
Influence of two different cover crops on soil N availability, N nutritional status, and grape...
14
REGULAR ARTICLE Influence of two different cover crops on soil N availability, N nutritional status, and grape yeast-assimilable N (YAN) in a cv. Tempranillo vineyard Eva P. Pérez-Álvarez & Teresa Garde-Cerdán & Pilar Santamaría & Enrique García-Escudero & Fernando Peregrina Received: 9 July 2014 /Accepted: 14 January 2015 # Springer International Publishing Switzerland 2015 Abstract Aim The aim was to study the effects of two different cover crops (leguminous versus gramineous) on soil NO 3 - -N availability, grapevine N status, and yeast- assimilable N (YAN) and amino acid profile in the must of a cv. Tempranillo vineyard from La Rioja (Spain) over 3 years. Methods Three treatments were established: gramine- ous (barley) and leguminous (Persian clover) cover crops and conventional tillage. Soil total N and extract- able NH 4 + -N at bloom, soil NO 3 - -N content from budbreak to postharvest, leaf N content at bloom, and free amino acids and ammonia in must were monitored. Results Each year, the barley cover crop treatment had lower soil NO 3 - -N than tillage, while the clover treatment showed lower soil NO 3 - -N than tillage in 2009 and showed higher values in 2010 and 2011. In 2011 leaf N content, yeast- assimilable N (YAN), and some amino acids in the grapes were lower in barley cover crop treat- ment thus affecting the amino acid profile. Mean- while, the clover cover crop did not have these effects. Furthermore, relationships between leaf N content and various amino acids were also found. Conclusions With the barley cover crop treatment, soil N availability decreased from the first year of the exper- iment, provoking a decrease in N grapevine status and in content of YAN and various amino acids at the third year. In contrast, clover cover crop maintained N grape- vine status, YAN, and amino acid profile similar to conventional tillage. Therefore, our study shows how soil N can be managed both within a year and from year to year when two different cover crops are sown and how the grapevine N nutritional status and YAN content and amino acid profile can be modified in the must. Keywords Grape amino acids . Leaf nitrogen content . Soilnitrogenavailability . Mediterraneanvineyard . Wine quality Abbreviations YAN yeast-assimilable nitrogen AOC appellation d’origine contrôlée Introduction Tillage is the common soil management method used in Mediterranean vineyards. Various authors have shown that cover crops decrease the soil N availability in Med- iterranean vineyards (Steenwerth and Belina 2008; Celette et al. 2009; Peregrina et al. 2012; Pérez-Álvarez et al. 2013a). The reduction of soil N availability caused Plant Soil DOI 10.1007/s11104-015-2387-7 Responsible Editor: Euan K. James. E. P. Pérez-Álvarez (*) : T. Garde-Cerdán : P. Santamaría : E. García-Escudero : F. Peregrina Instituto de Ciencias de la Vid y del Vino, ICVV (Gobierno de La Rioja-Universidad de La Rioja-CSIC). Servicio de Investigación y Desarrollo Tecnológico Agroalimentario (CIDA), Ctra. Logroño-Mendavia NA-134 Km. 90, 26071 Logroño, La Rioja, Spain e-mail: [email protected]
Transcript of Influence of two different cover crops on soil N availability, N nutritional status, and grape...

REGULAR ARTICLE
Influence of two different cover crops on soil N availability, Nnutritional status, and grape yeast-assimilable N (YAN) in a cv.Tempranillo vineyard
Eva P. Pérez-Álvarez & Teresa Garde-Cerdán &
Pilar Santamaría & Enrique García-Escudero &
Fernando Peregrina
Received: 9 July 2014 /Accepted: 14 January 2015# Springer International Publishing Switzerland 2015
AbstractAim The aim was to study the effects of two differentcover crops (leguminous versus gramineous) on soilNO3
−-N availability, grapevine N status, and yeast-assimilable N (YAN) and amino acid profile in the mustof a cv. Tempranillo vineyard from La Rioja (Spain)over 3 years.Methods Three treatments were established: gramine-ous (barley) and leguminous (Persian clover) covercrops and conventional tillage. Soil total N and extract-able NH4
+-N at bloom, soil NO3−-N content from
budbreak to postharvest, leaf N content at bloom, andfree amino acids and ammonia in must were monitored.Results Each year, the barley cover crop treatmenthad lower soil NO3
−-N than tillage, while theclover treatment showed lower soil NO3
−-N thantillage in 2009 and showed higher values in 2010and 2011. In 2011 leaf N content, yeast-assimilable N (YAN), and some amino acids inthe grapes were lower in barley cover crop treat-ment thus affecting the amino acid profile. Mean-while, the clover cover crop did not have these
effects. Furthermore, relationships between leaf Ncontent and various amino acids were also found.Conclusions With the barley cover crop treatment, soilN availability decreased from the first year of the exper-iment, provoking a decrease in N grapevine status and incontent of YAN and various amino acids at the thirdyear. In contrast, clover cover crop maintained N grape-vine status, YAN, and amino acid profile similar toconventional tillage. Therefore, our study shows howsoil N can be managed both within a year and from yearto year when two different cover crops are sown andhow the grapevine N nutritional status and YAN contentand amino acid profile can be modified in the must.
Keywords Grape amino acids . Leaf nitrogen content .
Soilnitrogenavailability .Mediterraneanvineyard .Winequality
AbbreviationsYAN yeast-assimilable nitrogenAOC appellation d’origine contrôlée
Introduction
Tillage is the common soil management method used inMediterranean vineyards. Various authors have shownthat cover crops decrease the soil N availability in Med-iterranean vineyards (Steenwerth and Belina 2008;Celette et al. 2009; Peregrina et al. 2012; Pérez-Álvarezet al. 2013a). The reduction of soil N availability caused
Plant SoilDOI 10.1007/s11104-015-2387-7
Responsible Editor: Euan K. James.
E. P. Pérez-Álvarez (*) : T. Garde-Cerdán : P. Santamaría :E. García-Escudero : F. PeregrinaInstituto de Ciencias de la Vid y del Vino, ICVV (Gobierno deLa Rioja-Universidad de La Rioja-CSIC). Servicio deInvestigación y Desarrollo Tecnológico Agroalimentario(CIDA), Ctra. Logroño-Mendavia NA-134 Km. 90,26071 Logroño, La Rioja, Spaine-mail: [email protected]

by the cover crops has potential negative effects ongrapevines such as limited vine vigor and yield reduc-tion (Tesic et al. 2007; Ripoche et al. 2011), but theseeffects can vary according to soil fertility, grapevinecultivar, and the cover crop species. However, thesepotentially negative effects can be judged as beneficialin fertile soils where vines have a high vigor. In thosecases, the cover crop can reduce the excessive grapevinevegetative growth that is unfavorable to high-qualitywine production. Moreover, a moderate N status invines reduces the potential for fermentation haze orconditions that can lead to excess thiol formation (Belland Henschke 2005).
In the appellation d’origine contrôlée (AOC) Riojaarea, which hasMediterranean climatic conditions, mostvineyards use tillage for soil-surface management, andthe norm of this appellation limits the red grape yield to6500 kg ha−1. In addition, the soils of La Riojavineyards have a low organic matter content (<1 %)and a loamy texture (Peregrina et al. 2010a), and con-sequently, they are highly susceptible to erosion. Recentstudies indicate that the use of permanent cover crops inAOC Rioja vineyard can increase both organic carbonand stable aggregates in the soil thus improving soilquality (Peregrina et al. 2010b, 2014). Also, cover cropscan decrease the soil N availability (Peregrina et al.2012; Pérez-Álvarez et al. 2013a). Therefore, in theAOC Rioja, cover crops can be an alternative to tillagein order to improve soil quality and also when areduction in vigor and an improvement in the mustquality are needed.
Must quality is influenced by nitrogen compositionwhich affects fermentation kinetics and the productionof ethanol, glycerol, and both aromatic and spoilagecompounds (Albers et al. 1996). Few reports exist onspecific relationships between individual free aminoacids and wine volatiles, but such associations are anarea of active research (Ugliano et al. 2009). The twomain sources of yeast-assimilable nitrogen (YAN) areprimary amino acids and ammonium (Butzke 1998).Consequently, YAN content in grapes can have a sig-nificant impact on the production of many active me-tabolites related to flavor (Hernández-Orte et al. 2005).
Xi et al. (2010) found that in a Cabernet Sauvignonvineyard, cover crops increased the wine aroma com-pounds when compared to tillage. By contrast, Lee andSteenwerth (2011), in a Californian vineyard with sim-ilar Mediterranean climatic conditions, found no signif-icant difference in the YAN composition of Cabernet
Sauvignon grapes under different cover crop manage-ment. However, the different rootstocks used in thatexperiment (110R high vigor, 420A low vigor) playeda significant role in YAN composition.
The reduction of the grapevine N uptake and theconsequent YAN decrease could be positive for winequality when it results in a lower concentration of aminoacids (i.e., histidine) which are precursors of biogenicamines such as histamine. However, if YAN is too low,the fermentation can be negatively affected. When at-mospheric N in the soil is fixed by using a leguminouscover crop, soil N availability and N content in the mustcan increase (Fourie et al. 2007a). The effects of Napplication on grape amino acid composition have beenstudied previously by Conradie (2001) and Linsenmeiret al. (2008). However, little research has been doneconcerning the different effects of gramineous and legu-minous cover crops on soil N availability, N grapevinestatus, and YAN grape composition in cv. Tempranillowhich is the main cultivar in the AOC Riojawinegrowing area. We hypothesized that a nonlegumi-nous cover crop could compete for soil N and decreaseN uptake by the grapevine and consequently affect thegrape YAN content and the production of aromatic andspoilage compounds during fermentation.
The objective of this study was to investigate theinfluence of two different cover crops, namely a gra-mineous [barley (Hordeum vulgare L.)] and a legumi-nous [Persian clover (Trifolium resupinatum L.)] crop,in the midterm (1 to 3 years after cover crop establish-ment) on soil N availability, N grapevine status, andYAN grape content in a highly vigorous cv. Tempranillovineyard in the AOC Rioja.
Materials and methods
Experimental design and site
The experiment began in 2009 in an experimental farmin Nájera, on a terrace of the Najerilla river (468 m ofelevation above sea level), in La Rioja region (northernSpain). Coordinates for the site were lat. 42° 26′ 34″ N;long. 2° 43′ 32″ W. The field slope was about 0.2 %facing west. The soil was classified as OxyaquicXerorthent (according to USDA Soil Taxonomy). Ad-ditional details for soil characteristics are shown inTable 1. The climate in the area is semiarid accordingto the UNESCO aridity index.
Plant Soil
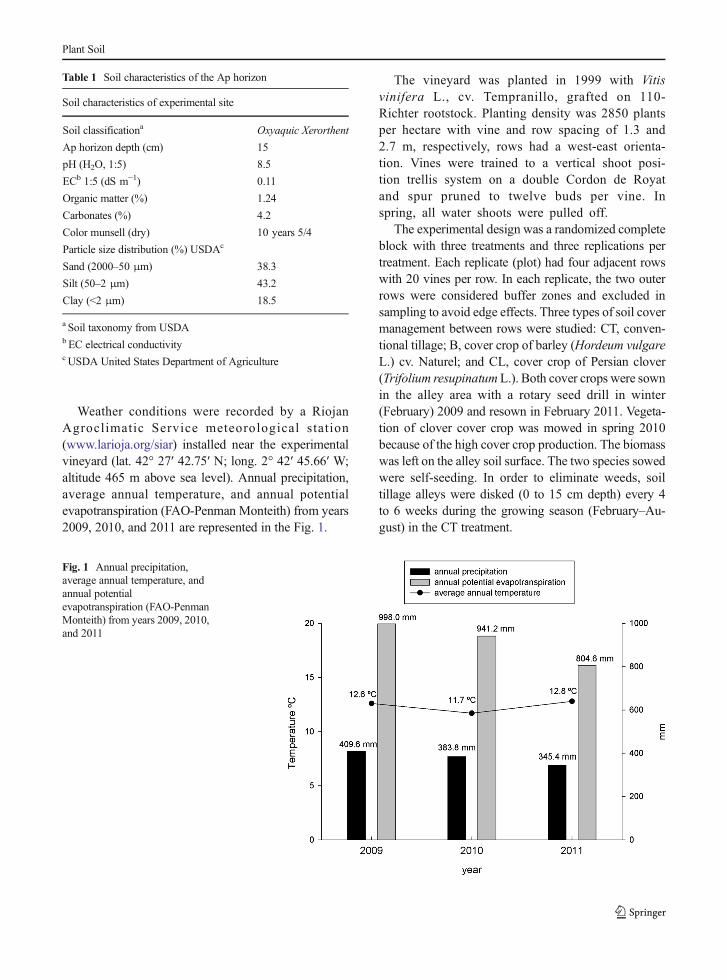
Weather conditions were recorded by a RiojanAgroclimatic Service meteorological station(www.larioja.org/siar) installed near the experimentalvineyard (lat. 42° 27′ 42.75′ N; long. 2° 42′ 45.66′ W;altitude 465 m above sea level). Annual precipitation,average annual temperature, and annual potentialevapotranspiration (FAO-Penman Monteith) from years2009, 2010, and 2011 are represented in the Fig. 1.
The vineyard was planted in 1999 with Vitisvinifera L., cv. Tempranillo, grafted on 110-Richter rootstock. Planting density was 2850 plantsper hectare with vine and row spacing of 1.3 and2.7 m, respectively, rows had a west-east orienta-tion. Vines were trained to a vertical shoot posi-tion trellis system on a double Cordon de Royatand spur pruned to twelve buds per vine. Inspring, all water shoots were pulled off.
The experimental design was a randomized completeblock with three treatments and three replications pertreatment. Each replicate (plot) had four adjacent rowswith 20 vines per row. In each replicate, the two outerrows were considered buffer zones and excluded insampling to avoid edge effects. Three types of soil covermanagement between rows were studied: CT, conven-tional tillage; B, cover crop of barley (Hordeum vulgareL.) cv. Naturel; and CL, cover crop of Persian clover(Trifolium resupinatumL.). Both cover crops were sownin the alley area with a rotary seed drill in winter(February) 2009 and resown in February 2011. Vegeta-tion of clover cover crop was mowed in spring 2010because of the high cover crop production. The biomasswas left on the alley soil surface. The two species sowedwere self-seeding. In order to eliminate weeds, soiltillage alleys were disked (0 to 15 cm depth) every 4to 6 weeks during the growing season (February–Au-gust) in the CT treatment.
Table 1 Soil characteristics of the Ap horizon
Soil characteristics of experimental site
Soil classificationa Oxyaquic Xerorthent
Ap horizon depth (cm) 15
pH (H2O, 1:5) 8.5
ECb 1:5 (dS m−1) 0.11
Organic matter (%) 1.24
Carbonates (%) 4.2
Color munsell (dry) 10 years 5/4
Particle size distribution (%) USDAc
Sand (2000–50 μm) 38.3
Silt (50–2 μm) 43.2
Clay (<2 μm) 18.5
a Soil taxonomy from USDAbEC electrical conductivityc USDA United States Department of Agriculture
Fig. 1 Annual precipitation,average annual temperature, andannual potentialevapotranspiration (FAO-PenmanMonteith) from years 2009, 2010,and 2011
Plant Soil

In all the treatments, a 0.8-m-wide herbicide strip waskept beneath the vines so the cover crop had awidth of 1.9 m.
The experimental vineyard had not been N fertilizedsince 2000 (9 years prior to the experiment starting), sothe N dynamics for cover crops and conventional tillagecould be evaluated. In the three treatments, after prun-ing, canes were shredded and left on the soil surfacebetween rows. The rest of the vineyard managementpractices were similar in all treatments. Grapevine can-opy management and control of vine pests and diseasesby chemical spraying were carried out by machines, andthe crop was manually harvested.
Soil total nitrogen, NH4+-N, and NO3
−-N analysisthroughout the growing season
In each plot, three subsamples were collected randomlyby an auger in the interrow, at 0 to 15 and 15 to 45 cmdepth and bulked to give a composite sample. Fivesamplings were carried out at each growing season; atbudbreak (end-April), bloom (mid-June), fruit set(mid-July), veraison (mid-August), and postharvest(early-November).
The soil samples were air dried and then ground andsieved to 2 mm. The percentage of coarse fragments(>2 mm) was determined in each sample. Soil totalnitrogen (organic N+inorganic N) was determined, insoil samples taken at bloom, by means of a dry com-bustionmethod (Matejovic 1997), using a TruSpec CNSElemental Analyzer (LECO Inc.; St. Joseph, MI, USA).Soil NH4
+-N and soil NO3−-N were extracted from the
soil samples with 2 M KCl and determined by colorim-etry at 660 and 550 nm, respectively, using a SEALAutoAnalyzer 3HR (Seal Analytical; Hamburg, Germa-ny), an elemental autoanalyzer of segmented flow. Inorder to express N content as NO3
−, NH4+, and total N in
kilograms per hectare, soil bulk density determined bythe core method, as well as the soil percentage of coarsefragments (>2 mm) at both 0 to 15 and 15 to 45 cm soildepths were employed.
Determination of leaf nitrogen content
A total of 20 healthy leaves per plot (only one leaf pervine, distributed randomly) were sampled from nodesopposite to the first bunch at bloom (mid-June). Atveraison (mid-August), 20 healthy leaves were alsosampled from nodes opposite to the second bunch.
In each growing season, blades and petioles wereseparated, washed with tap water, and rinsed with dis-tilled water. The plant material sample was dried at 70 °C in a forced-air oven (Selecta Dry-Big; J.P. Selecta,Barcelona, Spain) for 72 h and then ground and sievedto 0.5 mm in an ultracentrifuge mill (ZM1; Retsch,Haan, Germany). Nitrogen concentration in leaf bladesand petioles was determined with an elemental analyzerTruSpec CN (Leco Inc.; St. Joseph, MI, USA). Concen-trations were expressed on a dry weight basis (García-Escudero et al. 2013).
Yield, grape sampling, and grape must
At harvest in 2009, 2010, and 2011, 20 randomly cho-sen vines in each plot were harvested just prior to theAOC Rioja commercial harvest. The grape yield pro-duced per plot and the number of clusters per grapevinewere recorded. At the same time, random samples ofapproximately 500 berries were collected from 20grapevines distributed randomly throughout each plot.In the laboratory, berries were counted, weighed, andcrushed using a Masticator (IUL Instruments; GmbH,Königswinter, Germany) to obtain must which was keptfrozen (−20 °C) until analyses were carried out.
HPLC analysis of free amino acids and YANdetermination
Analysis of amino acids in must was performed accord-ing to the method described by Garde-Cerdán et al.(2014). Briefly, free amino acids (aspartic acid BASP,^glutamic acid BGLU,^ asparagine BASN,^ serineBSER,^ histidine BHIS,^ glycine BGLY,^ threonineBTHR,^ citrulline BCIT,^ arginine BARG,^ alanineBALA,^ tyrosine BTYR,^ valine BVAL,^ methionineBMET,^ tryptophan BTRP,^ phenylalanine BPHE,^ iso-leucine BILE,^ ornitine BORN,^ leucine BLEU,^ lysineBLYS,^ and proline BPRO^) were analyzed by reversephase using a Hewlett Packard Series 1100 LiquidChromatograph (Agilent Technologies; Waldbronn,Germany) equipped with an automatic liquid sampler(ALS), an Agilent 1100 fluorescence detector (FLD),and an ultraviolet diode array detector (DAD). An ali-quot (5 mL) of sample (previously centrifuged,4000 rpm, 10 min) was mixed with 100 μL of norvaline(internal standard to quantify all amino acids exceptPRO) and 100 μL of sarcosine (internal standard toquantify PRO). The mixture was filtered through a
Plant Soil

0.45-μmOlimPeak pore filter (Teknokroma; Barcelona,Spain) and sent to an automatic precolumn derivatiza-tion with o-phthaldialdehyde (OPA Reagent, AgilentTechnologies; Palo Alto, CA, USA) for primary aminoacids determination and with 9-fluorenylmethylchloroformate (FMOC Reagent, Agilent TechnologiesInc.; Palo Alto, CA, USA) for determination of second-ary amino acids. The injected amount from the derivatedsample was 10μL, and temperature was kept constant at40 °C. All separations were performed on a HypersilODS column (250×4.0 mm, I.D. 5 μm) (AgilentTechnologies Inc.; Palo Alto, CA, USA). Solventsand gradient conditions for amino acids analysisare described below.
Two eluents were used as mobile phase: eluent A,75 mM sodium acetate, 0.018 % triethylamine(pH 6.9)+0.3 % tetrahydrofuran; eluent B, water, meth-anol, and acetonitrile (10:45:45, v/v/v). All reagentswere first filtered through Millipore filters (0.45 μm).The gradient profile employed is described in Table 2.Detection was performed by FLD (λ excitation=340 nm, λ emission=450 nm for primary amino acids,and λ exCitation=266 nm, λ emission=305 nm forsecondary amino acids) and DAD (λ=338 nm for pri-mary amino acids and λ=262 nm for secondary aminoacids). Identification of compounds was carried out bycomparing their retention times with those of pure ref-erence standards. The pure reference compounds andinternal standards were from Sigma (Sigma-Aldrich; St.Louis, MO, USA). Water was obtained from a Milli-Qpurification system (Millipore Corporation; Billerica,MA, USA). Quantification of amino acids was per-formed employing norvaline and sarcosine as internalstandards, as mentioned above.
Unit for free amino acid was milligrams N per kilo-gram of whole berries (which will be referred to as mgNkg−1 for conciseness). Total free amino acids (FAN)were determined by summing the individual free aminoacid values.
Yeast-assimilable nitrogen values (mg N kg−1) weredetermined according to the method described by Aerny(1996). Ammonia concentration was calculated by sub-traction from the YAN content, the individual primaryfree amino acids, as recommended by Lee and Schreiner(2010) that can be easily utilized by yeast (all individualfree amino acids excluding HIS and PRO).
Statistical analysis
Treatment effects on measured variables were testedusing ANOVA (univariate linear model), and differ-ences between treatment means were determined usingthe least significant difference (LSD) multiple range testcalculated at P<0.05. The Pearson correlation coeffi-cient (r) was used to assess the relationship between Nconcentration in leaf tissues (blade and petiole) andammonia and free amino acids. Statistical procedureswere carried out with the software program StatgraphicsPlus for Windows (1998) (StatPoint Technologies Inc.;Warrenton, VA, USA).
Results
Cover crops effect on total N, extractable NH4+-N,
NO3−-N availability, leaf N content, and on yield com-
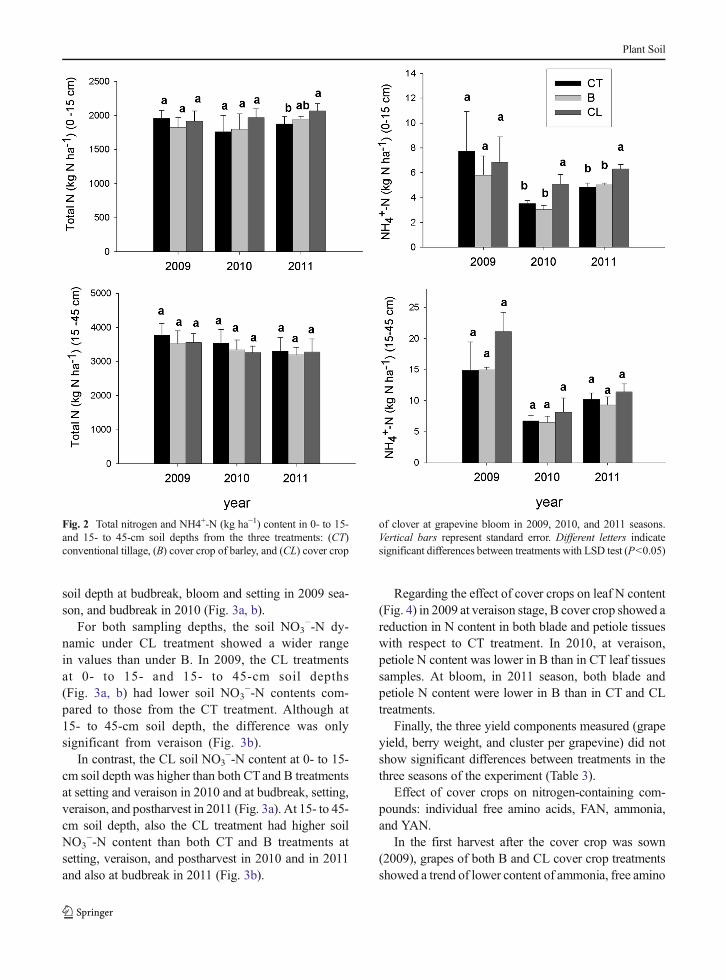
ponents throughout the 3 years monitored.At bloom in 2011, the CL cover crop showed higher
total soil N with respect to the CT treatment. Moreover,the CL cover crop had greater NH4
+-N with respect tothe CT and B treatments in the 0- to 15-cm soil depth in2010 and 2011 seasons (Fig. 2). There were no differ-ences between CT and B treatments for total soil N andNH4
+-N at either depth nor year.From the first soil sampling taken (budbreak, April
2009) to the last (postharvest, November 2011), soilNO3
−-N content in the B treatment was consistentlylower with respect to CT at both 0- to 15- and 15- to45-cm soil depths (Fig. 3a, b), although for the deepersoil layer, differences between treatments were smaller.A few exceptions where differences between treatmentswere not significant occurred for the 0- to 15-cm depthat budbreak and bloom in 2010, and for the 15- to 45-cm
Table 2 Gradient profile of eluent B (water, methanol, and ace-tonitrile in relation 10:45:45, v/v/v), employed in the determinationof amino acids with HPLC
Time (min) Eluent B (%) Flow rate (mL/min)
0 to 15 0 to 47.5 1.63
15 to 15.01 47.5 1.63 to 0.80
15.01 to 25 47.5 to 60 0.80
25 to 25.01 60 0.80 to 1.63
25.01 to 26.01 60 to 100 1.63
26.01 to 26.51 100 1.63 to 2.50
26.51 to 34.01 100 2.50
34.01 to 36.01 100 to 0 1.63
Plant Soil
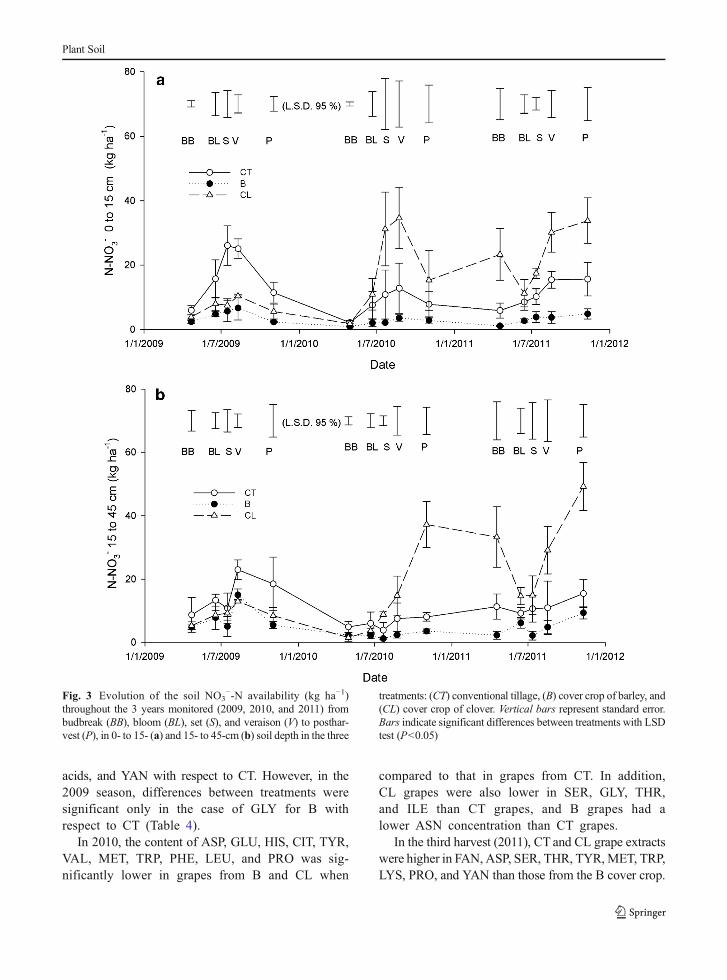
soil depth at budbreak, bloom and setting in 2009 sea-son, and budbreak in 2010 (Fig. 3a, b).
For both sampling depths, the soil NO3−-N dy-
namic under CL treatment showed a wider rangein values than under B. In 2009, the CL treatmentsat 0- to 15- and 15- to 45-cm soil depths(Fig. 3a, b) had lower soil NO3
−-N contents com-pared to those from the CT treatment. Although at15- to 45-cm soil depth, the difference was onlysignificant from veraison (Fig. 3b).
In contrast, the CL soil NO3−-N content at 0- to 15-
cm soil depth was higher than both CTand B treatmentsat setting and veraison in 2010 and at budbreak, setting,veraison, and postharvest in 2011 (Fig. 3a). At 15- to 45-cm soil depth, also the CL treatment had higher soilNO3
−-N content than both CT and B treatments atsetting, veraison, and postharvest in 2010 and in 2011and also at budbreak in 2011 (Fig. 3b).
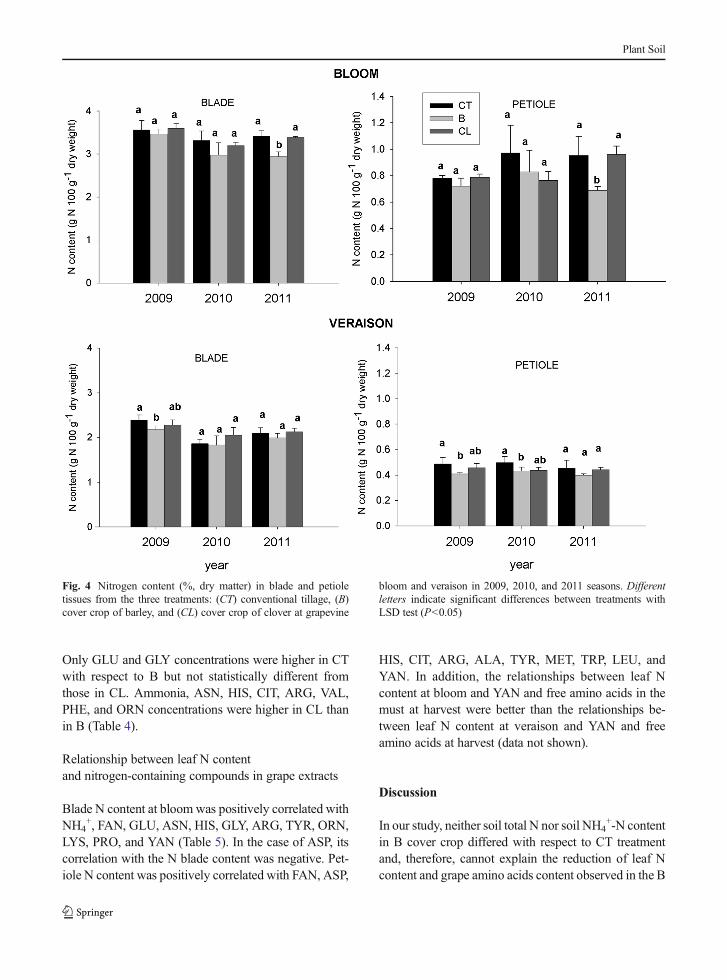
Regarding the effect of cover crops on leaf N content(Fig. 4) in 2009 at veraison stage, B cover crop showed areduction in N content in both blade and petiole tissueswith respect to CT treatment. In 2010, at veraison,petiole N content was lower in B than in CT leaf tissuessamples. At bloom, in 2011 season, both blade andpetiole N content were lower in B than in CT and CLtreatments.
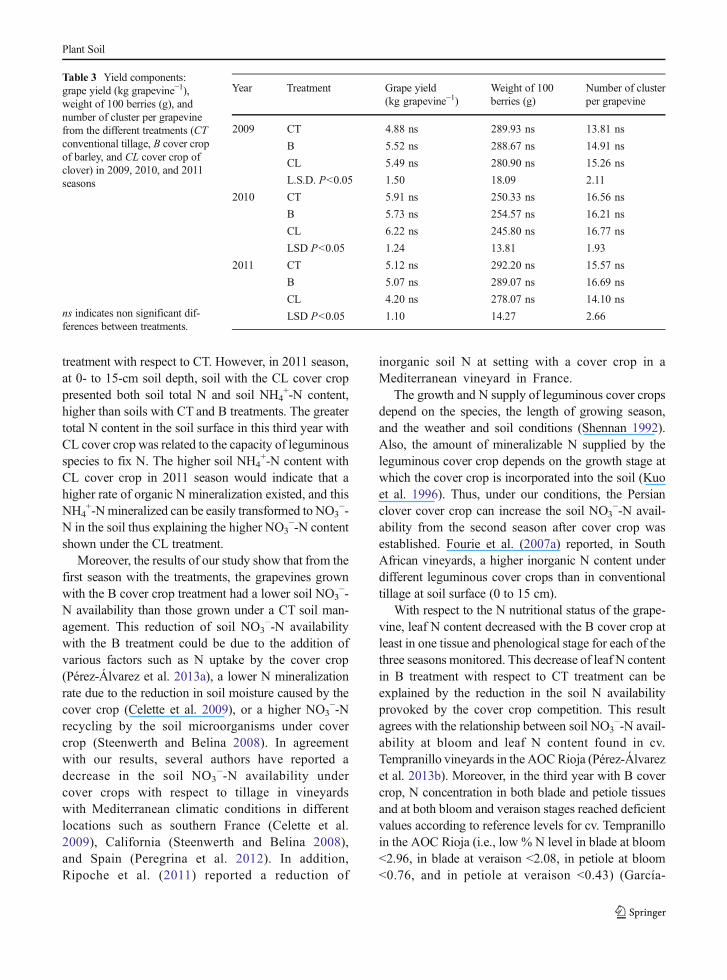
Finally, the three yield components measured (grapeyield, berry weight, and cluster per grapevine) did notshow significant differences between treatments in thethree seasons of the experiment (Table 3).
Effect of cover crops on nitrogen-containing com-pounds: individual free amino acids, FAN, ammonia,and YAN.
In the first harvest after the cover crop was sown(2009), grapes of both B and CL cover crop treatmentsshowed a trend of lower content of ammonia, free amino
Fig. 2 Total nitrogen and NH4+-N (kg ha−1) content in 0- to 15-and 15- to 45-cm soil depths from the three treatments: (CT)conventional tillage, (B) cover crop of barley, and (CL) cover crop
of clover at grapevine bloom in 2009, 2010, and 2011 seasons.Vertical bars represent standard error. Different letters indicatesignificant differences between treatments with LSD test (P<0.05)
Plant Soil
acids, and YAN with respect to CT. However, in the2009 season, differences between treatments weresignificant only in the case of GLY for B withrespect to CT (Table 4).
In 2010, the content of ASP, GLU, HIS, CIT, TYR,VAL, MET, TRP, PHE, LEU, and PRO was sig-nificantly lower in grapes from B and CL when
compared to that in grapes from CT. In addition,CL grapes were also lower in SER, GLY, THR,and ILE than CT grapes, and B grapes had alower ASN concentration than CT grapes.
In the third harvest (2011), CT and CL grape extractswere higher in FAN, ASP, SER, THR, TYR,MET, TRP,LYS, PRO, and YAN than those from the B cover crop.
Fig. 3 Evolution of the soil NO3−-N availability (kg ha−1)
throughout the 3 years monitored (2009, 2010, and 2011) frombudbreak (BB), bloom (BL), set (S), and veraison (V) to posthar-vest (P), in 0- to 15- (a) and 15- to 45-cm (b) soil depth in the three
treatments: (CT) conventional tillage, (B) cover crop of barley, and(CL) cover crop of clover. Vertical bars represent standard error.Bars indicate significant differences between treatments with LSDtest (P<0.05)
Plant Soil
Only GLU and GLY concentrations were higher in CTwith respect to B but not statistically different fromthose in CL. Ammonia, ASN, HIS, CIT, ARG, VAL,PHE, and ORN concentrations were higher in CL thanin B (Table 4).
Relationship between leaf N contentand nitrogen-containing compounds in grape extracts
Blade N content at bloom was positively correlated withNH4
+, FAN, GLU, ASN, HIS, GLY, ARG, TYR, ORN,LYS, PRO, and YAN (Table 5). In the case of ASP, itscorrelation with the N blade content was negative. Pet-iole N content was positively correlated with FAN, ASP,
HIS, CIT, ARG, ALA, TYR, MET, TRP, LEU, andYAN. In addition, the relationships between leaf Ncontent at bloom and YAN and free amino acids in themust at harvest were better than the relationships be-tween leaf N content at veraison and YAN and freeamino acids at harvest (data not shown).
Discussion
In our study, neither soil total N nor soil NH4+-N content
in B cover crop differed with respect to CT treatmentand, therefore, cannot explain the reduction of leaf Ncontent and grape amino acids content observed in the B
Fig. 4 Nitrogen content (%, dry matter) in blade and petioletissues from the three treatments: (CT) conventional tillage, (B)cover crop of barley, and (CL) cover crop of clover at grapevine
bloom and veraison in 2009, 2010, and 2011 seasons. Differentletters indicate significant differences between treatments withLSD test (P<0.05)
Plant Soil
treatment with respect to CT. However, in 2011 season,at 0- to 15-cm soil depth, soil with the CL cover croppresented both soil total N and soil NH4
+-N content,higher than soils with CT and B treatments. The greatertotal N content in the soil surface in this third year withCL cover crop was related to the capacity of leguminousspecies to fix N. The higher soil NH4
+-N content withCL cover crop in 2011 season would indicate that ahigher rate of organic N mineralization existed, and thisNH4
+-Nmineralized can be easily transformed to NO3−-
N in the soil thus explaining the higher NO3−-N content
shown under the CL treatment.Moreover, the results of our study show that from the
first season with the treatments, the grapevines grownwith the B cover crop treatment had a lower soil NO3
−-N availability than those grown under a CT soil man-agement. This reduction of soil NO3
−-N availabilitywith the B treatment could be due to the addition ofvarious factors such as N uptake by the cover crop(Pérez-Álvarez et al. 2013a), a lower N mineralizationrate due to the reduction in soil moisture caused by thecover crop (Celette et al. 2009), or a higher NO3
−-Nrecycling by the soil microorganisms under covercrop (Steenwerth and Belina 2008). In agreementwith our results, several authors have reported adecrease in the soil NO3
−-N availability undercover crops with respect to tillage in vineyardswith Mediterranean climatic conditions in differentlocations such as southern France (Celette et al.2009), California (Steenwerth and Belina 2008),and Spain (Peregrina et al. 2012). In addition,Ripoche et al. (2011) reported a reduction of
inorganic soil N at setting with a cover crop in aMediterranean vineyard in France.
The growth and N supply of leguminous cover cropsdepend on the species, the length of growing season,and the weather and soil conditions (Shennan 1992).Also, the amount of mineralizable N supplied by theleguminous cover crop depends on the growth stage atwhich the cover crop is incorporated into the soil (Kuoet al. 1996). Thus, under our conditions, the Persianclover cover crop can increase the soil NO3
−-N avail-ability from the second season after cover crop wasestablished. Fourie et al. (2007a) reported, in SouthAfrican vineyards, a higher inorganic N content underdifferent leguminous cover crops than in conventionaltillage at soil surface (0 to 15 cm).
With respect to the N nutritional status of the grape-vine, leaf N content decreased with the B cover crop atleast in one tissue and phenological stage for each of thethree seasons monitored. This decrease of leaf N contentin B treatment with respect to CT treatment can beexplained by the reduction in the soil N availabilityprovoked by the cover crop competition. This resultagrees with the relationship between soil NO3
−-N avail-ability at bloom and leaf N content found in cv.Tempranillo vineyards in the AOCRioja (Pérez-Álvarezet al. 2013b). Moreover, in the third year with B covercrop, N concentration in both blade and petiole tissuesand at both bloom and veraison stages reached deficientvalues according to reference levels for cv. Tempranilloin the AOC Rioja (i.e., low % N level in blade at bloom<2.96, in blade at veraison <2.08, in petiole at bloom<0.76, and in petiole at veraison <0.43) (García-
Table 3 Yield components:grape yield (kg grapevine−1),weight of 100 berries (g), andnumber of cluster per grapevinefrom the different treatments (CTconventional tillage, B cover cropof barley, and CL cover crop ofclover) in 2009, 2010, and 2011seasons
ns indicates non significant dif-ferences between treatments.
Year Treatment Grape yield(kg grapevine−1)
Weight of 100berries (g)
Number of clusterper grapevine
2009 CT 4.88 ns 289.93 ns 13.81 ns
B 5.52 ns 288.67 ns 14.91 ns
CL 5.49 ns 280.90 ns 15.26 ns
L.S.D. P<0.05 1.50 18.09 2.11
2010 CT 5.91 ns 250.33 ns 16.56 ns
B 5.73 ns 254.57 ns 16.21 ns
CL 6.22 ns 245.80 ns 16.77 ns
LSD P<0.05 1.24 13.81 1.93
2011 CT 5.12 ns 292.20 ns 15.57 ns
B 5.07 ns 289.07 ns 16.69 ns
CL 4.20 ns 278.07 ns 14.10 ns
LSD P<0.05 1.10 14.27 2.66
Plant Soil

Tab
le4
Ammonia,freeam
inoacid,and
yeast-assimilableN(m
gkg
−1)ingrapesextractsfrom
thedifferenttreatments(CTconventio
naltillage,Bcovercropofbarley,and
CLcovercropof
clover)in
2009,2010,and2011
seasons
2009
2010
2011
(mgN
kg−1)
CT
BCL
LSD
,P<0.05
CT
BCL
LSD,P
<0.05
CT
BCL
LSD,P
<0.05
Ammonia
26.75nsw
21.37ns
26.ns
18.29
28.48ns
25.48ns
22.37ns
14.23
16.13ab
7.50b
20.79a
9.38
FAN
89.6
ns79.02ns
86.33ns
17.36
106.99
ns76.84ns
83.16ns
11.03
76.95a
58.41b
79.29a
13.84
ASP
0.70ab
0.77a
0.63b
0.11
2.04a
1.65b
1.70b
0.16
1.28a
1.11b
1.32a
0.06
GLU
4.15
ns3.36
ns4.04
ns0.80
3.61a
3.00c
3.22b
0.22
2.72a
2.21b
2.56ab
0.40
ASN
1.16
ns0.87
ns1.06
ns0.52
1.11a
0.69b
0.84ab
0.36
0.43ab
0.31b
0.44a
0.12
SER
3.67
ns3.38
ns3.54
ns0.60
4.37a
3.98ab
3.69b
0.48
2.29a
1.82b
2.36a
0.40
HIS
5.00
ns3.45
ns4.99
ns2.51
6.24a
1.95b
3.59b
2.40
4.12ab
2.24b
4.29a
2.04
GLY
0.88a
0.60b
0.70ab
0.22
0.68a
0.52ab
0.51b
0.17
0.33a
0.22b
0.31ab
0.10
THR
2.86
ns2.79
ns2.80
ns0.74
3.49a
3.06ab
2.89b
0.52
1.96a
1.43b
2.04a
0.40
CIT
1.06
ns0.87
ns0.87
ns0.61
1.63a
0.84b
0.93b
0.42
0.92ab
0.61b
0.98a
0.33
ARG
26.41ns
22.78ns
25.26ns
4.59
25.97ns
21.92ns
20.92ns
5.58
26.65ab
19.90b
28.28a
8.06
ALA
5.11
ns4.67
ns4.89
ns0.71
5.26
ns4.99
ns4.67
ns0.69
6.64
ns5.60
ns6.91
ns1.51
TYR
0.41
ns0.39
ns0.40
ns0.15
0.49a
0.33b
0.33b
0.09
0.42a
0.28b
0.46a
0.13
VAL
1.65
ns1.87
ns1.72
ns0.38
3.26a
2.38b
2.21b
0.56
1.03ab
0.87b
1.16a
0.27
MET
0.33
ns0.30
ns0.34
ns0.11
0.60a
0.42b
0.36b
0.12
0.31a
0.18b
0.31a
0.12
TRP
2.21
ns2.19
ns2.21
ns0.42
4.18a
3.57b
3.57b
0.32
2.49a
1.92b
2.65a
0.46
PHE
0.43
ns0.44
ns0.44
ns0.13
0.92a
0.67b
0.60b
0.17
0.33ab
0.28b
0.37a
0.07
ILE
0.42
ns0.44
ns0.41
ns0.08
0.92a
0.77ab
0.66b
0.23
0.41
ns0.38
ns0.49
ns0.15
ORN
0.19
ns0.15
ns0.18
ns0.18
0.17
ns0.10
ns1.00
ns0.13
0.18ab
0.14b
0.23a
0.07
LEU
0.71
ns0.66
ns0.69
ns0.19
1.41a
0.99b
0.87b
0.32
0.83
ns0.61
ns0.87
ns0.30
LYS
0.25
ns0.26
ns0.24
ns0.13
0.16
ns0.15
ns0.12
ns0.08
0.26a
0.18b
0.27a
0.06
PRO
32.09ns
28.76ns
30.78ns
7.04
40.47a
24.86c
31.43b
4.94
23.36a
18.14b
23.01a
3.04
YAN
84.34ns
71.71ns
82.04ns
25.86
95.00ns
78.49ns
74.09ns
23.51
69.73a
47.77b
77.07a
10.22
nsindicatesnonsignificantd
ifferences
betweentreatm
ents.
FANfree
aminoacidsnitrogen,A
SPasparticacid,G
LUglutam
icacid,A
SNasparagine,SERserine,H
IShistidine,GLY
glycine,THRthreonine,CIT
citrullin
e,ARGarginine,A
LAalanine,
TYR
tyrosine,V
ALvalin
e,METmethionine,TRPtryptophan,P
HEphenylalanine,ILEisoleucine,O
RNornithine,LEUleucine,LY
Slysine,P
ROproline,YA
Nyeast-assimilablenitrogen
wIn
thesamerow,different
lettersam
ongtreatm
entsexpresssignificantd
ifferences
atP<0.05
usingtheLSD
test
Plant Soil

Escudero et al. 2013). However, this effect did not occurfor CT and CL treatments. Therefore, we consider that aN-deficient status existed in grapevines from B treat-ment at 2011. These results could be due to the fact thatcultural practices did not affect to the grapevine reservesat short term as showed Holzapfel and Smith (2012).Under B cover crop, the grapevine could employ the Nstored as reserves in its tissues (trunk and roots) in thefirst year, maintaining its N nutritional status; but after3 years with a lower soil N availability, the N reserves inthe grapevine could not compensate for this deficit, andthus, differences were found in the leaf N content withrespect to the CT. Tesic et al. (2007) also found lowerpetiole N content at bloom under cover cropwith respectto bare soil in cv. Chardonnay, and Sweet and Schreiner(2010) reported a lower N concentration in blade atbloom under resident vegetation with respect to tillageand a cover crop of a clover mixture in cv. Pinot noir.
Results of the grapevines under CL cover cropshowed a leaf N content similar to those under CTtreatment. The lower soil NO3
−-N availability showedduring the first season by the CL cover crop with respectto CT treatment was compensated by the ability of coverto fix N. The increase of soil NO3
−-N availability withrespect to CT started in the second season with the CLtreatment, provoking that leaf N content in CL reachedvalues similar to those of the CT treatment.
In contrast to our results, Fourie et al. (2007b) foundthat petiole NO3
−-N content at bloom in Chardonnayand Sauvignon blanc cultivars under a broad leaf le-gume cover crop increased with respect to a tilled soil.This different result could be due to the fact that in theexperiment carried out by Fourie et al. (2007b),the vineyard was irrigated while in our experiment,no irrigation was applied to the plots. In ourconditions, water competition of cover crops couldresult in the fact that soil moisture at soil surfaceunder CL cover crop was lower than that underthe CT treatment, as occurred in a vineyard withno irrigation under Mediterranean climatic condi-tions where cover crops reduce the soil moisture inthe surface layer of soil (Celette et al. 2008). Sowhen soil moisture is too low, soil N may not beavailable to the grapevine (González-Dugo et al.2010) and this effect could explain the differencesbetween our results with respect to those fromFourie et al. (2007b).
Regarding the yield components, both cover cropshad no effect on any yield components, indicating thatT
able5
Pearson
correlationcoefficientsandits
Pvalues
betweenleafN(%
N)tissues(blade
andpetio
leatbloom)and
nitrogen-containing(m
gNkg
−1)com
pounds
(NH4+,FAN,Y
AN,and
aminoacids)in
grapeextractsthroughout
the3yearsmonito
redin
thetreatm
ents(CTconventio
naltillage,Bcovercrop
ofbarley,and
CLcovercrop
ofclover)(n=27)
Bloom
NH4+
FAN
ASP
GLU
ASN
SER
HIS
GLY
THR
CIT
ARG
Blade
(%N)
0.4092*
0.4884**
−0.4601*
0.5272**
0.5653**
0.1927
0.5898**
0.5511**
0.2748
0.3726
0.5965**
Petio
le(%
N)
0.3587
0.4599*
0.4058*
−0.0201
0.1131
0.1308
0.4930*
0.0315
0.2086
0.5328**
0.6139**
ALA
TYR
VAL
MET
TRP
PHE
ILE
ORN
LEU
LYS
PRO
YAN
Blade
(%N)
0.1175
0.5710**
−0.0696
0.0972
−0.2294
−0.0982
−0.3661
0.5230**
−0.1421
0.6112**
0.4054*
0.5840**
Petio
le(%
N)
0.5515**
0.5987**
0.1323
0.4689*
0.4068*
0.3126
0.2093
0.2793
0.4508*
0.1979
0.2211
0.5510**
*Significant
correlationatP<0.05;*
*significant
correlationatP<0.01
FANfree
aminoacidsnitrogen,A
SPasparticacid,G
LUglutam
icacid,A
SNasparagine,SERserine,H
IShistidine,GLY
glycine,THRthreonine,CIT
citrullin
e,ARGarginine,A
LAalanine,
TYR
tyrosine,V
ALvalin
e,METmethionine,TRPtryptophan,P
HEphenylalanine,ILEisoleucine,O
RNornithine,LEUleucine,LY
Slysine,P
ROproline,YA
Nyeast-assimilablenitrogen
Plant Soil

differences found in the composition of N-containingcompounds in the grapes, would not be attributable toprocesses of concentration by reducing grape produc-tion or berry weight. In agreement with our results,Ingels et al. (2005) in California and Lopes et al.(2008) in Portugal found that the use of cover crop assoil management did not reduce grapevine yield.
In addition, under our experimental conditions, FANand YAN decreased under B cover crop in the thirdseason, and positive relationships between FAN andYAN with the leaf N content in grapevine at bloomwere found. In agreement with our results, Gaudillereet al. (2003) found correlations between blade N contentand N in must in vineyards under Atlantic andMediterranean climates in France, and Neilsen et al.(2010) described correlations between petiole-N atveraison and yeast-assimilable N in cv. Merlot underdifferent rate, timing, and form of N fertilization. Also,N concentration in must increased under a leguminouscover crop and decreased under a grain cover crop bothwith respect to conventional tillage in cultivars Char-donnay and Sauvignon blanc (Fourie et al. 2007b).Besides this, Lee and Schreiner (2010) found that incv. Pinot noir, lower N supplies reduced the YAN con-tent in grapes. Low YAN content in must could result insluggish fermentation and could produce residual sugar(Spayd et al. 1995). Nitrogen concentrations from 70 to270mgNL−1 YAN are reported to be adequate to insuregood fermentation (Bell and Henschke 2005). There-fore, in our experimental vineyard where CT had a YANconcentration near to the aforementioned minimum, thedecrease of YAN concentration in the third season underthe B cover crop could be undesirable for an adequatefermentation. From these results, studies could bedeveloped for a practical application in thevineyards, such as applying N fertilizer afterbloom with fertilizer rates calculated in functionto the leaf N content at this phenological stage,with the objective to insure an adequate level ofN-containing compounds in the must at harvest.
With respect to the grape amino acids profile in thedifferent treatments, we found a total of 20 free aminoacids with PRO and ARG being dominant. In our re-sults, the amino acid profile was modified by soil man-agement with the cover crops because 15 amino acidshad positive correlations with leaf N (blade or petiole)content at bloom. However, the N content in both bladeand petiole tissues at bloom did not correlate with thesame group of amino acids (i.e., GLU, ASN, GLY,
ORN, LYS, and PRO only was correlated with blade,and CIT, ALA, MET, TRP, and LEU only was correlat-ed with petiole). Therefore, blade or petiole tissue couldbe used depending on the specific amino acids to studyin grapes. Recently, Romero et al. (2013) reported thatblade, at both bloom and veraison stages, was the mostsuitable tissue to N nutrition diagnosis in the cv.Tempranillo in AOC Rioja. However, according to ourresults, the petiole should also be considered for thediagnosis of N-containing compounds (such as aminoacids) in the grapes.
Positive relations between the N supply and the freeamino acids concentration (ALA, HIS, GLY and GLU)were reported in cv. Riesling by Linsenmeir et al.(2008). Lee and Schreiner (2010) showed that onlyGLU, GLN, TRP, and HYP were not affected by Nsupply in cv. Pinot noir. The differences in those refer-ences with respect to our results between the free aminoacids that were affected byN availability could be due tothe fact that the amino acid profile and concentration canvary according to grape cultivar, rootstock, site,seasonal conditions, and level of maturity (Belland Henschke 2005).
Finally, the lower soil N availability and consequent-ly, the lower N status in the grapevine under B covercrop provoked lower concentrations of ASP, GLU,SER, GLY, THR, ARG, TYR, MET, TRP, LYS, andPRO in grapes from the B cover crop with respect to CTafter three seasons. This result could have relevant im-plications on wine quality since HIS, TYR, ORN, LYS,PHE, ARG, and TRP are precursors, by microbial de-carboxylation, of the biogenic amines histamine, tyra-mine, putrescine, cadaverine, 2-phenethylamine,agmatine, and tryptamine, respectively (Vincenziniet al. 2009). Therefore, the use of a B cover crop couldreduce the concentration of biogenic amines and im-prove the wine quality.
In contrast, after three seasons, the CL cover crop hadsimilar concentrations of free amino acids in grape withrespect to CT. This result is in agreement with the factthat B treatment decreased both the leaf N content andthe concentration of various free amino acids. In thisway, as the grapevine N status in the CL treatment didnot vary with respect to CT, the amino acid concentra-tions were, consequently, not affected.
Therefore, under our experimental conditions, the CLcover crop did not show any negative implicationslinked to a higher potential production of biogenicamines in the wine.
Plant Soil

Conclusions
Nitrogen availability as total N or extractable NH4+-N
showed a low sensitivity to the different soil manage-ment practices, and therefore, those parameters cannotexplain the changes in the grapevine N nutritional statusnor in the concentration of both YAN and free aminoacids in the grapes.
However, the two different cover crops studied mod-ify the soil N availability measured as NO3
−-N, althoughthey did so differently. The barley cover reduced theavailability of NO3
−-N in the soil with respect to con-ventional tillage from the first year in which it wasestablished; whereas, with respect to conventional till-age, clover cover reduced NO3
−-N when it wasestablished, but increases it afterwards.
These changes in the soil N availability affected thevine nutritional status and also the concentration of bothYAN and free amino acids in the grapes, which candirectly affect the biogenic amines biosynthetic path-way. Nevertheless, these changes show a delay (twoseasons) with respect to the observed change in the soilN availability.
YAN reduction under the barley cover crops cancause defective alcoholic fermentation of musts. How-ever, the lower concentration of amino acids which areprecursors of biogenic amines could improve the poten-tial quality of the wine. Meanwhile, the clover cover didnot have these effects because YAN and FAN concen-trations were similar to the ones found in the conven-tional tillage treatment.
Therefore, our study offers new knowledge showingthe implications of cover crops management. A gramin-eous cover crop such as barley can be suitable to reducethe N grapevine status and both YAN and freeamino acids in the grapes, although this effectwould occur in the midterm (3 years), but not inthe short term (1 year).
Acknowledgments This study was supported by the SpanishMinistry of Science, INIA (Instituto Nacional de Investigación yTecnología Agraria y Alimentaria) through project INIA-RTA2009-00101-00-00 and European Social Fund. F. Peregrina andT. Garde-Cerdán thank the INIA and European Social Fund fortheir postdoctoral grants and E. Pérez-Álvarez thanks the INIA forher predoctoral grant.
We particularly thank Ricardo Leza for lending us the vineyardin which we installed the experimental plots and for helping withthe field labor. Also, we thank Rosa López Martín (ICVV-CIDA,Government of La Rioja) for her help with the analysis method-ology of amino acids in grapes.
References
Aerny J (1996) Composés azotés desmoûts et des vins. Rev SuisseVitic Arboric Hortic 28:161–165
Albers E, Larsson C, Liden G, Niklasson C, Gustafsson L (1996)Influence of the nitrogen source on Saccharomycescerevisiae anaerobic growth and product formation. ApplEnviron Microbiol 62:3187–3195
Bell S, Henschke P (2005) Implications of nitrogen nutrition forgrapes, fermentation and wine. Aust J Grape Wine Res 11:242–295
Butzke CE (1998) Survey of yeast assimilable nitrogen status inmusts from California, Oregon and Washington. Researchnote. Am J Enol Vitic 49:220–224
Celette F, Gaudin R, Gary C (2008) Spatial and temporal changesto the water regime of a Mediterranean vineyard due to theadoption of cover cropping. Eur J Agron 29:153–162
Celette F, Findeling A, Gary C (2009) Competition for nitrogen inan unfertilized intercropping system: the case of an associa-tion of grapevine and grass cover in a Mediterranean climate.Eur J Agron 30:41–51
Conradie WJ (2001) Timing of nitrogen fertilisation and the effectof poultry manure on the performance of grapevines on sandysoil. I. Soil analysis, grape yield and vegetative growth. S AfrJ Enol Vitic 22:52–68
Fourie JC, Agenbag GA, Louw PJE (2007a) Cover crop manage-ment in a Chardonnay/99 Richter vineyard in the coastalregion, South Africa. 3. Effect of different cover crops andcover crop management practices on organic matter andmacro-nutrient content of a medium-textured soil. S Afr JEnol Vitic 28:61–68
Fourie JC, Louw PJE, Agenbag GA (2007b) Cover crop manage-ment in a Sauvignon blanc/Ramsey vineyard in the Semi-aridolifants River Valley, South Africa. 2. Effect of differentcover crops and cover crop management practices on grape-vine performance. S Afr J Enol Vitic 28:81–91
García-Escudero E, Romero I, Benito A, Domínguez N, Martín I(2013) Reference levels for leaf nutrient diagnosis of cv.Tempranillo Grapevine in the Rioja Appellation. CommunSoil Sci Plant Anal 44:645–654
Garde-Cerdán T, López R, Portu J, González-Arenzana L, López-Alfaro I, Santamaría P (2014) Study of the effects of proline,phenylalanine, and urea foliar application to Tempranillovineyards on grape amino acid content. Comparison withcommercial nitrogen fertilisers. Food Chem 163:136–141
Gaudillere JP, Chantelot E, Soyer JP, Molot C, Milin S(2003) Leaf and must nitrogen content two comple-mentary indicators of grapevine nitrogen status. J IntSci Vigne Vin 37:91–101
González-Dugo V, Durand JL, Gastal F (2010) Water deficit andnitrogen nutrition of crops. A review. Agron Sustain Dev 30:529–544
Hernández-Orte P, Ibarz MJ, Cacho J, Ferreira V (2005) Effect ofthe addition of ammonium and amino acids to must of Airenvariety on aromatic composition and sensory properties ofthe obtained wines. Food Chem 89:163–174
Holzapfel B, Smith J (2012) Developmental stage and cli-matic factors impact more on carbohydrate reservedynamics of Shiraz than cultural practice. Am J EnolVitic 63(3):333–342
Plant Soil

Ingels CA, ScowKM,Whisson DS, Drenovsky RE (2005) Effectsof cover crops on grapevines, yield, juice composition, soilmicrobial ecology, and gopher activity. Am J Enol Vitic 56:19–29
Kuo S, Sainju UM, Jellum EJ (1996) Winter cover croppinginfluence on nitrogen mineralization and corn yields. SoilBiol Fertil 22:310–317
Lee J, Schreiner RP (2010) Free amino acid profiles from ‘Pinotnoir’ grapes are influenced by vine N-status and samplepreparation method. Food Chem 119:484–489
Lee J, Steenwerth KL (2011) Rootstock and vineyard floor man-agement influence on ‘Cabernet Sauvignon’ grape yeast as-similable nitrogen (YAN). Food Chem 127:926–933
Linsenmeir AW, Loos U, Löhnertz O (2008) Must compositionand nitrogen uptake in a long-term trial as affected by timingof nitrogen fertilization in a cool-climate Riesling vineyard.Am J Enol Vitic 59:255–265
Lopes CM, Monteiro A, Machado JP, Fernandes N, Araújo A(2008) Cover cropping in a sloping non-irrigated vineyard.II. Effects on vegetative growth, yield, berry and wine qualityof Cabernet Sauvignon grapevines. Ciência Téc Vitic23:37–43
Matejovic I (1997) Determination of carbon and nitrogen in sam-ples of various soils by the dry combustion. Commun SoilSci Plant Anal 28:17–18
Neilsen GH, Neilsen D, Bowen P, Bogdanoff C, Usher K (2010)Effect of timing, rate, and form of N fertilization on nutrition,vigor, yield, and berry yeast-assimilable N of grape. Am JEnol Vitic 61:327–336
Peregrina F, López D, Zaballa O, Villar MT, González G, García-Escudero E (2010a) Soil quality of vineyards in the origindenomination Rioja: index of overcrusting risk (FAO-PNUMA), content of organic carbon and relation with soilfertility. Rev Ciênc Agrárias 33:338–345
Peregrina F, Larrieta C, Ibáñez S, García-Escudero E (2010b)Labile organic matter, aggregates, and stratification ratios ina semiarid vineyard with cover crops. Soil Sci Soc Am J 74:2120–2130
Peregrina F, Pérez-Álvarez EP, Colina M, García-Escudero E(2012) Cover crop and tillage influence soil organic matterand nitrogen availability in a semi-arid vineyard. Arch AgronSoil Sci 58:S95–S102
Peregrina F, Pérez-Álvarez EP, García-Escudero E (2014) Soilmicrobiological properties and its stratification ratios for soilquality assessment under different cover crop managementsystems in a semiarid vineyard. J Plant Nutr Soil Sci 177:548–559
Pérez-Álvarez EP, Pérez-Sotés JL, García-Escudero E, Peregrina F(2013a) Cover crop short-term effects on soil NO3
−-N avail-ability, nitrogen nutritional status, yield, and must quality in acalcareous vineyard of the AOC Rioja, Spain. Commun SoilSci Plant Anal 44:711–721
Pérez-Álvarez EP, Martínez-Vidaurre JM, Martín I, García-Escudero E, Peregrina F (2013b) Relationships among soilnitrate nitrogen and nitrogen nutritional status, yield compo-nents, and must quality in semi-arid vineyards from RiojaAOC, Spain. Commun Soil Sci Plant Anal 44:232–242
Ripoche A, Metay A, Celette F, Gary C (2011) Changing the soilsurface management in vineyards: immediate and delayedeffects on the growth and yield of grapevine. Plant Soil 339:259–271
Romero I, Garcia-Escudero E, Martin I (2013) Leaf blade versuspetiole analysis for nutritional diagnosis of Vitis vinifera L.cv. Tempranillo. Am J Enol Vitic 64:50–64
Shennan C (1992) Cover crops, nitrogen cycling, and soil proper-ties in semi-irrigated vegetable production systems. HortSci27:749–754
Spayd SE, Nagel CW, Edwards CG (1995) Yeast growth inRiesling juice as affected by vineyard nitrogen fertilization.Am J Enol Vitic 55:246–252
Steenwerth K, Belina KM (2008) Cover crops and cultivation:impacts on soil N dynamics andmicrobiological function in aMediterranean vineyard agroecosystem. Appl Soil Ecol 40:370–380
Sweet RM, Schreiner RP (2010) Alleyway cover crops have littleinfluence on Pinot noir grapevines (Vitis vinifera L.) in twowestern Oregon vineyards. Am J Enol Vitic 61:240–252
Tesic D, Keller M, Hutton RJ (2007) Influence of vineyard floormanagement practices on grapevine vegetative growth, yield,and fruit composition. Am J Enol Vitic 58:1–11
Ugliano M, Fedrizzi B, Siebert T, Travis B, Magno F, Versini Get al (2009) Effect of nitrogen supplementation andSaccharomyces species on hydrogen sulphide and other vol-atile sulphur compounds in Shiraz fermentation and wine. JAgric Food Chem 57:4948–4955
Vincenzini M, Guerrini S, Mangani S, Granchi L (2009) Aminoacid metabolisms and production of biogenic amines andethyl carbamate. In: Konig H, Unden G, Frohlch J (eds)Biology of microorganisms on grapes, in must and in wine.Springer, Berlin, pp 167–176
Xi ZM, Zhang ZW, Cheng YF, Li H (2010) The effect of vineyardcover crop on main monomeric phenols of grape berry andwine in Vitis vinifera L cv. Cabernet sauvignon. Agric SciChina 9:440–448
Plant Soil




















