
Living on the edge: demographic and phylogeographical patterns in the woodlouse-hunter spider...
17
Molecular Ecology (2007) 16, 3198–3214 doi: 10.1111/j.1365-294X.2007.03351.x © 2007 The Authors Journal compilation © 2007 Blackwell Publishing Ltd Blackwell Publishing Ltd Living on the edge: demographic and phylogeographical patterns in the woodlouse-hunter spider Dysdera lancerotensis Simon, 1907 on the eastern volcanic ridge of the Canary Islands LETICIA BIDEGARAY-BATISTA,* NURIA MACÍAS-HERNÁNDEZ,*† PEDRO OROMÍ † and MIQUEL A. ARNEDO * *Departament de Biologia Animal, Universitat de Barcelona, Av. Diagonal 645, 08028, Barcelona, Spain, †Departamento de Biologia Animal, Universidad de La Laguna, Universidad de La Laguna, 38206, La Laguna, Tenerife, Canary Islands Abstract The Eastern Canary Islands are the emerged tips of a continuous volcanic ridge running parallel to the northeastern African coast, originated by episodic volcanic eruptions that can be traced back to the Miocene and that, following a major period of quiescence and erosion, continued from the Pliocene to the present day. The islands have been periodically connected by eustatic sea-level changes resulting from Pleistocene glacial cycles. The ground-dwelling spider Dysdera lancerotensis Simon, 1907 occurs along the entire ridge, except on recent barren lavas and sand dunes, and is therefore an ideal model for studying the effect of episodic geological processes on terrestrial organisms. Nested clade and population genetic analyses using 39 haplotypes from 605 base pairs of mitochondrial DNA cytochrome c oxidase I sequence data, along with phylogenetic analyses including two additional mitochondrial genes, uncover complex phylogeographical and demographic patterns. Our results indicate that D. lancerotensis colonized the ridge from north to south, in contrast to what had been expected given the SSW-NNE trend of volcanism and to what had been reported for other terrestrial arthropods. The occurrence of several episodes of extinction, recolonization and expansion are hypothesized for this species, and areas that act as refugia during volcanic cycles are identified. Relaxed molecular clock methods reveal divergence times between main haplotype lineages that suggest an older origin of the northern islets than anticipated based on geological evidence. This study supports the key role of volcanism in shaping the distribution of terrestrial organisms on oceanic islands and generates phylogeographical predictions that warrant further research into other terrestrial endemisms of this fascinat- ing region. Keywords: Canary Islands, mitochondrial DNA, phylogeography, population fragmentation, volcanism Received 9 December 2006; revision accepted 20 March 2007 Introduction Volcanic islands have long captivated the attention of evolutionary biologists because of the experimental-like conditions they provide for the study of speciation (Mayr 1967; Wagner & Funk 1995; Grant 1998; Juan et al . 2000). More recently, volcanic islands have become fertile ground for phylogeography, the study of the causes behind the current distribution of genetic diversity (Avise 1998). The highly dynamic and accurately dated geology of volcanic islands make them ideal models for testing the role of historical fragmentation and recolonization in shaping contemporary patterns of the geographical distributions of species. Several studies have provided evidence of the close relationship between volcanic activity and lineage distributions. Forest fragmentation caused by recent lava flow from the Mauna Loa volcano on the island of Hawaii has affected the pattern of neutral genetic variation in several species of endemic Tetragnatha spiders, with a Correspondence: Miquel A. Arnedo Fax: +34 93 403-5740. E-mail: [email protected].
Transcript of Living on the edge: demographic and phylogeographical patterns in the woodlouse-hunter spider...

Molecular Ecology (2007)
16
, 3198–3214 doi: 10.1111/j.1365-294X.2007.03351.x
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Blackwell Publishing Ltd
Living on the edge: demographic and phylogeographical patterns in the woodlouse-hunter spider
Dysdera lancerotensis
Simon, 1907 on the eastern volcanic ridge of the Canary Islands
LETICIA BIDEGARAY-BATISTA,
*
NURIA MACÍAS-HERNÁNDEZ,
*†
PEDRO OROMÍ
†
and MIQUEL A. ARNEDO
*
*
Departament de Biologia Animal, Universitat de Barcelona, Av. Diagonal 645, 08028, Barcelona, Spain
, †
Departamento de Biologia Animal, Universidad de La Laguna, Universidad de La Laguna, 38206, La Laguna, Tenerife, Canary Islands
Abstract
The Eastern Canary Islands are the emerged tips of a continuous volcanic ridge runningparallel to the northeastern African coast, originated by episodic volcanic eruptions that canbe traced back to the Miocene and that, following a major period of quiescence and erosion,continued from the Pliocene to the present day. The islands have been periodically connectedby eustatic sea-level changes resulting from Pleistocene glacial cycles. The ground-dwellingspider
Dysdera lancerotensis
Simon, 1907 occurs along the entire ridge, except on recent barrenlavas and sand dunes, and is therefore an ideal model for studying the effect of episodicgeological processes on terrestrial organisms. Nested clade and population genetic analysesusing 39 haplotypes from 605 base pairs of mitochondrial DNA cytochrome
c
oxidase Isequence data, along with phylogenetic analyses including two additional mitochondrialgenes, uncover complex phylogeographical and demographic patterns. Our results indicatethat
D. lancerotensis
colonized the ridge from north to south, in contrast to what had beenexpected given the SSW-NNE trend of volcanism and to what had been reported for otherterrestrial arthropods. The occurrence of several episodes of extinction, recolonization andexpansion are hypothesized for this species, and areas that act as refugia during volcaniccycles are identified. Relaxed molecular clock methods reveal divergence times betweenmain haplotype lineages that suggest an older origin of the northern islets than anticipatedbased on geological evidence. This study supports the key role of volcanism in shaping thedistribution of terrestrial organisms on oceanic islands and generates phylogeographicalpredictions that warrant further research into other terrestrial endemisms of this fascinat-ing region.
Keywords
: Canary Islands, mitochondrial DNA, phylogeography, population fragmentation, volcanism
Received 9 December 2006; revision accepted 20 March 2007
Introduction
Volcanic islands have long captivated the attention ofevolutionary biologists because of the experimental-likeconditions they provide for the study of speciation (Mayr1967; Wagner & Funk 1995; Grant 1998; Juan
et al
. 2000).More recently, volcanic islands have become fertile groundfor phylogeography, the study of the causes behind the
current distribution of genetic diversity (Avise 1998). Thehighly dynamic and accurately dated geology of volcanicislands make them ideal models for testing the role ofhistorical fragmentation and recolonization in shapingcontemporary patterns of the geographical distributions ofspecies. Several studies have provided evidence of theclose relationship between volcanic activity and lineagedistributions. Forest fragmentation caused by recent lavaflow from the Mauna Loa volcano on the island of Hawaiihas affected the pattern of neutral genetic variation inseveral species of endemic
Tetragnatha
spiders, with a
Correspondence: Miquel A. Arnedo Fax: +34 93 403-5740. E-mail:[email protected].

P H Y L O G E O G R A P H Y O F
D Y S D E R A L A N C E R O T E N S I S
3199
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
greater influence on specialist than on generalist species(Vandergast
et al
. 2004). Research on
Brachyderes rugatusrugatus
, an endemic weevil from the island of La Palma inthe Canary Islands, has revealed correlations between phylo-geographical patterns and volcanic activity. However,some of the results did not match available geologicalevidence, which suggests that some of the volcanic terrainsare older than what geologists had previously assumed(Emerson
et al
. 2006). These results emphasize the positivefeedback between geological and phylogeographicalinferences. Phylogeographical analysis has identified asevere bottleneck in the giant tortoise
Geochelone nigravandenburghi
on Alcedo Volcano, in the Galapagos Islands,which it has been suggested, is the result of a contractionof the tortoise population caused by the prehistoric eruptionof the volcano, and the subsequent recolonization via anancestral haplotype (Beheregaray
et al
. 2003).The volcanic archipelago of the Canary Islands lies in the
Atlantic Ocean off the northwestern coast of Africa (Fig. 1).The archipelago was formed on a hot spot, hence thearrangement of its seven main islands in a linear geograph-ical and temporal cline: islands are older to the east andbecome younger towards the west (Carracedo
et al
. 1998).
The Eastern Canaries comprise the islands of Lanzaroteand Fuerteventura and the islet of Lobos (located betweenthese two larger islands), as well as La Graciosa, MontañaClara and Alegranza (to the north of Lanzarote). They arethe emerged parts of a volcanic ridge that runs roughlyparallel to the African coast, only 100 km away at its closestpoint (Coello
et al
. 1992). Although nowadays the mainislands and islets are separated, they have been periodicallyconnected and disconnected by marine transgressionsassociated with Pleistocene climate oscillations (Carracedo
et al
. 2003). The subaerial volcanism that formed the islandsstarted ~20 million years ago (Ma) and took place in twodistinct volcanic cycles with a SSW-NNE temporal andspatial polarity (Coello
et al
. 1992). The eruptive cycleswere separated by a significant gap, which resulted in along erosion period. Post-Miocene activity was restrictedto Lanzarote, the northern islets and the central and north-ern parts of Fuerteventura. Holocene volcanic activity hasbeen documented in the islands (Carracedo
et al
. 1992), andboth recent and historical eruptions have covered vastareas of the northeast and western central parts of Lanzarotewith lava flows (Carracedo
et al
. 2003). The Eastern Canariesare characterized by more arid and xerophilous habitatsthan their Western counterparts due to the joint effect ofstrong erosion, which prevents to capture the humidnortheast trade winds, and the dry and dusty winds blow-ing from the nearby Sahara Desert ( Juan
et al
. 2000).The geological and climatic features make the Eastern
Canary Islands an ideal model for investigating the influ-ence of volcanism and sea-level fluctuations in shaping thegeographical distribution of species, lineage diversification,population structure and demography. Lava flows andmarine transgression may cause local extinctions andpopulation fragmentations, but they also create newlyavailable recolonization sites and subsequent populationexpansions. In spite of this, very few studies have dealtwith the phylogeographical patterns of Eastern Canarianendemics, while in recent years the Western Islands havebeen the object of in-depth research in this field (Rees
et al
.2001a, b; Contreras-Díaz
et al
. 2003; Moya
et al
. 2004; Emerson
et al
. 2006; Moya
et al
. 2006). In the only example of popu-lation level molecular analysis of an Eastern Canarian endemic,Juan
et al
. (1998) found that the phylogeographical patternof the darkling beetle
Hegeter deyrollei
(formerly
Hegeter politus
)closely matched the SSW-NNE sequence of subaerialvolcanic activity and suggested a sequential colonizationof the eastern ridge from Jandía, in southern Fuerteventura,to the northernmost islet of Alegranza. More recently,Bloor & Brown (2005) examined the effect of recent lavaflows (locally known as
malpaís
) on the morphologicalvariation of the eastern endemic lizard
Gallotia atlantica
. Theyconcluded that the low level of morphological differentia-tion identified two major groups, one spread throughoutmost of Lanzarote and the islets, and a second restricted to
Fig. 1 Map of the Eastern Canary Islands showing samplinglocations of Dysdera lancerotensis. The population codes correspondto those listed in Table 1.

3200
L . B I D E G A R A Y - B A T I S T A
E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
two isolated populations in recent lava flows: Malpaís de laCorona and southeast of Haría, and two localities in thesouthwest of Timanfaya. The authors suggested that thisvariation in
Malpaís
populations would have developedrecently, rather than being the relicts of a former widespreadpopulation.
The woodlouse-hunter spider
Dysdera lancerotensis
Simon,1907 is endemic to the Eastern Canary Islands, where it iswidespread throughout Fuerteventura, Lanzarote and allthe islets. Like its congeneric species,
D. lancerotensis
is aground-dwelling, nocturnal-wandering hunter. The genus
Dysdera
has undergone a major evolutionary radiationprocess in the Canaries, where 43 endemic species havebeen identified (Arnedo & Ribera 1997; Arnedo & Ribera1999; Arnedo
et al
. 2000; Arnedo
et al
. 2001). Although thenumber of colonization events that account for this out-standing diversity remains a matter of debate, it has beenclearly shown that
D. lancerotensis
is not closely related toany of the other Canarian endemics, and that its closestrelatives are in fact found in Morocco (Arnedo
et al
. 2001).Hence,
D. lancerotensis
is most likely to be the result ofan independent colonization by African ancestors. It wasoriginally described as a subspecies of the cosmopolitan
Dysdera crocata
C.L. Koch, 1838 and subsequently elevatedto species status (Wunderlich 1991; Arnedo
et al
. 2000),which has been confirmed by DNA sequence data (Arnedo
et al
. 2001). The distribution of
D. lancerotensis
overlapswith five additional endemic species of
Dysdera
. However,the other endemics seem to have narrower ecologicalrequirements and are mostly restricted to the highest andmore humid parts of the islands, while
D. lancerotensis
hasalso been collected in xerophilous, sea-level habitats andshould be considered a generalist (Arnedo
et al
. 2000).In the present study, we use mitochondrial DNA (mtDNA)
sequence data to examine genetic variability and investi-gate levels of genetic structure in the Eastern Canarianendemic
D. lancerotensis
, in order to correlate the influenceof volcanic activity and marine transgressions in shapingphylogeographical patterns in a generalist species. Basedon geological history, we anticipate the following scenarios:first, a sequential colonization of the eastern ridge, follow-ing the SSW-NNE trend of Post-Miocene volcanic activity,as proposed for the darkling beetle
H. deyrollei
(Juan
et al
.1998); second, complex phylogeographical and demographicpatterns in
D. lancerotensis
, reflecting the occurrence offrequent extinction, expansion, fragmentation and recolon-ization processes as a result of long-term periodic volcanism.
Materials and methods
Sampling
Dysdera lancerotensis
specimens were collected from theEastern Canary Islands during 2004 and 2005, and stored
in absolute ethanol at 4
°
C. Seventeen localities weresampled: nine in Lanzarote, three in Fuerteventura, threein La Graciosa, one in Montaña Clara and one in Alegranza,and six additional sequences of
D. lancerotensis
were obtainedfrom GenBank (Fig. 1, Table 1). Samples of close morpho-logical relatives (several haplotypes of the cosmopolitan
Dysdera crocata
C. L. Koch, 1838 and two Moroccan species,
Dysdera mauritanica
Simon, 1909 and
Dysdera
sp.), alongwith two specimens of
Dysdera inermis
Ferrández, 1984 thatprovided a calibration point for estimating absolute lineageages (see below), as well as other
Dysdera
species endemicto the Eastern Canaries were included in the analyses(Table 1).
DNA extraction, PCR amplification and sequencing
Genomic DNA was extracted from two legs using theREALPURE Genomic DNA extraction kit from REALfollowing manufacturer’s guidelines. A 649-bp region ofthe mitochondrial cytochrome oxidase I (
cox1
) gene wasamplified for all samples using polymerase chain reaction(PCR), utilizing modified primers specifically designed forthis study C1-J-1546 (5
′
-GCTATAGTGGGTACGGCTATAAG-3
′
) and C1-N-2194 (5
′
-CTTCTGGATGACCAAAAAATC-3
′
).For a subset of samples, a region of about 1-Kb spanningfragments of the mitochondrial large ribosomal subunit(
rrnL
), tRNA leucine (L1) and the NADH dehydrogenasesubunit 1 (
nad1
) genes were amplified with primers LR-N-13398: (5
′
-CGCCTGTTTATCAAAAACAT-3
′
(Simon
et al
.1994)) and N1-J-12261: (5
′
-TCRTAAGAAATTATTTGAGC-3
′
(Hedin 1997).Amplifications were carried out in 25
µ
L reaction vol-ume in a final concentration of 1.25 U
Taq
polymerase(Promega), 1
×
buffer (Promega), 2.5 m
m
MgCl
2
(Promega),0.2 m
m
of each dNTP, 0.2
µ
m
of each primer and 2
µ
L ofDNA sample. PCR conditions were as follows: initial dena-turing step at 95
°
C for 5 min, 35 amplification cycles (94
°
Cfor 30 s, 45
°
C for 35 s, 72
°
C for 45 s for
cox1
fragment and94
°
C for 40 s, 45
°
C for 1 min, 72
°
C for 1 min for
rrnL-nad1
fragment) and a final step at 72
°
C for 5 min. PCR productswere purified using MultiScreen 96-Well Filter Plates fromMillipore. PCR products were cycle-sequenced in bothdirections using the same PCR primers and BigDye Termin-ator version 3.1 Cycle Sequencing Kit (Applied Biosystems).DNA sequences were edited using programs in the
staden
package (http://staden.sourceforge.net/) andassembled and managed using
bioedit
(Hall 1999).
Phylogenetic analyses
We performed maximum parsimony (MP), maximumlikelihood (ML) and Bayesian analyses to asses the internalstructure and the root of the
Dysdera lancerotensis
haplotypetree. Two data sets were analysed. The first data set included
P H Y L O G E O G R A P H Y O F
D Y S D E R A L A N C E R O T E N S I S
3201
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
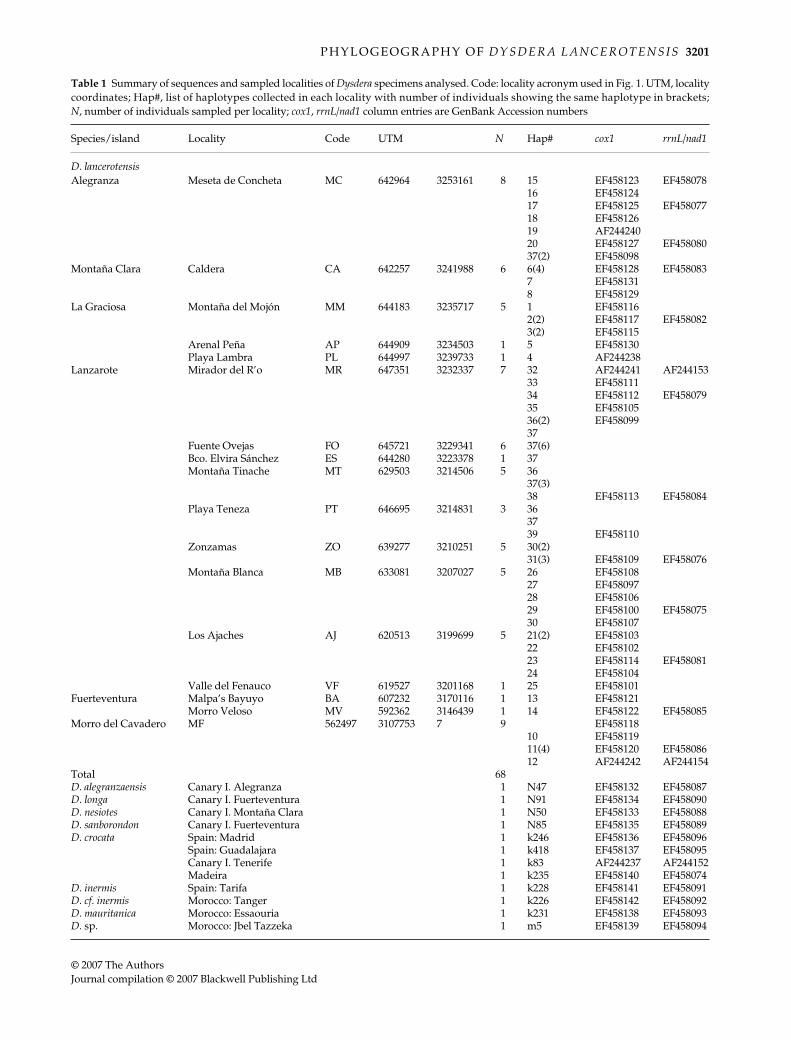
Table 1
Summary of sequences and sampled localities of
Dysdera
specimens analysed. Code: locality acronym used in Fig. 1. UTM, localitycoordinates; Hap#, list of haplotypes collected in each locality with number of individuals showing the same haplotype in brackets;
N
, number of individuals sampled per locality;
cox1
,
rrnL/nad1
column entries are GenBank Accession numbers
Species/island Locality Code UTM
N
Hap#
cox1 rrnL/nad1
D. lancerotensis
Alegranza Meseta de Concheta MC 642964 3253161 8 15 EF458123 EF45807816 EF45812417 EF458125 EF45807718 EF45812619 AF24424020 EF458127 EF45808037(2) EF458098
Montaña Clara Caldera CA 642257 3241988 6 6(4) EF458128 EF4580837 EF4581318 EF458129
La Graciosa Montaña del Mojón MM 644183 3235717 5 1 EF4581162(2) EF458117 EF4580823(2) EF458115
Arenal Peña AP 644909 3234503 1 5 EF458130Playa Lambra PL 644997 3239733 1 4 AF244238
Lanzarote Mirador del R’o MR 647351 3232337 7 32 AF244241 AF24415333 EF45811134 EF458112 EF45807935 EF45810536(2) EF45809937
Fuente Ovejas FO 645721 3229341 6 37(6)Bco. Elvira Sánchez ES 644280 3223378 1 37Montaña Tinache MT 629503 3214506 5 36
37(3)38 EF458113 EF458084
Playa Teneza PT 646695 3214831 3 363739 EF458110
Zonzamas ZO 639277 3210251 5 30(2)31(3) EF458109 EF458076
Montaña Blanca MB 633081 3207027 5 26 EF45810827 EF45809728 EF45810629 EF458100 EF45807530 EF458107
Los Ajaches AJ 620513 3199699 5 21(2) EF45810322 EF45810223 EF458114 EF45808124 EF458104
Valle del Fenauco VF 619527 3201168 1 25 EF458101Fuerteventura Malpa’s Bayuyo BA 607232 3170116 1 13 EF458121
Morro Veloso MV 592362 3146439 1 14 EF458122 EF458085Morro del Cavadero MF 562497 3107753 7 9 EF458118
10 EF45811911(4) EF458120 EF45808612 AF244242 AF244154
Total 68
D. alegranzaensis
Canary I. Alegranza 1 N47 EF458132 EF458087
D. longa
Canary I. Fuerteventura 1 N91 EF458134 EF458090
D. nesiotes
Canary I. Montaña Clara 1 N50 EF458133 EF458088
D. sanborondon
Canary I. Fuerteventura 1 N85 EF458135 EF458089
D. crocata
Spain: Madrid 1 k246 EF458136 EF458096Spain: Guadalajara 1 k418 EF458137 EF458095Canary I. Tenerife 1 k83 AF244237 AF244152Madeira 1 k235 EF458140 EF458074
D. inermis
Spain: Tarifa 1 k228 EF458141 EF458091
D. cf. inermis
Morocco: Tanger 1 k226 EF458142 EF458092
D. mauritanica
Morocco: Essaouria 1 k231 EF458138 EF458093
D.
sp. Morocco: Jbel Tazzeka 1 m5 EF458139 EF458094

3202
L . B I D E G A R A Y - B A T I S T A
E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
the haplotypes found in the 68 specimens analysed for the
cox1
gene fragment. A second data set included a sample of14 haplotypes (Table 1) for which the
cox1 fragment wascombined with additional sequences of the genes rrnL, L1and nad1 (three sequences retrieved from GenBank, Accessionnos AF244152–AF244154). The aim of the second data setwas to improve the phylogenetic signal, since some of thebasal relationships were ambiguous or poorly supportedby the cox1 fragment alone.
Sequence alignment of the cox1 was trivial since no inser-tions/deletions were observed. In contrast, sequences ofthe rrnL and L1 gene fragments showed length polymorphismand were aligned using the automatic multiple sequencealignment algorithm implemented in clustal x (Higginset al. 1992). The effect of different gap-opening and exten-sion values was investigated by building different multiplesequence alignments under the following parameter com-binations (GOP:GEP): 8:2, 8:4, 20:2, 24:4, 24:6 (transition/transversion ratio was kept constant at 0.5). Charactercongruence among gene partitions (cox1, nad1, rrnL-L1) asmeasured by the ILD index (Mickevich & Farris 1981) wasused to select one of the alignments for subsequent com-bined analyses. Gaps were scored as absence/presence datausing the simple method of Simmons & Ochoterena (2000)as implemented in gapcoder (Young & Healy 2002).Absence/presence scoring allows insertion/deletion in-formation to be incorporated into phylogenetic analyses,while minimizing the effect of increasing the weight ofoverlapping multiple nonhomologous gaps that result fromscoring gaps as a fifth character state. In addition, gapsscored as absence/presence are amenable to phylogeneticanalysis in a Bayesian framework.
The program modeltest version 3.6 (Posada & Crandall1998) was used to find the model of evolution that best fitthe data with lesser parameters, as suggested by the Akaikeinformation criterion (AIC; Posada & Buckley 2004), forsubsequent model-based phylogenetic analyses.
Parsimony analyses under equal weights were performedwith tnt version 1.0 (Goloboff et al. 2003) using a heuristicsearch based on 1000 replicates of random sequence addi-tion followed by tree-bisection–reconnection (TBR) branchswapping (five trees retained per iteration, and final roundof TBR branch swapping on all retained trees). Differentialweighting (e.g. down-weighting of third positions) wasnot implemented because of the lack of an objectivecriterion to assess phylogenetic reliability. It has also beenshown that the disadvantages of weighting schemesaccording to homoplasy levels may outweigh the benefits(Bjorklund 1999; Kaellersjö et al. 1999; Vogler et al. 2005).Standard nonparametric bootstrap technique provided ameasure of clade support based on 1000 pseudoreplicates(individual heuristic searches included 20 iterations of ran-dom sequence addition followed by TBR branch swappingand retaining five trees per replicate). Bayesian inference
analyses were conducted with mrbayes version 3.0(Ronquist & Huelsenbeck 2003). Four simultaneous MCMCruns (one cold, three heated) were performed for 2 millioniterations. Two independent runs were performed. Thestandard deviation of the split frequencies of the two runs(< 0.01) and the effective sample size (ESS) as measured bythe program tracer version 1.3 (Rambaut & Drummond2005) was used to ensure that the Markov chains hadreached stationarity. tracer was further used to deter-mine the correct number of generations to discard as aburn-in for the analysis. All analyses were run with specificand unlinked models selected by modeltest for each genefragment and a standard discrete model for gaps. Posteriorprobabilities of each clade were used as a support measure.Maximum-likelihood analyses were performed with phymlversion 2.4.4 (Guindon & Gascuel 2003), implementing themodel and parameter values obtained with modeltestand using the averaged Bayesian tree as starting point.Nonparametric bootstrap supports were calculated basedon 100 pseudoreplicates and using the best tree found inthe former analysis as starting point.
Estimations of substitution rate and genealogical and population divergence times
It is common practice in arthropod phylogeographicalstudies to rely on a supposedly universal 2.3% pairwisesequence divergence per million years of arthropod mtDNA(Brower 1994). However, it has been demonstrated thatsignificant rate of heterogeneity exists between even closelyrelated taxa. Therefore, we chose to estimate a lineage-specific substitution rate instead. Because of the absenceof a fossil record of the group, we use biogeographicalinformation based on the well-dated geochronology of theMediterranean. The vicariant events associated with theopening of the straight of Gibraltar about 5.3 Ma have beenused as calibration point to estimate lineage ages in otherorganisms, including both arthropods (Gomez-Zurita 2004)and vertebrates (Carranza & Arnold 2003). The speciesDysdera inermis Ferrández, 1984 is restricted to the south-ernmost part of the Iberian Peninsula, but closely relatedpopulations have been recently collected from the Rifregion in Morocco (Arnedo, unpublished data). Preliminarydata showed that mitochondrial genetic divergences betweenthe Moroccan and Iberian populations are in agreementwith a post-Messinian split of these populations. The cox1data set was reanalysed with mrbayes using the modelselected by modeltest (GTR + I + G) and constraining themain clades with high support obtained in the combinedanalyses of multiple data sets (see Results). The averagedtree was used to estimate clade ages and substitution ratesusing r8s (Sanderson 2003). The eastern clade representativeswere used to root the tree but were pruned beforeestimating time and rates. The split between the two trans-

P H Y L O G E O G R A P H Y O F D Y S D E R A L A N C E R O T E N S I S 3203
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Gibraltar sequences of D. inermis was fixed to 5.3 Ma. Across-validation analysis (Sanderson 2002) selected thepenalized likelihood with smoothing factor 316.23 as themost appropriate relaxed clock method for estimatingclade ages and rates. Confidence intervals were calculatedwith a sample of 100 trees obtained during the Bayesiansearches using the profile command in r8s.
We used a coalescent-based approach as implementedin mdiv (Nielsen & Wakeley 2001) to investigate themagnitude of the difference between time of populationdivergence and time of the most recent common ancestor(TMRCA) of the two groups of populations that corre-sponded to the two main haplotype clades (northern isletsvs. Lanzarote–Fuerteventura). mdiv provides a posteriordistribution of the demographic parameters theta (θ = 2Neu),migration rate (M = Nem), and time of population diver-
gence (T = t/Ne) using a likelihood framework, and alsoestimates the expected time to the most recent commonancestor (TMRCA = tu) (Nielsen & Wakeley 2001). mdivanalyses were run at Cornell’s CBSU computer cluster(http://cbsuapps.tc.cornell.edu/mdiv.aspx). We ran twoindependent simulations to ensure the convergence of theresults. Each simulation was run for 5 × 106 generationswith a 10% burn-in period under the ‘finite site model’ andusing a maximum prior of 10 for the scaled migration rate(M) and divergence time (T), as recommended by theauthor. The values with the highest posterior probabilitywere accepted as the best estimate. Values for T and TMRCAwere calculated using a generation time of 1.5 years (Cooke1965) and utilizing the lineage-specific substitution ratewith confidence intervals estimated in the present study(see above).
Fig. 2 The 50% majority-rule consensustree (left cladogram, right phylogram withoutgroup branches removed) of 3852 treessampled from the posterior distribution ofthe Bayesian analysis of cox1 after burn-in.Thick branches indicate clades supportedabove 95% Bayesian posterior probabi-lity. MP bootstrap support indicatedabove branches and ML bootstrap supportshown below (only values > 50% indicated).Asterisks denote contradicted groups. (MP:1818 trees of 720 steps, CI = 0.47, RI = 0.73;ML: 1 tree of –ln L = –3893.76821].

3204 L . B I D E G A R A Y - B A T I S T A E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Genetic diversity and population genetic structure
Genetic diversity indices, such as nucleotide and haplotypediversity and the average number of nucleotide differences,were calculated using arlequin version 3.01 (Excoffieret al. 2005). samova version 1.0 (Dupanloup et al. 2002) wasused to assess population structure. Given a priori numberof groups (k), samova implements a simulated annealingprocedure to define groups of geographically homogeneouspopulations that maximize the proportion of total geneticvariance due to differences between population groups.samova was run using 100 simulated annealing processesfor k values from 2 to 9. We used the Snn statistic (Hudson2000) implemented in dnasp 4.0 (Rozas et al. 2003) tocrosscheck the genetic differentiation between groupsselected by samova.
Demographic analyses
The demographic history of the studied populationswas investigated using mismatch distributions (Rogers &Harpending 1992) and statistics that test for departurefrom the neutral mutation model. Expansion parameterswere estimated using a nonlinear least-squares approach(Schneider & Excoffier 1999) as implemented in arlequinversion 3.01 under two different models: the suddenexpansion model (ι, θ0 and θ1; Harpending et al. 1998;Schneider & Excoffier 1999) and the spatial expansionmodel (τ, θ, M; Ray et al. 2003; Excoffier 2004). We estimatedthe approximate expansion time in generations and thenumber of females before and after the expansion fromthose parameters, assuming ι = 2ut is the expansion timein units of mutation time (where u is the mutation rate forthe whole sequence and t the number of generations sincethe expansion); θ0 = 2µN0 and θ1 = 2µN1 the mutationparameters before and after the expansion (where N0 andN1 are the number of females before and after the expansion;N0 = N1 under the spatial expansion model) and M = 2Nmthe migration parameter (m being the migrant exchangerate). The bootstrap approach (1000 replications) imple-mented in arlequin was used to obtain confidenceintervals for the estimated parameters and to test whetherthe observed data was consistent with the expansionmodels defined by the parameters (Schneider & Excoffier1999). The significance of the raggedness index (r) of theobserved distribution was also investigated with arlequinto further test the validity of the model.
As an alternative approach to the study of demographichistory, the Tajima’s D-test statistic (Tajima 1989) and Fu’sFS test (Fu 1997) were also computed using arlequin. Thesignificantly negative values for the FS and Tajima’s D-teststatistics are deviations from neutrality and are associatedwith signatures caused by population expansion and/ordirect selection (Tajima 1989; Fu 1997). A negative FS value
can be explained by an excess of recent mutations (Fu 1997)while a negative Tajima’s D-test value corresponds to anunequal frequency of polymorphic variants with an excessof the most common type and a deficiency of the lesscommon types (Hartl 2000).
Since demographic analyses are sensitive to populationsubdivision and haplotype frequencies, all analyses wereperformed in the homogeneous population groups definedby samova of considerable population size (at least 30individuals).
Network and nested clade analyses (NCA)
Haplotype networks were inferred using the statisticalparsimony analysis (Templeton et al. 1992) implemented intcs version 1.21 (Clement et al. 2000). Pairs of haplotypesare linked by the smallest number of differences definedby a 95% confidence limit that no homoplasic changes areincluded. Ambiguities in the haplotype network wereresolved following the guidelines proposed by Crandall& Templeton (1993). The final haplotype network wasmanually converted into a nested series of clades followingTempleton & Boerwinkle (1987) and Templeton & Sing(1993) to perform a nested clade phylogeographical analysis(NCPA) (Templeton et al. 1995; Templeton 1998, 2004).In cases where it was possible to use the tree topologyresulting from the combined analyses of the cox1 + rrnL +L1 + nad1 to link haplotype networks that could not beconnected at the 95% level of confidence using statisticalparsimony, we grouped individual networks as sister cladesat equal nesting levels (Templeton et al. 1987, e.g. Kozaket al. 2006). Additional criteria to establish polarity duringclade distance comparisons were obtained by lowering theconfidence limit of not including homoplasies provided.The NCPA was carried out using geodis version 2.4 (Posadaet al. 2000) and the distance pattern was interpreted usinga revised version of the inference key published byTempleton (2004) available on the geodis web page (http://darwin.uvigo.es/software/geodis.html).
Results
Sequences
We obtained 62 sequences of 605 bp of the cox1 mito-chondrial gene of Dysdera lancerotensis. Six additionalsequences were retrieved from GenBank, providing atotal of 68 cox1 sequences for subsequent analyses. Thesesequences corresponded to 39 haplotypes (GenBankAccession numbers in Table 1). All haplotypes wereexclusive to a single island except haplotype 37, which waspresent in both Alegranza and Lanzarote (Table 1). Amongthese 39 haplotypes, 109 nucleotide sites were variable (69.7%at the third codon position), 75 parsimony-informative

P H Y L O G E O G R A P H Y O F D Y S D E R A L A N C E R O T E N S I S 3205
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
and 34 singletons (nucleotide substitution present in onlyone sampled sequence). The A-T bias was 62%, and 68% ofnucleotide substitutions were transitions. Average uncor-rected pairwise genetic distance among all D. lancerotensishaplotypes was 4.05% (±1.8%) and the maximum sequencedivergence between any two haplotypes was 8.8%.
Phylogenetic structure
Results of the phylogentic analyses of the cox1 data matrixare summarized in Fig. 2. The analyses confirm that D.lancerotensis is not related to the other endemic speciesfrom the eastern islands but to Moroccan species. Allmethods strongly support monophyly of D. lancerotensishaplotypes. A single haplotype from Lanzarote (32) is thesister group to the remaining haplotypes sampled, althoughit only receives high support form the ML analyses. Theremaining haplotypes from Lanzarote are grouped in twoclades: L2, which receives high support in all analyses, andL1, with low support. Haplotypes from Fuerteventuraform a clade (F) that receives high support in all analyses.L2 and F are shown as sister groups but only receive highsupport in the ML analyses. Haplotypes sampled from the
northern islets form a clade that, again, only receives highsupport in the ML analyses. Inside this clade, haplotypesfrom Montaña Clara form a highly supported clade nestedwithin a clade that includes all haplotypes from La Graciosaand some from Alegranza (NI2). The remaining haplotypesfrom Alegranza are shown as basal to the former clade.Sequences of three additional mitochondrial gene fragments(nad1, rrnL and L1) from outgroups and representativesof the haplotype clades supported in the cox1 analyseswere included to improve resolution (GenBank Accessionnumbers in Table 1). Clustal alignment of the rrnL and L1fragments under gap opening cost 8 and gap extension cost 2,maximized congruence among genes (Table 2). Results ofthe phylogenetic analyses of the combined cox1 + L1 +rrnL + nad1 are summarized in Fig. 3. The combined analysesconfirm some of the clades already found in the cox1-onlyanalyses (NI2, L2, L1 and F), and provide strong supportfor some poorly resolved relationships. All methods agreein splitting haplotypes in two basal clades with highsupport, one that includes haplotypes from the northernislets, and another one that includes those from Lanzaroteand Fuerteventura. The northern islets clade includes,along with clade NI2, a clade exclusive to Alegranza (NI1).
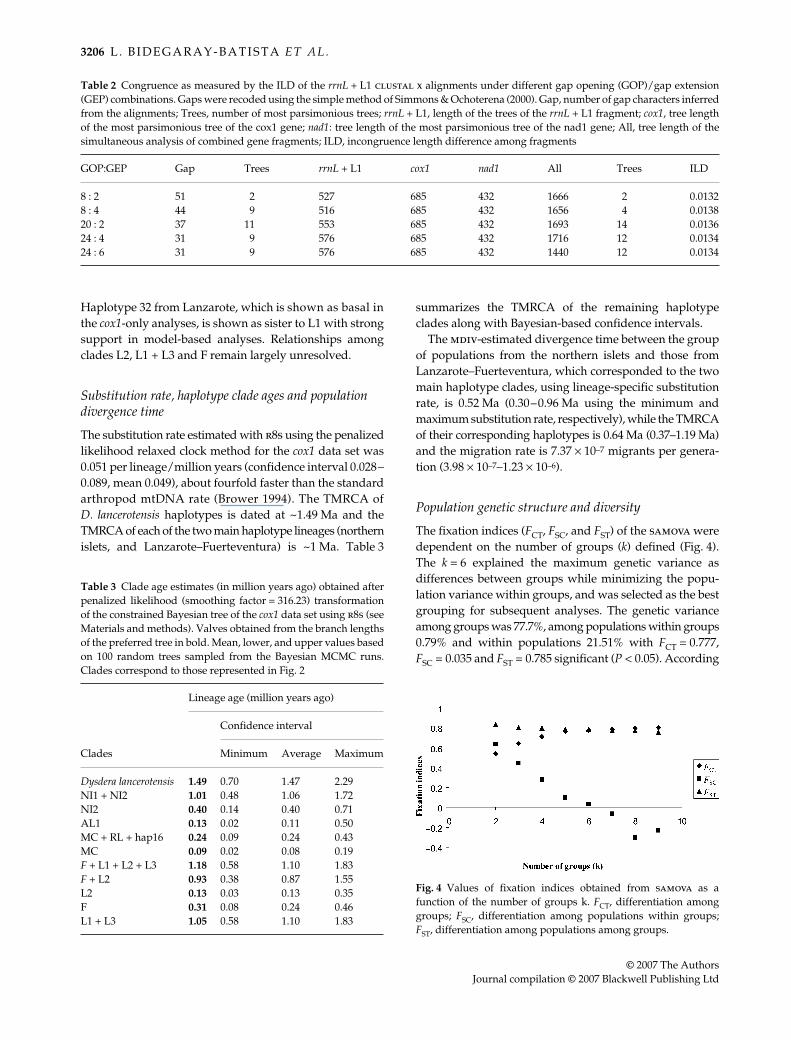
Fig. 3 The 50% majority-rule consensus phylogram of 6002 trees sampled from the posterior distribution of the Bayesian analysis of thecombined analyses of the cox1, L1, rrnL and nad1 partitions after burn-in. Thick branches indicate clades supported above 95% Bayesianposterior probability. Numbers near internal branches correspond to MP bootstrap support and ML bootstrap support, respectively (onlyvalues > 50% indicated). Arrows indicate those clades constrained in the Bayesian analyses of the cox1 only data set for lineage ageestimation (see Materials and methods). (MP: 2 trees of length 1666, CI = 0.57, RI = 0.68; ML: 1 tree of –ln L = –8934.28652).
3206 L . B I D E G A R A Y - B A T I S T A E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Haplotype 32 from Lanzarote, which is shown as basal inthe cox1-only analyses, is shown as sister to L1 with strongsupport in model-based analyses. Relationships amongclades L2, L1 + L3 and F remain largely unresolved.
Substitution rate, haplotype clade ages and population divergence time
The substitution rate estimated with r8s using the penalizedlikelihood relaxed clock method for the cox1 data set was0.051 per lineage/million years (confidence interval 0.028–0.089, mean 0.049), about fourfold faster than the standardarthropod mtDNA rate (Brower 1994). The TMRCA ofD. lancerotensis haplotypes is dated at ~1.49 Ma and theTMRCA of each of the two main haplotype lineages (northernislets, and Lanzarote–Fuerteventura) is ~1 Ma. Table 3
summarizes the TMRCA of the remaining haplotypeclades along with Bayesian-based confidence intervals.
The mdiv-estimated divergence time between the groupof populations from the northern islets and those fromLanzarote–Fuerteventura, which corresponded to the twomain haplotype clades, using lineage-specific substitutionrate, is 0.52 Ma (0.30–0.96 Ma using the minimum andmaximum substitution rate, respectively), while the TMRCAof their corresponding haplotypes is 0.64 Ma (0.37–1.19 Ma)and the migration rate is 7.37 × 10–7 migrants per genera-tion (3.98 × 10–7–1.23 × 10–6).
Population genetic structure and diversity
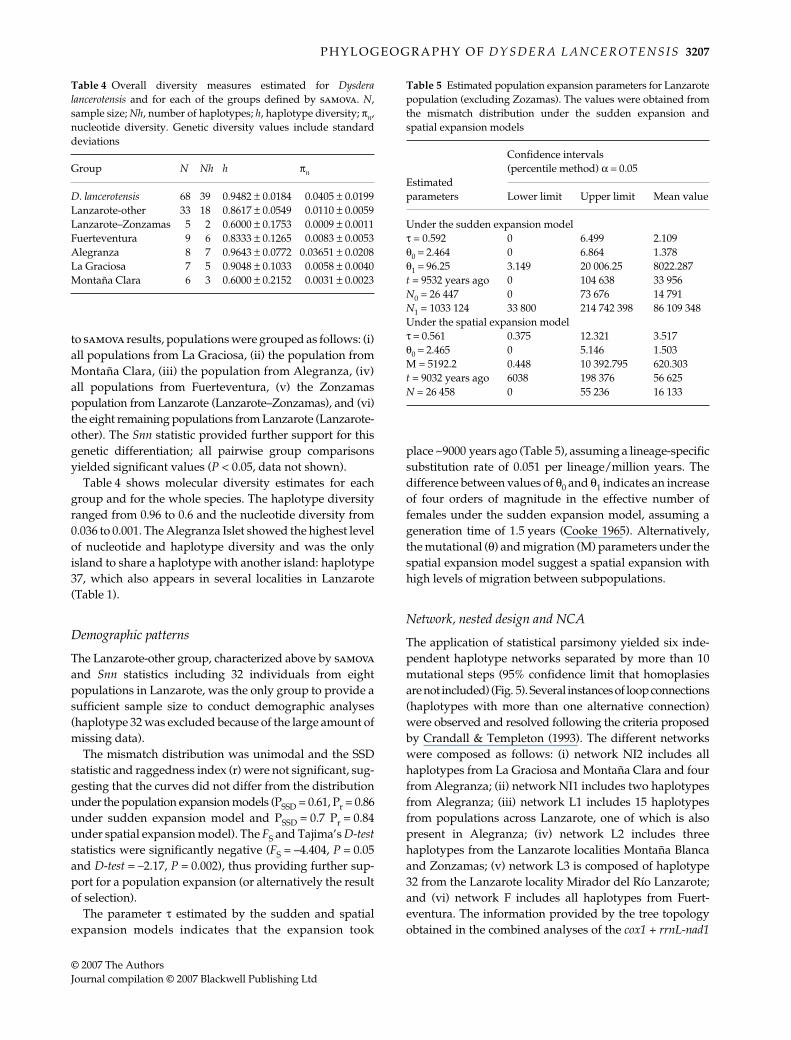
The fixation indices (FCT, FSC, and FST) of the samova weredependent on the number of groups (k) defined (Fig. 4).The k = 6 explained the maximum genetic variance asdifferences between groups while minimizing the popu-lation variance within groups, and was selected as the bestgrouping for subsequent analyses. The genetic varianceamong groups was 77.7%, among populations within groups0.79% and within populations 21.51% with FCT = 0.777,FSC = 0.035 and FST = 0.785 significant (P < 0.05). According
Table 3 Clade age estimates (in million years ago) obtained afterpenalized likelihood (smoothing factor = 316.23) transformationof the constrained Bayesian tree of the cox1 data set using r8s (seeMaterials and methods). Valves obtained from the branch lengthsof the preferred tree in bold. Mean, lower, and upper values basedon 100 random trees sampled from the Bayesian MCMC runs.Clades correspond to those represented in Fig. 2
Clades
Lineage age (million years ago)
Confidence interval
Minimum Average Maximum
Dysdera lancerotensis 1.49 0.70 1.47 2.29NI1 + NI2 1.01 0.48 1.06 1.72NI2 0.40 0.14 0.40 0.71AL1 0.13 0.02 0.11 0.50MC + RL + hap16 0.24 0.09 0.24 0.43MC 0.09 0.02 0.08 0.19F + L1 + L2 + L3 1.18 0.58 1.10 1.83F + L2 0.93 0.38 0.87 1.55L2 0.13 0.03 0.13 0.35F 0.31 0.08 0.24 0.46L1 + L3 1.05 0.58 1.10 1.83
Table 2 Congruence as measured by the ILD of the rrnL + L1 clustal x alignments under different gap opening (GOP)/gap extension(GEP) combinations. Gaps were recoded using the simple method of Simmons & Ochoterena (2000). Gap, number of gap characters inferredfrom the alignments; Trees, number of most parsimonious trees; rrnL + L1, length of the trees of the rrnL + L1 fragment; cox1, tree lengthof the most parsimonious tree of the cox1 gene; nad1: tree length of the most parsimonious tree of the nad1 gene; All, tree length of thesimultaneous analysis of combined gene fragments; ILD, incongruence length difference among fragments
GOP:GEP Gap Trees rrnL + L1 cox1 nad1 All Trees ILD
8 : 2 51 2 527 685 432 1666 2 0.01328 : 4 44 9 516 685 432 1656 4 0.013820 : 2 37 11 553 685 432 1693 14 0.013624 : 4 31 9 576 685 432 1716 12 0.013424 : 6 31 9 576 685 432 1440 12 0.0134
Fig. 4 Values of fixation indices obtained from samova as afunction of the number of groups k. FCT, differentiation amonggroups; FSC, differentiation among populations within groups;FST, differentiation among populations among groups.
P H Y L O G E O G R A P H Y O F D Y S D E R A L A N C E R O T E N S I S 3207
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
to samova results, populations were grouped as follows: (i)all populations from La Graciosa, (ii) the population fromMontaña Clara, (iii) the population from Alegranza, (iv)all populations from Fuerteventura, (v) the Zonzamaspopulation from Lanzarote (Lanzarote–Zonzamas), and (vi)the eight remaining populations from Lanzarote (Lanzarote-other). The Snn statistic provided further support for thisgenetic differentiation; all pairwise group comparisonsyielded significant values (P < 0.05, data not shown).
Table 4 shows molecular diversity estimates for eachgroup and for the whole species. The haplotype diversityranged from 0.96 to 0.6 and the nucleotide diversity from0.036 to 0.001. The Alegranza Islet showed the highest levelof nucleotide and haplotype diversity and was the onlyisland to share a haplotype with another island: haplotype37, which also appears in several localities in Lanzarote(Table 1).
Demographic patterns
The Lanzarote-other group, characterized above by samovaand Snn statistics including 32 individuals from eightpopulations in Lanzarote, was the only group to provide asufficient sample size to conduct demographic analyses(haplotype 32 was excluded because of the large amount ofmissing data).
The mismatch distribution was unimodal and the SSDstatistic and raggedness index (r) were not significant, sug-gesting that the curves did not differ from the distributionunder the population expansion models (PSSD = 0.61, Pr = 0.86under sudden expansion model and PSSD = 0.7 Pr = 0.84under spatial expansion model). The FS and Tajima’s D-teststatistics were significantly negative (FS = –4.404, P = 0.05and D-test = –2.17, P = 0.002), thus providing further sup-port for a population expansion (or alternatively the resultof selection).
The parameter τ estimated by the sudden and spatialexpansion models indicates that the expansion took
place ~9000 years ago (Table 5), assuming a lineage-specificsubstitution rate of 0.051 per lineage/million years. Thedifference between values of θ0 and θ1 indicates an increaseof four orders of magnitude in the effective number offemales under the sudden expansion model, assuming ageneration time of 1.5 years (Cooke 1965). Alternatively,the mutational (θ) and migration (M) parameters under thespatial expansion model suggest a spatial expansion withhigh levels of migration between subpopulations.
Network, nested design and NCA
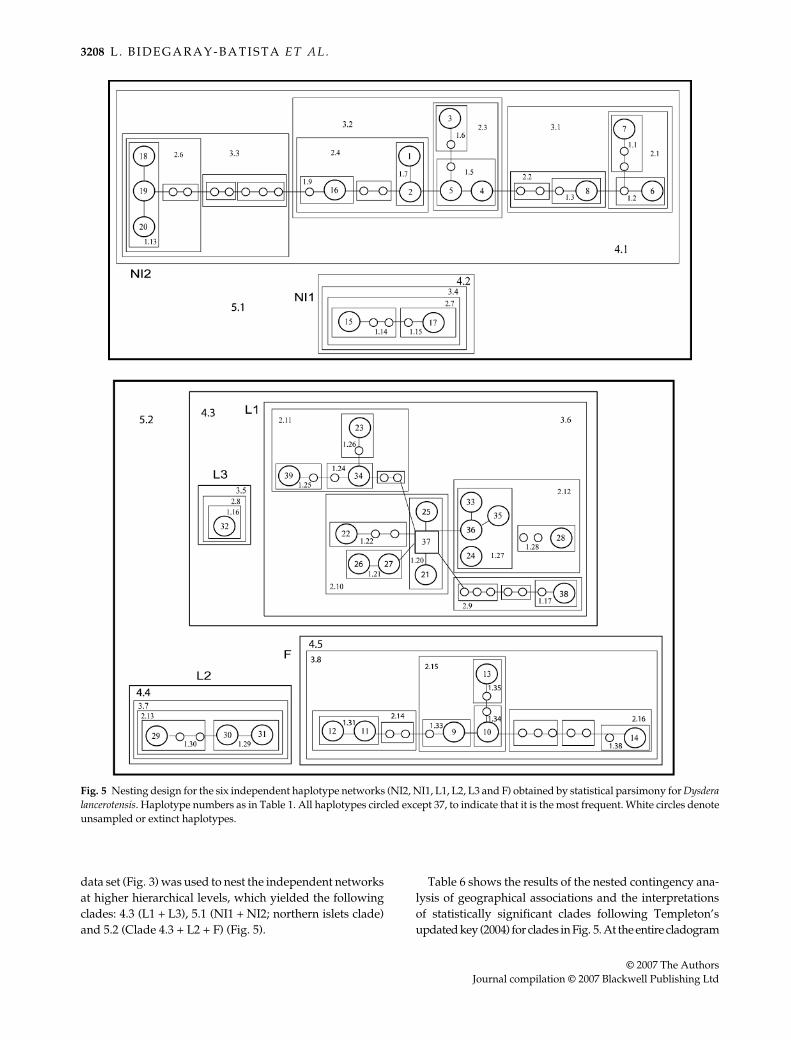
The application of statistical parsimony yielded six inde-pendent haplotype networks separated by more than 10mutational steps (95% confidence limit that homoplasiesare not included) (Fig. 5). Several instances of loop connections(haplotypes with more than one alternative connection)were observed and resolved following the criteria proposedby Crandall & Templeton (1993). The different networkswere composed as follows: (i) network NI2 includes allhaplotypes from La Graciosa and Montaña Clara and fourfrom Alegranza; (ii) network NI1 includes two haplotypesfrom Alegranza; (iii) network L1 includes 15 haplotypesfrom populations across Lanzarote, one of which is alsopresent in Alegranza; (iv) network L2 includes threehaplotypes from the Lanzarote localities Montaña Blancaand Zonzamas; (v) network L3 is composed of haplotype32 from the Lanzarote locality Mirador del Río Lanzarote;and (vi) network F includes all haplotypes from Fuert-eventura. The information provided by the tree topologyobtained in the combined analyses of the cox1 + rrnL-nad1
Table 4 Overall diversity measures estimated for Dysderalancerotensis and for each of the groups defined by samova. N,sample size; Nh, number of haplotypes; h, haplotype diversity; πn,nucleotide diversity. Genetic diversity values include standarddeviations
Group N Nh h πn
D. lancerotensis 68 39 0.9482 ± 0.0184 0.0405 ± 0.0199Lanzarote-other 33 18 0.8617 ± 0.0549 0.0110 ± 0.0059Lanzarote–Zonzamas 5 2 0.6000 ± 0.1753 0.0009 ± 0.0011Fuerteventura 9 6 0.8333 ± 0.1265 0.0083 ± 0.0053Alegranza 8 7 0.9643 ± 0.0772 0.03651 ± 0.0208La Graciosa 7 5 0.9048 ± 0.1033 0.0058 ± 0.0040Montaña Clara 6 3 0.6000 ± 0.2152 0.0031 ± 0.0023
Table 5 Estimated population expansion parameters for Lanzarotepopulation (excluding Zozamas). The values were obtained fromthe mismatch distribution under the sudden expansion andspatial expansion models
Estimated parameters
Confidence intervals (percentile method) α = 0.05
Lower limit Upper limit Mean value
Under the sudden expansion modelτ = 0.592 0 6.499 2.109θ0 = 2.464 0 6.864 1.378θ1 = 96.25 3.149 20 006.25 8022.287t = 9532 years ago 0 104 638 33 956N0 = 26 447 0 73 676 14 791N1 = 1033 124 33 800 214 742 398 86 109 348Under the spatial expansion modelτ = 0.561 0.375 12.321 3.517θ0 = 2.465 0 5.146 1.503M = 5192.2 0.448 10 392.795 620.303t = 9032 years ago 6038 198 376 56 625N = 26 458 0 55 236 16 133
3208 L . B I D E G A R A Y - B A T I S T A E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
data set (Fig. 3) was used to nest the independent networksat higher hierarchical levels, which yielded the followingclades: 4.3 (L1 + L3), 5.1 (NI1 + NI2; northern islets clade)and 5.2 (Clade 4.3 + L2 + F) (Fig. 5).
Table 6 shows the results of the nested contingency ana-lysis of geographical associations and the interpretationsof statistically significant clades following Templeton’supdated key (2004) for clades in Fig. 5. At the entire cladogram
Fig. 5 Nesting design for the six independent haplotype networks (NI2, NI1, L1, L2, L3 and F) obtained by statistical parsimony for Dysderalancerotensis. Haplotype numbers as in Table 1. All haplotypes circled except 37, to indicate that it is the most frequent. White circles denoteunsampled or extinct haplotypes.

P H Y L O G E O G R A P H Y O F D Y S D E R A L A N C E R O T E N S I S 3209
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
level, the separation between clades 5.1 (northern isletsclade) and 5.2 (Lanzarote–Fuerteventura clade) is explainedby allopatric fragmentation. However, the structurewithin these clades reflects different historical processes.Clade 5.2 is consistent with a contiguous range expansion,whereas the structure within clade 4.1, which includeshaplotypes from the northern islets, reflects a past gradualrange expansion followed by fragmentation.
Discussion
A tale of two lineages
Phylogenetic analyses of the combined mitochondrialmarkers suggest a basal split of Dysdera lancerotensis haplo-types into two lineages, one including haplotypes from thenorthern islets and the other including haplotypes fromLanzarote and Fuerteventura. The TMRCA estimated forthe two main haplotype lineages of D. lancerotensis usinga phylogenetic approach was 1.49 Ma (0.69–2.28 Ma) andthe TMRCA of each lineage was set around 1 Ma. Thesevalues are at odds with current geological evidence, whichsuggests that present-day subaerial volcanic terrains inthe northern islets resulted from volcanic activity startingat about 45 000 years ago (Nuez Pestana & Luisa AlonsoBlanco 1998), although our lineage-specific substitutionrate is fourfold faster than standard arthropod mtDNAestimates. A possible explanation of this puzzle is that thesplit of the main haplotype clades greatly preceded thedivergence between populations. However, the divergencetime between populations from the northern islets andpopulations from Lanzarote and Fuerteventura and theTMRCA of their haplotypes as estimated with mdiv areactually quite similar, 0.52 Ma (0.30–0.96 Ma) and 0.64 Ma(0.37–1.19 Ma), respectively. In addition, mdiv shows avery low level of migration between populations suggestingthat they diverged by isolation, which is further corroboratedby the NCA interpretation of the two main haplotypenetworks as the result of an allopatric fragmentation event(Fig. 5, Table 6). Broad difference between the TMRCAestimated by r8s and that of mdiv is most likely the resultof the deep intraspecific divergences of the cox1 (maximum~8.5%) in D. lancerotensis. r8s estimates were based on
branch lengths obtained using a GTR model with invariantand rate heterogeneity. Conversely, mdiv only implementsa finite site model based on a simpler HKY model withhomogenous rates, which most probably results in anunderestimation of divergences among haplotypes. Wetested this hypothesis by estimating TMRCA using analternative Bayesian approach implemented in the programbeast version 1.3. (Drummond & Rambaut 2003), whichincorporates an array of clock methods and evolutionarymodels. The TMRCA of the D. lancerotensis haplotypesusing the exponential clock method (Drummond et al. 2006)and a GTR + I + Γ was 1.12 Ma (0.58–1.85 Ma), whileimplementing the strict clock and a HKY model withhomogenous rates yielded 0.81 Ma (0.64–0.99 Ma). Theseresults illustrate that the use of simpler models may resultsin more recent estimates of the TMRCA. Therefore, ourpopulation divergence estimates hints at a much older ageof the northern islets, which let us to propose that they mayhave arisen during the volcanic series that formed theFamara edifice. Biogeographical events older than thosesuggested by geological data have also been proposed toexplain phylogeographical patterns in Brachyderes weevilsin the island of La Palma (Emerson et al. 2006).
The actual processes that may in fact have generatedsuch divergence remains unclear. The core of the northernpart of Lanzarote is the Famara volcanic edifice, which wasthe last part of the eastern ridge to be formed (around10 Ma) and has also experienced the longest and mostcontinuous cycles of volcanic activity (Coello et al. 1992).To the three initial pulses and subsequent erosive periodsof the series I subaerial volcanic activity were followed bya relatively long period of quiescence from 3.8 to 1.8 millionyears, when post-erosional volcanism started. Plio-Pleistoceneactivity in Lanzarote has been more or less continuous,reaching a peak in the Lower Pleistocene (Ancochea et al.2004). The estimated TMRCA of the two main D. lancerotensislineages fits well with this final pulse of volcanic activityand suggests that severe bottlenecks during periods ofvolcanic activity may have played a key role in generatingdeep genetic divergences in D. lancerotensis.
The Canaries do not have any significant subsidence,which explains why islands older than 20 million years arestill emergent, while islands in the Hawaiian chain become
Table 6 Significant clades obtained from the nested contingency analyses of geographical associations and interpretation based on theinference key (see test for details). *, significant at the 0.05 level
CladeChi-squared statistic Probability Inference chain Inference
1.20 1.00 0.0101* 1-19-20 Inadequate geographical sampling4.1 28.16 0.0000* 1-2-11-12-13-14 No Past gradual range expansion followed by fragmentation5.2 85.97 0.0000* 1-2-11-12- No Contiguous range expansionTotal cladogram 59.61 0.0000* 1-2-3-4-9 No Allopatric fragmentation

3210 L . B I D E G A R A Y - B A T I S T A E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
submerged by subsidence after about 7 million years(Carracedo 1999). Mass-wasting processes and marinetransgressions have shaped the present-day configurationof the geologically continuous eastern ridge into independ-ent islands and islets. However, most of the islands werecyclically connected during Pleistocene glacial maximawhen sea level dropped, which seems to rule out marinetransgression as the main factor in generating divergencebetween the main lineages. This suggestion is corroboratedby the internal phylogeographical pattern of the islets (seebelow) and the presence of at least one haplotype wide-spread across Lanzarote and in Alegranza, indicative ofpossible recent gene flow between the two areas.
The role of ecological factors in the separation of the twolineages also seems very unlikely. There are no obviousenvironmental differences between the collecting localitieson the islets and on the main islands, neither are there clearmorphological differences, although rigorous morphometricanalyses have not been performed. Moreover, high levelsof tolerance to a wide range of environmental conditionshave been reported in the closely related species Dysderacrocata (Cooke 1968), which seem to be confirmed in D.lancerotensis by its wider distribution range when comparedto other Dysdera endemic to the islands (Arnedo et al. 2000).
Simulation models based on coalescence predictionshave shown that deep phylogeographical breaks can formwithin a continuously distributed species even when thereare no barriers to gene flow (Irwin 2002). However, thesemodels require low levels of individual dispersal distanceand small population sizes, which do not seem consistentwith the widespread presence of D. lancerotensis through-out the eastern ridge.
The northern islets
The three main northern islets are separated by shallowwaters (200 m below sea level at its deepest points), whichhints at the existence of land bridges between the islandsduring Eustatic sea level changes. However, all but one ofthe haplotypes found in the northern islets are exclusive tothe area, and each of the three islets harbour geneticallydifferentiated populations, which suggests that suchconnections have not been as common as predicted by theperiodical drop of sea level during Pleistocene glaciations.The pattern of genealogical relationships of the haplotypes(Figs 2 and 3), showing successive paraphyly of Alegranzaexclusive haplotypes relative to La Graciosa and MontañaClara, and paraphyly of La Graciosa haplotypes relative toMontaña Clara, supports Alegranza as the source of thecurrent population on La Graciosa, and that Montaña Clarawas colonized from La Graciosa. The successive decreasein genetic diversity from Alegranza to Montaña Clara(Table 4) provides further support for this pattern ofcolonization. The NCA suggests that the phylogeographical
pattern of D. lancerotensis in the northern islets is the resultof a past gradual range expansion followed by fragmentation(Fig. 5, Table 6). The phylogeographical pattern observedin the northern islets could be explained by the occurrenceof major population extinctions and subsequent recoloni-zations. The present-day subaerial terrains of the isletsderive from the late Pleistocene–Holocene pulses of volcanicactivity. It is not difficult to imagine a scenario in whichmost of the biota of a small islet like Montaña Clara, ofabout 1 km2, could be completely wiped out by the effectof lava flows, and subsequently recolonized from the near-est islet, in this case La Graciosa. Our haplotype lineageage estimates are congruent with the proposed patternof colonization, although they suggest slightly older agesthan the available geological data.
The fact that Alegranza, the youngest and most remoteof the islets, has the highest levels of genetic diversity andhas been the source of colonization for the other isletsconstitutes something of a conundrum. However, thesepatterns are supported by further observations. Alegranzaharbours by far the highest number of terrestrial inverte-brate species on the islets, including several endemisms: 3gastropods, 2 spiders, 1 pseudoscorpion, 1 silverfish and 1beetle (also present in the Selvage Islands). The reasons forthe greater diversity found on Alegranza are unclear, sinceno additional habitats are apparent with respect to theother islets. Remoteness may have spared Alegranza fromdestructive geological events that affected Montaña Claraand La Graciosa. Giant landslides had a key role in shapingthe landscapes of the Canary Islands (Stillman 1999). Dur-ing the Quaternary, tsunamis caused by volcanic islandmass-slides had a significant impact on the world’s coast-lines (Whelan & Kelletat 2003). Evidence of such tsunamiscan possibly be seen in the limestone and conglomeraticlayers composed of rounded pebbles that are found in LaGraciosa, although this has been disputed (Acosta et al.2003).
A reversed colonization pattern
The pattern of relationships of D. lancerotensis haplotypessuggests a north-to-south colonization of the eastern ridge,which is consistent with the higher levels of genetic diversityfound in Lanzarote and Alegranza (Table 4). However, thispattern opposes the SSW-NNE trend of post-Miocenevolcanic activity in the ridge. It has been suggested that thistemporal migration of volcanism shaped the phylogeo-graphical patterns of the darkling beetle Hegeter deyrollei( Juan et al. 1998). The lack of congruence between thephylogeographical patterns of D. lancerotensis and H. deyrolleiis most likely to result from the colonization by these twoorganisms from two opposite extremes of the eastern ridge.Lanzarote was originally formed as two independentislands, the present-day massifs of Famara in the north and

P H Y L O G E O G R A P H Y O F D Y S D E R A L A N C E R O T E N S I S 3211
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Ajaches in the south (Coello et al. 1992), that were joinedinto a single mass during a new phase of volcanic activitystarting at the Late Pliocene and extending to the Holocene.D. lancerotensis may have been restricted to the northernmassif until a suitable land connection allowed them toprogress south.
According to NCA, the colonization of Fuerteventurafrom Lanzarote would have been the result of contiguousrange expansion, probably facilitated by land connectionsassociated to sea-level fluctuations. The TMRCA of thesampled haplotypes in Fuerteventura (0.31 Ma, 0.07–0.46 Ma)is less than half that of their split from their Lanzarote sisterlineage (0.93 Ma, 0.37–1.55 Ma). Population bottlenecksand/or reductions in effective population size in the pastare mechanisms that may account for this pattern. A reduc-tion in diversification rates and the lack of older lineages ofthe beetle Brachyderes rugatus in the Western Canaries havebeen explained by larger extinction events that would havereduced mtDNA diversity as a product of island coloniza-tion, high levels of volcanic activity and dramatic erosionalevents (Emerson et al. 2000). All three elements probablyconcurred during the contiguous range expansion ofD. lancerotensis towards Fuerteventura.
Population growth and expansion: the fingerprint of volcanism on demographic patterns?
Mismatch distribution and neutrality test analyses hint ata very recent population expansion in Lanzarote thatwould have occurred ~9000 years ago (Table 5). Based oncoalescent theory, haplotype 37 (with the highest frequencyin the Famara edifice) would be the ancestral haplotypeof the L1 network, because of its interior location in thenetwork, high frequency and widespread occurrence.Estimations for the TMRCA of the main haplotype networksin Lanzarote suggest that they became differentiated aftera maximum of volcanic activity in the Lower Pleistocene(Coello et al. 1992). The most recent volcanic activity is stillevident today in the large areas of barren lava known asMalpaís (badlands) found in the central-west and northeastof the island, corresponding to the two more recenteruptive events: the Montaña Corona, dated 21 000 yearsago (Carracedo et al. 2003), and the historical Timanfayaeruptions in 1730 and 1824 (Carracedo et al. 1992). Reconci-liation of the demographic and phylogeographical patternsexhibited by D. lancerotensis could suggest the followingscenario. During short erosive stages, D. lancerotensis wouldhave spread and diversified throughout the island, whereasvolcanic activity stages would have caused extinction ofmost populations and the one that survived sufferedsevere bottlenecks. Towards the Upper Pleistocene and theHolocene, the frequency of eruptive cycles was reduced,becoming residual. At that point, populations from Famarawould have recolonized most of the island through a range
expansion as suggested by the mismatch distribution andneutrality tests. Some isolated populations that survivedvolcanic events in other parts of the island may have beenincorporated into this late expansion, as suggested by thepresence of the divergent haplotype 32, while anothersurviving population may have remained isolated, asrepresented by Zonzamas.
Conclusion
The present study reveals the complex phylogeographicaland demographic patterns of Dysdera lancerotensis on theEastern Canary Ridge as expected from the history ofvolcanic activity in the region. The occurrence of longbranches separating groups of haplotypes with shortcoalescence times, and the discovery that most of thecurrent populations in Lanzarote could be the result of arecent population expansion, suggest that the history of D.lancerotensis has been shaped by successive episodes ofextinction, recolonization and expansion. Contrary to thecolonization patterns of the ridge that had been reportedfor the darkling beetle Hegeter deyrolley, which closelyfollow the SSW-NNE migration of subaerial volcanicactivity, D. lancerotensis has colonized the Eastern Canariesfrom north to south, reaching its maximum levels of geneticdiversity in Alegranza, the northernmost part of the ridge.Both species seem to share certain ecological requirements,since they occur in all areas except for recent barren lavasand sand dunes; the opposing colonization pathways ofthe two species should therefore be considered the result ofhistorical contingency due to the arrival of ancestors at theextreme tips of the ridge. The presence of ancient haplotypesin Alegranza and haplotypes in Zonzamas not belongingto the main expansion in Lanzarote suggest that these areasacted as possible refuges during periods of generalizedvolcanic activity in the rest of the islands.
In summary, our study identifies the key role of anextremely dynamic geology in shaping historical and demo-graphic patterns in a spider species. Future research intoother terrestrial organisms in the Eastern Canaries isnecessary to gain a better understanding of the historicaland ecological factors that shape phylogeographical patternsand drive population-level processes.
Acknowledgements
We are grateful to Julio Rozas, Jesús Gómez-Zurita and two anon-ymous reviewers for providing perceptive comments that greatlycontributed to improving the manuscript. We also wish toacknowledge further contributions by Hermans Contreras-Díazand Xavier Turón. Mercè Loukota helped us with the laboratorywork, and Heriberto López, Salvador de la Cruz and Antonio J.Pérez assisted in the fieldwork. The Cabildos from Fuerteventuraand Lanzarote authorized the collecting permits for the protectednatural areas, and the Cabildo of Lanzarote kindly provided boat

3212 L . B I D E G A R A Y - B A T I S T A E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
transport and accommodation to visit the islets. M.A. wassupported by the Program Ramón y Cajal funded by the SpanishMinistry of Education and Science and the EU FEDER program,and N.M. was supported by a graduate grant from the AutonomousGovernment of the Canary Islands. This project was funded by theSpanish Ministry of Education and Science grant BOS2003-05876(to M.A. and P.O.) and additional financial support was providedby project 2005SGR00045 from the Autonomous Government ofCatalonia.
References
Acosta J, Uchupi E, Muñoz A et al. (2003) Geologic evolution of theCanarian Islands of Lanzarote, Fuerteventura, Gran Canaria andLa Gomera and comparison of landslides at these islands withthose at Tenerife, La Palma and El Hierro. Marine GeophysicalResearches, 24, 1–40.
Ancochea E, Barrera JL, Bellido F et al. (2004) Canarias y el vulcan-ismo neógeno Peninsular. In: Geología de España (ed. Vera JA),pp. 635–682. Sociedad Geológica de España-Instituto Geológicoy Minero de España, Madrid.
Arnedo MA, Oromí P, Ribera C (2000) Systematics of the genusDysdera (Araneae, Dysderidae) in the Eastern Canaries. Journalof Arachnology, 28, 261–292.
Arnedo MA, Oromí P, Ribera C (2001) Radiation of the spidergenus Dysdera (Araneae, Dysderidae) in the Canary Islands:cladistic assessment based on multiple data sets. Cladistics, 17,313–353.
Arnedo MA, Ribera C (1997) Radiation of the genus. Dysdera(Araneae, Haplogynae, Dysderidae) in the Canary Islands: theisland of Gran Canaria. Zoologica Scripta, 26, 205–243.
Arnedo MA, Ribera C (1999) Radiation of the genus Dysdera(Araneae, Dysderidae) in the Canary Islands: the island ofTenerife. Journal of Arachnology, 27, 604–662.
Avise JC (1998) The history and purview of phylogeography: apersonal reflection. Molecular Ecology, 7, 371–379.
Beheregaray LB, Ciofi C, Geist D et al. (2003) Genes record aprehistoric volcano eruption in the Galapagos. Science, 302, 75.
Bjorklund M (1999) Are third positions really that bad? A testusing vertebrate cytochrome b. Cladistics, 15, 191–197.
Bloor P, Brown RP (2005) Morphological variation in Gallotia atlanticafrom the volcanic island of Lanzarote: subspecies designationsand recent lava flows. Biological Journa — Linnean Society, 85,395–406.
Brower AVZ (1994) Rapid morphological radiation and con-vergence among races of the butterfly Heliconius erato inferredfrom patterns of mitochondrial DNA evolution. Proceedings ofthe National Academy of Sciences, USA, 91, 6491–6495.
Carracedo JC (1999) Growth, structure, instability and collapse ofCanarian volcanoes and comparisons with Hawaiian volcanoes.Journal of Volcanology and Geothermal Research, 94, 1–19.
Carracedo JC, Badiola ER, Soler V (1992) The 1730–1736 eruptionof Lanzarote, Canary Islands: a long, high magnitude basalticfissure eruption. Journal of Volcanology and Geothermal Research,53, 239–250.
Carracedo JC, Day S, Guillou H et al. (1998) Hotspot volcanismclose to a passive continental margin: the Canary Islands.Geological Magazine, 135, 591–604.
Carracedo JC, Singer B, Jicha B et al. (2003) La erupción y el tubovolcánico del Volcán Corona (Lanzarote, Islas Canarias). EstudiosGeológicos, 59, 277–302.
Carranza S, Arnold EN (2003) History of West Mediterraneannewts, Pleurodeles (Amphibia: Salamandridae), inferred fromold and recent DNA sequences. Systematics and Biodiversity, 1,327–338.
Clement M, Posada D, Crandall KA (2000) tcs: a computer programto estimate gene genealogies. Molecular Ecology, 9, 1657–1660.
Coello J, Cantagrel JM, Hernán F et al. (1992) Evolution of the east-ern volcanic ridge of the Canary Islands based on new K-Ardata. Journal of Volcanology and Geothermal Research, 53, 251–274.
Contreras-Diaz HG, Moya O, Oromí P, Juan C (2003) Phylogeog-raphy of the endangered darkling beetle species of Pimeliaendemic to Gran Canaria (Canary Islands). Molecular Ecology,12, 2131–2143.
Cooke JAL (1965) A contribution to the biology of the Britishspiders belonging to the genus Dysdera. Oikos, 16, 20–25.
Cooke JAL (1968) Factors affecting the distribution of somespiders of the genus Dysdera (Araneae, Dysderidae). Entomologist’sMonthly Magazine, 103, 221–225.
Crandall KA, Templeton AR (1993) Empirical tests of some predic-tions from coalescent theory with applications to intraspecificphylogeny reconstruction. Genetics, 134, 959.
Drummond AJ, Ho SYW, Phillips MJ, Rambaut A (2006) Relaxedphylogenetics and dating with confidence. Public Library ofScience, Biology, 4, 699–710.
Drummond AJ, Rambaut A (2003) BEAST. Available from http://evolve.zoo.ox.ac.uk/beast/.
Dupanloup I, Schneider S, Excoffier L (2002) A simulated anneal-ing approach to define the genetic structure of populations.Molecular Ecology, 11, 2571–2581.
Emerson BC, Forgie S, Goodacre S, Oromí P (2006) Testing phylo-geographic predictions on an active volcanic island: Brachyderesrugatus (Coleoptera: Curculionidae) on La Palma (CanaryIslands). Molecular Ecology, 15, 449–458.
Emerson BC, Oromí P, Hewitt GM (2000) Colonization and diver-sification of the species Brachyderes rugatus (Coleoptera) on theCanary Islands: evidence from mitochondrial DNA COII genesequences. Evolution, 54, 911–923.
Excoffier L (2004) Patterns of DNA sequence diversity and geneticstructure after a range expansion: lessons from the infinite-islandmodel. Molecular Ecology, 13, 853–864.
Excoffier L, Laval G, Schneider S (2005) ARLEQUIN, Version 3.0: anIntegrated Software Package for Population Genetics Data Analysis.Evolutionary Bioinformatics Online, 1, 47–50.
Fu YX (1997) Statistical tests of neutrality of mutations againstpopulation growth, hitchhiking and background selection.Genetics, 147, 915–925.
Goloboff PA, Farris JS, Nixon KC (2003) TNT: tree analysis using newtechnologies. http://www.zmuc.dk/public/phylogeny/INT/.
Gomez-Zurita J (2004) Molecular systematics and time-scale forthe evolution of Timarcha, a leaf-beetle genus with a disjunctHolarctic distribution. Molecular Phylogenetics and Evolution, 32,647–665.
Grant PR (1998) Evolution on islands. In: Evolution on Islands (ed.Grant PR), pp. xiv+334. Oxford University Press, Oxford, UK/New York.
Guindon S, Gascuel O (2003) A simple, fast, and accurate algo-rithm to estimate large phylogenies by maximum likelihood.Systematic Biology, 52, 696–704.
Hall TA (1999) BIOEDIT: a User-Friendly Biological Sequence Alignmenteditor and Analysis Program for Windows 95/98/NT, 95–98.
Harpending HC, Batzer MA, Gurven M et al. (1998) Genetic tracesof ancient demography. PNAS, 95, 1961–1967.

P H Y L O G E O G R A P H Y O F D Y S D E R A L A N C E R O T E N S I S 3213
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Hartl DL (2000) A Primer of Population Genetics, 3rd edn. SinauerAssociates, Sunderland, Massachusetts.
Hedin MC (1997) Molecular phylogenetics at the population/species interface in cave spiders of the Southern Appalachians(Araneae: Nesticidae: Nesticus). Molecular Biology and Evolution,14, 309–324.
Higgins DG, Bleasby AJ, Fuchs R (1992) clustal v: improvedsoftware for multiple sequence alignment. Computational andApplied Bioscience, 8, 189–191.
Hudson RR (2000) A new statistic for detecting genetic differenti-ation. Genetics, 155, 2011–2014.
Irwin DE (2002) Phylogeographic breaks without geographicbarriers to gene flow. Evolution, 56, 2383–2394.
Juan C, Emerson BC, Oromí P, Hewitt GM (2000) Colonizationand diversification: towards a phylogeographic synthesis forthe Canary Islands. Trends in Ecology and Evolution, 15, 104–109.
Juan C, Ibrahim KM, Oromí P, Hewitt GM (1998) The phylo-geography of the darkling beetle, Hegeter politus, in the easternCanary Islands. Proceedings of the Royal Society of London. Series B,Biological Sciences, 265, 135–140.
Kaellersjö M, Albert VA, Farris JS (1999) Homoplasy increasesphylogenetic structure. Cladistics, 15, 91–93.
Kozak KH, Blaine RA, Larson A (2006) Gene lineages and easternNorth American palaeodrainage basins: phylogeography andspeciation in salamanders of the Eurycea bislineata speciescomplex. Molecular Ecology, 15, 191–207.
Mayr E (1967) The challenge of island faunas. Australian NaturalHistory, 15, 359–374.
Mickevich MF, Farris JS (1981) The implications of incongruencein Menidia. Systematic Zoology, 30, 351–370.
Moya O, Contreras-Diaz HG, Oromí P, Juan C (2004) Geneticstructure, phylogeography and demography of two ground-beetle species endemic to the Tenerife laurel forest (CanaryIslands). Molecular Ecology, 13, 3153–3167.
Moya O, Contreras-Diaz HG, Oromí P, Juan C (2006) Using statis-tical phylogeography to infer population history: case studieson Pimelia darkling beetles from the Canary Islands. Journal ofArid Environments, 66, 477–497.
Nielsen R, Wakeley J (2001) Distinguishing migration fromisolation: a Markov Chain Monte Carlo Approach. Genetics, 158,885–896.
Nuez Pestana Jdl Quesada Alvarez M, Luisa Alonso Blanco JJ(1998) Los Volcanes de Los Islotes Al Norte de Lanzarote, Islas Canarias.Fundación César Manrique, Teguise, Lanzarote.
Posada D, Buckley TR (2004) Model selection and model averagingin phylogenetics: advantages of Akaike information criterionand Bayesian approaches over likelihood ratio tests. SystematicBiology, 53, 793–808.
Posada D, Crandall KA (1998) modeltest: testing the model ofDNA substitution. Bioinformatics, 14, 817–818.
Posada D, Crandall KA, Templeton AR (2000) geodis: a programfor the cladistic nested analysis of the geographical distributionof genetic haplotypes. Molecular Ecology, 9, 487–488.
Rambaut A, Drummond A (2005) tracer, a program for ana-lysing results from Bayesian MCMC programs such as beastand mrbayes. Available online http://evolve.zoo.ox.ac.uk/software.html?id=tracer (last accessed 12 February 2006).
Ray N, Currat M, Excoffier L (2003) Intra-deme molecular diver-sity in spatially expanding populations. Molecular Biology andEvolution, 20, 76–86.
Rees DJ, Emerson BC, Oromí P, Hewitt GM (2001a) The diversifi-cation of the genus Nesotes (Coleoptera: Tenebrionidae) in the
Canary Islands: evidence from mtDNA. Molecular Phylogeneticsand Evolution, 21, 321–326.
Rees DJ, Emerson BC, Oromí P, Hewitt GM (2001b) MitochondrialDNA, ecology and morphology: interpreting the phylogeo-graphy of the Nesotes (Coleoptera: Tenebrionidae) of Gran Canaria(Canary Islands). Molecular Ecology, 10, 427–434.
Rogers AR, Harpending H (1992) Population growth makeswaves in the distribution of pairwise genetic differences. Mole-cular Biology and Evolution, 9, 552–569.
Ronquist F, Huelsenbeck JP (2003) mrbayes 3: Bayesian phylogeneticinference under mixed models. Bioinformatics, 19, 1572–1574.
Rozas J, Sánchez-De I, Barrio JC, Messeguer X, Rozas R (2003)dnasp, DNA polymorphism analyses by the coalescent andother methods. Bioinformatics, 19, 2496–2497.
Sanderson MJ (2002) Estimating absolute rates of molecular evo-lution and divergence times: a penalized likelihood approach.Molecular Biology and Evolution, 19, 101–109.
Sanderson MJ (2003) r8s: inferring absolute rates of molecularevolution and divergence times in the absence of a molecularclock. Bioinformatics, 19, 301–302.
Schneider S, Excoffier L (1999) Estimation of past demographicparameters from the distribution of pairwise differences whenthe mutation rates vary among sites: application to humanmitochondrial DNA. Genetics, 152, 1079–1089.
Simmons MP, Ochoterena H (2000) Gaps as characters in sequence-based phylogenetic analyses. Systematic Biology, 49, 369–381.
Simon C, Frati F, Beckenbach A et al. (1994) Evolution, weighting,and phylogenetic utility of mitochondrial gene sequences and acompilation of conserved polymerase chain reaction primers.Annals of the Entomological Society of America, 87, 651–701.
Stillman CJ (1999) Giant Miocene landslides and the evolution ofFuerteventura, Canary Islands. Journal of Volcanology and Geo-thermal Research, 94, 89–104.
Tajima F (1989) Statistical method for testing the neutral mutationhypothesis by DNA polymorphism. Genetics, 123, 585–595.
Templeton AR (1998) Nested clade analyses of phylogeographicdata: testing hypotheses about gene flow and population history.Molecular Ecology, 7, 381–397.
Templeton AR (2004) Statistical phylogeography: methods ofevaluating and minimizing inference errors. Molecular Ecology,13, 789–809.
Templeton AR, Boerwinkle ES, FC (1987) A cladistic analysis ofphenotypic associations with haplotypes inferred from restrictionendonuclease mapping. I. Basic theory and an analysis of alcoholdehydrogenase activity in Drosophila. Genetics, 117, 343–351.
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysis ofphenotypic associations with haplotypes inferred from restrictionendonuclease mapping and DNA sequence data. III. Cladogramestimation. Genetics, 132, 619.
Templeton AR, Routman E, Phillips CA (1995) Separating popula-tion structure from population history: a cladistic analysis of thegeographical distribution of mitochondrial DNA haplotypes in theTiger salamander, Ambystoma tigrinum. Genetics, 140, 767–782.
Templeton AR, Sing CF (1993) A cladistic analysis of phenotypicassociations with haplotypes inferred from restriction endo-nuclease mapping. IV. Nested analyses with cladogramuncertainty and recombination. Genetics, 134, 659–669.
Vandergast AG, Gillespie RG, Roderick GK (2004) Influence ofvolcanic activity on the population genetic structure of HawaiianTetragnatha spiders: fragmentation, rapid population growthand the potential for accelerated evolution. Molecular Ecology,13, 1729–1743.

3214 L . B I D E G A R A Y - B A T I S T A E T A L .
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd
Vogler AP, Cardoso A, Barraclough TG (2005) Exploring ratevariation among and within sites in a densely sampled tree:species level phylogenetics of North American Tiger beetles(Genus Cicindela). Systematic Biology, 54, 4–20.
Wagner WL, Funk VA (1995) Hawaiian Biogeography: Evolution ona Hot Spot Archipelago. Smithsonian Institution Press, Washington/London.
Whelan F, Kelletat D (2003) Submarine slides on volcanic islands— a source for mega-tsunamis in the Quaternary. Progress inPhysical Geography, 27, 198–216.
Wunderlich J (1991) Die Spinnen-fauna der MakaronesischenInseln. Beitr Araneology, 1, 1–619.
Young ND, Healy J (2002) GAPCODER. http://www.trinity.edu/nyoung/GapCoder/Download.html.
Leticia Bidegaray-Batista is a PhD student under the supervisionof Miquel Arnedo, studying the role of geological and envir-onmental factors on phylogeographic patterns of spiders. NuriaMacías-Hernández is PhD student under the supervision of PedroDromí and Miquel Arnedo, and her research focuses on the studyof factors shaping evolutionary patterns in Canarian Dysdera.Pedro Ororní is associate professor at the Universidad de LaLaguna, Canary Islands, and works on different projects dealingwith the terrestrial biodiversity of the Macaronesian archipelagoes.Miquel Arnedo is associate researcher at the Universitat deBarcelona, with interests in the systematics of spiders and the studyof species diversification in a phylogenetic framework.




















