
Late pleistocene/early holocene environments and climates, fauna, and human occupation in the...
22
' Late Pleistocene/Early Holocene Environments and Climates, Fauna, and Human Occupation in the Argentine Altiplano Jorge Fermindez Consejo Nacional de Investigaciones Cientificas y Tecnicas, CONICET and Instituto Nacional de Antropologia, 3 de Febrero 1370, 1427 Buenos Aires, Argentina Vera Markgraf Institute of Arctic and Alpine Research, University of Colorado, Boulder, Colorado, USA Hector 0. Panarello, Miguel Albero, Fernando E. Angiolini, Susana V alencio Instituto de Geocronologia y Geologia Isot6pica, INGEIS , Ciudad Universitaria, 1428 Buenos Aires, Argentina Mirta Arriaga Division Anatomia Vegetal, Museo Argentina de Ciencias Naturales, B. Rivadavia 470, 1405 Buenos Aires, Argentina The Barro Negro site (23°8 lat., 65.37'W long.) in the Altiplano (Puna) of northwestern Argentina contains a well stratified sequence of remains of Hippidion, the American extinct horse, camelids, and archaeological materials, which is the focus of this study. In addition to establishing a reliable chronology, paleoenvironmental information was obtained based on analyses of pollen and stable isotopes (oxygen and carbon) from bone and mar!. The data indicate that Hippidion was present at the site between 12,000 and 10,000 yr B.P., at a time when Altoandean grasslands had expanded to lower elevations. By 10,000 yr B.P., when modern semi-arid sub-puna scrub had replaced the Altoandean grasslands, only camelids (Lama or Vicugna) were present, simul taneous with the first evidence of local human occupation. This suggests that a climatic shift from cool and moist (winter rain regime) to warm and dry (summer rain regime) conditions took place simultaneously with the disappearance of the American horse and the appearance of camelids and man. INTRODUCTION In trying to understand the relation between early man and the environment during his conquest of the Americas, detailed analysis of the sequence of events has to be the primary focus. Of special interest are studies that analyze periods Geoarchaeology: An International Journal, Vol. 6, No. 3, 251-272 (1991) © 1991 by John Wiley & Sons, Inc. CCC 0883-6353/91/030251-22$04.00
Transcript of Late pleistocene/early holocene environments and climates, fauna, and human occupation in the...

'
Late Pleistocene/Early Holocene Environments and Climates, Fauna, and Human Occupation in the Argentine Altiplano
Jorge Fermindez Consejo Nacional de Investigaciones Cientificas y Tecnicas, CONICET and Instituto Nacional de Antropologia, 3 de Febrero 1370, 1427 Buenos Aires, Argentina
Vera Markgraf Institute of Arctic and Alpine Research, University of Colorado, Boulder, Colorado, USA
Hector 0. Panarello, Miguel Albero, Fernando E. Angiolini, Susana V alencio Instituto de Geocronologia y Geologia Isot6pica, INGEIS, Ciudad Universitaria, 1428 Buenos Aires, Argentina
Mirta Arriaga Division Anatomia Vegetal, Museo Argentina de Ciencias Naturales, B. Rivadavia 470, 1405 Buenos Aires, Argentina
The Barro Negro site (23°8 lat., 65.37'W long.) in the Altiplano (Puna) of northwestern Argentina contains a well stratified sequence of remains of Hippidion, the American extinct horse, came lids, and archaeological materials, which is the focus of this study. In addition to establishing a reliable chronology, paleoenvironmental information was obtained based on analyses of pollen and stable isotopes (oxygen and carbon) from bone and mar!. The data indicate that Hippidion was present at the site between 12,000 and 10,000 yr B.P., at a time when Altoandean grasslands had expanded to lower elevations. By 10,000 yr B.P., when modern semi-arid sub-puna scrub had replaced the Altoandean grasslands, only camelids (Lama or Vicugna) were present, simultaneous with the first evidence of local human occupation. This suggests that a climatic shift from cool and moist (winter rain regime) to warm and dry (summer rain regime) conditions took place simultaneously with the disappearance of the American horse and the appearance of camelids and man.
INTRODUCTION
In trying to understand the relation between early man and the environment during his conquest of the Americas, detailed analysis of the sequence of events has to be the primary focus. Of special interest are studies that analyze periods
Geoarchaeology: An International Journal, Vol. 6, No. 3, 251-272 (1991) © 1991 by John Wiley & Sons, Inc. CCC 0883-6353/91/030251-22$04.00

LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
having the greatest magnitude and rate of environmental change, because such studies may shed light on the potential and limitations of man's adaptability. One of the more recent major periods of large-scale paleoenvironmental and paleoclimatic change is the transition from Pleistocene to Holocene, a time when man began to leave his mark throughout the Americas. It was during this period that glaciers receded to their present locations, that the plant communities began to regroup and reestablish their interglacial-type distributions, and that a substantial number of large mammals became extinct. From a number of sites in central Peru (Lynch, 1983; MacNeish et al., 1981; Rick, 1988; Wheeler Pires-Ferreira et al. , 1976), southern Chile (Dillehay, 1989), and in Patagonia (Bird, 1988; Borrero, 1981; Cardich, 1985), there is evidence that early man coexisted between at least 12,000 and 10,000 yrs B.P. with the late Pleistocene extinct fauna. In the North American context th is gave rise to the overkill hypothesis (Martin, 1973) that was later extended to also apply globally (Martin, 1984). On the other hand, climatically induced environmental changes had also been quoted as a potential cause of extinction (Guilday, 1967; Graham, 1979), either by supposedly affecting the dietary habits of the animals, or altering the niche diversity in the environment (Guthrie, 1984).
The detailed study of the timing of the late Pleistocene faunal overturn and its relation to early man constitutes the first step to approach this archaeological and paleontological problem. In order to remove site-related ambiguities that could obscure the understanding of the extinction process, each local or individual recorded event would have to be calibrated (Mead and Meltzer, 1984). Radiocarbon dates on organic materials associated with extinct fauna frequen tly show a great range of ages. When radiometric dates from the Argentine Pampa region are compared with dates from Patagonia, both important areas in South America where late Pleistocene extinct fauna has been dated (Pascual et al., 1966; Marshall et al., 1984), a great range of ages is found. According to such records, the "death blow" that affected sabertooth cats, megatherids, giant ground sloths, horses, and giant armadillos, among many others, occurred in the time span between 12,000 and 5000 yr B.P., i.e., late Pleistocene to mid-Holocene times (Marshall et al., 1984, Table 1; Borrero, 1977). Such a chronological range, in some cases for the same taxa and in the same locality, represents a major challenge to the understanding of the extinction process. More likely, however, such age range is due to sampling errors, sample contamination, is stratigraphic reversals of faunal remains, especially when the material dated is fossil bones (Stafford et al., 1987, 1988).
The Barro Negro stratigraphic section in the Altiplano (Puna) of Northwestern Argentina, at 3820 m elevation, contains sediments which cover the late Pleistocene/Holocene transition. Beds oflacustrine and ftuviatile origin contain extinct fauna, overlain by extant fauna, and archaeological remains, providing a unique opportunity to study in detail the character and timing of the faunal turnover and ofpaleoenvironmental conditions. Apart from addressing the age of the events, we studied paleoenvironmental changes by analyzing 1) pollen
252 VOL. 6, NO. 3

:
LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
assemblage changes that could reflect changes in the regional vegetation and hence regional climate; 2) oxygen and carbon isotope ratios in the biogenic carbonate layers, which would reflect changes in local precipitation and temperature regime; and 3) carbon isotope ratios in the collagen of fossil bones that would allow us to identify differences in the animals' diet, reflecting differences in composition of the vegetation.
LOCATION OF THE SAMPLES AND FOSSIL CONTENT
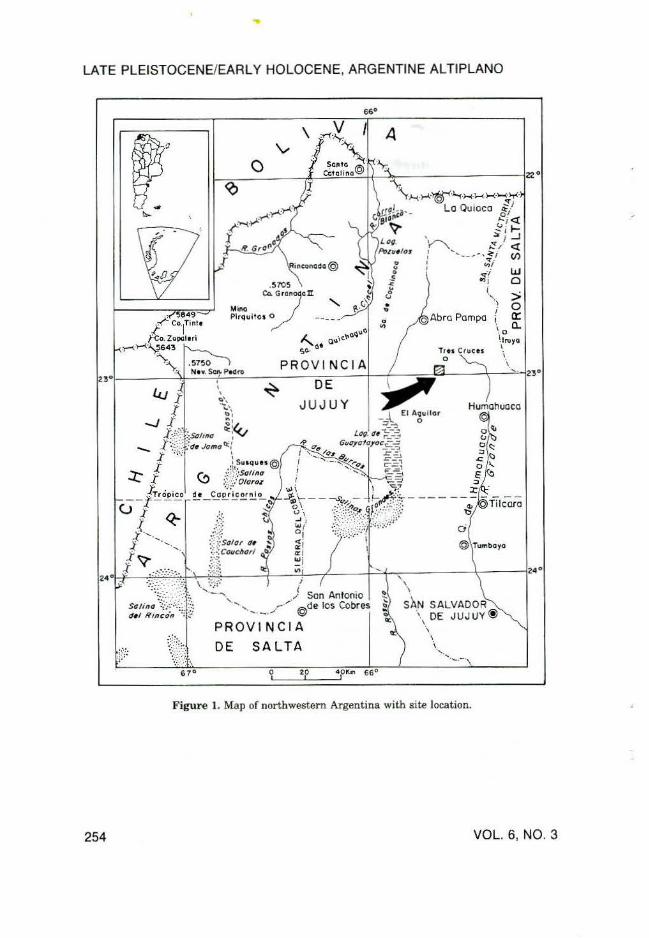
Sierra del Aguilar is a 5200 m high, isolated mountain range located to the east of the Central Andes in the province of Jujuy, northwestern Argentina (Figure 1). The base of the eastern flank of the range is a fioodplain with elevations between 3700 and 4000 m, dissected by many rivers and ephemeral streams, tributaries of the Humahuaca drainage system. In this barren, semiarid region, the annual rainfall of about 300 mm/yr falls only in the summer months, resulting in heavy stream erosion.
The arroyo Barro Negro is one of the streams draining the northeastern slope of the Aguilar range. Near the settlement of Barro Negro (23°S lat.; 65°37' W long., 3820 m elevation), the principal stream and its small tributaries (Quebrada Linda, Quebrada Las Piedras, and Zanjon Largo) cut a Quaternary sediment fill of a basin circumscribed by an outcrop of Cretaceous dolomitic limestone (Figure 2).
River downcutting in the valley, yet incomplete, exposed well stratified and fine-grained sediments with lateral facies changes from clays to peats and marls, suggesting deposition in shallow ponds and marsh environments along a slow-flowing stream. Three main sedimentary episodes are represented in the several sections studied which vary in total thickness between 7.5 m and 10 m, a basal stratum of primarily lacustrine sediments, peats, clays, and marls, a middle stratum of sands, and an upper stratum of peats, silts, and sands (Figures 3 and 4).
The ea. 2.5 m thick basal stratum I is composed of peaty silts, clays, sand layers, and marls towards the top (stratum la). The faunal content consists of bones and crania of a South American extinct equid, identified as Hippidion (Alberdi et al., 1986). So far at least five individuals have been recovered, and skeletal remains representing ten additional disarticulated or semiarticulated fossil equids have been more recently discovered.
No bones of other mammal species were associated with the Hippidion bones, nor cultural remains such as lithics or charcoal. The bones bear no marks of longitudinal or transverse cuts, striae, or other signs of human manipulation.
The overlying stratum II is about three m thick, and is composed of sand or silty sands without obvious stratification. It contains disarticulated and broken camelid bones (Lama or Vicugna), clearly associated with lithic industry and charcoal. The lithic artifacts were represented by flakes, blanks, and preforms, manufactured of local raw material (quartzite). A knife scraper found in the
GEOARCHAEOLOGY: AN INTERNATIONAL JOURNAL 253
.. LATE PLEISTOCENEIEARL Y HOLOCENE, ARGENTINE ALTIPLANO
A
0 20 40Km 66°
Figure 1. Map of northwestern Argentina with site location.
254 VOL. 6, NO. 3

LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
0~======~---------2Km Figure 2. Site location map of the Barro Negro sections. Elevations given in meters; contour intervals 25 m.
lower part of stratum II is an indicator of the type of activity, although no projectile points have been found indicative of a specific culture.
The uppermost stratum Ill is about two m thick, consists of layers of sand or silty sand, silts and interbedded peats, marls and buried soils, and also contains camelid bones.
Complete and broken camelid bones from strata II and Ill were compared with modern osteological material of Llama glama, L. pacos, L. guanicoe, and
GEOARCHAEOLOGY: ANINTERNATIONALJOURNAL 255

LATE PLEISTOCENEIEARLY HOLOCENE, ARGENTINE ALTIPLANO ..
0 Polletl section
e Hippidion bones
• '4C dote
A Camelitl bones
Figure 3. Sediment section Barro Negro, showing strata (in roman numbers), and location offossil bones, of samples for radiocarbon and pollen analysis.
Vicugna vicugna, but identification to species was considered impossible because of poor preservation. The majority of the camelid bones showed the typical spiral conchoidal intentional fractu ration of the epiphyses for marrow extraction, possibly indicating human use (Stanford et al., 1981).
MATERIALS AND METHODS
To establish a reliable chronology, sediments at several locations were collected for radiocarbon dating, including peats, bones, and marls. Five peat samples were obtained from the basal stratum I, three from the Barro Negro section, and two from the Quebrada Linda section, ea. 100 m upstream. In addition, bones of Hippidion from these peat strata were selected for collagen extraction, as well as three bulk samples from the marllayer in the upper part of stratum I. In stratum II datable material is scarce. Besides one wood sample, found in the Bano Negro section, two samples of organic mud from the Zanjon
256 VOL. 6, NO. 3

LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
@ Isotope sectio11
• "C tltul!
• Hippidion b011eii
! Cumelitl bones
Figure 4. Sediment section Barro Negro, near locality in Figure 3, showing strata (in roman numbers), location of fossil bones, and samples for radiocarbon and stable isotope analysis.
Largo section were dated. Ten charcoal and peat samples were dated from stratum III.
Radiocarbon and stable isotope analyses were done at the lnstituto de Geocronologia y Geologia Isot6pica (INGEIS) in Buenos Aires (Argentina).
Pretreatment for radiocarbon analysis consisted of the following steps: a) peat and organic muds were pretreated by several bot washes with 2% HCI and NaOH, to remove humates and carbonates, respectively; b) marl samples were heated four hours at 525°C to remove organic carbon; X-ray diffraction analysis revealed the absence of dolomite; c) bones were first thoroughly cleaned by picking and ultrasound treatment, followed by collagen extraction and purification according to the techniques used by Longin (1971), modified by DeNiro and Epstein (1978, 1981), and Schoeninger and DeNiro (1984).
The dating method employed was liquid scintillation counting of synthesized benzene described by Albero and Angiolini (1983, 1985). Samples were converted to C02; carbonates by perchloric acid dissolution, and organic carbon by combustion in an oxygen stream in a combustion system. Mud samples with
GEOARCHAEOLOGY: AN INTERNATIONAL JOURNAL 257

LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO
Table I. 1>13C and 14C ages of stratigraphically collected materials.
Sample No. Stratum Material o13C%o (PDB) Age (yr B.P.) Fauna! Association
AC-0748 m Peat - 25.3 510 ± 70 Camelidae AC-0681 m Peat 710± 80 Camelidae AC-0740 m Peat -27.6 940 ± 80 Camelidae AC-0747 Ill Peat -24.0 1000 :t 80 Camelidae AC-673 Ill charcoal -23.7 1140 ± 80 Camelidae AC-0738 III Peat - 25.4 1270 :t 80 Camelidae AC-0682 m Peat - 25.0 1680 ± 120 Camelidae AC-0739 Ill Peat -27.6 1690 ± 80 Camelidae AC-0680 m Peat -24.3 2890 ± 110 Camelidae AC-0746 ill Peat - 27.5 3470 :t 90 Camelidae AC-1154 II Wood 8320 :t 150 Camelidae AC-0742 II Organic mud - 24.8 9050 ± 140 Camelidae AC-0743 II Organic mud -28.8 9200 :t 140 Camelidae AC-0975 la Mar I -3.6 13,400 ± 400 Hippidion AC-0976 la Mar I -3.8 11,600 :t 160 Hippidion AC-743 la Mar I -5.0 9870 :t 220 Hippidion AC-0672 I Peat -25.2 10,200 ± 170 Hippidwn AC-0745 I Peat - 25.0 10,200 ± 140 Hippidwn AC-0679 I Peat - 23.4 10,740 ± 140 Hippidwn AC-0744 I Peat - 28.7 12,300 ± 170 Hippidion AC-0735 I Peat -26.8 12,530 ± 160 Hippidion AG-0969(1) I Bone Collagen -18.0 9120 ± 130 Hippidion AC-0969(2) I Bone Collagen -18.2 11,500 :t 400 Hippidion
low carbon content were "wet combusted" according to Albero and An.giolini (1985). All radiocarbon results have been normalized to ()13C = - 25%o (PDB) as recommended in Radiocarbon.
For the measurements of Stable Isotope Ratios in marl (180 /160 and 13Cf12C)-, 18 samples of marl from stratum la were crushed and ground to a grain size of less than 100 mesh. Powdered material was heated to 350°C under vacuum in order to destroy organic matter. Thirty mg of samples were hydrolyzed with two ml of concentrated phosphoric acid at 25°C according to the technique of McCrea (1950). The C02 gas was purified cryogenically and transferred to a Micromass 602-D McKinney type mass spectrometer using Carrara marble as a working reference.
Stable carbon isotopes were analyzed on collagen extracted from bones of Hippidion from strata I and la, of camelids from strata 11 and Ill, and of modern camelids from different elevations in the area. Only cortical tissue of diaphysis bone samples was chosen, falling into class I and II (very well to well preserved), according to Stafford et al. (1988). The samples were ultrasonically cleaned and powdered, and tested for their whole bone nitrogen content to assess collagen preservation during diagenesis (Stafford et al., 1988). Hippidion bone with less than 3.5% total nitrogen were not analyzed. Total collagen content and elementary carbon was measured as well (Ambrose and DeNiro, 1989).
258 VOL. 6, NO. 3

LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE AL TIPLANO
Table II. Stable isotope values of the analyzed mar! samples.
Sample No. 813C%o (PDB) 8180%o (PDB)
818 - 4.0 ± .1 -6.6 ± .2 817 -4.3 ± .1 -6.3 ± .2 8 16 -7.1 ± .1 -7.9 ± .2 815 -5.2 ± .1 -6.0 ± .1 814 -3.9 ± .1 - 5.2 ± .1 813 - 4.1 ± .1 - 5.4 ± .1 812 - 3.9 ± .1 - 5.4 ± .1 811 -3.8 ± .1 -5.4 ± .1 810 -3.8 ± .1 - 5.5 ± .1 S9 -3.8 ± .1 - 5.5 ± .1 88 -3.8 ± .1 -5.6 ± .1 87 - 3.8 ± .1 -6.1 ± .1 86 - 3.7 ± .1 - 6.0 ± .1 85 -2.2 ± .1 - 6.2 ± .2 S4 -2.2 ± .1 - 5.0 ± .2 83 -2.0 ± .1 - 4.9 ± .2 82 -1.9 ± .2 - 4.7 ± .2 81 - 1.7 ± .2 -5.8 ± .1
Ca. eight mg of sample of bone collagen was mixed with 100 mg vanadium pentoxide in a pyrex vial and flame sealed under a vacuum of about 10- 4 m bar. The sample was combusted for eight hours at 550°C in an electric furnace (Panarello, 1987). C02 gas was then purified by cryogenically removing water and non-condensable gases, trapped with liquid nitrogen, and transferred to a Micromass 602-D McK.inney type mass spectrometer. The same pretreatment was used for modern plant materials and animal feces.
Results are expressed as 8, defined as follows:
RS o :::; -R - 1 X 1000%o (PDB) ST
where 8 = 813C or o180 isotopic deviation in per roil R = 13CP2C or 180 J160 respective isotopic ratios S:::; sample
ST = standard
The standard used for 180 and 13C is PDB as defined by Craig (1957). The standard measurement error is ± 0.1%o; Table 11 shows the isotopic measurements from the marls of the Barro Negro site; Table Ill stable carbon isotope values from modern bone collagen, modern plant materials, and animal feces; and Table IV fossil bone collagen.
Pollen samples from the peat layers in stratum I and the overlying clay layers in stratum II of section B (Figure 2), comprising the interval between 12,300 and 9000 yr B.P. were pretreated and analyzed using standard techniques (Faegri and Iversen, 1975).
GEOARCHAEOLOGY: AN INTERNATIONAL JOURNAL 259

LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE AL TIPLANO
Table Ill. Stable carbon isotope ratios and estimated percent ofC3 plants, of modern collagen and feces of herbivores and carnivores from different elevations. Percentage ofC3 plants was estimated assuming a 813C of - 21.4%o for bone collagen of herbivore feeding on 100% C3 plants (Van der Merwe, 1982).
Animal Species Elevation (m) o13C%o (PDB) % C3 plants
COLLAGEN Lamaglama 3600 -14.1 48 Lamaglama 3700 - 16.8 67 Puma concolor 3700 - 16.1 Lamaglama 4000- 4400 -20.3* 92 Vicugna vicugna 4000-4700 - 19.3** 84 Lama guanicoe 4000-4500 - 19.2* 84 Aries ovis 3900-4200 - 18.3 78 Capra hircus 3900-4000 - 18.3 78 FECES Lama guanicoe 4300 - 26.8 100 Vicugna vicugna 4200 - 25.8 100 Vicugna vicugna 4400 - 26.4 100 Lamagla.ma 4000 - 26.7 100
*Average of 4 specimens **Average of 5 specimens
RESULTS. AND DISCUSSION
Radiocarbon Chronology
The radiocarbon dates (Table I) of 12,300 ::!: 170 (AC-0744) and 12,530 ± 160 (AC-0735) yr B.P. on peats near the base of stratum I represent the first and
Table IV. Whole bone nitrogen (%), carbon : nitrogen ra t io, stable carbon isotope ratios, and estimated percent of C3 plants in fossil bone collagen from the Barro Negro site.
Taxon Sediment Whole Bone C: N o13C%o (PDB) % C3 plants stratum % N
Hippidion I 4.48 2.6 - 18.9 82 Hippidion I 3.49 2.6 - 20.4 93 Hippidion I 3.57 2.6 -19.1 83 Hippidion I 2.5 -19.3 83 Hippidion la 2.6 - 18.9 82 Hippidion la 2.4 - 19.3 83
Camelidae II 2.7 -17.3 70 Camelidae II 2.6 - 17.3 70 Camelidae IT 2.6 - 16.6 65 Camelidae II 3.0 - 17.6 73 Camelidae III 2.4 - 17.2 70 Camelidae Ill 2.6 - 16.9 68 Camelidae III 2.8 -17.3 70 Camelidae Ill 2.7 -17.3 70
260 VOL. 6, NO. 3

LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO ...
oldest dates associated with Hippidion in the Argentine Puna, and the dates of 10,200 ± 140 (AC-0745) and 10,200 ± 170 (AC-0672) yr B.P. from the upper levels of stratum I the youngest dates.
Although concordant, the bone collagen dates are younger than dates on associated peats. Thus, the lower Hippidion bone collagen date of 11,500 ± 400 yr B.P. (AC-0969-2) compares to the peat age of 12,530 ± 160 yr B.P. (AC-0735), while the younger bone date of9120 ± 130 yr B.P. (AC-0969-1) compares to the peat age of 10,200 ± 140 yr B.P. (AC-0745).
When comparing peat and marl dates we would expect a similar problem with age differences. Except for the upper mar] age o£9870 ± 220 yr B.P. (AC-0974), which is not significantly different from the upper peat age of 10,200 ± 140 yr B.P., the other two marl samples, on the contrary, are significantly older than those of the stratigraphically correlated peat. This can be explained by a probable contamination with dissolved dead carbonates, originating from nearby outcrops of Cretaceous dolomitic limestones.
The oldest dates on bulk organic muds of stratum II containing broken camelid bones and lithic artifacts are 9200 ± 140 (AC-0743) and 9050 ± 140 (AC-0742) yr B.P. The overlying stratum ID was deposited between 3470 ± 90 (AC-0746) and 510 ± 70 (AC-0748) yr B.P. (Table I).
Thus, the boundary between strata I and II, which separates extinct from extant fauna (Table I), lies between 10,200 ± 140 yr B.P., and 9200 ± 140 yr B.P. The local extinction of the American horse in the Puna correlates with other dates of faunal extinction from other South American sites (Pascual et al., 1966; Marshall et al., 1984; Markgraf, 1985a).
Pollen Analysis
Pollen analysis of samples with statistically significant pollen counts (i.e., a minimum of 200 grains/sample) from the Barro Negro section dated between 12,300 and 9200 yr B.P., covering the transition between the strata with Hip pidion and with camelid remains. Unfortunately, many samples, especially between 11,000 and 10,000 yr B.P., are barren of pollen. Instead, charcoal particles are abundant, suggesting fires.
The five levels analyzed (Figure 5) are characterized by codominance of Gramineae (50 to 60%), Compositae (10 to 30%), and herbaceous taxa (15 to 40%), including (in order of decreasing frequency) Plantago, Umbelliferae (Azorella and othe1·s), Cruciferae, Caryophyllaceae, Acaena, Chenopodiaceae, Ranunculaceae, etc. Pollen from tree taxa (Myrtaceae, Podocarpus, Alnus), originating from the upper elevation forests to the east, only occur in traces.
Proportions of pollen taxa are different in the samples that predate 11,000 yr B.P . than in the younger samples. Prior to 11,000 yr B.P. herbaceous taxa are a prominent feature in the assemblages, characterized by a high taxa diversity, whereas Gramineae and Compositae (the latter especially in the 12,000 yr B.P. sample) are less abundant. After 11,000 yr B.P. Compositae and Gramineae are far more abundant, and herbaceous taxa occur in low amounts
GEOARCHAEOLOGY: AN INTERNATIONAL JOURNAL 261

LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO
STABLE ISOTOPES POLLEN (%)
d 13c (%o) PDB J 180 (%o) PDB
- 6 -4 -2 - 8 -6 - · •b 0 20 20 0 40 '1(,
Y r B. P. r----1--~--L-~-L--~--'--t::::::=::4::::::~~........,."1:=::::::::::-,ex cl.
10,000 S-18
11,000
12,000
Figure 5. Plot of stable isotope data W3C and 8180 ) and pollen data according to radiocarbon chronology developed independently for each section.
and with low diversity, primarily represented by Chenopodiaceae, Gomphrena, and Ephedra. Using floristic information from high elevation environments in northern Argentina (Ruthsatz and Movia, 1975; Halloy, 1983), high proportions and diversity of herbaceous taxa are characteristic for the high elevation AJtoandean vegetation, a cold and humid environment. Predominance of Compositae and Gramineae, on the other hand, suggests more xeric Prepuna.
When comparing this pollen sequence with the only other nearby high elevation pollen record, El Aguilar (Markgraf, 1985b), which is located about 200 m higher than the Barro Negro site (Figure 1), interesting paleoenvironmental aspects arise. There is a remarkable similarity between the early Holocene pollen spectra from El Aguilar (the record extends only back to 10,000 yr B.P.) and the pre-11,000 yr B.P. pollen spectra from Barro Negro. This could be taken to suggest that the Altoandean environment, today dominating above 4100 m elevation, in late Pleistocene times had expanded to lower elevations. Its up-
262 VOL. 6, NO. 3

LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO
ward retreat began at the Barro Negro site after 11,000 yr B.P. At the El Aguilar site Altoandean vegetation remained until after 7500 yr B.P.
Unfortunately, none of the Bolivian pollen records from comparable en vironments (Graf, 1979) covers this transitional period. The geographically closest record is from central Peru, from the J unin Plain at about latitude 11 os (Hansen et al., 1984). In contrast to the Barro Negro data this record shows prior to 11,000 yr B.P.less Gramineae, but more Compositae and other subpuna types instead, suggesting that climates were drier instead of wetter than today. This difference between the Pemvian and the Argentine records may actually be real, reflecting the different climatic response to the late Pleistocene climatic change (Markgraf, 1989).
STABLE ISOTOPE MEASUREMENTS
Biogenic Carbonates
The application of oxygen isotope ratios as a climatic indicator using fresh water carbonates was demonstrated by Stuiver (1970) who used the known relation between temperature and 180/160 ratios in modern precipitation. The technique was subsequently employed for paleoclimatic and paleohydrological studies in many parts of the world (Fritz and Poplawsky, 1974; Fritz et al., 1975; Eicher and Siegenthaler, 1976; 1983; Eicher et al., 1981; Fontes et al., 1981, 1983).
Chemical and X-ray diffraction analysis of the biogenic carbonate section from stratum la shows that calcium carbonate content ranges between 40 and 85%, mixed with sand, silt, clay, and organic matter; no clastic or reprecipitated magnesium limestone was found. A total of 18 samples were analyzed for stable isotopes: five from the basal portion, ten in the central, and three in the upper portion that forms the contact with the Holocene sediments.
The marl sediment analyzed for stable isotopes (Figure 4) was inferred to date between 10,000 and 11,000 yr B.P., using peat ages from corresponding levels in the adjacent sections. Results are listed in Table Ill and plotted in Figure 5. Two years of 8180 analysis of rain water at the IN GElS station at Tres Cruces, where mean annual temperature is 9°C, yielded an isotopic value of -15%o (Fernandez and Panarello, 1988).
Assuming that the water was in isotopic equilibrium at the time of carbonate deposition, the equation of O'Neill et al. (1969) allows inferences on past temperatures. However, other assumptions have to be considered as well, which could lead to contrasting climatic interpretations. Most oxygen isotope analyses from carbonate sediments in lakes (Fritz et al., 1975; Eicher et al., 1983) assume an increase in 180 to relate to either a warming of meteoric water, a decrease in glacial meltwater, or increased evaporation. In all these cases, a less negative 180f160 ratio would imply warmer temperatures. Effect of glacial meltwater can probably be excluded for the Barro Negro section. Differences in evaporation regime, on the other hand, must certainly have been involved, judging from
G EOARCHAEO LOGY: AN INTERNATIONAL JOURNAL 263

LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO
the carbonate precipitates. But these differences would probably only explain part of the oxygen isotope signal, because the system is not a closed basin. Probably the principal factor influencing the isotopic signal would relate to changes in seasonality of precipitation. This has also been suggested for the isotopic records of the Quelccaya ice cores (Thompson et al., 1985). There, the more negative 180 values were interpreted as corresponding to summer (warm, moist season) precipitation, and the less negative values to winter (cold, dry season) precipitation. Using this assumption, the climatic interpretation of the Barro Negro isotope data then calls for two intervals of primarily winter precipitation, around 11,000 (S-2 to 4; Table I!) and around 10,500 yr B.P. (S-8 to 14; Table Il) separated and followed by summer precipitation intervals.
Turning to the carbon isotopic data, the overall similarity between the trends of 180 and 13C (Figure 5) suggests that the mechanism of carbonate precipitation probably did not change substantially through time. Interpretation of the carbon 13 results, i.e., their meaning in terms ofpaleoenvironments, is still hotly debated. Commonly it is assumed to be related to bioactivity, which in turn is related to temperature. When plotting o180 values against o13C values (Figure 5), differences between the two curves emerge. During the earlier phase of less negative o180 values, o13C values are the least negative, suggesting low bioactivity in the aquatic system. During the following phase of less negative o180, o13C values are increasingly more negative, suggesting gradual increase in bioactivity. The most negative o13C values occur prior to 10,500 yr B.P., suggesting that in fact there is a substantial further shift to warmer temperatures at that time.
Bone Collagen
Fractionation during photosynthetic carbon fixation produces a change in the 13C/12C ratios of the atmospheric C02 (o
13C = - 7%o) due to the differential fixation of 12C, the lighter isotope. Plants that follow the C3 photosynthetic pathway average o13C of - 26.5%o, whereas plants that follow the C4 photosynthetic pathway average - 14.5%o (Bender, 1968, 1971; Smith and Epstein, 1971; Van der Menve, 1982). Plants that follow the CAM (Crassulacean Acid Metabolism) have intermediate 13CP2C ratios and are not considered here, because they are not a dietary source for camel or horse.
Distribution of C3 and C4 plants is related to differences in the plants' ecological tolerances in terms of moisture, temperature, and luminosity. C3 plants grow preferentially in cool, moist habitats, under winter rain regime (Hattersley, 1983) while c4 plants have a competitive advantage in warm, highly insolated habitats (Bjorkman and Berry, 1973) with summer rain regime (Hat tersley, 1983). Transects across ecologically different environments showed significant differences in the proportions of C3 vs. C4 plants, many of them in the grass family (Tiezsen et al., 1979; Ambrose and DeNiro, 1989).
The use of stable carbon isotopes in paleoecological research is based on the assumption that the isotopic composition of the plant is passed on to the tissue
264 VOL. 6, NO. 3
LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
S !3c modern Altitude
m
4500 0 bone collagen 1 I ~tT
8 feces
4100
3500L---.---.----.---,r---,---.---~----
s l3c - i 4 -16 ·i8 -20 - 22 -24 -26 (o/oo)PDB
8 13c fossil bone
14c a.P. Comelidoe
<~0.000 ye
) lO.OOOyrl._ ----.-~ ---.--..-~-~--,~-- Hippidion
S13c -14 -16 ·18 -20 lo/oo) PDB
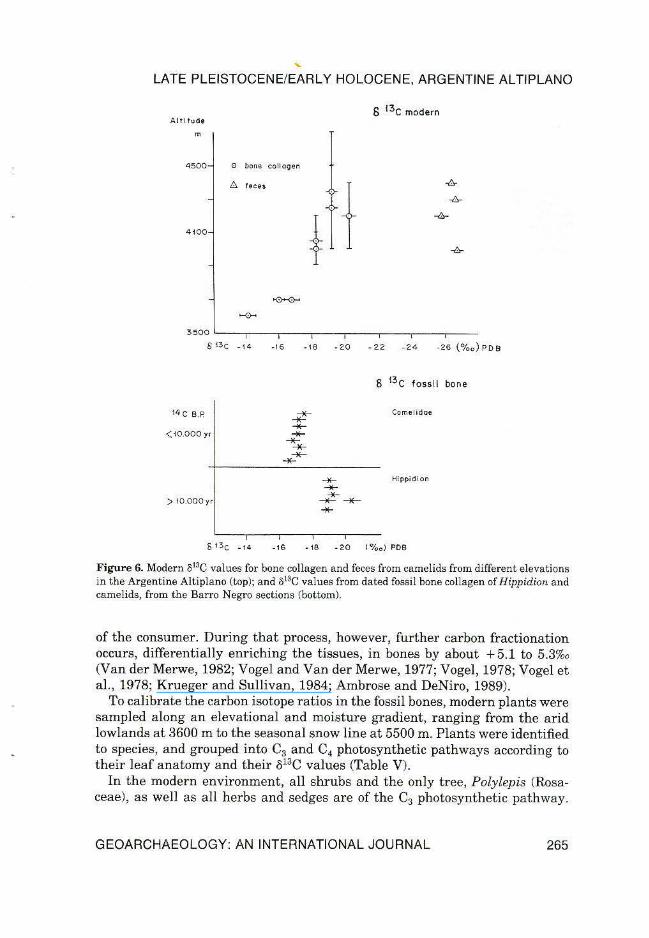
Figure 6. Modern 813C values for bone collagen and feces from camelids from different elevations in the Argentine Altiplano (top); and 813C values from dated fossil bone collagen of Hippidion and camelids, from the Barro Negro sections (bottom).
of the consumer. During that process, however, further carbon fractionation occurs, differentially enriching the tissues, in bones by about + 5.1 to 5.3%o (Van der Merwe, 1982; Vogel and Van der Merwe, 1977; Vogel, 1978; Vogel et al., 1978; Krueger and Sullivan, 1984; Ambrose and DeNiro, 1989).
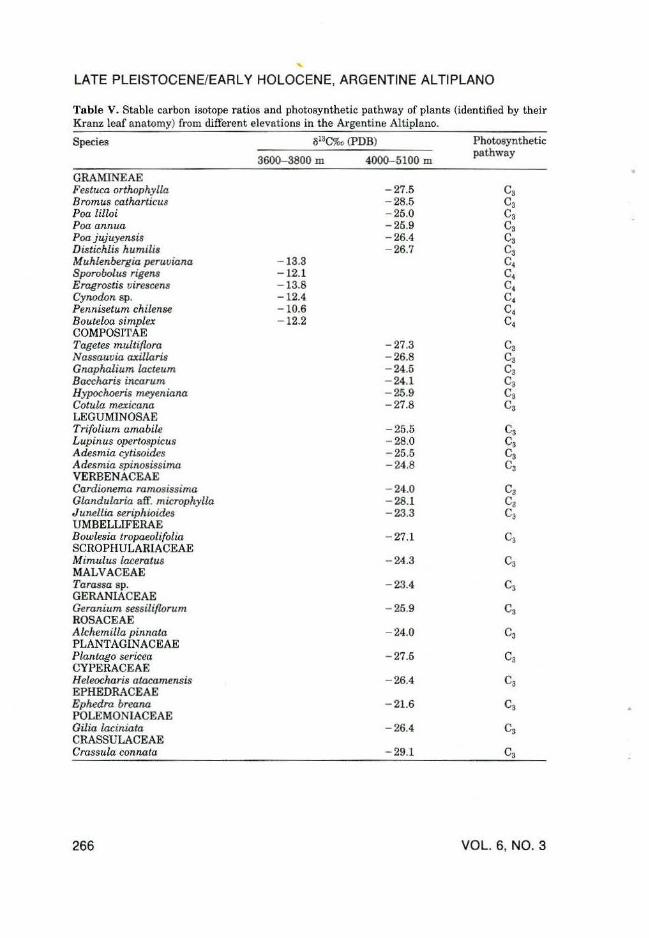
To calibrate the carbon isotope ratios in the fossil bones, modern plants were sampled along an elevational and moisture gradient, ranging from the arid lowlands at 3600 m to the seasonal snow line at 5500 m. Plants were identified to species, and grouped into c3 and c4 photosynthetic pathways according to their leaf anatomy and their 813C values (Table V).
In the modern environment, all shrubs and the only tree, Polylepis (Rosaceae), as well as all herbs and sedges are of the C3 photosynthetic pathway.
GEOARCHAEOLOGY: AN INTERNATIONAL JOURNAL 265
LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO
Table V. Stable carbon isotope ratios and photosynthetic pathway of plants (identified by their Kranz leaf anatomy) from different elevations in the Argentine Altiplano.
Species 813C%o (PDB)
GRAMINEAE Festuca orthophylla Bromus catharticus Poa lilloi Poa annua Poa jujuyensis Distichlis humilis M uhlenbergia peruviana Sporobolus rigens Erogrostis uirescens Cynodon sp. Pennisetum chilense Bouteloa simplex COMPOSITAE Tagetes multiflora Nassauuia axillaris Gnaphalium lacteum Baccharis incarum Hypochoeris meyeniana Cotula mexicana LEGUMINOSAE Trifolium amabile Lupinus opertospicus Adesmia cytisoides Adesmia spinosissima VERBENACEAE Cardionema ramosissima Glandularia at.f. microphylla J unellia seriphioides UMBELLIFERAE Bowlesia tropaeolifolia SCROPHULARIACEAE Mimulus laceratus MALVACEAE Tarassa sp. GERANIACEAE Geranium sessiliflorum ROSACEAE Alchemilla pinnata PLANTAGINACEAE Plantago sericea CYPERACEAE H eleocharis atacamensis EPHEDRACEAE Ephedra breana POLEMONIACEAE Gilia laciniata CRASSULACEAE Crassula connata
266
3600-3800 m 4000-5100 m
- 27.5 - 28.5 - 25.0 - 25.9 -26.4 - 26.7
- 13.3 - 12.1 - 13.8 -12.4 -10.6 - 12.2
- 27.3 - 26.8 - 24.5 - 24.1 - 25.9 - 27.8
- 25.5 - 28.0 - 25.5 - 24.8
- 24.0 - 28.1 - 23.3
- 27.1
- 24.3
- 23.4
- 25.9
- 24.0
- 27.5
- 26.4
- 21.6
- 26.4
- 29.1
Photosynthetic pathway
VOL. 6, NO. 3

LATE PLEISTOCENEJ~ARL Y HOLOCENE, ARGENTINE AL TIPLANO
Among the grasses, however, there are several taxa of the C4 pathway (Muhlenbergia atacamensis, Eragrostis virescens, Pennisetum chilensis, Sporobolus rigens, etc.; see Table V). At elevations above 4000 m Muhlenbergia atacamensis and Sporobolus rigens are the only C4 plants occurring quite rarely. In this environment plant materials in animal feces yielded a range of o13C between - 25.8%o and - 26.8%o, indicating 100% C3 diet (Table ill). Bones from these elevations (above 4000 m) for domestic (llama) and wild (vicugna) camelids gave cS13C values between -19.2%o and - 20.3%o, (i.e., - 24.5%o and - 25.4%o in the diet), indicating also essentially a 100% C3 diet (Table Ill). At elevations between 3600 and 4000 m, however, C4 plants are far more abundant (Table V) and the cS13C values in bones from domestic animals (llama, goat, and sheep) are isotopically heavier, ranging from - 14.1%o to -18.3%o (i.e., -19.2%o to - 23.4%o in the diet), which corresponds to a C3/C4 ratio between 50% and 80% (see Table liD.
Thus, the cS13C ratios of the fossil bone collagen of Hippidion (listed in Table IV and plotted in Figure 6), dated between 12,000 and 10,000 yr B.P. with their o13C range between -18.9%o and - 20.4%o, suggest a diet similar to the one of the high elevations today with 10% or less of C4 plants. The camelid bones, from the strata that dated younger than 10,000 yr B.P. have o13C values of - 16.6o/oo to - 17 .6%o, suggesting about 30% of c4 plants in their diet.
The overall conclusion from the carbon isotope ratios in fossil bone is that at the time when H ippidion populated the region, prior to 10,000 yr B.P., the higher elevation vegetation had expanded downslope. After 10,000 yr B.P. when camelids roamed instead, the vegetation resembled the modern one. In climatic terms, this implies that between 11,000 and 10,000 yr B.P. conditions shifted from a moister and cooler regime to a warmer and drier regime, comparable to the modern one.
CONCLUSIONS
The focus of this study was to search for an explanation for the timing and character of environmental change that could shed light on the extinction of Hippidion in the highlands of NW Argentina by about 10,000 yr B.P., and the subsequent appearance of camelids and man. The primary effort concentrated on the accuracy and reliability of the radiocarbon chronology by dating different types of materials represented in the sediment sections, including organic materials, marls, and bones. Questions concerning paleoenvironmental changes were addressed by analyzing pollen on one hand, and stable isotope ratios on the other hand, 180 /160 in biogenic carbonates, and 13C/12C in bone collagen. To interpret the fossil isotopic evidence, modern materials had to be analyzed as well.
In evaluating radiocarbon dates in the context of late Pleistocene faunal extinctions, Mead and Meltzer (1984) suggested a ranking scheme for different datable materials. Whereas bone collagen was formerly considered preferential to peat for dating, more recently serious doubts arose about the reliability of
GEOARCHAEOLOGY: ANINTERNATIONALJOURNAL 267

LATE PLEISTOCENE/EARL Y HOt OCENE, ARGENTINE ALTIPLANO
collagen dates, due to the presence of non-indigenous organic materials (Taylor, 1982). As noted by Mead and Meltzer (1984), e.g., all dubious dates younger than 10,000 yr B.P. related to late Pleistocene extinction in North America were obtained from bone collagen. When analyzing the reason for problems with bone dates, Stafford et al. (1987, 1988) found that beyond certain levels of degradation, bulk bone dates are unreliable unless the bone protein is co11agenous and methods are used to totally remove the humate contaminants.
Samples from the Barro Negro site confirm that bulk collagen dates are consistently younger than associated peat dates. Peat dates also seem more reliable than related marl dates, that through incorporation of old carbon appear too old by about 13%.
Based on the pollen data that show dominance of herbaceous grassland prior to 11,000 yr B.P. at elevations that today are characterized by puna scrub vegetation, climates had been moister and cooler than today. Support for this interpretation comes from the carbon isotope analysis of fossil bones, that reflects the paleodiet of the respective animal. Apparently, Hippidion's diet consisted primarily ofC3 plants with only minor proportions ofC4 plants. This suggests that during the time when Hippidion existed, the plant composition in terms of C3 vs. C4 plants at 3800 m resembled the one found today above 4000 m. After 10,000 yr B.P., when camelids roamed the area, and were hunted by man, the paleodiet resembled the modern one at these elevations.
Paleoclimatic interpretation of the oxygen isotope data is more complex because a combination of different factors can lead to the same isotopic ratio. Assuming that the most likely parameters determining the oxygen isotope ratios are evaporation and seasonality of precipitation, the results suggest that before 11,000 yr B.P. and to a lesser degree around 10,500 yr B.P. winter precipitation dominated. Closer to 10,000 yr B.P. summer-dominated precipitation patterns developed. The oxygen as well as the carbon isotope data, both showing changes towards higher temperatures after 11,000 yr B.P., seem to confirm the regional paleoenvironmental information derived from the pollen and paleodietary data.
In summary, paleoenvironmental information from several independent lines of evidence suggests that in the Altiplano of northwestern Argentina, the transition from late Pleistocene to Holocene was characterized by reduction of cool and moist Altoandean grasslands. The climatic interpretation calls for a shift in seasonality of precipitation from a winter rain regime to summer rain regime, coupled with an increase in temperature.
Comparable paleoclimatic information from Peru and Bolivia seem to support our data. Kessler (1984, 1985) estimated 30% higher precipitation coupled with 3°C lower temperatures between 12,500 and 11,000 yr B.P. for the Bolivian Altiplano, based on dated high lake levels of paleolake Tauca. On the other hand, Servant and Fontes (1978) interpreted the late glacial higher levels of lakes Titicaca and Minchin as due to higher glacial ablation and not higher precipitation. However, higher lake levels appear to coincide and not follow
268 VOL. 6, NO. 3

... LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
glacial advances in the Andes, suggesting to Hastenrath and Kutzbach (1985) 50% higher precipitation instead (of about 200 mm compared to today). Independent of the ultimate cause, the evidence suggests generally wetter and cooler conditions prior to 10,000 yr B.P., similar to the information from Barro Negro.
Subsequent desiccation of these lakes and rapid glacier shrinkage in the Peruvian Andes between 11,000 and 10,000 yr B.P. (Mercer, 1984) indicates increased temperatures and higher evaporation towards the end of the late Pleistocene, similar to the proposed interpretation of the Barro Negro record.
Whether this paleoenvironmental change alone was sufficiently severe enough to cause the disappearance of Hippidion is impossible to assess. There is no question that the shrinkage of the Altoandean grasslands presented a much larger problem to the horse, an obligatory grazer, than to the camelids, that are facultative grazers. It is interesting to note that at other sites in the Altiplano where an extinct fauna including sloth and deer has been found (MacNeish et al.,1981; Wheeler Pires-Ferreira et al., 1976), camelids are absent or quite rare until after the extinction event, just as at the Barro Negro site. Man was present in the Altiplano, although perhaps neither locally nor in great numbers, prior to the extinction. This led Lynch (1986, 1990) to suggest that human occupation of the Altiplano was perhaps lower because of the cooler and moister conditions. If on the other hand man had a preference for hunting camelids, his absence from the Altiplano and the Barro Negro site could be explained by the camelids' absence. As it stands, however, a combination of climatic change and human predation cannot be ruled out to explain the extinction of Hippidion in the Altiplano of northern Argentina, although the paleoenvironmental cause seems to present a more likely explanation.
This work is contribution No. 97 ofiNGEIS (lnstituto de Geocronologia y Geologia Isot6pica, Buenos Aires), and was supported under the International Science Cooperative Program between the National Science Foundation (grant ATM-8212836 to V. Markgraf) and CONICET (grant 065-A to J. Fernandez). Thanks are due to Dr. E. Linares, Director ofiNGEIS. Comments from Dr. F. Alonso (Radiocarbon Laboratory, Instituto Rocasolano, Madrid, Spain) on an earlier version of this manuscript, and from Drs. P. Fritz, J. White, T. Stafford, and T. F. Lynch on a later version are greatly appreciated. Gratitude is expressed for field work assistance from Compania Minera Aguilar, S.A. during the last four years, especially to engineers M. J. Rossetti, R. Panighi, T. Zurita, and Dr. J. Lanfranco. Lie. M. Do Campo provided x-ray diffraction analysis. Prof. M. Lopez identified the botanical specimens that are kept in the Museo Argentino de Ciencias Naturales, Buenos Aires.
REFERENCES
Alberdi, M.T., Fernandez, J., Menegaz, A.N., and Prado, J.L. (1986). Hippidion Owen 1869 (Mammalia, Perissodactyla) en sedimentos del Pleistoceno tardio de la localidad de Barro Negro (Jujuy, Argentina). Estudios Geologicos 42, 487-493.
Albero, M.C., and Angiolini, F.E. (1983). INGEIS radiocarbon laboratory dates. Radiocarbon 25, 831- 842.
Albero, M.C., and Angiolini, F.E. (1985). INGEIS radiocarbon laboratory dates Il. Radiocarbon 27, 314-337.
Ambrose, S.H., and DeNiro, M.J. (1989). Climate and habitat reconstruction using stable carbon
GEOARCHAEOLOGY:ANINTERNATIONALJOURNAL 269

LATE PLEISTOCENE/EARLY HOLOCENE, ARGENTINE ALTIPLANO
and nitrogen isotope ratios of collagen in prehistoric herbivore teeth from Kenya. Quaremary Research 31, 407-422.
Bender, M.M. (1968). Mass spectrometer studies of carbon-13 variations in corn and other grasses. Radiocarbon 10, 468-472.
Bender, M.M. (1971). Variations in the 13C/12C ratios of plants in relat ion to the pathway of photosynthetic carbon dioxide fixation. Phyrochemistry 10, 1239- 1244.
Bird, J .B. (1988). Travels and Archaeology in South Chile. Iowa City: University of Iowa Press, 246 pp.
Bjorkman, 0 ., and Berry, J. (1973). High-e fficiency photosynthesis. Scientific American 229,80-93. Borrero, L.A. (1977). La extinci6n de la megafauna: su explicaci6n por factores concurentes. La
situaci6n en Patagonia austral. Anales Instituto de la Patagonia 8, 81- 93. Borrero, L.A. (1981). El poblamiento de la Patagonia Austral: revalorizaci6n de Ios cazadores de
Mylodon. Aetas X . Congreso Inrernacional Ciencias Prehisrorioos y Protohisroricas, 90-93. Cardicb, A (1985). Paleoambientes y lamAs antigua presencia del hombre, in J.R. Barcena, Ed.,
Culturas Indigenas rk la Pala8oni4. Las Culturos de America en la Epoca del Descubrimiento, pp. 15- 36. Madrid: Biblioteca del V Centenario.
Craig, H. (1957). Isotopic standards for carbon and oxygen and correction factors for maas-spectrometric analysis of carbon dioxide. Geochimica Cosmochimica Acta 12, 133-149.
DeNiro, M.J ., and Epstein, S. (1978). Influence of diet on the distribution of carbon isotopes in animals. Geochimica C01mochimica A cta 42, 495-506.
DeNiro, M.J., and Epstein, S. (1981). Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica Cosmochimica Acta 45, 341- 351.
Dillehay, T.D. (1989). Monte Verde. A late Pleistocene settlement in Chile. Washington: Smithsonian Institution Press.
Eicher, U., and Siegenthaler, U. (1976). Palynological and oxygen isotope investigations on late Glacial sediment cores from Swiss lakes. Boreas IS, 109-117.
Eicher, U., and Siegenthaler, U. (1983). Paleoenvironnement tardiglaciaire d.es lacs de Pelleautier et Siguret (Hautes-Alpes, France) IV. Stable isotopes of oxygen and carbon in the carbonate sediments of lacs de Siguret (Hautes Alpes, France). Ecologia Mediterranea 9, 49- 53.
Eicher, U., Siegenthaler , U., and Wegmiiller, S. (1981). Pollen and oxygen isotope analysis on Late and Post-Glacial sediments of the Tourbiere de Chirens <Dauphine, France). Quaternary Research 15. 160- 170.
Faegri, K., and lvereen, J . (1975). Textbook ofPoUen Analysis. Kopenhagen: Munksgaard, 295 pp. Femlindez, J ., and Panarello, H.E. (1988). II!Otopos en Arqueologia, I: Valores isot6picos en aguas
mete6ricas y su paseje a la sangre humana y a la de camelidos de la Puna Argentina, a 4,000 de altitud. m.s., 13 pp.
Fontes, J .C., Boulanger, B., and Rodrigo, L.A. (1981). Carbone-14, carbone-13 and oxyglme-18 dans les sediments carbonnates du lac Titicaca. Premieres estimations des vitesses de sedimentation et essay de paleoclimatologie. Comptes Rendues Academie Science. Paris, ser. 2, 293, 53-56.
Fontes, J.C., Coque, R. , Dever, L., Filly, L., and Mamou, A. (1983). Paleohydrologie isotopique de l'Qued E l Akarit (Sug Tunisie) au Pleistocene superieur et a l'Holocene. Palaeogeography, Palaeoclimarology, Palaeoecology 43, 41- 62.
Fritz, P ., Anderson, 'f.W., and Lewis, C.F.M. (1975). Late Quaternary climatic trends and history of Lake Erie from stable isotope studies. Science 190, 267-269.
Fritz, P., and Poplawsky, S. (1974). 180 and 13C in the shells of freshwater molluscs and their environments. Earth and Planetary Science Letters 24, 91-98.
Graf, K. (1979). Untersuchungen zur rezenten Pollen- und Sporenflora in der nordlichen Zentralkordillere Boliviens und Versuch einer Auswertung von Profilen aus postglazialen Torfmooren, PhD Dissertation, University of Zurich, 104 pp.
Graham, R. W. (1979). Palaeoclimates and the Late Pleistocene Fauna! Provinces in North America, in R.L. Humphrey and D. Stanford, Eds., Pre-Llano Cultures of the Americas: Para~s and Possibilities, pp. 49-69. Washington: The Anthropological Societ¥·
270 VOL. 6, NO. 3

LATE PLEISTOCENEJEARLY HOLOCENE, ARGENTINE ALTIPLANO
Guilday, J.E. (1967). Differential Extinction during Late Pleistocene and Recent Times, in P.S. Martin and H.E. Wright, Eds., Pleistocene Extinctions: The Search for a Cause, pp. 121- 140. New Haven: Yale University Press.
Guthrie, R.D. (1984). Mosaics, Allelochemics and Nutrients: an Ecological Theory of Late Pleistocene megafaunal extinctions, in P .S. Martin and R.G. Klein, Eds., Quaternary Extinctions, A Prehistoric Revolution, pp. 259- 298. Tucson: University of Arizona Press.
Halloy, S. (1983). Contribucwn al estudio de la zona de Huaca Huasi, Cumbres de Calchaquies (Tucuman, Argentina), PhD Dissertation , University ofTucuman, Argentina, 839 pp.
Hansen, B.C.S., Wright, H.E., and Bradbury, J .P. (1984). Pollen studies in the Junin area, central Peruvian Andes. Geological Society of America, Bulletin 95, 1454-1465.
Hastenrath, S., and Kutzbach, J. (1985). Late Pleistocene climate and water budget of the South American Altiplano. Quaternary Research 24, 249- 256.
Hattersley, P.W. (1983). The distribution of Cs and C4 grasses in Australia in relation to climate. Oecologia 57, 113- 128.
KeSBler, A. (1984). The Paleohydrology of the Late Pleistocene Lake Tauca on the Southern Altiplano (Bolivia) and Recent Climatic Fluctuations, in J .C. Vogel, Ed., Late Cainozoic Palc.eoclimates of the Southern Hemisphere, pp. 115-122. Rotterdam: Balkema.
Keesler, A. (1985). Zur Rekonstruktion von spAtglazialem Klima und Wasserhaushalt auf dem peruanisch-bolivianischen Altiplano. Zeitschrift fur Gletscherkunde und Glazialgeologie 21, 107-114.
Krueger, H.W., and Sullivan, C.H. (1984). Models for Carbon Isotope Fractionation between Diet and Bone, in J.E. Turnlund and P.E. J ohnson, Eds ., Stable Isotopes in Nutrition, pp. 205-222. American Chemical Society, Symposium Series 258.
Longin, R. (1971). New method of collagen extraction for radiocarbon dating. Nature 230, 241- 242. Lynch, T.F. (1983). The Paleo-lndians, inJ .D. Jennings, Ed.,Ancient South Americans, pp. 87-137.
San Francisco: W.W. Freeman. Lynch, T.F. (1986). Climate change and human settlement around the late-glacial Laguna de · Punta Negra, northern Chile: the preliminary results. Geoarchaeology 1, 145-162. Lynch, T.F. (1990). Quaternary climate, environment, and the human occupation of the South
Central Andes. Geoarchaeology 5, 199- 228. MacNeish, R.S., Cook, A.G., Lumbreras, L.G., Vierra, R.K., and Nelken-Terner, A. (1981). Prehis
tory of the Ayacucho Basin, Peru. ll. Ann Arbor, University of Michigan Press, 104 pp. Mark.graf, V. (1985a). Late Pleistoce.ne fauna! extinctions in Southern Patagonia. Science 228,
1110- 1112. Markgraf, V. (1985b). Paleoenvironmental history of the last 10,000 years of northwestern Argen
tina. Zentrolbkztt filr Geologie und Palii.ontologie 11112, 1739-1749. Markgraf, V. (1989). Palaeoclimates in Central and South America since 18,000 BP based on pollen
a.nd lake-level records. Quaternary Science Reviews 8, 1-24. Marshall, L.G., Berta, A., Hoffstetter , R., Pascual, R. , Reig, O.A., Bombin, M., and Mones, A. (1984).
Mammals and stratigraphy: geochronology of the continental mammal-bearing Quaternary of South America. Memoirs Palcuovertebrata, pp. 1- 76. Mont9ellier, France.
Martin, P.S. (1973). The discovery of America. Science 179, 969- 974. Martin , P .S. (1984). Prehistoric Overkill: the Global Model, in P.S. Marwn and R.G. Klein, Eds.,
Quaternary E:~tinctions: A Prehistoric R evolution, pp. 354-403, Tucson: University of Arizona PreSB.
McCrea, J .M. (1950). The isotopic chemistry of carbonates and a temperature scale. Journal Chemical Physics 18, 849- 857.
Mead, J .L, and Meltzer, D.J. (1984). North American Late Quaternary Extinctions and the Radiocarbon Record, in P.S. Martin and R.G. Klein, Eds., Quaternary Extinctions: A Prehistoric Revolution, pp. 440-450. Tucson: University of Arizona Press.
Mercer, J .H. (1984). Late Cainozoic Glacial Variations in South America South of the Equator, in J.C. Vogel, Ed., Late Cainozoic Palaeoclimates of the Southern Hem.isphere, pp. 45-58. Rotter· dam: Balkema.
GEOARCHAEOLOGY: AN INTERNATIONAL JOURNAL 271

LATE PLEISTOCENE/EARL Y HOLOCENE, ARGENTINE ALTIPLANO
O'Neill, J.R., Clayton, R.N., and Mayeda, T.K. (1969). Oxygen isotope fractionation in divalent metal carbonates. JouriUJ.l Chemical Physics 51, 5547-5558.
Panarello, H.O. (1987). Relaci.ones entre concentraci.ones de is6topos liviancs utili.zados como indi.cadores ambientales y de pal~otemperaturas. PhD Dissertation, University of Buenos Aires, Ar· gentina.
Pascual, R., Espinosa, E.J. Ortega, Gondar, D., and Tonni, E. (1966). Paleontogratla Bonaerense IV. Vertebrate. Comisi.on de Investigaci.ones Cienti{u:as de Buencs Aires, pp. 1-202.
Rick, J . W. (1988). The Character and Context of Highland Preceramic Society, in R. W. Keatinge, Ed., Prehistoric Hunters, pp. 3-40, Cambridge, England: Cambridge University Press.
Ruthsatz, B., and Movia, C.P. (1975). Relevamiento de las estepas andinas del n.oroeste de la Provincia de Jujuy, pp. 1- 127, Buenos Aires: Fundaci6n para la Educacion, Ciencia y Cultura.
Schoeninger, M.J., and DeNiro, M.J . (1984). Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochimica Cosmochimica Acta 48, 625-639.
Servant, M., and Fontes, J.C. (1978). Les lacs quaternaires des hauts plateaux des Andes boliviennes. Premiilres interpretations paleoclimatiques. Cahier Orstcm, s~r. Geologie 10, 9-23.
Smith, B.N., and Epstein, S. (1971). Two categories of 13C/12C ratios for higher plants. Plant Physiology 47, 380-384.
Stanford, D., Wedel, W.R., and Scott, G.R. (1981). Archaeological investigations of the Lamb Spring Site. Southwestern Lore 47, 14-27.
Stafford, T.W., Jull, A.J.T., Brendel, K., Duhamel, R.C., and Donahue, D. (1987). Study of bone radiocarbon dating accuracy at the University of Arizona NSF accelerator facility for radioisotope analysis. Radiocarbon 29, 24-44.
Stafford, T.W., Brendel, K., and Duhamel, R.C. (1988). Radiocarbon, 13C and 15N analysis of fossil bone: removal of humates with XAD-2 resin. Geochimica Cosmochimica Acta 52, 2257-2267.
Stuiver, M. (1970). Oxygen and carbon isotope ratios of fresh water carbonates as climatic indicators. JouriUJ.l Geophysical Research 5, 5247-5257.
Taylor, R.E. (1982). Problems in the Radiocarbon Dating of Bone, in L.A. Currie, Ed., Nuclear and Chemical Dating Techniques: Interpreting the EnvironmenU1l Record, pp. 453-478, American Chemical Society.
Thompson, L.G., Mosley-Thompson, E., Bolzan, J.F., and Koci, B.R. (1985). A 1500-year record of tropical precipitation in ice cores from the Quelccaya ice cap, Peru. Science 229, 971-973.
Tieszen, L.L., Senyimba, M.M., lmbamba, S.K., and Troughton, J .H . (1979). The distribution ofC3 and C4 grasses and carbon isotope discrimination along an altitudinal and moisture gradient in Kenya. Oecologia 37, 337-350.
Van der Merwe, N.J . (1982). Carbon isotopes, photosynthesis, and archaeology. American Scientist 70, 596-606.
Vogel, J.C. (1978). Isotopic assessment of the dietary habits of ungulates. South African JournaL of Science 74, 298-301.
Vogel, J .C., and Van der Merwe, N.J . (1977). Isotopic evidence for early maize cultivation in New York State. American Antiquity 42, 238-242.
Vogel, J.C., Fuls, A., and Ellis, R.P. (1978). The geographical distribution of Kranz grasses in South Africa. South African Journal of Science 74, 209-215.
Wheeler Pires-Ferreira, J. Pires-Ferreira, E., and Kaulicke, P. (1976). Preceramic animal utilization in the Central Peruvian Andes. Science 194, 483-489.
Received October 13, 1990 Accepted for publication February 18, 1991
272 VOL. 6, NO. 3

![[Why are there few publications by the Argentine gastroenterology? Considerations on a bibliometric analysis of Argentine publications on gastroenterology]](https://static.fdokumen.com/doc/165x107/634608546cfb3d406409f4f4/why-are-there-few-publications-by-the-argentine-gastroenterology-considerations.jpg)


















