
In vitro and in vivo toxicological evaluation of extract and fractions from Baccharis trimera with...
10
Journal of Ethnopharmacology 138 (2011) 513–522 Contents lists available at SciVerse ScienceDirect Journal of Ethnopharmacology journa l h o me page: www.elsevier.com/locate/jethpharm In vitro and in vivo toxicological evaluation of extract and fractions from Baccharis trimera with anti-inflammatory activity N.P.A. Nogueira a , P.A. Reis a , G.A.T. Laranja a , A.C. Pinto a , C.A.F. Aiub b , I. Felzenszwalb b , M.C. Paes a , F.F. Bastos a , V.L.F.C. Bastos a , K.C.C. Sabino a , M.G.P. Coelho a,∗ a Departamento de Bioquímica, Instituto de Biologia Roberto Alcantara Gomes, Centro Biomédico, Universidade do Estado do Rio de Janeiro, Av. Professor Manoel de Abreu, 444, PAPC, 4o andar, CEP 20550-170, Rio de Janeiro, RJ, Brazil b Departamento de Biofísica e Biometria, Instituto de Biologia Roberto Alcantara Gomes, Centro Biomédico, Universidade do Estado do Rio de Janeiro, Av. Professor Manoel de Abreu, 444, PAPC, 4o andar, CEP 20550-170, Rio de Janeiro, RJ, Brazil a r t i c l e i n f o Article history: Received 11 May 2011 Received in revised form 5 September 2011 Accepted 8 September 2011 Available online 12 October 2011 Keywords: Baccharis trimera Cytotoxicity Genotoxicity Mutagenicity a b s t r a c t Ethnopharmacological relevance: Baccharis trimera (Less) DC. (Asteraceae), popularly known in Brazil as “carqueja”, have been used in folk medicine to treat gastrointestinal, hepatic and renal diseases, and inflammatory processes as rheumatism. Aim of the study: To evaluate the in vitro and in vivo toxicological effects of anti-inflammatory Baccharis trimera aqueous extract and fractions. Materials and methods: Aqueous extract of Baccharis trimera (AEBt) was produced by infusion in boiling water. After lyophylization AEBt was extracted with 80% ethanol, originating the ethanolic supernatant fraction (EFBt) and the aqueous sediment fraction (AFBt). Anti-inflammatory properties of AEBt, EFBt or AFBt (3, 30 or 300 g/kg b.w.) were evaluated by the carrageenan-induced mouse paw edema using indomethacin (10 mg/kg) as positive control. The growth of rat hepatoma cells (HTC) and human embryo kidney epithelial cells (HEK) was determined by protein staining assay. Cytotoxicity was assayed by the tetrazolium salt (MTT) reduction. Cyclosporin was used as reference cytotoxic drug for spleen cells and doxorubicin for HTC and HEK cells. For in vivo toxicological evaluation SW male mice were daily and oral (gavage) treated with extract/fractions at 4.2 mg/kg or 42 mg/kg during 15 days. After treatment liver or kidney cells were submitted to comet assay to determine the DNA damage index, and the glutathione S-transferase activity was assayed towards ETHA (class Pi) and CDNB (several classes). Mutagenicity was evaluated by the Ames test using Salmonella typhimurium strains TA97, TA98, TA100, and TA102. Results: The anti-inflammatory effects of EFBt were higher than those of AEBt or AFBt. Mice treat- ment (3–300 g/kg) with AFBt reduced the paw edema (3 h) at lower levels (29.2–37.3%; P < 0.01), than those observed for AEBt (44.7–54.2%; P < 0.001), EFBt (49.3–58.2%; P < 0.001) or indomethacin (64.6%, P < 0.001, 10 mg/kg). The growth of kidney cells (HEK) was inhibited by AEBt (IC 50 182.6 g/ml), EFBt (IC 50 78.1 g/ml) and AFBt (IC 50 86.2 g/ml), with lower effects on HTC hepatic cell (IC 50 308.8 g/ml, 396.5 g/ml and 167.9 g/ml, respectively). As evaluated by MTT test, AFBt exhibited cytotoxicity for HEK cells (IC 50 372.5 g/ml), but none for HTC ones; by the way, AFBt stimulated spleen cells (EC 50 2.2 g/ml) while cyclosporine, a cytotoxic reference drug inhibited them with IC 50 of 0.42 g/ml; the IC 50 for doxoru- bicin for HEK and HTC cells was 0.28 g/ml and 14.4 g/ml, respectively, at 96 h. No mutagenic potential was observed. Mice treatment with AEBt or AFBt at 42 mg/kg for 15 days altered the kidney relative weight, but not at 4.2 mg/kg. Baccharis trimera did not change liver, spleen or popliteal lymph node rela- tive weight. DNA damage index of kidney cells was observed on mice treated with AEBt/AFBt, but not on animals treated with EFBt, while DNA lesions were detected on liver cells only after AFBt treatment. The general activities of hepatic GST and Pi GST were reduced by EFBt and AFBt treatment, respectively. Conclusions: Baccharis trimera did not show mutagenicity, inhibited the GST activity, a hepatic detox- ification enzyme, and induced in vivo (genotoxicity) and in vitro toxicological effects to kidney cells. © 2011 Elsevier Ireland Ltd. All rights reserved. ∗ Corresponding author. Tel.: +55 21 2587 6143; fax: +55 21 2587 6136. E-mail addresses: [email protected], [email protected] (M.G.P. Coelho). 0378-8741/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.jep.2011.09.051
Transcript of In vitro and in vivo toxicological evaluation of extract and fractions from Baccharis trimera with...

It
NMa
4b
4
a
ARRAA
KBCGM
0d
Journal of Ethnopharmacology 138 (2011) 513– 522
Contents lists available at SciVerse ScienceDirect
Journal of Ethnopharmacology
journa l h o me page: www.elsev ier .com/ locate / je thpharm
n vitro and in vivo toxicological evaluation of extract and fractions from Baccharisrimera with anti-inflammatory activity
.P.A. Nogueiraa, P.A. Reisa, G.A.T. Laranjaa, A.C. Pintoa, C.A.F. Aiubb, I. Felzenszwalbb,.C. Paesa, F.F. Bastosa, V.L.F.C. Bastosa, K.C.C. Sabinoa, M.G.P. Coelhoa,∗
Departamento de Bioquímica, Instituto de Biologia Roberto Alcantara Gomes, Centro Biomédico, Universidade do Estado do Rio de Janeiro, Av. Professor Manoel de Abreu, 444, PAPC,o andar, CEP 20550-170, Rio de Janeiro, RJ, BrazilDepartamento de Biofísica e Biometria, Instituto de Biologia Roberto Alcantara Gomes, Centro Biomédico, Universidade do Estado do Rio de Janeiro, Av. Professor Manoel de Abreu,44, PAPC, 4o andar, CEP 20550-170, Rio de Janeiro, RJ, Brazil
r t i c l e i n f o
rticle history:eceived 11 May 2011eceived in revised form 5 September 2011ccepted 8 September 2011vailable online 12 October 2011
eywords:accharis trimeraytotoxicityenotoxicityutagenicity
a b s t r a c t
Ethnopharmacological relevance: Baccharis trimera (Less) DC. (Asteraceae), popularly known in Brazil as“carqueja”, have been used in folk medicine to treat gastrointestinal, hepatic and renal diseases, andinflammatory processes as rheumatism.Aim of the study: To evaluate the in vitro and in vivo toxicological effects of anti-inflammatory Baccharistrimera aqueous extract and fractions.Materials and methods: Aqueous extract of Baccharis trimera (AEBt) was produced by infusion in boilingwater. After lyophylization AEBt was extracted with 80% ethanol, originating the ethanolic supernatantfraction (EFBt) and the aqueous sediment fraction (AFBt). Anti-inflammatory properties of AEBt, EFBtor AFBt (3, 30 or 300 �g/kg b.w.) were evaluated by the carrageenan-induced mouse paw edema usingindomethacin (10 mg/kg) as positive control. The growth of rat hepatoma cells (HTC) and human embryokidney epithelial cells (HEK) was determined by protein staining assay. Cytotoxicity was assayed by thetetrazolium salt (MTT) reduction. Cyclosporin was used as reference cytotoxic drug for spleen cells anddoxorubicin for HTC and HEK cells. For in vivo toxicological evaluation SW male mice were daily and oral(gavage) treated with extract/fractions at 4.2 mg/kg or 42 mg/kg during 15 days. After treatment liver orkidney cells were submitted to comet assay to determine the DNA damage index, and the glutathioneS-transferase activity was assayed towards ETHA (class Pi) and CDNB (several classes). Mutagenicity wasevaluated by the Ames test using Salmonella typhimurium strains TA97, TA98, TA100, and TA102.Results: The anti-inflammatory effects of EFBt were higher than those of AEBt or AFBt. Mice treat-ment (3–300 �g/kg) with AFBt reduced the paw edema (3 h) at lower levels (29.2–37.3%; P < 0.01), thanthose observed for AEBt (44.7–54.2%; P < 0.001), EFBt (49.3–58.2%; P < 0.001) or indomethacin (64.6%,P < 0.001, 10 mg/kg). The growth of kidney cells (HEK) was inhibited by AEBt (IC50 182.6 �g/ml), EFBt(IC50 78.1 �g/ml) and AFBt (IC50 86.2 �g/ml), with lower effects on HTC hepatic cell (IC50 308.8 �g/ml,396.5 �g/ml and 167.9 �g/ml, respectively). As evaluated by MTT test, AFBt exhibited cytotoxicity for HEKcells (IC50 372.5 �g/ml), but none for HTC ones; by the way, AFBt stimulated spleen cells (EC50 2.2 �g/ml)while cyclosporine, a cytotoxic reference drug inhibited them with IC50 of 0.42 �g/ml; the IC50 for doxoru-bicin for HEK and HTC cells was 0.28 �g/ml and 14.4 �g/ml, respectively, at 96 h. No mutagenic potentialwas observed. Mice treatment with AEBt or AFBt at 42 mg/kg for 15 days altered the kidney relativeweight, but not at 4.2 mg/kg. Baccharis trimera did not change liver, spleen or popliteal lymph node rela-
tive weight. DNA damage index of kidney cells was observed on mice treated with AEBt/AFBt, but not onanimals treated with EFBt, while DNA lesions were detected on liver cells only after AFBt treatment. Thegeneral activities of hepatic GST and Pi GST were reduced by EFBt and AFBt treatment, respectively.Conclusions: Baccharis trimera did not show mutagenicity, inhibited the GST activity, a hepatic detox-ification enzyme, and induced in vivo (genotoxicity) and in vitro toxicological effects to kidney cells.© 2011 Elsevier Ireland Ltd. All rights reserved.
∗ Corresponding author. Tel.: +55 21 2587 6143; fax: +55 21 2587 6136.E-mail addresses: [email protected], [email protected] (M.G.P. Coelho).
378-8741/$ – see front matter © 2011 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.jep.2011.09.051

5 thnop
1
Akttpei(
Beerr2lBaip
fmRB(2tipilar(cgK
ieiTi(cG2fPca
tppvfttcop
14 N.P.A. Nogueira et al. / Journal of E
. Introduction
Baccharis trimera (Less) DC. (Asteraceae), a widespread Southmerican woody perennial shrub, 0.5–4.0 m in height is popularlynown in Brazil as “carqueja”. Flower and leave teas of Baccharisrimera and Baccharis genistelloides, have been used in folk medicineo treat gastrointestinal, liver and renal diseases, and inflammatoryrocesses as rheumatism (Pio Correa, 1984; Sousa et al., 1991; Verdit al., 2005). The popular use of Baccharis trimera consists of drink-ng 50–200 ml/day of an aqueous infusion (4–5 g) of the dried herbCoimbra, 1942).
Several species of the Baccharis genus as Baccharis articulate andaccharis crispa (Gené et al., 1992), Baccharis trinervis (De las Herast al., 1998), Baccharis calliprinos and Baccharis rethinoides (Gianellot al., 1999), Baccharis grisebachii, Baccharis incarum and Baccha-is latifolia (Perez-García et al., 2001), Baccharis medullosa and B.ufescens (Cifuente et al., 2001), Baccharis pentlandii (Parejo et al.,003), Baccharis grisebachii (Tapia et al., 2004), Baccharis genistel-
oides (Coelho et al., 2004), Baccharis obtusifolia, Baccharis latifolia,accharis pentlandii and Baccharis subulata (Abad et al., 2006),s well as Baccharis trimera (Gené et al., 1992, 1996) have beennvestigated for their chemical composition and anti-inflammatoryroperties.
On the other hand, toxic effects to animals have been reportedor Baccharis halimifolia, Baccharis cordifolia DC. and Baccharisegapotamica Sprengel (Manley et al., 1982; Jarvis et al., 1996;izzo et al., 1997) and toxicological studies have been evaluated foraccharis trinervis (Sanchez-Palomino et al., 2002), Baccharis illinitaBaggio et al., 2003), and Baccharis genistelloides (Coelho et al.,004). Baccharis trimera toxicological effects have been reported forhe hydroethanolic extract of the plant (Grance et al., 2008), whichnduced in vivo toxicity (8.4 mg/kg) for kidney and liver cells ofregnant rats; although such alterations are reversible once admin-
stration is discontinued. No genotoxic effects for blood cells oriver were observed after treatment of mice with Baccharis trimeraqueous extract, but the frequency of micronucleus in bone mar-ow cells increased, indicating a chromosomal mutagenic activityRodrigues et al., 2009). Nevertheless, antimutagenic effect of Bac-haris trimera has been described and attributed for the flavonesenkwanin, cirsimaritin, hispidulin, and apigenin (Nakasugi andomai, 1998).
The multigene family of glutathione S-transferases (GST)soenzymes acts by binding together reduced glutathione withlectrophilic metabolites formed by biotransformation, produc-ng conjugates generally more water soluble and less cytotoxic.his enzyme activity helps cells to detoxify harmful compounds,ncluding those sourced from endogenous reactive oxygen speciesNordberg and Arn’er, 2001) and environmental exogenous toxi-ants (Hayes and Pulford, 1995). The activity of different classes ofST isoenzymes may be assayed using different substrates, such as,4 dichloro-nitrobenzene (CDNB), a rather nonspecific substrate,or several GST classes (general activity); while GST class Pi (GST-i), the major class in the mouse liver (Raza et al., 1991), has aomparatively high activity with ethacrynic acid (ETHA) (McLellannd Hayes, 1987; Egaas et al., 1995).
Although aqueous preparations of Baccharis trimera, as decoc-ion and infusion, have been widely used to treat differentathologies in Brazil, there are still few reports of their toxicologicalotential. Thus, the aim of this work was to study the in vitro and inivo toxicological effects of Baccharis trimera aqueous extract andractions, both presenting anti-inflammatory properties. The in vivooxicity of Baccharis trimera was evaluated by its genotoxic poten-
ial for kidney and liver cells and by the level of general and Pi classytosolic GST activities in the liver after oral and daily treatmentf mice with the Baccharis trimera samples for 15 days. The sam-les effects on mutagenicity (Salmonella/mammalian microsomeharmacology 138 (2011) 513– 522
and survival assays) and on liver, kidney and immune system cellsin vitro toxicity were also evaluated.
The effects of these samples on mutagenic test (Salmonellamicrosome/mammals and tests of survival) and in vitro toxicitywith cells from liver, kidneys and immune system were also eval-uated.
2. Materials and methods
2.1. Chemicals and reagents
MTT (3-[4,5-dimethylthiazole-2-il]-2,5-dipheniltetrazoliumbromide), 2-anthramine (2-AA), 2-aminofluorene (2AF), mito-mycin C, 4-nitroquinoline 1-oxide (4NQNO), Benzo [a]pyren (BaP),1-chloro-2,4-dinitrobenzene (CDNB), ethacrynic acid (ETHA) andDulbecco’s modified Eagle’s medium (DMEM), were purchasedfrom Sigma Chemical Co. (USA) and dimethyl sulfoxide (DMSO)from Merck Indústria Química (Brazil).Vogel-Bonner E mediumand Bacto Nutrient Broth were purchased from Difco, BD (USA).Fetal bovine serum (FBS) was obtained from Gibco BRL (USA)and 1,2-dichloro-4-nitrobenzene (DCNB) was purchased fromFluka. All other chemicals and reagents were of the highest gradeavailable.
2.2. Preparation of plant extract and fractions
The Baccharis trimera specie was collected on August, 2001, inFriburgo, Rio de Janeiro, Brazil. A voucher sample was deposited(HB 86447) at the herbarium Bradeanum (Universidade do Estadodo Rio de Janeiro, Rio de Janeiro, Brazil). Aerial parts of the dry plantwere extracted by infusion (100 g/3 l) in boiling water for 45 minwith agitation. Then, it was filtered and lyophilized (Heto Drywin-ner, Denmark), yielding about 8% of the initial mass (AEBt). Thisprocedure was repeated five times. After homogeneity analysis ofindividual extracts by HPLC (Shim-pack C-8 column), they werepolled and stored at −20 ◦C. AEBt was submitted to extractions with80% ethanol, originating the ethanolic supernatant fraction (EFBt)and the aqueous sediment fraction (AFBt), yielding 76% and 24% ofAEBt initial mass, respectively.
2.3. Animals
Male Swiss Webster (SW) mice, 29–35 g body weight (b.w.), fedad libitum with a commercial rodent diet (Nuvilab Ltda., Curitiba,Brazil) and free access to drinking water were used in all exper-iments. For each experiment, mice were randomly selected intogroups (n = 6/group). All experiments were performed under theconsent and surveillance of the Ethical Committee for animals usein research of Biomedical Center, Universidade do Estado do Rio deJaneiro, Brazil (CEA-IBRAG/protocol 05/2009).
2.4. Carrageenan-induced mouse paw edema
The carrageenan assay was carried out according to Levy (1969)with modifications. Male SW mice, fasted for 1 h with free access towater, were randomly selected to perform the study groups: con-trol (vehicle); indomethacin (10 mg/kg b.w.); AEBt, EFBt or AFBt(3, 30 or 300 �g/kg b.w.). The extracts were dissolved in NaCl 0.9%(AEBt and AFBt) or ethanol 15% containing 1.25% Tween 20 (EFBt).One hour after the intraperitoneal (i.p.) administration of test solu-tions or of the appropriated vehicle (control with NaCl 0.9% orethanol 15% with 1.25% Tween 20), edema was induced by a sub
plantar injection of 50 �l of 0.6 g% (w/v) carrageenan suspension inNaCl 0.9% into the right hindpaw of each mouse. The swelling ofthe hindpaw was measured after 3 h (peak of edema) in a plethys-mometer (7150 Ugo Basile), being compared with the volume of
thnop
tTer
2
2
l(tscmMatt0w5(ad
2
(bmipw2(taf5IcapmTdt
2
3flgi(9ab
2
2
w
N.P.A. Nogueira et al. / Journal of E
he same foot prior to the carrageenan challenge (edema index).he results were analyzed using the mean of edema index ± SD ofach group and expressed as percentage of paw edema inhibitionelated to control group.
.5. In vitro toxicological evaluations
.5.1. Mammalian cell lines and murine splenocytesThe rat hepatoma cells (HTC), and human embryo kidney epithe-
ial cells (HEK), were purchased from Rio de Janeiro Cell BankBCRJ/UFRJ, Rio de Janeiro, Brazil). These adherent lines were rou-inely grown in Dulbecco’s modified Eagle’s medium (DMEM)upplemented with 10% fetal bovine serum (FBS), 100 IU/ml peni-illin, and 100 �g/ml streptomycin (supplemented DMEM), beingaintained at 37 ◦C in 7% CO2 by standard cell culture techniques.urine lymphocytes were isolated from SW male mice after euth-
nize with a CO2 chamber. The spleen was aseptically removed andhe isolated splenocytes suspended in 2 ml of RPMI-1640 medium,reated with hypotonic ammonium chloride buffer (0.16 M NH4Cl,.1 mM EDTA and 10 mM KHCO3, pH 7.4) for haemolysis, washedith RPMI 1640 containing 2 mM EDTA, centrifuged (400 × g,
min), and then, resuspended in supplemented RPMI medium5% FBS, 2 mM l-glutamine, 5 × 10−5 M 2-mercaptoethanol andntibiotics) and cultured at 37 ◦C with 5% CO2. Cell viability wasetermined by Trypan Blue dye (0.2%) exclusion.
.5.2. Cytotoxicity by MTT reduction assayAfter culture of HEK, HTC (5.0 × 105 cells/ml) or spleen cells
2.0 × 106 cells/ml; activated with Con A 10 �g/ml) in 96 flat-ottom well plates (Costar 3596, Cambridge, MA) in supplementededia (200 �l) for 72 h (splenocytes) or 96 h (HEK and HTC),
n the presence or not (controls) of different concentrations oflant preparations, the tetrazolium salt (MTT) reduction by cellsas determined according to Mossman (1983). After cell culture,
0 �l/well of MTT solution (5 mg/ml in phosphate buffered salinePBS), pH 7.4) were added to cells and then incubated for addi-ional 2 h at 37 ◦C under CO2 atmosphere. DMSO (200 �l/well) weredded to plate, after removal of 150 �l of culture supernatant, forormazan crystals dissolution and the absorbance determined at70 nm using a microplate reader (�Quant, Bio-Tek Instrumentsnc., USA). Doxorubicin was used as cytotoxic drug for tumoral lineells, and cyclosporine for spleen cells. The mitochondrial reductionctivity (MRA) was calculated as percent of control (no addition oflant sample). Incubation of AEBt, EFBt or AFBt with MTT in supple-ented media without cells showed no significant MTT reduction.
he formazan production is proportional to the total mitochondrialehydrogenase activity in the well, which in turn correlates withhe total number of viable cells.
.5.3. Cell growth determinationAfter culture of HTC (9.0 × 105) or HEK (6.75 × 105) cells onto
5 mm diameter culture dishes (Nunc) in supplemented DMEMor 24 h, medium was renewed and AEBt, EFBt or AFBt, solubi-ized in the same medium, added at different concentrations. Cellrowth was determined daily by cell fixation with 5% TCA and stain-ng with 1% bromophenol blue (BPB) in 1% acetic acid for 30 minLopes et al., 2000). Samples of 200 �l were then transferred to6 flat-bottom well plates (Costar 3596, Cambridge, MA) and thebsorbance determined at 570 nm by a microplate reader. Doxoru-icin was also used as reference drug.
.6. In vivo toxicological evaluation
.6.1. Treatment schedule and organ weights determinationsGroups of six animals were daily treated by gavage (100 �l)
ith AEBt/AFBt/EFBt at 4.2 mg/kg or 42 mg/kg during 15 days. The
harmacology 138 (2011) 513– 522 515
animals in the control group received only the vehicle. At the endof experiment all animals were submitted to autopsy for exami-nation of the anatomical localization and pathological changes ofthe organs. Selected organs like spleen, popliteal lymph node, liverand kidney were excised, trimmed of fat and connective tissue, andhad the organ-to-body weight ratio calculated (g organ or mg forpopliteal lymph node/g body weight × 100).
2.6.2. Single cell gel electrophoresis assay (comet assay)The alkaline version of the comet assay detects single and double
strand breaks in DNA (Fairbairn et al., 1995). To detect these lesions,10 �l of liver or kidney cell suspension from SW male mice weremixed with 120 ml of 0.5% low-melting-point agarose in PBS andadded to microscope slides pre-coated with 1.5% normal-melting-point agarose in PBS. Slides were covered with a microscope coverslip and keep at 4 ◦C for 5 min to gel, followed by immersion in ice-cold alkaline lysis solution (2.5 M NaCl, 10 mM Tris, 100 mM EDTA,20% DMSO, 2% Triton X-100, pH 10.0) for 1 h. Slides were thenincubated for 20 min in ice-cold electrophoresis solution (0.2 MNaOH, 1 mM EDTA), followed by electrophoresis at 25 V/300 mA,for 25 min. After electrophoresis, slides were rinsed with water,allowed to dry at 37 ◦C and was stained with ethidium bromide20 �g/ml. DNA of individual cells was viewed using an epifluo-rescence microscope (Olympus), with 516–560 nm emission froma 50-W mercury light source, and image magnification of 200×.Quantification of DNA breakage was achieved by visual scoringof 50 randomly-selected cells per slide, classifying them into fivecategories based on the migration length and/or the perceivedrelative proportion of DNA in the tail. These five categories rep-resent different degrees of DNA damage, allowing for qualitativeevaluation, ranging from the absence of comet (type 0, undam-aged cells) to maximum length comet (type 4, maximally damagedcells). Comets of type 1 are representative of cells with a mini-mal detectable frequency of DNA lesions while comets of types 2and 3 are representative of cells with moderate-low and moderate-high frequency of DNA lesions, respectively. Cellular comets wereclassified by investigators blinded to the animal experimental con-ditions from which the tissue samples were obtained. The resultsare expressed as DNA damage index, calculated as [(cell numberof score 0 × 0) + (cell number of score 1 × 1) + (cell number of score2 × 2) + (cell number of score 3 × 3) + (cell number of score 4 × 4)];the results can run between 0 and 200.
2.6.3. Glutathione S-transferase (GST) assaysConsidering that GST-Pi is the major class in mouse liver and a
number of GST isoenzymes from liver are well known for conjugat-ing CDNB, we assayed GST activities towards ETHA (class Pi) andCDNB (several classes). Activities of GST were determined in thehepatic cytosolic fraction (a 100,000 × g supernatant from homog-enized liver) in 0.1 M Na-phosphate buffer containing 1 mM EDTA,following Habig et al. (1974) and Egaas et al. (1995) with minormodifications. The optimum pH for CDNB was 7.5, whereas ETHAwas best conjugated at pH 6.5. The general GST activity was deter-mined using 10 �g of protein, 1 mM CDNB in ethanol 4% and 1 mMGSH. The activity towards ETHA was determined with 500 mg ofprotein, 0.25 mM GSH and 0.2 mM ETHA. Every assay was carriedout in triplicate with correction for non-enzymatic conjugation(Egaas et al., 1995). Protein concentration was determined accord-ing to Peterson (1977) using bovine serum albumin as standard(10–100 �g).
2.7. Salmonella/mammalian microsome and survival assays
The Salmonella typhimurium strains TA97, TA98, TA100, andTA102 described by Maron and Ames (1983) were provided by Dr.B.N. Ames (University of California, Berkeley, CA, USA) and their

516 N.P.A. Nogueira et al. / Journal of Ethnopharmacology 138 (2011) 513– 522
Table 1Salmonella typhimurium strains used in this study.
Strain Relevant genotype Kind of effect observed References
TA97 hisD 6610 pKM101 �uvrB rfa Frameshift mutation Levin et al. (1982)TA98 hisD 3052 pKM101 �uvrB rfa Frameshift mutation Ames et al. (1975)TA100 hisG 46 pKM101 �uvrB rfa Base-pair substitution mutation Ames et al. (1975)TA102 hisG 428 (pAQ1) pKM101 rfa Base-pair substitution mutation Levin et al. (1982)
T nce gene; rfa mutations cause partial loss of the lipopolysaccharide barrier and an increasei s the nitrate reductase (chl) and biotin (bio) genes.
gppfaAsm
2
MTtTt(oomahaaTcitdea(rco
2
s2te7isewtt
2
ep
Fig. 1. Effects of AEBt, its fractions and indomethacin (INDO) on carrageenan-induced mouse paw edema. Hindpaw edema was induced 1 h after i.p.administration of test solutions or vehicle by sub plantar injection of carrageenan(0.6%, w/v in NaCl 0.9%). Foot paw volume was measured at the peak of edema (3 h)by plethysmography. Each point represents the mean inhibition index of hindpawedema of each group (n = 6/group) calculated as described in Section 2. The edema
he multicopy plasmid pAQ1 carries the hisG428 mutation and a tetracycline resistan permeability to large molecules. The deletion (�) through uvrB gene also include
enetic characteristics are listed in Table 1. The S9 fraction, pre-ared from the liver of Sprague-Dawley rats pretreated with aolychlorinated biphenyl mixture (Aroclor 1254), was purchasedrom Molecular Toxicology Inc. (Moltox TM, USA). The S9 metabolicctivation mixture (S9 mix) was prepared according to Maron andmes (1983) and was used in Salmonella/microsome tests andurvival assays as described elsewhere (Lopes et al., 2000), withodifications.
.7.1. Salmonella/mammalian microsome assayMutagenicity was measured using the procedure described by
aron and Ames (1983) with Salmonella typhimurium strains TA97,A98, TA100, and TA102 preincubated with different concentra-ions of extract/fractions Baccharis trimera with or without S9 mix.he assay mixture consisted of 100 �l extract/fractions Baccharisrimera (final concentration of 1, 10 or 100 �g/ml), 500 �l S9 mix2.4, 4.78, 9.55, and 19.1 mg total protein/ml) or the same volumef 0.2 M sodium phosphate buffer, pH 7.4, in experiments with-ut S9 mix, and 100 �l bacterial suspension (2 × 109 cells/ml). Theixture was preincubated at 37 ◦C with shaking. After 20, 60, 90
nd 120 min, 2 ml top agar (0.6% agar, 0.6% NaCl w/v, 50 �M l-istidine, 50 �M biotin, pH 7.4, 45 ◦C) was added to the test tubend the final mixture was poured onto a Petri dish with minimalgar (1.5% agar, Vogel-Bonner E medium, containing 2% glucose).he plates were incubated at 37 ◦C for 72 h, and the His + revertantolonies were counted. The results were expressed as a mutagenicndex (MI = number of His + induced in the sample/number of spon-aneous His + in the negative control). Positive controls were asescribed elsewhere (Aiub et al., 2003). Mutagenicity was consid-red positive when: (a) the number of revertant colonies in thessay was at least twice the number of spontaneous revertantsMI ≥ 2), (b) analysis of variance (ANOVA) revealed a significantesponse (P ≤ 0.05), and (c) a reproducible positive dose–responseurve (P ≤ 0.01) was present. This evaluation was based on previ-usly established criteria (Vargas et al., 1993, 1995).
.7.2. Survival experimentsTo determine the cytotoxicity, cultures of each bacterial
train were preincubated in a final volume of 700 �l (100 �l of × 109 cells/ml; 100 �l of 0.9% NaCl or extract/fractions Baccharisrimera for final concentration of 1, 10 or 100 �g/ml; and 500 �l ofach concentration of S9 mix or 0.2 M sodium phosphate buffer, pH.4). After 60, 90 and 120 min, the cells were washed and diluted
n 0.9% NaCl to obtain a suspension containing 2 × 103 cells/ml. Auitable aliquot (100 �l) of this suspension was plated on nutri-nt agar (0.8% Bacto nutrient broth, 0.5% NaCl and 1.5% agar), afterhich the plates were incubated at 37 ◦C for 24 h and the colonies
hen counted. Each experiment was repeated at least twice, withriplicate.
.8. Statistics
The results were reported as mean ± SD. Variance analysis wasstablished by one-way ANOVA. Significant differences betweenairs of groups were calculated using Tukey’s multiple comparison
index of control groups were: 100.6 ± 5.9 (NaCl 0.9%) and 104.1 ± 16.2 (ethanol 15%with 1.25% Tween 20). bP < 0.01, and cP < 0.001, related to control group (vehicle) byTukey’s test.
tests or Student’s t-test, with significance level reported at P ≤ 0.05,as indicated. When appropriate, the mean effective concentra-tion IC50 values (concentration that reduces 50% of control groupresponse) or EC50 (concentration that stimulate 50% of controlgroup response) were determined by non-linear regression usingGraph Pad Prism, 5.0 (GraphPad Software Inc., San Diego, CA, USA).
3. Results
3.1. Antiedematogenic activity
The aqueous extract of Baccharis trimera (AEBt), and its ethanolic(EFBt) and aqueous (AFBt) fractions, exhibited significantly inhibi-tion of the paw edema induced by carrageenan for all doses tested,related to the control group (Fig. 1). The treatment with differentdoses (3–300 �g/kg) of AFBt reduced the paw edema at 3 h (peakof edema in the control group, vehicle) at lower levels (29.2–37.3%;P < 0.01), than that observed for AEBt (44.7–54.2%; P < 0.001) andEFBt (49.3–58.2%; P < 0.001). Indomethacin (10 mg/kg), used as acontrol drug, reduced (P < 0.001) 64.6% of the edema at 3 h (datanot shown).
3.2. In vitro toxicological effects on mammalian cells
The toxicological evaluations were initiated by the in vitroassays. The effects of AEBt, EFBt or AFBt on human embryo kid-
ney epithelial cells (HEK) and rat hepatoma cells (HTC) growthwere studied (Figs. 2 and 3). Different behavior of cell growth wasobserved for the cell lines along the culture. No cytotoxic effect forHEK cells was shown with AEBt up to 100 �g/ml (Fig. 2A), while
N.P.A. Nogueira et al. / Journal of Ethnopharmacology 138 (2011) 513– 522 517
0 24 48 72 96 120
0
10
20
30
40 Control1 μg/ml10 μg/ml100 μg/ml500 μg/ml
c c c c
AEBt
Time (h)
HE
K c
ell g
row
th(A
bs 5
70 n
m)
0 24 48 72 96 120
0
5
10
15Control1 μg/ ml
10 μg /ml50 μg/ ml100 μg/ ml
a c c c
Time (h)
HE
K c
ell g
row
th(A
bs 5
70 n
m)
EFBt
0 24 48 72 96 120
0
5
10
15 Control1 μg/ ml10 μg/ ml50 μg/ ml100 μg/ ml
c cc
Time (h)
HE
K c
ell g
row
th(A
bs 5
70 n
m)
AFBt
A
C
B
Fig. 2. Growth kinetics of HEK cells in the presence of AEBt (A), EFBt (B) and AFBt(C). Exponentially growing cells were cultured in the absence (control cells) or in thepresence of different concentrations of extracts/fractions. At the indicated times, thetotal cells number was determined by bromophenol blue staining (BPB) of proteins,as described in Section 2. Each point represents the mean absorbance at 570 nm ± SDof culture duplicates (representative of three experiments). The calculated IC50 at9c
sbol2trl5ecioiOo3ard
0 24 48 72 96 120
0
5
10
15
20 Control1 μg/ ml10 μg/ ml100 μg/ ml500 μg/ml
bb
a a
AEBt
Time (h)
HT
C c
ell g
row
th(A
bs 5
70 n
m)
0 24 48 72 96 120
0
5
10
15
20 Control1 μg/ ml10 μg/ ml50 μg/ ml100 μg/ml
EFBt
Time (h)
HT
C c
ell g
row
th(A
bs 5
70 n
m)
0 24 48 72 96 120
0
5
10
15
20 Control1 μg/ ml10 μg/ ml50 μg/ ml100 μg/ml
a
AFBt
Time (h)
HT
C c
ell g
row
th(A
bs 5
70 n
m)
A
B
C
Fig. 3. Growth kinetics of HTC cells in the presence of AEBt (A), EFBt (B) and AFBt (C).Exponentially growing cells in DMEM with 10% FBS were cultured in the absence(control cells) or in the presence of different concentrations of extracts/fractions. Atthe indicated times, the total number of cells were determined by bromophenol bluestaining (BPB) of proteins, as described in methods. Each point represents the meanabsorbance at 570 nm ± SD of culture duplicates (representative of three experi-
6 h were 182.6 �g/ml (AEBt), 78.1 �g/ml (EFBt) and 86.2 �g/ml (AFBt). aP < 0.05,P < 0.001, related to control culture by Tukey’s test.
ignificant inhibition can be observed at 500 �g/ml, reaching inhi-ition rates of 94.3% just after 24 h of culture (P < 0.001), with IC50f 182.6 �g/ml. EFBt (Fig. 2B) and AFBt (Fig. 2C) inhibited cell pro-iferation at the higher tested concentration (100 �g/ml) just after4 h of culture, with reduction indexes (P < 0.001) after 96 h of cul-ure of 94.9% and 79.9% and IC50 of 78.1 �g/ml and 86.2 �g/ml,espectively. The AEBt (Fig. 3A) has inhibited HTC cell line pro-iferation only at 500 �g/ml, exhibiting cell growth inhibition of7.3% (P < 0.001), after 96 h of culture and IC50 308.8 �g/ml. Differ-nt from HEK cells, EFBt induced no significant difference on HTCells proliferation (Fig. 3B) with IC50 396.5 �g/ml while AFBt inhib-ted HTC cell growth only at 96 h of culture (Fig. 3C) with reductionf 53.9% (P < 0.05) and IC50 167.9 �g/ml. The EFBt or AFBt cytotox-city was also studied on HEK, HTC and mice spleen cells (Fig. 4).nly the AFBt fraction induced cytotoxic effect (viability inhibitionf 30%) and only HEK cells were sensitive to it (Fig. 4C), with IC50
72.5 �g/ml. Doxorubicin, used as positive control drug, exhibitedt 96 h IC50 14.4 �g/ml and 0.28 �g/ml for HTC cells and HEK cells,espectively. By the way, AFBt treatment stimulated mitochon-rial reduction activity of spleen cells in a concentration dependentments). The calculated IC50 at 96 h were 308.8 �g/ml (AEBt), 396.5 �g/ml (EFBt) and167.9 �g/ml (AFBt). aP < 0.05, bP < 0.01, related to control culture by Tukey’s test.
manner (Fig. 4A), with EC50 2.2 �g/ml, reaching 104% of stimula-tion with 100 �g/ml, while no significant difference was observedwith any dose of EFBt. The IC50 of cyclosporine for spleen cells was0.42 �g/ml.
3.3. In vivo toxicological studies
3.3.1. Effects on the body and organ weightOral and daily administration of AEBt or its fractions to healthy
mice for 15 days did not induce clinical symptoms of toxicity ordeath (data not shown). All groups increased the body weight about3.2 ± 1.6 g during the treatment. Table 2 shows no significant dif-ference in the spleen or liver relative weights after treatment with4.2 mg/kg of AEBt or its fractions AFBt and EFBt related to the control
group. At the higher dose (42 mg/kg), slight variation on the kidneyrelative weights (P < 0.05) was observed in mice treated with AEBtor AFBt.
518 N.P.A. Nogueira et al. / Journal of Ethnopharmacology 138 (2011) 513– 522
Table 2Effects of the AEBt/fractions treatment on relative organ weights of healthy mice.
Group Relative weight (%)
Liver Kidneys Spleen aPopliteal lymph node
Control 6.7 ± 0.8 0.62 ± 0.03 0.40 ± 0.05 NDAEBt 4.2 mg/kg 6.7 ± 0.5 0.59 ± 0.02 0.37 ± 0.07 NDAEBt 42 mg/kg 5.9 ± 0.6 0.67 ± 0.04a,d 0.43 ± 0.04 NDControl 5.8 ± 0.3 0.55 ± 0.07 0.56 ± 0.05 7.17 ± 2.67AFBt 4.2 mg/kg 5.4 ± 0.5 0.50 ± 0.03 0.51 ± 0.10 13.46 ± 3.07AFBt 42 mg/kg 5.4 ± 0.4 0.46 ± 0.02a 0.44 ± 0.07 10.88 ± 4.42Control 5.4 ± 0.8 0.56 ± 0.03 0.57 ± 0.07 6.90 ± 4.80EFBt 4.2 mg/kg 5.8 ± 0.2 0.58 ± 0.02 0.53 ± 0.07 7.62 ± 2.23EFBt 42 mg/kg 5.5 ± 0.2 0.59 ± 0.05 0.64 ± 0.10 10.26 ± 0.11
Animals have been treated orally and daily for 15 days. Relative weight was calculated as organ weight (g or amg)/body weight (g) × 100 and are expressed as means ± SD(n = 6 per group). ND – not determined. aP < 0.05 vs. control group (vehicle) and dP < 0.01 vs. AEBt 4.2 mg group, by Student’s t-test.
0.1 1 10 1000
50
100
150
200
250 AFBt EFBt
ac c c
Cyclosporin
c
cc c
c
c
Concentration (μg/ml)
MTT
redu
ctio
n (%
)Sp
leen
cel
ls
0.1 1 10 100
0
50
100
150
200 AFBt EFBt
Concentration ( μg/ml)
MTT
redu
ctio
n (%
)H
TC c
ells
Doxorubicin
cc
c a
0.1 1 10 100
0
50
100
150
200 AFBt EFBt
a
cc
cc
Doxorubicin
Concentration ( μg/ml)
MTT
redu
ctio
n (%
)H
EK c
ells
A
B
C
Fig. 4. Mitochondrial reduction activity (MRA) of spleen cells (A), HTC cells (B),or HEK cells (C) in the presence of EFBt or AFBt. The cells were incubated in theabsence (control) or presence of different concentrations of AEBt fractions for 72 h(splenocytes) or 96 h (HEK and HTC). MRA was determined by the MTT assay, asdescribed in Section 2. EC50 AFBt 2.2 �g/ml and IC50 cyclosporine 0.42 �g/ml forspleen cells. IC50 doxorubicin 14.4 �g/ml and 0.28 �g/ml for HTC and HEK cells,respectivelly. No inhibition was observed for EFBt and AFBt for HTC or HEK cells.Td
3
t
Fig. 5. GST activity towards: (A) 1-chloro-2,4-dinitrobenzene (CDNB, total GSTactivity); (B) ethachrynic acid (ETHA, GST class Pi activity), in hepatic microsomes
3.3.3. Glutathione S-transferase levels
he results express the mean percentage of control ± S.D. of three experiments withuplicates. aP < 0.05, cP < 0.001, related to control culture by Tukey’s test.
.3.2. Single cell gel electrophoresis assayTo detect potential in vivo genotoxic effects of AEBt/fractions
o liver and kidney cells, the DNA of these tissues from healthy
from SW male healthy mice treated during 15 days with oral doses of AFBt and EFBt(n = 5/group). All analysis was performed in duplicate. aP < 0.05 vs. control; #P < 0.05vs. 4.2 mg/kg group by Tukey’s test.
animals treated with vehicle (control), AEBt, AFBt and EFBt wereexamined by the comet assay. We did not observe significant dif-ference in liver DNA damage index between groups treated withAEBt (Table 3), with predominance of undamaged cells (score 0)and low levels of maximal DNA lesions (score 4). However, AEBttreatment induced kidney DNA damage at 4.2 and 42 mg/kg. Thetreatment of healthy mice with AFBt at both doses increased thenumber of liver and kidney cell lesions when related to controland 4.2 mg/kg treated group (P < 0.05), as can be observed by theirhigher DNA damage index and the increase in the number of livercells with score 4. No significant differences in the kidney and liverDNA damage were observed on mice groups treated with EFBt inrelation to untreated groups.
As can be seen in Fig. 5A, the general GST activity (assayed withCDNB) was reduced 37.6% with the treatment of mice with 42 mgEFBt/kg (P < 0.05). Class Pi GST activity was reduced 47.8% in mice
N.P.A. Nogueira et al. / Journal of Ethnopharmacology 138 (2011) 513– 522 519
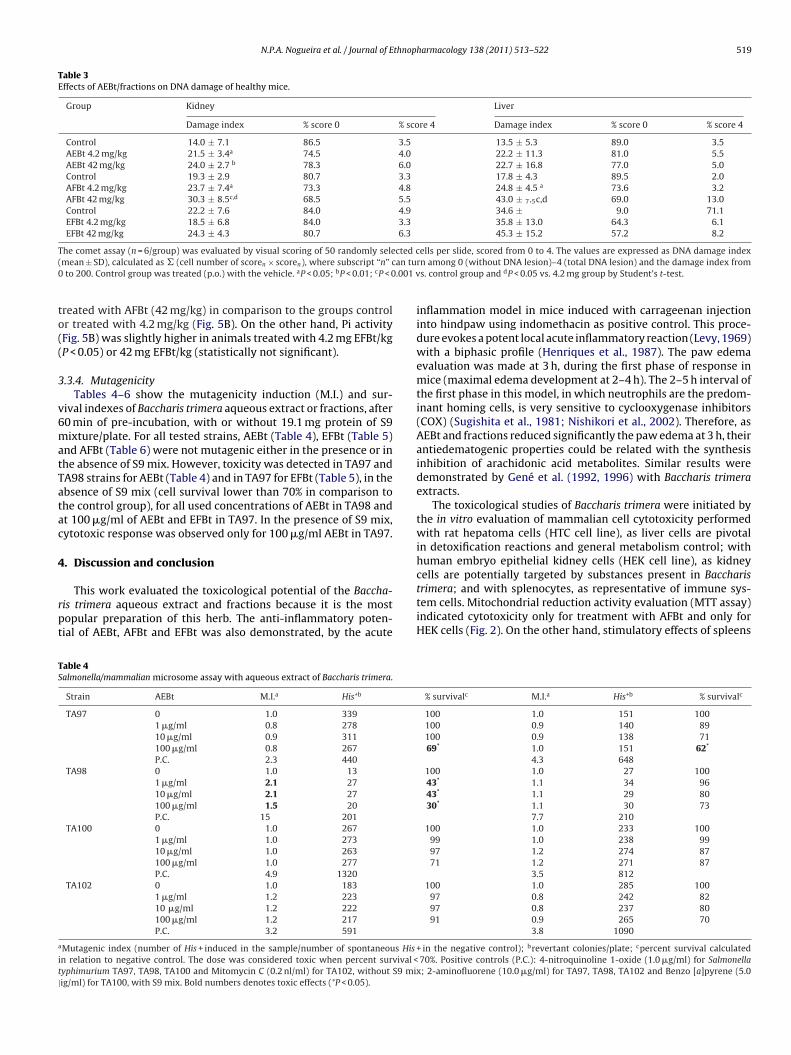
Table 3Effects of AEBt/fractions on DNA damage of healthy mice.
Group Kidney Liver
Damage index % score 0 % score 4 Damage index % score 0 % score 4
Control 14.0 ± 7.1 86.5 3.5 13.5 ± 5.3 89.0 3.5AEBt 4.2 mg/kg 21.5 ± 3.4a 74.5 4.0 22.2 ± 11.3 81.0 5.5AEBt 42 mg/kg 24.0 ± 2.7 b 78.3 6.0 22.7 ± 16.8 77.0 5.0Control 19.3 ± 2.9 80.7 3.3 17.8 ± 4.3 89.5 2.0AFBt 4.2 mg/kg 23.7 ± 7.4a 73.3 4.8 24.8 ± 4.5 a 73.6 3.2AFBt 42 mg/kg 30.3 ± 8.5c,d 68.5 5.5 43.0 ± 7.5c,d 69.0 13.0Control 22.2 ± 7.6 84.0 4.9 34.6 ± 9.0 71.1EFBt 4.2 mg/kg 18.5 ± 6.8 84.0 3.3 35.8 ± 13.0 64.3 6.1EFBt 42 mg/kg 24.3 ± 4.3 80.7 6.3 45.3 ± 15.2 57.2 8.2
The comet assay (n = 6/group) was evaluated by visual scoring of 50 randomly selected cells per slide, scored from 0 to 4. The values are expressed as DNA damage index( an tu0 .001 v
to((
3
v6matTatac
4
rpt
TS
a
it|
mean ± SD), calculated as � (cell number of scoren × scoren), where subscript “n” c to 200. Control group was treated (p.o.) with the vehicle. aP < 0.05; bP < 0.01; cP < 0
reated with AFBt (42 mg/kg) in comparison to the groups controlr treated with 4.2 mg/kg (Fig. 5B). On the other hand, Pi activityFig. 5B) was slightly higher in animals treated with 4.2 mg EFBt/kgP < 0.05) or 42 mg EFBt/kg (statistically not significant).
.3.4. MutagenicityTables 4–6 show the mutagenicity induction (M.I.) and sur-
ival indexes of Baccharis trimera aqueous extract or fractions, after0 min of pre-incubation, with or without 19.1 mg protein of S9ixture/plate. For all tested strains, AEBt (Table 4), EFBt (Table 5)
nd AFBt (Table 6) were not mutagenic either in the presence or inhe absence of S9 mix. However, toxicity was detected in TA97 andA98 strains for AEBt (Table 4) and in TA97 for EFBt (Table 5), in thebsence of S9 mix (cell survival lower than 70% in comparison tohe control group), for all used concentrations of AEBt in TA98 andt 100 �g/ml of AEBt and EFBt in TA97. In the presence of S9 mix,ytotoxic response was observed only for 100 �g/ml AEBt in TA97.
. Discussion and conclusion
This work evaluated the toxicological potential of the Baccha-is trimera aqueous extract and fractions because it is the mostopular preparation of this herb. The anti-inflammatory poten-ial of AEBt, AFBt and EFBt was also demonstrated, by the acute
able 4almonella/mammalian microsome assay with aqueous extract of Baccharis trimera.
Strain AEBt M.I.a His+b
TA97 0 1.0 339
1 �g/ml 0.8 278
10 �g/ml 0.9 311
100 �g/ml 0.8 267
P.C. 2.3 440
TA98 0 1.0 13
1 �g/ml 2.1 27
10 �g/ml 2.1 27
100 �g/ml 1.5 20
P.C. 15 201
TA100 0 1.0 267
1 �g/ml 1.0 273
10 �g/ml 1.0 263
100 �g/ml 1.0 277
P.C. 4.9 1320
TA102 0 1.0 183
1 �g/ml 1.2 223
10 �g/ml 1.2 222
100 �g/ml 1.2 217
P.C. 3.2 591
Mutagenic index (number of His + induced in the sample/number of spontaneous His
n relation to negative control. The dose was considered toxic when percent survival <yphimurium TA97, TA98, TA100 and Mitomycin C (0.2 nl/ml) for TA102, without S9 mixig/ml) for TA100, with S9 mix. Bold numbers denotes toxic effects (*P < 0.05).
rn among 0 (without DNA lesion)–4 (total DNA lesion) and the damage index froms. control group and dP < 0.05 vs. 4.2 mg group by Student’s t-test.
inflammation model in mice induced with carrageenan injectioninto hindpaw using indomethacin as positive control. This proce-dure evokes a potent local acute inflammatory reaction (Levy, 1969)with a biphasic profile (Henriques et al., 1987). The paw edemaevaluation was made at 3 h, during the first phase of response inmice (maximal edema development at 2–4 h). The 2–5 h interval ofthe first phase in this model, in which neutrophils are the predom-inant homing cells, is very sensitive to cyclooxygenase inhibitors(COX) (Sugishita et al., 1981; Nishikori et al., 2002). Therefore, asAEBt and fractions reduced significantly the paw edema at 3 h, theirantiedematogenic properties could be related with the synthesisinhibition of arachidonic acid metabolites. Similar results weredemonstrated by Gené et al. (1992, 1996) with Baccharis trimeraextracts.
The toxicological studies of Baccharis trimera were initiated bythe in vitro evaluation of mammalian cell cytotoxicity performedwith rat hepatoma cells (HTC cell line), as liver cells are pivotalin detoxification reactions and general metabolism control; withhuman embryo epithelial kidney cells (HEK cell line), as kidneycells are potentially targeted by substances present in Baccharis
trimera; and with splenocytes, as representative of immune sys-tem cells. Mitochondrial reduction activity evaluation (MTT assay)indicated cytotoxicity only for treatment with AFBt and only forHEK cells (Fig. 2). On the other hand, stimulatory effects of spleens% survivalc M.I.a His+b % survivalc
100 1.0 151 100100 0.9 140 89100 0.9 138 7169* 1.0 151 62*
4.3 648100 1.0 27 10043* 1.1 34 9643* 1.1 29 8030* 1.1 30 73
7.7 210100 1.0 233 100
99 1.0 238 9997 1.2 274 8771 1.2 271 87
3.5 812100 1.0 285 100
97 0.8 242 8297 0.8 237 8091 0.9 265 70
3.8 1090
+ in the negative control); brevertant colonies/plate; cpercent survival calculated 70%. Positive controls (P.C.): 4-nitroquinoline 1-oxide (1.0 �g/ml) for Salmonella; 2-aminofluorene (10.0 �g/ml) for TA97, TA98, TA102 and Benzo [a]pyrene (5.0
520 N.P.A. Nogueira et al. / Journal of Ethnopharmacology 138 (2011) 513– 522
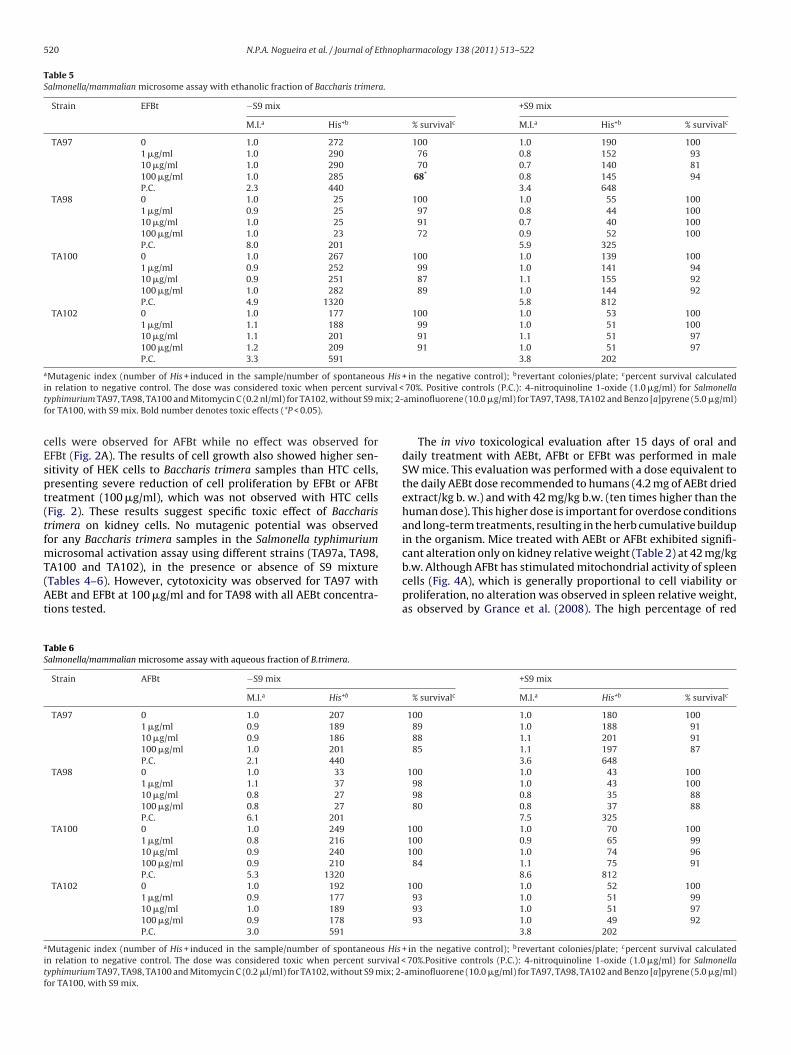
Table 5Salmonella/mammalian microsome assay with ethanolic fraction of Baccharis trimera.
Strain EFBt −S9 mix +S9 mix
M.I.a His+b % survivalc M.I.a His+b % survivalc
TA97 0 1.0 272 100 1.0 190 1001 �g/ml 1.0 290 76 0.8 152 9310 �g/ml 1.0 290 70 0.7 140 81100 �g/ml 1.0 285 68* 0.8 145 94P.C. 2.3 440 3.4 648
TA98 0 1.0 25 100 1.0 55 1001 �g/ml 0.9 25 97 0.8 44 10010 �g/ml 1.0 25 91 0.7 40 100100 �g/ml 1.0 23 72 0.9 52 100P.C. 8.0 201 5.9 325
TA100 0 1.0 267 100 1.0 139 1001 �g/ml 0.9 252 99 1.0 141 9410 �g/ml 0.9 251 87 1.1 155 92100 �g/ml 1.0 282 89 1.0 144 92P.C. 4.9 1320 5.8 812
TA102 0 1.0 177 100 1.0 53 1001 �g/ml 1.1 188 99 1.0 51 10010 �g/ml 1.1 201 91 1.1 51 97100 �g/ml 1.2 209 91 1.0 51 97P.C. 3.3 591 3.8 202
aMutagenic index (number of His + induced in the sample/number of spontaneous His + in the negative control); brevertant colonies/plate; cpercent survival calculatedi vival <t ix; 2-f
cEspt(tfmT(At
TS
a
itf
n relation to negative control. The dose was considered toxic when percent suryphimurium TA97, TA98, TA100 and Mitomycin C (0.2 nl/ml) for TA102, without S9 mor TA100, with S9 mix. Bold number denotes toxic effects (*P < 0.05).
ells were observed for AFBt while no effect was observed forFBt (Fig. 2A). The results of cell growth also showed higher sen-itivity of HEK cells to Baccharis trimera samples than HTC cells,resenting severe reduction of cell proliferation by EFBt or AFBtreatment (100 �g/ml), which was not observed with HTC cellsFig. 2). These results suggest specific toxic effect of Baccharisrimera on kidney cells. No mutagenic potential was observedor any Baccharis trimera samples in the Salmonella typhimurium
icrosomal activation assay using different strains (TA97a, TA98,
A100 and TA102), in the presence or absence of S9 mixtureTables 4–6). However, cytotoxicity was observed for TA97 withEBt and EFBt at 100 �g/ml and for TA98 with all AEBt concentra-ions tested.
able 6almonella/mammalian microsome assay with aqueous fraction of B.trimera.
Strain AFBt −S9 mix
M.I.a His+b
TA97 0 1.0 207
1 �g/ml 0.9 189
10 �g/ml 0.9 186
100 �g/ml 1.0 201
P.C. 2.1 440
TA98 0 1.0 33
1 �g/ml 1.1 37
10 �g/ml 0.8 27
100 �g/ml 0.8 27
P.C. 6.1 201
TA100 0 1.0 249
1 �g/ml 0.8 216
10 �g/ml 0.9 240
100 �g/ml 0.9 210
P.C. 5.3 1320
TA102 0 1.0 192
1 �g/ml 0.9 177
10 �g/ml 1.0 189
100 �g/ml 0.9 178
P.C. 3.0 591
Mutagenic index (number of His + induced in the sample/number of spontaneous His
n relation to negative control. The dose was considered toxic when percent survival <yphimurium TA97, TA98, TA100 and Mitomycin C (0.2 �l/ml) for TA102, without S9 mix; 2-or TA100, with S9 mix.
70%. Positive controls (P.C.): 4-nitroquinoline 1-oxide (1.0 �g/ml) for Salmonellaaminofluorene (10.0 �g/ml) for TA97, TA98, TA102 and Benzo [a]pyrene (5.0 �g/ml)
The in vivo toxicological evaluation after 15 days of oral anddaily treatment with AEBt, AFBt or EFBt was performed in maleSW mice. This evaluation was performed with a dose equivalent tothe daily AEBt dose recommended to humans (4.2 mg of AEBt driedextract/kg b. w.) and with 42 mg/kg b.w. (ten times higher than thehuman dose). This higher dose is important for overdose conditionsand long-term treatments, resulting in the herb cumulative buildupin the organism. Mice treated with AEBt or AFBt exhibited signifi-cant alteration only on kidney relative weight (Table 2) at 42 mg/kg
b.w. Although AFBt has stimulated mitochondrial activity of spleencells (Fig. 4A), which is generally proportional to cell viability orproliferation, no alteration was observed in spleen relative weight,as observed by Grance et al. (2008). The high percentage of red+S9 mix
% survivalc M.I.a His+b % survivalc
100 1.0 180 10089 1.0 188 9188 1.1 201 9185 1.1 197 87
3.6 648100 1.0 43 100
98 1.0 43 10098 0.8 35 8880 0.8 37 88
7.5 325100 1.0 70 100100 0.9 65 99100 1.0 74 96
84 1.1 75 918.6 812
100 1.0 52 10093 1.0 51 9993 1.0 51 9793 1.0 49 92
3.8 202
+ in the negative control); brevertant colonies/plate; cpercent survival calculated 70%.Positive controls (P.C.): 4-nitroquinoline 1-oxide (1.0 �g/ml) for Salmonellaaminofluorene (10.0 �g/ml) for TA97, TA98, TA102 and Benzo [a]pyrene (5.0 �g/ml)

thnop
bcttt
ttlDSposAbtffooh(cc
agwoctwtmicFmdpba
pt1aapGpeAr2tcoit2t
an
Gianello, J.C., Cifuente, D.A., Giordano, O.S., Tonn, C.E., 1999. Bioactive flavones andterpenes from Baccharis calliprinos and B. rethinoides (Asteraceae). Acta Farma-
N.P.A. Nogueira et al. / Journal of E
lood cells in spleen, compared to lymphocytes and macrophages,an be contributing for the non altered spleen relative weight. Theendency of lymph node to increase its relative weight reinforceshis hypothesis, since lymphocytes and antigen presenting cells arehe major cells in this lymphoid organ.
To evaluate if Baccharis trimera could contain substances poten-ially genotoxic to mammalian cells, DNA of liver and kidney cells ofhe treated animals was examined by the comet assay. In the alka-ine version of the comet assay, which detects a broad spectrum ofNA lesions, including double and single strand breaks (Brendler-chwaab et al., 2005), DNA evaluation of kidney cells indicated theresence of higher damage index in animals treated with both dosesf AEBt and AFBt, but none on those treated with EFBt. Liver cellshowed no detectable DNA lesions after treatment of animals withEBt or EFBt, while AFBt treatment induced genotoxic effect onoth doses. Thus, kidney cells showed higher in vivo sensitivityo AEBT genotoxic effects than liver ones. These results are rein-orced by the absence of genotoxic effect (comet assay) in liverrom mice treated orally for three consecutive days with 500, 1000r 2000 mg/kg of Baccharis trimera aqueous extract, and presencef toxic effects for kidney cells of pregnant rats orally treated withydroethanolic extract of Baccharis trimera (8.4 mg/kg) for 19 daysGrance et al., 2008). The results also indicate that AFBt seems tooncentrate the genotoxic agents of AEBt for both liver and kidneysells.
As the conjugation activity of xenobiotics with glutathione isn important detoxifying reaction, the Baccharis trimera effects oneneral and specific GST activities in the liver were evaluated in thisork. Although reduction in the hepatic GST-Pi activity has been
bserved after treatment of mice with 42 mg AFBt/kg (Fig. 5B), nohange was observed in the GST general activity with the samereatment (Fig. 5A). This finding indicates that the AFBt might beeakening cytosolic GST antioxidant activity in the liver of mice. On
he other hand, as hepatic GST-Pi activity was slightly increased inice treated with EFBt (Fig. 5B) and general GST activity was dimin-
shed after mice treatment with 42 mg EFBt/kg (Fig. 5A), one mayonjecture that GST activities other than Pi were reduced by EFBt.urthermore, the increase of GST-Pi activity after EFBt treatmentight be indicating that this Baccharis trimera fraction stimulates
etoxification and antioxidant activities. In this regard, it has beenublished that extracts from leaves of Ginkgo biloba, which haseen used for their high antioxidant property, also induced GST-Pictivity in human and mice hepatoma cell lines (Liu et al., 2009).
The flavonoids represent a large and important group ofolyphenolic compounds. They have been reported to haveherapeutic potential effects by their antioxidant (Chen et al.,990), anti-inflammatory, antihepatotoxic, antiulcer (Ferrandiznd Alcaraz, 1991), and antimutagenic (Edenhardder et al., 1993)ctivities. Furthermore, several works have been reporting thatlant polyphenols inhibit the GST activity (Kawabata et al., 2000;yamfi et al., 2004; Krajka-Kuzniak et al., 2008). Notably, a plantolyphenol, the tannic acid, injected i.p. in mice, decreased the gen-ral liver GST activity (Krajka-Kuzniak and Baer-Dubowska, 2003).s flavonoids have been reported in several species of the Baccha-is genus (Weimann et al., 2002; Akaike et al., 2003; Feresin et al.,003; Rodrigues et al., 2009), the anti-inflammatory action andhe inhibitory effects of Baccharis trimera on GST activities (Fig. 2)ould be at least partially attributed to these substances. However,ther polyphenolic compounds (tannins, saponins), alkaloids andsoprenoids (diterpenes, triterpenes) have also been described inhis specie (Simões-Pires et al., 2005; Verdi et al., 2005; Silva et al.,007; Lago et al., 2008). Current studies are on the way to identify
hese substances.In conclusion, the Baccharis trimera samples showed similarnti-inflammatory potential as indomethacin at lower doses, didot show mutagenicity, inhibited the activity of an important
harmacology 138 (2011) 513– 522 521
detoxification hepatic enzyme, the glutathione S-transferase, andinduced in vivo (genotoxicity) and in vitro toxicological effect forkidney cells. These results suggest that care should be taken interms of overdose and prolonged treatments, which result in accu-mulation of the medicinal herb in the organism.
Conflict of interest
The authors declare that there are no conflicts of interest.
Acknowledgments
We would like to thank the technical assistance from LIA-BPPN Laboratory. This work was supported by grants from theConselho Nacional de Desenvolvimento Científico e Tecnológico(CNPq), Fundac ão Carlos Chagas Filho de Amparo à Pesquisa doEstado do Rio de Janeiro (FAPERJ) and Universidade do Estado doRio de Janeiro (UERJ).
References
Abad, M.J., Bessa, A.L., Ballarin, B., Aragón, O., Gonzales, E., Bermejo, P., 2006. Anti-inflammatory activity of four Bolivian Baccharis species (Compositae). Journalof Ethnopharmacology 103, 338–344.
Aiub, C.A.F., Ribeiro Pinto, L.F., Felzenszwalb, I., 2003. N-Nitrosodiethylamine muta-genicity at low concentrations. Toxicology Letters 145, 36–45.
Akaike, S., Sumino, M., Sekine, T., Seo, S., Kimura, N., Ikegami, F., 2003. A new ent-clerodane diterpene from the aerial parts of Baccharis gaudichaudiana. Chemicaland Pharmaceutical Bulletin 51, 197–199.
Ames, B.N., McCann, J., Yamasaki, E., 1975. Methods for detecting carcinogensand mutagens with the Salmonella/mammalian-microsome mutagenicity test.Mutation Research 31, 347–364.
Baggio, C.H., Freitas, C.S., Rieck, L., Marques, M.C., 2003. Gastroprotective effects of acrude extract of Baccharis illinita DC in rats. Pharmacological Research 47, 93–98.
Brendler-Schwaab, S., Hartmann, A., Pfuhler, S., Speit, G., 2005. The in vivo cometassay: use and status in genotoxicity testing. Mutagenesis 20, 245–254.
Chen, Y.T., Zheng, R.L., Jia, Z.J., Ju, Y., 1990. Flavonoids as super oxide scavengers andantioxidant. Free Radical Biology and Medicine 9, 19–21.
Cifuente, D.A., Simirgiotis, M.J., Favier, L.S., Rotelli, A.E., Pelzer, L.E., 2001.Anti-inflammatory activity from aerial parts of Baccharis medullosa, Baccha-ris rufescens and Laennecia sophiifolia in mice. Phytotherapy Research 15,529–531.
Coelho, M.G.P., Reis, P.A., Gava, V.C.B., Gayer, C.R.M., Felzenszwalb, I., Sabino, K.C.C.,2004. Anti-arthritic effect and subacute toxicological evaluation of Baccharisgenistelloides aqueous extract. Toxicology Letters 154, 69–80.
Coimbra, R., 1942. Notas de fitoterapia. Catálogos dos dados principais sobre plantasutilizadas em medicina e farmácia, Araújo, C.S. Ed., 1st ed., p. 222.
De las Heras, B., Slowing, K., Benedi, J., Carretero, E., Ortega, T., Toledo, C., Bermejo,P., Iglesias, I., Abad, M.J., Gomez-Serranillos, P., Liso, P.A., Villar, A., Chiriboga,X., 1998. Anti-inflammatory and antioxidant activity of plant used in traditionalmedicine in Ecuador. Journal of Ethnopharmacology 61, 161–166.
Edenhardder, R., Von Petersdorff, I., Rauscher, R., 1993. Antimutagenic effects offlavonoids, chalcones and structurally related compounds on the activity of 2-amino-3-methylimidazole [4,5-f]quinoline (IQ) and other heterocyclic aminemutagens from cooked food. Mutation Research 287, 261–274.
Egaas, E., Falls, J.G., Dauterman, W.C., 1995. A study of gender, strain and age differ-ences in mouse liver glutathione S-transferase. Comparative biochemistry andphysiology. Part C. Pharmacology. Toxicology and Endocrinology 110, 35–40.
Fairbairn, D.W., Olive, P.L., O’neill, K.L., 1995. The comet assay: a comprehensivereview. Mutation Research 339, 37–59.
Feresin, G.E., Tapia, A., Gimenez, A., Ravelo, A.G., Zacchino, S., Sortino, M., Schmeda-Hirschmann, G., 2003. Constituents of the Argentinian medicinal plant Baccharisgrisebachii and their antimicrobial activity. Journal of Ethnopharmacology 89,73–80.
Ferrandiz, M.L., Alcaraz, M.J., 1991. Anti-inflammatory activity and inhibition ofarachidonic acid metabolism by flavonoid. Agents and Actions 32, 283–288.
Gené, R.M., Cartna, C., Adzet, T., Marín, E., Parella, T., Canigueral, S., 1996. Anti-inflammatory and analgesic activity of Baccharis trimera: identification of itsactive constituents. Planta Medica 62, 232–235.
Gené, R.M., Marim, E., Adzet, T., 1992. Anti-inflammatory effect of aqueous extractsof three species of the genus Baccharis. Planta Medica 58, 565–566.
ceutica Bonaerense 18, 99–102.Grance, S.R.M., Teixeira, M.A., Leite, R.S., Guimarães, E.B., Siqueira, J.M., Filiu, W.F.O.,
Vasconcelos, S.B.S., Vieira, M.C., 2008. Baccharis trimera: effect on hematolog-ical and biochemical parameters and hepatorenal evaluation in pregnant rats.Journal of Ethnopharmacology 117, 28–33.

5 thnop
G
H
H
H
J
K
K
K
L
L
LL
L
M
M
M
M
N
22 N.P.A. Nogueira et al. / Journal of E
yamfi, A.M., Ohtani, I.I., Shinno, E., Aniya, Y., 2004. Inhibition of glutathioneS-transferases by thonningianin A, isolated from the African medicinalherb, Thonningia sanguinea, in vitro. Food and Chemical Toxicology 42,1401–1408.
abig, W.H., Pabst, M.J., Jakoby, W.B., 1974. Glutathione S-transferases: the firstenzymatic step in mercapturic acid formation. Journal of Biological Chemistry249, 7130–7139.
ayes, J.D., Pulford, D.J., 1995. The glutathione S-transferase supergene family: regu-lation of GST and the contribution of the isoenzymes to cancer chemopreventionand drug resistance. Critical Reviews in Biochemistry and Molecular Biology 30,445–600.
enriques, M.G., Silva, P.M., Martins, M.A., Flores, C.A., Cunha, F.Q., Assreuy-Filho, J., Cordeiro, R.S., 1987. Mouse paw edema. A new model forinflammation? Brazilian Journal of Medical and Biological Research 20,243–249.
arvis, B.B., Wang, S., Cox, C., Rao, M.M., Philip, V., Varaschin, M.S., Barros, C.S., 1996.Brazilian Baccharis toxins: livestock poisoning and the isolation of macrocyclictrichothecene glucosides. Natural Toxins 4, 58–71.
awabata, K., Yamamoto, T., Hara, A., Shimizu, M., Yamada, Y., Matsunaga, K., Tanaka,T., Mori, H., 2000. Modifying effects of ferulic acid on azoxymethane-inducedcolon carcinogenesis in F344 rats. Cancer Letters 157, 15–21.
rajka-Kuzniak, V., Baer-Dubowska, W., 2003. The effect of tannic acid oncytochrome P450 and phase II enzymes in mouse liver and kidney. ToxicologyLetters 143, 209–216.
rajka-Kuzniak, V., Kaczmarek, J., Baer-Dubowska, W., 2008. Effect of natu-rally occurring phenolic acids on the expression of glutathione S-transferaseisozymes in the rat. Food and Chemical Toxicology 46, 1097–1102.
ago, J.H.G., Romoff, P., Fávero, O.A., Soares, M.G., Baraldi, P.T., Corrêa, A.G., Souza,F.O., 2008. Composic ão química dos óleos essenciais das folhas de seis espéciesdo gênero Baccharis. Química Nova 31, 727–730.
evin, D.E., Yamasaki, E., Ames, B.N., 1982. A new Salmonella tester strain, TA97,for the detection of frameshift mutagens: A run of cytosines as a mutationalhot-spot. Mutation Research/Fundamental and Molecular Mechanisms of Muta-genesis 94, 315–330.
evy, L., 1969. Carrageenan paw edema in mouse. Life Science 8, 601–606.iu, X.-P., Goldring, C.E.P., Wang, H.-Y., Copple, I.M., Kitteringham, N.R., Park, B.K.,
2009. Extract of Ginkgo biloba induces glutathione S-transferase subunit-Piin vitro. Phytomedicine 16, 451–455.
opes, L.C., Albano, F., Laranja, G.A.T., Alves, L.M., Silva, L.F.M., Souza, G.P., Araujo, I.M.,Nogueira-Neto, J.F., Felzenszwalb, I., Kovary, K., 2000. Toxicological evaluationby in vitro and in vivo assays of an aqueous extract prepared from Echinodorusmacrophyllus leaves. Toxicology Letters 116, 189–198.
anley, G.D., Edds, G.T., Sundlof, S.F., 1982. Cattle deaths from poisonous plant.Florida Veterinary Journal 11, 20.
aron, D.M., Ames, B.N., 1983. Revised methods for the Salmonella mutagenicitytest. Mutation Research 113, 173–215.
cLellan, L.I., Hayes, J.D., 1987. Sex-specific constitutive expression of the pre-neoplastic marker glutathione S-transferase, YfYf, in mouse liver. BiochemicalJournal 245, 399–406.
ossman, T., 1983. Rapid colorimetric assay for cellular growth and survival:
application to proliferation and cytotoxicity assays. Journal of ImmunologicalMethods 65, 55–63.akasugi, T., Komai, K., 1998. Antimutagens in the Brazilian folk medicinal plantcarqueja (Baccharis trimera Less.). Journal of Agricultural and Food Chemistry46, 2560–2564.
harmacology 138 (2011) 513– 522
Nishikori, T., Irie, K., Suganuma, T., Ozaki, M., Yoshioka, T., 2002. Anti-inflammatorypotency of FR167653, a p38 mitogen-activated protein kinase inhibitor, inmouse models of acute inflammation. European Journal of Pharmacology 451,327–333.
Nordberg, J., Arn’er, E.S.J., 2001. Reactive oxygen species, antioxidants and the mam-malian thioredoxin system. Free Radical Biology and Medicine 31, 1287–1312.
Parejo, I., Viladomat, F., Bastida, J., Rosas-Romero, A., Saavedra, G., Murcia, M.A.,Jimıenez, A.M., Codina, C., 2003. Investigation of Bolivian plant extracts for theirradical scavenging activity and antioxidant activity. Life Science 73, 1667–1681.
Perez-García, F., Marín, E., Adzet, T., Canigueral, S., 2001. Activity of plant extracts onthe respiratory burst and the stress protein synthesis. Phytomedicine 8, 31–38.
Peterson, G.L., 1977. A simplification of the protein assay method of Lowry et al.which is more generally applicable. Analytical Biochemistry 83, 346–356.
Pio Correa, M., 1984. Dicionário das plantas úteis do Brasil e das exóticas culti-vadas, vol. III, Ministério da Agricultura, Instituto Brasileiro de DesenvolvimentoFlorestal, Rio de Janeiro, p. 129.
Raza, H., Awasthi, Y.C., Zaim, M.T., Eckert, R.L., Mukhtar, H., 1991. Glutathione S-transferases in human and rodent skin: multiple forms and species-specificexpression. Journal of Investigative Dermatology 96, 463–467.
Rizzo, I., Varsavky, E., Haidukowski, M., Frade, H., 1997. Macrocyclic trichothecenesin Baccharis coridifolia plants and endophytes and Baccharis artemisioides plants.Toxicon 35, 753–757.
Rodrigues, C.R.F., Dias, J.H., Mello, R.N., Richter, M.F., Picada, J.N., Ferraz, A.B.F., 2009.Genotoxic and antigenotoxic properties of Baccharis trimera in mice. Journal ofEthnopharmacology 125, 97–101.
Sanchez-Palomino, S., Abad, M.J., Bedoya, L.M., Garcia, J., Gonzales, E., Chiriboga,X., Bermejo, P., Alcami, J., 2002. Screening of South American plants againsthuman immunodeficiency virus: preliminary fractionation of aqueous extractfrom Baccharis trinervis. Biological & Pharmaceutical Bulletin 25, 1147–1150.
Silva, F.G., Oliveira, C.B.A., Pinto, J.E.B.P., Nascimento, V.E., Santos, S.C., Seraphin, J.C.,Ferri, P.H., 2007. Seasonal Variability in the Essential Oils of Wild and CultivatedBaccharis trimera. Journal of the Brazilian Chemical Society 18, 990–997.
Simões-Pires, C.A., Debenedetti, S., Spegazzini, E., Mentz, L.A., Matzenbacher, N.I.,Limberger, R.P., Henriques, A.T., 2005. Investigation of the essential oil from eightspecies of Baccharis belonging to section Caulopterae (Asteraceae, Astereae): ataxonomic approach. Plant Systematics and Evolution 253, 23–32.
Sousa, M.P., Matos, N.E.O., Matos, M.F.J.A., Machado, M.I.L., Craveiro, A.A., 1991. Con-stituintes químicos ativos de plantas medicinais brasileiras, UFC Eds, Fortaleza.
Sugishita, E., Amagaya, S., Ogihara, Y., 1981. Anti-inflammatory testing methods:comparative evaluation of mice and rats. Journal of Pharmacobiodynamics 4,565–575.
Tapia, A., Rodriguez, J., Theoduloz, C., Lopez, S., Feresin, G.E., Schmeda-Hirschmann,G., 2004. Free radical scavengers and antioxidants from Baccharis grisebachii.Journal of Ethnopharmacology 95, 155–161.
Vargas, V.M.F., Guidobono, R.R., Jordão, C., Henriques, J.A.P., 1995. Use of two shortterm tests to evaluate the genotoxicity of river water treated with differentconcentration/extraction procedures. Mutation Research 343, 31–52.
Vargas, V.M.F., Motta, V.E.P., Henriques, J.A.P., 1993. Mutagenic activity detected bythe Ames test in river water under the influence of petrochemical industries.Mutation Research 319, 31–45.
Verdi, L.G., Brighent, I.M.C., Pizzolatti, M.G., 2005. Gênero Baccharis (Asteraceae):aspectos químicos, econômicos e biológicos. Química Nova 28, 85–94.
Weimann, C., Goransson, U., Pongprayoon-Claeson, P., Bohlin, L., Rimpler, H.,Heinrich, M., 2002. Spasmolytic effects of Baccharis conferta and some of itsconstituents. Journal of Pharmacy and Pharmacology 54, 99–104.




















