
In vivo neutralization of anti-A and successful transfusion of A antigenincompatible red blood cells...
12
IMMUNOHEMATOLOGY In vivo neutralization of anti-A and successful transfusion of A antigen–incompatible red blood cells in an animal modelCaroline Oliver, Deborah Blake, and Stephen Henry BACKGROUND: Our aim was to determine if the his- torical principle of Lewis glycolipid neutralization of anti- body and subsequent Lewis-incompatible transfusion could be extended and applied to the ABO blood group system using synthetic glycolipid-like constructs. STUDY DESIGN AND METHODS: In vitro experiments with human blood and blood group A function-spacer- lipid constructs (FSL-A) were used to determine rates and concentrations that caused antigen transformation and anti-A neutralization. FSL-A constructs were intra- venously infused into naive and anti-A–immunized mice to determine in vivo antigen transformation, anti-A inhi- bition, and tolerance to A antigen–incompatible transfu- sions (A+biotin kodecytes). RESULTS: FSL-A was able to cause in vivo transfor- mation of circulating mouse cells into A antigen–positive cells (in vivo A kodecytes) without consequence in animals either with or without circulating anti-A. FSL-A was able to neutralize circulating anti-A and allow for successful transfusion of incompatible A kodecytes. In the absence of FSL-A neutralization incompatible cells were rapidly destroyed. CONCLUSIONS: FSL constructs have the potential to neutralize circulating antibodies and allow for, or miti- gate, the consequences of ABO-incompatible red blood cell transfusion. T he majority of the population has antibodies directed against ABO antigens and accidental incompatible transfusions, although rare, still do occur. 1 The reaction to transfused ABO- incompatible blood can range from no observable reac- tion through to renal failure, shock, and disseminated intravascular coagulation causing death. 1,2 The outcome is not usually fatal, although the death rate is higher if more than 50 mL are transfused. 1,2 If an ABO-incompatible red blood cell (RBC) transfusion occurs usually prompt clini- cal intervention, particularly discontinuing the transfu- sion, will minimize morbidity and mortality. 1 Clinical management usually includes prompt recognition, pre- venting hypotension, promoting urine output, improving renal blood flow, and vasopressive medication. 1 No therapy to halt further antibody-antigen–mediated destruction is currently used. Not all transfused incom- patible blood will be rapidly and intravascularly hemo- lyzed, because the incompatible transfusion itself effectively reduces antibody and complement levels thereby temporarily mitigating the reaction. 2,3 Antibody can also induce antigen loss from RBCs, 4,5 and some cells are more robust. 6 The deliberate use of ABO-incompatible blood trans- fusions to reduce antibody titers as a preparative treat- ment for marrow transplants has been safely achieved in patients with normal renal function, 7,8 but it is not an alternative transfusion regime. ABO antigen destroying enzymes or antigen masking systems have been proposed as possible future options to allow transfusion across the ABO barrier. As yet these techniques are not in current use. 9,10 In contrast, deliberately transplanting across the ABO barrier has to some extent been achieved by actively reducing the level of ABO antibodies (as reviewed 11 ). Historically, Lewis-incompatible RBCs have been deliberately transfused without adverse consequences into patients with complement-binding hemolytic anti- bodies. It was a reported practice in the past century to transfuse Lewis-positive plasma into recipients with Lewis antibodies and then safely follow this with incompatible Lewis-positive blood. 12,13 The underlying principle was that the Lewis substances present in the plasma were able to neutralize the circulating Lewis antibodies and thus ABBREVIATION: FSL = function-spacer-lipid KODE technology construct. From the Biotechnology Research Institute, AUT University, Auckland, New Zealand. Address correspondence to: Steve Henry, Biotechnology Research Institute, AUT University, Private Bag 92006, Auckland 1142, New Zealand; e-mail: [email protected]. Received for publication December 12, 2010; revision received March 17, 2011, and accepted March 24, 2011. doi: 10.1111/j.1537-2995.2011.03184.x TRANSFUSION 2011;51:2664-2675. 2664 TRANSFUSION Volume 51, December 2011
Transcript of In vivo neutralization of anti-A and successful transfusion of A antigenincompatible red blood cells...

I M M U N O H E M A T O L O G Y
In vivo neutralization of anti-A and successful transfusion of Aantigen–incompatible red blood cells in an animal model_3184 2664..2675
Caroline Oliver, Deborah Blake, and Stephen Henry
BACKGROUND: Our aim was to determine if the his-torical principle of Lewis glycolipid neutralization of anti-body and subsequent Lewis-incompatible transfusioncould be extended and applied to the ABO blood groupsystem using synthetic glycolipid-like constructs.STUDY DESIGN AND METHODS: In vitro experimentswith human blood and blood group A function-spacer-lipid constructs (FSL-A) were used to determine ratesand concentrations that caused antigen transformationand anti-A neutralization. FSL-A constructs were intra-venously infused into naive and anti-A–immunized miceto determine in vivo antigen transformation, anti-A inhi-bition, and tolerance to A antigen–incompatible transfu-sions (A+biotin kodecytes).RESULTS: FSL-A was able to cause in vivo transfor-mation of circulating mouse cells into A antigen–positivecells (in vivo A kodecytes) without consequence inanimals either with or without circulating anti-A. FSL-Awas able to neutralize circulating anti-A and allow forsuccessful transfusion of incompatible A kodecytes. Inthe absence of FSL-A neutralization incompatible cellswere rapidly destroyed.CONCLUSIONS: FSL constructs have the potential toneutralize circulating antibodies and allow for, or miti-gate, the consequences of ABO-incompatible red bloodcell transfusion.
The majority of the population has antibodiesdirected against ABO antigens and accidentalincompatible transfusions, although rare, stilldo occur.1 The reaction to transfused ABO-
incompatible blood can range from no observable reac-tion through to renal failure, shock, and disseminatedintravascular coagulation causing death.1,2 The outcome isnot usually fatal, although the death rate is higher if morethan 50 mL are transfused.1,2 If an ABO-incompatible redblood cell (RBC) transfusion occurs usually prompt clini-cal intervention, particularly discontinuing the transfu-sion, will minimize morbidity and mortality.1 Clinicalmanagement usually includes prompt recognition, pre-venting hypotension, promoting urine output, improvingrenal blood flow, and vasopressive medication.1 Notherapy to halt further antibody-antigen–mediateddestruction is currently used. Not all transfused incom-patible blood will be rapidly and intravascularly hemo-lyzed, because the incompatible transfusion itselfeffectively reduces antibody and complement levelsthereby temporarily mitigating the reaction.2,3 Antibodycan also induce antigen loss from RBCs,4,5 and some cellsare more robust.6
The deliberate use of ABO-incompatible blood trans-fusions to reduce antibody titers as a preparative treat-ment for marrow transplants has been safely achieved inpatients with normal renal function,7,8 but it is not analternative transfusion regime. ABO antigen destroyingenzymes or antigen masking systems have been proposedas possible future options to allow transfusion across theABO barrier. As yet these techniques are not in currentuse.9,10 In contrast, deliberately transplanting across theABO barrier has to some extent been achieved by activelyreducing the level of ABO antibodies (as reviewed11).
Historically, Lewis-incompatible RBCs have beendeliberately transfused without adverse consequencesinto patients with complement-binding hemolytic anti-bodies. It was a reported practice in the past century totransfuse Lewis-positive plasma into recipients with Lewisantibodies and then safely follow this with incompatibleLewis-positive blood.12,13 The underlying principle wasthat the Lewis substances present in the plasma were ableto neutralize the circulating Lewis antibodies and thus
ABBREVIATION: FSL = function-spacer-lipid KODE technology
construct.
From the Biotechnology Research Institute, AUT University,
Auckland, New Zealand.
Address correspondence to: Steve Henry, Biotechnology
Research Institute, AUT University, Private Bag 92006, Auckland
1142, New Zealand; e-mail: [email protected].
Received for publication December 12, 2010; revision
received March 17, 2011, and accepted March 24, 2011.
doi: 10.1111/j.1537-2995.2011.03184.x
TRANSFUSION 2011;51:2664-2675.
2664 TRANSFUSION Volume 51, December 2011

allow Lewis-positive blood to be immediately tolerated.The subsequent elution of the Lewis antigens from thetransfused cells allowed for their prolonged survival.12
Recently a technology known as KODE technology, whichutilizes synthetic glycolipid-like constructs, has been usedto attach blood group antigens onto RBCs,14,15 inhibitviruses and toxins,16 and to determine in vivo cell sur-vival.17 The possibility that these glycolipid-like constructscould also be used to inhibit circulating ABO antibodiesand allow for deliberate transfusion of ABO-incompatibleblood (in a mouse model) is described.
MATERIALS AND METHODS
FSL constructsThree function-spacer-lipid (FSL) constructs wereobtained from KODE Biotech Materials Ltd (Auckland,New Zealand; http://www.kodebiotech.com): FSL-Awith a blood group A trisaccharide epitopeGalNAca3(Fuca2)Galb (FSL-A(GALNa3[Fa2]GALb)-SA1-L1, item 421604), FSL-B with a blood group B trisaccharideepitope Gala3(Fuca2)Galb (FSL-B(GAL3[Fa2]GALb)-SA1-L1, item 199283), and FSL-biotin with a single biotin Fmoiety (FSL-CONJ(1Biotin)-SC2-L1, item 187786). Theseconstructs are also available from Sigma-Aldrich (St Louis,MO) and have been previously used to determine in vivocell survival.17 The process of inserting an FSL into a cellmembrane is called transformation, while the modifiedcell is termed a kodecyte if the construct is detectable.
Serology and human blood samplesUnless stated otherwise cell suspensions and FSL con-struct dilutions were made in antibiotic-containing cellstorage phosphate-buffered saline (PBS; Celpresol, CSL,Melbourne, Australia)—hereafter referred to as PBS. Serol-ogy was in the Diamed-ID Micro Typing System, withLISS/antiglobulin test (low-ionic-strength saline withanti-immunoglobulin [Ig]G plus anti-C3d) and NaCl(saline only) cards (Diamed AG, Cressier-sur-Morat, Swit-zerland) and used according to the manufacturer instruc-tions and scoring system. Tables report score numbers assymbols for visual clarity, for example, 3+ = +++. Titers areexpressed as the reciprocal of dilution. Blood group Aantigen was detected in saline gel cards with IgM mono-clonal anti-A (Epiclone CSL, Melbourne, Australia)—titer1024 versus A1 RBCs. The presence [+] or absence [–] ofanti-A in mice was determined using a serologically vali-dated solid phase A antigen enzyme-linked immunosor-bent assay (ELISA) technique and an anti-IgM plus IgGplus IgA enzyme conjugate.18
Three human samples (volunteers) were used as asource of RBCs and plasma or serum. Because only bloodgroup A incompatibility was studied, antibody activity
against A antigen is defined simply as anti-A, which in thecase of the O serum also encompasses anti-A,B. Anti-Atiters of the group O and B samples were determinedagainst group A1 RBC suspensions at 37°C—with directsaline agglutination classified as simply agglutinin activity(expected to be predominantly IgM but with some contri-bution from IgG) and indirect agglutination with anti-IgG/C3d as IgG/C3d activity (only if activity was greater thanthat observed in saline direct agglutination reactions). Thegroup O sample was a “high titer IgG” anti-A, with activityof 1024, while the B sample had an IgG anti-A titer of 256.Both O and B samples had agglutinin anti-A titers of 64.
In vitro formation of human A kodecytesWashed group O human RBCs (200 mL) were mixed withan equal volume of either blood group O serum, A serum,or PBS, each of which had been spiked with decreasingconcentrations of FSL-A (final concentration range, 4.0-0.025 mg/mL). Reactants were mixed and incubated at37°C. At time points 0.1, 1, 2, 4, and 8 hours, 50 mL wasremoved from each master dilution and immediatelywashed three times with PBS. Samples were prepared ascell suspensions and tested in parallel against monoclonalanti-A.
In vitro antibody sensitization of A kodecytesduring transformationUsing the same method as above in vitro kodecytes wereprepared in group O serum spiked with 4.0, 2.0, 1.0, and0 mg/mL FSL-A. Antigen transformation was confirmedwith monoclonal anti-A. Sensitization of the A kodecyteswith anti-A, which occurred during transformation, wasmeasured at the various time intervals in saline and anti-IgG plus C3d gel cards, immediately after the washingstep. In addition the washed A kodecytes from each timepoint were subsequently incubated for 15 minutes at 37°Cwith unspiked group O serum to determine their ability tobe sensitized with anti-A present in the untreated Oserum.
FSL-A in vitro inhibition of anti-A activity againstA1 cellsBased on the same 2:7 volume ratio of the FSL-A solutionto plasma later given in vivo to mice, a series of in vitroexperiments with fresh human serum were conducted.FSL-A, FSL-B, and FSL-A plus B solutions each containinga total of 20, 10, 5, 2, 1, and 0.5 mg/mL FSL were preparedin PBS (note: FSL-A plus B has a total of 20 mg/mL as10 mg/mL FSL-A and 10 mg/mL FSL-B). In duplicate,10 mL of each FSL solution was added to 35 mL of eithergroup O or group B serum and immediately followed by45 mL (80% packed cells) of blood group A1 RBCs. Tubeswere incubated at 37°C for 1 hour and a 20-mL sample wastaken from the mixture, examined for evidence of aggluti-
ABO-INCOMPATIBLE TRANSFUSION
Volume 51, December 2011 TRANSFUSION 2665

nation or hemolysis, and then washed four times in PBSand prepared as cell suspensions for serology.
In vitro elution of anti-A from A1 cells with FSL-AAnti-A–sensitized cells were prepared by mixing serialdilutions of group O serum (400 mL) with an equal volumeof washed A1 RBCs and incubating at 37°C for 30 minutes.From this mix 175 mL was transferred into two new tubesto which was added either 35 mL of FSL-A (20 mg/mL togive a final concentration of 4 mg/mL) or PBS. Both sus-pensions were incubated at 37°C with 50-mL samplesremoved (and immediately washed four times) every 30minutes, for 2 hours, and the level of agglutinin and IgG/C3d sensitization was measured.
Animal proceduresC57 black 6 laboratory mice (C57BL/6) at least 6 weeks oldwith implanted RFID devices were used. All procedureswere approved by the University of Auckland AnimalEthics Committee (AEC#C500 and AEC#R629). All infu-sions and transfusions were given within 120 seconds andrespiration rate, behavior, urine output, and color (animalwas placed on white filter paper), and the color of extremi-ties were continuously monitored throughout the proce-dure and for the first 2 hours and then hourly for the next4 hours.
Hemoglobinuria detectionTo establish our ability to detect hemoglobinuria, 40 mL ofhemolyzed (frozen-thawed) RBCs in 100 mL was infusedinto six anesthetized mice. Five mice were euthanized at 5,15, 30, 45, and 60 minutes and bladders were inspected.The sixth mouse was allowed to urinate on the filter paperbefore necropsy.
Statistical analysisStandard deviations and one-way analysis of variance pvalues were calculated using a computer statisticalpackage (SPSS, v17, SPSS, Inc., Cary, NC).
In vitro murine kodecytes for transfusionMouse A+biotin kodecytes for transfusions were preparedby incubating a mixture of 100 mg/mL FSL-A and50 mg/mL of FSL-biotin with 1 volume of washed, hardpacked (approx. 95%) RBCs from the same animal orstrain, and incubated (with intermittent mixing) at 37°Cfor 120 minutes. The resultant kodecytes were washedthree times with sterile saline and stored overnight at 4°Cin PBS (Celpresol, CSL). Kodecytes were washed and sus-pended in sterile, endotoxin-free saline for injection
(DEMO S.A., Athens, Greece) and infused as 60% suspen-sions within 1 hour of washing. FSL-A transformation wasconfirmed by routine serology with monoclonal anti-Awhile biotinylation was determined with avidin AlexaFluor 488 (Invitrogen, Carlsbad, CA) and fluorescentmicroscopy.
FSL-A infusions and A+biotin transfusionsAnti-A was stimulated in some animals by subcutaneousinjection of 4¥ 100-mL volumes of concentrated bloodgroup A Le(a–b+) saliva mixed 1:1 with Titermax Goldadjuvant (TiterMax USA, Norcross, GA) three times atthree weekly intervals. Relative antibody levels of someserum samples were determined from the last dilution(titer range, 1-128) to give a clear positive ELISA result.Anesthetized mice (with anti-A present or absent) weregiven 100 mL of A+biotin kodecyte transfusions and/or200 mL of FSL-A infusions (2:7 volume ratio to serum anddose range of 50 to 200 mg FSL-A/kg body mass) via thesubclavian vein. If both an infusion and a transfusion wereto be given, the FSL-A infusion was given first, followed at2 minutes by the kodecyte transfusion. At 6 minutes aftertransfusion or infusion a blood sample was taken from thesubclavian vein (using new equipment and at a new site).The animal was then surgically closed and monitored. At2-, 8-, 24-, 48-, 72-, and 96-hour intervals, a 50-mL bloodsample was collected into a heparinized capillary tubefrom the tail vein for survival studies.17 To minimizedamage to the tail veins (so later sampling could beachieved) only a single attempt to obtain blood was made,which occasionally resulted in a failure to obtain anadequate sample at a given time point. Plain capillary tubesamples were also collected from selected animals toobtain serum to determine the presence or absence ofanti-A before and after transfusion. All animals wereexsanguinated 1 to 3 weeks after transfusion. Kodecytesurvival was determined by the detection of kodecytes viatheir biotin label (Fig. 1) as previously described.17
Mouse FSL-A toxicityFive mice were given infusions of 200 mg of FSL-A/kgbody mass. Five untreated control animals were testedin parallel. Health and behavior were monitored andrecorded by a veterinarian immediately after infusion for 1hour and at 24 hours, 48 hours, 1 week, and monthly inter-vals. All mice including controls were exsanguinated fornecropsy at 3 months with histologic investigations con-ducted on their organs (liver, spleen, hepatic lymphnodes, lung, kidney). Histologic samples were blinded andperformed independently by Gribbles Veterinary Pathol-ogy Ltd. Blood films were also prepared and Giemsastained, and differentials were conducted. Serum was ana-lyzed for the presence of anti-A.
OLIVER ET AL.
2666 TRANSFUSION Volume 51, December 2011
RESULTS
A two-pronged experimental approach was taken. Thefirst was to generate in vitro data with human samplesthat could be extrapolated to a human setting, while thesecond was to model the process in vivo in mice usingkodecytes. Results from the in vivo experiments wereclustered into similar groups (Clusters I-V) based ontheir experimental design and/or outcome. For theprimary in vitro reactions we generally used a serum-to-RBC ratio of 1:1. As a consequence, the level of in vitroantibody sensitization should relate more to an in vivoscenario, but will be less than that of standard serologicmethods that have a serum-to-RBC ratio of more than20:1.
In vitro formation of human A kodecytes: serumversus PBS
To determine the amount of FSL-A that would probablyinsert into RBCs in vivo, transformation in serum wascompared with PBS (the usual in vitro FSL transformationdiluent). FSL-A in PBS could be shown to have insertedinto RBCs within 6 minutes with concentrations as low as0.10 mg/mL (Table 1). Increasing the incubation time upto 2 hours increased the amount of construct inserted butafter that no further transformation was achieved. Basedon these and other unpublished results, more than 95% ofFSL-A has inserted into the RBCs at 2 hours. Identicalresults with FSL-A in PBS were obtained for mouse RBCs(data not shown).
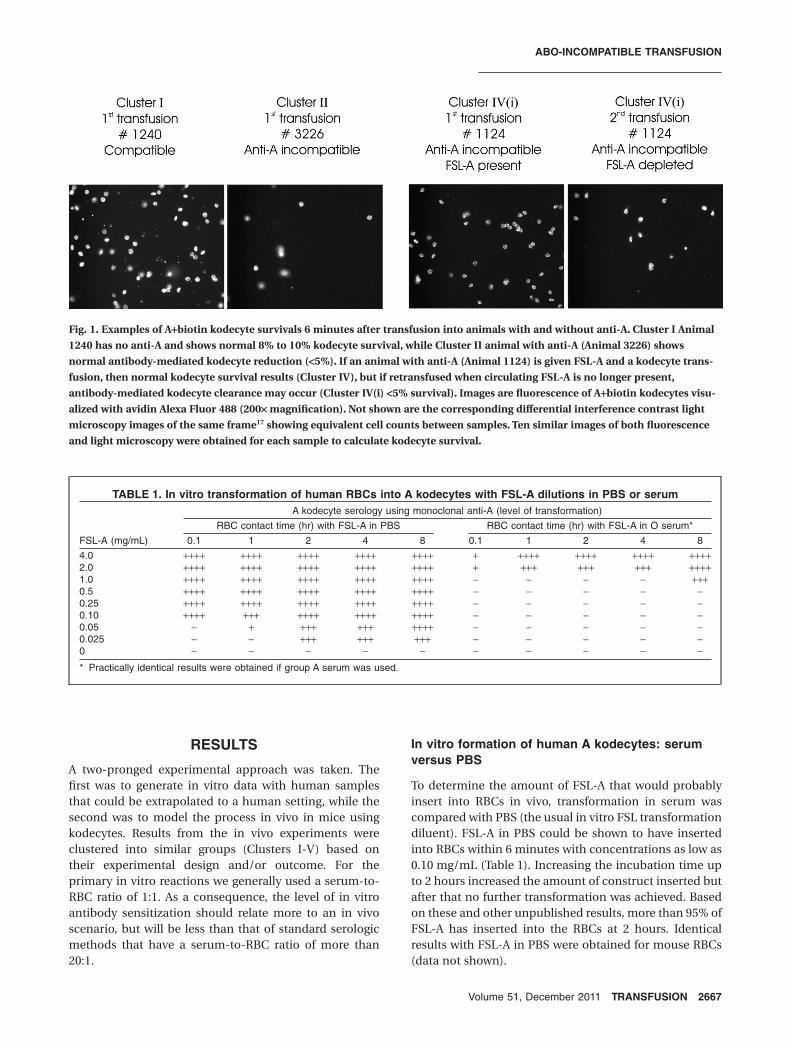
Fig. 1. Examples of A+biotin kodecyte survivals 6 minutes after transfusion into animals with and without anti-A. Cluster I Animal
1240 has no anti-A and shows normal 8% to 10% kodecyte survival, while Cluster II animal with anti-A (Animal 3226) shows
normal antibody-mediated kodecyte reduction (<5%). If an animal with anti-A (Animal 1124) is given FSL-A and a kodecyte trans-
fusion, then normal kodecyte survival results (Cluster IV), but if retransfused when circulating FSL-A is no longer present,
antibody-mediated kodecyte clearance may occur (Cluster IV(i) <5% survival). Images are fluorescence of A+biotin kodecytes visu-
alized with avidin Alexa Fluor 488 (200¥ magnification). Not shown are the corresponding differential interference contrast light
microscopy images of the same frame17 showing equivalent cell counts between samples. Ten similar images of both fluorescence
and light microscopy were obtained for each sample to calculate kodecyte survival.
TABLE 1. In vitro transformation of human RBCs into A kodecytes with FSL-A dilutions in PBS or serum
FSL-A (mg/mL)
A kodecyte serology using monoclonal anti-A (level of transformation)
RBC contact time (hr) with FSL-A in PBS RBC contact time (hr) with FSL-A in O serum*
0.1 1 2 4 8 0.1 1 2 4 8
4.0 ++++ ++++ ++++ ++++ ++++ + ++++ ++++ ++++ ++++2.0 ++++ ++++ ++++ ++++ ++++ + +++ +++ +++ ++++1.0 ++++ ++++ ++++ ++++ ++++ - - - - +++0.5 ++++ ++++ ++++ ++++ ++++ - - - - -0.25 ++++ ++++ ++++ ++++ ++++ - - - - -0.10 ++++ +++ ++++ ++++ ++++ - - - - -0.05 - + +++ +++ ++++ - - - - -0.025 - - +++ +++ +++ - - - - -0 - - - - - - - - - -
* Practically identical results were obtained if group A serum was used.
ABO-INCOMPATIBLE TRANSFUSION
Volume 51, December 2011 TRANSFUSION 2667

The transformation of RBCs with FSL-A in serum wassubstantially less and slower. In contrast to FSL-A in PBS,which at 6 minutes and a concentration of 0.1 mg/mL hadachieved 4+ serology, serum with a 40¥ higher concentra-tion of FSL-A could only achieve a 1+ reaction. At 1 hourFSL-A in serum at the 2.0 and 4.0 mg/mL levels hadresulted in strong A antigen–expressing kodecytes and by8 hours could be seen down to 1.0 mg/mL concentration.
Assuming the FSL-A in serum result achieved with1.0 mg of FSL-A at 8 hours is at completion and equivalentto the 0.025 mg/mL FSL-A in PBS result, then these resultssuggest that only 2.5% of the FSL-A in serum inserts intocells compared with more than 95% when the FSL-A is inPBS.
If the O serum was substituted for A serum then theresults were identical, indicating that the presence ofanti-A does not influence the transformation process(data not shown).
In vitro sensitization of A kodecytesThe possibility that anti-A present in the O serum maysensitize the A kodecytes as they formed was evaluated(Table 2). When FSL-A concentrations in serum weregreater than 1.0 mg/mL, then A kodecytes resulted, inagreement with previous results (Table 1). At all timepoints during the transformation in group O serum (1-8 hr)at no stage could either agglutinins or IgG/C3d be detectedon the A kodecytes. To ensure the kodecytes formed werereactive with the anti-A in the O serum, untreated serum(without FSL-A) was tested against the washed A kode-cytes. Positive serology due to IgG/C3d was present on all A
kodecytes. Despite the IgM monoclonal anti-A reagentreacting with the A kodecytes, the IgM anti-A activity of theuntreated serum (agglutinin titer 64 at 37°C) was negative.This is probably due to the relative low density of FSL-Aantigens on in vitro kodecytes when created with FSL-A inserum (it can be shown that this O serum will react stronglywith in vitro kodecytes if formed in PBS). These resultsshow that IgG anti-A is capable of reacting with the Akodecytes but the high level FSL-A in serum required toinduce the creation of kodecytes was more than adequateto also neutralize the serum IgG anti-A thereby preventingit from interacting with the kodecytes.
In vitro inhibition of anti-AFresh group O and B serum was neutralized with FSL-Asolutions (at an FSL-A solution to serum volume ratio of2:7) to determine the amount of FSL-A that could berequired to prevent polyclonal anti-A activity againstblood group A1 RBCs (Table 3). The treated group Oserum was incubated with packed human A1 cells at aratio of 1:1 (without consideration to the volume contri-bution of the FSL-A) and the level of agglutinin and IgG/C3d sensitization of the RBCs determined. There was noevidence of hemolysis in any tubes. In the absence ofFSL-A (0 mg/mL tube) positive reactivity was observedwith both agglutinin and IgG/C3d serology. FSL-A at allconcentrations was able to completely neutralize all 64titer agglutinin anti-A activity in both the group B and thegroup O sera. FSL-B was able to partially neutralize theagglutinin anti-A, presumably due to neutralization ofsome anti-A,B.
TABLE 2. Anti-A sensitization of human A kodecytes during in vitro formation with FSL-A spiked O serum
FSL-Aserum(mg/mL)
Serologic results obtained after contact hours of FSL-A–spiked group O serum with O RBCs
In vitro kodecyte formation,anti-A (monoclonal)
Sensitization duringtransformation, IgM/IgG/C3d
Indirect sensitization (15 min at 37°C) againstposttransformation A kodecytes
Agglutinin IgG/C3d
1 2 4 8 1, 2, 4, 8 1, 2, 4, 8 1 2 4 8
4.0 ++++ ++++ ++++ ++++ - - ++ ++ ++ +++2.0 ++ +++ ++++ ++++ - - ++ ++ ++ +++1.0 - - ++ ++ - - - + +0 - - - - - - - - -
TABLE 3. In vitro inhibition of human anti-A with FSL-A
ABO serum Serology (initial titer) Neutralizer
FSL concentration (mg/mL serum), serologic activity against A1 cells—human
20 10 5 2 1 0.5 0
B Agglutinin (64) FSL-A - - - - - - ++++Anti-IgG/C3d (256) FSL-A - - - - + + ++++
O Agglutinin (64) FSL-A - - - - - - ++++Anti-IgG/C3d (1024) FSL-A + ++ +++ +++ +++ +++ ++++
O Agglutinin (64) FSL-A + B - - - - - - ++++Anti-IgG/C3d (1024) FSL-A + B ++ ++ +++ +++ +++ ++++ ++++
O Agglutinin (64) FSL-B ++ ++ ++ ++ ++ ++ ++++Anti-IgG/C3d (1024) FSL-B +++ +++ ++++ ++++ ++++ ++++ ++++
OLIVER ET AL.
2668 TRANSFUSION Volume 51, December 2011

The 256 titer IgG anti-A of the group B serum wasneutralized by FSL-A at concentrations as low as 2 mg/mLbut the 1024 titer IgG anti-A of the group O sample wasmore difficult to neutralize. The FSL-A 20 mg/mL solutionwas able to reduce the 4+ serologic activity score to 1+ whilethe 10 mg/mL solution reduced it to 2+. The FSL-A+B com-bination appeared to have no advantages in neutralizationof anti-A in O serum and the FSL-B had little effect.
In vitro dissociation of anti-A fromsensitized A1 cellsThe ability of FSL-A to dissociate anti-A from A1 RBCspresensitized with serial dilutions of group O serum wasevaluated over time (Table 4). The PBS reaction representsthe sensitization score obtained in the absence of FSL-A. Itwas clear that the cell-bound agglutinin anti-A was com-pletely dissociated from the RBCs within 30 minutes. Incontrast, IgG anti-A activity was more resilient and onlyresulted in a minor reduction in score. Identical resultswere obtained at each 30-minute interval up to 120minutes, indicating that no increased elution occurredafter 30 minutes.
Symptoms and hemoglobinuriaIn the trial to establish our ability to detect intravascularhemolysis, we observed no symptoms or changes inbehavior after a 40-mL infusion of hemolyzed RBCs in anymice. However, bladder urine samples of all these animalsshowed hemoglobinuria ranging in color from pink at 5
minutes to deep red after 15 minutes. Red urine wasclearly observed on the filter paper of the sixth mouse.Because 40 mL of hemolyzed RBCs represents two-thirdsof the 100-mL transfusion of 60% kodecytes (see below),the detection of pink- or red-colored urine on filter paperfor up to 2 hours after transfusion was considered indica-tive of an acute intravascular hemolytic transfusion reac-tion. There were no observable symptoms, includinghemoglobinuria, detected in any mice receiving intactA+biotin kodecytes or FSL-A infusions, regardless of thepresence or absence of anti-A.
In vivo FSL-A infusions (mice)FSL-A constructs were infused directly into the venouscirculation of 17 mice (doses up to 200 mg of FSL-A/kgbody mass or about 5.7 mg/mL of plasma). All infusionsproduced no symptoms while anesthetized or aftersurgery. The administered volume of 200 to 250 mLequates with a ratio of approximately 2:7 of FSL-A infusionto the estimated plasma volume.19
Three months after infusion histologic examinationof major organs and hematologic examination of the fivemice receiving infusions (in the specific toxicity series) didnot find any evidence of toxicity, nor could they be distin-guished from the five untreated control animals.
In all mice receiving an infusion of approximately3 mg or more of FSL-A/mL of plasma, their circulatingRBCs acquired the blood group A phenotype—and becameso-called in vivo A kodecytes (Table 5). This acquisition of
TABLE 4. FSL-A in vitro dissociation of anti-A from group O serum sensitized A1 RBCs*
Serology detection system Neutralizer
Anti-A reactivity (created by serial dilution)
1 2 4 8 16 32
Direct agglutination PBS +++ ++ -FSL-A (4 mg/mL) - - -
Indirect agglutination (anti-IgG/C3d) PBS ++++ ++++ ++ ++ + -FSL-A (4 mg/mL) +++ ++ + - - -
*Identical results were obtained for 30, 60, 90, and 120 minutes.
TABLE 5. In vivo transformation of mouse RBCs with FSL-A infusions
Cluster IDInitial anti-A
status
FSL-A dose Anti-A serology hours after FSL-A infusion (in vivo A kodecytes)
mg/kg body mass mg/mL plasma 0.1 2 8 24 48 72
1091 - 200 5.7 ++++ ++++ ++++ ++++ ++ -7058 - 200 5.7 ++++ ++++ +++ ++++ - -4424 - 100 2.9 +++ ++++ +++ +++ -9570 - 100 2.9 +++ ++++ ++++ +++ -4310 - 50 1.4 - - - - -0343 - 50 1.4 - - - - -
IV(i) 1124 + 200 5.7 ++++ ++++ ++++ - -IV(i) 1389 + 200 5.7 ++++IV(i) 9239 + 200 5.7 ++++ ++++ ++++ +++ -IV(ii) 8998 + 200 5.7 ++++ ++++ +++ ++ -IV(ii) 6014 + 200 5.7 ++++ ++++ ++++ ++ -V 3649 + 200 5.7 ++++ ++++ ++++ +++ -
ABO-INCOMPATIBLE TRANSFUSION
Volume 51, December 2011 TRANSFUSION 2669

FSL-A appeared to be rapid, with the first sample takenat 6 minutes having a 4+ serologic reaction againstanti-A. However, as approximately 30 minutes elapsedafter collection and washing, in vitro transformationwill have occurred (see above for in vitro serumtransformation).
This in vivo creation of kodecytes occurred regardlessof whether the animal had circulating anti-A or not(Table 5) and the incompatible A kodecytes appeared tobe tolerated without consequence. If pretransfusionserum was tested against washed in vivo A kodecytes, thenthey gave strong 3+ serologic reactions, clearly indicatingthat circulating in vivo A kodecytes were serologicallyincompatible with preneutralized antibodies.
The in vivo A kodecyte status was transient and gen-erally detectable for less than 48 hours. The similaritybetween the clearance rate of the 200 and 100 mg/kg invivo created A kodecytes suggests that clearance may bemore strongly influenced by time than concentration(Table 5). Although not unambiguous there is some sug-gestion that the level of FSL-A detected at 24 hours on thein vivo kodecytes in the animals with anti-A (Clusters IVand V) was slightly less than that in the animals withoutanti-A. It is possible that less FSL-A was available for invivo modification due to its prior interaction with anti-Aand possible systemic clearance.
The period of retention of the FSL-A construct in thecirculating cells does not necessarily reflect the plasmalevels as the construct is probably retained longer in thecell membrane than free in plasma (see discussion). Wecould not specifically establish the time FSL-A constructsremained in circulatory plasma or its concentration.However, after 8 hours, the levels of plasma FSL-A musthave decreased below 1 mg/mL (the minimum level atwhich in vivo transformation can occur), thereby allowingthe kodecytes to begin to lose constructs from their mem-branes. In agreement with this is the observation that afterFSL-A infusion, anti-A did not become detectable inplasma until after 8 hours (Table 6), suggesting that theFSL-A levels of plasma had fallen below that capable ofcausing total anti-A neutralization.
Compatible transfusion (mice)The transfusion of a 100-mL 60% suspension of A+biotinkodecytes into two mice that had no anti-A (Cluster I) didnot result in any observable symptoms. Starting from thetheoretical 10% point (contribution of the kodecyte trans-fusion to cell count) the kodecytes count of both animalsreduced by approximately 10% at the first sampling point(0.1 hr) before reducing further and then stabilizing into agradual decline at approximately 8 hours (Table 6, Fig. 2).A similar pattern over the first 2 hours was observed ina single animal (Animal 7994, Cluster III) who was alsogiven FSL-A (albeit less abrupt).
TAB
LE
6.S
urv
ivin
gm
ou
seko
dec
ytes
and
anti
-Ap
rese
nce
or
abse
nce
atti
me
inte
rval
s,so
rted
into
rela
ted
clu
ster
s
Clu
ster
IDIn
itial
anti-
AF
SL-
Am
g/m
Lpl
asm
a
Per
cent
age
surv
ivin
gA
+bio
tinko
decy
tes
and
anti-
Apr
esen
ce[+
]or
abse
nce
[–]
attim
ein
terv
als
(hr)
Nec
rops
yan
ti-A
Hou
rsaf
ter
first
tran
sfus
ion
(10%
)M
idw
ayan
ti-A
Hou
rsaf
ter
seco
ndtr
ansf
usio
n(1
0%)
0.1
28
2448
0.1
28
2448
I00
86[-
]0
9.5
9.6
7.9
7.6
6.6
I12
40[-
]0
8.9
7.5
5.6
5.4
4.5
II05
06[+
]0
5.2
3.6
2.3
II48
62[+
]0
4.7
3.9
3.0
II44
67[+
]0
3.5
3.4
3.3
II33
97[+
]0
3.2
1.9
1.4
II09
54[+
]0
2.2
1.3
II61
61[+
]0
2.1
[+]
2.1
2.1
0.4
II32
26[+
]0
1.5
[+]
1.2
[+]
1.2
1.1
0.7
III79
94[-
]5.
710
.0[-
]9.
67.
87.
05.
6IV
(i)11
24[+
]5.
710
.0[-
]9.
4[-
]7.
67.
2[+
]6.
2[+
][+
]1.
7[+
]1.
61.
31.
41.
4[+
]>1
28IV
(i)13
89[+
]5.
79.
5[-
]6.
3[-
]6.
45.
4[+
]5.
24.
53.
7[+
]2.
31.
61.
3[+
]>1
28IV
(i)92
39[+
]5.
78.
86.
46.
46.
04.
4[+
][+
]2.
62.
52.
61.
8[+
]>1
28IV
(ii)
8998
[+]
5.7
9.4
[-]
7.4
5.4
[-]
5.6
4.9
[+]
[+]
6.4
6.3
6.1
5.9
5.8
[+]
=8
IV(ii
)60
14*
[+]
5.7
8.4
[-]
8.3
8.2
[-]
7.3
[+]
5.4
[+]
[+]
9.5
8.3
8.2
7.4
7.2
[+]
=12
8V
3649
*[+
]5.
77.
67.
53.
40.
50.
50.
20.
10.
1[+
]>1
28
*S
econ
dtr
ansf
usio
nw
asgi
ven
1w
eek
afte
rth
efir
sttr
ansf
usio
n;al
loth
ers
wer
e2
wee
ksaf
ter
the
first
tran
sfus
ion.
OLIVER ET AL.
2670 TRANSFUSION Volume 51, December 2011

Incompatible transfusion (mice)The transfusion of a 100-mL 60% suspension of A+biotinkodecytes into seven mice that had anti-A (Cluster II) alsodid not result in any observable symptoms. Starting fromthe theoretical 10% point, the kodecytes count of allanimals reduced by approximately 50% to 85% at the firstsampling point (0.1 hr) then stabilized into a gradualdecline (Table 6, Fig. 2).
FSL-A neutralized incompatible transfusion (mice)The first transfusion of a 100-mL 60% suspension ofA+biotin kodecytes into six mice that had anti-A (ClustersIV and V) and had been given 5.7 mg of FSL-A/mL ofplasma did not produce any observable symptoms. It canbe seen that anti-A was neutralized in these animals(Table 6, [–]) for at least 8 hours. The mean shape of the
kodecyte survival curve for the fiveanimals with anti-A and FSL-A neutral-ized (Cluster IV) was very dissimilar tothe untreated animals with anti-A(Cluster II; p < 0.001 for all points;Fig. 2, Table 6). In fact the curve was notsignificantly different (p > 0.05 for allpoints) from that of Cluster I animalswithout anti-A (Fig. 2). At 24 hourswhen the anti-A had started again tobecome detectable (Table 6, [+]) the invivo A kodecyte status was also chang-ing to negative (Table 5). It needs to benoted that the level of FSL-A in thetransfused A kodecytes is at least 20¥higher than that in the in vivo formedkodecytes and remains detectable inthe surviving kodecytes for at least 3days.17
One animal, Animal 3649, had anunusual survival profile (Table 6) andwas split into its own cluster (Cluster V).In this animal the rate of A+biotin kode-cyte loss was markedly greater than theothers and by 24 hours had crashed to0.5%. At first glance it appears thatthe reappearance of anti-A at 24 hours(Table 6, [+]) may have been res-ponsible; however, anti-A was alsodetectable in other Cluster IV animalswho did not respond so adversely. Atthis time point, the animal would beexpected to be losing its in vivo A kode-cyte status, and there was no evidenceof a hemolytic event involving its in vivoA kodecytes. Kodecyte destruction wasextremely rapid when the second Akodecyte transfusion was given 1 week
later without FSL-A anti-A neutralization, reaching 0.2%within 6 minutes (yet without evidence of hemoglo-buinuria). Anti-A levels and isotype were similar in otheranimals with anti-A, and there was no immunologic ELISAevidence that this animal had an antibody to either biotinor the FSL construct per se (not shown).
The second (rechallenge) transfusion with A kode-cytes without FSL-A anti-A neutralization in the Cluster IVanimals, whose serum could be demonstrated to haveanti-A (Table 6, [+]), also did not result in any observablesymptoms. However, this time there appeared to be twodifferent outcomes. Starting from the theoretical 10%point the kodecyte count of three animals (Cluster IV(i))reduced by approximately 50% to 85% at the first samplingpoint (0.1 hr) then stabilized into a gradual decline(Table 6, Fig. 2). The average shape of the kodecyte sur-vival curve (Fig. 2) for these three animals (Cluster IV(i))
Fig. 2. Average cluster survival curves of A+biotin kodecytes transfused into animals
with and without anti-A and with and without FSL-A infusions. Upper bar graph
indicates average kodecyte first transfusion survival (with 1 SD error bars) obtained
at each sampling time point for Cluster II (anti-A) and Cluster IV (FSL-A neutralized
anti-A) animals. Line diagram compares trends over time. Control animals are
Cluster I, which did not have anti-A and showed normal survival, while Cluster II
animals with anti-A showed antibody-mediated kodecyte clearance. FSL-A neutral-
ized anti-A in Cluster IV animals showed normal survival of their first kodecyte
transfusion. The second transfusion into these Cluster IV animals, when their
plasma FSL-A was absent, resulted in two different patterns. Cluster IV(i) animals
resulted in the expected antibody-mediated kodecyte clearance pattern, while
Cluster IV(ii) animals showed a normal survival pattern. All curves are means of the
data in their clusters and are plotted from the theoretical 10% transfusion starting
point, indicated by an open circle.
ABO-INCOMPATIBLE TRANSFUSION
Volume 51, December 2011 TRANSFUSION 2671

was very similar to that of animals with anti-A (Cluster II)not receiving FSL-A anti-A neutralization and was evi-dence that in the absence of FSL-A protection, A kode-cytes would not survive normally in these specificanimals. Unexpectedly, despite being positive for anti-Aby ELISA, two animals, Animals 8998 and 6014 (classifiedas Cluster IV(ii)), had second kodecyte transfusion sur-vival curves very similar to that of animals without anti-Aor with neutralized anti-A (Fig. 2). The reason for thisapparent normal survival in Animal 8998 can perhaps beexplained by a low anti-A titer (8) and, although the 128titer of Animal 6014 was high, it was not more than 128 asobserved in the other animals. It is not known whetherthis reduced level of antibody in these two animals was anindirect consequence of FSL-A treatment or a naturalvariation.
DISCUSSION
Breaching the ABO barrier can be met with significant andsometimes fatal consequences.1,2 The clearance of ABO-incompatible RBCs is clearly mediated by the actions ofantibody and complement2 and incompatible blood oncetransfused is usually destined to a reduced life span. Therate of vascular clearance has been shown to be depen-dent on antibody levels2,3 and a reduction in antibody by avariety of approaches including immunoabsorption andplasmapheresis are current strategies for allowing ABO-incompatible transplantation.11,20 There is substantial evi-dence that RBCs can also survive in the presence of lowlevels of ABO antibodies, including antibody sensitization,as seen by the practice of group O whole blood transfu-sions into group A and B individuals2,21 and the occasionaltolerated accidental transfusions of ABO-incompatibleblood.1,2 Deliberately transfusing RBCs across the ABObarrier has been used to reduce antibody levels beforebone marrow transplantation,7,8 and platelets that poorlyexpress ABO antigens can be safely given unmatched forABO,22 albeit with reduced efficacy.23
Attempting to in vivo neutralize ABO antibodies hasbeen experimentally attempted, where intravenous (IV)trisaccharide infusions allowed for experimental incom-patible transplants in baboons24 and in treatment of ABOhemolytic disease of the newborn.25 There are, however,no reports of attempting to neutralize recipient ABO anti-bodies and giving a deliberate ABO-incompatible transfu-sion. Historical evidence of the ability to neutralizecomplement-activating antibodies with glycolipids orblood group substances, and then successfully transfuseincompatible RBCs, has been established for the Lewisblood group system.12,13 Lewis-positive plasma (whichcontains only approx. 2 mg/mL Lewis glycolipids26,27) willeffectively neutralize circulating Lewis antibodies andallow for the subsequent transfusion of Lewis-positiveRBCs.12,13 Because FSL constructs have properties like gly-
colipids, a propensity to associate with plasma27,28 and anability to inhibit viruses and toxins,16 we considered thepossibility that these constructs could also be used toinhibit ABO antibodies.
To undertake this study in humans was not feasible sowe established an animal model, guided by in vitro dataderived from human cell or serum experiments. Unfortu-nately, because of the approaches and/or reagents used,we cannot be certain of the relative contributions of IgM/IgG/C3d to the results of some in vitro experiments.
The animal modeling required several steps includingstimulating ABO antibodies in animals without anti-A, arti-ficially creating ABO-incompatible RBCs, and measuringtheir posttransfusion survival. The first step of stimulatingABO antibodies was relatively simple although the choiceof mouse strain was an important consideration.29,30 Weelected only to study blood group A antigen incompatibil-ity due to the fact that animals have a restricted antibodyresponse to the blood group B antigen, because they havethe linear B antigen.31
The second step was that ABO antigens are notpresent on the RBCs of small laboratory animals.32 Trans-fusing human RBCs into laboratory animals is not usuallyan option due to antispecies activity, substantial antigenicdifferences, and physical differences.33 For example, thehuman RBC is two times larger in diameter than themouse RBC and is known to have reduced survivalin mice. We therefore had to create artificial ABO-incompatible mouse RBCs by attaching blood group Aantigens onto them with FSL constructs, thus creatingin vitro A kodecytes. By using FSL constructs and mouseRBCs the only foreign antigen(s) are those introduced bythe FSL construct(s).
The third step was the ability to detect and measure atransfusion reaction at multiple time points in the smallblood volume of a mouse. To achieve this we included inthe kodecyte a biotin label.17 Thus incompatible A+biotinkodecytes for transfusion were created from compatiblemouse RBCs modified with FSL-A and FSL-biotin. Thesurvival of these cells could be determined by visualizingA+biotin kodecytes after staining with avidin fluorophoreconjugate, with respect to method limitations.17 As previ-ously reported, data for up to 24 hours only should beconsidered as the FSL constructs dissociate from kode-cytes at a rate of about 20% per day.17
Based on natural glycolipid data,27,28,34 the infused FSLconstructs will be expected to predominately associatewith plasma lipids (e.g., high-density lipoproteins and low-density lipoproteins) where they will be mobile and flexibleand thus become multivalent neutralizing lipid particles.The preferred association of FSLs with plasma rather thanfor cells was demonstrated both in vitro (Table 1) and invivo (Table 5) where approximately 40 times more FSL-Awas required in serum to bring about the same level oftransformation as seen with cells modified in PBS. It was
OLIVER ET AL.
2672 TRANSFUSION Volume 51, December 2011

estimated that only 2.5% of the FSLs in plasma will associ-ate with RBCs and that is why FSLs which dissociate fromkodecytes are unlikely to detectably relabel other circulat-ing cells. The association with cells, so-called in vivo trans-formation, occurred regardless of the presence or absenceof anti-A and always where the FSL-A dose was greater than2 mg/mL plasma. This is similar to the phenomenondescribed when Lewis-positive plasma is transfused toLewis-negative individuals.35
Our in vitro experiments suggest that at least 5 to10 mg of FSL-A/mL of plasma would be required to ensuresignificant antibody neutralization of a human group Oplasma with high titer of anti-A (Table 3). Tolerance ofincompatible A kodecytes in the in vivo mouse model sug-gested doses as low as 4 mg/mL plasma could be adequate(Table 6). Similarly in vivo kodecytes spontaneouslyforming in anti-A–positive animals with levels of FSL-Agreater than 1 mg/mL were tolerated without any evi-dence of cell destruction or sensitization. But thesein vivo–formed kodecytes are probably tolerated becauseof their much lower antigen density than in vitro kodecytes.In contrast, natural blood group A1 cells appeared to be lessin vitro tolerant of FSL-A neutralized antibody (Table 3)probably because they have more A antigen than A kode-cytes, and the natural antigens may also present in a biva-lent configuration. Results from in vitro neutralizationassays, where the antibody is still present but neutralizedby FSL-A, may not necessarily reflect in vivo neutralizationdynamics. As a consequence the speculated actual dosethat could prevent an incompatible transfusion reaction inhumans is probably somewhere between 1 and 10 mg/mLplasma. If the dose was 5 mg/mL of plasma (or 0.2 g/kgbody mass), then this would equate with a 13.5-g infusion(e.g., 675 mL of 20 mg/mL) for a 70-kg human. This dose iscompatible with IV infusion practice where for example0.7 g/kg body mass of saccharides in the form of glucose,36
or 2.5 g/kg body mass of lipid in the form of intralipid37 canbe safely given to a human. Additionally doses of A and Btrisaccharides at 1 to 2 g/kg body mass have been safelygiven to baboons to prevent incompatible transplant rejec-tion24 and approximately 0.025 mg/kg body mass tohuman newborns.25
The in vivo experiments here show the administra-tion of the FSL construct immediately before the transfu-sion of incompatible blood. This, of course, would only beof value if a deliberate transfusion of incompatible blood(or organs) was anticipated. FSL intervention to mitigatean accidental incompatible transfusion reaction wouldonly be of value if surviving incompatible cells werepresent and destined for destruction. This is often thecase, as ABO incompatibility is to some extent self-limiting, due to the consumption of antibody and/orcomplement.2 FSL infusions may further mitigate anyreactions particularly if they able to dissociate bound anti-body as suggested by our in vitro data (Table 4) and the
literature.25,38 The exact period of time the FSL-A con-structs will remain in plasma is uncertain, but studies withrelated 125I-radiolabeled FSLs injected into rats show thatthey are cleared from the circulation within 24 hours(unpublished). This appears to correlate with the in vivoantibody neutralization data here, which show reappear-ance of detectable antibody at approximately 24 hours. Incontrast to simple trisaccharides, which are excretedrapidly in urine,38 having the neutralant as an FSL con-struct appears to significantly extend its circulating half-life. Furthermore, the fate of the anti-A that hascomplexed with FSL-A is unknown, and the possibilitythat these antibodies may also be cleared has yet to beestablished.
A significant concern raised by the Lewis blood groupsystem is the neutralization process can induce a second-ary antibody response.12 In the case of Lewis incompatibil-ity this is not a problem as the transfused cells becomecompatible through antigen loss.12 But for cell-boundantigens such as ABO this secondary response would posea significant issue (despite the fact that cell-bound anti-gens may undergo antigenic modulation4,5). However, thematerial historically used to neutralize Lewis antibodieswas plasma13,39 or Lewis blood group substance12 and notpurified glycolipids. Plasma alone (glycolipids plus glyco-proteins) appeared not to change the antibody titersubstantially13,39 but the purified glycoprotein Lewis sub-stance neutralization resulted in a substantial secondaryantibody response.12 Glycolipids are immunologicallyprocessed very differently from glycoproteins40 andglycoproteins would be expected to induce a significantantibody response, while glycolipids as poor immunogensprobably would not.40-42 There is one further distinction tobe made here, which is that FSL constructs are notglycolipids—they have a spacer and a different lipid tail14,16
and as a consequence probably have different immuno-logic attributes than glycolipids. In unpublished work wehave attempted to induce an antibody response to theFSL-A constructs by mixing them with adjuvant andimmunizing several series of mice and none producedanti-A (while salivary glycoprotein A control animals did).To further evaluate the immunology of FSL constructs weconstructed FSL-Gala. Unlike the glycolipid Gala ceram-ide that powerfully activates human invariant NKT cellsafter binding to CD1d, FSL-Gala showed no ability tostimulate human invariant NKT cell clones (R. Dunbar,unpublished), further evidence that FSLs behave differ-ently than natural glycolipids. Based on the crystal struc-ture,43 it appears that the spacer of the FSL constructwould probably impair its ability to complex with CD1dand glycolipids are only immunogenic when bound toCD1 molecules.40
Finally it is also possible that FSL constructs may insome circumstances be able to down regulate the immunesystem by inducing auto regulation, thereby causing down
ABO-INCOMPATIBLE TRANSFUSION
Volume 51, December 2011 TRANSFUSION 2673

regulation of specific antibody production. The ClusterIV(ii) mice potentially showed this effect, which is alsosupported by the literature where prolonged immuniza-tions with glycolipids has resulted in decreased specificantibody titers.41,42
This proof-of-concept trial showed that FSL-A canneutralize anti-A in vitro and in vivo. The opportunity toalso use FSL constructs in an in vitro setting with rare butABO-incompatible reagent antibodies is also possible. Itnow remains to be established if these or related con-structs (including those bearing peptides or in multivalentconfigurations) can be translated into a clinical setting.
ACKNOWLEDGMENTS
Dr Sandy Ferguson is acknowledged for his support and veteri-
nary expertise and Katie Barr for the inkjet printing antibody
ELISA technology.
CONFLICT OF INTEREST
SH is the CEO/CSO and a stockholder of KODE Biotech Ltd, the
patent owner of KODE cell surface engineering technology. CO
and DB are stockholders in KODE Biotech Ltd.
REFERENCES
1. Janatpour KA, Kalmin ND, Jensen HM, Holland PV. Clinical
outcomes of ABO-incompatible RBC transfusions. Am J
Clin Pathol 2008;129:276-81.
2. Mollison PL. The transfusion of red cells. In: Klein HG,
Anstee DJ, editors. Mollison’s blood transfusion in clinical
medicine. 11th ed. Oxford: Blackwell Scientific
Publications; 2005. p. 412-75.
3. Chaplin H. Studies on the survival of incompatible cells in
patients with hypogammaglobulinaemia. Blood 1959;14:
24-36.
4. Zimring JC, Hair GA, Chadwick TE, Deshpande SS, Ander-
son KM, Hillyer CD, Roback JD. Nonhemolytic antibody-
induced loss of erythrocyte surface antigen. Blood 2005;
106:1105-12.
5. Zimring JC, Caldwell CM, Spitalnik SL. Antigen loss from
antibody-coated red blood cells. Transfus Med Rev 2009;23:
189-204.
6. Liepkalns JS, Zimring JC. Crossmatch incompatible RBCs
have an intrinsic range of susceptibility to hemolysis.
Transfusion 2009;49:28A.
7. Nussbaumer W, Schwaighofer H, Gratwohl A, Kilga S,
Schönitzer D, Nachbaur D, Niederwieser D. Transfusion of
donor-type red cells as a single preparative treatment for
bone marrow transplants with major ABO incompatibility.
Transfusion 1995;35:592-5.
8. Scholl S, Klink A, Mügge LO, Schilling K, Höffken K, Sayer
HG. Safety and impact of donor-type red blood cell trans-
fusion before allogeneic peripheral blood progenitor cell
transplantation with major ABO mismatch. Transfusion
2005;45:1676-83.
9. Olsson ML, Clausen H. Modifying the red cell surface:
towards an ABO-universal blood supply. Br J Haematol
2007;140:3-12.
10. Garratty G. Modulating the red cell membrane to produce
universal/stealth donor red cells suitable for transfusion.
Vox Sang 2008;94:87-95.
11. Crew RJ, Ratner LE. ABO-incompatible kidney transfusion:
current practice and the decade ahead. Curr Opin Org
Transplant 2010;15:526-30.
12. Mollison PL, Polley MJ, Crome P. Temporary suppression
of Lewis blood-group antibodies to permit incompatible
transfusion. Lancet 1963;1:909-12.
13. Pelosi MA, Bauer JL, Langer A, Hung CT. Transfusion of
incompatible blood after neutralization of Lewis antibod-
ies. Obstet Gynaecol 1974;44:590-3.
14. Frame T, Carroll T, Korchagina E, Bovin N, Henry S. Syn-
thetic glycolipid modification of red blood cell mem-
branes. Transfusion 2007;47:876-82.
15. Heathcote D, Carroll T, Wang JJ, Flower R, Rodionov I,
Tuzikov A, Bovin N, Henry S. Novel antibody screening
cells, MUT+Mur kodecytes, created by attaching peptides
onto red blood cells. Transfusion 2010;50:635-41.
16. Harrison AL, Olsson ML, Jones RB, Ramkumar S, Sakac D,
Binnington B, Henry S, Lingwood CA, Branch DR. A syn-
thetic globotriaosylceramide analogue inhibits HIV-1
infection in vitro by two mechanisms. Glycoconj J 2010;27:
515-24.
17. Oliver C, Blake D, Henry S. Modeling transfusion reactions
and predicting in vivo cell survival with kodecytes. Trans-
fusion 2011;51:1723-30.
18. Barr K, Diegel O, Parker S, Bovin N, Henry S. Function-
Spacer-Lipid (FSL) constructs enable inkjet printing of
blood group antigens [abstract E1.12]. FEBS J 2010;277
Suppl 1:235.
19. Hoff J. Methods of blood collection in the mouse. Lab
Anim 2000;29:47-53.
20. Rydberg L. ABO-incompatiblity in solid organ transfusion.
Transfus Med 2001;11:325-42.
21. Lefebre J, McLellan BA, Coovadia AS. Seven years experi-
ence with group O unmatched packed red blood cells
in a regional trauma unit. Ann Emerg Med 1987;16:
1344-8.
22. British Committee for Standards in Haematology, Blood
Transfusion Task Force (Chairman P Kelsey). Guidelines for
the use of platelet transfusions. Br J Haematol 2003;122:10-
23.
23. Shehata N, Tinmouth A, Naglie G, Freedman J, Wilson K.
ABO-identical versus nonidentical platelet transfusion: a
systematic review. Transfusion 2009;49:2442-53.
24. Cooper DK, Ye Y, Niekrasz M, Kehoe M, Martin M, Neeth-
ling FA, Kosanke S, DeBault LE, Worsley G, Zudhi N, Oriol
R, Romano E. Specific intravenous carbohydrate therapy. A
new concept in inhibiting antibody-mediated rejection—
OLIVER ET AL.
2674 TRANSFUSION Volume 51, December 2011

experience with ABO-incompatible cardiac allografting in
the baboon. Transplantation 1993;56:769-77.
25. Romano EL, Soyano A, Montano RF, Ratcliffe M, Olson M,
Suarez G, Martinez N, Worsely G. Treatment of ABO
hemolytic disease with synthetic blood group trisaccha-
rides. Vox Sang 1994;66:194-9.
26. Hammar L, Månsson S, Rohr T, Chester MA, Ginsburg V,
Lundblad A, Zopf D. Lewis phenotype of erythrocytes and
Leb-active glycolipid in serum of pregnant women. Vox
Sang 1981;40:27-33.
27. Rohr TE, Smith DF, Zopf DA, Ginsburg V. Leb-active gly-
colipid in human plasma: measurement by radioimmu-
noassay. Arch Biochem Biophys 1980;199:265-9.
28. Swarzmann G. Uptake and metabolism of exogenous gly-
cosphingolipids by cultured cells. Cell Dev Biol 2001;12:
163-71.
29. Larkin JM, Porter CD. Mice are unsuitable for modeling
ABO discordance despite strain-specific A cross reactive
natural IgM. Br J Haematol 2005;130:310-7.
30. Néron S, Lemieux R. Type 2 T-cell-independent murine
immune response to the human ABO blood group anti-
gens. Vox Sang 1994;67:68-74.
31. Galili U. Immune response, accommodation, and toler-
ance to transplantation carbohydrate antigens. Transplant
2004;78:1093-8.
32. Oriol R, Le Pendu J, Mollicone R. Genetics of ABO, H,
Lewis, X and related antigens. Vox Sang 1986;51:161-71.
33. Hod EA, Zimring JC, Spitalnik SL. Lessons learned from
mouse models of hemolytic transfusion reactions. Curr
Opin Haematol 2008;15:601-5.
34. Marcus DM, Cass LE. Glycosphingolipids with Lewis blood
group activity: uptake by human erythrocytes. Science
1969;164:553-5.
35. Sneath JS, Sneath PH. Transformation of the Lewis groups
of human red cells. Nature 1955;176:172.
36. Moreno A, Blüml S, Hwang JH, Ross BD. Alternative 1-13C
glucose infusion protocols for clinical 13C MRS examina-
tions of the brain. Magn Reson Med 2001;46:39-48.
37. Josephson DL. Intravenous nutritional support. In: Joseph-
son DL, editor. Intravenous infusion therapy for nurses.
2nd ed. Clifton Park (NY): Thomson Delmar Learning;
2004. p. 399.
38. Romano EL, Soyano A, Linares J, Lauzon GJ. Neutralization
of ABO blood group antibodies by specific oligosaccha-
rides. Transplant Proc 1987;19:4426-30.
39. Andorka DW, Arosemena A, Harris JL. Neutralization in
vivo of Lewis antibodies. Report of two cases. Am J Clin
Pathol 1974;62:47-51.
40. De Libero G. The Robin Hood of antigen presentation.
Science 2004;303:485-7.
41. Lingwood CA, Murray RK, Schachter H. The preparation of
rabbit antiserum specific for mammalian testicular sul-
fogalactoglycerolipid. J Immunol 1980;124:769-74.
42. Alving CR, Richards RL. Immune reactivities of antibodies
against glycolipids—II. Comparative properties, using lipo-
somes, of purified antibodies against mono-, di and tri-
hexosyl ceramide haptens. Immunochemistry 1977;14:
383-9.
43. Koch M, Stronge VS, Shepherd D, Gadola SD, Mathew B,
Ritter G, Fersht AR, Besra GS, Schmidt RR, Jones EY,
Cerundolo V. The crystal structure of human CD1d with
and without alpha-galactosylceramide. Nat Immunol
2005;6:819-26.
ABO-INCOMPATIBLE TRANSFUSION
Volume 51, December 2011 TRANSFUSION 2675




















