
Long-lasting behavioral effects of juvenile trauma in an animal model of PTSD associated with a...
14
Long-lasting behavioral effects of juvenile trauma in an animal model of PTSD associated with a failure of the autonomic nervous system to recover Hagit Cohen a, ⁎ , Zeev Kaplan a , Michael A. Matar a , Uri Loewenthal a , Joseph Zohar b , Gal Richter-Levin c a Ministry of Health Mental Health Center, Anxiety and Stress Research Unit, Faculty of Health Sciences, Ben-Gurion University of the Negev, Beer-Sheva 84170, Israel b The Chaim Sheba Medical Center, Sackler Medical School, Tel-Aviv University 52621, Israel c Department of Psychology, University of Haifa 31905, Israel Received 13 June 2006; received in revised form 12 October 2006; accepted 7 November 2006 Abstract Background: Early life exposure to potentially traumatic experiences (PTEs) significantly increases the risk of responding more severely to stressful events experienced in adulthood. The aim of this study was to assess the autonomic nervous system (ANS) response to exposure to two PTEs in youth and again in adulthood, in terms of heart rate and heart rate variability in animals that responded to the PTE dramatically as compared to those that displayed virtually no behavioral response and to control animals. Methods: The prevalence of individuals displaying extreme anxiety-like behavioral responses to the PTE (predator urine or elevated platform) was assessed in the elevated plus-maze and startle response paradigms. Behavioral paradigms were complemented by assessment of the involvement of the ANS in relation to changes in behavior. Results: Juvenile trauma increases the vulnerability for developing long-term behavioral disruptions, taken to represent post-traumatic stress symptoms, after a second exposure to the same stressor in adulthood. PTSD-like behaviors and persisting physiological abnormalities resulted from disturbed recovery from the initial stress response. Conclusions: Exposure to a PTE during youth can have significant and long-lasting effects in adulthood and predispose the individual to PTSD upon subsequent re-exposure. Monitoring of ANS parameters confirms that development of extreme long-term (PTSD-like) behavioral changes is associated with a failure of recovery from the initial ANS responses to stress exposure. © 2006 Elsevier B.V. and ECNP. All rights reserved. KEYWORDS Post-traumatic stress disorder; Animal model; Autonomic nervous system; Early life trauma; Juvenile stress; Heart rate; Heart rate variability 1. Introduction The emotional and physical response of the individual to stress involves the activation of a complex cascade of ⁎ Corresponding author. Anxiety and Stress Research Unit, Ministry of Health Mental Health Center, Faculty of Health Sciences, Ben- Gurion University of the Negev, P.O. Box 4600, Beer-Sheva 84170 Israel. Tel.: +972 8 6401743; fax: +972 8 6401742. E-mail address: [email protected] (H. Cohen). 0924-977X/$ - see front matter © 2006 Elsevier B.V. and ECNP. All rights reserved. doi:10.1016/j.euroneuro.2006.11.003 www.elsevier.com/locate/euroneuro European Neuropsychopharmacology (2007) 17, 464–477
Transcript of Long-lasting behavioral effects of juvenile trauma in an animal model of PTSD associated with a...

www.e l sev i e r. com/ l oca te /eu roneu ro
European Neuropsychopharmacology (2007) 17, 464–477
Long-lasting behavioral effects of juvenile trauma inan animal model of PTSD associated with a failure ofthe autonomic nervous system to recoverHagit Cohen a,⁎, Zeev Kaplan a, Michael A. Matar a, Uri Loewenthal a,Joseph Zohar b, Gal Richter-Levin c
a Ministry of Health Mental Health Center, Anxiety and Stress Research Unit, Faculty of Health Sciences,Ben-Gurion University of the Negev, Beer-Sheva 84170, Israelb The Chaim Sheba Medical Center, Sackler Medical School, Tel-Aviv University 52621, Israelc Department of Psychology, University of Haifa 31905, Israel
Received 13 June 2006; received in revised form 12 October 2006; accepted 7 November 2006
⁎ Corresponding author. Anxiety andof Health Mental Health Center, FacuGurion University of the Negev, P.O.Israel. Tel.: +972 8 6401743; fax: +972
E-mail address: [email protected] (H
0924-977X/$ - see front matter © 200doi:10.1016/j.euroneuro.2006.11.003
Abstract
Background: Early life exposure to potentially traumatic experiences (PTEs) significantlyincreases the risk of responding more severely to stressful events experienced in adulthood. Theaim of this study was to assess the autonomic nervous system (ANS) response to exposure to twoPTEs in youth and again in adulthood, in terms of heart rate and heart rate variability in animalsthat responded to the PTE dramatically as compared to those that displayed virtually nobehavioral response and to control animals.Methods: The prevalence of individuals displaying extreme anxiety-like behavioral responses tothe PTE (predator urine or elevated platform) was assessed in the elevated plus-maze and startleresponse paradigms. Behavioral paradigms were complemented by assessment of theinvolvement of the ANS in relation to changes in behavior.Results: Juvenile trauma increases the vulnerability for developing long-term behavioraldisruptions, taken to represent post-traumatic stress symptoms, after a second exposure to thesame stressor in adulthood. PTSD-like behaviors and persisting physiological abnormalitiesresulted from disturbed recovery from the initial stress response.Conclusions: Exposure to a PTE during youth can have significant and long-lasting effects inadulthood and predispose the individual to PTSD upon subsequent re-exposure. Monitoring of ANSparameters confirms that development of extreme long-term (PTSD-like) behavioral changes isassociated with a failure of recovery from the initial ANS responses to stress exposure.© 2006 Elsevier B.V. and ECNP. All rights reserved.
KEYWORDSPost-traumatic stressdisorder;Animal model;Autonomic nervoussystem;Early life trauma;Juvenile stress;Heart rate;Heart rate variability
Stress Research Unit, Ministrylty of Health Sciences, Ben-Box 4600, Beer-Sheva 841708 6401742.. Cohen).
6 Elsevier B.V. and ECNP. All right
1. Introduction
The emotional and physical response of the individual tostress involves the activation of a complex cascade of
s reserved.

465Lack of ANS recovery in juvenile trauma
physiological systems (McEwen, 2001). Exposure to stressrepresents a threat to homeostasis, triggering the activationof the stress cascade, beginning with corticotropin-releasinghormone (CRH) and locus caeruleus–norepinephrine (LC/NE)-autonomic systems centrally and culminating in theirperipheral effectors, the pituitary–adrenal axis and theautonomic system. The consequences of the stress responseare generally adaptive in the short run, representing ahomeostatic reaction (allostasis) (McEwen, 2002). When thetemporary dysregulation is not overcome by the organism tore-establish and maintain homeostasis, long-term patholog-ical changes can occur. This is exemplified in human subjectsby post-traumatic stress disorder (PTSD), a clinical conditioncharacterized by both psychological and physiologicalcomponents, resulting in symptoms involving intrusive re-experiencing of events, avoidance of reminders, emotionalnumbing and hyper-arousal (American Psychiatric Associa-tion, 1994).
In humans and in animals, potentially traumatizingstressful experiences (PTEs) occurring in early life havelong-term effects on the individual (Breslau, 2002; Ford andKidd, 1998; Heim and Nemeroff, 2001; Heim et al., 2002;Shea et al., 2005). Numerous studies in rodents havedemonstrated substantial long-term bio-physiological con-sequences of environmental manipulations in immature (pre-weaning) animals, in which the formation of neuroendocrineregulatory mechanisms has not been completed and asso-ciative neuronal circuits are not yet fully elaborated (Caldjiet al., 2000; Francis et al., 1999; Goldberg et al., 2003;Heidbreder et al., 2000; Maslova et al., 2002a,b). Juvenile(post-weaning, but pre-pubertal) animals have also beenshown to demonstrate long-term behavioral consequencessubsequent to early exposure to PTEs (Avital and Richter-Levin, 2005).
There is a considerable degree of variability in individualresponses to potentially traumatic events in humans and,over time, a minority is found to fulfill criteria for PTSD; thediagnosis requires a certain number of symptoms, from eachof three symptom clusters, to have been manifested for aminimal period of time. Animal studies have routinelyincluded the entire exposed population as the studypopulation. Since animals also display significant individualvariance in their responses, in our lab, we routinelysegregate the study population into groups according todegree of individual response to the stressor in order tomodel the approach to the clinical condition more closely.
The application of maximal and minimal cutoff behavioralcriteria (CBC), paralleling clinical study inclusion/exclusioncriteria, enables the most clearly affected extreme behav-ioral response (EBR) group to be set apart from their minimalbehavioral response (MBR) counterparts. In this manner, it ispossible to focus on the physiological data for animals withextremely disrupted behavior patterns as distinct from thedata for animals virtually unaffected behaviorally by thestressor. The overall proportion of EBR animals in a largeseries of studies has been approximately 25%, a figureapproximating the proportion of individuals fulfilling criteriafor PTSD from among those exposed to potentially traumaticexperiences in human studies (Breslau et al., 1991; Resnicket al., 1993). These behaviorally distinct groups of animalshave consistently demonstrated differing concomitant phys-iological responses, in terms of the hypothalamic–pituitary–
adrenal axis and the autonomic nervous system (ANS), themore marked physiological changes being associated withmore extremely disrupted behavioral responses and minimalphysiological changes with almost no behavioral response(Cohen et al., 2003, 2004a, 2005, 2006a,b,c; Cohen andZohar, 2004).
Despite the wealth of studies on neuroendocrineresponses to early life stress, there is only limited informa-tion regarding the autonomic nervous response to early lifestressors. The ANS, expressed in terms of cardiac measuressuch as heart rate (HR) and heart rate variability (HRV), hasbeen a major focus of attention in the study of thephysiological component of PTSD.
1.1. Heart rate variability
Heart rate variability is a physiological phenomenon thatreflects the influence of the ANS on cardiac parameters.Efferent links in the neural control of heart rhythm consist ofsympathetic and parasympathetic fibers innervating thesinus node. Because sympathetic and parasympathetic firingalter spontaneous sinus node depolarization, cardiac rateand rhythm convey information about autonomic influenceson the heart (Elghozi et al., 2001). Heart rate is not constantbut oscillates around a mean value determined by modulat-ing effects of ANS activity. These effects may be assessed bymeans of measuring changes in sinus rate over time or, moreprecisely, the standard deviation of intervals betweensuccessive R waves (SDRR) of the cardiac cycle, which aretermed heart rate variability (HRV) (Akselrod et al., 1981;Malliani et al., 1991; Stein and Kleiger, 1999; Task Force,1996).
Depression of HRV has been observed in a variety ofcardiac and non-cardiac clinical conditions, includingautonomic neuropathy, heart transplantation, congestiveheart failure and myocardial infarction (Task Force, 1996).The clinical implications of HRV analysis include its clearlyrecognized value as a predictor of risk for arrhythmicevents and/or sudden cardiac death after acute MI and as aclinical marker of evolving diabetic neuropathy (TaskForce, 1996).
Subjects suffering from both psychiatric and cardiovas-cular disorders may be at increased risk owing to increasedANS involvement. For example, Carney et al. (1995) reportedan increased incidence of ventricular tachycardia amongcoronary artery disease patients with clinical depression,compared to that among non-depressed coronary arterydisease patients. This increase was attributed to reduce HRVin the group with both diagnoses.
The overall aim of this study was to examine theconsequences of juvenile stress on behavior and physiologyat later stages of life, applying the CBC principle for groupingexposed individuals according to degree of response.Physiological consequences were assessed by measuring HRand HRV. The trials aimed to examine the relationshipbetween immediate and long-term behavioral and ANSresponses to single versus repeated PTE exposure, distin-guishing between exposure early in life and later on andrepeated exposure in adulthood. The PTEs consisted of twodistinct stress paradigms—elevated platform and the scent ofa prime predator (soiled cat litter). The working hypothesiswas that the long-term behavioral changes and predisposition
466 H. Cohen et al.
to more marked responses to stress exposure in adulthoodresulting from early life exposure would be reflected in thephysiological data of the individual.
2. Materials and methods
All treatment and testing procedures were approved by the AnimalCare Committee of Ben-Gurion University of the Negev and werecarried out in accordance with the NIH Guide for the Care and Use ofLaboratory Animals.
2.1. Animals
Adult male Wistar rats (45–49 g) (Harlan, Jerusalem, Israel) werehoused four per cage in 75×55×15 cm Plexiglas cages intemperature-controlled (22±1°C) animal quarters on a 12:12light–dark cycle, with free access to standard Purina Rat Chowpellets and water. Rats were weighed and treated by the sameinvestigator.Male rats were selected for this study in order toeliminate the possible confounding effects of sex hormones. This iscommon practice; male animals have predominated in themajority of animal studies for this very reason. Moreover, althoughsome studies in human subjects indeed demonstrate a predomi-nance of female vs. male patients, this has not been a uniformfinding at all and numerous humans studies do not demonstratedgender differences for incidence and/or recovery rates of PTSDpatterns (Freedman et al., 2002).
2.2. Experimental design
The animals were randomly assigned to one of 16 groups as describedin Table 1.
There were 20 animals in each group.Stress procedure: Two different stressors were used: elevated
platform and predator scent stress.Platform exposure (EP): Animals were placed on a 12 × 12 cm
platform, located 7–10 cm above the water level in the center of apool of water for 10 min in a brightly lit room.
Table 1 Experimental design
Groups Age atexposure(PND)⁎
Type ofstressor
PSSa EPa
Unexposed control (Con)Juvenile stress (JS) 28 xAdulthood stress (AS) 60 xJuvenile+adulthood stress (JS+AS) 28 + 60 xUnexposed control (Con)Juvenile stress (JS) 28 xAdulthood stress (AS) 60 xJuvenile+adulthood stress (JS+AS) 28+60 xUnexposed control (Con)Juvenile stress (JS) 28 xAdulthood stress (AS) 60 xJuvenile+adulthood stress (JS+AS) 28+60 xUnexposed control (Con)Juvenile stress (JS) 28 xAdulthood stress (AS) 60 xJuvenile+adulthood stress (JS+AS) 28+60 xa PND=post-natal day; PSS=predator scent stress; EP=elevated plat
Predator stress scent (PSS): The test animals were placed onwell-soiled cat litter for 10 min (in use by the cat for 2 days, siftedfor stools). Control animals were exposed to fresh, unused litter.
2.3. Behavioral paradigms
The rats were tested in two behavioral models—the elevated plus-maze (EPM) and the acoustic startle response (ASR) paradigm.
The elevated plus-maze: This maze consists of a plus-shapedplatform with two open arms and two closed arms, with arms of thesame type located opposite each other (File, 1993).Closed arms aresurrounded by 14-cm-high opaque walls on three sides. Each rat wasplaced on the central platform facing an open arm and was allowedto explore the maze for 5 min. Each test was videotaped and scoredby an independent observer. Arm entry was defined as entering anarm with all four paws.
The following terms were used: durations in open and closedarms; open and closed arm entries.
Acoustic startle response: Startle response was measured usingtwo ventilated startle chambers (SR-LAB system, San Diego Instru-ments, San Diego, CA). Each chamber consisted of a Plexiglascylinder resting on a platform inside a sound-attenuated, ventilatedchamber. Movement inside the tube was detected by a piezoelectricaccelerometer below the frame. Sound levels within each testchamber were measured routinely using a sound level meter (RadioShack) to ensure consistent presentation. Chambers were calibrateddaily. The animals were placed inside the tube and allowed a 5-minacclimatization period with background noise only. Following a 5-min acclimatization period, 30 acoustic startle trials (110 dB whitenoise; 40 ms duration; 30 or 45 s inter-trial interval) were presentedover the 68-dB white noise background.
Two measures of behavior were assessed: (1) mean startleamplitude and (2) percent of startle responses habituation overrepeated presentation of the acoustic pulse. Startle amplitude wasaveraged over all 30 trials. Percent habituation was calculated bycomparing the responses in the first and last blocks of six trials each.
Percent habituation=100×[(average startle amplitude in Block1)− (average startle amplitude in Block 5]/(average startle ampli-tude in Block 1).
Age ofbehavioralassessment(PND)⁎
Age at ANS evaluation (PND) a
Day 60 Day 80 Baseline During Day 60 Day 80
x x xx x x xx x x xx x x x
x x xx x x xx x x xx x x x
x x xx x x xx x x xx x x x
x x xx x x xx x x xx x x x
form stress.

467Lack of ANS recovery in juvenile trauma
2.4. Behavioral assessments for determination of cutoffbehavioral criteria (CBCs)
We used two consecutive tests whose outcomes from the basis dothe CBCs, each of which represents a marker for a diagnosticcluster on the conceptual level. Fearful behaviors on the elevatedplus-maze and non-habituated startle reaction were selected toconstitute the basis for the cutoff behavioral criteria (CBCs) for anumber of reasons. Firstly, each has been shown to be a validmeasure of stress responses in numerous studies. Secondly, theyare each well defined and straightforward to score. Extremeresponses to both of these serial CBCs were required for“inclusion” into the EBR group, whereas the reverse was requiredfor inclusion in the MBR group.
The assessments were designed in two steps:
2.4.1. Step I: Assessment of overall behavioral defectPrior to attempting to distinguish the differentially affectedsubgroups, we routinely performed a preliminary assessment ofthe overall response of the exposed population, to ascertain thatexposure to the stressor did, in fact, have significant overallbehavioral effects on the exposed rats as a group compared tocontrols. As a standard procedure, the data are examined to verifythat there is a sufficient distribution of behavioral changes for validapplication of the CBCs—i.e., that the potential yield of individualsin the extreme groups suffices.
Behavioral changes such as extremely compromised exploratorybehavior on the plus-maze and markedly increased startle reactionthat does not undergo any adaptation, reflect anxiety-like beha-viors, i.e., fearfulness and hypervigilance.
2.4.2. Step II: Application of the CBCs to the dataHaving established that the stressor had an effect on the studypopulation as a whole, and that not all animals responded to it in thesame manner, we focused only on animals that demonstratedextremes of behavioral change on the one hand (extreme behavioralresponse), or virtually no change on the other hand (minimalbehavioral response). Data for rats with partial responses were setaside for reasons of clarity.
In order to maximize the resolution and minimize false-positives,we defined the behavioral cutoff criteria to represent the mostextreme degree of behavioral disturbance in each of the twoconsecutive behavioral paradigms. Thus, in order to be defined as“affected” (EBR), the individual animal had to have conformed toboth sets of criteria, consecutively. Conversely, in order to beconsidered to have responded hardly at all, animals must haveconformed to equally extreme criteria for “near normal” (minimalbehavioral response (MBR)) behaviors. The validity of the criteriawas reaffirmed in each study by ascertaining that the vast majorityof unexposed control animals conformed to the criteria for MBR andnone to the criteria for EBR.
The CBCs, determined as above, were as follows:Extreme behavioral response:
(1) 5 min (entire session) spent in closed arms and no entries intothe open arms;
(2) mean amplitude of the startle response (at 110 dB) N1200 Uand startle response shows no habituation over time.
Minimal behavioral response:
(1) 0–1 min spent in closed arms and ≥8 open-arm entries;(2) mean amplitude of the startle response (at 110 dB) b700 U and
normal habituation of the acoustic startle response.
Weight: The rats' weights were monitored over the experimentalperiod. A one-way ANOVA was performed for repeated measuredesigns (data not shown).
2.5. Evaluation of autonomic nervous system
ANS effects of the stress were assessed at a number of time points toassess immediate, short-term and enduring effects: at baseline,during exposure and at days 60 and 80. Data were analyzed for theentire population exposed to each stressor and then looked at againaccording to the CBCs—i.e., for the selected groups of EBR and MBRrats.
All electrocardiographic (ECG) data were registered at the sametime of day (between 3 and 4 p.m.). The ECG signal was acquiredover 180 s on a PC-based system, equipped with a 16-channel A/Dacquisition card (Axotape, Axon Instruments, Inc., Foster City, CA,USA) at a sampling rate of 2 kHz (Oxford 4-24). This ECG sampleduration is a good compromise between the need for a large timeseries to achieve sufficient accuracy in terms of computation and theneed to obtain stationary recordings, which require short periods oftime. To avoid “noise” and ectopic beats, each ECG recording wascarefully reviewed visually (Task Force, 1996).
HR and HRV were analyzed in the time domain. The statisticalparameters were calculated as mean value of HR and the standarddeviation of the successive difference in RR intervals (HRV).
2.6. Statistical analyses
For the HR and HRV results, the statistical analyses were performedusing one-way analysis of variance (ANOVA). Where significant groupeffects were detected, Scheffe's test (pb0.05) was used to test forsignificant post hoc differences between individual groups.
The prevalence of the affected rats as a function of study groupwas tested using cross-tabulation and non-parametric Fisher exacttests.
Linear correlations between behavioral parameters and HR andHRV data were assessed.
3. Results
3.1. The effect of the predator scent stress (PSS)paradigm on overall anxiety-like behavior andstartle response
As shown in Fig. 1 (A and B), within the PSS-exposedpopulation a broad range of variation in behavioral responsewas observed and several subgroups were identifiable.Based on the results of this phase of the study, the animalswere subdivided into groups reflecting magnitude ofresponse according to the CBCs, focusing selectively onEBR and MBR.
3.2. Acute and enduring behavioral responsepatterns to the predator scent stress paradigm (Fig. 2)
There were significant differences in the prevalence rates ofindividuals displaying an extreme degree of behavioralresponses (EBR) among the various groups. For day 60:Pearson χ2=32.7, df=3, pb0.00001; and for day 80: Pearsonχ2=26.8, df=3, pb0.00001.
As shown in Fig. 2A, there were no EBR individuals amongthe unexposed controls. Fifty percent of each of the twopopulations of rats exposed to either juvenile (JS—28 days)or adulthood (AS—60 days) stress demonstrated extremebehavioral responses when tested at 60 days, a significanteffect as compared to controls (Fisher exact pb0.0002). Incontrast, 90% of those exposed to both juvenile andadulthood trauma (JS+AS) demonstrated EBR, significantly
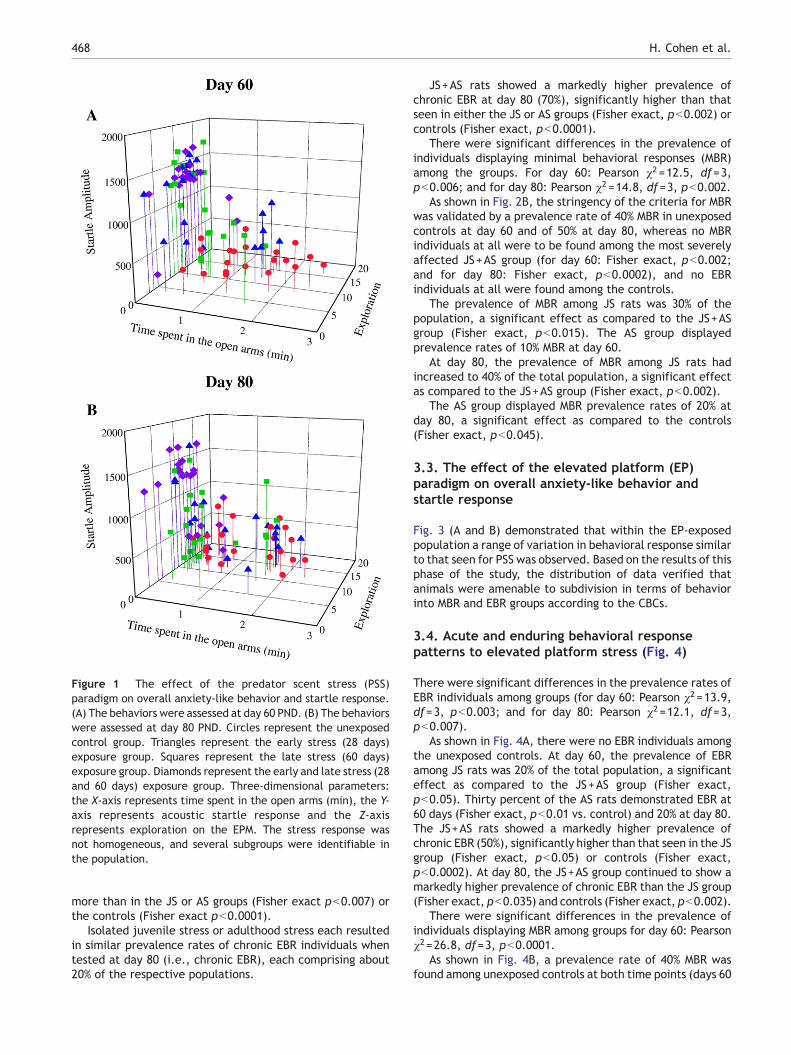
Figure 1 The effect of the predator scent stress (PSS)paradigm on overall anxiety-like behavior and startle response.(A) The behaviors were assessed at day 60 PND. (B) The behaviorswere assessed at day 80 PND. Circles represent the unexposedcontrol group. Triangles represent the early stress (28 days)exposure group. Squares represent the late stress (60 days)exposure group. Diamonds represent the early and late stress (28and 60 days) exposure group. Three-dimensional parameters:the X-axis represents time spent in the open arms (min), the Y-axis represents acoustic startle response and the Z-axisrepresents exploration on the EPM. The stress response wasnot homogeneous, and several subgroups were identifiable inthe population.
468 H. Cohen et al.
more than in the JS or AS groups (Fisher exact pb0.007) orthe controls (Fisher exact pb0.0001).
Isolated juvenile stress or adulthood stress each resultedin similar prevalence rates of chronic EBR individuals whentested at day 80 (i.e., chronic EBR), each comprising about20% of the respective populations.
JS+AS rats showed a markedly higher prevalence ofchronic EBR at day 80 (70%), significantly higher than thatseen in either the JS or AS groups (Fisher exact, pb0.002) orcontrols (Fisher exact, pb0.0001).
There were significant differences in the prevalence ofindividuals displaying minimal behavioral responses (MBR)among the groups. For day 60: Pearson χ2=12.5, df=3,pb0.006; and for day 80: Pearson χ2=14.8, df=3, pb0.002.
As shown in Fig. 2B, the stringency of the criteria for MBRwas validated by a prevalence rate of 40% MBR in unexposedcontrols at day 60 and of 50% at day 80, whereas no MBRindividuals at all were to be found among the most severelyaffected JS+AS group (for day 60: Fisher exact, pb0.002;and for day 80: Fisher exact, pb0.0002), and no EBRindividuals at all were found among the controls.
The prevalence of MBR among JS rats was 30% of thepopulation, a significant effect as compared to the JS+ASgroup (Fisher exact, pb0.015). The AS group displayedprevalence rates of 10% MBR at day 60.
At day 80, the prevalence of MBR among JS rats hadincreased to 40% of the total population, a significant effectas compared to the JS+AS group (Fisher exact, pb0.002).
The AS group displayed MBR prevalence rates of 20% atday 80, a significant effect as compared to the controls(Fisher exact, pb0.045).
3.3. The effect of the elevated platform (EP)paradigm on overall anxiety-like behavior andstartle response
Fig. 3 (A and B) demonstrated that within the EP-exposedpopulation a range of variation in behavioral response similarto that seen for PSS was observed. Based on the results of thisphase of the study, the distribution of data verified thatanimals were amenable to subdivision in terms of behaviorinto MBR and EBR groups according to the CBCs.
3.4. Acute and enduring behavioral responsepatterns to elevated platform stress (Fig. 4)
There were significant differences in the prevalence rates ofEBR individuals among groups (for day 60: Pearson χ2=13.9,df=3, pb0.003; and for day 80: Pearson χ2=12.1, df=3,pb0.007).
As shown in Fig. 4A, there were no EBR individuals amongthe unexposed controls. At day 60, the prevalence of EBRamong JS rats was 20% of the total population, a significanteffect as compared to the JS+AS group (Fisher exact,pb0.05). Thirty percent of the AS rats demonstrated EBR at60 days (Fisher exact, pb0.01 vs. control) and 20% at day 80.The JS+AS rats showed a markedly higher prevalence ofchronic EBR (50%), significantly higher than that seen in the JSgroup (Fisher exact, pb0.05) or controls (Fisher exact,pb0.0002). At day 80, the JS+AS group continued to show amarkedly higher prevalence of chronic EBR than the JS group(Fisher exact, pb0.035) and controls (Fisher exact, pb0.002).
There were significant differences in the prevalence ofindividuals displaying MBR among groups for day 60: Pearsonχ2=26.8, df=3, pb0.0001.
As shown in Fig. 4B, a prevalence rate of 40% MBR wasfound among unexposed controls at both time points (days 60
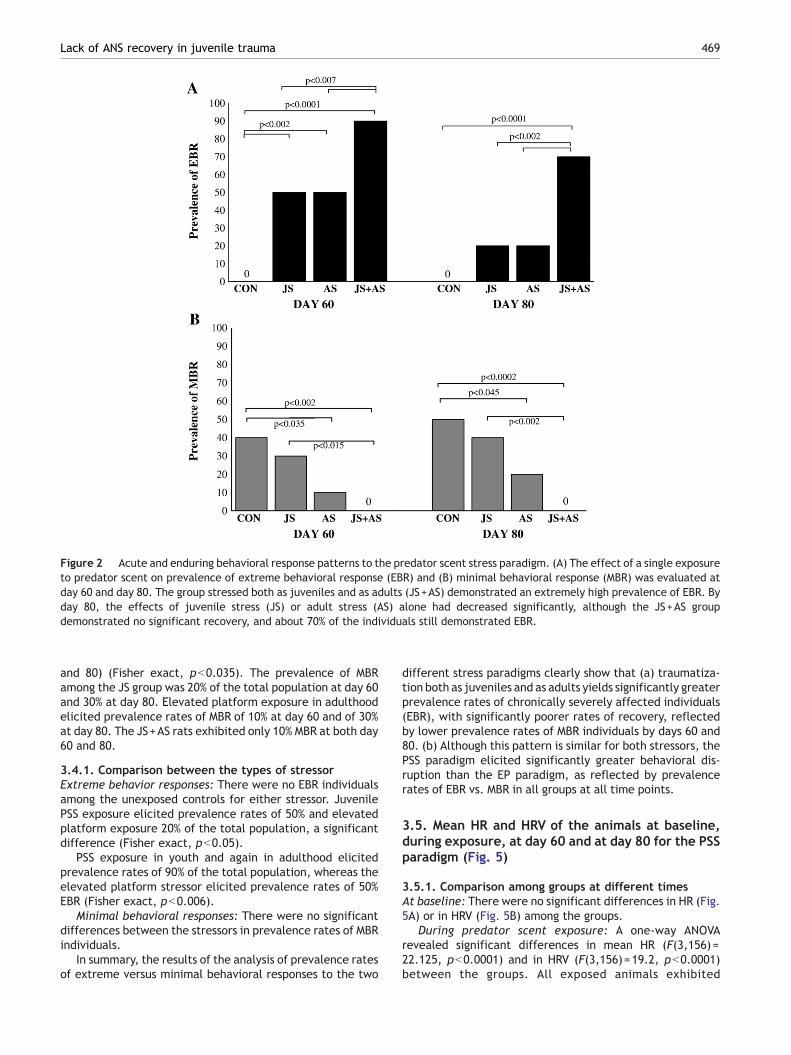
Figure 2 Acute and enduring behavioral response patterns to the predator scent stress paradigm. (A) The effect of a single exposureto predator scent on prevalence of extreme behavioral response (EBR) and (B) minimal behavioral response (MBR) was evaluated atday 60 and day 80. The group stressed both as juveniles and as adults (JS+AS) demonstrated an extremely high prevalence of EBR. Byday 80, the effects of juvenile stress (JS) or adult stress (AS) alone had decreased significantly, although the JS+AS groupdemonstrated no significant recovery, and about 70% of the individuals still demonstrated EBR.
469Lack of ANS recovery in juvenile trauma
and 80) (Fisher exact, pb0.035). The prevalence of MBRamong the JS group was 20% of the total population at day 60and 30% at day 80. Elevated platform exposure in adulthoodelicited prevalence rates of MBR of 10% at day 60 and of 30%at day 80. The JS+AS rats exhibited only 10% MBR at both day60 and 80.
3.4.1. Comparison between the types of stressorExtreme behavior responses: There were no EBR individualsamong the unexposed controls for either stressor. JuvenilePSS exposure elicited prevalence rates of 50% and elevatedplatform exposure 20% of the total population, a significantdifference (Fisher exact, pb0.05).
PSS exposure in youth and again in adulthood elicitedprevalence rates of 90% of the total population, whereas theelevated platform stressor elicited prevalence rates of 50%EBR (Fisher exact, pb0.006).
Minimal behavioral responses: There were no significantdifferences between the stressors in prevalence rates of MBRindividuals.
In summary, the results of the analysis of prevalence ratesof extreme versus minimal behavioral responses to the two
different stress paradigms clearly show that (a) traumatiza-tion both as juveniles and as adults yields significantly greaterprevalence rates of chronically severely affected individuals(EBR), with significantly poorer rates of recovery, reflectedby lower prevalence rates of MBR individuals by days 60 and80. (b) Although this pattern is similar for both stressors, thePSS paradigm elicited significantly greater behavioral dis-ruption than the EP paradigm, as reflected by prevalencerates of EBR vs. MBR in all groups at all time points.
3.5. Mean HR and HRV of the animals at baseline,during exposure, at day 60 and at day 80 for the PSSparadigm (Fig. 5)
3.5.1. Comparison among groups at different timesAt baseline: There were no significant differences in HR (Fig.5A) or in HRV (Fig. 5B) among the groups.
During predator scent exposure: A one-way ANOVArevealed significant differences in mean HR (F(3,156)=22.125, pb0.0001) and in HRV (F(3,156)=19.2, pb0.0001)between the groups. All exposed animals exhibited
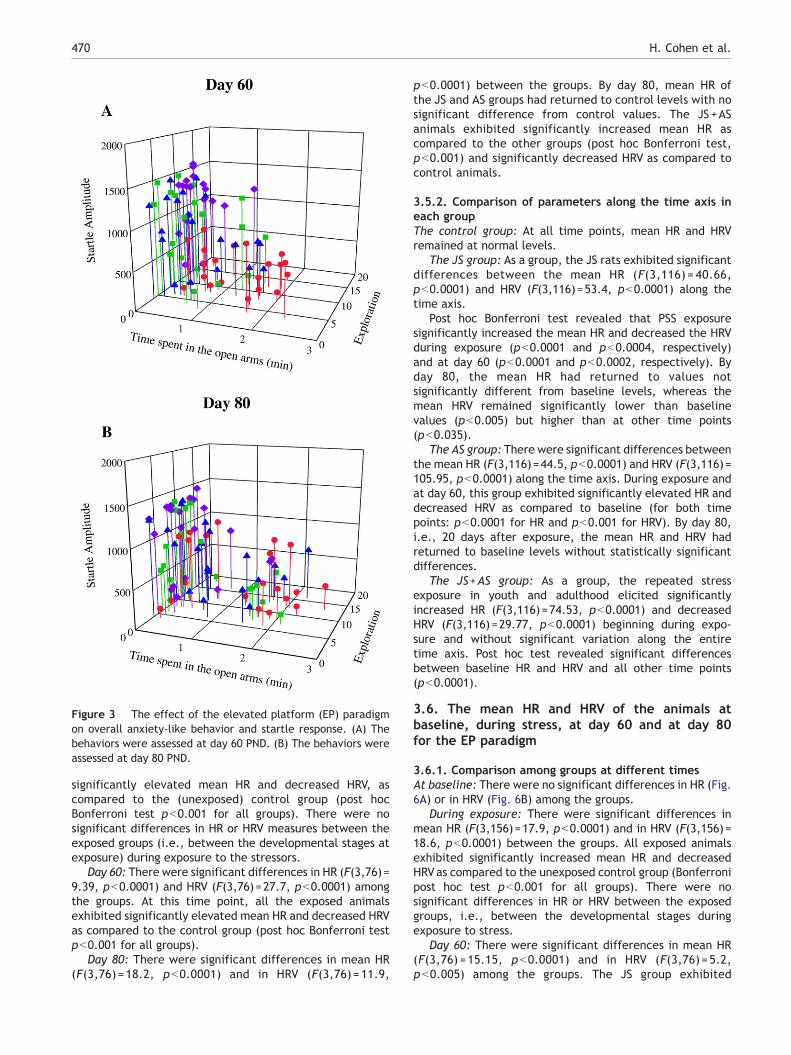
Figure 3 The effect of the elevated platform (EP) paradigmon overall anxiety-like behavior and startle response. (A) Thebehaviors were assessed at day 60 PND. (B) The behaviors wereassessed at day 80 PND.
470 H. Cohen et al.
significantly elevated mean HR and decreased HRV, ascompared to the (unexposed) control group (post hocBonferroni test pb0.001 for all groups). There were nosignificant differences in HR or HRV measures between theexposed groups (i.e., between the developmental stages atexposure) during exposure to the stressors.
Day 60: There were significant differences in HR (F(3,76)=9.39, pb0.0001) and HRV (F(3,76)=27.7, pb0.0001) amongthe groups. At this time point, all the exposed animalsexhibited significantly elevated mean HR and decreased HRVas compared to the control group (post hoc Bonferroni testpb0.001 for all groups).
Day 80: There were significant differences in mean HR(F(3,76)=18.2, pb0.0001) and in HRV (F(3,76)= 11.9,
pb0.0001) between the groups. By day 80, mean HR ofthe JS and AS groups had returned to control levels with nosignificant difference from control values. The JS+ASanimals exhibited significantly increased mean HR ascompared to the other groups (post hoc Bonferroni test,pb0.001) and significantly decreased HRV as compared tocontrol animals.
3.5.2. Comparison of parameters along the time axis ineach groupThe control group: At all time points, mean HR and HRVremained at normal levels.
The JS group: As a group, the JS rats exhibited significantdifferences between the mean HR (F(3,116) = 40.66,pb0.0001) and HRV (F(3,116)=53.4, pb0.0001) along thetime axis.
Post hoc Bonferroni test revealed that PSS exposuresignificantly increased the mean HR and decreased the HRVduring exposure (pb0.0001 and pb0.0004, respectively)and at day 60 (pb0.0001 and pb0.0002, respectively). Byday 80, the mean HR had returned to values notsignificantly different from baseline levels, whereas themean HRV remained significantly lower than baselinevalues (pb0.005) but higher than at other time points(pb0.035).
The AS group: There were significant differences betweenthe mean HR (F(3,116)=44.5, pb0.0001) and HRV (F(3,116)=105.95, pb0.0001) along the time axis. During exposure andat day 60, this group exhibited significantly elevated HR anddecreased HRV as compared to baseline (for both timepoints: pb0.0001 for HR and pb0.001 for HRV). By day 80,i.e., 20 days after exposure, the mean HR and HRV hadreturned to baseline levels without statistically significantdifferences.
The JS+AS group: As a group, the repeated stressexposure in youth and adulthood elicited significantlyincreased HR (F(3,116)=74.53, pb0.0001) and decreasedHRV (F(3,116)=29.77, pb0.0001) beginning during expo-sure and without significant variation along the entiretime axis. Post hoc test revealed significant differencesbetween baseline HR and HRV and all other time points(pb0.0001).
3.6. The mean HR and HRV of the animals atbaseline, during stress, at day 60 and at day 80for the EP paradigm
3.6.1. Comparison among groups at different timesAt baseline: There were no significant differences in HR (Fig.6A) or in HRV (Fig. 6B) among the groups.
During exposure: There were significant differences inmean HR (F(3,156)=17.9, pb0.0001) and in HRV (F(3,156)=18.6, pb0.0001) between the groups. All exposed animalsexhibited significantly increased mean HR and decreasedHRVas compared to the unexposed control group (Bonferronipost hoc test pb0.001 for all groups). There were nosignificant differences in HR or HRV between the exposedgroups, i.e., between the developmental stages duringexposure to stress.
Day 60: There were significant differences in mean HR(F(3,76) =15.15, pb0.0001) and in HRV (F(3,76)= 5.2,pb0.005) among the groups. The JS group exhibited
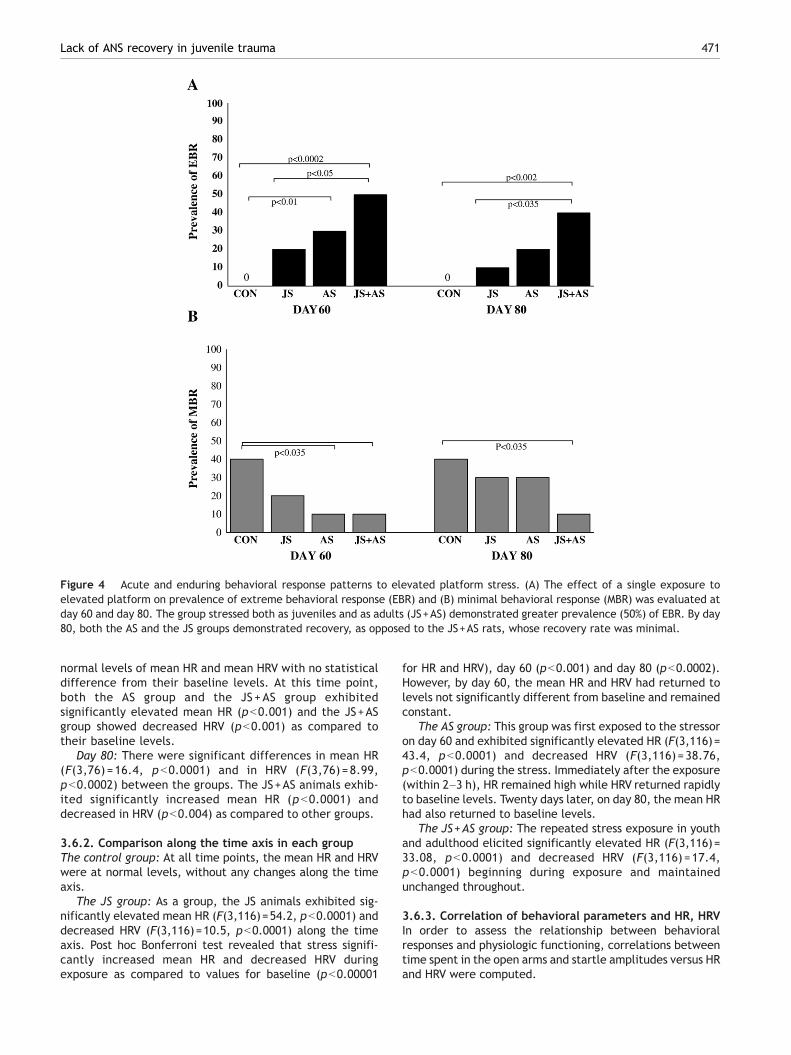
Figure 4 Acute and enduring behavioral response patterns to elevated platform stress. (A) The effect of a single exposure toelevated platform on prevalence of extreme behavioral response (EBR) and (B) minimal behavioral response (MBR) was evaluated atday 60 and day 80. The group stressed both as juveniles and as adults (JS+AS) demonstrated greater prevalence (50%) of EBR. By day80, both the AS and the JS groups demonstrated recovery, as opposed to the JS+AS rats, whose recovery rate was minimal.
471Lack of ANS recovery in juvenile trauma
normal levels of mean HR and mean HRV with no statisticaldifference from their baseline levels. At this time point,both the AS group and the JS +AS group exhibitedsignificantly elevated mean HR (pb0.001) and the JS+ASgroup showed decreased HRV (pb0.001) as compared totheir baseline levels.
Day 80: There were significant differences in mean HR(F(3,76) =16.4, pb0.0001) and in HRV (F(3,76) =8.99,pb0.0002) between the groups. The JS+AS animals exhib-ited significantly increased mean HR (pb0.0001) anddecreased in HRV (pb0.004) as compared to other groups.
3.6.2. Comparison along the time axis in each groupThe control group: At all time points, the mean HR and HRVwere at normal levels, without any changes along the timeaxis.
The JS group: As a group, the JS animals exhibited sig-nificantly elevated mean HR (F(3,116)=54.2, pb0.0001) anddecreased HRV (F(3,116)=10.5, pb0.0001) along the timeaxis. Post hoc Bonferroni test revealed that stress signifi-cantly increased mean HR and decreased HRV duringexposure as compared to values for baseline (pb0.00001
for HR and HRV), day 60 (pb0.001) and day 80 (pb0.0002).However, by day 60, the mean HR and HRV had returned tolevels not significantly different from baseline and remainedconstant.
The AS group: This group was first exposed to the stressoron day 60 and exhibited significantly elevated HR (F(3,116)=43.4, pb0.0001) and decreased HRV (F(3,116)=38.76,pb0.0001) during the stress. Immediately after the exposure(within 2–3 h), HR remained high while HRV returned rapidlyto baseline levels. Twenty days later, on day 80, the mean HRhad also returned to baseline levels.
The JS+AS group: The repeated stress exposure in youthand adulthood elicited significantly elevated HR (F(3,116)=33.08, pb0.0001) and decreased HRV (F(3,116)=17.4,pb0.0001) beginning during exposure and maintainedunchanged throughout.
3.6.3. Correlation of behavioral parameters and HR, HRVIn order to assess the relationship between behavioralresponses and physiologic functioning, correlations betweentime spent in the open arms and startle amplitudes versus HRand HRV were computed.
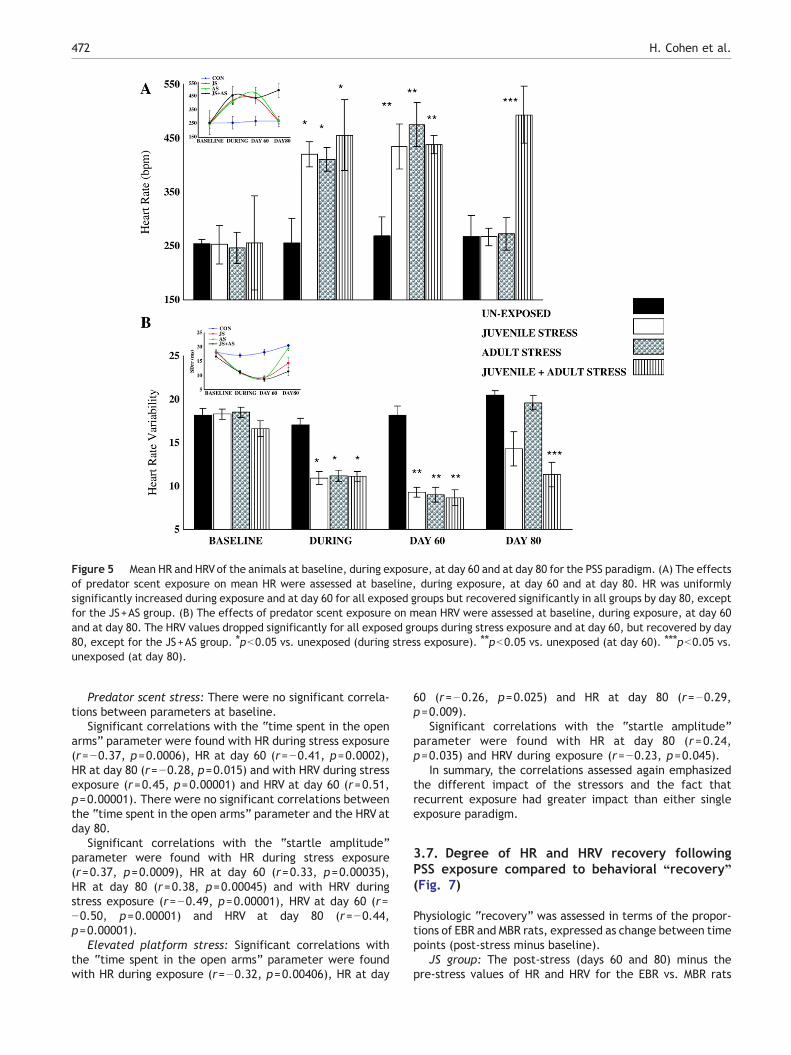
Figure 5 Mean HR and HRVof the animals at baseline, during exposure, at day 60 and at day 80 for the PSS paradigm. (A) The effectsof predator scent exposure on mean HR were assessed at baseline, during exposure, at day 60 and at day 80. HR was uniformlysignificantly increased during exposure and at day 60 for all exposed groups but recovered significantly in all groups by day 80, exceptfor the JS+AS group. (B) The effects of predator scent exposure on mean HRV were assessed at baseline, during exposure, at day 60and at day 80. The HRV values dropped significantly for all exposed groups during stress exposure and at day 60, but recovered by day80, except for the JS+AS group. ⁎pb0.05 vs. unexposed (during stress exposure). ⁎⁎pb0.05 vs. unexposed (at day 60). ⁎⁎⁎pb0.05 vs.unexposed (at day 80).
472 H. Cohen et al.
Predator scent stress: There were no significant correla-tions between parameters at baseline.
Significant correlations with the “time spent in the openarms” parameter were found with HR during stress exposure(r=−0.37, p=0.0006), HR at day 60 (r=−0.41, p=0.0002),HR at day 80 (r=−0.28, p=0.015) and with HRV during stressexposure (r=0.45, p=0.00001) and HRV at day 60 (r=0.51,p=0.00001). There were no significant correlations betweenthe “time spent in the open arms” parameter and the HRV atday 80.
Significant correlations with the “startle amplitude”parameter were found with HR during stress exposure(r=0.37, p=0.0009), HR at day 60 (r=0.33, p=0.00035),HR at day 80 (r=0.38, p=0.00045) and with HRV duringstress exposure (r=−0.49, p=0.00001), HRV at day 60 (r=−0.50, p=0.00001) and HRV at day 80 (r=−0.44,p=0.00001).
Elevated platform stress: Significant correlations withthe “time spent in the open arms” parameter were foundwith HR during exposure (r=−0.32, p=0.00406), HR at day
60 (r=−0.26, p=0.025) and HR at day 80 (r=−0.29,p=0.009).
Significant correlations with the “startle amplitude”parameter were found with HR at day 80 (r=0.24,p=0.035) and HRV during exposure (r=−0.23, p=0.045).
In summary, the correlations assessed again emphasizedthe different impact of the stressors and the fact thatrecurrent exposure had greater impact than either singleexposure paradigm.
3.7. Degree of HR and HRV recovery followingPSS exposure compared to behavioral “recovery”(Fig. 7)
Physiologic “recovery” was assessed in terms of the propor-tions of EBR and MBR rats, expressed as change between timepoints (post-stress minus baseline).
JS group: The post-stress (days 60 and 80) minus thepre-stress values of HR and HRV for the EBR vs. MBR rats
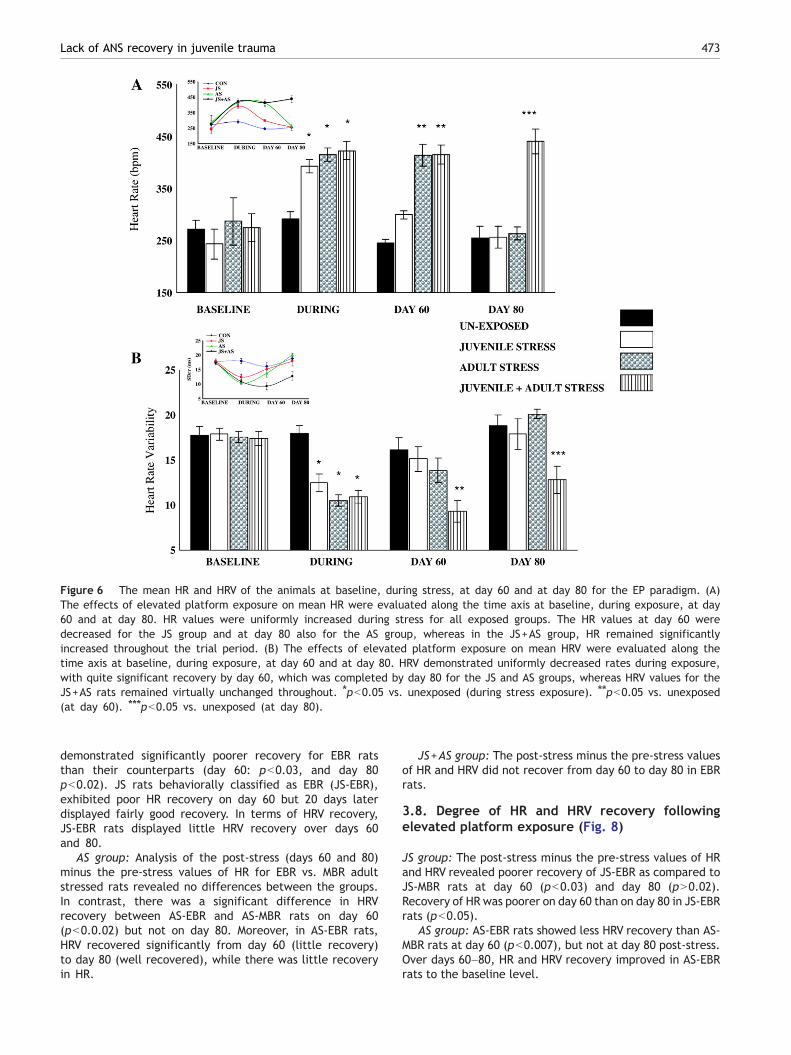
Figure 6 The mean HR and HRV of the animals at baseline, during stress, at day 60 and at day 80 for the EP paradigm. (A)The effects of elevated platform exposure on mean HR were evaluated along the time axis at baseline, during exposure, at day60 and at day 80. HR values were uniformly increased during stress for all exposed groups. The HR values at day 60 weredecreased for the JS group and at day 80 also for the AS group, whereas in the JS+AS group, HR remained significantlyincreased throughout the trial period. (B) The effects of elevated platform exposure on mean HRV were evaluated along thetime axis at baseline, during exposure, at day 60 and at day 80. HRV demonstrated uniformly decreased rates during exposure,with quite significant recovery by day 60, which was completed by day 80 for the JS and AS groups, whereas HRV values for theJS+AS rats remained virtually unchanged throughout. ⁎pb0.05 vs. unexposed (during stress exposure). ⁎⁎pb0.05 vs. unexposed(at day 60). ⁎⁎⁎pb0.05 vs. unexposed (at day 80).
473Lack of ANS recovery in juvenile trauma
demonstrated significantly poorer recovery for EBR ratsthan their counterparts (day 60: pb0.03, and day 80pb0.02). JS rats behaviorally classified as EBR (JS-EBR),exhibited poor HR recovery on day 60 but 20 days laterdisplayed fairly good recovery. In terms of HRV recovery,JS-EBR rats displayed little HRV recovery over days 60and 80.
AS group: Analysis of the post-stress (days 60 and 80)minus the pre-stress values of HR for EBR vs. MBR adultstressed rats revealed no differences between the groups.In contrast, there was a significant difference in HRVrecovery between AS-EBR and AS-MBR rats on day 60(pb0.0.02) but not on day 80. Moreover, in AS-EBR rats,HRV recovered significantly from day 60 (little recovery)to day 80 (well recovered), while there was little recoveryin HR.
JS+AS group: The post-stress minus the pre-stress valuesof HR and HRV did not recover from day 60 to day 80 in EBRrats.
3.8. Degree of HR and HRV recovery followingelevated platform exposure (Fig. 8)
JS group: The post-stress minus the pre-stress values of HRand HRV revealed poorer recovery of JS-EBR as compared toJS-MBR rats at day 60 (pb0.03) and day 80 (pN0.02).Recovery of HR was poorer on day 60 than on day 80 in JS-EBRrats (pb0.05).
AS group: AS-EBR rats showed less HRV recovery than AS-MBR rats at day 60 (pb0.007), but not at day 80 post-stress.Over days 60–80, HR and HRV recovery improved in AS-EBRrats to the baseline level.
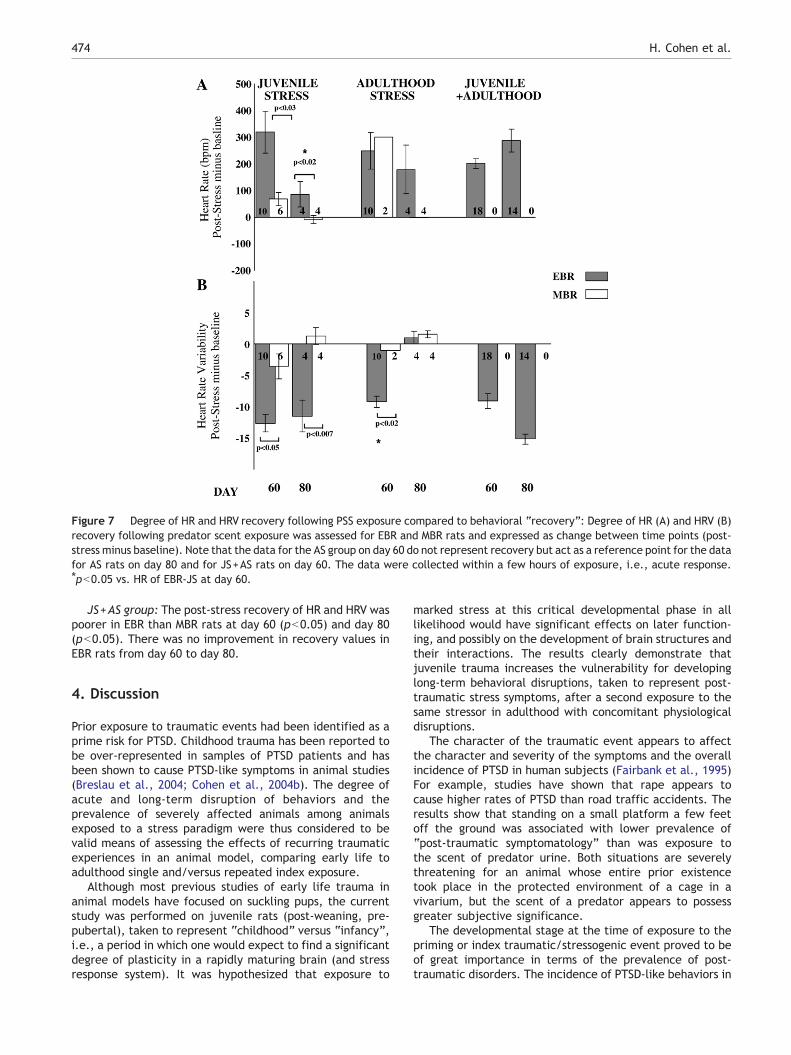
Figure 7 Degree of HR and HRV recovery following PSS exposure compared to behavioral “recovery”: Degree of HR (A) and HRV (B)recovery following predator scent exposure was assessed for EBR and MBR rats and expressed as change between time points (post-stress minus baseline). Note that the data for the AS group on day 60 do not represent recovery but act as a reference point for the datafor AS rats on day 80 and for JS+AS rats on day 60. The data were collected within a few hours of exposure, i.e., acute response.⁎pb0.05 vs. HR of EBR-JS at day 60.
474 H. Cohen et al.
JS+AS group: The post-stress recovery of HR and HRV waspoorer in EBR than MBR rats at day 60 (pb0.05) and day 80(pb0.05). There was no improvement in recovery values inEBR rats from day 60 to day 80.
4. Discussion
Prior exposure to traumatic events had been identified as aprime risk for PTSD. Childhood trauma has been reported tobe over-represented in samples of PTSD patients and hasbeen shown to cause PTSD-like symptoms in animal studies(Breslau et al., 2004; Cohen et al., 2004b). The degree ofacute and long-term disruption of behaviors and theprevalence of severely affected animals among animalsexposed to a stress paradigm were thus considered to bevalid means of assessing the effects of recurring traumaticexperiences in an animal model, comparing early life toadulthood single and/versus repeated index exposure.
Although most previous studies of early life trauma inanimal models have focused on suckling pups, the currentstudy was performed on juvenile rats (post-weaning, pre-pubertal), taken to represent “childhood” versus “infancy”,i.e., a period in which one would expect to find a significantdegree of plasticity in a rapidly maturing brain (and stressresponse system). It was hypothesized that exposure to
marked stress at this critical developmental phase in alllikelihood would have significant effects on later function-ing, and possibly on the development of brain structures andtheir interactions. The results clearly demonstrate thatjuvenile trauma increases the vulnerability for developinglong-term behavioral disruptions, taken to represent post-traumatic stress symptoms, after a second exposure to thesame stressor in adulthood with concomitant physiologicaldisruptions.
The character of the traumatic event appears to affectthe character and severity of the symptoms and the overallincidence of PTSD in human subjects (Fairbank et al., 1995)For example, studies have shown that rape appears tocause higher rates of PTSD than road traffic accidents. Theresults show that standing on a small platform a few feetoff the ground was associated with lower prevalence of“post-traumatic symptomatology” than was exposure tothe scent of predator urine. Both situations are severelythreatening for an animal whose entire prior existencetook place in the protected environment of a cage in avivarium, but the scent of a predator appears to possessgreater subjective significance.
The developmental stage at the time of exposure to thepriming or index traumatic/stressogenic event proved to beof great importance in terms of the prevalence of post-traumatic disorders. The incidence of PTSD-like behaviors in
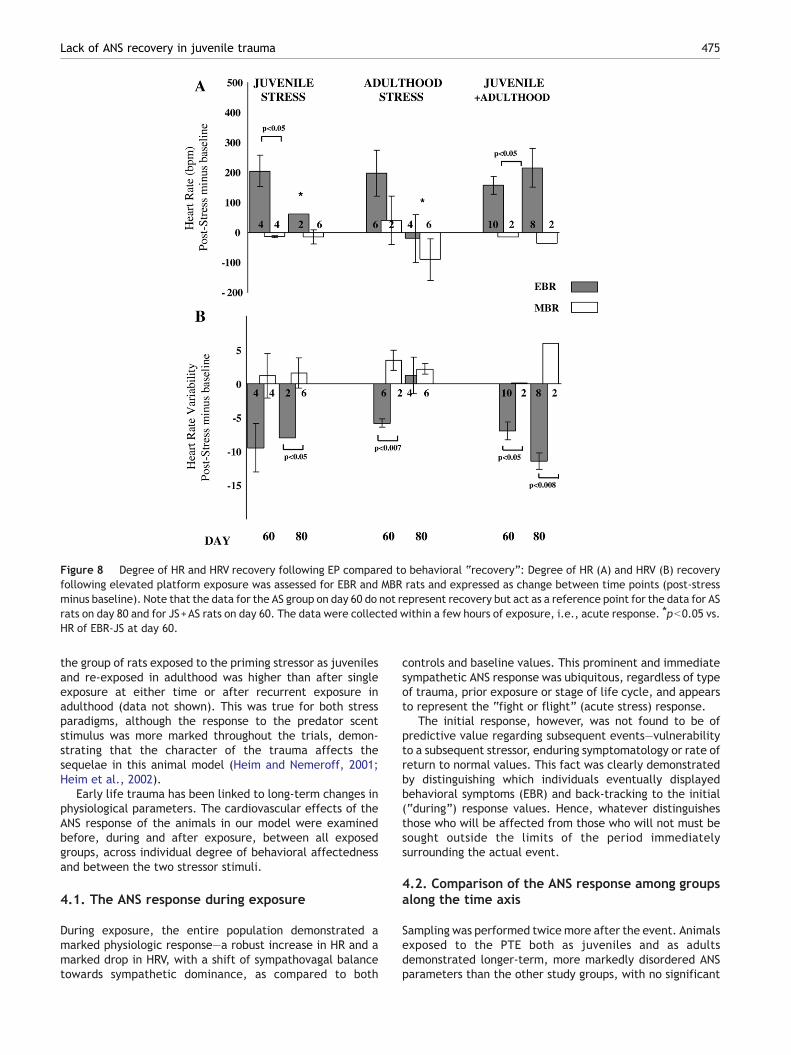
Figure 8 Degree of HR and HRV recovery following EP compared to behavioral “recovery”: Degree of HR (A) and HRV (B) recoveryfollowing elevated platform exposure was assessed for EBR and MBR rats and expressed as change between time points (post-stressminus baseline). Note that the data for the AS group on day 60 do not represent recovery but act as a reference point for the data for ASrats on day 80 and for JS+AS rats on day 60. The data were collected within a few hours of exposure, i.e., acute response. ⁎pb0.05 vs.HR of EBR-JS at day 60.
475Lack of ANS recovery in juvenile trauma
the group of rats exposed to the priming stressor as juvenilesand re-exposed in adulthood was higher than after singleexposure at either time or after recurrent exposure inadulthood (data not shown). This was true for both stressparadigms, although the response to the predator scentstimulus was more marked throughout the trials, demon-strating that the character of the trauma affects thesequelae in this animal model (Heim and Nemeroff, 2001;Heim et al., 2002).
Early life trauma has been linked to long-term changes inphysiological parameters. The cardiovascular effects of theANS response of the animals in our model were examinedbefore, during and after exposure, between all exposedgroups, across individual degree of behavioral affectednessand between the two stressor stimuli.
4.1. The ANS response during exposure
During exposure, the entire population demonstrated amarked physiologic response—a robust increase in HR and amarked drop in HRV, with a shift of sympathovagal balancetowards sympathetic dominance, as compared to both
controls and baseline values. This prominent and immediatesympathetic ANS response was ubiquitous, regardless of typeof trauma, prior exposure or stage of life cycle, and appearsto represent the “fight or flight” (acute stress) response.
The initial response, however, was not found to be ofpredictive value regarding subsequent events—vulnerabilityto a subsequent stressor, enduring symptomatology or rate ofreturn to normal values. This fact was clearly demonstratedby distinguishing which individuals eventually displayedbehavioral symptoms (EBR) and back-tracking to the initial(“during”) response values. Hence, whatever distinguishesthose who will be affected from those who will not must besought outside the limits of the period immediatelysurrounding the actual event.
4.2. Comparison of the ANS response among groupsalong the time axis
Sampling was performed twice more after the event. Animalsexposed to the PTE both as juveniles and as adultsdemonstrated longer-term, more markedly disordered ANSparameters than the other study groups, with no significant

476 H. Cohen et al.
tendency towards recovery. The exposed animals in the othertwo groups demonstrated significant rates of physiologicaland behavioral recovery, with HR and HRV returning back tobaseline values (pre-exposure) more rapidly for adult thanfor juvenile exposure.
The degrees of increase in HR and of decrease in HRVdid not differ between the types of trauma/stressor duringexposure, yet recovery after exposure to the “stronger”stressor (predator odor) took longer, as measured byprevalence of PTSD-like response behavior and ANS para-meters. In the rats exposed to juvenile trauma only,exposure to the elevated platform produced fewer EBRrats, with a more rapid recovery curve, than theircounterparts exposed to the predator scent stimulus, whodemonstrated a higher prevalence of extremely affected(EBR) individuals, with significantly slower rates of recov-ery (day 80).
These data imply that PTSD can be thought of asstemming, at least in part, from a faulty recovery processand appear to demonstrate distinctions related to the impactof the event (akin to a “dose–response”).
4.3. The ANS response across individual differencesas expressed by degree of behavioral affectedness
Since the proportions of behaviorally and physiologicallymore affected animals appeared to be correlated, “recov-ery”, defined as post-stress minus pre-stress values of HRand HRV, was assessed for the rats whose behavior wasgrossly affected (EBR) and for those whose behavior wasunaffected (MBR). The post-stress (day 60 and day 80) minusthe pre-stress values of HR and HRV for the EBR vs. MBR ratsdemonstrated significantly poorer physiologic recovery forthe behaviorally EBR rats than for their behaviorallyunaffected counterparts.
Rats exposed to the PSS paradigm in youth and again inadulthood and whose behavior was significantly affected(EBR), displayed only minimal HR recovery or none at all andcould be said to have demonstrated an exacerbatedresponse. Animals exposed once, either in youth or inadulthood only, displayed fairly good recovery along thetime axis.
In terms of HRV, the overall patterns demonstrateenduring effects after juvenile exposure. The data forexposed juveniles that exhibited EBR show no HRV recoveryat all over the entire study period. Thus, for rats exposed inyouth and re-exposed in adulthood, the second exposure hada very obvious exacerbating effect and occurred whichoccurs before there had been significant recovery from theinitial exposure.
In general, HR values tended to drop back toward baselinevalues sooner than HRV values, especially in the behaviorallyaffected individuals. This finding concurs with data fromstudies on human subjects and (although a population sizeeffect may have affected these findings) the trend was quiterobust and has physiological implications.
The ANS links the central nervous system (CNS) and thecardiovascular system. HRV reflects fluctuations in autonom-ic inputs to the heart rather than the mean level ofautonomic inputs (Task Force, 1996). Thus, both autonomicdown-regulation and a high level of sympathetic input to thepoint of saturation lead to diminished HRV. One may
speculate that our findings reflect a dysfunction of CNSregulation in juvenile exposed animals rather than aperipheral abnormality. A variable HR may act as a bufferagainst challenges to homeostasis. Animals exposed totrauma in youth and re-traumatized in adulthood exhibitabnormal HR at day 80. This may reflect a cortical control ofthe ANS that is insufficiently buffered to withstand stressfulchallenges.
The results of the present study suggest that exposure tostressogenic/traumatic events during youth can have signif-icant and long-lasting effects on the autonomic cardiovas-cular system in adulthood correlated with gross behavioraldisruption. Rats exposed to juvenile stress and re-exposedlater in life suffered from a sensitization response expressedas an excessive cardiovascular responsiveness of the auto-nomic neural pathways.
4.4. Limitations
One must take care not to be too literal in interpretinganimal models. They are not to be taken to comprehen-sively reflect the human disorder but rather to approxi-mate certain aspects of it. The human clinical syndrome ofPTSD involves factors beyond the scope of a physiologicalimbalance or change in behavior alone, which this animalmodel is unable to approximate. It would be presumptuousto assume that the “criteria” applied in this study in factreflect behavioral–physiological parameters in the life ofthe rat, which are commensurate with the emotional,physiological and existential implications of criteria forPTSD in humans. The authors are not proposing that thecriteria described or the behavioral paradigms used are theonly appropriate ones for defining CBCs. The rationale ofthe principle is to make better sense the de facto range ofbehavioral observations.
The HR sampling method used required the insertion of asubcutaneous probe. The procedure causes little or no painand requires only brief immobilization; however, it isinvasive and a little uncomfortable, in and of itself. Futurestudies will be performed using telemetric probes insertedunder anesthesia.
The overall sample size is sufficient for statisticalanalysis, but larger samples would be preferable.
4.5. Conclusions
Exposure to traumatic events in childhood has resoundingeffects on behavioral morbidity and the ANS in later life andpredisposes subjects to more significant pathology uponadulthood re-exposure. The predominant difference in thecourse of the sequelae of traumatic events between ratswhose behavior is significantly affected and those whoappear unaffected is the rate of behavioral and physiologicalrecovery from the (ubiquitous) immediate response duringthe event.
Acknowledgment
The work was supported by the Israel Academy of Sciencesand Humanities (#429/03) and the NARSAD independentaward to H.C.

477Lack of ANS recovery in juvenile trauma
References
Akselrod, S., Gordon, D., Shannon, D., Barger, A., Cohen, R., 1981.Power spectral analysis of heart rate fluctuation: a quantitativeprobe of beat-to beat cardiovascular control. Science 213,220–222.
American Psychiatric Association, 1994. Diagnostic and StatisticalManual of Mental Disorders, 4 ed. American Psychiatric Associ-ation, Washington, DC.
Avital, A., Richter-Levin, G., 2005. Exposure to juvenile stressexacerbates the behavioural consequences of exposure to stressin the adult rat. Int. J. Neuropsychopharmacol. 8, 163–173 (Epub2004 Nov 17).
Breslau, N., 2002. Psychiatric morbidity in adult survivors ofchildhood trauma. Semin. Clin. Neuropsychiatry 7, 80–88.
Breslau, N., Davis, G.C., Andreski, P., Peterson, E., 1991. Traumaticevents and posttraumatic stress disorder in an urban populationof young adults. Arch. Gen. Psychiatry 48, 216–222.
Breslau, N., Wilcox, H.C., Storr, C.L., Lucia, V.C., Anthony, J.C.,2004. Trauma exposure and posttraumatic stress disorder: astudy of youths in urban America. J. Urban Health 81, 530–544.
Caldji, C., Diorio, J., Meaney, M.J., 2000. Variations in maternal carein infancy regulate the development of stress reactivity. Biol.Psychiatry 48, 1164–1174.
Carney, R.M., Saunders, R.D., Freedland, K.E., Stein, P., Rich, M.W.,Jaffe, A.S., 1995. Association of depression with reduced heartrate variability in coronary artery disease. Am. J. Cardiol. 76,562–564.
Cohen, H., Joseph, Z., Matar, M., 2003. The relevance of differentialresponse to trauma in an animal model of post-traumatic stressdisorder. Biol. Psychiatry 15, 463–473.
Cohen, H., Maayan, R., Touati-Werner, D., et al., 2006a. Decreasedcirculatory levels of neuroactive steroids in behaviourally moreextremely affected rats subsequent to exposure to a potentiallytraumatic experience. Int. J. Neuropsychopharmacol. 1–7.
Cohen, H., Matar, M.A., Richter-Levin, G., Zohar, J., 2006b. Thecontribution of an animal model toward uncovering biologicalrisk factors for PTSD. Ann. N.Y. Acad Sci. 1071, 335–350.
Cohen, H., Zohar, J., 2004. Animal models of post traumatic stressdisorder: the use of cut off behavioral criteria. Ann. N.Y. Acad.Sci. 1032, 167–178.
Cohen, H., Zohar, J., Gidron, Y., et al., 2006c. Blunted HPA axisresponse to stress influences susceptibility to posttraumaticstress response in rats. Biol. Psychiatry 59, 1208–1218.
Cohen, H., Zohar, J., Matar, M.A., Kaplan, Z., Geva, A.B., 2005.Unsupervised fuzzy clustering analysis supports behavioral cutoffcriteria in an animal model of posttraumatic stress disorder. Biol.Psychiatry 58, 640–651.
Cohen, H., Zohar, J., Matar, M.A., Zeev, K., Loewenthal, U., Richter-Levin, G., 2004a. Setting apart the affected: the use ofbehavioral criteria in animal models of post traumatic stressdisorder. Neuropsychopharmacology 29, 1962–1970.
Cohen, J., Goodman, R.F., Brown, E.J., Mannarino, A., 2004b.Treatment of childhood traumatic grief: contributing to a newlyemerging condition in the wake of community trauma. Harv. Rev.Psychiatry 12, 213–216.
Elghozi, J.L., Girard, A., Laude, D., 2001. Effects of drugs on theautonomic control of short-term heart rate variability. Auton.Neurosci. 90, 116–121.
Fairbank, J.A., Schlenger, W.E., Saigh, P.A., Davidson, J.R.T., 1995.An epidemiologic profile of post-traumatic stress disorder:prevalence, comorbidity, and risk factors. In: M.J.F., D.S.C., A.Y.D. (Eds.), Neurobiological and Clinical Consequences of Stress:
From Normal Adaptation to PTSD. Lippincott-Raven, Philadel-phia, pp. 415–427.
File, S.E., 1993. The interplay of learning and anxiety in the elevatedplus-maze. Behav. Brain Res. 58, 199–202.
Ford, J.D., Kidd, P., 1998. Early childhood trauma and disorders ofextreme stress as predictors of treatment outcome with chronicposttraumatic stress disorder. J. Trauma. Stress 11, 743–761.
Francis, D.D., Caldji, C., Champagne, F., Plotsky, P.M., Meaney, M.J.,1999. The role of corticotropin-releasing factor-norepinephrinesystems in mediating the effects of early experience on thedevelopment of behavioral and endocrine responses to stress.Biol. Psychiatry 46, 1153–1166.
Freedman, S.A., Gluck, N., Tuval-Mashiach, R., Brandes, D., Peri, T.,Shalev, A.Y., 2002. Gender differences in responses to traumaticevents: a prospective study. J. Trauma. Stress 15, 407–413.
Goldberg, S., Levitan, R., Leung, E., et al., 2003. Cortisolconcentrations in 12- to 18-month-old infants: stability overtime, location, and stressor. Biol. Psychiatry 54, 719–726.
Heidbreder, C.A., Weiss, I.C., Domeney, A.M., et al., 2000.Behavioral, neurochemical and endocrinological characteriza-tion of the early social isolation syndrome. Neuroscience 100,749–768.
Heim, C., Nemeroff, C.B., 2001. The role of childhood trauma in theneurobiology of mood and anxiety disorders: preclinical andclinical studies. Biol. Psychiatry 49, 1023–1039.
Heim, C., Newport, D.J., Wagner, D., Wilcox, M.M., Miller, A.H.,Nemeroff, C.B., 2002. The role of early adverse experience andadulthood stress in the prediction of neuroendocrine stressreactivity in women: a multiple regression analysis. Depress.Anxiety 15, 117–125.
Malliani, A., Pagani, M., Lombardi, F., Cerutti, S., 1991. Cardiovas-cular neural regulation explored in the frequency domain.Circulation 84, 482–492.
Maslova, L.N., Bulygina, V.V., Markel, A.L., 2002a. Chronic stressduring prepubertal development: immediate and long-lastingeffects on arterial blood pressure and anxiety-related behavior.Psychoneuroendocrinology 27, 549–561.
Maslova, L.N., Bulygina, V.V., Popova, N.K., 2002b. Immediate andlong-lasting effects of chronic stress in the prepubertal age onthe startle reflex. Physiol. Behav. 75, 217–225.
McEwen, B.S., 2001. Plasticity of the hippocampus: adaptation tochronic stress and allostatic load. Ann. N.Y. Acad Sci. 933,265–277.
McEwen, B.S., 2002. The neurobiology and neuroendocrinology ofstress. Implications for post-traumatic stress disorder from abasic science perspective. Psychiatr. Clin. North Am. 25,469–494.
Resnick, H.S., Kilpatrick, D.G., Dansky, B.S., Saunders, B.E., Best, C.L., 1993. Prevalence of civilian trauma and posttraumatic stressdisorder in a representative national sample of women. J.Consult. Clin. Psychol. 61, 984–991.
Shea, A., Walsh, C., Macmillan, H., Steiner, M., 2005. Childmaltreatment and HPA axis dysregulation: relationship to majordepressive disorder and post traumatic stress disorder infemales. Psychoneuroendocrinology 30, 162–178.
Stein, P., Kleiger, R., 1999. Insights from the study of heart ratevariability. Annu. Rev. Med. 50, 249–261.
Task Force, 1996. Heart rate variability. Standards of measurement,physiological interpretation, and clinical use. Task Force of theEuropean Society of Cardiology and the North American Societyof Pacing and Electrophysiology. Eur. Heart J. 17, 354–381.




















