
Is the distribution of Fiji leaf gall in Australian sugarcane explained by variation in the vector...
10
CSIRO PUBLISHING www.publish.csiro.au/journals/app Australasian Plant Pathology, 2006, 35, 103–112 Is the distribution of Fiji leaf gall in Australian sugarcane explained by variation in the vector Perkinsiella saccharicida? A. W. Ridley A,B , K. Dhileepan A,D , K. N. Johnson A,B , P. G. Allsopp B,C , K. A. Nutt C , G. H. Walter A and B. J. Croft B,C,E A School of Integrative Biology, The University of Queensland, Qld 4072, Australia. B CRC for Tropical Plant Protection, The University of Queensland, Qld 4072, Australia. C BSES Limited, PO Box 86, Indooroopilly, Qld 4068, Australia. D Current address: Alan Fletcher Research Station, PO Box 36, Sherwood, Qld 4075, Australia. E Corresponding author. Email: [email protected] Abstract. Fiji leaf gall (FLG) is an important virally induced disease in Australian sugarcane. It is confined to southern canegrowing areas, despite its vector, the delphacid planthopper Perkinsiella saccharicida, occurring in all canegrowing areas of Queensland and New South Wales. This disparity between distributions could be a result of successful containment of the disease through quarantine and/or geographical barriers, or because northern Queensland populations of Perkinsiella may be poorer vectors of the disease. These hypotheses were first tested by investigating variation in the ITS2 region of the rDNA fragment among eastern Australian and overseas populations of Perkinsiella. The ITS2 sequences of the Western Australian P. thompsoni and the Fijian P. vitiensis were distinguishable from those of P.saccharicida and there was no significant variation among the 26 P.saccharicida populations. Reciprocal crosses of a northern Queensland and a southern Queensland population of P. saccharicida were fertile, so they may well be conspecific. Single vector transmission experiments showed that a population of P. saccharicida from northern Queensland had a higher vector competency than either of two southern Queensland populations. The frequency of virus acquisition in the vector populations was demonstrated to be important in the vector competency of the planthopper. The proportion of infected vectors that transmitted the virus to plants was not significantly different among the populations tested. This study shows that the absence of FLG from northern Queensland is not due to a lack of vector competency of the northern population of P. saccharicida. Additional keywords: acquisition, Fiji disease virus, Perkinsiella, Reovirus, transmission. Introduction Fiji leaf gall (FLG) is a virally induced disease of sugarcane that results in stunting and death of infected plants. The definitive symptom of FLG (formerly known as Fiji disease) is the presence of raised, whitish-yellow galls on the abaxial side of the leaf blade and midrib. The galls result from hypertrophy of the phloem and xylem cells in the vascular bundle following infection and replication of Fiji disease virus (family Reoviridae, genus Fijivirus, FDV) within the cells (Egan et al. 1989; Smith 2000). Growth of the stalk slows, newly formed leaves are shorter, darker green and stiffer, and the spindle leaf often dies. Symptoms appear quickest on young, rapidly growing plants of susceptible cultivars, with the minimum time to symptom expression being 15–29 days after infection (Mungomery 1946; Egan et al. 1989). In more resistant cultivars or slow-growing plants, symptom expression can take many months. Plants grown from infected cuttings are usually stunted and often die. FDV is vectored by delphacid planthoppers of the genus Perkinsiella Kirkaldy. Epidemics of FLG have occurred in Australian sugarcane, most recently in Bundaberg during the 1970s and 1980s. At that time, the FLG-susceptible cultivar NCo310 comprised more than 90% of the area’s crop and P. saccharicida was present at high densities (Bull 1977; Ryan 1988). The disease reduced yields in plant crops by 50% and in second ratoons by 100%. Additional costs were incurred in the production of disease-free planting material and replanting crops prematurely (Ryan 1988). Resistant cultivars and disease-free planting material were used to bring the epidemic under control (Ryan 1988). FLG is still active in two of the three mill areas in New South Wales and in the © Australasian Plant Pathology Society 2006 10.1071/AP06011 0815-3191/06/020103
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Is the distribution of Fiji leaf gall in Australian sugarcane explained by variation in the vector...

CSIRO PUBLISHING
www.publish.csiro.au/journals/app Australasian Plant Pathology, 2006, 35, 103–112
Is the distribution of Fiji leaf gall in Australian sugarcane explainedby variation in the vector Perkinsiella saccharicida?
A. W. RidleyA,B, K. DhileepanA,D, K. N. JohnsonA,B, P. G. AllsoppB,C, K. A. NuttC,G. H. WalterA and B. J. CroftB,C,E
ASchool of Integrative Biology, The University of Queensland, Qld 4072, Australia.BCRC for Tropical Plant Protection, The University of Queensland, Qld 4072, Australia.
CBSES Limited, PO Box 86, Indooroopilly, Qld 4068, Australia.DCurrent address: Alan Fletcher Research Station, PO Box 36, Sherwood, Qld 4075, Australia.
ECorresponding author. Email: [email protected]
Abstract. Fiji leaf gall (FLG) is an important virally induced disease in Australian sugarcane. It is confined tosouthern canegrowing areas, despite its vector, the delphacid planthopper Perkinsiella saccharicida, occurringin all canegrowing areas of Queensland and New South Wales. This disparity between distributions could bea result of successful containment of the disease through quarantine and/or geographical barriers, or becausenorthern Queensland populations of Perkinsiella may be poorer vectors of the disease. These hypotheses were firsttested by investigating variation in the ITS2 region of the rDNA fragment among eastern Australian and overseaspopulations of Perkinsiella. The ITS2 sequences of the Western Australian P. thompsoni and the Fijian P. vitiensiswere distinguishable from those of P. saccharicida and there was no significant variation among the 26 P. saccharicidapopulations. Reciprocal crosses of a northern Queensland and a southern Queensland population of P. saccharicidawere fertile, so they may well be conspecific. Single vector transmission experiments showed that a population ofP. saccharicida from northern Queensland had a higher vector competency than either of two southern Queenslandpopulations. The frequency of virus acquisition in the vector populations was demonstrated to be important in thevector competency of the planthopper. The proportion of infected vectors that transmitted the virus to plants wasnot significantly different among the populations tested. This study shows that the absence of FLG from northernQueensland is not due to a lack of vector competency of the northern population of P. saccharicida.
Additional keywords: acquisition, Fiji disease virus, Perkinsiella, Reovirus, transmission.
IntroductionFiji leaf gall (FLG) is a virally induced disease of sugarcanethat results in stunting and death of infected plants. Thedefinitive symptom of FLG (formerly known as Fiji disease)is the presence of raised, whitish-yellow galls on the abaxialside of the leaf blade and midrib. The galls result fromhypertrophy of the phloem and xylem cells in the vascularbundle following infection and replication of Fiji diseasevirus (family Reoviridae, genus Fijivirus, FDV) within thecells (Egan et al. 1989; Smith 2000). Growth of the stalkslows, newly formed leaves are shorter, darker green andstiffer, and the spindle leaf often dies. Symptoms appearquickest on young, rapidly growing plants of susceptiblecultivars, with the minimum time to symptom expressionbeing 15–29 days after infection (Mungomery 1946; Eganet al. 1989). In more resistant cultivars or slow-growing
plants, symptom expression can take many months. Plantsgrown from infected cuttings are usually stunted and oftendie. FDV is vectored by delphacid planthoppers of the genusPerkinsiella Kirkaldy.
Epidemics of FLG have occurred in Australian sugarcane,most recently in Bundaberg during the 1970s and 1980s. Atthat time, the FLG-susceptible cultivar NCo310 comprisedmore than 90% of the area’s crop and P. saccharicidawas present at high densities (Bull 1977; Ryan 1988).The disease reduced yields in plant crops by 50% and insecond ratoons by 100%. Additional costs were incurredin the production of disease-free planting material andreplanting crops prematurely (Ryan 1988). Resistant cultivarsand disease-free planting material were used to bring theepidemic under control (Ryan 1988). FLG is still active intwo of the three mill areas in New South Wales and in the
© Australasian Plant Pathology Society 2006 10.1071/AP06011 0815-3191/06/020103

104 Australasian Plant Pathology A. W. Ridley et al.
canegrowing areas close to Brisbane (Fig. 1). It is currentlymanaged in Queensland and NSW through the compulsoryuse of resistant cultivars. The inability to grow superior butsusceptible cultivars in FLG-affected areas is a continuingcost to the industry.
FLG was originally recognised in Fiji but probablyoriginated in New Guinea (Smith 2000), where it iswidespread. FLG also occurs in Malaysia, Philippines,Thailand, Vanuatu, Fiji and Samoa. It has been recorded inMadagascar, but not since 1971 (Egan et al. 1989; Smith2000). In eastern Australia, FLG has never been found insugarcane areas from the Burdekin north (Fig. 1) even thoughthe vectors are present. Northern Queensland has similarconditions to other tropical sugarcane growing areas whereFLG occurs so it is unlikely that environmental conditionsinhibit the spread of the disease. Currently cultivars
Fig. 1. The Australian sugar industry (Anon. 2003).
susceptible to FLG are grown in northern Queensland fortheir superior agronomic value.
The disparity between vector and virus distribution inAustralia is consistent with alternative interpretations. Firstly,quarantine boundaries, in conjunction with geographicalseparation of sugarcane areas, may restrict the disease tosouthern Queensland and New South Wales. FDV did breachthe Bundaberg–Mackay barrier (Fig. 1) during the epidemicin Bundaberg, but rapid implementation of control measureshas resulted in no further reports for more than 10 years.Secondly, the vector competency of the northern Queenslandpopulation of P. saccharicida may be much lower than that ofthe southern populations, to the extent that even if the diseasereached the north, the vectors may be unable to transmitit. Correct identification of pest species and understandingtheir vector status is important for management of FLG

Perkinsiella vector competency Australasian Plant Pathology 105
in Australia. If there is only one species of Perkinsiellapresent in sugarcane on the east coast, then movementof insects from FDV-free areas to an area where FDVoccurs is of little importance. However, rigorous quarantinemust be maintained to restrict the movement of FLG. If,on the other hand, an undetected species of Perkinsiellaexists in northern Queensland, and it is significantly moreefficient at transmitting FDV than P. saccharicida populationscurrently present in southern Queensland, then movementof insects from northern Queensland becomes a risk forsouthern regions.
These hypotheses were tested using a number ofapproaches. First, rDNA internal transcribed spacer 2 (ITS2)sequence data were used to determine if there is geneticdifferentiation among sugarcane populations of Perkinsiellafrom eastern Australia. Reciprocal crossing of a northernQueensland and a southern Queensland population was usedto further investigate potential species differences. Second,the differences in the vector competency of P. saccharicidafrom Tully (northern Queensland) with that of two southernpopulations (Woodford and Bundaberg) were tested. Thisstudy also determined whether any differences in vectoracquisition are maintained over prolonged periods by rearingthe three vector populations for five generations on plantsinfected with FDV.
The virus
Fiji disease virus is a member of the Reoviridae family ofviruses, a diverse group of double-stranded RNA virusesinfecting plants, vertebrates and arthropods. It is the typespecies of the genus Fijivirus that includes nine speciesdivided into five groups based on vectors, plant hosts andsimilarities in serological and nucleotide sequence. FDV isthe sole member of group 1 and is most closely related to theviruses in group 2: Rice black-streaked dwarf virus, Maizerough dwarf virus, Pangola stunt virus and Mal del RioCuarto virus (Mertens et al. 2000; McQualter et al. 2003).FDV particles are icosahedral and contain a 10-segmenteddsRNA genome (Mertens et al. 2000).
FDV proliferates within the insect vector and the plant host(Francki and Grivell 1972). Phylogenetic and transmissionrelationships suggest that FDV may be an insect virus witha secondary plant host (Ikeda et al. 2001; McQualter et al.2003; Smith and Candy 2004). Comparison of data foracquisition, latency, infectivity and transmission within theFDV-Perkinsiella-sugarcane system with the parameters thatdefine plant-virus transmission classes (Madden et al. 2000;Nault 1997) suggests that the system is representative of apropagative, persistently transmitted plant virus.
FDV is spread by the use of infected planting material,but is transmitted between plants only by the insectvectors. All attempts to mechanically transmit FDV havefailed, including injection of ‘infected’ juice, infiltrationof ‘infected’ juice into test stalks before planting, grafting
diseased and healthy plants, and replacing holes made in thecentre of the internodes of test plants with correspondingplugs from a diseased plant (Hughes and Robinson 1961;Mungomery and Bell 1933).
The vector
Three species of Perkinsiella are demonstrated vectorsof FDV: P. vitiensis Kirkaldy in Fiji (Hussain et al.1965), P. vastatrix (Breddin) in the Philippines (Ocfemia1934) and P. saccharicida in Australia, Malaysia andThailand (Mungomery and Bell 1933; Egan et al. 1989).Circumstantial evidence suggests that P. lakokensis Muiris a vector (Buzacott 1953) and other Perkinsiella speciesmay also be vectors. The genus appears to have originatedin New Guinea and adjacent islands, with 13 species knownfrom Papua New Guinea (Fennah 1979). P. saccharicidahas been moved with sugarcane and now has a widerdistribution than the virus, occurring in Hawaii, theAmericas, Mauritius, Reunion, southern Africa, China andTaiwan (FitzGibbon et al. 1998). Two Perkinsiella speciesare thought to occur on sugarcane in sugarcane-producingareas in Australia: P. saccharicida in eastern Australia andP. thompsoni Muir, a species of unknown vector potentialthat occurs in the Ord River Irrigation Area (ORIA) ofWestern Australia (Fletcher and Lariviere 2003) (Fig. 1). Themorphology of all known specimens from eastern Australiais consistent with the description of P. saccharicida, buta non-vector population or cryptic species (Paterson1991) could explain the absence of FLG fromnorthern Queensland.
Smith and Candy (2004) summarise the characteristicsof transmission for the FDV–Perkinsiella–sugarcane system.Only the early instars (1–3) of the vector can acquire thevirus from infected plants (Anon. 1979; Egan et al. 1989).Why later instars and adults do not become infected withFDV is not known. Nymphs transmit about 12–14 days afteracquiring the virus (Chang 1977; Egan et al. 1989), butless than 50% of plants develop FLG when fed on by aninfected vector (Egan et al. 1989; Francki et al. 1986). Theacquisition and transmission of FDV by P. saccharicida wasinvestigated by caging adult vectors (from a population bredon diseased plants) on 3-week-old plants for 7 days (Franckiet al. 1986). Only 15% of the vectors were diagnosed byimmunoelectron microscopy as having a FDV infection. Ofthose infected vectors, 41% transmitted FDV to test sugarcaneplants, a vector competency of <10%. The failure of someof the infected vectors to transmit FDV to test plants maybe related to the feeding behaviour of P. saccharicida or theinstability of FDV (Francki et al. 1986). Even though vectorsmay have introduced FDV into a test plant, the plant neednot necessarily have developed FLG. This implies that onlythose vectors whose host plant goes on to develop FLG wereconsidered to have transmitted FDV. The term ‘transmission’was used strictly in relation to development of FLG in test

106 Australasian Plant Pathology A. W. Ridley et al.
plants by Francki et al. (1986), and this definition was usedfor this study.
P. saccharicida from the Ingham area in northernQueensland can transmit FLG (Osborn and Forteath 1972).However, the vector competency of the northern Queenslandpopulation has never been compared with that of southernpopulations. Differences in vector competency could explainthe absence of FLG from northern Queensland.
MethodsMolecular phylogeny
Putative P. saccharicida, P. thompsoni and P. vitiensis were obtainedfrom localities given in Table 1. On collection, all insects were storedin >80% ethanol and identifications were confirmed morphologicallyby Murray Fletcher, NSW Department of Primary Industries,Orange, NSW.
DNA was extracted from individual adults using a 96-wellmodification of the protocol of Miller et al. (1998). Variation wasassessed among a subset of the collections by sequencing the
Table 1. Species, collection codes and collection localities for specimens examinedMultiple listings for the one locality are collections made at different times
Species Collection code Location Latitude, longitude GenBank accessionnumbers
Perkinsiella saccharicida PsAirQLD Airlie Beach Qld 20◦16′S 148◦43′E AY895060PsBrucQLD Ayr, Qld 19◦34′S 147◦24′E AY895061PsBundQLD Bundaberg, Qld 24◦52′S 152◦21′E AY895062PsChilQLD Childers, Qld 25◦14′S 152◦17′E AY895063PsClewUSA Clewiston, Florida, USA 26◦45′N 80◦56′W AY895064PsDunUSA Dunwody Farm, Florida, USA 26◦46′N 80◦53′W AY895065PsHarNSW Harwood, NSW 29◦26′S 153◦15′E AY895066PsHomQLD Home Hill, Qld 19◦40′S 147◦25′E AY895067PsKN Mount Edgecombe, KwaZulu-Natal, 29◦55′S 30◦56′E AY895068
South AfricaPsLouUSA Lafourche Parish, Louisiana, USA 29◦46′N 90◦46′W AY895069PsMacQLD1 Mackay, Qld 21◦09′S 149◦11′E AY895070PsMacQLD2 AY895071PsMaNSW Maclean, NSW 29◦28′S 153◦12′E AY895072PsMarQLD Maryborough, Qld 25◦32′S 152◦43′E AY895073PsProQLD Proserpine, Qld 20◦24′S 148◦35′E AY895074PsSarQLD Sarina, Qld 21◦26′S 149◦13′E AY895075PsSaReun Savannah, Reunion 20◦50′S 55◦26′E AY895076PsSMReun Sainte-Marie, Reunion 20◦52′S 55◦30′E AY895077PsTLouUSA Terrebonne Parish, Louisiana, USA 29◦30′N 91◦30′W AY895078PsTulQLD Tully, Qld 17◦55′S 145◦55′E AY895079PsWalQLD Wallaville, Qld 25◦05′S 151◦59′E AY895080PsWood1 Woodford, Qld 26◦57′S 152◦47′E AY895081PsWood2 AY895082PsWood3 AY895083PsYanQLD Yandina, Qld 26◦34′S 152◦57′E AY895084PsYukQLD Yukan via Sarina, Qld 21◦31′S 149◦14′E AY895085
Perkinsiella thompsoni PtOrdWA Waringarri Aboriginal Corporation, 15◦47′S 128◦44′E AY895086Kununurra, WA
Perkinsiella vitiensis PvTaFiji1 Tauarau, Lautoka, Fiji 17◦37′S 177◦27′E AY895087PvTaFiji2 AY895088PvViFiji1 Vitogo, Lautoka, Fiji 17◦36′S 177◦26′E AY895089PvViFiji2 AY895090
Cytochrome oxidase two (COII) mitochondrial gene, 16S and ITS2rDNA genes from five individuals from each of P. vitiensis (Tauarau,Fiji), P. vitiensis (Vitogo, Fiji) and P. saccharicida (Clewiston, USA;Mount Edgecombe, KwaZulu-Natal, South Africa; Savannah, Reunion;Airlie Beach, Qld; Sarina, Qld; Mackay, Qld; Harwood, NSW; Maclean,NSW). The ITS2 fragment was then sequenced from five individualsfrom all of the remaining collections in Table 1.
PCR of the COII DNA fragment was carried out in 25 µL totalreaction volume containing 4 mM MgCl2, 20 mM Tris-HCl, 100 mM KCl,0.2 mM each dNTP, 0.2 µM each primer mtD16 and mtD20 (Liu andBeckenbach 1992), 1U Taq polymerase (Qiagen) and 20 ng DNA.Thermal cycling was performed in a PC960 Thermal Cycler (CorbettResearch) using the following cycling conditions: 35 cycles of 94◦C for30 s, 50◦C for 60 s and 72◦C for 60 s.
PCR of the 16S DNA fragment was carried out in 25 µLtotal reaction volume containing 2 mM MgCl2, 20 mM Tris-HCl,100 mM KCl, 0.2 mM each dNTP, 0.2 µM each primer (16ScbF:5′-AAGATTTTAATGATCGAACAG-3′, 16ScbR: 5′-TGACTGTACAAAGGTAGCATA-3′), 1U Taq polymerase and 20 ng DNA. Thermalcycling was performed using the following conditions: 40 cycles of92◦C for 45 s, 50◦C for 60 s and 72◦C for 90 s.

Perkinsiella vector competency Australasian Plant Pathology 107
PCR of the ITS2 DNA fragment was carried out in 25 µLtotal reaction volume containing 1.5 mM MgCl2, 20 mM Tris-HCl,100 mM KCl, 0.2 mM each dNTP, 0.2 µM each primer (ITSB:5′-TCCTCCGCTTATTGATATGC-3′, ITSN2: 5′-TTCTCGCATCGATGAAGAACG-3′), 1U Taq polymerase and 20 ng DNA. Thermalcycling was performed using the following conditions: 30 cycles of94◦C for 20 s, 50◦C for 20 s and 72◦C for 30 s.
Three µL of each PCR reaction was run on a 1% TBE agarose gelto confirm amplification. The remaining PCR product was purified byMultiScreen PCR plates (Millipore, North Ryde, NSW). Sequencingwas performed in a 12 µL total reaction volume containing 4 µL ofABI dye terminator chemistry (Applied Biosystems), 3.2 pM of primerand 50 ng of PCR product in a PC960 Thermal Cycler using a cyclingprogram of 94◦C for 5 min followed by 30 cycles of 96◦C for 10 s, 50◦Cfor 5 s and 60◦C for 4 min. Sequences were purified using MontageSEQ96 sequencing reaction clean-up kits (Millipore, North Ryde,NSW) and run on an ABI 3700 DNA sequencer (Australian GenomeResearch Facility, Brisbane). Consensus sequences for each taxon(derived from aligned forward and reverse complemented sequencesof multiple individuals) were aligned using BioEdit (Hall 1999) andrefined manually to improve positional homology. The dataset wasanalysed with PAUP* 4.0 (Swofford 2002), using the Neighbour-joiningalgorithm (Saito and Nei 1987), with optimality criterion set for distanceusing log-determinant (Steel 1994; Lockhart et al. 1994) parameters.The reliability of the branching order was determined by 1000 bootstrapreplications at a confidence level of 95% (Felsenstein 1985). Thephylogenetic tree was rooted with P. thompsoni as a monophyletic sisteroutgroup to the remaining taxa.
Population crossing
Late-instar nymphs from the Tully and Woodford populations werecaged onto individual plants to obtain virgin adults. Crosses were madewith individual females and males of both populations. Six Tully femaleswere crossed with Woodford males and two Tully males were crossedwith Woodford females. Crosses were examined 5 weeks later foroffspring production.
Vector competency
Approximately 100 P. saccharicida were collected from each of threesites in Queensland with various histories of FLG. Site 1—Bundaberg:the centre of an epidemic in the 1970–80s, but a rigorous managementscheme, including the use of resistant cultivars, brought the diseaseunder control but the last record of FLG in Bundaberg was in 1986.Site 2—Tully: no record of FLG since first sugarcane grown in the1920s. Site 3—Woodford: FLG has been actively cultivated at BSESresearch station Woodford near Brisbane since 1997 and continues FLGscreening experiments that commenced in the 1950s at another BSESresearch station based at Brisbane. The P. saccharicida population atboth of these Brisbane research stations has been continually exposedto FDV.
Vector competency was tested for each of the three populationsby establishing five replicates each of 10 female and five male adultP. saccharicida in cages containing 3–5 plants of cultivar NCo310showing obvious symptoms of FLG infection. The adults were removedfrom the source plants 1 week after release and the plants weremaintained in the glasshouse to allow the eggs laid by the adultsto hatch and mature. From each cage, 15 third instar nymphs wererandomly selected and placed individually onto test plants of cultivarNCo310. The NCo310 was planted using cuttings from the BSESdisease-free isolation plot at Kallangur and they were 4-weeks-old whenthe nymphs were released. After 7 days, the nymphs were removedand frozen at –20◦C for RT–PCR diagnosis of FDV infection. Theplants were sprayed at weekly intervals with imidacloprid (Confidor50 g kg/a.i.) insecticide and were placed in an insect-proof glasshouse
and examined for FLG-symptom development every 2 months for6 months. Differences in infection levels were determined using analysisof variance of the proportion of test plants from each replicate to developFLG symptoms.
To determine if the planthoppers were infected with FDV, nucleicacid was extracted from frozen specimens. Each was ground in100 µL of TPS buffer (100 mM Tris pH 9.5, 1 M KCl, 10 mM EDTA)using a micropestle and the extract was stored on ice. Thehomogenate was boiled for 10 min then returned to ice. Tubeswere vortexed briefly after the addition of 50 µL of chloroformand followed by centrifugation at 15 000 g for 5 min. Twenty µLof the supernatant were transferred to a new tube for storageat −20◦C.
A C.therm polymerase one-step RT–PCR kit from Roche wasused to determine if samples contained FDV. The primers usedwere FDV7R 5′-CAAGTGGTGACGAAATGA-3′, FDV7F 5′-CCGATTACGGTCAGACTGTTCTT-3′ (Smith and Van de Velde 1994). Theprimers and template were boiled at 99◦C for 2 min to denature thedsRNA, and then cooled rapidly with ice for 5 min before the kit reagentswere added. The cycling conditions were: 60◦C for 30 min, 94◦C for2 min, 35 cycles of 94◦C for 1 min, 60◦C for 1 min, 72◦C for 1 min anda final hold of 72◦C for 7 min. Samples were screened with the insectCOII primer pair A298 and B-tLys (Liu and Beckenbach 1992) as apositive control for the extraction procedure. Any negative samples forthis control were seen as extraction failures and were excluded fromall analyses to avoid false negatives. Differences in infection rates weredetermined using an analysis of variance on the proportion of vectorsinfected with FDV from each replicate.
Exposure of FDV to P. saccharicida
Planthoppers from Tully, Woodford and Bundaberg were reared onsugarcane grown from FDV-infected planting material of cultivarNCo310 for five generations. For each population, five caged potswere used with 4–5 diseased plants per pot to rear the planthoppers.Adults were collected from each pot and 10 individuals from eachcage were taken for RT–PCR diagnosis for FDV. A sample of insectsfrom each population was pooled and then released onto five new plantswith FLG. The protocols for FDV diagnosis were as above. The meaninfection rate for each population in each generation was analysed usingtwo-way ANOVA.
Results
Molecular phylogeny
Of the three DNA regions sequenced from the initial set ofpopulations, the ITS2 fragment had the most variation andwas used to analyse the phylogeny of all 26 populationsof P. saccharicida. Only two of the 26 collections showedpolymorphisms that differed from the consensus, both by asingle base change. This level of variation was not sufficientto identify any one population of P. saccharicida as beingdifferent from other populations originating from differentlocales. The sequence of the ITS2 region from P. thompsoniwas distinctly different from all of the P. saccharicida andP. vitiensis samples. Samples of P. vitiensis fell into twogroups, corresponding to their collection locations, but thesequences differed in only a few bases (Fig. 2). Sequences ofall populations have been lodged in GenBank. The accessionnumbers are in Table 1.

108 Australasian Plant Pathology A. W. Ridley et al.
Fig. 2. Phylogeny of samples of Perkinsiella saccharicida (from Australia and overseas),P. thompsoni and P. vitiensis based on ITS2 sequence. Location codes are given in Table 1.The scale bar indicates the distance between taxa based on the number of nucleotidesubstitutions per site.
Population crosses
Seven of the eight crosses between Tully and WoodfordP. saccharicida produced offspring. One of the Tullyfemale/Woodford male crosses did not produce offspring.
Vector competency
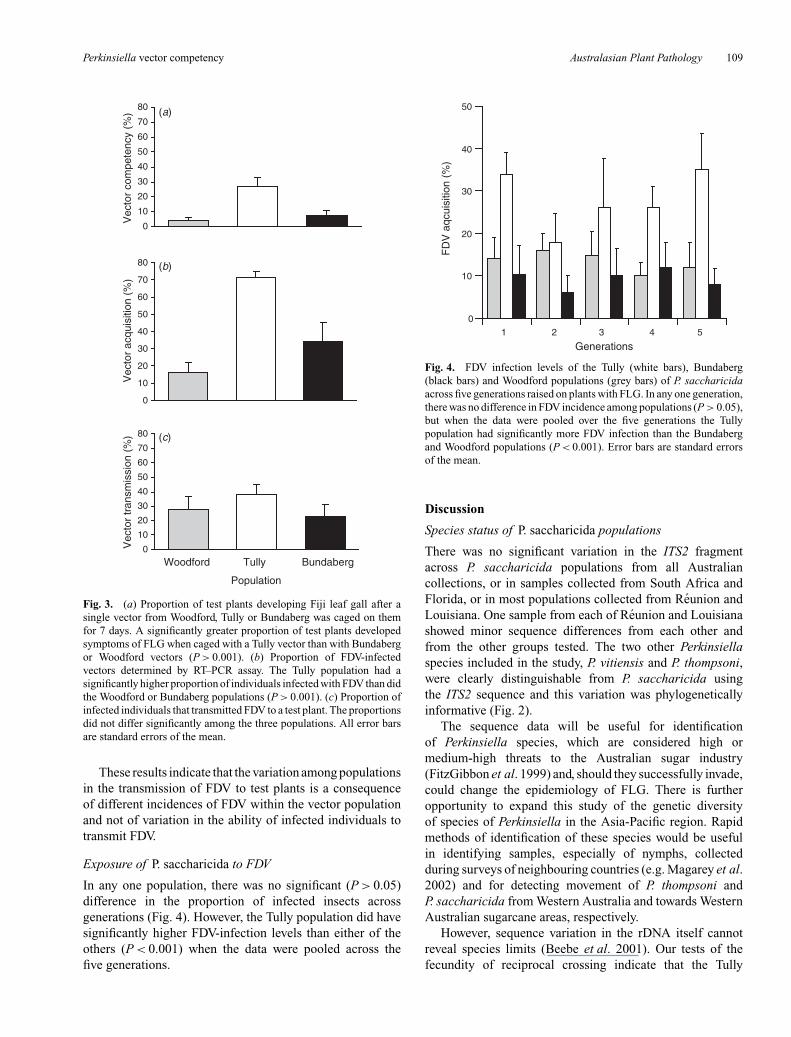
Significantly more test plants infested with TullyP. saccharicida developed symptoms of FLG than did plantsinfested with planthoppers from Bundaberg or Woodford
(P > 0.001) (Fig. 3a). There was no significant differencein infection levels across the Woodford and Bundabergpopulations. The Tully population had a significantly higherproportion of planthoppers infected with FDV than dideither the Woodford or Bundaberg populations (P > 0.001)(Fig. 3b). However, all individuals collected from plants thatdeveloped FLG symptoms were confirmed as infected withFDV. The proportion of FDV-positive vectors that transmittedFDV to test plants did not differ significantly among the threepopulations (Fig. 3c).
Perkinsiella vector competency Australasian Plant Pathology 109
0
10
20
30
40
50
60
70
80
Vec
tor
com
pete
ncy
(%)
Vec
tor
acqu
isiti
on (
%)
0
10
20
30
40
50
60
70
80
Woodford Tully Bundaberg
Vec
tor
tran
smis
sion
(%
)
Population
0
10
20
30
40
50
60
70
80
(a)
(b)
(c)
Fig. 3. (a) Proportion of test plants developing Fiji leaf gall after asingle vector from Woodford, Tully or Bundaberg was caged on themfor 7 days. A significantly greater proportion of test plants developedsymptoms of FLG when caged with a Tully vector than with Bundabergor Woodford vectors (P > 0.001). (b) Proportion of FDV-infectedvectors determined by RT–PCR assay. The Tully population had asignificantly higher proportion of individuals infected with FDV than didthe Woodford or Bundaberg populations (P > 0.001). (c) Proportion ofinfected individuals that transmitted FDV to a test plant. The proportionsdid not differ significantly among the three populations. All error barsare standard errors of the mean.
These results indicate that the variation among populationsin the transmission of FDV to test plants is a consequenceof different incidences of FDV within the vector populationand not of variation in the ability of infected individuals totransmit FDV.
Exposure of P. saccharicida to FDV
In any one population, there was no significant (P > 0.05)difference in the proportion of infected insects acrossgenerations (Fig. 4). However, the Tully population did havesignificantly higher FDV-infection levels than either of theothers (P < 0.001) when the data were pooled across thefive generations.
0
10
20
30
40
50
1 2 3 4 5
Generations
FD
V a
qcui
sitio
n (%
)
Fig. 4. FDV infection levels of the Tully (white bars), Bundaberg(black bars) and Woodford populations (grey bars) of P. saccharicidaacross five generations raised on plants with FLG. In any one generation,there was no difference in FDV incidence among populations (P > 0.05),but when the data were pooled over the five generations the Tullypopulation had significantly more FDV infection than the Bundabergand Woodford populations (P < 0.001). Error bars are standard errorsof the mean.
Discussion
Species status of P. saccharicida populations
There was no significant variation in the ITS2 fragmentacross P. saccharicida populations from all Australiancollections, or in samples collected from South Africa andFlorida, or in most populations collected from Reunion andLouisiana. One sample from each of Reunion and Louisianashowed minor sequence differences from each other andfrom the other groups tested. The two other Perkinsiellaspecies included in the study, P. vitiensis and P. thompsoni,were clearly distinguishable from P. saccharicida usingthe ITS2 sequence and this variation was phylogeneticallyinformative (Fig. 2).
The sequence data will be useful for identificationof Perkinsiella species, which are considered high ormedium-high threats to the Australian sugar industry(FitzGibbon et al. 1999) and, should they successfully invade,could change the epidemiology of FLG. There is furtheropportunity to expand this study of the genetic diversityof species of Perkinsiella in the Asia-Pacific region. Rapidmethods of identification of these species would be usefulin identifying samples, especially of nymphs, collectedduring surveys of neighbouring countries (e.g. Magarey et al.2002) and for detecting movement of P. thompsoni andP. saccharicida from Western Australia and towards WesternAustralian sugarcane areas, respectively.
However, sequence variation in the rDNA itself cannotreveal species limits (Beebe et al. 2001). Our tests of thefecundity of reciprocal crossing indicate that the Tully

110 Australasian Plant Pathology A. W. Ridley et al.
and Woodford populations represent a single species,but evidence of this nature is also not conclusive(Fernando and Walter 1997). The only definitive method tocharacterise a species is to study its specific mate-recognitionsystem (SMRS) (Paterson 1991). The use of laboratory-basedmate crossing experiments may exclude components of thesibling species’ SMRS. Restriction of the SMRS of the twosibling species to the components that are identical betweenthe two species will result in mating that would not occurin nature. Characterisation of the SMRS of P. saccharicidashould be undertaken to provide definitive evidence of thespecies status of P. saccharicida in Australia. Crossingexperiments involving P. saccharicida and P. thompsoni fromthe ORIA would be useful to determine the robustness of suchcrossing experiments in distinguishing different species.
Vector competency across Australianplanthopper populations
These results also support the earlier finding thatP. saccharicida from northern Queensland can vector FDV(Osborn and Forteath 1972). In addition, the comparisons inthis study show that the Tully population has a higher vectorcompetency than either of the two southern populations(Fig. 3a). In the FDV–Perkinsiella–sugarcane system, vectorcompetency involves at least two processes: the vector mustacquire the virus and the virus must then be transmitted tothe test plant.
The Tully population acquired the virus at a greaterfrequency than the southern populations (Fig. 3b), but theproportion of infected Tully vectors that transmitted thevirus to test plants was not significantly different fromthe rates recorded for the other populations. The higherlevel of infection of the population of Tully vectors wasresponsible for a significantly greater proportion of the testplants becoming infected.
Intraregional variation in vector competency has beendemonstrated in a number of studies on the mosquitoAedes aegypti, a vector of dengue fever Flaviviruses (Knoxet al. 2003 and references therein). Mapping of loci thataffect the expression of quantitative traits controlling thedevelopment of the midgut infection barrier and the midgutescape barrier suggests that variation in the development ofthese barriers across populations of A. aegypti explains thevariation in vector competency across populations of thismosquito (Bosio et al. 2000). The factors that contribute toP. saccharicida acquiring FDV have not been establishedand the results of this study have revealed no geneticdifference across the populations of P. saccharicida inthe ITS2 fragment. Further investigation of the process ofinfection and the genetics of the Australian P. saccharicidapopulations is needed. This study suggests that the Tullypopulation may develop a midgut barrier later in the lifecycle than do planthoppers in the southern populations,thus providing more time for the Tully vectors to become
infected. Susceptibility of leafhopper midgut epithelial cellsto infection and gut permeability to Wound tumour virusdecreases with increasing vector age (Sinha 1963), providingsome precedent for the hypothesis challenged in this study.Alternatively, variation in the midgut microbiota acrosspopulations could lead to variation in the acquisition ofvirus as proposed, on indirect evidence, for the uptake ofarboviruses by Culicoides spp. (bitting midges) (Campbellet al. 2004). A clear understanding of the mechanismby which P. saccharicida becomes infected with FDVis important to understanding differential infection ratesacross populations.
Differential transmission across planthopper individuals
This study has demonstrated that acquisition is an importantfactor in determining the vector competency of differentP. saccharicida populations (Fig. 3a, b and c), but the factorsthat determine why some individual vectors transmit andothers do not, remains unknown. Low levels of transmissionhave been recorded in other closely related virus/vectorsystems (Milne and Lovisolo 1977; Arneodo et al. 2002;Wang et al. 2002). Extending the time that an FDV-infectedP. saccharicida planthopper remains on a sugarcane plant hasbeen suggested to increase the probability of transmission(Smith and Candy 2004) but, in this study, the planthopperswere allowed to feed for 7 days. It is unlikely that any vectorsthat did not transmit during that time would have done sogiven more time.
The study of transmission by infected vectors is difficult,because the only non-destructive method to identity infectedvectors is to put them on test plants and wait for symptomdevelopment; this can take months, by which time the vectorsare dead. Future studies could compare the virus titre ininsects that had been exposed to test plants, to compare thetitre of those that transmit against those that failed to transmit.Furthermore, the tissues in the vector that are infected withthe FDV in the two groups should be identified to determineif salivary gland infection is critical for FDV transmission.Studies in other vector systems have shown that insects thatare infected but fail to transmit do not have virus in thesalivary glands (Assis Filho et al. 2004; Moritz et al. 2004;Shikata and Kitagawa 1977). For example, only those thripsthat acquired Tomato spotted wilt virus (TSWV) as larvaecan transmit the virus. Although adults continue to becomeinfected with TSWV, the virus does not travel to the salivaryglands and so cannot be transmitted. The process that inhibitsTSWV from entering the salivary glands of adult thrips is stillin dispute (Assis Filho et al. 2004; Moritz et al. 2004).
Francki et al. (1986) conducted a study on the vectorcompetency of P. saccharicida. There is no indication ofwhere the vectors in their study were collected, but it islikely to have been from a southern Queensland population.The vector competency, vector acquisition rates and vectortransmission rates are similar to our results for the two

Perkinsiella vector competency Australasian Plant Pathology 111
southern populations. However, the age and cultivar of thetest plants do influence transmission rates of FDV andcomparisons of vector competency and transmission ratesacross populations should be controlled for these factors.The assay used to detect FDV infection needs to be sensitiveenough to identify all infected individuals. Francki et al.(1986) misdiagnosed some of the planthoppers they usedas being non-infected but transmitting FDV. Their use ofimmunoelectronmicroscopy was not sensitive enough toinvestigate FDV acquisition accurately. The RT–PCR used inthis study has increased the sensitivity of detection methodssignificantly. All of the individuals that transmitted FDV totest plants in these experiments were confirmed positive byRT–PCR.
This study shows that the northern population ofP. saccharicida is a competent vector of FDV and, therefore,the resident population of the insect could spread the diseasein northern Queensland. This suggests that the absence ofthe disease from this area is a consequence of a combinationof natural geographic barriers to the movement of FDV-infected P. saccharicida and strict quarantine restrictionson the movement of sugarcane in Queensland. UnderQueensland legislation, it is an offence to move sugarcanefrom the southern districts, where FLG is present, to northerndistricts. FLG-susceptible cultivars that are grown in northernQueensland could suffer severe yield losses if FLG spreadto this region and the use of FLG-susceptible germplasm inplant breeding would need to be restricted, reducing geneticgain. The greater vector competency of the northern vectorpopulation may mean that the disease would spread even morerapidly in this region than it did in southern Queensland.Preventing the spread of FLG to northern Queensland is vitalas more than 50% of the Australian sugar industry is basedin this region.
Acknowledgements
We thank Regis Goebel (Reunion), Bill White (Louisiana),Graeme Leslie (South Africa), David Hall (Florida) andRupeni Tamanikaiyaroi (Fiji) for sending planthoppers,Murray Fletcher for the assistance with the morphologicalidentification of planthoppers, Kirsten Scott, Corinna Langeand Glenn Graham for some of the DNA sequences,and Audrey Anderson for assistance with the transmissionexperiments.
References
Anon. (1979) ‘79th annual report. 1979.’ (Bureau of Sugar ExperimentStations: Brisbane)
Anon. (2003) ‘Annual review 2003.’ (Australian Sugar Milling Council:Brisbane)
Arneodo JD, Guzman FA, Conci LR, Laguna IG, Truol GA (2002)Transmission features of Mal de Rıo Cuarto virus in wheat by itsplanthopper vector Delphacodes kuscheli. The Annals of AppliedBiology 141, 195–200.
Assis Filho FMD, Deom CM, Sherwood JL (2004) Acquisitionof Tomato spotted wilt virus by adults of two thrips species.Phytopathology 94, 333–336.
Beebe NW, Maung J, van den Hurk AF, Ellis JT, Cooper RD (2001)Ribosomal DNA spacer genotypes of the Anopheles bancroftiigroup (Diptera: Culicidae) from Australia and Papua New Guinea.Insect Molecular Biology 10, 407–413. doi: 10.1046/j.0962-1075.2001.00278.x
Bosio CF, Fulton RE, Salasek ML, Beaty BJ, Black WC (2000)Quantitative trait loci that control vector competence for dengue-2 virus in the mosquito Aedes aegypti. Genetics 156, 687–698.
Bull RM (1977) Leafhopper populations on some sugarcane varieties.Proceedings of the Queensland Society of Sugar Cane Technologists44, 83–87.
Buzacott JH (1953) Insects associated with sugar cane in New Guinea.Technical Communications, Bureau of Sugar Experiment Stations,Queensland 1953, 23–30.
Campbell CL, Mummey DL, Schmidtmann ET, Wilson WC (2004)Culture-independent analysis of midgut microbiota in the arbovirusvector Culicodes sonorensis (Diptera: Ceratopogonidae). Journal ofMedical Entomology 41, 340–348.
Chang VCS (1977) Transovarial transmission of the Fiji disease virus inPerkinsiella saccharicida Kirk. Sugarcane Pathologists’ Newsletter18, 22–23.
Egan BT, Ryan CC, Francki RIB (1989) Fiji disease. In ‘Diseases ofsugarcane. Major diseases’. (Eds C Ricaud, BT Egan, AGJ Gillaspie,CG Hughes) pp. 263–288. (Elsevier: Amsterdam)
Felsenstein J (1985) Confidence limits on phylogenies: An approachusing the bootstrap. Evolution 39, 783–791.
Fennah RG (1979) New species and new records of Perkinsiella(Hemiptera: Delphacidae) from Papua New Guinea. Bulletin ofEntomological Research 69, 507–517.
Fernando LCP, Walter GH (1997) Species status of two-associatedpopulations of Aphytis lingnanensis (Hymenoptera: Aphelinidae)in citus. Bulletin of Entomological Reseach 87, 137–144.
FitzGibbon F, Allsopp PG, DeBarro PJ (1998). ‘Sugarcane exoticpests—pest risk analysis database.’ (Bureau of Sugar ExperimentStations: Brisbane)
FitzGibbon F, Allsopp PG, DeBarro PJ (1999) Chomping, boring andsucking on our doorstep – the menace from the north. Proceedingsof the Australian Society of Sugar Cane Technologists 21,149–155.
Fletcher MJ, Lariviere M-C (2003) ‘Identification keys andchecklists for the leafhoppers, planthoppers and theirrelatives occurring in Australia and New Zealand (Hemiptera:Auchenorrhyncha).’ Available at http://www.agric.nsw.gov.au/Hort/ascu/start.htm (verified 31 January 2006)
Francki RIB, Grivell CJ (1972) Occurence of similar particles on Fijidisease virus-infected sugar cane and insect vector cells. Virology48, 305–307. doi: 10.1016/0042-6822(72)90145-6
Francki RIB, Ryan CC, Hatta T, Rohozinski J, Grivell CJ (1986)Serological detection of Fiji disease virus antigens in the planthopperPerkinsiella saccharicida and its inefficient ability to transmit thevirus. Plant Pathology 35, 324–328.
Hall TA (1999) BioEdit: a user-friendly biological sequence alignmenteditor and analysis program for Windows 95/98/NT. Nucleic AcidsSymposium Series 41, 95–98.
Hughes CG, Robinson PE (1961) Fiji disease. In ‘Sugar-cane diseasesof the world’. (Eds JP Martin, EV Abbott, CG Hughes) pp. 389–405.(Elsevier: Amsterdam)
Hussain AAA, Brown AHD, Hutchinson PB, Wismer CA (1965)The testing of sugarcane varieties for resistance to Fiji diseasein Fiji. Proceedings of the International Society of Sugar CaneTechnologists 12, 1154–1164.

112 Australasian Plant Pathology A. W. Ridley et al.
Ikeda K, Nagaoka S, Winkler S, Kotani K, Yagi H, et al.(2001) Molecular characterization of Bombyx mori cytoplasmicpolyhedrosis virus genome segment 4. Journal of Virology 75,988–995. doi: 10.1128/JVI.75.2.988-995.2001
Knox TB, Kay BH, Hall RA, Ryan PA (2003) Enhanced vectorcompetence of Aedes aegypti (Diptera: Culicidae) from the TorresStrait compared with mainland Australia for dengue2 and 4 viruses.Journal of Medical Entomology 40, 950–956.
Liu H, Beckenbach AT (1992) Evolution of the mitochondrialcytochrome oxidase II gene among ten orders of insects. MolecularPhylogenetics and Evolution 1, 41–52. doi: 10.1016/1055-7903(92)90034-E
Lockhart PJ, Steel MA, Hendy MD, Penny D (1994) Recoveringevolutionary trees under a more realistic model of sequenceevolution. Molecular Biology and Evolution 11, 605–612.
McQualter R, Smith GR, Dale JL, Harding RM (2003) Molecularanalysis of Fiji disease Fijivirus genome segments 1 and 3. VirusGenes 26, 283–289. doi: 10.1023/A:1024403431373
Madden LV, Jeger MJ, van den Bosch F (2000) A theoretical assessmentof the effects of vector-virus transmission mechanism on plant virusdisease epidemics. Phytopathology 90, 576–594.
Magarey RC, Suma S, Irawan, Kuniata LS, Allsopp PG (2002) Sikna binatang bilong suka – diseases and pests encountered during asurvey of Saccharum germplasm ‘in the wild’ in Papua New Guinea.Proceedings of the Australian Society of Sugar Cane Technologists24, 219–227.
Mertens P, Arella M, Attoui H, Belloncik S, Bergoin M, et al. (2000)Reoviridae. In ‘Virus taxonomy: classification and nomenclature ofviruses: 7th report of the International Committee on Taxonomy ofViruses’. (Eds MHV Van Regenmortel, CM Fauquet, DJ McGeogh,CR Pringle, RB Wickner) pp. 395–480. (Academic Press:San Diego)
Miller SA, Dykes DD, Polesky HF (1998) A simple salting out procedurefor extracting DNA from human nucleated cells. Nucleic AcidsResearch 16, 1215.
Milne RC, Lovisolo O (1977) Maize rough dwarf and related viruses.Advances in Virus Research 21, 267–341.
Moritz G, Kumm S, Mound L (2004) Tospovirus transmissiondepends on thrips ontogeny. Virus Research 100, 143–149.doi: 10.1016/j.virusres.2003.12.022
Mungomery RW (1946) Report of the Division of Entomology andpathology. In ‘46th annual report of the Bureau of Sugar ExperimentStations’. pp. 31–38. (Bureau of Sugar Experiment Stations:Brisbane)
Mungomery RW, Bell AF (1933) Fiji disease of sugar-cane and itstransmission. Bulletin, Division of Pathology, Bureau of SugarExperiment Stations 4, 1–28.
Nault LR (1997) Arthropod transmission of plant viruses: A newsynthesis. Annals of the Entomological Society of America 90,521–541.
Ocfemia GO (1934) An insect vector of the Fiji disease of sugarcane.American Journal of Botany 21, 113–120.
Osborn AW, Forteath GNR (1972) Transmission of Fiji diseaseby Perkinsiella saccharicida Kirkaldy collected from northernQueensland. Journal of the Australian Entomological Society 11,344–346.
Paterson HEH (1991) The recognition of cryptic species amongeconomically important insects. In ‘Heliothis: methods andprospects’. (Ed. MP Zalucki) pp. 1–10. (Springer-Verlag: New York)
Ryan CC (1988) Epidemiology and control of Fiji disease virus ofsugarcane. Advances in Disease Vector Research 4, 163–176.
Saito N, Nei M (1987) The neighbour-joining method: a new method forreconstructing phylogenetic trees. Molecular Biology and Evolution4, 406–452.
Shikata E, Kitagawa Y (1977) Rice black-streaked dwarf virus: itsproperties, morphology and intracellular localization. Virology 77,826–842. doi: 10.1016/0042-6822(77)90502-5
Sinha RC (1963) Effect of age of vector and of abdomen puncture onvirus transmission. Phytopathology 53, 1170–1173.
Smith GR (2000) Fiji disease. In ‘A guide to sugarcane diseases’.(Eds P Rott, RA Bailey, JC Comstock, BJ Croft, A Salem Saumtally)pp. 239–244. (CIRAD and ISSCT: Montpellier)
Smith GR, Candy JM (2004) Improving Fiji disease resistancescreening trials in sugarcane by considering virus transmission classand possible origin of Fiji disease virus. Australian Journal ofAgricultural Research 55, 665–672. doi: 10.1071/AR03241
Smith GR, Van de Velde R (1994) Detection of sugarcane mosaic virusand Fiji disease virus in diseased sugarcane using the polymerasechain reaction. Plant Disease 78, 557–561.
Steel M (1994) Recovering a tree from the Markov leaf colourations itgenerates under a Markov model. Applied Mathematics Letters 7,19–23. doi: 10.1016/0893-9659(94)90024-8
Swofford DL (2002) ‘PAUP*. Phylogenetic analysis using parsimony(*and other methods), version 4.0.’ (Sinauer Associates:Sunderland)
Wang Z, Zhou Y, Fan Y, Cheng Z, Zhang W (2002) Rapid methodsfor the detection of rice black streaked dwarf Fijivirus in a singleplanthopper. Journal of Shanghai Jiaotong University 20, 340–343.
Received 16 March 2005, accepted 25 July 2005
http://www.publish.csiro.au/journals/app




















