorescent-oligonucleotide probing of whole cells for determinativephylogenetic and environmental studies in microbiology. JBacteriol 172:762–770
Blumenberg M, Seifert R, Reitner J, Pape T, Michaelis W (2004)Membrane lipid patterns typify distinct anaerobic methan-otrophic consortia. Proc Natl Acad Sci USA 101:11111–11116
Boetius A, Ravenschlag K, Schubert CJ, Rickert D, Widdel F,Gieseke A, Amann R, Jørgensen BB, Witte U, Pfannkuche O(2000) A marine microbial consortium apparently mediatinganaerobic oxidation of methane. Nature 407:623–626
Brown BV (1993) A further chemical alternative to critical-point-drying for preparing small (or large) flies. Fly Times 11:10
Burggraf S, Mayer T, Amann R, Schadhauser S, Woese C, StetterK (1994) Identifying members of the domain Archaea withrRNA-targeted oligonucleotide probes. Appl Environ Microbiol60:3112–3119
Burne RV, Moore LS (1987) Microbialites: organosedimentarydeposits of benthic microbial communities. Palaios 2:241–254
Burton EA (1993) Controls on marine carbonate cement mineralogy:review and reassessment. Chem Geol 105:163–179
Campbell KA, Farmer JD, Des Marais D (2002) Ancient hydrocar-bon seeps from the Mesozoic convergent margin of California:carbonate geochemistry, fluids and paleoenvironments.Geofluids 2:63–94
Cazenave S, Chapoulie R, Villeneuve G (2003) Cathodolumines-cence of synthetic and natural calcite: the effects of manganeseand iron on orange emission. Mineral Petrol 78:243–253
Egorov VN, Luth U, Luth C, Gulin MB (1998) Gas seeps in thesubmarine Dnieper Canyon, Black Sea: acoustic, video andtrawl data. In: Luth U, Luth C, Thiel H (eds) Methane gas seepexplorations in the Black Sea (MEGASEEBS), Project Report.Ber Zent Meeres Klimaforsch, Univ Hamburg, Hamburg, pp 11–21
Frankel RB, Bazylinski DA, Johnson MS, Taylor BL (1997)Magneto-aerotaxis in marine coccoid bacteria. Biophys J73:994–1000
Gal’chenko VF (2004) On the problem of anaerobic methaneoxidation. Microbiology (translated from Mikrobiologiya)73:698–707
Goedert JL, Squires RL (1990) Eocene deep-sea communities inlocalized limestones formed by subduction-related methaneseeps, southwestern Washington. Geology 18:1182–1185
Greinert J, Bohrmann JG, Suess E (2001) Gas hydrate-associatedcarbonates and methane-venting at Hydrate Ridge: classifi-cation, distribution, and origin of authigenic lithologies. In:Natural gas hydrates: occurrence, distribution, and detection.Geophys Monogr 124, American Geophysics Union
Hinrichs K-U, Hayes JM, Sylva SP, Brewer PG, DeLong EF (1999)Methane-consuming archaebacteria in marine sediments.Nature 398:802–805
Hoffmann F, Janussen D, Drose W, Arp G, Reitner J (2003)Histological investigation of organisms with hard skeletons: acase study of siliceous sponges. Biotech Histochem 78

79
Ivanov MV, Polikarpov GG, Lein AY, Galtchenko VF, Egorov VN,Gulin SB, Gulin MB, Rusanov II, Miller YM, Kuptsov VI(1991) Biogeochemistry of the carbon cycle in the region ofmethane gas seeps of the Black Sea. Dokl Akad Nauk USSR320:1235–1240
Kruger M, Meyerdierks A, Glockner FO, Amann R, Widdel F,Kube M, Reinhardt R, Kahnt J, Bocher R, Thauer RK, ShimaS (2003) A conspicuous nickel protein in microbial mats thatoxidize methane anaerobically. Nature 426:878–881
Lein AY, Ivanov MV, Pimenov NV, Gulin MB (2002) Geochemicalcharacteristics of the carbonate constructions formed duringmicrobial oxidation of methane under anaerobic conditions.Microbiology 70:78–90
Luth C, Luth U, Gebruk AV, Thiel H (1999) Methane gas seepsalong the oxic/anoxic gradient in the Black Sea: manifestations,biogenic sediment compounds, and preliminary results onbenthic ecology. Mar Ecol Prog Ser 20:221–249
Manz W, Eisenbrecher M, Neu TR, Szewzyk U (1998) Abundanceand spatial organization of gram negative sulfate-reducingbacteria in activated sludge investigated by in situ probing withspecific 16S rRNA targeted oligonucleotides. FEMS MicrobiolEcol 25:43–61
Manz W, Arp G, Schumann-Kindel G, Szewzyk U, Reitner J (2000)Widefield deconvolution epifluorescence microscopy combinedwith fluorescence in situ hybridization reveals the spatialarrangement of bacteria in sponge tissue. J Microbiol Methods40:125–134
Michaelis W, Seifert R, Nauhaus K, Treude T, Thiel V, BlumenbergM, Knittel K, Gieseke A, Peterknecht K, Pape T, Boetius A,Amann R, Jørgensen BB, Widdel F, Peckmann J, Pimenov NV,Gulin MB (2002) Microbial reefs in the Black Sea fueled byanaerobic oxidation of methane. Science 297:1013–1015
Nauhaus K, Treude T, Boetius A, Kruger M (2005) Environmentalregulation of the anaerobic oxidation of methane: a comparisonof ANME-I and ANME-II communities. Environ Microbiol7:98–106
Orphan V, Hinrichs K-U, Ussler W III, Paull CK, Taylor LT, Sylva SP,Hayes JM, DeLong EF (2001) Comparative analysis of methane-oxidizing archaea and sulfate-reducing bacteria in anoxic marinesediments. Appl Environ Microbiol 67:1922–1934
Orphan V, House CH, Hinrichs K-U, McKeegan KD, DeLong EF(2002) Multiple archaeal groups mediate methane oxidationin anoxic cold seep sediments. Proc Natl Acad Sci USA99:7663–7668
Pagel M, Barbin V, Blanc P, Ohnenstetter D (2000) Cathodolumi-nescence in Geosciences. Springer, Heidelberg
Peckmann J, Thiel V (2004) Carbon cycling at ancient methane-seeps. Chem Geol 205:443–467
Peckmann J, Thiel V, Michaelis W, Clari P, Gaillard C, Martire L,Reitner J (1999) Cold seep deposits of Beauvoisin (Oxfordian;southeastern France) and Marmorito (Miocene; northern Italy):microbially induced, authigenic carbonates. Int J Earth Sci88:60–75
Peckmann J, Reimer A, Luth U, Luth C, Hansen BT, HeinickeC, Hoefs J, Reitner J (2001) Methane-derived carbonates andauthigenic pyrite from the northwestern Black Sea. Mar Geol177:129–150
Pimenov NV, Rusanov II, Poglazova MN, Mityushina LL, SorokinDY, Khmelenina VN, Trotsenko YA (1997) Bacterial matson coral-like structures at methane seeps in the Black Sea.Microbiology (translated from Mikrobiologiya) 66:354–360
Podda F, Zuddas P, Minacci A, Pepi M, Baldi F (2000) Heavy metalcoprecipitation with hydrozincite [Zn5(CO3)2(OH)6] from minewaters caused by photosynthetic microorganisms. Appl EnvironMicrobiol 66:5092–5098
Polikarpov GG, Egorov VN, Nezhdanov AI, Gulin SB, Kulev YD,Gulin MB (1989) The phenomenon of active gas escapes frommounds on the slope of the western Black Sea. Dokl AkadNauk USSR pp 13–16
Posfai M, Buseck PR, Bazylinsky DA, Frankel RB (1998) Ironsulfides from magnetotactic bacteria: structure, composition,and phase transitions. Am Mineral 83:1469–1481
Reitner J, Peckmann J, Blumenberg M, Michaelis W, Reimer A,Thiel V (2005) Concretionary methane-seep carbonates andassociated microbial communities in Black Sea sediments.Palaeogeogr Palaeoclimatol Palaeoecol (in press)
Riding R (2000) Microbial carbonates: the geological record ofcalcified bacterial-algal mats and biofilms. Sedimentology47:179–214
Romeis B (1989) Mikroskopische Technik. Urban and Schwarzen-berg, Munchen
Schlegel HG (1985) Allgemeine Mikrobiologie. Georg Thieme,Stuttgart, New York
Schuler D (1999) Formation of magnetosomes in magnetotacticbacteria. J Mol Microbiol Biotechnol 1:79–86
Shapiro R, Fricke H (2002) Tepee Buttes: fossilized methane-seepecosystems. GSA Field Guide 3:94–101
Thiel V, Peckmann J, Richnow HH, Luth U, Reitner J, MichaelisW (2001) Molecular signals for anaerobic methane oxidationin Black Sea seep carbonates and a microbial mat. Mar Chem73:97–112
Tourova TP, Kolganova TP, Kusnetsov KB, Pimenov N (2002) Phylo-genetic diversity of the archaeal component of bacterial mats oncoral-like structures in zones of methane seeps in the Black Sea.Microbiology (translated from Mikrobiologiya) 71:196–201
Valentine DL (2002) Biogeochemistry and microbial ecology ofmethane oxidation in anoxic environments: a review. Antonievan Leeuwenhook 81:271–282
Walter LN (1986) Relative efficiency of carbonate dissolution andprecipitation during diagenesis: a progress report on the role ofsolution chemistry. SEPM Spec Publ 38:1–11




















