
Pinus thunbergii bark extract rich in flavonoids promotes hair ...
Upload
independentCategory
view
1download
0
Biological Control 54 (2010) 206–212
Contents lists available at ScienceDirect
Biological Control
journal homepage: www.elsevier .com/ locate/ybcon
Is there intraguild predation between winter-active spiders (Araneae) on appletree bark?
S. Korenko, S. Pekár *
Department of Botany and Zoology, Faculty of Science, Masaryk University, Kotlárská 2, 611 37 Brno, Czech Republic
a r t i c l e i n f o a b s t r a c t
Article history:Received 7 January 2010Accepted 12 May 2010Available online 11 June 2010
Keywords:IGPBiological controlApple orchardAnyphaenaPhilodromus
1049-9644/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.biocontrol.2010.05.008
* Corresponding author. Tel.: +420 549496464.E-mail address: [email protected] (S. Pekár).
We studied whether there si intraguild predation among overwintering invertebrates in bark trapsinstalled on apple trees. We found that 86% of the overwintering invertebrates in traps were predators,which were dominated by spiders (72%) of different guilds and different body sizes. Spiders had spatiallysegregated in the traps so that smaller specimens hid in an interstitial space that was inaccessible to lar-ger specimens. Spiders of the bark-dwelling guild, Anyphaena and Philodromus, are winter-active and canprey on other smaller spiders. Analysis of their abundance in traps indicated a negative relationshipbetween Anyphaena and Philodromus or other spiders. Inspection of traps during winter revealed an influxof spider specimens confirming their winter-activity. In a semi-field experiment, where overwintering incardboard bands was studied under controlled conditions, we observed mortality of both winter-activePhilodromus and Anyphaena spiders due to their interspecific predation. The mortality was affected bytheir size ratio – larger specimens consumed smaller ones. Presence of alternative prey, theridiid spiders,did not affect their mortalities, but the presence of shelters (interstitial spaces) did. In boards with inter-stitial spaces smaller spiders suffered lower mortality than in boards without the interstitial spaces.Obtained results suggest that intraguild predation between overwintering spiders can decrease theirpopulation density during winter.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
In central Europe, in lowland and submontane areas of theCzech Republic, the ambient conditions are unfavorable (i.e., aver-age temperatures <5 �C) from late autumn to early spring for thepopulation development of invertebrates. Many pests and theirnatural enemies pass such an unfavorable period in a state of dor-mancy (e.g., Schaefer, 1977). They hide in refuges, found in the soilor in leaf litter, which are good insulators against the cold (Foelix,1996). In fruit orchards many arboreal pests and their predatorsoverwinter in the crevices of bark (Bower and Snetsinger, 1985).However, few species of spiders remain active during winter. Thesespecies not only move but catch prey at temperatures close to zeroor even at temperatures below zero (Aitchison, 1984, 1987; Kore-nko et al., 2010). Yet, it is not known what prey they catch, pests orother predators?
Spiders are a major component of the generalist predator guildin various ecosystems (Moulder and Reichle, 1972), and one of themost important natural enemies of arthropod pest species in com-mercial fruit orchards (Olszak et al., 1992; Riechert and Lockley,1984; Nyffeler and Benz, 1987; Markó et al., 2009). A few win-
ll rights reserved.
ter-active spider species were found to catch important pests offruits, namely leafrollers (Bogya, 1999) and psyllids (Horton,2004). Winter-active spiders could also have high bio-control po-tential during early spring, when some pests, such as psyllids, arealready active but their specific predators are still inactive (Bogya,1999). Therefore, understanding the ecology of overwinteringarthropod predators and determining the mechanisms that influ-ence their bio-control efficiency may provide new insight intothe regulation of pests.
However, winter-active predators can also perform intraguildpredation (IGP), i.e., consumption of a guild member, therebyreducing the density of natural enemies (e.g., Polis et al., 1989).This is often the case with polyphagous spiders (Dinter, 1998;Guillebeau and All, 1989; Rosenheim et al., 1995) that can preyupon each other (e.g., Polis et al., 1989; Gunnarsson, 1985) or onother natural enemies, such as beetles, lacewings (Dinter, 1998,2002), lygaeid bugs (Guillebeau and All, 1989) and other arachnids(e.g., Polis and McCormick, 1986). IGP thus disrupts bio-control(Wise, 1993; Polis and McCormick, 1986; Polis et al., 1989; Rosen-heim et al., 1995) and can even entail predator species exclusion(Holt and Polis, 1997; McCann et al., 1998).
Here, we studied intraguild interactions between the mostabundant overwintering arthropod predators on apple trees. Previ-ously, we found a negative relationship between the densities of

S. Korenko, S. Pekár / Biological Control 54 (2010) 206–212 207
large and small spiders overwintering on tree bark (Pekár, 1999a)and suspected intraguild predation of large spiders on small ones.The occurrence of intraguild predation in the field is often con-cluded indirectly from the relationship between the density of pre-dators (Wise and Chen, 1999; Meyhöfer and Hindayana, 2000). Tounravel IGP it is necessary to find that predators (1) co-occur dur-ing overwintering at the same microhabitat; (2) are active duringwinter; and (3) capture each other. Therefore, we installed trapson bark to obtain evidence for co-occurrence in the study habitatand for sharing the same microhabitat; we inspected the traps sev-eral times during winter to see if there is an influx of winter-activepredators; and we performed semi-field experiments to test howthe presence of a shelter and alternative prey influences the fre-quency of intraguild predation.
In these experiments, we focused on the role of two dominantspider predators, Anyphaena accentuata Walckenaer (Anyphaeni-dae) and species of the genus Philodromus (Philodromidae). Thesespecies are common in apple trees in central Europe (Bogya andMols, 1996). In summer A. accentuata is a nocturnal wanderer,while Philodromus is a diurnal wanderer (Marc, 1991; Marc andCanard, 1997), thus there is limited overlap in their niche. Butduring winter both species are confined to limited spaces on thetree trunks (Pekár, 1999a; Miliczky et al., 2008) so ample opportu-nities for interactions are expected. Recently, we estimated thatthese spiders should be actively hunting for a few winter months(Korenko et al., 2010). As both species are large and adapted tohunt at low temperatures, we expect these species to feed notonly on insects but also on other rather smaller spiders duringwinter, which may result in a decrease of the overall spider pop-ulation density.
2. Materials and methods
The field study was carried out in a commercial apple orchardunder integrated pest management in Brno (49�0903800N, 16�330
3800E) during the winters of 2006/07 and 2007/08. A variety of barktraps have been designed but corrugated cardboard trap bandswere found to be most efficient for arachnids (Mizell and Schiff-hauer, 1987). The traps (13 corrugations per 10 cm) were installedon tree trunks (Golden Delicious) in October 2006 and 2007. Thebands were 30 cm wide strips with a length equal to the trunkperimeter (mean = 70.5 cm). The bands were fixed on tree trunkswith wire at a height of 5–15 cm from the ground.
The outdoor temperature was measured continuously in theorchard throughout the winter with a digital thermometer (DallasSemiconductor DS18B20) installed 2 m above the ground. The
Fig. 1. (A) Cross-section of the trunk and the corrugated band showing the inner ancorrugations. The boards were 5 mm apart. (C) Cross-section of a board without shelter
average temperature between November and January in winter2006/07 was 4.24 �C (26 days with maximum day temperatureabove 10 �C) and in winter 2007/2008 it was 1.45 �C (only 9 dayswith maximum day temperature above 10 �C).
The survey of the overwintering community was based on thebands (N = 20) installed in October and exposed until the end ofJanuary in both winters. Each band provided two types of shelter:an outer and an inner layer (Fig. 1A). The outer layer included thespace between the bark and the cardboard band. The inner layerwas the interstitial space within corrugations. The diameter ofthe interstitial space was about 2.5 mm. When removing the bandsspiders were collected separately from the outer and inner layersand the bark below bands. Spiders were categorized visually intotwo groups by body size: small (total body length <2 mm) andlarge (total body length >2 mm). The spiders were identified usingHeimer and Nentwig (1991) and Roberts (1995). Philodromusspiders overwintered as juveniles and could not be identified tospecies. However, during the season we found two species in theorchard, P. cespitum (Walckenaer) and P. dispar Walckenaer.Because the former species was dominant we consider specimensused in experiments (see below) to have been P. cespitum.
The temporal change of abundance and composition of the spi-der overwintering community was studied by exposing cardboardtraps for different periods in the same apple orchard in winter2006/2007. Twenty bands were installed at the end of October2006 and exposed for 120 days. Sixty other bands were installedin the beginning of February 2007 and were exposed for 1, 5, and25 days (20 bands for each period). The average temperature forthese periods was 5.44 �C for the 1-day period, 6.8 �C for the 5-day period, 3.94 �C for the 25-day period, and 4.24 �C for the120 day period.
We measured the size (prosoma length) of three principal spi-der taxa found overwintering in the traps with an Olympus SZ 40stereomicroscope equipped with an ocular micrometer.
A semi-field experiment was performed in the winter of 2007/2008. The effects of three variables on the outcome of intraguildinteractions between the two principal winter-active spiders, A.accentuata and Philodromus sp., was studied. The variables wereas follows: (1) presence or absence of alternative prey, (2) presenceor absence of shelters, and (3) Anyphaena/Philodromus size ratio(0.7–3.2). Spiders were sorted by their prosoma size (range 1.1–2.5 mm for Anyphaena and 0.6–2.5 mm for Philodromus) in orderto reach homogeneous size classes with SE < 0.02 (see Table 1 forthe ratios used). Spiders of a similar size within each species werereleased to boards (described below). This allowed us to excludeintraspecific predation because these spider species were found
d outer layers. (B) Cross-section of a board with shelters indicating space insides.
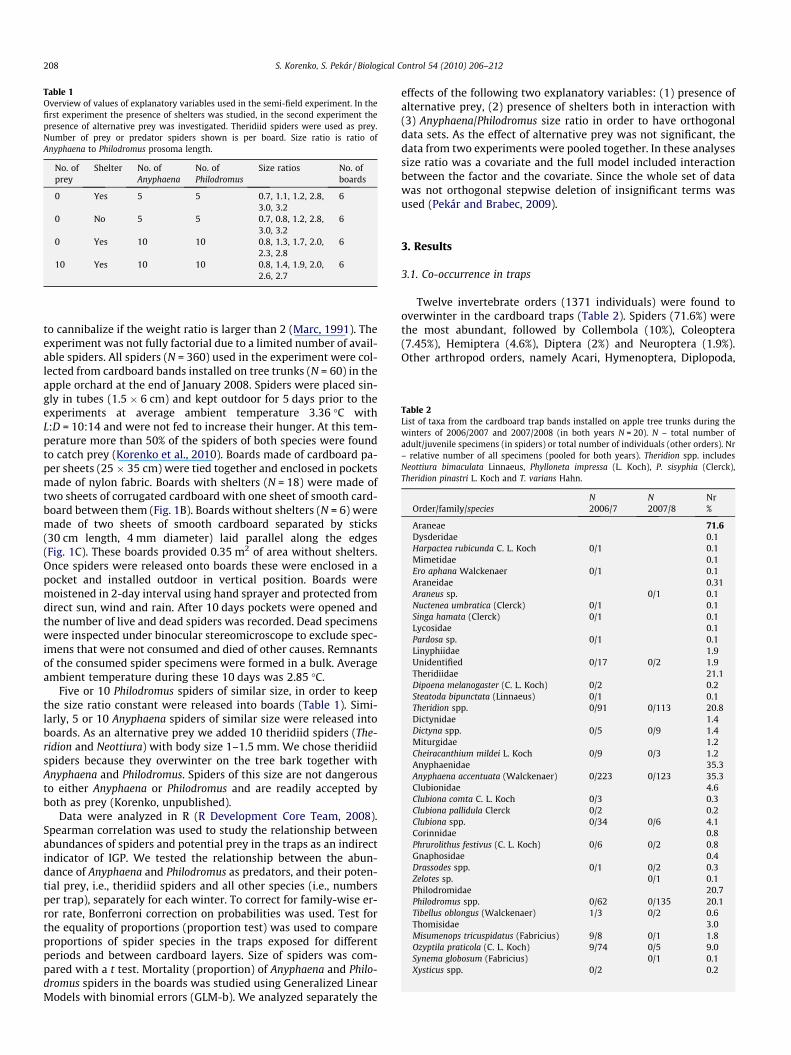
Table 1Overview of values of explanatory variables used in the semi-field experiment. In thefirst experiment the presence of shelters was studied, in the second experiment thepresence of alternative prey was investigated. Theridiid spiders were used as prey.Number of prey or predator spiders shown is per board. Size ratio is ratio ofAnyphaena to Philodromus prosoma length.
No. ofprey
Shelter No. ofAnyphaena
No. ofPhilodromus
Size ratios No. ofboards
0 Yes 5 5 0.7, 1.1, 1.2, 2.8,3.0, 3.2
6
0 No 5 5 0.7, 0.8, 1.2, 2.8,3.0, 3.2
6
0 Yes 10 10 0.8, 1.3, 1.7, 2.0,2.3, 2.8
6
10 Yes 10 10 0.8, 1.4, 1.9, 2.0,2.6, 2.7
6
Table 2List of taxa from the cardboard trap bands installed on apple tree trunks during thewinters of 2006/2007 and 2007/2008 (in both years N = 20). N – total number ofadult/juvenile specimens (in spiders) or total number of individuals (other orders). Nr– relative number of all specimens (pooled for both years). Theridion spp. includesNeottiura bimaculata Linnaeus, Phylloneta impressa (L. Koch), P. sisyphia (Clerck),Theridion pinastri L. Koch and T. varians Hahn.
N N NrOrder/family/species 2006/7 2007/8 %
Araneae 71.6Dysderidae 0.1Harpactea rubicunda C. L. Koch 0/1 0.1Mimetidae 0.1Ero aphana Walckenaer 0/1 0.1Araneidae 0.31Araneus sp. 0/1 0.1Nuctenea umbratica (Clerck) 0/1 0.1Singa hamata (Clerck) 0/1 0.1Lycosidae 0.1Pardosa sp. 0/1 0.1Linyphiidae 1.9Unidentified 0/17 0/2 1.9Theridiidae 21.1Dipoena melanogaster (C. L. Koch) 0/2 0.2Steatoda bipunctata (Linnaeus) 0/1 0.1Theridion spp. 0/91 0/113 20.8Dictynidae 1.4Dictyna spp. 0/5 0/9 1.4Miturgidae 1.2Cheiracanthium mildei L. Koch 0/9 0/3 1.2Anyphaenidae 35.3Anyphaena accentuata (Walckenaer) 0/223 0/123 35.3Clubionidae 4.6Clubiona comta C. L. Koch 0/3 0.3Clubiona pallidula Clerck 0/2 0.2Clubiona spp. 0/34 0/6 4.1Corinnidae 0.8Phrurolithus festivus (C. L. Koch) 0/6 0/2 0.8Gnaphosidae 0.4Drassodes spp. 0/1 0/2 0.3Zelotes sp. 0/1 0.1Philodromidae 20.7Philodromus spp. 0/62 0/135 20.1Tibellus oblongus (Walckenaer) 1/3 0/2 0.6Thomisidae 3.0Misumenops tricuspidatus (Fabricius) 9/8 0/1 1.8Ozyptila praticola (C. L. Koch) 9/74 0/5 9.0Synema globosum (Fabricius) 0/1 0.1Xysticus spp. 0/2 0.2
208 S. Korenko, S. Pekár / Biological Control 54 (2010) 206–212
to cannibalize if the weight ratio is larger than 2 (Marc, 1991). Theexperiment was not fully factorial due to a limited number of avail-able spiders. All spiders (N = 360) used in the experiment were col-lected from cardboard bands installed on tree trunks (N = 60) in theapple orchard at the end of January 2008. Spiders were placed sin-gly in tubes (1.5 � 6 cm) and kept outdoor for 5 days prior to theexperiments at average ambient temperature 3.36 �C withL:D = 10:14 and were not fed to increase their hunger. At this tem-perature more than 50% of the spiders of both species were foundto catch prey (Korenko et al., 2010). Boards made of cardboard pa-per sheets (25 � 35 cm) were tied together and enclosed in pocketsmade of nylon fabric. Boards with shelters (N = 18) were made oftwo sheets of corrugated cardboard with one sheet of smooth card-board between them (Fig. 1B). Boards without shelters (N = 6) weremade of two sheets of smooth cardboard separated by sticks(30 cm length, 4 mm diameter) laid parallel along the edges(Fig. 1C). These boards provided 0.35 m2 of area without shelters.Once spiders were released onto boards these were enclosed in apocket and installed outdoor in vertical position. Boards weremoistened in 2-day interval using hand sprayer and protected fromdirect sun, wind and rain. After 10 days pockets were opened andthe number of live and dead spiders was recorded. Dead specimenswere inspected under binocular stereomicroscope to exclude spec-imens that were not consumed and died of other causes. Remnantsof the consumed spider specimens were formed in a bulk. Averageambient temperature during these 10 days was 2.85 �C.
Five or 10 Philodromus spiders of similar size, in order to keepthe size ratio constant were released into boards (Table 1). Simi-larly, 5 or 10 Anyphaena spiders of similar size were released intoboards. As an alternative prey we added 10 theridiid spiders (The-ridion and Neottiura) with body size 1–1.5 mm. We chose theridiidspiders because they overwinter on the tree bark together withAnyphaena and Philodromus. Spiders of this size are not dangerousto either Anyphaena or Philodromus and are readily accepted byboth as prey (Korenko, unpublished).
Data were analyzed in R (R Development Core Team, 2008).Spearman correlation was used to study the relationship betweenabundances of spiders and potential prey in the traps as an indirectindicator of IGP. We tested the relationship between the abun-dance of Anyphaena and Philodromus as predators, and their poten-tial prey, i.e., theridiid spiders and all other species (i.e., numbersper trap), separately for each winter. To correct for family-wise er-ror rate, Bonferroni correction on probabilities was used. Test forthe equality of proportions (proportion test) was used to compareproportions of spider species in the traps exposed for differentperiods and between cardboard layers. Size of spiders was com-pared with a t test. Mortality (proportion) of Anyphaena and Philo-dromus spiders in the boards was studied using Generalized LinearModels with binomial errors (GLM-b). We analyzed separately the
effects of the following two explanatory variables: (1) presence ofalternative prey, (2) presence of shelters both in interaction with(3) Anyphaena/Philodromus size ratio in order to have orthogonaldata sets. As the effect of alternative prey was not significant, thedata from two experiments were pooled together. In these analysessize ratio was a covariate and the full model included interactionbetween the factor and the covariate. Since the whole set of datawas not orthogonal stepwise deletion of insignificant terms wasused (Pekár and Brabec, 2009).
3. Results
3.1. Co-occurrence in traps
Twelve invertebrate orders (1371 individuals) were found tooverwinter in the cardboard traps (Table 2). Spiders (71.6%) werethe most abundant, followed by Collembola (10%), Coleoptera(7.45%), Hemiptera (4.6%), Diptera (2%) and Neuroptera (1.9%).Other arthropod orders, namely Acari, Hymenoptera, Diplopoda,

Table 2 (continued)
N N NrOrder/family/species 2006/7 2007/8 %
Salticidae 0.8Carrhotus xanthogramma (Latreille) 0/4 0.4Dendryphantes rudis (Sundevall) 0/1 0.1Neon reticulates (Blackwall) 0/1 0.1Salticus zebraneus (C. L. Koch) 2/0 0.2Acari 0.8Trombidiformes 8 3 0.8Crustacea 0.1Oniscoidea 1 0.1Diplopoda 0.2Glomerida 1 0.1Julida 2 0.1Collembola 68 69 10.0Dermaptera 0.1Forficula auricularia Linnaeus 1 0.1Hemiptera 4.6Auchenorrhyncha 9 4 0.9Aphididae 3 1 0.3Psyllidae 3 0.2Anthocoris sp. 5 13 1.3Orius spp. 6 6 0.9Unidentified 10 3 0.9Neuroptera 1.9Chrysoperla spp. 21 4 1.8Unidentified 1 0.1Coleoptera 7.5Coccinellidae 7 9 1.2Curculionidae 3 4 0.5Unidentified 15 6 1.5Larvae 37 21 4.2Hymenoptera 0.9Lasius sp. 4 5 0.7Unidentified 4 0.3Diptera 2.0Unidentified 21 6 2.0Lepidoptera 0.4Caterpillars 3 2 0.4
S. Korenko, S. Pekár / Biological Control 54 (2010) 206–212 209
Lepidoptera, Dermaptera and Oniscoidea, represented less than 1%of the overwintering community. Thus 85.8% of overwinteringindividuals were predators.
The great majority of spiders overwintered as juveniles (98%,N = 981). There were three dominant taxa: A. accentuata (35.3%),Philodromus spp. (20.7%) and Theridiidae (20.8%). All these speciesoverwintered as juveniles. Theridiid spiders had the smallest pro-
0
2
4
6
8
10
12
0 5 10 15 20 25
No. of Philodromus
No
. of
Any
ph
aen
a
A B
Fig. 2. Relationship between abundance of Anyphaena and Philodromus spiders (A) and otrespectively.
soma size (mean = 0.54 mm, SD = 0.07, N = 277). Philodromus pro-soma size (mean = 1.1 mm, SD = 0.4, N = 308) was significantlylarger than that in Theridiidae (t583 = 24.7, P < 0.0001). Anyphaenaoverwintered also at juvenile stage and had the largest prosomasize (mean = 2.2 mm, SD = 0.3, N = 211), significantly larger thanthat in Philodromus (t517 = 26.1, P < 0.0001).
Both Anyphaena and Philodromus could feed during winter oninsects. There was a slightly positive or no relationship betweenthe abundance of Anyphaena or Philodromus and Collembola, Cole-optera and Hemiptera (Spearman correlation, P > 0.1). As spiderswere clearly the dominant component, we expected that Anyphae-na could feed on both Philodromus and theridiid spiders and Philo-dromus could feed on theridiid spiders. No significant relationship,even at uncorrected probability level, between the number of spi-ders was found. But there was a negative trend between the num-ber of Anyphaena and the number of Philodromus in winter 2006/07(Spearman correlation, rho = �0.37, P = 0.11, Fig. 2A), and the num-ber of Anyphaena and the number of other spiders in winter 2007/08 (Spearman correlation, rho = �0.41, P = 0.07, Fig. 2B).
3.2. Microhabitat segregation
Although both Anyphaena and Philodromus were found to sharethe same microhabitat, we observed a difference in preference forthe two trap layers (Fig. 3). While the inner layer was dominatedby small Philodromus and theridiid spiders (87.3%), the outer layerwas dominated by large Anyphaena spiders (41.9%). Small spidersthus preferred the inner layer to the outer one (Proportion test,v2
1 = 108.5, P < 0.0001).
3.3. Activity during winter
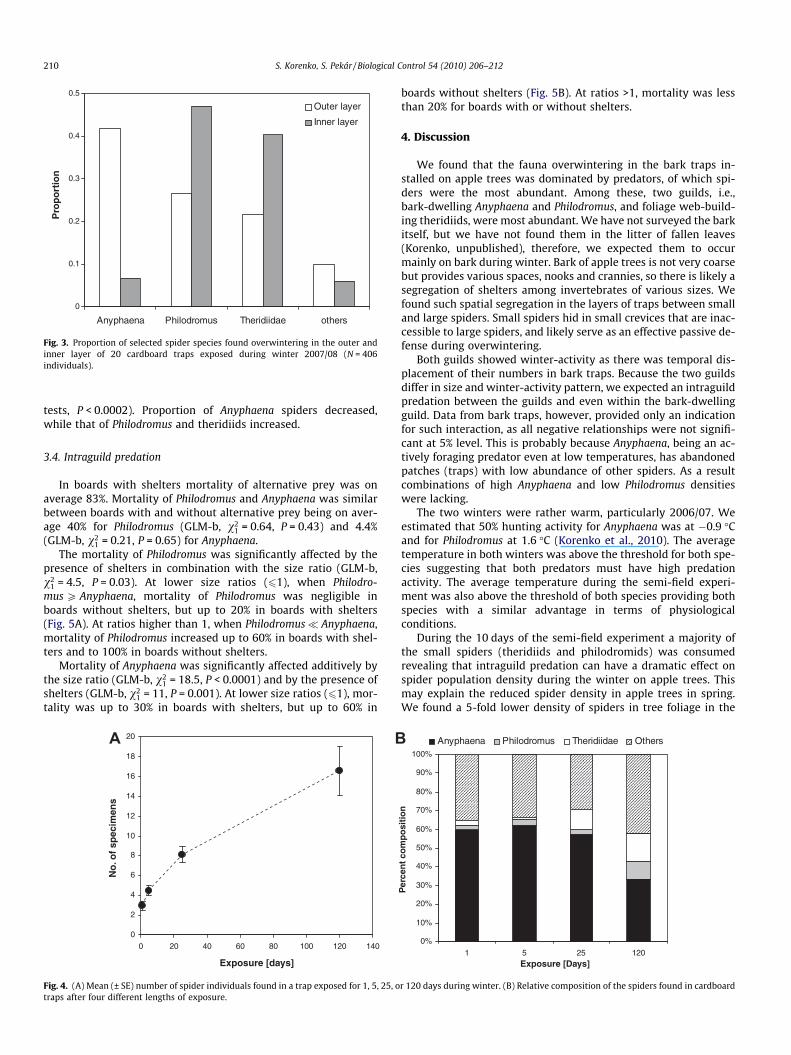
The exposure period of traps affected the density and composi-tion of the overwintering spider fauna. The number of overwinter-ing specimens increased linearly with the exposure period(Fig. 4A). After 1 day on average three specimens were found, whileafter 120 days 16.6 specimens were found in the bands, suggestingthat there is constant movement into bands with a rate of 0.14specimen per day. The composition of three major spider taxawas similar for 1 and 5 days of exposure (Proportion tests,P > 0.75, Fig. 4B). After 25 days only the proportion of Theridiidaeincreased significantly (Proportion test, v2
1 = 17.4, P < 0.0001).And after 120 days all groups changed their proportion (Proportion
0
1
2
3
4
5
6
0 2 4 6 8 10 12
No. of other spiders
No
. of
Any
ph
aen
a
her spiders (B) overwintering in 20 traps in the winter 2006/07 and winter 2007/08,
0
0.1
0.2
0.3
0.4
0.5
Anyphaena Philodromus Theridiidae others
Pro
po
rtio
n
Outer layer
Inner layer
Fig. 3. Proportion of selected spider species found overwintering in the outer andinner layer of 20 cardboard traps exposed during winter 2007/08 (N = 406individuals).
210 S. Korenko, S. Pekár / Biological Control 54 (2010) 206–212
tests, P < 0.0002). Proportion of Anyphaena spiders decreased,while that of Philodromus and theridiids increased.
3.4. Intraguild predation
In boards with shelters mortality of alternative prey was onaverage 83%. Mortality of Philodromus and Anyphaena was similarbetween boards with and without alternative prey being on aver-age 40% for Philodromus (GLM-b, v2
1 = 0.64, P = 0.43) and 4.4%(GLM-b, v2
1 = 0.21, P = 0.65) for Anyphaena.The mortality of Philodromus was significantly affected by the
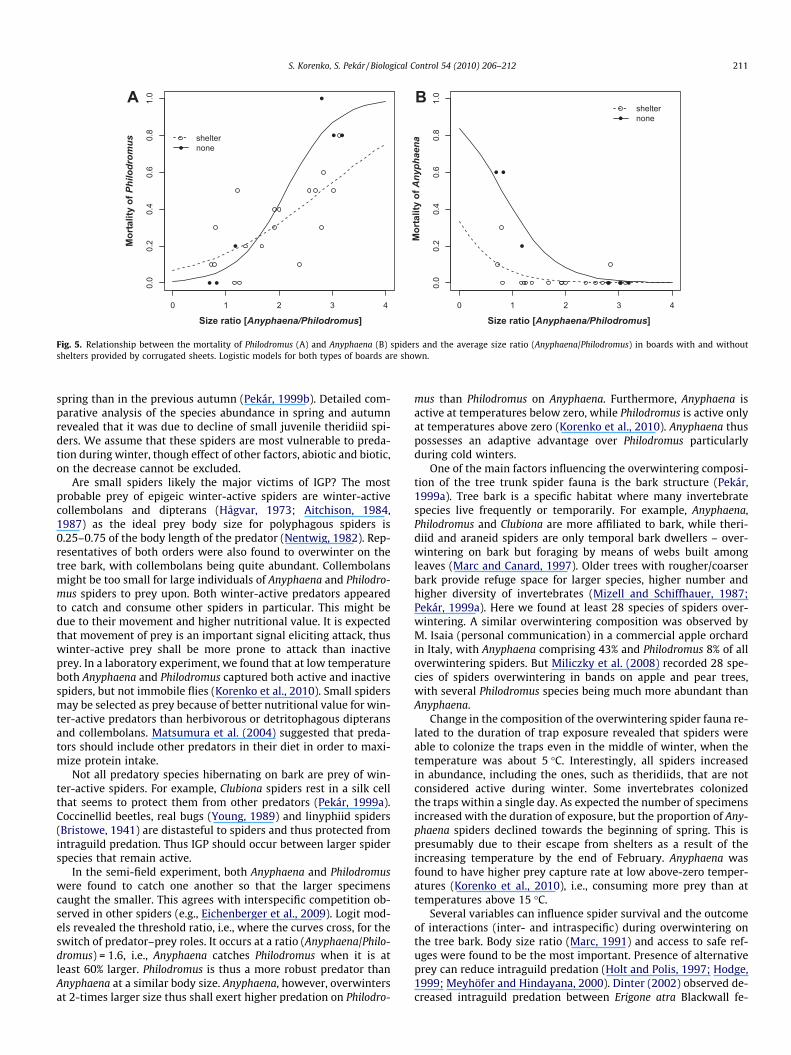
presence of shelters in combination with the size ratio (GLM-b,v2
1 = 4.5, P = 0.03). At lower size ratios (61), when Philodro-mus P Anyphaena, mortality of Philodromus was negligible inboards without shelters, but up to 20% in boards with shelters(Fig. 5A). At ratios higher than 1, when Philodromus� Anyphaena,mortality of Philodromus increased up to 60% in boards with shel-ters and to 100% in boards without shelters.
Mortality of Anyphaena was significantly affected additively bythe size ratio (GLM-b, v2
1 = 18.5, P < 0.0001) and by the presence ofshelters (GLM-b, v2
1 = 11, P = 0.001). At lower size ratios (61), mor-tality was up to 30% in boards with shelters, but up to 60% in
0
2
4
6
8
10
12
14
16
18
20
0 20 40 60 80 100 120 140
Exposure [days]
No
. of
spec
imen
s
A B
Fig. 4. (A) Mean (± SE) number of spider individuals found in a trap exposed for 1, 5, 25, otraps after four different lengths of exposure.
boards without shelters (Fig. 5B). At ratios >1, mortality was lessthan 20% for boards with or without shelters.
4. Discussion
We found that the fauna overwintering in the bark traps in-stalled on apple trees was dominated by predators, of which spi-ders were the most abundant. Among these, two guilds, i.e.,bark-dwelling Anyphaena and Philodromus, and foliage web-build-ing theridiids, were most abundant. We have not surveyed the barkitself, but we have not found them in the litter of fallen leaves(Korenko, unpublished), therefore, we expected them to occurmainly on bark during winter. Bark of apple trees is not very coarsebut provides various spaces, nooks and crannies, so there is likely asegregation of shelters among invertebrates of various sizes. Wefound such spatial segregation in the layers of traps between smalland large spiders. Small spiders hid in small crevices that are inac-cessible to large spiders, and likely serve as an effective passive de-fense during overwintering.
Both guilds showed winter-activity as there was temporal dis-placement of their numbers in bark traps. Because the two guildsdiffer in size and winter-activity pattern, we expected an intraguildpredation between the guilds and even within the bark-dwellingguild. Data from bark traps, however, provided only an indicationfor such interaction, as all negative relationships were not signifi-cant at 5% level. This is probably because Anyphaena, being an ac-tively foraging predator even at low temperatures, has abandonedpatches (traps) with low abundance of other spiders. As a resultcombinations of high Anyphaena and low Philodromus densitieswere lacking.
The two winters were rather warm, particularly 2006/07. Weestimated that 50% hunting activity for Anyphaena was at �0.9 �Cand for Philodromus at 1.6 �C (Korenko et al., 2010). The averagetemperature in both winters was above the threshold for both spe-cies suggesting that both predators must have high predationactivity. The average temperature during the semi-field experi-ment was also above the threshold of both species providing bothspecies with a similar advantage in terms of physiologicalconditions.
During the 10 days of the semi-field experiment a majority ofthe small spiders (theridiids and philodromids) was consumedrevealing that intraguild predation can have a dramatic effect onspider population density during the winter on apple trees. Thismay explain the reduced spider density in apple trees in spring.We found a 5-fold lower density of spiders in tree foliage in the
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
1 5 25 120Exposure [Days]
Per
cen
t co
mp
osi
tio
n
Anyphaena Philodromus Theridiidae Others
r 120 days during winter. (B) Relative composition of the spiders found in cardboard
0 1 2 3 4
0.0
0.2
0.4
0.6
0.8
1.0
foytila tr o
Msu
mo rdolihPshelternone
Size ratio [Anyphaena/Philodromus]
A
0 1 2 3 4
0.0
0.2
0.4
0.6
0.8
1.0
shelternone
Size ratio [Anyphaena/Philodromus]
foytilatro
Man eahpyn
A
B
Fig. 5. Relationship between the mortality of Philodromus (A) and Anyphaena (B) spiders and the average size ratio (Anyphaena/Philodromus) in boards with and withoutshelters provided by corrugated sheets. Logistic models for both types of boards are shown.
S. Korenko, S. Pekár / Biological Control 54 (2010) 206–212 211
spring than in the previous autumn (Pekár, 1999b). Detailed com-parative analysis of the species abundance in spring and autumnrevealed that it was due to decline of small juvenile theridiid spi-ders. We assume that these spiders are most vulnerable to preda-tion during winter, though effect of other factors, abiotic and biotic,on the decrease cannot be excluded.
Are small spiders likely the major victims of IGP? The mostprobable prey of epigeic winter-active spiders are winter-activecollembolans and dipterans (Hågvar, 1973; Aitchison, 1984,1987) as the ideal prey body size for polyphagous spiders is0.25–0.75 of the body length of the predator (Nentwig, 1982). Rep-resentatives of both orders were also found to overwinter on thetree bark, with collembolans being quite abundant. Collembolansmight be too small for large individuals of Anyphaena and Philodro-mus spiders to prey upon. Both winter-active predators appearedto catch and consume other spiders in particular. This might bedue to their movement and higher nutritional value. It is expectedthat movement of prey is an important signal eliciting attack, thuswinter-active prey shall be more prone to attack than inactiveprey. In a laboratory experiment, we found that at low temperatureboth Anyphaena and Philodromus captured both active and inactivespiders, but not immobile flies (Korenko et al., 2010). Small spidersmay be selected as prey because of better nutritional value for win-ter-active predators than herbivorous or detritophagous dipteransand collembolans. Matsumura et al. (2004) suggested that preda-tors should include other predators in their diet in order to maxi-mize protein intake.
Not all predatory species hibernating on bark are prey of win-ter-active spiders. For example, Clubiona spiders rest in a silk cellthat seems to protect them from other predators (Pekár, 1999a).Coccinellid beetles, real bugs (Young, 1989) and linyphiid spiders(Bristowe, 1941) are distasteful to spiders and thus protected fromintraguild predation. Thus IGP should occur between larger spiderspecies that remain active.
In the semi-field experiment, both Anyphaena and Philodromuswere found to catch one another so that the larger specimenscaught the smaller. This agrees with interspecific competition ob-served in other spiders (e.g., Eichenberger et al., 2009). Logit mod-els revealed the threshold ratio, i.e., where the curves cross, for theswitch of predator–prey roles. It occurs at a ratio (Anyphaena/Philo-dromus) = 1.6, i.e., Anyphaena catches Philodromus when it is atleast 60% larger. Philodromus is thus a more robust predator thanAnyphaena at a similar body size. Anyphaena, however, overwintersat 2-times larger size thus shall exert higher predation on Philodro-
mus than Philodromus on Anyphaena. Furthermore, Anyphaena isactive at temperatures below zero, while Philodromus is active onlyat temperatures above zero (Korenko et al., 2010). Anyphaena thuspossesses an adaptive advantage over Philodromus particularlyduring cold winters.
One of the main factors influencing the overwintering composi-tion of the tree trunk spider fauna is the bark structure (Pekár,1999a). Tree bark is a specific habitat where many invertebratespecies live frequently or temporarily. For example, Anyphaena,Philodromus and Clubiona are more affiliated to bark, while theri-diid and araneid spiders are only temporal bark dwellers – over-wintering on bark but foraging by means of webs built amongleaves (Marc and Canard, 1997). Older trees with rougher/coarserbark provide refuge space for larger species, higher number andhigher diversity of invertebrates (Mizell and Schiffhauer, 1987;Pekár, 1999a). Here we found at least 28 species of spiders over-wintering. A similar overwintering composition was observed byM. Isaia (personal communication) in a commercial apple orchardin Italy, with Anyphaena comprising 43% and Philodromus 8% of alloverwintering spiders. But Miliczky et al. (2008) recorded 28 spe-cies of spiders overwintering in bands on apple and pear trees,with several Philodromus species being much more abundant thanAnyphaena.
Change in the composition of the overwintering spider fauna re-lated to the duration of trap exposure revealed that spiders wereable to colonize the traps even in the middle of winter, when thetemperature was about 5 �C. Interestingly, all spiders increasedin abundance, including the ones, such as theridiids, that are notconsidered active during winter. Some invertebrates colonizedthe traps within a single day. As expected the number of specimensincreased with the duration of exposure, but the proportion of Any-phaena spiders declined towards the beginning of spring. This ispresumably due to their escape from shelters as a result of theincreasing temperature by the end of February. Anyphaena wasfound to have higher prey capture rate at low above-zero temper-atures (Korenko et al., 2010), i.e., consuming more prey than attemperatures above 15 �C.
Several variables can influence spider survival and the outcomeof interactions (inter- and intraspecific) during overwintering onthe tree bark. Body size ratio (Marc, 1991) and access to safe ref-uges were found to be the most important. Presence of alternativeprey can reduce intraguild predation (Holt and Polis, 1997; Hodge,1999; Meyhöfer and Hindayana, 2000). Dinter (2002) observed de-creased intraguild predation between Erigone atra Blackwall fe-

212 S. Korenko, S. Pekár / Biological Control 54 (2010) 206–212
males and lacewing larvae by about 44% when additional prey waspresent. In our experiment, the presence of alternative prey did notdecrease intraguild predation. The alternative prey was presum-ably consumed by smaller individuals of Anyphaena and Philodro-mus spiders that could access interstitial space of the cardboardwhere theridiids likely hid.
Suppression of pest populations by polyphagous predators issuccessful when the cumulative impact of the predator guilds ex-ceeds the impact of antagonistic interactions between predators(Losey and Denno, 1998; Wise, 1993; Polis et al., 1989; Rosenheimet al., 1995). There are several mechanisms which can be used toreduce these antagonistic interactions. This study suggests that in-creased spatial heterogeneity of overwintering sites decreasesantagonistic interactions between Anyphaena and Philodromusresulting in higher survival of both predators. Therefore, installa-tion of artificial cardboard bands with various interstitial spacesshall increase the amount of safe space available for overwinteringresulting in an increase of the number of spiders on the tree barkthat could lead to enhanced biological control. Anyphaena andPhilodromus are known to be polyphagous predators (Klein,1988; Korenko, unpublished). Therefore, we expect them to feedon diverse insect pests that occur in apple orchards.
Acknowledgments
We would like to thank J. Viktorín from Fruit-growing companyin Brno for allowing us to collect spiders and perform our study attheir commercial orchard. Two anonymous referees provided use-ful comments to the manuscript. This study was supported by thegrant 1G58081 from the Grant Agency of the Ministry of Agricul-ture of the Czech Republic. SP was supported by the Grant No.MSM0021622416 provided by the Ministry of Education, Youthand Sports of the Czech Republic. S.K. was supported by the GrantNo. 526/09/H025 of the Czech Science Foundation.
References
Aitchison, C.W., 1984. Low temperature feeding by winter-active spiders. Journal ofArachnology 12, 297–305.
Aitchison, C.W., 1987. Feeding ecology of winter-active spiders. In: Nentwig, W.(Ed.), Ecophysiology of Spiders. Springer-Verlag, Berlin, pp. 264–273.
Bogya, S., 1999. Spiders (Araneae) as Polyphagous Natural Enemies in Orchards. PhDThesis, Landbouwuniversiteit Wageningen.
Bogya, S., Mols, P.J.M., 1996. The role of spiders as predators of insect pests withparticular reference to orchards: a review. Acta Phytopathologica etEntomologica Hungarica 31, 83–159.
Bower, L.F., Snetsinger, R., 1985. Tree bark as an overwintering niche for spiders(Araneae) in an oak-maple forest edge. Melsheimer Entomological Series 35, 1–5.
Bristowe, W.S., 1941. The Comity of Spiders II. Ray Society, London.Dinter, A., 1998. Intraguild predation between erigonid spiders, lacewing larvae,
and carabids. Journal of Applied Entomology 122, 163–167.Dinter, A., 2002. Microcosm studies on intraguild predation between female
erigonid spiders and lacewing larvae and influence of single versus multiplepredators on cereal aphids. Journal of Applied Entomology 126, 249–257.
Eichenberger, B., Siegenthaler, E., Schmidt-Entling, M.H., 2009. Body sizedetermines the outcome of competition for webs among alien and nativesheetweb spiders (Araneae: Linyphiidae). Ecological Entomology 34, 363–368.
Foelix, R.F., 1996. Biology of Spiders, second ed. Oxford University Press, New York.Guillebeau, L.P., All, J.N., 1989. Geocoris spp. (Hemiptera: Lygaeidae) and the striped
lynx spider (Araneae: Oxyopidae): cross predation and prey preferences.Journal of Economic Entomology 82, 1106–1110.
Gunnarsson, B., 1985. Interspecific predation as a mortality factor amongoverwintering spiders. Oecologia 65, 498–502.
Hågvar, S., 1973. Ecological studies on a winter-active spider, Bolyphantes index(Thorell) (Araneida, Linyphiidae). Norsk Entomologisk Tidsskrift 20, 309–314.
Heimer, S., Nentwig, W., 1991. Spinnen Mitteleuropas. Paul Parey, Berlin, Hamburg.
Hodge, M.A., 1999. The implications of intraguild predation for the role of spiders inbiological control. Journal of Arachnology 27, 351–362.
Holt, R.D., Polis, G.A., 1997. A theoretical framework for intraguild predation.American Naturalist 149, 745–764.
Horton, D.R., 2004. Phenology of emergence from artificial overwintering sheltersby some predatory arthropods common in pear orchards of the PacificNorthwest. Journal of Entomological Society of British Columbia December101, 108.
Klein, W., 1988. Erfassung und bedetung der in den apfelanlangen aufgetretenenspinnen (Araneae) als nützlinge im grossraum Bonn. PhD Theiss, RheinischenFriedrich Wilhelms Universität Bonn.
Korenko, S., Pekár, S., Honek, A., 2010. Predation activity of two winter-activespiders (Araneae: Anyphaenidae, Philodromidae). Journal of Thermal Biology35, 112–116.
Losey, J.E., Denno, R.F., 1998. Factors facilitating synergistic predation: the centralrole of synchrony. Ecological Applications 9, 378–386.
Marc, P., 1991. Interspecific and intraspecific interactions between spider speciesfrom apple orchards. Bulletin de la Societe Neuchateloise des SciencesNaturelles 116 (1), 177–184.
Marc, P., Canard, A., 1997. Maintaining spider biodiversity in agroecosystems as atool in pest control. Agriculture Ecosystems and Environment 62, 229–235.
Markó, V., Keresztes, B., Fountain, M.T., Cross, J.V., 2009. Prey availability, pesticidesand the abundance of orchard spider communities. Biological Control 48, 115–124.
Matsumura, M., Trafelet-Smith, G.M., Gratton, C., Finke, D.L., Fagan, W.F., Denno,R.F., 2004. Does intraguild predation enhance predator performance? Astoichiometric perspective. Ecology 85 (9), 2601–2615.
McCann, K., Hastings, A., Huxel, G.R., 1998. Weak trophic interactions and thebalance of nature. Nature 395, 794–798.
Meyhöfer, R., Hindayana, D., 2000. Effect of intraguild predation on aphid parasitoidsurvival. Entomologia Experimentalis et Applicata 97, 115–122.
Miliczky, E.R., Horton, D.R., Calkins, C.O., 2008. Observations on phenology andoverwintering of spiders associated with apple and pear orchards in south-central Washington. Journal of Arachnology 36, 565–573.
Mizell, R.F., Schiffhauer, D.E., 1987. Trunk traps and overwintering predators inpecan orchards: survey of species and emergence time. Florida Entomologist 70(2), 238–244.
Moulder, B.C., Reichle, D.E., 1972. Significance of spider predation in the energydynamics of forest floor arthropod communities. Ecological Monographs 42,473–498.
Nentwig, W., 1982. Epigeic spiders: their potential prey and competitors:relationship between size and frequency. Oecologia 55, 130–136.
Nyffeler, M., Benz, G., 1987. Spiders in natural pest control: a review. Journal ofApplied Entomology 103, 321–339.
Olszak, W., Luczak, J., Niemczyk, E., Zajac, R., 1992. The spider communityassociated with apple trees under different pressure of pesticides. EkologiaPolska 40 (2), 265–286.
Pekár, S., 1999a. Some observations on overwintering of spiders (Araneae) in twocontrasting orchards in the Czech Republic. Agriculture Ecosystems andEnvironment 73, 205–210.
Pekár, S., 1999b. Effect of IPM practices and conventional spraying on spiderpopulation dynamics in an apple orchard. Agriculture Ecosystems andEnvironment 73, 155–166.
Pekár, S., Brabec, M., 2009. Modern Analysis of Biological Data. 1. Generalised LinearModels in R. Scientia, Praha. (in Czech).
Polis, G.A., McCormick, S.J., 1986. Scorpions, spiders and solpugids: predation andcompetition among distantly related taxa. Oecologia 71, 111–116.
Polis, G.A., Myers, C.A., Holt, R.D., 1989. The ecology and evolution of intraguildpredation – potential competitors that eat each other. Annual Review of Ecologyand Systematics 20, 297–330.
R Development Core Team, 2008. R: A Language and Environment for StatisticalComputing. R Foundation for Statistical Computing Vienna, Austria. Availablevia <http://www.R-project.org>.
Riechert, S.E., Lockley, T., 1984. Spiders as biological control agents. Annual Reviewof Entomology 29, 299–320.
Roberts, M.J., 1995. Spiders of Britain and Northern Europe. Harper CollinsPublishers, Bath.
Rosenheim, J.A., Kaya, H.K., Ehler, L.E., Marois, J.J., Jaffee, B.A., 1995. Intraguildpredation among biological control agents: theory and evidence. BiologicalControl 5, 303–335.
Schaefer, M., 1977. Winter ecology of spiders (Araneida). Zeitschrift für AngewandteEntomologie 83, 113–134.
Wise, D.H., 1993. Spiders in Ecological Webs. Cambridge University Press,Cambridge.
Wise, D.H., Chen, B., 1999. Impact of intraguild predators on survival of a forest-floor wolf spider. Oecologia 121, 129–137.
Young, O.P., 1989. Predation by Pisaurina mira (Araneae, Pisauridae) on Lyguslineolaris (Heteroptera, Miridae) and other arthropods. Journal of Arachnology17, 43–48.
Copyright © 2022 FDOKUMEN




















