
Improved efficacy of DNA vaccination against breast cancer by boosting with the repeat beta-hCG...
10
Vaccine 24 (2006) 2575–2584 Improved efficacy of DNA vaccination against breast cancer by boosting with the repeat beta-hCG C-terminal peptide carried by mycobacterial heat-shock protein HSP65 He Yi, Yan Rong, Zhang Yankai, Liu Wentao, Zhou Hongxia, Wu Jie, Cao Rongyue, Li Taiming, Liu Jingjing ∗ Minigene Pharmacy Laboratory, Biopharmaceutical College, China Pharmaceutical University, Tongjia Xiang 24, Nanjing 210009, PR China Received 23 October 2005; received in revised form 27 November 2005; accepted 7 December 2005 Available online 18 January 2006 Abstract Studies have demonstrated that active-specific immunotherapy has potential for controlling mammary tumor progression. Human chorionic gonadotropin (hCG) is expressed and extremely sensitive, easily detectable and highly correlated with breast cancer. We developed a gene vaccine using a plasmid vector to deliver the six copies of 10-amino acid residues of -hCG 109–118 and hCG C-terminal 37-amino acid (CTP37). BALB/c female mice were immunized with a combination of pCR-HBc-X6-hCGCTP37 DNA vaccine and HSP-X6-hCGCTP37 protein vaccine. pCR-HBc-X6-hCGCTP37 DNA vaccine were injected intramuscularly three times, on days −46,−25 and −11 and HSP- X6-hCGCTP37 protein were applied two times, 21 and 14 days before tumor cell challenge. We assessed a combined DNA and protein vaccine for its effect of against murine EMT6 mammary tumor cells. In this study, animals vaccinated DNA vaccination boosting with the repeat -hCG C-terminal peptide carried by mycobacterial heat-shock protein HSP65 induced higher avidity antibodies and effectively inhibited the growth of tumor, compared with treatment using DNA alone or BCG priming HSP-X6-hCGCTP37 protein boosting. The data presented demonstrate that improve immunogenicity of DNA vaccination by boosting with the repeat -hCG C-terminal peptide carried by mycobacterial heat-shock protein HSP65, which should prove useful in the development of new DNA vaccine against growth factors for cancer immunotherapy. © 2005 Elsevier Ltd. All rights reserved. Keywords: Human chorionic gonadotropin; DNA vaccine; Heat shock protein; Self-peptide; Prime-boost strategy; Protein vaccine; Breast cancer; Immunother- apy 1. Introduction Like the other members of the glycoprotein hormone fam- ily, human Chorionic Gonadotropin (hCG) is a heterodimeric molecule consisting of an -chain common to all members of the glycoprotein hormone family non-covalently associated with a hormone specific beta-chain [1]. Human CG is ini- ∗ Corresponding author. Tel.: +86 25 83271242; fax: +86 25 83240240. E-mail address: [email protected] (L. Jingjing). tially expressed in the early blastocyst by syncytiotrophoblast and produces oestrogen and progesterone which are required for the implantation and maintenance of the ovum. How- ever, numerous studies have demonstrated the presence of free -hCG in the serum and urine of patients with both tro- phoblastic and nontrophoblastic tumors and define a class of solid tumors with a particularly poor prognosis [2–4]. The role of hCG and mammary tumor seem to be para- doxically. Discoveries revealed that both pregnancy and hCG treatment induce differentiation of the mammary gland, this 0264-410X/$ – see front matter © 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2005.12.030
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Improved efficacy of DNA vaccination against breast cancer by boosting with the repeat beta-hCG...

Vaccine 24 (2006) 2575–2584
Improved efficacy of DNA vaccination against breastcancer by boosting with the repeat beta-hCGC-terminal peptide carried by mycobacterial
heat-shock protein HSP65
He Yi, Yan Rong, Zhang Yankai, Liu Wentao,Zhou Hongxia, Wu Jie, Cao Rongyue,
Li Taiming, Liu Jingjing∗
Minigene Pharmacy Laboratory, Biopharmaceutical College, China Pharmaceutical University,Tongjia Xiang 24, Nanjing 210009, PR China
Received 23 October 2005; received in revised form 27 November 2005; accepted 7 December 2005Available online 18 January 2006
n chorionicoped a geneid
-and proteinting withffectively
atabyfactors for
munother-
blastquiredow-nce oftro-ss of
ara-hCGthis
Abstract
Studies have demonstrated that active-specific immunotherapy has potential for controlling mammary tumor progression. Humagonadotropin (hCG) is expressed and extremely sensitive, easily detectable and highly correlated with breast cancer. We develvaccine using a plasmid vector to deliver the six copies of 10-amino acid residues of�-hCG 109–118 and� hCG C-terminal 37-amino ac(CTP37). BALB/c female mice were immunized with a combination of pCR-HBc-X6-�hCGCTP37 DNA vaccine and HSP-X6-�hCGCTP37protein vaccine. pCR-HBc-X6-�hCGCTP37 DNA vaccine were injected intramuscularly three times, on days−46,−25 and−11 and HSPX6-�hCGCTP37 protein were applied two times, 21 and 14 days before tumor cell challenge. We assessed a combined DNAvaccine for its effect of against murine EMT6 mammary tumor cells. In this study, animals vaccinated DNA vaccination boosthe repeat�-hCG C-terminal peptide carried by mycobacterial heat-shock protein HSP65 induced higher avidity antibodies and einhibited the growth of tumor, compared with treatment using DNA alone or BCG priming HSP-X6-�hCGCTP37 protein boosting. The dpresented demonstrate that improve immunogenicity of DNA vaccination by boosting with the repeat�-hCG C-terminal peptide carriedmycobacterial heat-shock protein HSP65, which should prove useful in the development of new DNA vaccine against growthcancer immunotherapy.© 2005 Elsevier Ltd. All rights reserved.
Keywords: Human chorionic gonadotropin; DNA vaccine; Heat shock protein; Self-peptide; Prime-boost strategy; Protein vaccine; Breast cancer; Imapy
1. Introduction
Like the other members of the glycoprotein hormone fam-ily, human Chorionic Gonadotropin (hCG) is a heterodimericmolecule consisting of an�-chain common to all members ofthe glycoprotein hormone family non-covalently associatedwith a hormone specific beta-chain[1]. Human CG is ini-
∗ Corresponding author. Tel.: +86 25 83271242; fax: +86 25 83240240.E-mail address: [email protected] (L. Jingjing).
tially expressed in the early blastocyst by syncytiotrophoand produces oestrogen and progesterone which are refor the implantation and maintenance of the ovum. Hever, numerous studies have demonstrated the presefree�-hCG in the serum and urine of patients with bothphoblastic and nontrophoblastic tumors and define a clasolid tumors with a particularly poor prognosis[2–4].
The role of hCG and mammary tumor seem to be pdoxically. Discoveries revealed that both pregnancy andtreatment induce differentiation of the mammary gland,
0264-410X/$ – see front matter © 2005 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2005.12.030

2576 H. Yi et al. / Vaccine 24 (2006) 2575–2584
process is considered to be essential for the inhibition ofthe neoplastic process[5]. However, women diagnosed withbreast cancer during pregnancy did not seem to have betterprognoses and often had worse effects[6]. Furthermore, it hasbeen reported that hCG may play a role in cancer progres-sion, such as cell transformation, angiogenic, metastatic, andimmune escape phenomena[7]. Then studies are in progressto develop a vaccine approach that will best target this expres-sion. Our laboratory have found that hCG protein vaccinein the presence of adjuvant failed to significantly inhibit thegrowth of breast cancer xenografts (data unpublished), due tothe lack of higher avidity antibodies induced. While studieshave reported that DNA immunization induce higher avid-ity antibodies and examination of the IgG subclass responseshowed that IgG2a was induced by i.m. DNA immunization,coinciding with elevated interferon y production, and cyto-toxic T cell (CTL) response could be detected after DNAimmunization[8]. Then we have developed a plasmid expres-sion vector pCR-HBc which contain hepatitis B virus core(HBc) protein as an epitope carrier for its particular abilityto self-assemble into highly organized virus like particles,consequently largely improving the immunogenicity to elicitstrong immune responses in human and rodent[9]. And othersequences in this vector that may function as immunostimula-tory sequences (ISS). ISS are unmethylated CpG motifs withthe context of certain franking bases, which could activatet lulari
ule( oxylt anti-b ody( ence( i-c e of� sixr thep oseds rmD
ina ownt evedw ie elec-te anti-b vantb edr rialh -t Bc-X vesw tiona withp
2. Materials and methods
2.1. Tumor cell line and mice
EMT6 mammary tumor cells were maintained inPRM1640 medium supplemented with 10% fetal calf serum(FCS), penicillin G (100 units/ml), streptomycin sulfate(100 ug/ml) and 2 mMl-glutamine. For all experimentsfemale, 6–8-week-old, BALB/c mice were housed under spe-cific pathogen-free conditions at Animal Center of NanjingUniversity (Nanjing, PR China). The implantation of subcu-taneous tumors was carried out by injection of EMT6 cell2× 104 for the prophylactic approach.
2.2. Plasmids construction and transfection in vitro
Plasmid pCR3.1 (Invittrogen, Paisley, UK) was used as thebackbone plasmid, into which the whole gene of HBc and arestriction enzyme site (AgeI) into HBc fragment at a posi-tion corresponding to aa 80–81 was subcloned, to generatepCR-HBc. ISS containing eight 5′-GACGTT-3′ CpG motifswas synthesized by recombinant PCR and inserted into back-bone of plasmid pCR-HBc. Plasmids encoding six copies of�-hCG 109–118 and the C-terminal of 37 peptide of hCGwere made by PCR with the sense primer with (5′-ATG GGTGGC ATG GAT TTC ACC GGT ACT TGC G-3′) and theaT at tog oi xin-p edb
edi ionR cellsw -c ns-f trols.I utedi ofl andi esw byoa anti-m
2c
ss (5G -G 2( -
he innate immune system and facilitate humoral or celmmune responses as vaccine adjuvant[10,11].
Studies reported that a totally synthetic molec109–145 peptide) analogous to the beta-subunit carberminus was used as an antigen in the development ofodies by hybridoma technique. And a monoclonal antib702 D7) specifically recognized the 10 amino acid sequ109–118) free native�-hCG.[12]. All of these studies indate that there are special epitopes in CTP37 peptid-hCG. So, we have inserted the DNA fragment coding
epeat�-hCG C-terminal and CTP37 peptide into HBc oflasmid pCR-HBc at a position corresponding to the expite within its immunodominent region (aa 80–81) to foNA vaccine pCR-HBc-X6-�hCG CTP37.When DNA and virus were first used successivelyPrime-boost protocol, immune responses were sh
o be of much greater magnitude than could be achiith either vector used separately[13]. Nadia Terrazzint al. have reported that subjecting the muscles to
ric pulses following intramuscular injection of�-hCG-xpressing plasmids increased the levels of hCG specificodies, which could be further enhanced by protein-adjuoosts[14]. In the following study, we have constructepeat�-hCG C-terminal peptide carried by mycobacteeat-shock protein HSP65 (HSP-X6-�hCGCTP37) as pro
ein vaccine in absence of adjuvant to boost the pCR-H6-�hCG CTP37 DNA vaccine. In this study, our objectiere to assess whether the efficacy of DNA vaccinagainst breast cancer could be improved by boostingeptide.
ntisense primer (5′-GAC GAC CGG TCT GCG GCA GGACG GAG TGT C-3′) using pHSP-X6-�hCGCTP37 as
emple, followed by insertion into AgeI site of pCR-HBcenerate pCR-HBc-X6-�hCGCTP37. All plasmids used t
mmunize mice were purified using the Qiagen endotourification Kit (Qiagen, Valencia, CA, USA) and quantifiy measuringA260 using 1 mg/ml equal toA260= 50.
The plasmid pCR-HBc-X6-�hCGCTP37 was transfectnto CHO cells using DOTAP Liposomal Transfecteagent (Roche, Germany). Two days later, transfectedere lysed for the presence of X6-�hCGCTP37 by a speific ELISA analysis. Lysates prepared from cells traected by empty plasmids were used as negative conn brief, cells lysates or culture supernatants were diln buffer (0.1 mmol/l Na2CO3–NaHCO3, pH 9.5), applied tat-bottom ELISA plates (Costar, USA) at 100 ul per wellncubated overnight for 4◦C. Ninty-six-plates were incubatith a anti-�hCG antibody (1:50) previously developedur laboratory, followed by detection of a bound anti-�hCGntibody with a horseradish peroxidase-labeled sheepouse antibody from Wuhan Boster (Wuhan, China).
.3. Construction of HSP65-X(0,2,4,6)-βhCG CTP37himeric protein
The partial encoding sequence of�-hCGCTP37 waynthesized by PCR using the oligonucleotides: h1′-ACGACCCGCGTTTCCAGGACTCTTCTTCTTCTAAACTCCGCCGCCATCTCTG-3′) as the forward primer; h
ACGGACCCGGCAGACGAGACGGAGACGGCAGAGA

H. Yi et al. / Vaccine 24 (2006) 2575–2584 2577
TGGCGGCGGAGC-3′) as the reverse primer. PCR reactionwas carried out in 50 ul volumes containing 100 pmolh1, 100 pmol h2 in the standard reaction buffer (dNTPs0.2 mM; 50 mM KCl; 10 mM Tris–HCl, pH 9.0; 0.1%Triton X-100). Typically, the reaction was taken through30 cycles with 30 s at 94◦C, 2 min at 58◦C and 2 min at72◦C. The resulting products after purification were usedas DNA templates for amplification of the full encodingsequence of�-hCGCPT37 using the upstream primer h3 (5′-GGGGGATCCGACTGGTGGCACTTGCGACGACCCGC-GTTTCCAGGAC-3′) and the downstream primer h4 (5′-AAAAAGCTTACTGCGGCAGGATCGGAGTGTCAGAC-GGACCCGGCAG-3′). Then the resulting PCR fragmentwith a BamHI site located at the 5′ end, and aHindIII sitelocated just after the 3′ end to the termination codon TAAwas digested withBamHI andHindIII inserted into pET-28awhich was cut withBamHI and HindIII. The plasmid waspET-�hCGCTP37.
The HSP65 gene containing the signal peptidesequence was amplified by PCR fromMycobacteriumbovis BCG genomic DNA template using H1 (5′-TTGACCATGGCCAAGACAATTGCGTAC-3′) containingan ATG start codon at aNcolI site; reverse primer H2 (5′-TAAAAGATCTGCGCTAGCGAAATCCATGCCACCCAT-3′) contains aNheI site and aBgelII (which is a isocaudamer
e
h
hemrt
-h5
e
extending the C-terminal part of HSP 65 using the methoddescribed above. On the analogy of this, the plasmid ofHSP-X6-�hCGCTP37 containing six copies of 10-aminoacid residues of�-hCG 109–118 was constructed frompHSP-X4-�hCGCTP37 by extending the C-terminal part ofHSP65.
2.4. Expression and purification ofHSP65-X6-βhCGCTP37 chimeric protein
A preculture, inoculated from a single bacterial colony,was grown for 10 h at 37◦C. The pre-culture was diluted50-fold in fermenter medium (maize plasma 2.5% (v/v), beefextraction 1.5% (v/v), monosodium glutamate 1% (v/v)) con-taining kanomycin (50 ug/ml), and the bacterial were grownat 37◦C with an aeration rate of 1 (v/v/min) and agitationat 600 rpm in a 10 l steel fermentor (Taixing BiochemicalInstrument Plant, Jiangsu Province, China). The expressionof HSP65-X6-�hCGCTP37 was induced atA550 of 1.3–1.5by addition of lactose to a final concentration of 5 mM. Theculture, incubated for 4 h after induction, reached a finalA550 of 2.6–3.5. The bacterial were harvested at this pointand were kept frozen until used. Polyacrylamide gel elec-trophoresis in the presence of SDS (SDS–polyacrylamide gelelectrophoresis (PAGE) described by Laemmli (1970) wasused for determining the expression level of the recombinantp
es sedw14 ctionc dt ra-p tedw nw hene .5 MN ingt thep .
2m
ndp gneda at ntrolgo go 5-X e( tra-mt smid
rotein.BL21 (DE3) cells harboring HSP-X6-�hCGCTP37 wer
uspended in buffer of 50 mM Tris–HCl pH 7.4 and lyith 0.4 ug/ml in the presence of 1 U/mlDNaseI at 37◦C forh. Centrifugation was carried out at 12,000× g for 20 min at◦C. The pellets were discarded and the supernatant fraontaining soluble HSP65-X6-�hCGCTP37 protein filterehrough a 14 KDa cutoff point filter prior to chromatoghy on a DEAD-Sephacel (Pharmacia, USA) equilibraith 50 mM Tris–HCl (equilibration buffer). The columas washed with 300 ml of the equilibration buffer and tluted with a linear gradient buffer between 50 mM and 0aCl in equilibration buffer. The peak fractions contain
he HSP65-X6-�hCGCTP37 were pooled. A sample ofooled solution was taken for 12% SDS–PAGE analysis
.5. Immunization procedure and mammary tumorodel
In order to analyze the antitumor effect of DNA prime arotein boost strategy, a vaccination protocol was desis shown inFig. 3 A. We set four group as follow while
otal often was used for each experimental group: PBS coroup, treated with either pCR-HBc-X6-�hCGCTP37 DNAr BCG priming HSP65-X6-�hCGCTP37 protein boostinr a combination of pCR-HBc-X6-�hCGCTP37 and HSP66-�hCGCTP37. pCR-HBc-X6-�hCGCTP37 DNA vaccin
100 ug in a final volume of 100 ul PBS) was injected inuscularly three times, on days−46, −25 and−11 before
umor cells challenge. To enhance the cell uptake of pla
of BamHI) site. Amplification was performed using thfollowing parameters: 40 cycles with 1 min at 94◦C, 2 min at62, and 4 min at 72◦C. The PCR product was digested witNcolI and BgelII, purified and ligated to pET-�hCGCTP37which had been cut withNcolI and BamHI. The resultingplasmid was pET-HSP65-�hCGCTP37.
The chimeric plasmid pET-HSP-X6-�hCGCTP37encodes six copies of 10-amino acid residues of�-hCG109–118, the C-terminal of 37 peptide of hCG and tfusion protein of HSP65. This plasmid was constructed fropET-HSP65-�hCGCTP37 by extending the C-terminal paHSP65. The DNA fragment ofX2, which encodes two copiesof 10-amino acid residues of�-hCG 109–118, was synthesized by PCR amplification using the oligonucleotides:(5′-ATATCTAGAACTTGCGACGACCCGCGTTTCCAG-GACTCTACTTGCGACGACCCG-3′) as the forward primerand h6 (5′-AAAGCTAGCAGAGTCCTGGAAACGCGG-GTCGTCGCAAGTTGAGTCCTGGAAACG-3′) as thereverse primer. Amplification was performed using thfollowing parameters: 30 cycles with 1 min 94◦C, 50 s at58◦C and 30 s at 72◦C. The resulting PCR fragment with aXbaI (which is a isocaudamer ofNheI) site located at the 5′end, and aNheI site located at the 3′ end, was digested withrestriction endouncleasesXbaI and NheI and subsequentlyinserted into pHSP65-�hCGCTP37 digested withNheI. TheX2 was located at the 3′ end of the encoding region for HSPand the 5′ end of the encoding region for�hCGCTP37. Theresulting plasmid was HSP-X2-�hCGCTP37, which alsocontains aNheI site. The plasmid of HSP-X4-�hCGCTP37containing four copies of 10-amino acid residues of�-hCG109–118 was constructed from pHSP-X2-�hCGCTP37 by

2578 H. Yi et al. / Vaccine 24 (2006) 2575–2584
DNA, the quadriceps were injected with a total of 100 ulof 0.25% bupivacaine three days before each DNA admin-istration. HSP65-X6-�hCGCTP37 were applied intra twotimes, 21 and 14 days before tumor cells challenge. Micetreated by HSP65-X6-�hCGCTP37 group received intraperi-toneal injection of 106 CFU of BCG (Bacillus Calmette-Guerin; attenuatedMycobacterium tuberculosis var. bovis).After 0 day, EMT6 mammary tumor cells (2× 104) wereinjected subcutaneously in treatment groups and in PBS con-trol group. Fourteen days after the tumor cells challenge, allmice were sacrificed for analysis the weights and volumes oftumors. Two bisecting diameters of each tumor were mea-sured with calipers. The volume was calculated using theformulaV = 0.4ab2, with “a” as the larger diameter and “b” asthe smaller diameter. And tumors were fixed in 10% neutral-buffered formalin.
2.6. Specificity of anti-βhCG antibodies
To demonstrate that the antibody from the mice immu-nized was specific for hCG, the carboxyl terminus fragmentof the l-asparaginasev (ansB-C) was used as the fusionpartner for constructing a ansB-C–�hCGCTP37 fusion pro-tein. The purified fusion protein and the native hCG wereelectrophoresed on 15% SDS–polyacrlaminde gel and trans-ferred to nitrocellulose membrane at 4◦C overnight. Equall ingt werew fatd3 ilki da 5%3 xidei
2
n sor-bb nanta epto ma hedt ST)a erao et atedw abbita om-p ure,t eredwt l/LN s
stopped with 50 ul/well of 2 mol/L H2SO4 and absorbanceswere read atA450nm.
2.8. Determination of relative avidity of antibodies byELISA
The method for avidity assays were similar to thatdescribed by Romero-Steiner et al.[16]. Briefly, ELISAplates were coated with 50 ul/well of antigen in PBS, pH 7.4,and kept overnight at 4◦C. Antigen was then removed fromthe wells and 150 ul of 5% (w/v) BSA in PBS added for 1 h.After incubation with the sera, the wells were washed PBST,and covered with 50 ul of serial dilutions of 0–10 M sodiumthiocyanate (NaSCN, Sigma, USA), a chaotropic compoundthat interfered with antigen–antibody reaction in 0.1 M phos-phate buffer, pH 7.0. The wells were incubated for 20 minat room temperature before washing three times with PBST.The remainder of the assay was done similar to the ELISAmethod for detecting anti-hCG specific antibodies.
3. Results
3.1. Expression of HBc-X6-βhCGCTP37 in transfectedCHO cell lines
X6-� ctedC werecp CHOc sa -t tantso
F s;e as-m tsnt c-X6-� ctedb ysa
oading of individual lanes were confirmed by stainhe membranes with 1% Ponceau Red. Membranesashed with water and blocked with 5% (w/v) lowry milk in PBST for 1 h. The expression of�hCGCTP7 was detected with mice antiserum in 2% dry m
n PBST for 1 h at 37◦C followed by HRP-conjugatenti-rat IgG. The reaction was completed with 0.0,3′-diaminobenzidine and 0.012% hydrogen pero
n PBS.
.7. Detection of anti-βhCG antibodies
The formation of antibodies against�hCG in immu-ized mice was tested using enzymelinked immunoent assay (ELISA) as published[15]. Briefly, 96-well flat-ottomed plates were coated with 100 ul/well of recombinsB-C–�hCGCTP37 proteins (10 ug/well) in PBS and kvernight at 4◦C. Wells were blocked with 5% bovine serulbumin (BSA) in PBS at room temperature for 2 h, was
hree times with PBS containing 0.05% Tween-20 (PBnd incubated with 100 ul of serial dilutions of individual sbtained from immunized mice for 1 h at 37◦C. The sera wer
hen removed, the wells washed with PBST, and incubith 100 ul horseradish peroxidase (HRP)-conjugated rnti-IgG secondary diluted at 1:20,000 (Wuhan Boster Cany, PR China) with PBST. After 1 h at room temperat
he wells were washed three times with PBS and covith 100 uL of the peroxidade substrate (0.01% 3,3′,5,5-
etramethylbenzidine in 50 mmol/L citric acid–100 mmoaH2PO4, pH 5.2 and 0.24% (v/v) H2O2). The reaction wa
CHO cells were transfected with plasmids pCR-HBc-hCGCTP37 and empty vectors pCR-HBc. After transfeHO cells lysing, the supernatants of culture and lysateoated into 96-well ELISA plates. HBc-X6-�hCGCTP37roteins were expressed in the lysate supernatants ofells transfected by pCR-HBc-X6-�hCGCTP37 two dayfter transfection. In contrast, HBc-X6-�hCGCTP37 pro
eins were undetectable in cell culture and lysate supernaf CHO cells transfected by empty vectors (Fig. 1).
ig. 1. Expression of HBc-X6-�hCGCTP37 in transfected CHO cell linexpression of HBc-X6-�hCGCTP37 of CHO cells transfected with plid pCR-HBc-X6-�hCGCTP37 by ELISA. ( ) Positive control represenative hCG as coated protein. () Expression of HBc-X6-�hCGCTP37 in
he cell lysate supernatant two days after transfected with pCR-HBhCGCTP37. ( ) The culture supernatants from CHO cells transfey pCR-HBc-X6-�hCGCTP37. ( ) The cell lysate supernatant two dafter transfected with empty vectors pCR-HBc.
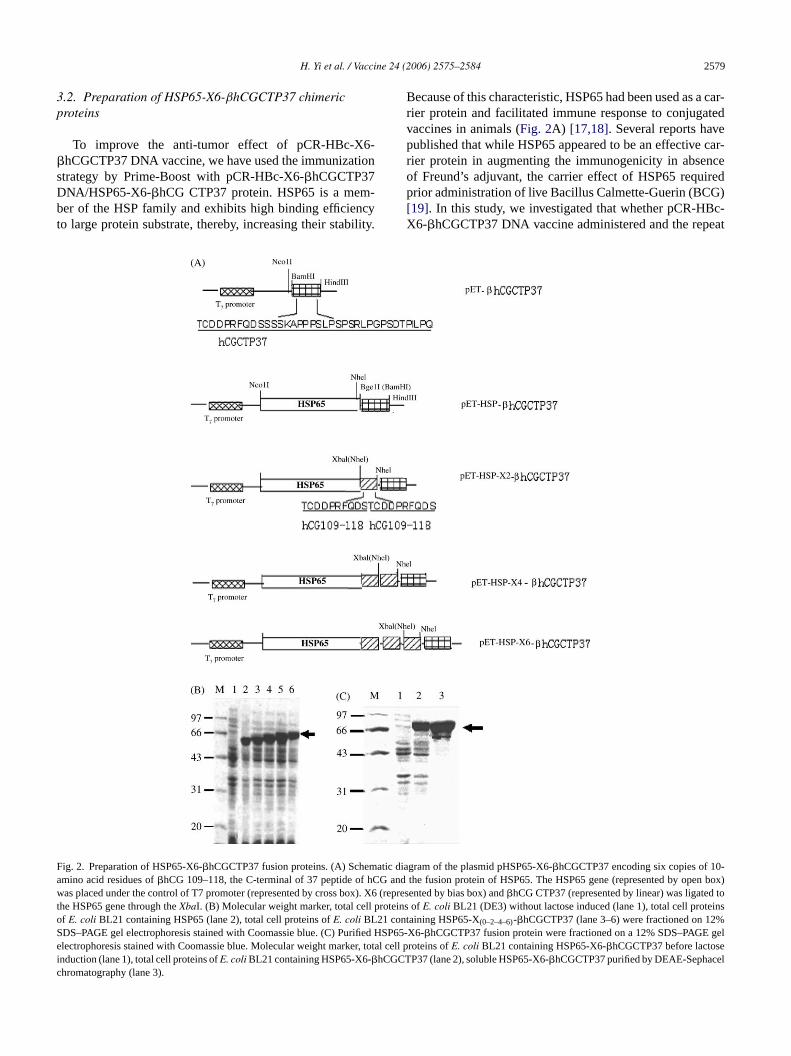
H. Yi et al. / Vaccine 24 (2006) 2575–2584 2579
3.2. Preparation of HSP65-X6-βhCGCTP37 chimericproteins
To improve the anti-tumor effect of pCR-HBc-X6-�hCGCTP37 DNA vaccine, we have used the immunizationstrategy by Prime-Boost with pCR-HBc-X6-�hCGCTP37DNA/HSP65-X6-�hCG CTP37 protein. HSP65 is a mem-ber of the HSP family and exhibits high binding efficiencyto large protein substrate, thereby, increasing their stability.
Because of this characteristic, HSP65 had been used as a car-rier protein and facilitated immune response to conjugatedvaccines in animals (Fig. 2A) [17,18]. Several reports havepublished that while HSP65 appeared to be an effective car-rier protein in augmenting the immunogenicity in absenceof Freund’s adjuvant, the carrier effect of HSP65 requiredprior administration of live Bacillus Calmette-Guerin (BCG)[19]. In this study, we investigated that whether pCR-HBc-X6-�hCGCTP37 DNA vaccine administered and the repeat
FawtoSeic
ig. 2. Preparation of HSP65-X6-�hCGCTP37 fusion proteins. (A) Schematicmino acid residues of�hCG 109–118, the C-terminal of 37 peptide of hCGas placed under the control of T7 promoter (represented by cross box). X6
he HSP65 gene through theXbaI. (B) Molecular weight marker, total cell protef E. coli BL21 containing HSP65 (lane 2), total cell proteins ofE. coli BL21 conDS–PAGE gel electrophoresis stained with Coomassie blue. (C) Purified Hlectrophoresis stained with Coomassie blue. Molecular weight marker, tota
nduction (lane 1), total cell proteins ofE. coli BL21 containing HSP65-X6-�hCGChromatography (lane 3).
diagram of the plasmid pHSP65-X6-�hCGCTP37 encoding six copies of 10-and the fusion protein of HSP65. The HSP65 gene (represented by open box)(represented by bias box) and�hCG CTP37 (represented by linear) was ligated toins ofE. coli BL21 (DE3) without lactose induced (lane 1), total cell proteinstaining HSP65-X(0–2–4–6)-�hCGCTP37 (lane 3–6) were fractioned on 12%SP65-X6-�hCGCTP37 fusion protein were fractioned on a 12% SDS–PAGE gell cell proteins ofE. coli BL21 containing HSP65-X6-�hCGCTP37 before lactoseTP37 (lane 2), soluble HSP65-X6-�hCGCTP37 purified by DEAE-Sephacel
2580 H. Yi et al. / Vaccine 24 (2006) 2575–2584
�-hCG C-terminal peptide carried by mycobacterial HSP65injected following can generate synergistic effect of againstbreast tumor. The 65 kDa of HSP65 and the 7.2 kDa HSP65-X6-�hCGCTP37 fusion proteins were expressed in recombi-nantE. coli BL21 (DE3) (Fig. 2B). Cells harboring HSP-X6-�hCGCTP37 were suspended in buffer of 50 mM Tris–HClpH 7.4 and lysed with lyszome in the presence ofDNaseI.After centrifugal, the pellets were discarded and the super-natant fraction containing soluble HSP65-X6-�hCGCTP37protein filtered through a 14 KDa cutoff point filter prior tochromatography on a DEAD-Sephacel (Pharmacia, USA). Asample near to homogeneity of the pooled solution was takenfor 12% SDS–PAGE analysis (Fig. 2C).
3.3. Induction of anti-hCG antibodies in immunizedmice
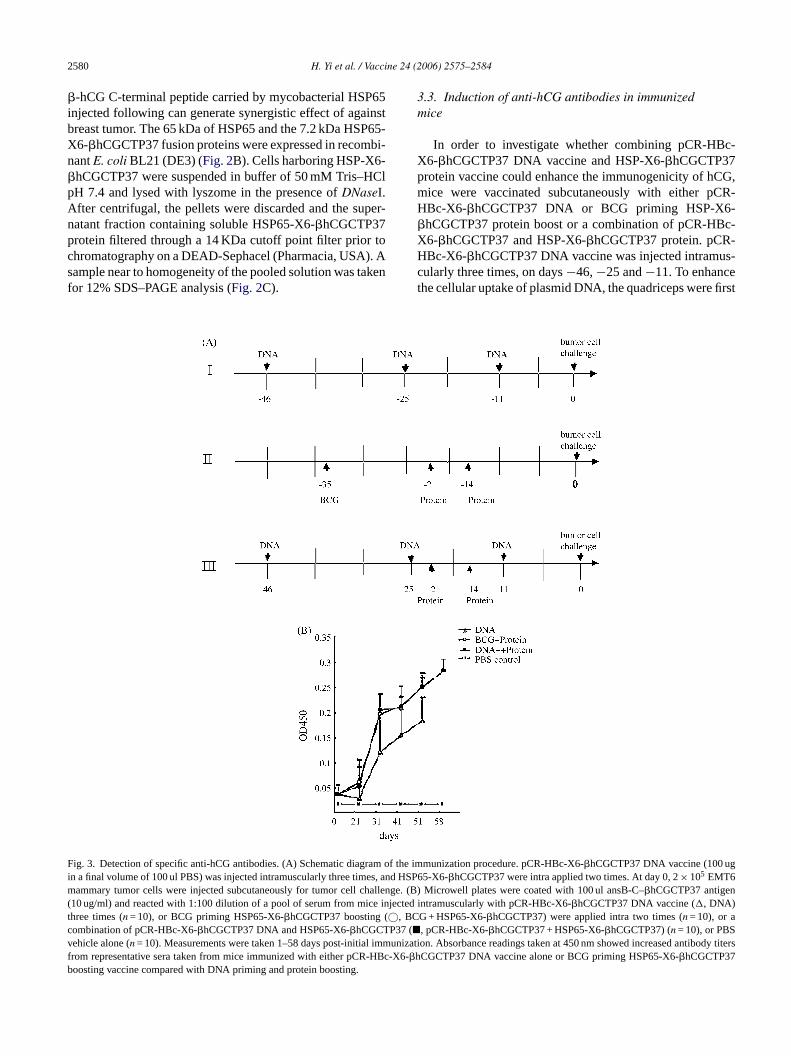
In order to investigate whether combining pCR-HBc-X6-�hCGCTP37 DNA vaccine and HSP-X6-�hCGCTP37protein vaccine could enhance the immunogenicity of hCG,mice were vaccinated subcutaneously with either pCR-HBc-X6-�hCGCTP37 DNA or BCG priming HSP-X6-�hCGCTP37 protein boost or a combination of pCR-HBc-X6-�hCGCTP37 and HSP-X6-�hCGCTP37 protein. pCR-HBc-X6-�hCGCTP37 DNA vaccine was injected intramus-cularly three times, on days−46,−25 and−11. To enhancethe cellular uptake of plasmid DNA, the quadriceps were first
Fim(tcvfb
ig. 3. Detection of specific anti-hCG antibodies. (A) Schematic diagram ofn a final volume of 100 ul PBS) was injected intramuscularly three times, and
ammary tumor cells were injected subcutaneously for tumor cell challeng10 ug/ml) and reacted with 1:100 dilution of a pool of serum from mice injehree times (n = 10), or BCG priming HSP65-X6-�hCGCTP37 boosting (©, BCombination of pCR-HBc-X6-�hCGCTP37 DNA and HSP65-X6-�hCGCTP37 (�ehicle alone (n = 10). Measurements were taken 1–58 days post-initial immurom representative sera taken from mice immunized with either pCR-HBc-�hoosting vaccine compared with DNA priming and protein boosting.
the immunization procedure. pCR-HBc-X6-�hCGCTP37 DNA vaccine (100 ugHSP65-X6-�hCGCTP37 were intra applied two times. At day 0, 2× 105 EMT6e. (B) Microwell plates were coated with 100 ul ansB-C–�hCGCTP37 antigen
cted intramuscularly with pCR-HBc-X6-�hCGCTP37 DNA vaccine (�, DNA)G + HSP65-X6-�hCGCTP37) were applied intra two times (n = 10), or a, pCR-HBc-X6-�hCGCTP37 + HSP65-X6-�hCGCTP37) (n = 10), or PBSnization. Absorbance readings taken at 450 nm showed increased antibody titersX6-CGCTP37 DNA vaccine alone or BCG priming HSP65-X6-�hCGCTP37

H. Yi et al. / Vaccine 24 (2006) 2575–2584 2581
injected with a total of 100 ul of 0.25% bupivacaine three daysbefore each DNA administration. HSP-X6-�hCGCTP37 pro-teins were applied two times, 21 and 14 days before tumorcell challenge (Fig. 3A) [20]. Antibody formation againsthCG was verified using ELISA. Animals injected withpCR-HBc-X6-�hCGCTP37 DNA or BCG-priming HSP-X6-�hCGCTP37 protein boosting or a combination of DNAand protein showed increasing titers of anti-hCG antibodiesby ELISA as early as 3 weeks following initial inoculationand remaining high for 7 weeks. The anti-hCG antibodiestiters in mice immunized with pCR-HBc-X6-�hCGCTP37DNA priming HSP-X6-�hCGCTP37 protein boosting andBCG-priming HSP-X6-�hCGCTP37 protein boosting werenot significantly different; while the anti-hCG antibodiestiters of this two groups mice appeared to be higher than inpCR-HBc-X6-�hCGCTP37 DNA alone immunized mice. Incontrast, injections with PBS vehicle failed to elicit antibodyformation (Fig. 3B). These findings suggested that combin-ing DNA vaccine and the repeat�-hCG C-terminal peptidecarried by mycobacterial heat-shock protein HSP65 couldenhance the immunogenicity.
3.4. Anti-hCG antibodies analysis by Western blot
To demonstrate that the antibodies from the miceimmunized with a combination of pCR-HBc-X6-� nw t oft artnerf in[ np gela micei ein( e� tingt .
3
nti-b nes,a mals( 6-� 6-� andp adt rydr izedw ryl M),r izedwa erN mu-
Fig. 4. SDS-PAGE and Western blot analyses of anti-hCG antibodies, Todetermine the specificity of the anti-hCG antibodies, ansB-C–�hCGCTP37and ansB-C vehicle proteins expressed in bacteria were purified by ureawashing. These proteins and native hCG were electrophoresed on a 15%SDS–polyacrylamide gel and Western blot. ansB-C–�hCGCTP37 andansB-C protein, native hCG reacted with serum (diluted 1:50) from miceimmunized with pCR-HBc-X6-�hCGCTP37 DNA priming and HSP65-X6-�hCGCTP37 boosting. Reaction was completed using DAB and H2O2. Lane1, Arrow A refers to ansB-C–�hCGCTP37; lane 2, arrow B refer to nativehCG(reduced); lane 3, ansB-C vehicle without hCG epitopes.
nized with DNA vaccine alone (p < 0.1) and BCG prim-ing HSP-X6-�hCGCTP37 protein boosting (p < 0.001). Theresults of these suggest the presence of high-avidity antibod-ies in the sera of mice immunized with a combination of DNApriming and protein boosting.
3.6. Effect on mammary tumor model
To test whether a combination of pCR-HBc-X6-�hCGCTP37 DNA and HSP-X6-�hCGCTP37 protein cangenerate synergistic effect of against tumor, we applied the
Fig. 5. The relative avidity of anti-hCG antibodies. Sera from mice immu-nized with pCR-HBc-X6-�hCGCTP37 DNA (n = 10) alone or BCG-primingH dp eree ultsw quiredt eena Stu-d ina-t nw miceia
hCGCTP37DNA and HSP-X6-�hCGCTP37 proteiere specific for hCG, the carboxyl terminus fragmen
he l-asparaginase (ansB-C) was used as the fusion por constructing a ansB-C–�hCGCTP37 fusion prote21]. This fusion protein ansB-C–�hCGCTP37, the fusioarter and native hCG were fractioned in SDS–PAGEnd processed for western blot using antiserum from
mmunized with a combination of DNA vaccine and protFig. 4). Antibodies denoted ansB-C–�hCGCTP37, nativhCG in the Western blot, but not fusion parter, sugges
hat the antibodies specifically recognized the�hCG antigen
.5. Relative avidities of antibodies
To further investigate the relationship between the aody titers and the biological effects of the hCG vaccividity assays were performed on the anti-sera of anin = 10 per group) immunized with either pCR-HBc-XhCGCTP37 DNA vaccine alone or BCG priming HSP-XhCGCTP37 protein boosting or a combination of DNArotein (Fig. 5). The concentration of NaSCN required to le
o the value of OD450 dropping down to half of the primaefines relative avidities of antibodies[22]. The inhibitioneaction between antigen and anti-sera from mice immunith DNA or BCG priming protein boosting required ve
ow concentrations of NaSCN (2–6 M) and NaSCN (2–4espectively. In contrast, antibodies from mice immunith a combination of pCR-HBc-X6-�hCGCTP37 DNAnd HSP-X6-�hCGCTP37 protein were inhibited by highaSCN concentration (3–7 M) than those from mice im
SP65-X6-�hCGCTP37 boosting (n = 10) or a combination of DNA anrotein (n = 10) and obtained 40 days after the initial immunization wxamined for their binding avidity to hCG in a modified ELISA. The resere expressed as the concentration of sodim thiocyanate (NaSCN) re
o disrupt antibody binding to hCG. Any significance in differences betwvidities obtained from the two groups was determined using pairedent’st-test. In contrast, antibodies from mice immunized with a comb
ion of pCR-HBc-X6-�hCGCTP37 DNA and HSP-X6-�hCGCTP37 proteiere inhibited by higher NaSCN concentration (3–7 M) than those from
mmunized with pCR-HBc-X6-�hCGCTP37 DNA vaccine alone (p < 0.1)nd BCG priming HSP-X6-�hCGCTP37 protein boosting (p < 0.001).
2582 H. Yi et al. / Vaccine 24 (2006) 2575–2584
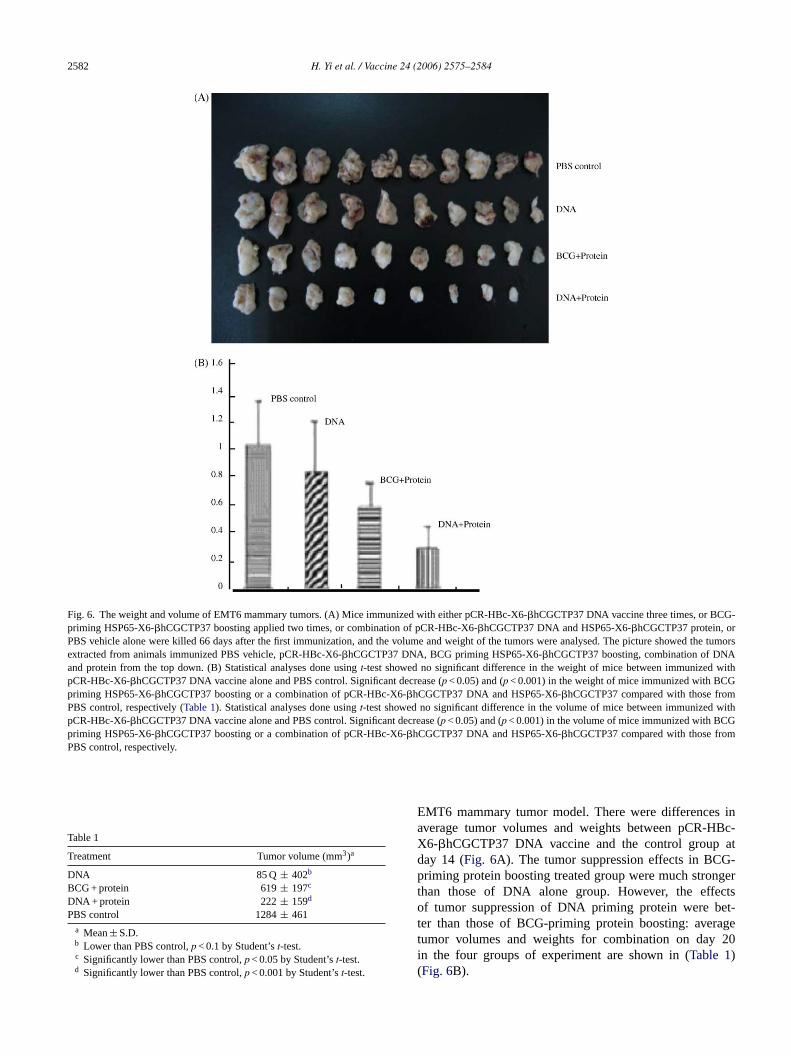
Fig. 6. The weight and volume of EMT6 mammary tumors. (A) Mice immunized with either pCR-HBc-X6-�hCGCTP37 DNA vaccine three times, or BCG-priming HSP65-X6-�hCGCTP37 boosting applied two times, or combination of pCR-HBc-X6-�hCGCTP37 DNA and HSP65-X6-�hCGCTP37 protein, orPBS vehicle alone were killed 66 days after the first immunization, and the volume and weight of the tumors were analysed. The picture showed the tumorsextracted from animals immunized PBS vehicle, pCR-HBc-X6-�hCGCTP37 DNA, BCG priming HSP65-X6-�hCGCTP37 boosting, combination of DNAand protein from the top down. (B) Statistical analyses done usingt-test showed no significant difference in the weight of mice between immunized withpCR-HBc-X6-�hCGCTP37 DNA vaccine alone and PBS control. Significant decrease (p < 0.05) and (p < 0.001) in the weight of mice immunized with BCGpriming HSP65-X6-�hCGCTP37 boosting or a combination of pCR-HBc-X6-�hCGCTP37 DNA and HSP65-X6-�hCGCTP37 compared with those fromPBS control, respectively (Table 1). Statistical analyses done usingt-test showed no significant difference in the volume of mice between immunized withpCR-HBc-X6-�hCGCTP37 DNA vaccine alone and PBS control. Significant decrease (p < 0.05) and (p < 0.001) in the volume of mice immunized with BCGpriming HSP65-X6-�hCGCTP37 boosting or a combination of pCR-HBc-X6-�hCGCTP37 DNA and HSP65-X6-�hCGCTP37 compared with those fromPBS control, respectively.
Table 1
Treatment Tumor volume (mm3)a
DNA 85 Q ± 402b
BCG + protein 619± 197c
DNA + protein 222± 159d
PBS control 1284± 461a Mean± S.D.b Lower than PBS control,p < 0.1 by Student’st-test.c Significantly lower than PBS control,p < 0.05 by Student’st-test.d Significantly lower than PBS control,p < 0.001 by Student’st-test.
EMT6 mammary tumor model. There were differences inaverage tumor volumes and weights between pCR-HBc-X6-�hCGCTP37 DNA vaccine and the control group atday 14 (Fig. 6A). The tumor suppression effects in BCG-priming protein boosting treated group were much strongerthan those of DNA alone group. However, the effectsof tumor suppression of DNA priming protein were bet-ter than those of BCG-priming protein boosting: averagetumor volumes and weights for combination on day 20in the four groups of experiment are shown in (Table 1)(Fig. 6B).

H. Yi et al. / Vaccine 24 (2006) 2575–2584 2583
4. Discussion
Immunization with plasmid-based DNA vaccines is a use-ful way due to their stability and ease of production and purifi-cation. However, for their low relatively immnuogenicity,intramuscular injection of expression plasmids often resultsin a heterogeneous antibody response with some animals pro-ducing appreciable levels of antibodies whereas others barelydevelop detectable levels[23]. A number of potential strate-gies have been devised in order to circumvent this problemof poor immnogenicity, including of adding cytokines andother adjuvants or taking special immune scheme such asprime-boost. Some studies report that a prime-boost strategyis a promising one[24–26]. hCG is extremely sensitive, eas-ily detectable and highly correlated with breast cancer[7].It has been established that vaccines which can attack hCG-producing and cancer/promoting bacteria are able to deprivecancer cells of a key source of hCG; as the levels of hCG arelowered, the immune-system’s ability to launch an assaulton cancer cells increases. It is dependent on the ability toneutralize hCG[27].
In this study, we have evaluated the effects of the vaccineon EMT6 murine breast cancer. Mice immunized with pCR-HBc-X6-�hCGCTP37 DNA priming were clearly effectivein enhancing immune response against�hCG followingHSP-X6-�hCGCTP37 protein boost. Higher relative avid-i fort an-t ime,t thes to-t nala np end teredi y Bc rtedu the-r beent -d ikelyt HBVc nse-q ngi ,t vac-c lln thatD orya tionoI engeo terl c-X th hock
protein HSP65 have higher relative avidities of antibod-ies than pCR-HBc-X6-�hCG CTP37 DNA vaccine or BCGpriming HSP-X6-�hCG CTP37 protein boost. In contrast,EMT6 tumor cell growth was completely inhibited (n = 1)and substantially reduced (n = 9) when animals were immu-nized pHBc-X6-�hCGCTP37 DNA vaccine boosting withthe repeat�-hCG C-terminal peptide carried by mycobacte-rial heat-shock protein HSP65. The weights of these tumorswere substantially reduced in comparison to those from ani-mals injected with pHBc-X6-�hCG CTP37 DNA vaccineor BCG priming HSP-X6-�hCG CTP37 protein boosting orthe control. The efficiency of cancer treatment was due tothe high titer�hCG-specific antibodies which exhibited anexceptional and unique characteristic dose dependent cyto-toxic activity.
In this study, we synthesized recombinant proteinsthat contain six copies of 109–118 of�-hCGCTP37,�-hCGCTP37 peptide and mycobacterial heat-shock proteinHSP65. The repeat�-hCG C-terminal peptide was designedto further increase the immunogenecity of this protein. Self-peptides exhibit low immunogenecity, which have beenpartially obviated by administering immunogens contain-ing multiple copies of the self-peptides in linear alignment.Administering immunogens containing multiple copies ofthe self-peptides in linear alignment have been used, due toits well-known ability to enhance self-peptide low immuno-g ingr -celle nRHcI tidew als.T ac-c peat� eat-s oldp
A
ch-n 2);C ant#3 ienceF 1).
R
f theRes
of aitro.
ty of anti-�hCG antibodies may be the direct reasonhe inhibition of breast cancer by cytotoxic effect. Kalarov and Acevedo have demonstrated, for the first that only the MoAbs CTP103, which directed againstynthetic CTP of the�hCG, exhibited dose dependent cyoxic activity and this type of activity was an exceptiond unique characteristic[28]. However, genetic vaccinatioCR-HBc-X6-�hCGCTP37 encoding a cytoplasmic antigo not produce a very strong antibody response adminis
ntramuscularly. This is not due to a lack of necessarell epitopes, since a World Health Organization-reposing a hCG-specific 37 CTP peptide coupled to diphia toxoid has induced effective immune response andhrough a phase II trial[29]. The failure of the mice to prouce strong antibody response to this nucleic acid is unl
o the lack of T cell epitopes, because the core gene ofan assemble into highly organized virus like particles, couently largely improving the immunogenicity to elicit stro
mmune responses in human and rodent[9]. In conclusionhe weak heterogeneous antibody response to the DNAination of pCR-HBc-X6-�hCGCTP37 is not due to T ceor B cell epitopes. It has been previously demonstratedNA vaccination is as efficient in generating B cell mems conventional immunization, but the memory generaccurs in the absence of an overt effector responses[30].
t is accordance with our result that a subsequent challf the DNA-immunized mice with protein induced a grea
evel of anti-�hCG antibodies. Animals immunized pHB6-�hCG CTP37 DNA vaccine boosting with the repea�-CG C-terminal peptide carried by mycobacterial heat-s
enecity [31]. Our laboratory have reported that usepeated GnRH units conjugated to hinge region and Tpitope induce strong antibody response than single Gonjugated to the same hinge region and T-cell epitope[32].ncreasing the content of the B epitope in antigenic pepas efficient in inducing an antibody response in animaken together, we first report that the efficacy of DNA vination against breast cancer by boosting with the re-hCG C-terminal peptide carried by mycobacterial hhock protein HSP65, providing a useful method to hromise for cancer treat.
cknowledgements
This work was supported by the National High Teology “863” Programs of China (No. 2002 AA217031-hina National Natural Science Fund Committee (Gr0500458; Grant# 30270298); and Jiangsu Natural Scund Committee (Grant# BK 95092309 and BG200101
eferences
[1] Fiddes JC, Tamadge K. Structure, expression and envolution ogenes for the human glycoprotein hormones. Recent Prog Horm1984;40:43–74.
[2] Gawronska B, Leuschner C, Enright FM, Hansel W. Effectslytic peptide conjugated to beta HCG on ovarian: studies in vGynecol Oncol 2002;86(1):45–52.

2584 H. Yi et al. / Vaccine 24 (2006) 2575–2584
[3] Lundin M, Nordling S, Carpelan-Holmstrom M, Louhimo J, AlfthanH, Stenman UH, et al. A comparison of serum and tissuehCG as prognostic markers in colorectal cancer. Anticancer Res2000;20(60):4949–51.
[4] Marcillac I, Cottu P, Theodore C, Terieer-Lacombe MJ, Bellet D,Droz JP. Free hCG-beta subunit as tumor marker in urothelian cancer.Lancet 1993;341:1354–5.
[5] Russo J, Russo IH. The etiopathogenesis of breast cancer prevention.Cancer Lett 1995;90(1):81–9.
[6] Rao CV. Does full-term pregnancy at a young age protect womenagainst breast cancer through hCG? Obstet Gynecol 2000;96:783–6.
[7] Triozzi PL, Stevens VC. Human chorionic gonadotropin as a targetfor cancer vaccines. Oncol Rep 1999;6(1):7–17.
[8] Boyle JS, Silva A, Brady JL, Lew AM. DNA immunization: induc-tion of higher avidity antibody and effect of roution T cell cytotox-icity. Proc Natl Acad Sci USA 1997;94(26):14626–31.
[9] Pumpens P, Grens E. HBV core particles as a carrier for B cell/Tcell epitopes. Intervirology 2001;44(2–3):98–114.
[10] Zhang A, Jin H, Zhang F, Ma Z, Ren Z, Tu Y, et al. Effects of mul-tiple copies of CpG on DNA vaccinati. Cell Biol 2005;24(5):292–8.
[11] Vollmer J, Jurk M, Samulouitz U, Lip ford G, Forsbarch A, Wull-ner M, et al. CpG oligodeoxynucleotides stimulate IFN-gamma-inducible protein-10 production in human B cells. J Endotoxin Res2004;10(6):431–8.
[12] Bellet D, Biard JM, Jolivet M, Tartar A, Caillaud JM, Ozturk M,et al. A monoclonal antibody against a synthetic peptide is spe-cific for the free native human chorionic gonadotropin beta-subunit.Endocrinology 1984;115(1):330–6.
[13] Richmond JF, Mustafa F, Lu S, Santoro JC, Weng J, O’ ConnellM, et al. Screening of HIV-1 Env glycoproteins for the ability toraise neutralizing antibody using DNA immunization and recombi-
[ NAccine
[ s ofnol
[ r JF,instlyDis
[ teinsunol
[ rdiniarrier
[ gnenju-
gated to purified protein derivative or mycobacterial heat shockproteins in the absence of adjuvants. Clin Exp Immunol 1993;93(3):382–6.
[20] Nawrath M, Pavlovic J, Moelling K, et al. Synergistic effectof a combined DNA and peptide vaccine against gp100 in amalignant melanoma mouse model [J]. J Mol Med 2001;79(2–3):133–42.
[21] Gaofu Q, Dan M, Jie W, Liao Z, Li Z, Roques RS, jingjing L, et al.Long-lasting specific antibodies against CETP induced by subcuta-neous and mucosal administration of a 26-amino acid CETP epitopecarried by heat shock protein 65 kDa in the absence of adjuvants.Vaccine 2004;22(23–4):3187–94.
[22] Romero-Steiner S, Holder PF, Gomez de Leno P, Hennessy TW,Carlone GM. Avidity determinations forHaemophilus influenzaeType b anti-polyribosylribitol phosphate antibodies. Clin Diagn LabImmunol 2005;12(9):1029–35.
[23] Gurunathan S, Klinman DM, Seder RA. DNA vaccines: immunology,application, and optimization. Annu Rev Immunol 2000;18:927–74.
[24] Lasaro MO, Luiz WB, Sbrogio-Almeida ME, Ferreira LC. Prime-boost vaccine regimen confers protective immunity to human-derivedenterotoxigenicEscherichia coli. Vaccine 2005;23(19):2430–8.
[25] Putz MM, Ammerlaan W, Schneider F, Jung G, Muller CP.Humoral immune responses to a protective peptide-conjugateagainst measles after different prime-boost regimens. Vaccine2004;22(31–2):4173–82.
[26] Scheerlinck JP, Casey G, McWaters P, Kelly J, Woollard D,Lightowlers MW, et al. The immune response to a DNA vaccinecan be modulated by co-delivery of cytokine genes using a DNAprime-protein boost strategy. Vaccine 2001;19(28–9):4053–60.
[27] Livingston VW, Livingston AM. Some cultural, immunological, andbiochemical properties of Progenitor cryptocides. Trans N Y Acad
[ dentnic.
[ zzionicorec-vival.
[ PJ,r B
[ t al.usingr Res
[ hesingH or2.
nant vaccinia virus boosting. Virology 1997;230(2):265–74.14] Terrazzini N, Hannesdottir S, Dleves PJ, Lund T, et al. D
immunization with plasmids expressing hCGbeta-chimeras. Va2004;22(17–8):2146–53.
15] Ghosh S, Jackson DC. Antigenic and immunogenic propertietotally synthetic peptide-based anti-fertility vaccines. Int Immu1999;11(7):1103–10.
16] Romero-Steiner S, Musher DM, Cetron MS, Pais LB, GrooveFoire AE, et al. Reduction in functional antibody activity agaStreptococcus pneumoniae in vaccinated elderly individuals highcorrelates with decreased IgG antibody avidity. Clin Infect1999;29(2):281–8.
17] Li Z, Menoret A, Srivastara P, et al. Roles of heat-shock proin antigen presentation and cross-presentation. Curr Opin Imm2002;14:45.
18] Lussow AR, Barrios C, van Embden J, Van der Zee R, VeAS, Pessi A, et al. Mycobacterial heat-shock proteins as cmolecules. Eur J Immunol 1991;21(10):2297–302.
19] Perraut R, Lussow AR, Gavollie S, Garraud O, Matile H, TouC, et al. Successful primate immunization with peptides co
Sci 1974;36(6):569–82.28] Kalantarov G, Acevedo HF. Demonstration of dose depen
cytotoxic activity in cancer cells by specific human choriogonadotropin monoclonal antibodies. Cancer 1998;83(4):783–7
29] Moulton HM, Yoshihara PH, Mason DH, Iversen PL, TrioPL. Active specific immunotherapy with a beta-human chorigonadotropin peptide vaccine in patients with metastatic coltal cancer: antibody response is associated with improved surClin Cancer Res 2002;8(7):2044–51.
30] Laylor R, Porakishvili N, De Souza JB, Playfair JH, DelvesLund T. DNA vaccination favours memory rather than effectocell responses. Clin Exp Immunol 1999;117(1):106–12.
31] Hsu CT, Ting CY, Ting CJ, Chen TY, Lin CP, Whang-Peng J, eVaccination against gonadotropin-releasing hormone (GnRH)toxin receptor-binding domain-conjugated GnRH repeats. Cance2000;60(14):3701–5.
32] Jinshu X, Jingjing L, Duan P, Zheng Z, Ding M, Jie W, et al. Timmunogenicity of recombinant and dimeric gonadotrophin-releahormone vaccines incorporating a T-helper epitope and GnRrepeated GnRH units. Immunol Methods 2004;289(1–2):111–2




















