
efeitos do exercicio físico sobre a expressão de receptores de ...
Upload
khangminh22Category
view
1download
0
,PSRUWkQFLD�GD�DQiOLVH�GD�H[SUHVVmR�JrQLFD�HP�WHFLGRV H
FpOXODV�~QLFDV�QR�HVWXGR�GDV�QHXURFLrQFLDV��GR�GHVHQYROYLPHQWR GR
QHRFyUWH[�DR�HVWXGR�GD�GRHQoD�GH�$O]KHLPHU
7HVH�VXEPHWLGD�DR�GHSDUWDPHQWR�GH�%LRLQIRUPiWLFDGD�8QLYHUVLGDGH�)HGHUDO�GR�5LR�*UDQGH�GR�1RUWH
FRPR�SDUWH�GRV�SUp�UHTXLVLWRVSDUD�D�REWHQomR�GR�JUDX�GH�'RXWRU�HP�%LRLQIRUPiWLFD
'LHJR�0��&RHOKR0DUoR������

',(*2�0$548(6�&2(/+2
,PSRUWkQFLD�GD�DQiOLVH�GD�H[SUHVVmR�JrQLFD�HP�WHFLGRV H
FpOXODV�~QLFDV�QR�HVWXGR�GDV�QHXURFLrQFLDV��GR
GHVHQYROYLPHQWR�GR�QHRFyUWH[�DR�HVWXGR�GD�GRHQoD�GH
$O]KHLPHU
'HIHVD GH 'RXWRUDGR DSUHVHQWDGD DR3URJUDPD GH 3yV�*UDGXDomR HP%LRLQIRUPiWLFD SHOD 8QLYHUVLGDGH )HGHUDO GR5LR�*UDQGH�GR�1RUWH
ÈUHD�GH�FRQFHQWUDomR��%LRLQIRUPiWLFD
/LQKD GH 3HVTXLVD� 1HXURFLrQFLD H7UDQVFULSW{PLFD
2ULHQWDGRU��3URI��0DUFRV�5RPXDOGR�&RVWD&R�RULHQWDGRU��3URI��6DQGUR�-RVp�GH�6RX]D
1$7$/�51����

Marques-Coelho, Diego. Importância da análise da expressão gênica em tecidos ecélulas-únicas no estudo das neurociências: do desenvolvimentodo neocórtex ao estudo da doença de Alzheimer / Diego MarquesCoelho. - 2021. 205f.: il.
Tese (Doutorado) - Universidade Federal do Rio Grande doNorte, Instituto Metrópole Digital, Programa de Pós-Graduação emBioinformática, Natal, 2021. Orientador: Dr. Marcos Romualdo Costa. Coorientador: Dr. Sandro José de Souza.
1. Transcriptômica - Tese. 2. Análise da expressão gênica -Tese. 3. Doença de Alzheimer - Tese. 4. Desenvolvimento deneocórtex - Tese. 5. scRNAseq -Tese. I. Costa, Marcos Romualdo.II. Souza, Sandro José de. III. Título.
RN/UF/BCZM CDU 004:577
Universidade Federal do Rio Grande do Norte - UFRNSistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Central Zila Mamede
Elaborado por RAIMUNDO MUNIZ DE OLIVEIRA - CRB-15/429

'HGLFR�HVVD�WHVH�D�PLQKD�IDPtOLD�SDVVDGD�
SUHVHQWH�HIXWXUD

³6H�YLP�DR�PXQGR��IRL�VRPHQWH�SDUD�GHVIORUDU�IORUHVWDV YLUJHQV�
(�GHVHQKDU�PHXV�SUySULRV�SpV�QD�DUHLD�LQH[SORUDGD�
2�PDLV�TXH�IDoR��QmR�YDOH�QDGD´
��-RVp�5pJLR

$JUDGHFLPHQWRV
0DLV�GR�TXH�VDEHU�RV�VDEHUHV��p�WHU�JUDWLGmR��'DGR LVVR��VRX�JUDWR�
¬�)DLJD�0DUTXHV�H�&OiXGLR�&RHOKR��VHP�HOHV�QDGD�VHULD �OLWHUDOPHQWH��
¬ PLQKD TXHULGD IDPtOLD� SRU PH DPDU LPHQVDPHQWH PHVPR HX VHQGR WmR
FKDWR�H�LQGLIHUHQWH�
¬ -XOLD SRFR�SRFR $SRO{QLR� PLQKD HVSRVD� SRU VHU PLQKD FRQILGHQWH H IRUoD
PRWUL]�HP�WHPSRV�TXH�QHP�HX�DFUHGLWDYD�HP�PLP�
$R $JXVWLQ 5LTXHOPH H /HR 0RULWD� SRU PH RXYLU H LQVSLUDU D EXVFDU QRYRV
FDPLQKRV�
$R PHX DPLJR -XOLR 0HQGRQoD� SRU PH ID]HU DFUHGLWDU WDPEpP QD
H[LVWrQFLD�GH�XPD�YLGD�QmR�DFDGrPLFD�
$R 3URI� 0DUFRV &RVWD� SHOD RULHQWDomR� IRL XP LPHQVR SUD]HU DSUHQGHU�
SURGX]LU�FLrQFLD�H�WHU�SOHQD�OLEHUGDGH�GH�HVFROKHU PHXV�FDPLQKRV�
¬ 3URID� &HFtOLD +HGLQ� 3URI� -RmR 0HQH]HV� 3URI� 5RGULJR 'DOPROLQ� SHORV
HQVLQDPHQWRV�TXH�IRUDP�RV�SLODUHV�SDUD�FULDU�PHX�SHQVDPHQWR FLHQWtILFR�
$R 3URI� 8OL 0XHOOHU H 3URID� &ULVWLQD 6DQ] SHOR YRWR GH FRQILDQoD QR PHX
WUDEDOKR�
$R 'DQLOR ,PSDUDWR� $QGUp )DXVWLQR� 5DIIDHO $]HYHGR� ,DUD 6RX]D� .DUOD
7DERVD� &OyYLV 5HLV� 3DXOR %UDQFR� 9LYLDQH 1RJXHLUD H WRGRV RV DPLJRV TXH
GLYLGLUDP�DV�GRUHV�H�DOHJULDV�GLiULDV�GH�VH�WRUQDU XP�ELRLQIRUPDWD�
$R�5HQ]R�7RUUHFXVR�H�$QGUp�6DOOHV��SHOR�ERP�FKRUR �H�DOJXQV�FLJDUURV��
¬ -pVVLFD $OYHV� 6RUDLD %DUmR H &KLK�PLQJ� SRU VHPSUH PH DMXGDU H SHODV
LQFRQWiYHLV�KRUDV�RXYLQGR�DV�PLQKDV�DWURFLGDGHV�VHP HIHWLYDPHQWH�PH�PDWDU�
¬ WRGRV QmR�FLWDGRV GR %LRPH � 8)51� %LRItVLFD � 8)5-� 'HSDUWPHQW RI
1HXURVFLHQFH � -+8 H HPSUHVD 0HQGHOLFV TXH FRQWULEXtUDP SDUD D H[LVWrQFLD GHVWD
WHVH��SHoR�GHVFXOSDV�SRU�QmR�FLWi�ORV�H�YRV�DJUDGHoR�
2 SUHVHQWH WUDEDOKR IRL UHDOL]DGR FRP DSRLR GD &RRUGHQDomR GH
$SHUIHLoRDPHQWR GH 3HVVRDO GH 1tYHO 6XSHULRU � %UDVLO �&$3(6� � &yGLJR GH
)LQDQFLDPHQWR�����

5HVXPR
2 VHTXHQFLDPHQWR GH 51$ PHQVDJHLUR HP ODUJD HVFDOD �51$VHT� SHUPLWHDYDOLDU D GLYHUVLGDGH GH WUDQVFULWRV H[SUHVVRV HP XP GHWHUPLQDGR PRPHQWR GH XPVLVWHPD ELROyJLFR� $WUDYpV GD ELRLQIRUPiWLFD� SRGHPRV DQDOLVDU RV GDGRV GHVHTXHQFLDPHQWR SDUD REWHU LQIRUPDo}HV TXDQWLWDWLYDV VREUH D H[SUHVVmR JrQLFD�WDLV FRPR D H[SUHVVmR GLIHUHQFLDO GH JHQHV H VXDV LVRIRUPDV �VSOLFHV DOWHUQDWLYRV��1HVWD WHVH� DSUHVHQWDPRV GRLV HVWXGRV LQGHSHQGHQWHV TXH VH YDOHUDP GDELRLQIRUPiWLFD SDUD REWHU LQIRUPDo}HV UHOHYDQWHV VREUH GLIHUHQWHV IHQ{PHQRVELROyJLFRV� 1R SULPHLUR FDVR� QyV XWLOL]DPRV GDGRV GH VHTXHQFLDPHQWR GH 51$PHQVDJHLUR HP FpUHEURV GH SDFLHQWHV FRP D GRHQoD GH $O]KHLPHU SDUD HVWXGDU DH[SUHVVmR GLIHUHQFLDO GH JHQHV H WUDQVFULWRV DVVRFLDGDV FRP D SURJUHVVmR GHVWDGRHQoD� 1yV GHPRQVWUDPRV TXH D DQiOLVH GH WUDQVFULWRV SHUPLWH D LGHQWLILFDomR GHDOWHUDo}HV JrQLFDV LJQRUDGDV HP HVWXGRV DQWHULRUHV DYDOLDQGR DSHQDV D H[SUHVVmRJOREDO GRV JHQHV� 8WLOL]DQGR GDGRV GH VHTXHQFLDPHQWR GH 51$ PHQVDJHLUR HPFpOXODV ~QLFDV �VF51$VHT�� QyV WDPEpP PDSHDPRV DV DOWHUDo}HV GD H[SUHVVmRJrQLFD QR FpUHEUR GH SDFLHQWHV FRP D GRHQoD GH $O]KHLPHU SDUD WLSRV FHOXODUHVHVSHFtILFRV� 2V UHVXOWDGRV GHVWH SULPHLUR WUDEDOKR FRQWULEXHP SDUD XPD PHOKRUFRPSUHHQVmR GD SDWRILVLRORJLD GD GRHQoD GH $O]KHLPHU H LQGLFDP SRWHQFLDLVDOWHUDo}HV PROHFXODUHV DVVRFLDGDV FRP D GRHQoD HP WLSRV FHOXODUHV LQGLYLGXDLV� 1RVHJXQGR WUDEDOKR GHVHQYROYLGR QHVWD WHVH� QyV XWLOL]DPRV D WpFQLFD GH VF51$VHTSDUD HVWXGDU D GLYHUVLGDGH GH FpOXODV SURJHQLWRUDV HP HVWiJLRV LQLFLDLV GRGHVHQYROYLPHQWR GR QHRFyUWH[� $WUDYpV GH DQiOLVHV GH H[SUHVVmR GLIHUHQFLDO GHJHQHV H D XWLOL]DomR GH XPD DERUGDJHP XWLOL]DQGR UHGHV UHJXODWyULDV WUDQVFULFLRQDLV�QyV LGHQWLILFDPRV R IDWRU GH WUDQVFULomR 6R[� FRPR XP UHJXODGRU GR FRPSRUWDPHQWRGH GLIHUHQWHV VXEWLSRV GH SURJHQLWRUHV QHXUDLV �+HV�� YV� +HV���� &RQILUPDQGRHVWHV DFKDGRV GD ELRLQIRUPiWLFD� H[SHULPHQWRV JHQpWLFRV SDUD PDQLSXODU RV QtYHLVGH H[SUHVVmR GH 6R[� HP SURJHQLWRUHV QHXUDLV GHPRQVWUDUDP D LPSRUWkQFLD GHVWHIDWRU GH WUDQVFULomR QD UHJXODomR GD SUROLIHUDomR H GLIHUHQFLDomR FHOXODU� (PFRQMXQWR� RV UHVXOWDGRV GHVWD WHVH GHPRQVWUDP D LPSRUWkQFLD GD DQiOLVHWUDQVFULSW{PLFD DWUDYpV GH PpWRGRV FRPSOHPHQWDUHV SDUD XPD PHOKRU LGHQWLILFDomRGDV�DOWHUDo}HV�GD�H[SUHVVmR�UHOHYDQWHV�HP�GLIHUHQWHV FRQWH[WRV�ELROyJLFRV�
3DODYUDV�FKDYHV� WUDQVFULSW{PLFD� DQiOLVH GD H[SUHVVmR JrQLFD� GRHQoD GH
$O]KHLPHU��GHVHQYROYLPHQWR�GR�QHRFyUWH[��VF51$VHT

$EVWUDFW
/DUJH�VFDOH PHVVHQJHU 51$ VHTXHQFLQJ �51$VHT� DOORZV WKH HYDOXDWLRQ RIWKH GLYHUVLW\ RI WUDQVFULSWV H[SUHVVHG DW D JLYHQ PRPHQW LQ D ELRORJLFDO V\VWHP�7KURXJK ELRLQIRUPDWLFV� ZH FDQ DQDO\]H WKH VHTXHQFLQJ GDWD WR REWDLQ TXDQWLWDWLYHLQIRUPDWLRQ DERXW JHQH H[SUHVVLRQ� VXFK DV WKH GLIIHUHQWLDO H[SUHVVLRQ RI JHQHV DQGWKHLU LVRIRUPV �DOWHUQDWLYH VSOLFHV�� ,Q WKLV WKHVLV� ZH SUHVHQW WZR LQGHSHQGHQW VWXGLHVWKDW XVHG ELRLQIRUPDWLFV WR REWDLQ UHOHYDQW LQIRUPDWLRQ DERXW GLIIHUHQW ELRORJLFDOSKHQRPHQD� ,Q WKH ILUVW FDVH� ZH XVHG PHVVHQJHU 51$ VHTXHQFLQJ GDWD LQ WKHEUDLQV RI SDWLHQWV ZLWK $O]KHLPHUV GLVHDVH WR VWXG\ WKH GLIIHUHQWLDO H[SUHVVLRQ RIJHQHV DQG WUDQVFULSWV DVVRFLDWHG ZLWK WKH SURJUHVVLRQ RI WKLV GLVHDVH� :H KDYHVKRZQ WKDW WKH DQDO\VLV RI WUDQVFULSWV DOORZV WKH LGHQWLILFDWLRQ RI JHQHWLF FKDQJHVLJQRUHG LQ SUHYLRXV VWXGLHV E\ HYDOXDWLQJ RQO\ WKH JOREDO H[SUHVVLRQ RI JHQHV� 8VLQJVLQJOH�FHOO PHVVHQJHU 51$ VHTXHQFLQJ GDWD �VF51$VHT�� ZH DOVR PDS FKDQJHV LQJHQH H[SUHVVLRQ LQ WKH EUDLQ RI SDWLHQWV ZLWK $O]KHLPHUV GLVHDVH WR VSHFLILF FHOOW\SHV� 7KH UHVXOWV RI WKLV ILUVW ZRUN FRQWULEXWH WR D EHWWHU XQGHUVWDQGLQJ RI WKHSDWKRSK\VLRORJ\ RI $O]KHLPHUV GLVHDVH DQG SLQSRLQWV SRVVLEOH FHOO�W\SH VSHFLILFPROHFXODU PHFKDQLVPV RI WKH GLVHDVH� ,Q WKH VHFRQG ZRUN GHYHORSHG LQ WKLV WKHVLV�ZH XVHG WKH VF51$VHT WHFKQLTXH WR VWXG\ WKH GLYHUVLW\ RI SURJHQLWRU FHOOV LQ WKHHDUO\ VWDJHV RI WKH GHYHORSPHQW RI WKH QHRFRUWH[� 7KURXJK DQDO\VLV RI GLIIHUHQWLDOJHQH H[SUHVVLRQ DQG WKH XVH RI DQ DSSURDFK XVLQJ UHJXODWRU\ WUDQVFULSWLRQDOQHWZRUNV� ZH LGHQWLILHG WKH WUDQVFULSWLRQ IDFWRU 6R[� DV D UHJXODWRU RI WKH EHKDYLRU RIGLIIHUHQW VXEW\SHV RI QHXUDO SURJHQLWRUV �+HV�� YV� +HV���� &RQILUPLQJ WKHVH ILQGLQJVIURP ELRLQIRUPDWLFV� JHQHWLF H[SHULPHQWV WR PDQLSXODWH 6R[� H[SUHVVLRQ OHYHOV LQQHXUDO SURJHQLWRUV GHPRQVWUDWHG WKH LPSRUWDQFH RI WKLV WUDQVFULSWLRQ IDFWRU LQ WKHUHJXODWLRQ RI FHOO SUROLIHUDWLRQ DQG GLIIHUHQWLDWLRQ� 7RJHWKHU� WKH UHVXOWV RI WKLV WKHVLVGHPRQVWUDWH WKH LPSRUWDQFH RI WUDQVFULSWRPLF DQDO\VLV WKURXJK FRPSOHPHQWDU\PHWKRGV IRU D EHWWHU LGHQWLILFDWLRQ RI UHOHYDQW JHQH H[SUHVVLRQ FKDQJHV LQ GLIIHUHQWELRORJLFDO�FRQWH[WV�
.H\ZRUGV� WUDQVFULSWRPLFV� JHQH H[SUHVVLRQ DQDO\VLV� $O]KHLPHUV GLVHDVH�
QHRFRUWH[�GHYHORSPHQW��VF51$VHT

6XPiULR
/LVWD�GH�ILJXUDV�H�WDEHODV [
/LVWD�GH�DEUHYLDo}HV [L
,QWURGXomR �
'R�HVWXGR�GD�WUDQVFULSW{PLFD �
'HILQLo}HV�H�FRQFHLWRV�EiVLFRV �
'D�WpFQLFD�H�VXDV�QXDQFHV �
$OLQKDGRUHV���,GHQWLILFDQGR�R�FyGLJR�JHQpWLFR �
$OLQKDPHQWR�GLUHWR�GH�UHDGV �
3VHXGR�DOLQKDPHQWR �
([SUHVVmR�GLIHUHQFLDO�GH�JHQHV�H�WUDQVFULWRV �
$QiOLVH�GD�H[SUHVVmR�GLIHUHQFLDO �
8VR�GLIHUHQFLDO�GH�WUDQVFULWRV ��
5HGHV�GH�LQWHUDomR�H�RQWRORJLDV ��
6HTXHQFLDPHQWR�GH�FpOXOD�~QLFD��XPD�QRYD�IURQWHLUD ��
3URYD�GH�FRQFHLWR�H�SULPHLURV�SDVVRV�GR�VHTXHQFLDPHQWR GH�FpOXOD�~QLFD ��
3ULPHLURV�HVWXGRV�SRSXODFLRQDLV ��
&pOXODV�HP�JRWDV�H�D�H[SDQVmR�GDV�IURQWHLUDV ��
$QiOLVHV�GH�VF51$VHT ��
&DVR�GH�HVWXGR�,���$O]KHLPHU�H�VHXV�WUDQVFULWRV ��
$�GRHQoD�GH�$O]KHLPHU�FRPR�D�SULQFLSDO�FDXVD�GH�GHPrQFLD ��
0HFDQLVPRV�GD�SDWRORJLD ��
)DWRUHV�JHQpWLFRV�GD�SDWRORJLD ��
'DGRV�VHTXHQFLDGRV�H�VXDV�GHPRJUDILDV ��
&DVR�GH�HVWXGR�,,���1HXURELRORJLD�GR�GHVHQYROYLPHQWR H�VHXV�WUDQVFULWRV ��
6XUJLPHQWR�GR�QHRFyUWH[�FHUHEUDO ��
2EMHWLYRV ��
&DStWXOR�,���'LIIHUHQWLDO�WUDQVFULSW�XVDJH�XQUDYHOV JHQH�H[SUHVVLRQ�DOWHUDWLRQV�LQ$O]KHLPHU¶V�GLVHDVH�KXPDQ�EUDLQV
&DStWXOR�,,���'LIIHUHQWLDO�H[SUHVVLRQ�OHYHOV�RI�6R[� LQ�HDUO\�QHRFRUWLFDO�UDGLDO�JOLDO�FHOOVUHJXODWH�WKH�GHFLVLRQ�EHWZHHQ�VWHP�FHOO�PDLQWHQDQFH DQG�GLIIHUHQWLDWLRQ
'LVFXVVmR ���
&RQFOXVmR ���
5HIHUrQFLDV ���

$3Ç1',&( ���

/LVWD�GH�ILJXUDV�H�WDEHODV
)LJXUD �� 5HSUHVHQWDomR HVTXHPiWLFD GD IXQFLRQDOLGDGH GRV
SVHXGRDOLQKDGRUHV
)LJXUD������ 7URFD�GH�LVRIRUPDV
)LJXUD������ (VWXGRV�FRP�VHTXHQFLDPHQWR�GH�FpOXOD�~QLFD
)LJXUD������ 3ULQFLSDLV�PDUFDGRUHV�SDWROyJLFRV�GD GRHQoD�$O]KHLPHU
)LJXUD �� )RUPDomR GRV QHXU{QLRV GH SURMHomR QR FyUWH[ GH
FDPXQGRQJR
[

/LVWD�GH�DEUHYLDo}HV
$����6tWLR�GH�VSOLFLQJ�DOWHUQDWLYR��¶�$����6tWLR�GH�VSOLFLQJ�DOWHUQDWLYR��¶�$'���'RHQoD�GH�DO]KHLPHU�$2'���LGDGH�GRV�LQGLYtGXRV�QR�PRPHQWR�GD�PRUWH�$3$���3ROLDGHQLODomR�DOWHUQDWLYD�$776���6tWLR�GH�LQtFLR�WUDQVFULomR�DOWHUQDWLYR�$766���6tWLR�GH�ILP�WUDQVFULomR�DOWHUQDWLYR�F'1$���'1$�FRPSOHPHQWDU�&'5���&ODVVLILFDomR�FOtQLFD�SDUD�GHPrQFLD�&(5$'���&RQVyUFLR�SDUD�HVWDEHOHFHU�XP�UHJLVWUR�GD GRHQoD�GH�DO]KHLPHU�&30���&RQWDJHQV�SRU�PLOK}HV�GH�NLOREDVH�'$���'RHQoD�GH�$O]KHLPHU�'*(���([SUHVVmR�GLIHUHQFLDO�GH�JHQHV�'1$���ÈFLGR�GHVR[LUULERQXFOHLFR�'78���8VR�GLIHUHQFLDO�GH�WUDQVFULWRV�(2$'���'RHQoD�GH�$O]KHLPHU�GH�LQtFLR�FHGR�(6���6DOWR�GH�H[RQ�)3.0���)UDJPHQWRV�GH�UHDGV�PDSHDGRV�SRU�NLOREDVH�,5���5HWHQomR�GH�LQWURQ�/2$'���'RHQoD�GH�$O]KHLPHU�GH�LQtFLR�WDUGLR�0((���([RQ�PXWXDPHQWH�H[FOXVLYRV�0(6���6DOWR�P~OWLSOR�GH�H[RQV�P51$���51$�PHQVDJHLUR�0$<2�� 0D\R�&OLQLF�06%%�� 0RXQW�6LQDL�%UDLQ�%DQN�QF51$���51$�QmR�FRGDQWH�1)7���(PDUDQKDGRV�QHXURILEULODUHV�1*6���6HTXHQFLDPHQWR�GH�VHJXQGD�JHUDomR�13���3ODFDV�DPLOyLGHV�3&5���5HDomR�HP�FDGHLD�GD�SROLPHUDVH�30,���LQWHUYDOR SRVW�PRUWHP GH�FROHWD�5*���JOLD�UDGLDO�5,1���YDORU�GH�LQWHJULGDGH�GR�51$�51$���ÈFLGR�ULERQXFOpLFR�51$�VHT���6HTXHQFLDPHQWR�GH�51$�5260$3���5HOLJLRXV�2UGHUV�6WXG\���5XVK�0HPRU\�DQG $JLQJ�3URMHFW�U51$���51$�ULERVV{PLFR�571���5HGHV�UHJXODWyULDV�WUDQVFULFLRQDLV�
[L

VF51$VHT���6HTXHQFLDPHQWR�GH�51$VHT�GH�FpOXOD�~QLFD�V51$���51$�SHTXHQR�69=���=RQD�VXEYHQWULFXODU�730���7UDQVFULWRV�SRU�PLOK}HV�GH�NLOREDVH�766���6tWLRV�GH�LQtFLR�GH�WUDQVFULomR�875���5HJLmR�QmR�WUDGX]LGD�9=���=RQD�YHQWULFXODU�
[LL

,QWURGXomR
2 GRJPD FHQWUDO GD ELRORJLD PROHFXODU GHILQH FRPR RV WUrV SLODUHV GD
WUDQVPLVVmR GD LQIRUPDomR HQWUH RV VHUHV YLYRV� iFLGR GHVR[LUULERQXFOpLFR �'1$� GR
LQJOrV� GHR[\ULERQXFOHLF DFLG�� iFLGR ULERQXFOpLFR �51$� GR LQJOrV� ULERQXFOHLF DFLG� H
SURWHtQDV��1D�SUHVHQWH�WHVH��YDPRV�IRFDU�HP�PHFDQLVPRV GD�PROpFXOD�51$�
'R�HVWXGR�GD�WUDQVFULSW{PLFD
'HILQLo}HV�H�FRQFHLWRV�EiVLFRV
$ WUDQVFULSW{PLFD ID] SDUWH GR EORFR GDV FLrQFLDV ³{PLFDV´ �FRPR JHQ{PLFD�
SURWH{PLFD� HSLJHQ{PLFD� HWF�� VHQGR HOD D UHVSRQViYHO SRU HVWXGDU R WUDQVFULSWRPD
GRV VHUHV YLYRV� 7UDQVFULSWRPD p R FRQMXQWR GH WRGRV RV WUDQVFULWRV HP XP VLVWHPD
ELROyJLFR HP XP GHWHUPLQDGR SHUtRGR H�RX HVWDGR ILVLROyJLFR� 1D WUDQVFULSW{PLFD VH
EXVFD FDWDORJDU WRGRV RV WLSRV GH WUDQVFULWR� FRPR 51$ PHQVDJHLUR �P51$� GR
LQJOrV� PHVVHQJHU 51$�� 51$ QmR�FRGDQWH �QF51$� GR LQJOrV� QRQ�FRGLQJ 51$� H
51$ SHTXHQR �V51$� GR LQJOrV� VPDOO 51$� �:DQJ HW DO� ������ 2 VHTXHQFLDPHQWR
GH VHJXQGD JHUDomR �1*6� GR LQJOrV� QH[W JHQHUDWLRQ VHTXHQFLQJ� SHUPLWLX WDQWR R
PDSHDPHQWR TXDQWR D TXDQWLILFDomR GR 51$� HVVH PpWRGR p FKDPDGR
VHTXHQFLDPHQWR GH ODUJD HVFDOD GH 51$ �51$�VHT� GR LQJOrV� 51$ VHTXHQFLQJ��
0DLV HVSHFLILFDPHQWH� FRP D TXDQWLILFDomR GR P51$ FRQVHJXLPRV REWHU GDGRV GD
H[SUHVVmR JrQLFD H� LQGLUHWDPHQWH� SURWHtQDV� $ SDUWLU GHVWHV GDGRV� SRGH�VH ID]HU
�

LQIHUrQFLDV VREUH RV HYHQWRV PROHFXODUHV H FHOXODUHV RSHUDQGR HP XP VLVWHPD
ELROyJLFR��FRQVLGHUDQGR�RV�FRQKHFLPHQWRV�SUpYLRV�VREUH DV�SURWHtQDV�HP�TXHVWmR�
'D�WpFQLFD�H�VXDV�QXDQFHV
$LQGD TXH D WpFQLFD GH PLFURDUUD\ WHQKD SHUPLWLGR D LGHQWLILFDomR H
TXDQWLILFDomR GH WUDQVFULWRV QmR GHVFULWRV HP GLYHUVRV PRGHORV �%HUWRQH HW DO� �����
<DPDGD HW DO� ������ DOJXPDV OLPLWDo}HV� WDLV FRPR R DOWR QtYHO GH KLEULGL]DomR
FUX]DGD �FURVV�K\EULGL]DWLRQ�� D DPSOLWXGH GRV WUDQVFULWRV H D EDL[D FDSDFLGDGH GH
FRPSDUDomR HQWUH GLIHUHQWHV H[SHULPHQWRV �%HUWRQH HW DO� ����� 2NRQLHZVNL HW DO�
������ OLPLWDP R XVR GHVWD WpFQLFD SDUD D DYDOLDomR GD H[SUHVVmR JrQLFD HP ODUJD
HVFDOD��
3RU RXWUR ODGR� R 51$�VHT SHUPLWH D LGHQWLILFDomR GR P51$ GH WRGRV RV
WUDQVFULWRV SUHVHQWHV QXP PHLR� VHQGR TXH DWXDOPHQWH� R 51$�VHT Mi SRVVXL FXVWRV
HTXLSDUiYHLV H XPD LPSOHPHQWDomR PDLV FRQILiYHO TXH XP H[SHULPHQWR FRP
PLFURDUUD\ �6WDUN HW DO� ������ $OpP GLVVR� D WpFQLFD 51$�VHT WDPEpP DEULX QRYDV
IURQWHLUDV SDUD D GHVFREHUWD GH QRYDV LVRIRUPDV UDUDV� FRQVHTXHQWHPHQWH� PDLV
VtWLRV GH VSOLFLQJ H H[SUHVVmR GLIHUHQFLDO GH LVRIRUPDV� R TXH VHULD LPSURYiYHO FRP
PLFURDUUD\ GHYLGR D QHFHVVLGDGH GH DOWD TXDQWLGDGH GH 51$ �6XOWDQ HW DO� �����
:DQJ�HW�DO��������
'LIHUHQWH GH XP VHTXHQFLDPHQWR 1*6 GH '1$� SDUD 51$�VHT VmR
QHFHVViULRV FHUWRV SDVVRV TXH LQIOXHQFLDP GLUHWDPHQWH QRV UHVXOWDGRV GR
VHTXHQFLDPHQWR� VmR HOHV� FDSWXUD GR 51$� SUHSDUDomR GH ELEOLRWHFDV GH '1$
FRPSOHPHQWDU �F'1$� GR LQJOrV� FRPSOHPHQWDU\ '1$� EDQGD�HVSHFtILFD�
�

HQULTXHFLPHQWR �¶ RX �¶� XVR GH LGHQWLILFDGRUHV PROHFXODUHV ~QLFRV �80,� GR LQJOrV�
XQLTXH�PROHFXODU�LGHQWLILHUV��H�IUDJPHQWDomR�
$ FDSWXUD GH 51$ DIHWD GLUHWDPHQWH TXDO WLSR GH 51$ GHYHUi VHU
HQULTXHFLGR� &RPR a��� GR 51$ SUHVHQWH HP DPRVWUDV p GH 51$ ULERVV{PLFR
�U51$� GR LQJOrV� ULERQXFOHLF 51$�� p HVVHQFLDO TXH H[LVWD D VHSDUDomR GH P51$
GRV RXWURV WLSRV �0RUODQ HW DO� ������ &DEH VDOLHQWDU TXH D FDSWXUD GH P51$� SRU
PHLR GD FDXGD SROL$� JHUD XP YLpV QD UHJLmR QmR�WUDGX]LGD �¶ ��¶�875� GR LQJOrV� �¶
XQWUDQVODWHG UHJLRQ�� RX VHMD� VRPHQWH D UHJLmR SUy[LPD D SDUWH ILQDO GR P51$ p
VHTXHQFLDGD� &RPR RV JHQHV QmR QHFHVVLWDP GH WRGR R VHX P51$ VHTXHQFLDGR
SDUD D H[HFXomR GH DQiOLVHV FRPR H[SUHVVmR GLIHUHQFLDO �'*(� GR LQJOrV�
GLIIHUHQWLDO JHQH H[SUHVVLRQ�� HVVH p XP PpWRGR EDUDWR H LGHDO SDUD WDLV ILQDOLGDGHV
�%HFN HW DO� ������ $ UHPRomR GLUHWD GH U51$ �RX GHSOHomR� VH SURYRX XPD ERD
DOWHUQDWLYD SDUD R HQULTXHFLPHQWR GH P51$ �0RUODQ HW DO� ������ DSHVDU GH VHU
QHFHVViULR XPD PDLRU FREHUWXUD �UHDGV SRU JHQH�WUDQVFULWR VHTXHQFLDGDV� SRU
H[SHULPHQWR �=KDR HW DO� ������ e LPSRUWDQWH DILUPDU TXH DPEDV DV WpFQLFDV
SRGHUmR VHU XVDGDV HP H[SHULPHQWRV YROWDGRV SDUD D H[SUHVVmR GLIHUHQFLDO GH
JHQHV�
$ IUDJPHQWDomR GR PDWHULDO JHQpWLFR p RXWUR SDVVR LPSRUWDQWH� H[LVWHP GRLV
WLSRV� IUDJPHQWDomR GR F'1$ RX IUDJPHQWDomR GR 51$� $ SULPHLUD FDXVD XPD
SUHIHUrQFLD QDWXUDO GD UHJLmR �¶ SDUD UHJLmR �¶ GR WUDQVFULWR �1DJDODNVKPL HW DO�
������ VHQGR D VHJXQGD PDLV KRPRJrQHD� PDV FRP SHUGD QDV H[WUHPLGDGHV GRV
WUDQVFULWRV��0RUWD]DYL�HW�DO��������
�

2 HQULTXHFLPHQWR GDV UHJL}HV ��¶ H ��¶ VmR GXDV SURSRVWDV YiOLGDV SDUD R
VHTXHQFLDPHQWR GH P51$� 2 HQULTXHFLPHQWR GH 51$ ��¶ ��¶�VHT� p EDVWDQWH XVDGR
SDUD DQiOLVHV GH H[SUHVVmR GLIHUHQFLDO �0ROO HW DO� ������ HQTXDQWR R HQULTXHFLPHQWR
GH 51$ ��¶ ��¶�VHT� p PDLV DSURSULDGR SDUD LGHQWLILFDomR GH SURPRWRUHV H VtWLRV GH
LQtFLR GD WUDQVFULomR �766V� GR LQJOrV� WUDQVFULSWLRQ VWDUW VLWHV� �%DWXW HW DO� �����
,VODP�HW�DO��������
&DEH QRWDU TXH �¶�VHT SRGHUi GLVSHQVDU R XVR GD FDXGD SROL$� R TXH WRUQD R
PpWRGR DLQGD PDLV EDUDWR FRP XPD SUHFLVmR PDLRU FRP PHQRV UHDGV �+HU]RJ HW DO�
������ $OpP GLVVR� H[LVWH D SRVVLELOLGDGH GH GLIHUHQFLDU UHDGV QR �¶�VHT TXDQWR D
SROLDGHQLODomR DOWHUQDWLYD �$3$� GR LQJOrV� SRO\DGHQ\ODWLRQ DOWHUQDWLYH� �7LDQ HW DO�
������ $LQGD DVVLP� H[LVWH XPD JUDQGH OLPLWDomR SDUD GLVWLQJXLU LVRIRUPDV XVDQGR R
PpWRGR �¶�VHT� R TXH R WRUQDULD XPD RSomR LQGHVHMDGD SDUD HVWXGRV TXH EXVFDP
GLVWLQJXLU�H�GHVFREULU�QRYDV�LVRIRUPDV�
3RU ILP� R XVR GH 80, HP H[SHULPHQWRV GH 51$�VHT p UHFRPHQGDGR SDUD
GLPLQXLU XP ELDV FDXVDGR SHOD DPSOLILFDomR GR F'1$ GXUDQWH D HWDSD GH UHDomR HP
FDGHLD GD SROLPHUDVH �3&5� GR LQJOrV� SRO\PHUDVH FKDLQ UHDFWLRQ� FRQKHFLGR FRPR
WD[D�GH�GXSOLFDomR��.LYLRMD�HW�DO��������6PLWK�HW DO��������
8PD YH] TXH D LQIRUPDomR ELROyJLFD p VHTXHQFLDGD� Ki D QHFHVVLGDGH GH
LQWHUSUHWDUPRV WRGD HVVD LQIRUPDomR EUXWD� EDVLFDPHQWH DV VHTXrQFLDV GH SDUHV GH
EDVHV� 7DLV VHTXrQFLDV VHUmR FKDPDGDV GH UHDGV� TXH VmR DV OHLWXUDV DGYLQGDV GRV
VHTXHQFLDGRUHV� 3DUDOHODPHQWH� GXDV RQGDV VXUJLUDP� GH XP ODGR� DOLQKDGRUHV
SUySULRV SDUD 51$VHT� DOJRULWPRV FDSD]HV GH TXDQWLILFDU TXDQWRV UHDGV HVWDULDP
UHODFLRQDGRV D JHQHV H�RX WUDQVFULWRV� H GR RXWUR ODGR� DQiOLVH GH H[SUHVVmR
�

GLIHUHQFLDO SURSULDPHQWH GLWD� DOJRULWPRV TXH DYDOLDP DV PDWUL]HV JHQH �RX
WUDQVFULWRV� YV� DPRVWUDV H GHWHUPLQDP DV DOWHUDo}HV HQWUH GRLV VLVWHPDV RX
FRQGLo}HV�ELROyJLFDV�
6HQGR DVVLP� QD SUy[LPD SDUWH� YDPRV HQWHQGHU FRPR LQWHUSUHWDU RV GDGRV
EUXWRV H WDPEpP YHUHPRV FRPR RV QRYRV DOJRULWPRV TXH VXUJLUDP PROGDUDP D
DQiOLVH�GR�WUDQVFULSWRPD�GHVGH�R�DOLQKDPHQWR�DWp�D H[SUHVVmR�GLIHUHQFLDO�
$OLQKDGRUHV���,GHQWLILFDQGR�R�FyGLJR�JHQpWLFR
'HSRLV TXH R GDGR EUXWR p JHUDGR� SUHFLVDPRV UHODFLRQDU DV UHDGV�
VHTXrQFLDV GH EDVHV JHUDGDV SHOR VHTXHQFLDPHQWR� D VHXV UHVSHFWLYRV WUDQVFULWRV�
3DUD WDO WDUHID XWLOL]DPRV RV DOLQKDGRUHV TXH VmR EDVLFDPHQWH DOJRULWPRV
GHVHQYROYLGRV�SDUD�GDU�D�ORFDOL]DomR�GDV UHDGV QR JHQRPD�
$OLQKDPHQWR�GLUHWR�GH�UHDGV
$OLQKDGRUHV GLUHWRV VmR DTXHOHV TXH ID]HP XP DOLQKDPHQWR GDV UHDGV
VHTXHQFLDGDV D XP JHQRPD�WUDQVFULSWRPD UHIHUrQFLD� 7DLV DOLQKDGRUHV WHP XP
FXVWR FRPSXWDFLRQDO HOHYDGR H WHQGHP D OHYDU KRUDV D GLDV SDUD FRPSOHWDU VXD
WDUHID� +RMH� RV WUrV DOLQKDGRUHV PDLV XWLOL]DGRV VmR� 7RS+DW� �.LP HW DO� ������
+LVDW���.LP�HW�DO��������H�67$5��'RELQ�HW�DO��������
'HSRLV GR DOLQKDPHQWR SURSULDPHQWH GLWR DLQGD VmR QHFHVViULDV GXDV
HWDSDV� FRQWDJHP H QRUPDOL]DomR� $ FRQWDJHP p UHVSRQViYHO SRU IRUQHFHU TXDQWDV
�

UHDGV IRUDP DOLQKDGDV SDUD FDGD JHQH�WUDQVFULWR� VHQGR DOJXQV DOJRULWPRV
FRQKHFLGRV��+76HT��$QGHUV�HW�DO��������H�&XIIOLQNV �7UDSQHOO�HW�DO��������
-i D QRUPDOL]DomR SRQGHUD SHFXOLDULGDGHV GH FDGD JHQH �SRU H[HPSOR�
WDPDQKR� GLIHUHQWHV GH WUDQVFULWRV� FRQWH~GR *&� HWF� H JHUD XP YDORU FRPSDUiYHO
SDUD DQiOLVHV IXWXUDV� 7DLV YDORUHV GHYHUmR UHWLUDU R YLpV JHUDGR SRU
VHTXHQFLDPHQWRV GLIHUHQWHV� WRUQDQGR H[SHULPHQWRV GLYHUVRV FRPSDUiYHLV�
$OJXPDV PHGLGDV FRQKHFLGDV VmR IUDJPHQWRV GH UHDGV PDSHDGRV SRU NLOREDVH
�)3.0� GR LQJOrV� IUDJPHQWV SHU NLOREDVH RI PDSSHG UHDGV�� WUDQVFULWRV SRU PLOK}HV
GH NLOREDVH �730� GR LQJOrV� WUDQVFULSWV SHU PLOOLRQ NLOREDVH� H FRQWDJHQV SRU
PLOK}HV GH NLOREDVH �&30� GR LQJOrV� FRXQWV SHU PLOOLRQ NLOREDVH�� e LPSRUWDQWH
VDOLHQWDU TXH PHGLGDV FRPR )3.0 QmR GHYHUmR VHU XVDGDV HP DQiOLVHV IXWXUDV
FRPR H[SUHVVmR GLIHUHQFLDO� VHQGR 730 XPD PHGLGD DSURSULDGD SDUD HVVH WLSR GH
DQiOLVH�
3VHXGR�DOLQKDPHQWR
(P ����� %UD\ H FRODERUDGRUHV LQWURGX]LUDP XP QRYR FRQFHLWR GH SVHXGR
DOLQKDGRUHV TXH VmR EDVLFDPHQWH TXDQWLILFDGRUHV GH DEXQGkQFLD GH WUDQVFULWRV
�%UD\ HW DO� ������ $ LGHLD QmR HUD PDLV DOLQKDU DV UHDGV D XPD GHWHUPLQDGD UHJLmR
GH�XP�WUDQVFULWR�H�VLP�GHVFREULU�D�FRPSDWLELOLGDGH GH�FDGD�WUDQVFULWR��)LJXUD����
2 DOJRULWPR NDOOLVWR GHVHQYROYLGR SHOR JUXSR FRQVHJXLD REWHU XPD DFXUiFLD
VLPLODU DRV SULQFLSDLV DOLQKDGRUHV GD pSRFD� H PDLV LPSRUWDQWH� FRP XP WHPSR GH
H[HFXomR PLOKDUHV GH YH]HV PHQRU H FRP XPD FDSDFLGDGH FRPSXWDFLRQDO
UHODWLYDPHQWH EDL[D �%UD\ HW DO� ������ $OpP GLVVR� DV WDUHIDV GH FRQWDJHP H
�

QRUPDOL]DomR Mi VmR H[HFXWDGDV MXQWR FRP R SVHXGR DOLQKDPHQWR� R TXH DFHOHUD
DLQGD�PDLV�R�SURFHVVR�
$LQGD FDEH FLWDU TXH� HP ����� 3DWUR H FRODERUDGRUHV Mi KDYLDP LQWURGX]LGR
R FRQFHLWR FRP R DOJRULWPR 6DLOILVK �3DWUR HW DO� ����� H� HP ����� KRXYH XPD
DWXDOL]DomR�GR�PHVPR�FRP�R�DOJRULWPR 6DOPRQ �3DWUR HW�DO��������
'DGR TXH DV UHDGV IRUDP LGHQWLILFDGDV H Mi VDEHPRV D TXDO WUDQVFULWR�JHQH
HVVDV UHDGV SHUWHQFHP� QRV FDEH LQWHUSUHWDU TXDLV IRUDP DV DOWHUDo}HV HQFRQWUDGDV
QDV FRPSDUDo}HV� /RJR� QD SUy[LPD SDUWH� YDPRV HQWHQGHU FRPR D DQiOLVH GH
H[SUHVVmR�GLIHUHQFLDO�QRV�SHUPLWH�LGHQWLILFDU�DV�DOWHUDo}HV HQWUH�DPRVWUDV�
�

)LJXUD �� 5HSUHVHQWDomR HVTXHPiWLFD GD IXQFLRQDOLGDGH GRVSVHXGRDOLQKDGRUHV� 1HVVH H[HPSOR� �$� XP UHDG �SUHWR� WHQWDULD VH DOLQKDU HP �SRVVtYHLV WUDQVFULWRV �FRORULGRV� GLIHUHQWHV� �%� 8P LQGH[ p IRUPDGR FRP QyV�OHLD�VH� N�PHUV� FRPSDUWLOKDGRV HQWUH GLYHUVRV WUDQVFULWRV �FRORULGR�� 2X VHMD�HQWHQGD FDGD WUDQVFULWR FRPR XP FDPLQKR� �&� 3DUD H[HPSOLILFDU� QHVVH PDSD RV �SULPHLURV QyV FRUUHVSRQGHP DR UHDG SUHWR� (P �'�� YHULILFDPRV D HILFLrQFLD GRSVHXGRDOLQKDPHQWR GHYLGR DR VDOWR SRVVtYHO HQWUH RV QyV UHGXQGDQWHV �GH 9� SDUD9��� �(� 3RU ILP� D LQWHUVHomR HQWUH DV SRVVLELOLGDGHV GRV QyV p HVFROKLGD� $GDSWDGRGH�%UD\�HW�DO�������
�

([SUHVVmR�GLIHUHQFLDO�GH�JHQHV�H�WUDQVFULWRV
$ DQiOLVH GD H[SUHVVmR GLIHUHQFLDO p D PHWRGRORJLD PDLV FRPXP DGRWDGD QD
WUDQVFULSW{PLFD� 6mR QHFHVViULRV DOJR HQWUH �� D �� PLOK}HV GH ³UHDGV´ SRU DPRVWUD
SDUD XP H[SHULPHQWR FRP '*( �6WDUN HW DO� ������ $OpP GLVVR� Mi VRPRV FDSD]HV
GH YHU DOWHUDo}HV HP QtYHLV GH WUDQVFULWRV� VHMD FRP H[SUHVVmR GLIHUHQFLDO RX XVR
GLIHUHQFLDO��/RYH�HW�DO��������9LWWLQJ�6HHUXS�HW�DO� ������
$QiOLVH�GD�H[SUHVVmR�GLIHUHQFLDO
$ DQiOLVH GH H[SUHVVmR GLIHUHQFLDO p XPD PHWRGRORJLD TXH EXVFD HQFRQWUDU
DV GLIHUHQoDV QD H[SUHVVmR GRV JHQHV�WUDQVFULWRV GH XP GHWHUPLQDGR HYHQWR
ELROyJLFR� RX VHMD� HVVD GLIHUHQoD SRGH VHU HQFRQWUDGD HQWUH� VDXGiYHO YV�
SDWROyJLFR� GLIHUHQWHV LGDGHV GR GHVHQYROYLPHQWR� HQWUH WLSR FHOXODUHV GLIHUHQWHV�
GHQWUH�RXWURV�
$ DQiOLVH GH H[SUHVVmR GLIHUHQFLDO FRPHoRX FRP R XVR GH GDGRV GH
PLFURDUUD\ H D WHQWDWLYD GH WRUQDU DQiOLVHV JHQ{PLFDV PDLV DFHVVtYHLV H
UHSURGXWtYHLV� YLGH R UHSRVLWyULR ELRFRQGXFWRU �*HQWOHPDQ HW DO� ����� 6P\WK HW DO�
����� 3HDUW HW DO� ������ 2 VRIWZDUH GH 6P\WK H FRODERUDGRUHV� OLPPD� IRL SLRQHLUR
QHVVH WLSR GH DQiOLVH� VHQGR HOH XWLOL]DGR WDPEpP SDUD H[SHULPHQWRV GH 3&5�
DUUDQMR GH SURWHtQD H DGDSWDGR SRVWHULRUPHQWH SDUD DQiOLVH GH 51$VHT HP JHQHV H
VtWLRV GH VSOLFLQJ �GHVGH TXH KRXYHVVH D LQIRUPDomR D QtYHO GH p[RQ� �6P\WK HW DO�
������+XEHUW�HW�DO��������0DQQVSHUJHU�HW�DO������� 5LWFKLH�HW�DO��������
�

3RVWHULRUPHQWH� VXUJLUDP RXWURV DOJRULWPRV FRPR '(6HT� �/RYH HW DO�
������ HGJH5 �5RELQVRQ HW DO� ������ VHQGR TXH FRP R OLPPD �5LWFKLH HW DO� ������
HVVHV VmR RV WUrV DOJRULWPRV PDLV XVDGRV DWXDOPHQWH� 7DQWR '(6HT� TXDQWR HGJH5
VHJXHP XP PRGHOR VHPHOKDQWH XVDQGR XPD GLVWULEXLomR ELQRPLDO QHJDWLYD�
HQTXDQWR TXH R OLPPD DVVXPH XPD GLVWULEXLomR ORJDUtWPLFD� &DEH QRWDU TXH SDUD
GHVHQKRV H[SHULPHQWDLV FRP XPD TXDQWLGDGH FRQVLGHUiYHO GH UXtGR� PpWRGRV FRPR
HGJH5 H '(6HT� VmR PDLV UHFRPHQGDGRV �%DLN HW DO� ������ $OpP GLVVR� QRYDV
PHWRGRORJLDV EXVFDP XOWUDSDVVDU D UHOHYkQFLD GD H[SUHVVmR QR QtYHO GH JHQHV H
HQWHQGHU WDPEpP R TXDQWR RV WUDQVFULWRV SRGHP DMXGDU HP GLIHUHQWHV FRPSDUDo}HV
�<L�HW�DO��������
$SHVDU GD H[SUHVVmR GLIHUHQFLDO GH JHQHV VHU XPD DQiOLVH EDVWDQWH GLUHWD H
H[WHQVDPHQWH XVDGD� D DQiOLVH GLIHUHQFLDO GH WUDQVFULWRV DLQGD p XPD WpFQLFD FRP
EDVWDQWH UXtGR H SRXFR XWLOL]DGD� 1R SUy[LPR WUHFKR� YDPRV YHU FRPR RV WUDQVFULWRV
SRGHP VHU DQDOLVDGRV SDUD JHUDU XPD LQIRUPDomR VREUH HYHQWRV FRPR VSOLFLQJ
DOWHUQDWLYR�
8VR�GLIHUHQFLDO�GH�WUDQVFULWRV
&RP R DYDQoR GDV WpFQLFDV GH 51$VHT H IHUUDPHQWDV DOLQKDPHQWR FRPR
.DOOLVWR� H[LVWH D SRVVLELOLGDGH GD TXDQWLILFDomR GR WUDQVFULSWRPD D QtYHO GH
LVRIRUPDV� &RP LVVR� SRGHPRV GHVFUHYHU D DOWHUDomR LQGLYLGXDO GH LVRIRUPDV H D
SRVVtYHO WURFD GH LVRIRUPDV �9LWWLQJ�6HHUXS HW DO� ������ 2 XVR GLIHUHQFLDO GH
WUDQVFULWRV �'78� GR LQJOrV� GLIIHUHQWLDO WUDQVFULSW XVDJH� QDGD PDLV p TXH D
��

TXDQWLILFDomR GD DOWHUDomR GH WUDQVFULWRV SRU PHLR GR XVR GD IUDomR GDV LVRIRUPDV
�,)��GR�LQJOrV� LVRIRUP�IUDFWLRQ���)LJXUD����
$ ELEOLRWHFD ,VRIRUP6ZLWFK$QDO\]H5 �9LWWLQJ�6HHUXS HW DO� ����� XQLILFD
GLYHUVDV DERUGDJHQV GH XVR GLIHUHQFLDO GH WUDQVFULWRV FRPR '5,06HT �1RZLFND HW
DO� ����� H '(;6HT �$QGHUV HW DO� ������ DVVLP FRPR DQiOLVHV SDUD SUHGLomR GH
GRPtQLRV SURWpLFRV� FRQVHTXrQFLD GH XVR IXQFLRQDO GDV VHTXrQFLDV H VSOLFLQJ
DOWHUQDWLYR �9LWWLQJ�6HHUXS HW DO� ������ &DEH QRWDU TXH R JDQKR RX SHUGD GH XPD
LVRIRUPD� VHP XPD WURFD �JDQKR H SHUGD VLPXOWkQHD�� SRGHUi VHU WDPEpP
LQWHUSUHWDGR�FRPR�XVR�GLIHUHQFLDO�
e UHOHYDQWH GL]HU TXH QmR VH GHYH FRQIXQGLU D �H[SUHVVmR GLIHUHQFLDO GH
WUDQVFULWRV� �'7(� GR LQJOrV� GLIIHUHQWLDO WUDQVFULSW H[SUHVVLRQ� FRP ³XVR GLIHUHQFLDO
GH WUDQVFULWRV´ �'78� GR LQJOrV� GLIIHUHQWLDO WUDQVFULSW XVDJH�� $PERV IRUQHFHP
LQIRUPDomR ³GLIHUHQFLDO´ VREUH RV WUDQVFULWRV �/RYH HW DO� ������ FRQWXGR RV PpWRGRV
GH FiOFXOR VmR EDVWDQWH GLIHUHQWHV� '7( IXQFLRQD H[DWDPHQWH LJXDO DR '*(� Vy TXH
FRP WUDQVFULWRV� '78 OHYD HP FRQVLGHUDomR D IUDomR GH XP WUDQVFULWR �,)� GR LQJOrV�
LVRIRUP IUDFWLRQ� SDUD XP GHWHUPLQDGR JHQH �)LJXUD ��� &RP R XVR GD GLIHUHQoD GH
,) p SRVVtYHO REWHU YDORUHV UHOHYDQWHV SDUD LGHQWLILFDU VH KRXYH DOWHUDo}HV QR QtYHO
GH�GHWHUPLQDGR�WUDQVFULWR�
��

)LJXUD �� 7URFD GH LVRIRUPDV� 1D SDUWH HVTXHUGD� WHPRV GXDV LVRIRUPDV� XPDVDXGiYHO H XPD SDWROyJLFD� $ WURFD GHPRQVWUD R DXPHQWR SUHIHUHQFLDO GH XPD GDVLVRIRUPDV QD FRQGLomR SDWRJrQLFD YV� VDXGiYHO� 1D SDUWH GLUHLWD� WHPRV R H[HPSORGR FiOFXOR FRP R XVR GDV LVRIRUPDV� $ GLIHUHQoD QD IUDomR GD LVRIRUPD �G,)� LQGLFD RDXPHQWR�RX�GLPLQXLomR�GH�XPD�LVRIRUPD�QD�FRPSDUDomR HQWUH�FRQGLo}HV�
��

5HGHV�GH�LQWHUDomR�H�RQWRORJLDV
$SyV D DQiOLVH GH H[SUHVVmR GLIHUHQFLDO� p VHPSUH DGHTXDGR WHQWDU DYDQoDU
QD DYDOLDomR GD VLJQLILFkQFLD GDV DOWHUDo}HV JrQLFDV HQFRQWUDGDV� 3DUD LVVR�
H[LVWHP GXDV PHWRGRORJLDV GH JUDQGH YDORU SDUD R PRPHQWR DSyV DQiOLVH
GLIHUHQFLDO��DV�UHGHV�GH�LQWHUDomR�H�DV�RQWRORJLDV�
$V UHGHV GH LQWHUDomR EXVFDP HQWHQGHU D GLQkPLFD GH DomR HQWUH GLIHUHQWHV
JHQHV�SURWHtQDV� 2 JUXSR GH /DQJIHOGHU H FRODERUDGRUHV IRL FDSD] GH FULDU XP
DOJRULWPR� :*&1$ �GR LQJOrV� ZHLJKWHG FRUUHODWLRQ QHWZRUN DQDO\VLV�� H[WHQVDPHQWH
XVDGR HP YDULDGDV DQiOLVHV� GHVGH WHFLGR DWp FpOXOD ~QLFD �/DQJIHOGHU HW DO� ������
2 :*&1$ VH EDVHLD QXPD UHGH GH FRH[SUHVVmR QmR�GLUHFLRQDGD� RQGH RV ³QyV´
VmR D H[SUHVVmR GR JHQH H RV YpUWLFHV D FRUUHODomR HQWUH GRLV JHQHV GLIHUHQWHV�
0yGXORV GH FRH[SUHVVmR VmR EDVLFDPHQWH RV DJUXSDPHQWRV GH JHQHV DOWDPHQWH
FRUUHODFLRQDGRV HQFRQWUDGRV GHQWUR GH XPD UHGH� RX VHMD� VmR XP GHWHUPLQDGR
JUXSR�GH�JHQHV�TXH�DWXDP�FRQMXQWDPHQWH�HP�XPD�UHGH GH�FRH[SUHVVmR�LQIHULGD�
2XWUR WLSR GH DERUGDJHP VHPHOKDQWH VmR DV UHGHV UHJXODWyULDV
WUDQVFULFLRQDLV H R DOJRULWPR GHVHQYROYLGR SRU &DVWUR H FRODERUDGRUHV �&DVWUR HW DO�
������ 1HVVD UHGH WHPRV RV IDWRUHV GH WUDQVFULomR FRPR FHQWUDLV QD UHGH GH
LQWHUDomR� VHQGR HOD XPD UHGH GLUHFLRQDO RQGH RV ³QyV´ VmR IRUPDGRV SRU IDWRUHV GH
WUDQVFULomR H VHXV YpUWLFHV LQGLFDP D SUR[LPLGDGH HQWUH ³QyV´ SRU PHLR GH PHGLGDV
FRPR D GLVWkQFLD GH -DFFDUG� 8PD LQIRUPDomR UHOHYDQWH QHVVD UHGH p TXH RV
IDWRUHV GH WUDQVFULomR VmR LQWHUSUHWDGRV FRPR UHJXORQV TXH p R FRQMXQWR GR IDWRU GH
WUDQVFULomR H WRGRV RV JHQHV DR TXDO HOH LQIOXHQFLD� ,QIRUPDo}HV FRPR R WDPDQKR
��

GR Qy SRGHP LQGLFDU D TXDQWLGDGH GH JHQHV TXH VmR LQIOXHQFLDGRV SRU GHWHUPLQDGR
IDWRU�GH�WUDQVFULomR�
$V RQWRORJLDV VmR FRQMXQWRV GH JHQHV UHODFLRQDGRV D XP GHWHUPLQDGR WHUPR
ELROyJLFR� FRPR H[HPSOR WHPRV� UHJXODomR GD GLIHUHQFLDomR GH QHXU{QLRV
GRSDPLQpUJLFRV �*2��������� FRP DSUR[LPDGDPHQWH �� JHQHV HQYROYLGRV� RX
GRHQoD GH DO]KHLPHU �KVD������ FRP GH]HQDV GH JHQHV HQYROYLGRV� 2QWRORJLDV VmR
DOWDPHQWH GHSHQGHQWHV GH XPD H[WHQVLYD H FRQVWDQWH FXUDGRULD� VHQGR DOJXQV
FDVRV FRQKHFLGRV GH EDQFRV GH GDGRV FRP RQWRORJLDV� .(** �GR LQJOrV� .\RWR
(QF\FORSHGLD�RI�*HQHV�DQG�*HQRPHV��H�*2��GR�LQJOrV� JHQH�RQWRORJ\��
)HUUDPHQWDV FRPR JSURILOHU� �.ROEHUJ HW DO� ����� H (QULFK5 �&KHQ HW DO�
����� IDFLOLWDUDP R XVR GHVVHV EDQFRV QD PHGLGD HP TXH HOHV XQLILFDP GLYHUVRV
EDQFRV GH GDGRV GH RQWRORJLD H DSUHVHQWDP UHVXOWDGRV GH WRGDV DV RQWRORJLDV
HQFRQWUDGDV�HP�XPD�~QLFD�DQiOLVH�
4XDQGR WHPRV XPD DVVLQDWXUD �JHQHV UHVXOWDQWHV GD DQiOLVH GLIHUHQFLDO�
SRGHPRV SHUJXQWDU� DR XQLYHUVR GH JHQHV GDV RQWRORJLDV� TXDLV WHUPRV HVWDULDP
HQULTXHFLGRV SDUD XPD DVVLQDWXUD TXDOTXHU� $V RQWRORJLDV HQULTXHFLGDV LQGLFDP
TXDLV HYHQWRV ELROyJLFRV HVWmR VHQGR LQIOXHQFLDGRV SHOR PHX JUXSR GH JHQHV� 2XWUR
GHWDOKH LPSRUWDQWH p TXH D H[SUHVVmR GRV JHQHV SRGH VHU OHYDGD HP FRQVLGHUDomR
SDUD�KDYHU�RXWURV�GHWDOKHV�FRPR�DXPHQWR�RX�GLPLQXLomR QD�UHJXODomR�GH�XP�WHUPR�
&DEH QRWDU TXH DQiOLVHV HQYROYHQGR WHUPRV SRVVXHP FHUWDV OLPLWDo}HV H
ELDV FRPR R DXPHQWR GHVSURSRUFLRQDO GH WHUPRV HQYROYHQGR WHPDV UHFRUUHQWHV
�FRPR FkQFHU GH PDPD� H JHQHV PDLV FLWDGRV �FRPR %5&$��%5&$��� R TXH
��

SRGHULD MXVWLILFDU XP DXPHQWR GH WHUPRV IDOVR QHJDWLYRV HP UHVXOWDGRV �+D\QHV HW
DO��������
1HVWH WUHFKR� IRL YLVWR TXH D H[SUHVVmR GLIHUHQFLDO QRV Gi DVVLQDWXUDV FRP
RV JHQHV LPSRUWDQWHV HP XP GHWHUPLQDGR HYHQWR H RXWUDV WpFQLFDV FRPR UHGHV GH
LQWHUDomR H RQWRORJLDV DXPHQWDP QRVVD FRPSUHHQVmR DFHUFD GHVVH JUXSR GH
JHQHV� 1D SUy[LPD SDUWH� YDPRV HQWHQGHU XPD WpFQLFD TXH DXPHQWRX D UHVROXomR
GRV DFKDGRV DQWHV HQFRQWUDGRV VRPHQWH HP WHFLGR� R VHTXHQFLDPHQWR GH
FpOXOD�~QLFD�
��

6HTXHQFLDPHQWR�GH�FpOXOD�~QLFD��XPD�QRYD�IURQWHLUD
3URYD�GH�FRQFHLWR�H�SULPHLURV�SDVVRV�GR�VHTXHQFLDPHQWR GH�FpOXOD�~QLFD
2 VHTXHQFLDPHQWR GR 51$ GH FpOXOD�~QLFD �GR LQJOrV� VLQJOH�FHOO 51$
VHTXHQFLQJ� VXUJH HP ���� FRP 7DQJ H FRODERUDGRUHV �)LJXUD ��� -i QHVVH SULPHLUR
H[SHULPHQWR� IRL HQFRQWUDGR XP DXPHQWR GH H[SUHVVLYR Q~PHUR GH JHQHV
LGHQWLILFDGRV �a���� VH FRPSDUDGR FRP PLFURDUUD\ �.XULPRWR HW DO� ����� HP
EODVW{PHURV� $OpP GLVVR� P~OWLSORV WUDQVFULWRV SRU JHQHV IRUDP LGHQWLILFDGRV� H
FRQVHTXHQWHPHQWH� GLYHUVRV VtWLRV GH VSOLFLQJ �7DQJ HW DO� ������ $QRV PDLV WDUGH�
DWp PHVPR QXP HVWiJLR HPEULRQiULR GH EODVW{PHUR FRP DSHQDV � FpOXODV
SOXULSRWHQWHV� GLIHUHQoDV FRQVLVWHQWHV QD H[SUHVVmR FHOXODU VHULDP HQFRQWUDGDV
�*RRODP�HW�DO��������
2XWUDV PHWRGRORJLDV FRPR &(/VHT �+DVKLPVKRQ\ HW DO� ����� H VPDUW6HT
�5DPVNROG HW DO� ����� WDPEpP GHPRQVWUDUDP D YLDELOLGDGH GR VHTXHQFLDPHQWR GH
DOJXPDV GH]HQDV GH FpOXODV SRU H[SHULPHQWR �)LJXUD ��� $LQGD QHVVH PRPHQWR� D
DQiOLVH GR VHTXHQFLDPHQWR GH FpOXOD�~QLFD HUD WUDWDGD FRPR XPD DQiOLVH GH
51$VHT VHPHOKDQWH D DQiOLVH GH WHFLGR� 6HQGR DVVLP� D H[SUHVVmR GLIHUHQFLDO
DLQGD HUD SRXFR YLiYHO H R IRFR GDV DQiOLVHV HVWDYD QD GHVFULomR GD YDULDELOLGDGH GH
WUDQVFULWRV�H�VtWLRV�GH�VSOLFLQJ�
��

)LJXUD �� (VWXGRV FRP VHTXHQFLDPHQWR GH FpOXOD�~QLFD� 'LIHUHQWHV WHFQRORJLDVH VHXV DYDQoRV DR ORQJR GR WHPSR PRVWUDP XP DXPHQWR H[SRQHQFLDO QR Q~PHUR GHFpOXODV VHTXHQFLDGDV SRU HVWXGR HP PHQRV GH XPD GpFDGD� 1D SDUWH VXSHULRU GDILJXUD HVWmR UHSUHVHQWDGRV RV SULQFLSDLV SURWRFRORV H VXDV UHVSHFWLYDV SXEOLFDo}HV�1D SDUWH LQIHULRU H[LVWH D UHODomR HQWUH RV DQRV GH SXEOLFDomR H D TXDQWLGDGH GHFpOXODV�VHTXHQFLDGDV�HP�FDGD�HVWXGR��$GDSWDGR�GH�6YHQVVRQ HW�DO�������
��

3ULPHLURV�HVWXGRV�SRSXODFLRQDLV
(P SRXFR WHPSR� QRYDV WpFQLFDV FRPR VPDUW6HT� �3LFHOOL HW DO� ����� H
0$56�VHT DXPHQWDUDP D VHQVLELOLGDGH �a����� JHQHV GHWHFWDGRV� H D TXDQWLGDGH
GH FpOXODV SDUD DOJXPDV FHQWHQDV D PLOKDUHV �)LJXUD ��� 1HVVH PRPHQWR� DOJXQV
HVWXGRV Mi EXVFDYDP UHSUHVHQWDU D GLYHUVLGDGH FHOXODU FRP FLWRPHWULD GH IOX[R H
51$VHT QR VLVWHPD KHPDWRSRpWLFR DGXOWR �0F.LQQH\�)UHHPDQ HW DO� ������ QR
FpUHEUR GH FDPXQGRQJR H KXPDQR DGXOWR �=KDQJ HW DO� ����� =KDQJ HW DO� ������
&RQWXGR� HP WRGRV HVVHV FDVRV� DV GLIHUHQoDV LQWUtQVHFDV HQWUH SRSXODo}HV
FHOXODUHV IRUDP LPHQVDPHQWH H[SDQGLGDV FRP RV SULPHLURV H[SHULPHQWRV
SRSXODFLRQDLV�GH�VF51$VHT�
&pOXODV�HP�JRWDV�H�D�H[SDQVmR�GDV�IURQWHLUDV
(P ����� 0DFRVNR H FRODERUDGRUHV DXPHQWDUDP H[SRQHQFLDOPHQWH R
Q~PHUR GH FpOXODV VHTXHQFLDGDV FRP D FULDomR GR VHTXHQFLDPHQWR XVDQGR
QDQRJRWDV �GR LQJOrV� GURSOHW�EDVHG�� RQGH FpOXODV VHULDP FRORFDGDV HP SHTXHQRV
JRWDV�GH�yOHR��)LJXUD����
(P VHX H[SHULPHQWR� 0DFRVNR UHYHORX D H[LVWrQFLD GH a���� JHQHV
H[SUHVVRV QD UHWLQD GH FDPXQGRQJR� PDLV D H[LVWrQFLD GH �� VXESRSXODo}HV
SXWDWLYDV� $Wp DTXHOH PRPHQWR� R VHTXHQFLDPHQWR VH OLPLWDYD D SRXFRV PLOKDUHV GH
FpOXODV� KDYHQGR XPD H[SDQVmR SDUD PDLV GH a�� PLO FpOXODV H HP SRXFR DQRV ���
PLOK}HV GH FpOXODV VHULDP VHTXHQFLDGDV ���[ JHQRPLFV� &KURPLXP70 6LQJOH FHOO �¶
Y���VHP�SXEOLFDomR��
��

$LQGD DVVLP� R VHTXHQFLDPHQWR GURSOHW�EDVHG VRIUH DOJXQV SUHMXt]RV� FRPR
SRU H[HPSOR� FRQWDPLQDomR FUX]DGD GH EDUFRGHV H �¶�ELDV QDV UHDGV� $SHVDU GH
H[LVWLU XPD FRQWDPLQDomR FUX]DGD HVSHUDGD QR XVR GH GURSOHW�EDVHG HQWUH
EDUFRUGHV GH FpOXODV GLIHUHQWHV �HVWLPDGR HQWUH ���� GDV UHDGV�� HVVH GDGR SRGHUi
VHU GHWHFWDGR H UHPRYLGR FRP R XVR GH 80, �*ULIILWKV HW DO� ������ 1R HQWDQWR� R
�¶�ELDV p XPD OLPLWDomR GR VHTXHQFLDPHQWR GURSOHW�EDVHG� DLQGD VHP VROXomR� R TXH
LPSHGH D GHVFULomR GH QRYRV WUDQVFULWRV H WRUQD SUDWLFDPHQWH LPSRVVtYHO D
LGHQWLILFDomR�GH�HYHQWRV�GH�VSOLFLQJ�QHVVH�PRGHOR�
$QiOLVHV�GH�VF51$VHT
(P SDUDOHOR� GLYHUVRV SLSHOLQHV VXUJLUDP FRP R DXPHQWR GR Q~PHUR GH
FpOXODV� H[SORUDQGR QRYDV SHUVSHFWLYDV GD GLYHUVLGDGH FHOXODU� FRPR� LGHQWLILFDomR
GH WLSRV FHOXODUHV FRP DOJRULWPRV GH DJUXSDPHQWR H H[SUHVVmR GLIHUHQFLDO�
FRQVWUXomR GH PDSDV SVHXGR WHPSRUDLV H UHFRQVWUXomR HVSDFLDO GD GLYHUVLGDGH
FHOXODU��FULDomR�GH�UHGHV�UHJXODWyULDV�GH�JHQHV�
'LYHUVDV ELEOLRWHFDV FRPR 6HXUDW Y� �%XWOHU HW DO� ������ 0RQRFOH� �4LX HW DO�
����� H VFDQS\ �:ROI HW DO� ����� SHUPLWLUDP R DJUXSDPHQWR GH FpOXODV HP JUXSRV
SXWDWLYRV� JHUDOPHQWH WDO DQiOLVH SDVVD SRU � SDVVRV� VHOHomR GH JHQHV DOWDPHQWH
YDULiYHLV� UHGXomR OLQHDU GH GLPHQVLRQDOLGDGH FRP R XVR GH FRPSRQHQWHV SULQFLSDLV
H D GLYLVmR GDV FpOXODV HP JUXSRV FRP R XVR GH DOJRULWPRV FRP EDVH HP JUDIRV
FRPR /RXYDLQ� .11 �GR LQJOrV� .�QHDUHVW QHLJKERU�� 7DO PHWRGRORJLD IRL FRQFHELGD
SRU /HYLQH H FRODERUDGRUHV QR GHVHQYROYLPHQWR GR DOJRULWPR 3KHQR*UDSK �/HYLQH
HW DO� ������ RQGH Ki D UHGXomR GH PDWUL]HV GH DOWD GLPHQVLRQDOLGDGH �OHLD�VH�
��

PDWUL]HV FRP JUDQGH Q~PHUR GH OLQKDV GH JHQHV YV� FROXQDV GH FpOXODV� SDUD
SHTXHQRV�JUXSRV�GH�FpOXODV�
'HSRLV TXH RV JUXSRV SXWDWLYRV VmR HQFRQWUDGRV� p UHDOL]DGD XPD DQiOLVH GH
H[SUHVVmR GLIHUHQFLDO SDUD LGHQWLILFDU DVVLQDWXUDV GH WDLV JUXSRV� &RP DV
DVVLQDWXUDV� RV JUXSRV VmR PDQXDOPHQWH FODVVLILFDGRV SHOR FRQKHFLPHQWR SUpYLR
TXH WHPRV GR PRGHOR� &DEH VDOLHQWDU TXH R DJUXSDPHQWR p XP DOJRULWPR
QmR�VXSHUYLVLRQDGR� RX VHMD� RV JUXSRV VmR HQFRQWUDGRV VHP LQIOXrQFLD GR
SHVTXLVDGRU� FRQWXGR� D FODVVLILFDomR GRV JUXSRV p XPD HVWUDWpJLD WRWDOPHQWH
VXSHUYLVLRQDGD�
2XWUD DERUGDJHP EDVWDQWH FRPXP p D FULDomR GH PDSDV SVHXGR WHPSRUDLV
TXH VmR EDVLFDPHQWH D RUGHQDomR GDV FpOXODV QXPD VpULH�JUDGLHQWH GH H[SUHVVmR
JrQLFD� 2X VHMD� R DOJRULWPR VDL GR SUHVVXSRVWR TXH H[LVWH XP ³FDPLQKR´ WHPSRUDO
HP TXH DV FpOXODV SRGHP VHU HQFDL[DGDV� 2 DOJRULWPR PDLV FRQKHFLGR p R
PRQRFOH� �4LX HW DO� ����� TXH� VHPHOKDQWH DRV DOJRULWPRV GH DJUXSDPHQWR� XVD
XP PpWRGR GH JUDIR FRPR 5*( �GR LQJOrV� UHYHUVHG JUDSK HPEHGGLQJ� SDUD FULDU
XP FDPLQKR EDVHDGR QD YDULHGDGH� H HP VHJXLGD� FRORFDU DV FpOXODV QHVVH PHVPR
FDPLQKR� 9DOH GL]HU TXH TXDOTXHU PDSD GH GLIXVmR VHJXH D PHVPD LGpLD GRV PDSDV
SVHXGR WHPSRUDLV� FRPR SRU H[HPSOR� R TXH p IHLWR SHOR DOJRULWPR GHVWLQ\ GH
$QJHUHU�H�FRODERUDGRUHV��$QJHUHU�HW�DO��������
e LPSRUWDQWH FLWDU TXH RV PDSDV SVHXGRWHPSRUDLV VmR XP DOJRULWPR
VHPL�VXSHUYLVLRQDGR� R TXH LQGLFD TXH R FRQKHFLPHQWR SUpYLR GR PRGHOR H GRV
GDGRV�VmR�HVVHQFLDLV�SDUD�VH�REWHU�DOJXP�DFKDGR�UHOHYDQWH�
��

7DPEpP WHPRV D UHFRQVWUXomR HVSDFLDO TXH VmR PpWRGRV EDVWDQWHV
SDUHFLGRV FRP RV PDSDV SVHXGR WHPSRUDLV� RQGH Ki D WHQWDWLYD GH SRVLFLRQDU DV
FpOXODV QXP SODQR DQDW{PLFR� 6DWLMD H FRODERUDGRUHV Mi KDYLDP WHQWDGR UHSUHVHQWDU
DV FpOXODV QXP FRQWH[WR DQDW{PLFR �6DWLMD HW DO� ����� QD SULPHLUD YHUVmR GR 6HXUDW�
FRQWXGR�� 7DO PHWRGRORJLD H[LJH XP FRQKHFLPHQWR SUpYLR GDV UHJL}HV HVWXGDGDV� SRU
H[HPSOR� GDGRV GH KLEULGL]DomR LQ VLWX� PLFURGLVVHFomR� GHQWUH RXWURV TXH DSRQWDP
SLVWDV DQDW{PLFDV TXH VHUmR FRUUHODFLRQDGDV FRP R GDGR GH H[SUHVVmR� +RMH� D
UHFRQVWUXomR HVSDFLDO WDPEpP SRGHUi VHU D QtYHO LQWUDFHOXODU� FRPR Mi p YLVWR QD
WpFQLFD�6HT),6+��6KDK�HW�DO��������
$ FRQVWUXomR GH UHGHV UHJXODWyULDV WUDQVFULFLRQDLV �571� GR LQJOrV� UHJXODWRU\
WUDQVFULSWLRQDO QHWZRUNV� VmR XPD IRUPD HILFLHQWH GH SUHGL]HU D GLQkPLFD GH GLYHUVRV
JHQHV HP GLIHUHQWHV WLSRV FHOXODUHV� $OJRULWPRV FRPR 6&(1,& �$LEDU HW DO� ����� RX
6HXUDW Y� �6WXDUW HW DO� ����� Mi IRUDP GHVHQYROYLGRV H[FOXVLYDPHQWH SDUD D
FRQVWUXomR GHVVDV UHGHV HP GDGRV GH VF51$VHT� $OpP GLVVR� DOJXQV DOJRULWPRV
SDUD DQiOLVH GH WHFLGR FRPR :*&1$ �/DQJIHOGHU HW DO� ����� Mi IRUDP XVDGRV GH
PDQHLUD EHP VXFHGLGD SDUD VF51$VHT� e LPSRUWDQWH VDOLHQWDU TXH D UHVROXomR GD
H[SUHVVmR JrQLFD IRUQHFLGD SHOR VF51$VHT DXPHQWD H[WHQVLYDPHQWH D TXDOLGDGH
GDV UHGHV UHJXODWyULDV Mi TXH QmR Ki HIHLWRV FRIXQGDGRUHV FRPR D PLVWXUD GH
GLIHUHQWHV�WLSRV�FHOXODUHV�HP�WHFLGRV��.XPDU�HW�DO� ������
5HVXPLGDPHQWH� YLPRV FRPR DFRQWHFHX R VXUJLPHQWR GR VHTXHQFLDPHQWR GH
FpOXOD�~QLFD� DV PHOKRULDV TXH HVVD WpFQLFD QRV IRUQHFHX H DOJXQV FDPLQKRV SDUD D
DQiOLVH GRV GDGRV GH VF51$VHT� 1RV SUy[LPRV GRLV WUHFKRV� VHJXLUi DOJXQV WySLFRV
��

VREUH RV GRLV HVWXGRV GH FDVR TXH IRUDP LQYHVWLJDGRV SRU DQiOLVHV GR WUDQVFULSWRPD
QHVWD�WHVH�
&DVR�GH�HVWXGR�,���$O]KHLPHU�H�VHXV�WUDQVFULWRV
$�GRHQoD�GH�$O]KHLPHU�FRPR�D�SULQFLSDO�FDXVD�GH�GHPrQFLD
$ GRHQoD GH $O]KHLPHU �'$� p XPD QHXURSDWRORJLD TXH DFRPHWH DOJXQV
PLOK}HV GH SHVVRDV QD SRSXODomR KXPDQD� (VVD QHXURSDWRORJLD p XP WLSR GH
GHPrQFLD� WHUPR XVDGR SDUD GHVLJQDU XP JUXSR KHWHURJrQHR GH GRHQoDV
FDUDFWHUL]DGDV SRU GpILFLWV FRJQLWLYRV SURJUHVVLYRV� e UHOHYDQWH QRWDU TXH D '$ p
UHVSRQViYHO SRU ������ GR FDVRV GH GHPrQFLD �.XPDU HW DO� ������ VHQGR TXH
RXWUDV QHXURSDWRORJLDV FRPR GHPrQFLD YDVFXODU� GHPrQFLD IURQWRWHPSRUDO H
GHPrQFLD FRP FRUSRV GH /HZLV SRVVXHP XPD IRUWH VREUHSRVLomR GH VLQWRPDV H
FDXVDV �0DFOLQ HW DO� ������ (P ����� R Q~PHUR GH FDVRV GH GHPrQFLD GHYH FKHJDU
D DSUR[LPDGDPHQWH �� PLOK}HV GH FDVRV� VHQGR TXH D SURMHomR p TXH R DXPHQWR
HP�SDtVHV�FRP�PHQRU�UHQGD�VHMD�PDLV�SURQXQFLDGR��3ULQFH HW�DO��������
0HFDQLVPRV�GD�SDWRORJLD
+RMH� H[LVWHP GRLV WLSRV GH DFKDGRV SDWROyJLFRV SULQFLSDLV SDUD R $O]KHLPHU�
VHQGR HOHV� D IRUPDomR GH SODFDV DPLOyLGHV �13� GR LQJOrV� QHXULWLF SODTXHV� H RV
HPDUDQKDGRV QHXURILEULODUHV �1)7� GR LQJOrV� QHXURILEULOODU\ WDQJOH� �)LJXUD ��� 1R
HQWDQWR� D UHODomR FDXVDO HQWUH HVWHV DFKDGRV H RV GpILFLWV FRJQLWLYRV HP SDFLHQWHV
SHUPDQHFH�XP�WHPD�GH�LQWHQVR�GHEDWH�
��

$ IRUPDomR GDV 13 p FDUDFWHUL]DGR SHOR DF~PXOR LQGHYLGR GRV SHSWtGHRV
$��� H $��� TXH VmR UHVXOWDQWHV GD FOLYDJHP GD SURWHtQD SUHFXUVRUD GH DPLOyLGH
�$33� GR LQJOrV� DP\ORLG SUHFXUVRU SURWHLQ� SHODV HQ]LPDV �� DQG ��VHFUHWDVHV� (VVH
GHVEDODQoR GD IRUPDomR GH SHSWtGHRV DVVRFLDGR D SUREOHPDV GH OLPSH]D GHVVDV
PROpFXODV UHVXOWDP QD IRUPDomR GDV SODFDV DPLOyLGHV �.XPDU HW DO� ������ e
LPSRUWDQWH QRWDU TXH R PHGLFDPHQWR DGXFDQXPDE Mi IRL FDSD] GH UHGX]LU D
TXDQWLGDGH GH SODFDV QR FpUHEUR� VHQGR TXH VXD HILFLrQFLD QD UHGXomR GRV FDVRV GH
$O]KHLPHU�DLQGD�p�FRQWHVWDGD��+RZDUG�HW�DO��������
2XWUR PDUFDGRU SDWROyJLFR FOiVVLFR VmR RV 1)7� TXH VmR WDPEpP XP
GHSyVLWR JHUDGR SHOR DF~PXOR GH SURWHtQD 7DX� &RPR XPD SURWHtQD LQWUDFHOXODU� 7DX
HVWi DVVRFLDGD QD IRUPDomR H HVWDELOLGDGH GH PLFURW~EXORV� p HQFRQWUDGD
SULQFLSDOPHQWH QRV D[{QLRV GRV QHXU{QLRV �5HJDQ HW DO� ����� H p SURGX]LGD SHOR
VSOLFLQJ DOWHUQDWLYR GR JHQH 0$37 �1HYH HW DO� ������ &RPR IRVIRSURWHtQD HOD SRGH
HVWDU IRVIRULODGD RX QmR� VHQGR TXH Ki XPD FRUUHODomR GLUHWD HQWUH D VDWXUDomR GR
VHX�HVWDGR�IRVIRULODGR�FRP�D�SDWRORJLD�$O]KHLPHU��0RFDQX HW�DO��������
��

)LJXUD �� 3ULQFLSDLV PDUFDGRUHV SDWROyJLFRV GD GRHQoD $O]KHLPHU� $ IRUPDomR
1R ODGR HVTXHUGR� D UHSUHVHQWDomR GD IRUPDomR GDV SODFDV DPLOyLGHV H� QR ODGR
GLUHLWR��RV�HPDUDQKDGRV�QHXURILEULODUHV���$GDSWDGR GH�3RVSLFK�HW�DO�������
��

$OpP GLVVR� QDV ~OWLPDV GpFDGDV� KRXYH XP HVIRUoR SDUD GHILQLU PpWULFDV
TXDQWLWDWLYDV H VHPL TXDQWLWDWLYDV TXH DX[LOLHP D FODVVLILFDomR GRV HVWiJLRV GD
GRHQoD GH DO]KHLPHU� (VVDV FODVVLILFDo}HV EXVFDP DYDOLDU WDQWR DV DOWHUDo}HV
FRJQLWLYDV�FRPR�DV�DOWHUDo}HV�ILVLROyJLFDV�GRV�SDFLHQWHV� 6HQGR�HODV�
&ODVVLILFDomR FOtQLFD SDUD GHPrQFLD �&'5� GR LQJOrV� FOLQLFDO GHPHQWLD
UDWLQJ�� )RL FULDGR SRU +XJKHV H FRODERUDGRUHV H DWXDOL]DGR SRU 0RUULV �+XJKHV HW
DO� ����� 0RUULV HW DO� ����� FRP R LQWXLWR GH DYDOLDU FRJQLomR H FRPSRUWDPHQWR�
DVVLP FRPR D DWXDomR GD SHUGD FRJQLWLYD QR GLD�D�GLD GRV SDFLHQWHV� (VVD
FODVVLILFDomR p GLYLGLGD HP � FDWHJRULDV� QHQKXP �� TXHVWLRQiYHO ���� OHYH ��
PRGHUDGR � H VHYHUR �� 6y GHYHUi VHU FRQILJXUDGR FRPR HVWiJLR LQLFLDO GH
GHPrQFLD� &'5 PDLRU RX LJXDO D �� GHYLGR D QHFHVVLGDGH GH DMXGD SDUD DWLYLGDGHV
GLiULDV�
8P FRQVyUFLR SDUD HVWDEHOHFHU XP UHJLVWUR GD GRHQoD GH DO]KHLPHU
�&(5$'� GR LQJOrV� &RQVRUWLXP WR (VWDEOLVK D 5HJLVWU\ IRU $O]KHLPHU¶V 'LVHDVH�
WDPEpP IRL FULDGR HP ���� SHOR 1,$ �GR LQJOrV� 1DWLRQDO ,QVWLWXWH RI $JLQJ� �0LUUD HW
DO� ����� )LOOHQEDXP HW DO� ������ &(5$' p RXWUD PpWULFD VHPLTXDQWLWDWLYD TXH
HVWLPD D GHQVLGDGH GH SODFDV DPLOyLGHV �%HQQHWW HW DO� ����� DVVLP FRPR XPD RXWUD
EDWHULD GH H[DPHV FRPR FOtQLFR� QHXURSVLFROyJLFR� KLVWyULFR IDPLOLDU� GHQWUH RXWURV�
(OH�p�GLYLGLGR�HP��GHILQLWLYR� ����SURYiYHO� ����SRVVtYHO ���H�QmR�DO]KHLPHU� ���
(VWiJLRV GH %UDDN IRUDP FULDGRV %UDDN H %UDDN HP ���� �%UDDN HW DO� �����
%UDDN HW DO� ����� VHQGR XPD PpWULFD VHPL TXDQWLWDWLYD TXH PHQVXUD D GLVWULEXLomR
GRV HPDUDQKDGRV QHXURILEULODUHV QR FpUHEUR� ([LVWHP � HVWiJLRV GH EUDDN� VHQGR
HOHV�
��

� �� �QRUPDO��TXDQGR�QmR�Ki�SUHVHQoD�GH�1)7V�
� ,�,,��LQGLFDQGR�D�SUHVHQoD�GH�1)7V�VRPHQWH�QR�FyUWH[ HQWRUULQDO�
� ,,,�,9� FRP D SUHVHQoD GH 1)7V HP UHJL}HV OtPELFDV �SRU H[HPSOR�
KLSRFDPSR��
� 9�9,��HQYROYLPHQWR�WDPEpP�GH�UHJL}HV�GR�QHRFyUWH[�
&DEH VDOLHQWDU D H[LVWrQFLD GH RXWUDV FODVVLILFDo}HV FRPR 006( �GR LQJOrV�
0LQL�PHQWDO 6WDWH ([DPLQDWLRQ�� *'5 �GR LQJOrV� *HULDWULF 'HSUHVVLRQ 6FDOH�� GHQWUH
RXWURV� TXH QmR IRUDP XWLOL]DGRV QHVWD WHVH RX HVWmR LQFRUSRUDGDV LQGLUHWDPHQWH QDV
PpWULFDV�Mi�FLWDGDV�DFLPD�
)DWRUHV�JHQpWLFRV�GD�SDWRORJLD
$ GRHQoD GH $O]KHLPHU SRGH VHU VXEGLYLGLGD HP GRLV JUDQGHV JUXSRV GH
DFRUGR FRP D VXD KHUGDELOLGDGH� �� IDPLOLDU� �I$'� GR LQJOrV� IDPLOLDO $O]KHLPHUV
GLVHDVH�� H �� HVSRUiGLFR �V$'� GR LQJOrV� VSRUDGLF $O]KHLPHUV GLVHDVH�� 8PD RXWUD
FODVVLILFDomR SRGH VHU IHLWD EDVHDGD QD LGDGH GH LQtFLR GH VLQWRPDV� �� FHGR �(2$'�
GR LQJOrV� HDUO\�RQVHW DO]KHLPHU GLVHDVH� FRP SDFLHQWHV DQWHV GRV �� DQRV� H ��
WDUGLR �/2$'� GR LQJOrV� ODWH�RQVHW DO]KHLPHU GLVHDVH� FRP SDFLHQWHV GHSRLV GRV ��
DQRV�
I$' p XPD GRHQoD DXWRVV{PLFD GRPLQDQWH FDXVDGD JHUDOPHQWH SRU DOJXQV
JHQHV GLUHWDPHQWH HQYROYLGRV FRP D SDWRORJLD FRPR $33� 36(1� H 36(1�� 7DLV
YDULDQWHV VmR H[WUHPDPHQWH UDUDV H FRPS}HP PHQRV TXH �� GRV FDVRV� VHQGR
��

TXH R GHVHQYROYLPHQWR GD SDWRORJLD FRPHoD SRU YROWD GRV �� DQRV GH LGDGH �5\PDQ
HW�DO��������
2 (2$' p LGHQWLILFDGR SHORV FDVRV GH $O]KHLPHU TXH DFRQWHFHP DQWHV GRV
�� DQRV GH LGDGH� 2V JHQHV $33� 36(1� H 36(1� WDPEpP HVWmR HQYROYLGRV FRP
(2$'� FRQWXGR� D VREUHSRVLomR (2$' H )$' VmR QR Pi[LPR ��� GRV FDVRV
�-DUPRORZLF] HW DO� ������ $OpP GLVVR� Mi IRL REVHUYDGR TXH HVVHV FDVRV ID]HP SDUWH
GH�DSUR[LPDGDPHQWH������GRV�FDVRV��0HQGH]�HW�DO��������
3RU ~OWLPR� D JUDQGH PDLRULD GRV FDVRV GH V$' VmR GR WLSR /2$'�
FRUUHVSRQGHQGR D PDLV GH ��� GH WRGRV RV FDVRV GH $'� (VWHV FDVRV WDPEpP
DSUHVHQWDP XP IRUWH FRPSRQHQWH JHQpWLFR� TXH SRGH H[SOLFDU DWp ��� GR ULVFR GH
$' �%HOOHQJXH] HW DO�� ������ 'H IDWR� HVWXGRV GH DVVRFLDomR JHQpWLFD GR WLSR
*:$6 �GR LQJOrV� JHQRPH�ZLGH DVVRFLDWLRQ VWXGLHV� UHYHODUDP LQ~PHUDV YDULDQWHV
DVVRFLDGDV D XP ULVFR DXPHQWDGR GH GHVHQYROYHU $' �9HUJKHVH HW DO� �����
/DPEHUW HW DO�� ����� .DUFK HW DO� ����� -DQVHQ HW DO� ����� .XQNOH HW DO�� �����
%HOOHQJXH] HW DO�� ������ $ PDLRU SDUWH GHVWDV YDULDQWHV IRUDP LGHQWLILFDGDV HP
UHJL}HV QmR�FRGDQWHV GR JHQRPD H SURYDYHOPHQWH DWXDP PRGLILFDQGR RV QtYHLV GH
H[SUHVVmR GH JHQHV DOYR� GHQRPLQDGRV JHQHV GH ULVFR SDUD $' �/DPEHUW HW DO��
����� .XQNOH HW DO�� ������ 1R HQWDQWR� DLQGD SRXFR VH VDEH VREUH RV SURFHVVRV
ELROyJLFRV UHJXODGRV SRU PXLWRV GHVWHV JHQHV RX VH RV VHXV QtYHLV GH H[SUHVVmR
VmR DIHWDGRV QR FXUVR GD SDWRORJLD GD $'� 2 SULPHLUR DUWLJR DSUHVHQWDGR QHVWD WHVH
�0DUTXHV�&RHOKR HW DO�� ����� DSUHVHQWD UHVXOWDGRV TXH YLVDP FRQWULEXLU SDUD R
SUHHQFKLPHQWR�GHVWD�ODFXQD�GR�QRVVR�FRQKHFLPHQWR�
��

'DGRV�VHTXHQFLDGRV�H�VXDV�GHPRJUDILDV
3DUD R QRVVR HVWXGR REWLYHPRV GDGRV GH � FRQVyUFLRV VmR HOHV� 06%%
�0RXQW 6LQDL %UDLQ %DQN� ZDQJ HW DO� ������ 0$<2 �0D\R &OLQLF� $OOHQ HW DO� ����� H
5260$3 �5HOLJLRXV 2UGHUV 6WXG\ �526�� 5XVK 0HPRU\ DQG $JLQJ 3URMHFW �0$3��
'H -DJHU HW DO ������ 7RGRV RV WHFLGRV VHTXHQFLDGRV IRUDP REWLGRV GH FpUHEURV
SRVW�PRUWHP FRP XP LQWHUYDOR SRVW�PRUWHP GH FROHWD �30,� SRVW�PRUWHP LQWHUYDO�
PpGLR HQWUH ��� KRUDV H VHPSUH PHQRU TXH �� KRUDV� $ LGDGH GRV LQGLYtGXRV QR
PRPHQWR GD PRUWH �$2'� DJH RI GHDWK� ILFRX HP PpGLD SUy[LPR DRV �� DQRV GH
LGDGH� 2 YDORU GH LQWHJULGDGH GR 51$ �5,1� 51$ LQWHJULW\ QXPEHU� ILFRX SUy[LPR D ��
$OpP�GLVVR��WRGRV�RV�LQGLYtGXRV�VmR�UHVLGHQWHV�GRV (VWDGRV�8QLGRV�
&DEH GL]HU TXH R GDWDVHW 06%% SRVVXL ��� LQGLYtGXRV VHQGR HOHV
FODVVLILFDGRV FRPR� HXURSHXV ������ DIULFDQRV ����� ODWLQRV ����� DVLiWLFR H UDoD
GHVFRQKHFLGD ���� VHQGR TXH DV DPRVWUDV QR 06%% VmR GH � UHJL}HV GR FpUHEUR
VmR HODV DV iUHDV GH EURDGPDQQ %0��� %0��� %0�� H %0��� 2 GDWDVHW 0$<2
SRVVXL ��� LQGLYtGXRV� WRGRV EUDQFRV FDXFDVLDQRV� VHQGR TXH WRGDV DV DPRVWUDV
VmR GD UHJLmR WHPSRUDO� 2 GDWDVHW 5260$3 SRVVXL ��� LQGLYtGXRV� WRGRV
DXWRGHFODUDGRV EUDQFRV QmR�ODWLQRV� VHQGR WRGDV DV DPRVWUDV GD UHJLmR GR FyUWH[
SUHIURQWDO�GRUVRODWHUDO�
1HVWH WUDEDOKR IRUDP HQFRQWUDGRV ��� '(*V H ��� '78V QD UHJLmR
WHPSRUDO GR FpUHEUR HP GRLV GDWDVHWV GLIHUHQWHV� HQTXDQWR TXH VRPHQWH �� '(*V H
�� '78V IRUDP HQFRQWUDGRV QD UHJLmR IURQWDO GR FpUHEUR HP RXWURV GRLV GDWDVHWV�
&RP HVVHV '(*V H '78V WDPEpP IRL LGHQWLILFDGR XPD JUDQGH TXDQWLGDGH GH
RQWRORJLDV UHODFLRQDGDV D VLQDOL]DomR FHOXODU H PHPEUDQD FHOXODU� $OpP GLVVR� IRL
��

GHVFULWR TXH RV HYHQWRV WrP XPD SUHIHUrQFLD D DFRQWHFHUHP HP HVWiJLR PDLV WDUGLR
GD�GRHQoD�
7DPEpP IRUDP LGHQWLILFDGRV R DXPHQWR GH GLYHUVRV WLSRV GH HYHQWRV GH
VSOLFLQJ �FRPR $766 H $776� DVVLP FRPR SRVVtYHLV FRQVHTXrQFLDV GHVVHV HYHQWRV
QD UHJLmR WHPSRUDO GR FpUHEUR� $OJXQV GRPtQLRV GH SURWHtQD UHOHYDQWHV IRUDP
GHVFULWRV FRP D DMXGD GD DQiOLVH GRV '78V� 3RU ILP� FRP D DMXGD GH VF51$VHT�
IRUDP HOXFLGDGRV JHQHV H WLSRV FHOXODUHV TXH SRGHULDP HVWDU VHQGR DIHWDGRV
GXUDQWH�D�GRHQoD�$O]KHLPHU�
&DVR GH HVWXGR ,, � 1HXURELRORJLD GR GHVHQYROYLPHQWR H VHXV
WUDQVFULWRV
$ IRUPDomR GR QHRFyUWH[ FHUHEUDO GH PDPtIHURV p XP HYHQWR FRPSOH[R H
DOWDPHQWH VLQFURQL]DGR� 1HOH� FDPDGDV JHUPLQDWLYDV FULDP HP RQGDV GH
SUROLIHUDomR� PLJUDomR H GLIHUHQFLDomR� WRGD D GLYHUVLGDGH GH FpOXODV QHXURQDLV H
PDFURJOLDLV�SUHVHQWHV�QR�FyUWH[�FHUHEUDO�
��

6XUJLPHQWR�GR�QHRFyUWH[�FHUHEUDO
'XUDQWH D IRUPDomR GR FyUWH[ FHUHEUDO� QHXU{QLRV GH SURMHomR H[FLWDWyULRV
LUmR SRSXODU GLIHUHQWHV FDPDGDV GR FyUWH[ FHUHEUDO �)LJXUD ��� 2 VXUJLPHQWR GHVVHV
QHXU{QLRV H[FLWDWyULRV SDUWH GDV FpOXODV GD JOLD UDGLDO �5*� GR LQJOrV� UDGLDO JOLD� QD
]RQD YHQWULFXODU �9=� GR LQJOrV� YHQWULFXODU ]RQH�� $OpP GLVVR� D IRUPDomR GR FyUWH[
FHUHEUDO VHUi DX[LOLDGR SRU FpOXODV SURJHQLWRUDV LQWHUPHGLiULDV �,3&� GR LQJOrV�
LQWHUPHGLDWH SURJHQLWRU FHOO� SUHVHQWHV QD FDPDGD JHUPLQDWLYD VHFXQGiULD� ]RQD
VXEYHQWULFXODU �69=� GR LQJOrV� VXEYHQWULFXODU ]RQH�� �0RO\QHDX[ HW DO� ����� *UHLJ
HW DO� ������ 9DOH VDOLHQWDU TXH RV LQWHUQHXU{QLRV FRUWLFDLV TXH WDPEpP ID]HP SDUWH
GR�FyUWH[�FHUHEUDO�QmR�IRUDP�FLWDGRV�SRLV�QmR�VmR R�IRFR�GR�QRVVR�WUDEDOKR�
$ IRUPDomR GDV FDPDGDV PDLV SURIXQGDV DFRQWHFH HQWUH (�����(����
VHQGR TXH DV FDPDGDV PDLV VXSHUILFLDLV RFRUUHP HQWUH (�����(���� �)LJXUD ��� $
GLVWLQomR HQWUH HVVDV VXESRSXODo}HV GH QHXU{QLRV GH SURMHomR VH Gi D SDUWLU GDV
FpOXODV SURJHQLWRUDV GD JOLD UDGLDO �5*�� 6HQGR HVWDV� FDSD]HV GH DXPHQWDU D
TXDQWLGDGH GH FpOXODV IRUPDGDV QR QHRFyUWH[ SRU PHLR GH VXFHVVLYDV GLYLV}HV
VLPpWULFDV�QD�UHJLmR�EDVDO�GD�]RQD�YHQWULFXODU��+DXEHQVDN HW�DO��������
$LQGD DVVLP� D GLYHUVLGDGH GH SURJHQLWRUHV QHRFRUWLFDLV SHUPDQHFH SRXFR
HQWHQGLGD� GLYLGLGD HP GXDV KLSyWHVHV SULQFLSDLV� DV FpOXODV GD JOLD UDGLDO VmR XPD
SRSXODomR KRPRJrQHD TXH DR ORQJR GR GHVHQYROYLPHQWR PXGDP VXD HVSHFLILFLGDGH
SDUD IRUPDomR GH GLIHUHQWHV FDPDGDV RX H[LVWH XPD GLVWLQomR GHVVDV FpOXODV Mi HP
SHUtRGRV HPEULRQiULRV QR LQtFLR GR GHVHQYROYLPHQWR �)UDQFR HW DO�� ����� *DR HW DO��
������*LO�6DQ]�HW�DO���������
��

)LJXUD �� )RUPDomR GRV QHXU{QLRV GH SURMHomR QR FyUWH[ GH FDPXQGRQJR� $JOLD UDGLDO �5*�� SUHYLDPHQWH QHXURHSLWpOLR �1(�� QD ]RQD YHQWULFXODU �9=� FRPHoD DSURGX]LU QHXU{QLRV HP (����� &RQFRPLWDQWH� 5* FRPHoD D IRUPDU SURJHQLWRUHVLQWHUPHGLiULRV �,3V� H D 5* H[WHUQD �R5*�� HVWDEHOHFHQGR DVVLP D ]RQDVXEYHQWULFXODU �69=�� &pOXODV &DMDO�5HW]LXV �&5� PLJUDP SDUD FDPDGD , GH UHJL}HVQmR�FRUWLFDLV� DX[LOLDQGR QD IRUPDomR GDV FDPDGDV SRVWHULRUHV� (QTXDQWR LVVR�WRGRV RV QHXU{QLRV GH SURMHomR IRUDP IRUPDGRV QDV UHJL}HV QHRFRUWLFDLV 9= RX 69=H PLJUDUDP SDUD VXDV UHVSHFWLYDV FDPDGDV� 'HSRLV GLVVR� GLIHUHQWHV SLFRV GHIRUPDomR IRUPDUDP QHXU{QLRV GH SURMHomR� VXESODFD �631�� FRUWLFRWDOkPLFR�&7K31�� VXEFHUHEHODUHV �6&31�� JUDQXODUHV �*1� H FDORVDLV �&31�� $GDSWDGR GH*UHLJ�HW�DO�������
��

1HVWH WUDEDOKR IRL GHVFULWR D H[SUHVVmR JrQLFD GH VXESRSXODo}HV GH
SURJHQLWRUHV GXUDQWH D IRUPDomR GR FyUWH[ HP FDPXQGRQJRV� &RP D DQiOLVH GD
H[SUHVVmR GLIHUHQFLDO H UHJXODGRUHV WUDQVFULFLRQDLV� R IDWRU GH WUDQVFULomR 6R[� IRL
VHOHFLRQDGR FRPR ERP FDQGLGDWR SDUD D VHSDUDomR HQWUH SURJHQLWRUHV VXSUD H
LQIUD�JUDQXODUHV� $OpP GLVVR� IRL GHPRQVWUDGR TXH 6R[� SRGH SURORQJDU R SHUtRGR GH
ODWrQFLD�GRV�SURJHQLWRUHV�H�FDXVDU�XP�DXPHQWR�GDV FDPDGDV�VXSUD�JUDQXODUHV�
��

2EMHWLYRV
2EMHWLYR�JHUDO
(VWD WHVH YLVD GHPRQVWUDU GLIHUHQWHV PDQHLUDV H IRUPDV GH HQFRQWUDUPRV
PDUFDGRUHV SDUD HYHQWRV ELROyJLFRV H�RX SDWRORJLDV FRP R XVR GH GDGRV GH
51$VHT�H�VF51$VHT�
2EMHWLYRV�HVSHFtILFRV
�� (QFRQWUDU DV DOWHUDo}HV GD H[SUHVVmR GH JHQHV H LVRIRUPDV HP
GLIHUHQWHV iUHDV GR FpUHEUR GH SDFLHQWHV FRP $O]KHLPHU XWLOL]DQGR GDGRV GH
51$VHT�
�� ,GHQWLILFDU HYHQWRV ELROyJLFRV SRWHQFLDOPHQWH DIHWDGRV QD SDWRORJLD GD
GRHQoD GH $O]KHLPHU DWUDYpV GD DQiOLVH GH HQULTXHFLPHQWR GH JUXSRV GH
JHQHV�RQWRORJLDV�
�� 0DSHDU DOWHUDo}HV GD H[SUHVVmR JrQLFD SDUD WLSRV FHOXODUHV HVSHFtILFRV
GR FpUHEUR KXPDQR DWUDYpV GD DQiOLVH GH VF51$VHT H FRUUHODFLRQDU FRP RV
JHQHV�HQFRQWUDGRV�QD�H[SUHVVmR�GLIHUHQFLDO�GH�JHQHV H�LVRIRUPDV�
�� 8WLOL]DU VF51$VHT SDUD LGHQWLILFDU JHQHV GLIHUHQFLDOPHQWH H[SUHVVRV HP
FpOXODV SURJHQLWRUDV QHXUDLV +HV��SRVLWLYDV HP FRPSDUDomR FRP DV
+HV��QHJDWLYDV�QR�FyUWH[�FHUHEUDO�GH�FDPXQGRQJRV�HP GHVHQYROYLPHQWR�
�� 8WLOL]DU D DERUGDJHP GH UHGHV UHJXODWyULDV WUDQVFULFLRQDLV SDUD
LGHQWLILFDU�UHJXODGRUHV�GD�H[SUHVVmR�JrQLFD�HP�SURJHQLWRUHV QHXUDLV�+HV���
��

&DStWXOR�,
'LIIHUHQWLDO�WUDQVFULSW�XVDJH�XQUDYHOV�JHQH�H[SUHVVLRQ DOWHUDWLRQV�LQ$O]KHLPHU¶V�GLVHDVH�KXPDQ�EUDLQV
$XWRUHV� 'LHJR�0DUTXHV�&RHOKR� /XNDV�,RKDQ�GD�&UX] &DUYDOKR��$QD�5DTXHO�0HORGH�)DULDV��1HXUR&(%�%UDLQ�%DQN��-HDQ�&KDUOHV�/DPEHUW� 0DUFRV�5RPXDOGR�&RVWD
$UWLJR�SXEOLFDGR�QR�SHULyGLFR QSM�$JLQJ�DQG�0HFKDQLVPV RI�'LVHDVH
GRL� LXXTW���HSM�SVK���������W�����������������

ARTICLE OPEN
Differential transcript usage unravels gene expression
alterations in Alzheimer’s disease human brainsDiego Marques-Coelho 1,2, Lukas da Cruz Carvalho Iohan1,2, Ana Raquel Melo de Farias1,3, Amandine Flaig3 and The Brainbank
Neuro–CEB Neuropathology Network*, Jean-Charles Lambert 3 and Marcos Romualdo Costa 1,3✉
Alzheimer’s disease (AD) is the leading cause of dementia in aging individuals. Yet, the pathophysiological processes involved in AD
onset and progression are still poorly understood. Among numerous strategies, a comprehensive overview of gene expression
alterations in the diseased brain could contribute for a better understanding of the AD pathology. In this work, we probed the
differential expression of genes in different brain regions of healthy and AD adult subjects using data from three large
transcriptomic studies: Mayo Clinic, Mount Sinai Brain Bank (MSBB), and ROSMAP. Using a combination of differential expression of
gene and isoform switch analyses, we provide a detailed landscape of gene expression alterations in the temporal and frontal lobes,
harboring brain areas affected at early and late stages of the AD pathology, respectively. Next, we took advantage of an indirect
approach to assign the complex gene expression changes revealed in bulk RNAseq to individual cell types/subtypes of the adult
brain. This strategy allowed us to identify previously overlooked gene expression changes in the brain of AD patients. Among these
alterations, we show isoform switches in the AD causal gene amyloid-beta precursor protein (APP) and the risk gene bridging
integrator 1 (BIN1), which could have important functional consequences in neuronal cells. Altogether, our work proposes a novel
integrative strategy to analyze RNAseq data in AD and other neurodegenerative diseases based on both gene/transcript expression
and regional/cell-type specificities.
npj Aging and Mechanisms of Disease (2021) 7:2 ; https://doi.org/10.1038/s41514-020-00052-5
INTRODUCTION
Changes in gene expression characterize a multitude of humandiseases and have been successfully used to predict molecularand cellular mechanisms associated with pathological processes1.Alzheimer’s disease (AD) is the most prevalent type of dementiaand causes a progressive cognitive decline, for which there is noeffective treatment or cure. Although expression analyses in braindiseases are generally limited by tissue availability, RNA sequen-cing (RNAseq) data have been generated from postmortem brainsamples of healthy and AD individuals2–4. However, a compre-hensive description of the gene expression alterations in the ADbrain remains elusive.Recent work has begun to address this important gap in the
study of AD pathology using bulk brain tissue RNA sequencing(RNAseq)5 or single-cell RNA sequencing (scRNAseq)6,7. However,these studies have focused on samples obtained from differentbrain regions, namely the dorsolateral prefrontal5,7 and entorhinalcortices6, which could lead to important discrepancies in theresults. In fact, AD pathology shows a progressive impact ondifferent brain regions, characterized at early stages by thepresence of TAU protein inclusions in the locus coeruleus, thetransentorhinal and entorhinal regions (stages I and II). This isfollowed by the presence of TAU inclusions in the hippocampalformation and some parts of the neocortex (stages III and IV),followed by large parts of the neocortex (stages V and VI)8. Thistemporal progression of AD pathology could differently impactgene expression in those brain areas. Accordingly, a recent studyhas shown that changes in protein expression are much moreprominent in areas affected at early and intermediate stages, suchas the hippocampus, entorhinal cortex, and cingulate cortex in the
temporal lobe, compared to other brain regions affected at laterstages of AD pathology, such as sensory cortex, motor cortex, andcerebellum9.Another important aspect to consider is the descriptive
relevance of gene expression analysis based solely on theidentification of differentially expressed genes (DEG), whichfails to detect dynamics in the expression of multiple relatedtranscripts10. Recently, new approaches using transcripts-levelanalysis, so-called differential transcript usage (DTU), enablesidentification of alternative splicing and isoform switches with theprediction of functional consequences11,12. Therefore, importantgene expression modifications in the AD brain could occur at thetranscript level and be overlooked in classical DEG analyses.Here, we took advantage of three available RNAseq datasets,
generated using samples from different brain regions, to system-atically probe gene expression changes (DEG and DTU) in AD. InMayo’s clinic study, both the temporal cortex and cerebellum wereused to obtain bulk RNAseq2. In the Religious Orders Study (ROS)and Memory and Aging Project (MAP), henceforth called ROSMAP,the dorsolateral prefrontal cortex was used3. Finally, in the MountSinai/JJ Peters VA Medical Center Brain Bank (MSBB), four differentBrodmann areas of the brain were studied: areas 22 and 36 fromthe temporal lobe, areas 10 and 44 in the frontal lobe4. We alsoadded another level of complexity using an indirect approach toassign DEGs and gDTUs to unique cell types in order to identifyAD gene expression signatures for neural cells, microglia, andendothelial cells. Finally, we linked these alterations with ADcausal and risk genes, identifying novel isoform switches inBIN1 and APP genes of potential functional consequences forpathology progression.
1Brain Institute, Federal University of Rio Grande do Norte, Av. Nascimento de Castro, 2155 Natal, Brazil. 2Bioinformatics Multidisciplinary Environment (BioME), Federal University
of Rio Grande do Norte, Natal, Brazil. 3Unité INSERM 1167, RID-AGE-Risk Factors and Molecular Determinants of Aging-Related Diseases, Institut Pasteur de Lille, University of Lille,
Lille Cedex, France. *A list of authors and their affiliations appears at the end of the paper. ✉email: [email protected]
www.nature.com/npjamd
Published in partnership with the Japanese Society of Anti-Aging Medicine
1234567890():,;

RESULTS
Regional gene expression alterations in the AD brain correlateswith pathological progression
Several consortia have generated RNAseq data from brains ofindividuals with a clinical and/or pathological diagnostic of AD2–4.Considering the regional progression of AD pathology8, we set outto identify and compare differentially expressed genes (DEG) inthe temporal lobe (TL), encompassing brain regions affect at earlystages of the AD such as the hippocampus and entorhinal cortex,and in the frontal lobe (FL), affect at more advanced stages ofthe pathology (Fig. 1). Comparisons between control and ADindividuals were performed independently for each dataset andonly genes with fold change >1.3 and FDR > 0.01 were consideredas DEGs. We found 3348 (1244 down- and 2104 upregulatedgenes) and 2172 (1170 down and 999 upregulated genes in BM22and BM36; three genes regulated in opposite directions in thesetwo areas) DEGs in the TL of AD individuals compared to theirrespective controls in the MSBB_TL and Mayo datasets, respec-tively (Fig. 2A, B and Supplementary Table 1). Of those DEGs, 734genes (145 down and 520 up) were commonly regulated in bothMayo and MSBB_TL (88.4% of genes altered in the same direction;15.33% of overlap; P= 8.56 × 10−59, hypergeometric test). Incontrast, only 327 (113 down and 214 up) and 209 (97 downand 112 up) DEGs were detected in the MSBB_FL and ROSMAP,respectively. Of those, 31 genes (18 down and 13 up) were foundin both datasets (7.34% of overlap; P= 1.67 × 10−14, hypergeo-metric test) (Fig. 2A, B and Supplementary Table 1). This smallnumber of DEGs in the FL is in agreement with previous dataobtained from the DLPFC (106 down- and 158 upregulated geneswith FC > 1.3)13. Among DEGs detected in the FL, 62.5% were alsodetected in the TL (Fig. 2B), suggesting that similar molecularchanges occur in these brain areas, but at different stages of thedisease progression. The differences in the number of DEGs
detected in the FL and TL can neither be attributed to lack ofstatistical power nor potential biases due to tissue processing,since the number of samples in the FL is larger than in the TLgroups (Fig. 1) and differences are observed even in samplesobtained from the same donors (compare MSBB_TL and MSBB_FLin Fig. 2). Thus, changes in gene expression are much moreprominent in brain areas affected at the early stages of ADpathology.To select genes consistently altered in AD brains, considering
the several sources of measurement variations in RNAseqexperiments14, we decided to focus only on DEGs replicated inat least two independent datasets obtained from related brainareas. This resulted in a set of 734 DEGs detected in both Mayoand MSBB TL (temporal lobe intersection—TLI), and 31 DEGsshared between ROSMAP and MSBB FL (frontal lobe intersection—FLI) (Supplementary Table 2). Among TLI DEGs, we observedABCA1 and 2 (ATP-binding cassette subfamily A member 1 and 2),primarily involved in the maintenance of normal brain home-ostasis and associated with AD and other neurological diseases15;Complement C1R and C1S, involved in the immune/inflammatoryresponse and previously shown to be upregulated in the brain of a3 × Tg mouse model of AD when Aβ plaques start to accumu-late16; RE1 silencing transcription factor (REST), which regulatesneural circuit activity during aging17; glutamate decarboxylase 1and 2 (GAD1 and 2), solute carrier family 32 GABA vesiculartransporter, member 1 (SLC32A1), calbindin 1 (CALB1), parvalbu-min (PVALB), somatostatin (SST), and vasoactive intestinal peptide(VIP), all expressed in GABAergic neurons and involved incognitive decline in AD and other neurological diseases18. Amongthe few DEGs common to TLI and FLI, we observed a significantdownregulation of the neurosecretory protein VGF (VGF nervegrowth factor inducible), recently suggested as a key regulator ofAlzheimer’s disease19.
Fig. 1 Schematic summary of the methodology. A Datasets obtained from three consortia (Mayo, MSBB, and ROSMAP) were groupedaccording to the brain region sampled in the frontal lobe (FL) or temporal lobe (TL). Next, RNAseq data were pseudo-aligned using Kallisto.Clinico-pathological classifications were included as metada. B scRNAseq data from the midle temporal gyrus (MTG, Allen Brain Atlas) wereanalyzed using the R package SEURAT. C Gene expression analyses were performed using the R packages DESeq2, IsoformSwitchAnalyzeR(ISAR), and gene set enrichment analysis (GSEA). Assignment of differentially expressed genes or isoform switches to specific cell types/subtypes was performed indirectly using scRNAseq signatures obtained from the MTG (B).
D. Marques-Coelho et al.
2
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine
1234567890():,;

Next, we used gene set enrichment analyses (GSEA) to assessthe functional profile of the DEGs identified in our analysis. Again,we used only genes commonly altered in two datasets (TLI or FLI)to avoid inaccurate results associated with the use of large genesets in functional analysis20. We found that TLI DEGs weresignificantly enriched for terms (GO:BP, GO:CC and KEGG)associated with generic biological processes, such as cell-signaling pathways and cell-cell signaling, whereas the smallnumber of DEGs in the FLI were not significantly enriched for anyterm (Fig. 2C and Supplementary Table 3). The limited number ofsignificant gene set enrichment observed in our analysis afterinputting DEGs is in disagreement with results reported by Canchiet al.13. This discrepancy can likely be explained by the use ofstringent criteria to detect TLI DEGs in our study (only genesdetected in at least two independent datasets with FC > 1.3 and
FDR < 0.01), which significantly reduce the number of genes usedin the GSEA.
Differential transcript usage analysis reveals novel genesassociated with AD pathology
Gene-level expression analysis lacks the sensitivity to detectpossible changes at the transcript-level caused, for example, byalterations in alternative splicing10,21. To overcome this limitation,we used differential transcript usage (DTU) analysis to identifyadditional alterations of gene expression in the AD brainscompared to controls. We observed 2509 and 1843 genes withdifferential transcript usage (gDTU) in the temporal lobe of ADbrains studied in the Mayo and MSBB datasets, respectively(Fig. 3A, B and Supplementary Table 1). Similar to what weobserved for DEGs, a much smaller number of gDTUs were
Fig. 2 Gene expression alterations are more prominent in the temporal than the frontal lobe of AD patients. A Volcano plots showingdifferentially expressed genes (DEG, red dots; FC > 1.3 and FDR < 0.01) in the frontal lobe (ROSMAP and MSBB FL - BM10 and BM44) andtemporal lobe (Mayo and MSBB TL - BM22 and BM36). B Upset plot showing the total number of DEGs identified in each dataset (horizontalbars) and the number of DEGs exclusive of one dataset (first four vertical bars) or shared by different datasets (other vertical bars). Black dotsbelow vertical bars indicate datasets quantified. Venn diagram illustrates the same results in colors and circle sizes. C Gene ontology termsenriched for DEGs identified in the TL or FL intersections (TLI and FLI, respectively).
D. Marques-Coelho et al.
3
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

detected in the frontal lobe, both in ROSMAP and MSBB studies (59and 855 genes with transcripts altered, respectively). We found 435gDTUS in TLI (11.1% of overlap; P= 6.16 × 10−25, hypergeometrictest) and 13 gDTUs in FLI (1.47% of overlap; P= 2.56 × 10−3,hypergeometric test) (Supplementary Table 2). In TLI, most gDTUsdid not overlap with DEGs (TL—34 gDTUs that are DEGs out of 435gDTUs, Fig. 4A), whereas in FLI, we found no overlap at all.Consistent with this small overlap, GSEA using only DEGs, onlygDTUs or both showed complementary results (Fig. 4B). GSEAusing gDTUs (alone or in combination with DEGs) showedsignificant enrichment for vesicle-mediated transport and other
synapse-related terms, which were not observed while inputtingonly DEGs (Figs. 3C and 4B; Supplementary Table 3). The functionalenrichment annotation using both DEGs and gDTUs is in agreementwith previous studies using scRNAseq to identify gene expressionalterations in unique cell types6,7 and clearly improves theannotation observed using only DEGs, suggesting that the use ofDTU analysis could contribute to unraveling gene expressionalterations overlooked in the classical DEG analysis.Among genes with isoform switches enriched in synaptic-
related terms, we observed the AD causal gene APP, previouslyassociated with regulation of synapse transmission and long-term
Fig. 3 Differential transcript usage analysis identifies gene expression alterations in AD associated with synapse transmission. A Volcanoplots showing genes with differential transcript usage (gDTU, yellow dots; Differential isoform fraction (dIF) >0.05 and FDR < 0.05) in thefrontal lobe (ROSMAP and MSBB_FL - BM10 and BM44) and temporal lobe (Mayo and MSBB_TL - BM22 and BM36). B Upset plot showing thetotal number of gDTUs identified in each dataset (horizontal bars) and the number of gDTUs exclusive of one dataset (first four vertical bars)or shared by different datasets (other vertical bars). Black dots below vertical bars indicate datasets quantified. Venn diagram illustrates thesame results in colors and circle sizes. C Synapse-related terms enriched for gDTUs in the TLI are not observed in the FLI.
D. Marques-Coelho et al.
4
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine

plasticity in AD22; neuronal vesicle trafficking associated 1 (NSG1),which has been implicated in the regulation of AMPA receptors(AMPAR) and APP trafficking, thus affecting synaptic transmission,plasticity, and Aβ production23,24; RELN (Reelin) that plays
important role in synaptic transmission and has been associatedwith AD25; gamma-aminobutyric acid type A receptor subunitalpha 1 (GABRA1), which encodes for a subunit of the mainionotropic GABA receptor in the brain and has previously been
shown to be downregulated in the AD brain26.
Alternative splice events in AD brains and functionalconsequences
To identify the causes subjacent to gene isoform switches in theAD brain, we quantified the frequency of splicing events
associated with the isoform switches detected in AD comparedto control brains (Fig. 5 and Supplementary Table 4). We foundthat alternative transcription start site (ATSS), alternative transcrip-tion termination site (ATTS), and exon skipping (ES) were the mostfrequent splicing events in AD brains (Fig. 5B). Other commonsplicing events observed were alternative 3′ or 5′ splice sites (A3and A5, respectively), multiple exons skipping (MES) and intronretention (IR) (Fig. 5B). These observations suggest that changes inalternative splicing could be implicated in AD pathogenesis,corroborating previous analyses in the ROSMAP cohort usingintronic usage ratios to identify abnormal splicing events in theAD brain5.Alternative splicing events may have diverse functional
consequences for protein expression, such as shifting thefrequency of transcripts containing introns (noncoding) or mRNA
Fig. 4 Differential transcript usage analysis in AD brains reveals gene expression alterations overlooked in DEG analysis. A Venn diagramshowing DEGs and gDTUs identified in the TLI. B Comparison of GO and KEGG terms enriched for DEG, gDTU, or DEG+ gDTU identified inthe TLI.
D. Marques-Coelho et al.
5
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

stability (nonsense-mediated decay) or leading to gain/loss ofprotein domains, intrinsically disordered regions, or signalingpeptides12. Quantification of these consequences revealed someinteresting differences between Mayo and MSBB BM36 (Fig. 5C),the two datasets with the largest numbers of gDTUs. Whereas inthe Mayo dataset, a high number of isoforms showed loss ofcoding potential and protein domains, in the MSBB BM36 isoformsshowed an even distribution of loss and gain of coding potentialor protein domains (Fig. 5B). These differences could be at leastpartly explained by the larger number of gDTUs detected in theMayo compared to MSBB TL (Fig. 3) and are likely related to thedifferent median read depth of these datasets (Mayo—12.58billion bases; MSBB BM22—3.23 billion bases; MSBB BM36—3.56billion bases)27.
Differential expression of genes involved in alternative splicingcorrelates with isoform switches during disease progression
To evaluate whether the emergence of gDTUs could be correlatedwith AD pathology hallmarks, we quantified the total of gDTUsobserved at different disease stages in the MSBB dataset using theBraak classification (Fig. 6 and Supplementary Table 5). For this
purpose, we subdivided samples into three groups: low Braak (0, 1,and 2)— 196 samples (clinical diagnosis: 15 AD and 181 controls);mid-Braak (3 and 4)—133 samples (clinical diagnosis: 58 AD and75 controls); and high Braak (5 and 6)—308 samples (clinicaldiagnosis: 305 AD and 3 controls). Next, we evaluated the numberof gDTUs when comparing individuals at these different stages(Fig. 6). We observed that most gDTUs were detected only whilecomparing high with either low or mid-Braak stages (Fig. 6A–D).This pattern was observed both in the FL (BM10 and BM44) and TL(BM22 and BM36), suggesting that gene isoform switchespositively correlate with AD pathology progression.Next, we set out to evaluate alterations in the expression of
genes encoding for proteins of the splicing machinery betweenthe same Braak stages. We found that among 441 genes related to“splicing” or “spliceosome” terms (Supplementary Table 6), 79were DEGs at high compared to low or mid-Braak stages (Fig. 6E).In contrast, we could not detect any DEG in the comparison of midvs low Braak stages. Among DEGs detected in the comparisonbetween high and low/mid-Braak stages, we observed that severalgenes specifically associated with the neuronal splicing regulatorynetwork28, such as RBFOX1 and 2 (RNA binding Fox-1 homolog 1and 2), ELAVL2 (ELAV like RNA binding protein 2), MBNL3
Fig. 5 Alternative splicing mechanisms associated with isoform switches and consequences for protein expression. A Schematic showingdifferent splicing events that can lead to gene isoform switches. B Quantification of the number of isoforms showing more or less splicingevents in AD compared to controls for each dataset. C Quantification of the number of isoforms showing (i) gain or loss of coding potential,domains/signal peptides identified, intrinsically disordered regions (IDR), intron retention, open-reading frame (ORF) sequencing similarity; (ii)switch (simultaneous gain and loss) of domains identified or IDR; (iii) sensitive or insensitive to nonsense-mediated decay (NMD); and (iv)longer or shorter ORF sequencing similarity.
D. Marques-Coelho et al.
6
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine

A
B
C
D
E
F
Fig. 6 Coincidence between altered expression of splicing-related genes and gDTUs in advanced pathologic stages of AD. A–D Upsetplots showing the total number of gDTU identified in the comparison between different Braak stages (low vs. high, low vs. mid, and mid vs.high) in BM10 (A), BM44 (B), BM22 (C), or BM36 (D). Horizontal bars show the total number of gDTUs identified in each comparison (low vs.high, low vs. mid, and mid vs. high), whereas vertical bars indicate the gDTUs exclusive or common to different comparisons. Black dots belowvertical bars indicate stages analyzed. E, F Differential expression of genes associated with splicing/spliceosome after comparison of differentBraak stages (E) or AD vs controls in different datasets (F). Red and blue squares indicate, respectively, up- and downregulated genes. Genesymbols highlighted in red indicate genes belonging to the neuronal splicing machinery.
D. Marques-Coelho et al.
7
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

(muscleblind like splicing regulator 3), PTBP1 (polypyrimidine tractbinding protein 1), and NOVA2 (NOVA alternative splicingregulator 2) (Fig. 6E, highlighted in red). A similar correlationbetween pathological burden and differential expression of thesame 441 splice-related genes was observed in the comparisonbetween all AD versus control subjects of the different datasets(Fig. 6F). Changes in the expression of those genes were hardlyobserved in FL (low number of gDTUs—Fig. 3), but were frequentin TL samples (high number of gDTUs—Fig. 3), albeit to a lesserextent than that observed in the comparison between differentBraak stages (likely due to the effects of combining low, mid andhigh Braak stages in the AD group). Remarkably, the majority ofthe splicing-related genes with altered expression in the Mayodataset was not reproduced in the MSBB BM36 dataset, and viceversa (Fig. 6F). This could help to explain the dissimilarconsequences of alternative splicing events observed in thosedatasets (Fig. 5C) and suggest that a myriad of proteins could beinvolved in altered splicing in the AD brains.
Differential gene expression in separate cell types of the humanbrain
Considering the cellular diversity in the brain, we took an indirectapproach to sort DEGs and gDTUs according to individual celltypes. To that, we used scRNAseq data obtained from the adulthuman brain to identify cell types expressing the genes altered inour DEG/gDTU analysis (Fig. 7 and Supplementary Fig. 1). Wefound that, out of the 1135 genes with altered expression, i.e.,gDTU + DEG, in the TLI (Figs. 2 and 3), 839 were found in at leastone cell-type using as cutoff the expression in more than 10% ofcells assigned for a specific cell-type (Supplementary Table 7).From these, 239 were identified in unique cell-types/subtypes, 396in multiple (2–4 cell-types), and 211 in all cell-types analyzed(Fig. 7A, Supplementary Figs. 1 and 2, Supplementary Table 7).Confirming the efficacy of our strategy, GO analyses using cell-type-specific genes revealed that DEGs/gDTUs in the TLI of ADpatients were significantly enriched for biological processesassociated with inflammation in microglial cells, whereas thoseassociated with cell adhesion were enriched in endothelial cells(Fig. 7B and Supplementary Table 8). Similarly, DEGs/gDTUsidentified in neuronal cells were enriched for GO terms such assynaptic signaling, synaptic plasticity, and synapse vesicle cycle(Fig. 7C). Notably, these enrichments were more significant inGABAergic neurons, which could suggest a more pronouncedpathological burden on these cells compared to glutamatergicneurons (Fig. 7C). Comparison of the cell-type gene expressionsignatures identified in our work with previous studies usingscRNAseq in AD6,7 showed a similar degree of overlap (Supple-mentary Figs. 3 and 4; Supplementary Table 9), further supportingthe effectiveness of our strategy to assign gene expressionalterations to unique cell types in the AD brain.
DEG/gDTU analyses identify cell-type-specific alterations in ADrisk/causal genes
Genomic association studies have revealed about 45 loci contain-ing variants related to an increased or decreased probability ofdeveloping AD29,30. However, the functional variants and theirtarget genes remain mostly elusive31. To contribute to theidentification of target genes, we first evaluate the expression of176 genes located within the 45 loci associated with the AD risk(Supplementary Table 10)31 and 3 causal AD genes—PSEN1,PSEN2, and APP—in individual cell types of the adult human brain.We found that 116 out of the 179 AD risk/causal genes wereexpressed by at least one of the major cell types identified inthe brain (Fig. 8A and Supplementary Table 11). Subsets of thesegenes were exclusively expressed either in microglial cells (14 outof 116), neurons (12), astrocytes (2), oligodendrocytes (6), or
endothelial cells (6), suggesting cell-type specific roles for theseAD risk/causal genes.Next, we set out to evaluate the differential expression or
transcript usage for these genes. Out of the 116 AD risk/causalgenes expressed by brain cell types (Fig. 8A), we observed that 54were also DEGs/gDTUs in at least one of the bulk RNAseq datasetsanalyzed. Among those genes, two were exclusively identified inthe FL (Fig. 8B). We, therefore, decided to focus on the 52 AD risk/causal genes identified in the temporal lobe for further analyses.In this region, we identified 27 and 17 DEGs/gDTUs in the MSBB_TLand Mayo datasets, respectively, including some well-characterizedAD risk genes, such as ADAM10 (ADAM metallopeptidase domain10), BIN1, CLU (Clusterin), and TREM2 (triggering receptorexpressed on myeloid cells 2), and the causal AD genes APP,PSEN1 and 2 (presenilin 1 and 2) (Fig. 8A, B). Eight genes werealtered in both datasets (Fig. 8B, yellow box; 15,38% of overlap)and were selected for further analysis of isoform switch. Using ISARto identify the isoforms altered in the AD brains compared tocontrols, we observed some patterns of isoform switch thatcould have important functional relevance (Fig. 8C, D). For instance,while BIN1 transcripts ENST00000316724.9 (NP_647593.1—isoform1) and ENST00000409400.1 (NP_647600.1—isoform 9) weredownregulated, transcripts ENST00000393040.7 (NP_647598.1—isoform 6) and ENST00000462958.5, ENST0000046611.5 andENST00000484253.1 (intron retention) were upregulated (Fig. 8C).This pattern could lead to a decrease of the neuronal-specific BIN1isoform 1 expression32, given that retained introns are noncodingsequences. Using western blotting analysis, we confirmed thisdecrease of BIN1 isoform 1 protein in the frontal cortex andhippocampus of AD brain samples compared to controls (Supple-mentary Fig. 5).We also observed isoform switches in the AD causal gene APP
with possible functional consequences in neuronal cells. Whiletwo APP isoforms were downregulated (ENST00000348990 andENST00000354192), the isoforms ENST00000346798 and ENST00000357903 were upregulated in Mayo and MSBB datasets (Fig. 8D).Noteworthy, significantly downregulated APP isoforms lack exon7, which contains the Kunitz protease inhibitor (KPI) domain. KPI isone of the main serine protease inhibitors and increased KPI(+ )APP mRNA and protein expression levels have been described inAD brains and are associated with increased amyloid-betadeposition33–35. At the exception of ENST00000354192, the othertranscripts are mostly expressed in neurons (Marques-Coelho andCosta, unpublished data), indicating that these cells may have aselective increase in the expression of KPI(+)APP and, conse-quently, enhanced production of Aβ1–42.
DISCUSSION
Comprehensive knowledge of gene expression alterations asso-ciated with the onset and progression of human diseases is a keystep toward the understanding of their cellular and molecularmechanisms36. In this work, we provide a novel framework toidentify cell-type-specific gene expression alterations in AD usingpatient-derived bulk RNAseq. Comparing RNA sequencing dataobtained from distinct brain regions of control and AD patients,we show that changes in gene expression are more significant inthe temporal than frontal lobe. We also show that a large numberof genes present isoform switches without changes in the globalexpression levels. As a consequence, these genes are overlookedin classical differential expression analysis but can be detectedthrough differential transcript usage analysis. Gene isoformswitches are mostly evident at late stages of the pathology andcorrelate with altered expression of genes encoding for splicing-related proteins. Using an indirect approach to assign genes tounique cell types, we are also able to map DEGs/gDTUs to uniquecell populations of the adult brain, and our results are comparableto previously published scRNAseq data6,7. Finally, we show that a
D. Marques-Coelho et al.
8
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine

subset of AD causal/risk factors such as APP or BIN1 is differentiallyexpressed in the AD brain. Altogether, our work provides acomprehensive description of regional and cell-type-specific geneexpression changes in the AD brain and suggests that alternativesplicing could be an important mechanism for pathologicalprogression.Despite the availability of RNAseq datasets generated from
healthy subjects and AD patients2–4, a systematic evaluation of thegene expression changes in the AD brain, as well as comparisonsof these changes in distinct brain regions, was missing. To the bestof our knowledge, only one study aimed at comparing geneexpression levels in different AD brain regions37, but this workwas based on microarray data which has limited gene coverage.
We show, using bulk tissue RNAseq data, that alterations in geneexpression are highly prominent in biological samples obtainedfrom the temporal lobe, which harbors the first brain regionsaffected in the AD pathogenesis8. Conversely, few changes arepresent in biological samples derived from the frontal lobe, wherecells are affected only at advanced stages of AD. Theseobservations are in line with recent data showing that changesin protein expression levels in AD brains are much moreprominent in the temporal lobe (hippocampus, entorhinal cortex,and cingulate gyrus) than in the frontal lobe (motor cortex)9. Theycan also help to explain the low number of DEGs identified inscRNAseq data obtained from the frontal lobe7 compared to asimilar study in the entorhinal cortex6.
Fig. 7 Cell-type expression pattern for genes altered in AD brains. A Schematic representation showing our strategy to assign DEGs andgDTUs identified in the TLI to specific cell types of the adult human brain (see also Supplementary Fig. 1). Out of 839 single-cell TLI genes(scTLI), 281 were expressed in a unique cell-type, 249 in 2–4 cell-types, and 77 in all cell-types/subtypes analyzed. B Gene ontology termsenriched for scTLI DEGs, gDTUs, or both per cell type. C Selected GO terms associated with synaptic transmission.
D. Marques-Coelho et al.
9
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

Fig. 8 Expression of AD risk/causal genes is mostly altered in the TL of patients. A Heatmap showing the expression of predicted ADrisk/causal genes in different cell types of the adult human brain. DEGs and gDTUs in at least one dataset are highlighted in red. B Venndiagram showing the number of AD risk/causal DEGs or gDTUs identified in the different datasets analyzed. The intersection betweenMayo and MSBB TL is highlighted in yellow, and genes identified are shown in the yellow box. C Representation of the 6 most significantBIN1 isoforms altered (left) and quantification of the differential isoform fraction (dIF) in AD brains compared to controls (right). Mainprotein domains are indicated with different colors. D Similar representation for APP. * dIF >0.05 and FDR > 0.01.
D. Marques-Coelho et al.
10
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine

In order to minimize the variability in RNAseq experiments36,we here focused on DEGs (genes with FC > 1.3 and FDR < 0.01 inAD versus control) detected independently in at least twodatasets containing samples of similar brain regions (TLI or FLI).These stringent criteria limited the number of DEGs used insubsequent analyses, but still allowed the uncovering of severalgenes previously associated with AD pathologies, such asABCA1, ABCA2, CALB1, C1R, C1S, GAD1/2, PVALB, REST,SLC32A1, SST, VGF, and VIP15–19 The reduced number of DEGsin FLI and TLI likely explains our failure to detect functionalannotations associated with synaptic transmission and immuneresponse in GSEA, as previously reported13. However, this studyanalyzed only the ROSMAP dataset and considered genes withFDR < 0.05 as significant, regardless of the fold change,identifying 1722 DEGs in AD versus control brains. Besides thequestionable meaning of DEGs with very small fold changes, theuse of such a large set of genes for GSEA can artificially increasethe number of significantly enriched functional annotations andis not advised20.Nevertheless, our failure to detect key functional annotations
associated with AD pathology while inputting TLI DEGs is puzzlingand could suggest that DEG analysis fails to detect relevantalterations in gene expression in the AD brain. Indeed, classicalDEG analysis using DESeq or edgeR, which rank all genetranscripts, including noncoding sequences38, are insensitive tothe dynamics of gene expression that could, for example, lead toisoform switches with important functional consequences21.Therefore, important gene expression alterations could occur atthe level of transcripts, without significant changes in the globalexpression of genes. According to this possibility, we provideconvincing evidence that a high number of genes in the AD brainshow isoform switches (DTU) but are not detected by DEGanalysis, including several genes associated with the regulation ofsynapse transmission, such as APP, NSG1, RELN, GABRA122–26.Moreover, gDTUs identified in two independent datasets (TLI),alone or in combination with TLI DEGs, were enriched for keybiological processes involved in AD pathogenesis, such as synapticcommunication, immune response, inflammation, endocytosis,and cell-signaling39. Similar gene set enrichment has beendescribed using the analysis of co-expression modules in bulkRNAseq27,40 or DEG analysis DEGs in unique cell types inscRNAseq6,7. This could suggest that the combination of DEGand DTU to analyze bulk RNAseq is comparable to scRNAseqregarding the sensitivity to detect gene expression alteration inAD brains. In agreement with this possibility, we were able toassign DEGs and gDTUs to unique cell types and confirm thesimilarities among cell-type-specific functional annotationsobserved in our work compared to previous scRNAseq studies6,7.Notably, we show that several DEGs/gDTUs associated with AD
pathogenesis, such as NSG1, CALB1, RELN23–25,41 are exclusivelyassigned to GABAergic neurons. These genes may be particularlyrelevant for AD pathogenesis, given the central role of GABAergicneurons for the generation of oscillatory rhythms, networksynchrony, and memory in different animal models of AD42.Isoform switches in the APP gene could particularly affectGABAergic neurons, which express high levels of that gene,contributing to AD pathogenesis. According to this possibility,conditional knockout of APP/APLP2 only in GABAergic forebrainneurons using DlxCre mice leads to cognitive deficits inhippocampus-dependent spatial learning and memory tasks,associated with altered neuronal morphology and synapticplasticity43. It is tempting to speculate that GABAergic neuronscould be particularly vulnerable in AD, contributing to theincreased neuronal activity and synapse downscaling observedin AD brains39,44.The high number of gDTUs observed in AD brains compared to
controls can likely be explained by altered expression of genesencoding for proteins of the splicing machinery, affecting
alternative splicing. According to this interpretation, we showthat a high number of isoform switches is associated withalternative transcription start site, alternative transcription termi-nation site, exon skipping, alternative 3′ or 5′ splice sites, multipleexon skipping and intron retention. Moreover, we show thatseveral genes encoding for proteins of the splicing machineryhave their expression altered in AD brains, especially thoseshowing a high degree of pathology (Braak >5). Also in agreementwith the regional differences in gene expression described above,alterations in the splicing machinery are more prominent in the TLthan in the FL, which could help to explain the low number ofgDTUs in the latter brain region identified in our work and in theprevious study using a different strategy to detect isoform switch5.Particularly interesting, several genes encoding for proteins
involved in the control of alternative splicing in neurons aredifferently expressed in the TL of AD brains. For instance,RBFOX1 and 2 are downregulated in the MSBB BM36 and couldcontribute to the altered rate of exon skipping observed in thisregion28,45. Noteworthy, reduced expression of RBFOX1 hasbeen associated with increased inclusion of exon 7 in the APPgene, leading to an enhanced expression of APP isoforms 770and 751 containing the KPI domain45. A similar switch in the APPisoforms has also been associated with somatic gene recombi-nation in AD46, indicating that increased ratios of APP isoformscontaining the KPI domain could be detrimental to neurons.Considering these findings and the well-established associationsbetween KPI(+)APP expression levels, amyloid plaque deposi-tion, and AD pathology progression33–35, it is tempting tospeculate that controlling APP isoform switches by manipulatingRBFOX family proteins could be a potential therapeutic strategyto hamper disease progression.Altered exon skipping could also help to explain the isoform
switch observed for BIN1, which is a major risk factor for AD29,30.BIN1 comprises a N-BAR domain involved in membrane curvaturesensing, an SH3 domain that binds to proline-rich motifs, and aclathrin-binding domain (CLAP) specific of the neuronal isoform132. We show that the transcript encoding for this latter isoform issignificantly reduced in the temporal lobe, suggesting thatexpression of BIN1 isoform 1 in neurons could be reduced. Thisobservation is in line with decreased BIN1 isoform 1 proteinexpression in the AD brain compared with controls (our ownresults)47. This would be also in agreement with the observationthat overexpression of the BIN1 isoform 1 may be protective in amodel of Tauopathy48.Although we cannot formally rule out that a stage-dependent
increase in the number of DEGs and gDTUs could be due to theloss of neuronal cells in brain regions affected by the pathology,several lines of evidence indicate that this is not the mostparsimonious explanation for the data described here. First, weobserve that the percentage of up- and downregulated genes inGABAergic and glutamatergic neurons are close to 50%, ruling outthe possibility that changes in cell numbers could explain thesechanges. Secondly, previous scRNAseq studies in AD observed aconsistent fraction of cell types isolated across control and ADindividuals6,7, ruling out significant changes in the cellularcomposition of AD brains. Lastly, a large number of genes withtotal expression levels unchanged but presenting isoformswitches in the AD brains may likely presuppose a steady cellularcomposition of the tissue.Altogether, our work proposes a novel strategy to analyze bulk
RNAseq data and identify meaningful gene expression alterationsin the diseased brain. It also corroborates previous workimplicating alternative splicing in AD susceptibility5 and suggeststhat isoform switches in the gene BIN1 are involved in the reducedexpression of the main neuronal BIN1 isoform 1 in AD brains.
D. Marques-Coelho et al.
11
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

METHODS
Bulk RNAseq data from human control and disease banks
RNAseq datasets obtained from different brain regions were used (Mayo2;MSBB3; ROSMAP4). Datasets were downloaded from AMP-AD KnowledgePortal (https://www.synapse.org) following all terms and conditions to usethe data. The brain area analyzed and the number of individuals percondition was the following: Mayo - Temporal cortex, which neuroanato-mically subdivides into the inferior, middle, and superior temporal gyri(STG), and cytoarchitectonically can be subdivided into Brodmann areas(BM, instead of BA) 20/21/22/41/4249, N= 160 subjects (82 AD and 78controls); MSBB - BM22, which is part of the Wernicke’s area in the STG,N= 159 subjects (98 AD and 61 controls); MSBB BM36, corresponding tothe lateral perirhinal cortex, N= 154 subjects (88 AD and 64 controls);MSBB BM10, corresponding to the anterior prefrontal cortex, N=176 subjects (105 AD and 71 controls); MSBB BM44, corresponding tothe inferior frontal gyrus, N= 153 subjects (90 AD and 63 controls); andROSMAP - Dorsolateral prefrontal cortex (DLPFC), containing BM46 andpart of BM9, N= 423 subjects (222 AD and 201 controls). Unless statedotherwise, data obtained from different analyses were grouped in“temporal lobe” (TL) - Mayo, MSBB BM22 and MSBB BM26; or “frontallobe” (FL) - ROSMAP, MSBB BM10 and MSBB BM44.Metadata obtained from each study was used to classify patients into
Control and Alzheimer’s disease groups (Supplementary Table 12). Briefly,for the MSBB dataset, we used patients with Neuropathology Category(NP.1) labeled as “Control” and “definitive Alzheimer”. For the Mayo
dataset, we used the “Diagnosis” column of the metadata, selecting only
“AD” and “Control” patients. For the ROSMAP dataset, we also used the
column “Diagnosis” of the metadata, selecting only “Control” (value = 1)
and “Alzheimer with no other conditions” (value = 4). In all those datasets,
subjects marked as “AD” showed Braak stage values higher than 4. In the
MSBB dataset, CDR scores of AD patients were consistently higher than 2.
In the Mayo and ROSMAP datasets, all AD patients had also a definitive
diagnosis according to NINCDS criteria. Covariates such as “Postmortem
interval (PMI)”, “RNA integrity number (RIN)”, “Age of death”, and “Sex”
were balanced among the different groups (Table 1; Chi-square test, p >
0.05). We used RIN and PMI as covariates in our model to control for
possible “batch effects” (linear regression). For detailed information of
all individual samples used in this study, please refer to Supplementary
Table 13.For single-cell RNA sequencing (scRNAseq) analyses, we used processed
data obtained from the middle temporal gyrus (MTG), available at the Allen
Brain Atlas consortium (https://celltypes.brain-map.org/rnaseq).
Realignment of human reads into single pseudoaligner pipeline
Using human GRCh38 cDNA release 94 (ftp://ftp.ensembl.org/pub/release-94)
as a reference, we built an index to align all our fastq files. Next, we used
pseudoaligner Kallisto50 (version 0.43.1) with our pre-built index to align
fastq files.
Table 1. Summary of clinical, demographic, and technical variables of samples analyzed from different datasets.
Diagnosis Sex n Braak AOD CDR RIN PMI
Mayo–TL
AD F 49 5.55 ± 0.53 83.33 ± 7.23 – 8.58 ± 0.58 6.34 ± 5.8
AD M 33 5.42 ± 0.55 81.67 ± 8.16 – 8.59 ± 0.5 8.6 ± 5.83
Control F 37 2.03 ± 0.78 84.81 ± 8.04 – 7.59 ± 1 7 ± 7.69
Control M 41 2.36 ± 0.87 80.27 ± 9.13 – 7.64 ± 1.05 5.51 ± 6.42
MSBB–BM10
AD F 86 5.18 ± 1.21 85.94 ± 6 3.17 ± 1.35 6.05 ± 1.64 6.01 ± 3.97
AD M 53 5.48 ± 0.99 79.26 ± 7.67 3.23 ± 1.28 6.19 ± 1.42 7.38 ± 4.73
Control F 52 2.02 ± 1.09 83.04 ± 7.43 0.81 ± 1.1 6.37 ± 1.28 8.69 ± 5.73
Control M 42 1.62 ± 0.94 78.93 ± 8.56 1.23 ± 1.45 6.73 ± 1.14 11.31 ± 7.47
MSBB–BM22
AD F 88 5.25 ± 1.2 85.81 ± 5.95 3.17 ± 1.39 6 ± 1.5 5.34 ± 3.71
AD M 64 5.41 ± 0.97 79.44 ± 7.67 3.48 ± 1.35 5.53 ± 1.28 7.2 ± 4.8
Control F 45 2 ± 1.26 82.09 ± 8.1 0.94 ± 1.29 6 ± 1.23 9.18 ± 6.18
Control M 41 1.72 ± 0.97 79.29 ± 8.66 1.22 ± 1.42 5.72 ± 1.06 11.01 ± 7.29
MSBB–BM36
AD F 95 5.35 ± 1.1 86.79 ± 5.2 3.22 ± 1.28 5.59 ± 1.71 5.96 ± 4.14
AD M 53 5.47 ± 0.91 79.26 ± 7.98 3.42 ± 1.41 5.69 ± 1.61 6.65 ± 4.9
Control F 44 2.14 ± 1.15 83.45 ± 7.23 0.86 ± 1.28 6.15 ± 1.32 8.3 ± 5.66
Control M 40 1.65 ± 0.92 78.28 ± 8.79 0.85 ± 1.18 6.2 ± 1.2 12.47 ± 7.92
MSBB–BM44
AD F 84 5.2 ± 1.29 85.62 ± 6.02 3.11 ± 1.41 6.9 ± 2.51 5.74 ± 3.82
AD M 51 5.49 ± 0.98 79.69 ± 7.59 3.2 ± 1.25 6.57 ± 2.41 7.19 ± 4.58
Control F 45 2.11 ± 1.19 81.82 ± 9.06 0.93 ± 1.26 7.43 ± 2.15 8.19 ± 5.74
Control M 47 1.82 ± 0.95 78.13 ± 8.16 0.96 ± 1.22 7.35 ± 2.14 11.48 ± 7.3
ROSMAP–DLPFC
AD F 147 4.18 ± 0.93 88.55 ± 3.05 – 6.97 ± 0.88 7.25 ± 4.51
AD M 64 3.92 ± 1.28 87.6 ± 3.18 – 7 ± 1.09 6.58 ± 4.24
Control F 118 3.04 ± 1.07 85.66 ± 4.83 – 7.31 ± 0.99 7.25 ± 5.18
Control M 74 2.58 ± 1.4 83.69 ± 5.69 – 7.19 ± 1.08 7.34 ± 4.05
AOD age of death, CDR clinical dementia rate, n number of samples, RIN RNA integrity number, PMI postmortem interval (in hours), AD Alzheimer disease,
F female, M, male.
Values are mean ± SD. See supplementary Table 13 for individual values.
D. Marques-Coelho et al.
12
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine

Differential gene expression analyses. Differentially expressed genes(DEGs) were identified using differential gene expression at transcript-level using DESeq2 R library51,52. To facilitate kallisto output import,transcript-level estimated counts, length, and abundance were extractedusing tximport function53. As described by Michael Love group, transcript-level differential gene expression enhances analysis resolution52. UsingDESeqDataSetFromTximport, a DESeq2 object was created and filteredusing rows with sum of all counts bigger than 10. Next, DESeq function wasused with default values. Using the results function, we selected all geneswith a false discovery rate (FDR) < 0.01 and fold change (FC) > 1.3. We alsoused RIN and PMI as covariates (linear regression).Differential transcript usage (DTU) analysis was performed using the R
library IsoformSwitchAnalyzeR12. Following pipeline instructions, kallistoabundance tables were imported using importIsoformExpression andimportRdata functions to create a switchAnalyzeRlist object. Same cDNArelease used in kallisto alignment and correspondent annotation (ftp://ftp.ensembl.org/pub/release-94/gtf/homo_sapiens/Homo_sapiens.GRCh38.94.chr_patch_hapl_scaff.gtf.gz) were applied as input. We filtered data using agene expression cutoff= 10, isoform expression cutoff= 3, differentialisoform fraction (dIF) cutoff= 0.05 and removed single isoform genes.Although DEXSeq is recommended to test differential isoform usage, itdoes not work efficiently for large datasets (more than 100 samples)11. Forthat reason, we chose isoformSwitchAnalysisPart1 function using DRIM-Seq53 to test differential transcript usage. Using part1 fasta files, all externalanalysis was performed and used as input to isoformSwitchAnalysisPart2function. We used CPC2, Pfam, SignalIP and Netsurfp2 as indicated in thepipeline. Next, we performed a confirmation stage using stageR14 togenerate isoforms overall false discovery rate (OFDR). We selected allisoforms with OFDR < 0.01 and dIF >0.05. RIN and PMI metadata were usedas covariates (linear regression).Statistical significance of the intersections among different datasets was
calculated using the hypergeometric test (phyper).
Splicing events and event consequences
We used extractSplicingSummary and extractConsequenceSummary func-
tions to quantify gain/loss of predicted splicing events (such as exon
skipping and intron retention); and gain/loss (also sensitive/insensitive,
shorter/longer and switch) of predicted functional consequences (such as
coding potential and domain identified), respectively.
Single-cell RNAseq
Using R library seurat54, we created a seurat object (CreateSeuratObject),
normalized data (NormalizeData), found variable genes (FindVariableFeatures),
and rescaled data using a linear model (ScaleData, use.umi= F). After that, we
generated 50 PC’s (RunPCA) but only used 35 of them based on the PC’s
visualization distribution (ElbowPlot). Since Allen data were already anno-
tated, we only used tSNE (RunTSNE) to facilitate visualization. A group
classified as “None” by Allen metadata were removed from our analysis. This
strategy generated 7 main different cell types: Astrocytes, Endothelial cells,
Glutamatergic Neurons, GABAergic Neurons, Microglia, Oligodendrocytes
and oligodendrocyte precursor cells (OPCs). To assign genes to specific cell
types, we used the AverageExpression function. Using pct.exp bigger than
0.1, we created a list of genes that were expressed by each cell type.
Gene set enrichment analysis (GSEA)
For gene ontology analysis, R library gprofiler255 was used. Using gost
function, correction_method= “fdr” and significant=TRUE. To minimize
the enrichment of gene ontologies based on a small set of genes, we used
three conditions for significance assessment: false discovery rate (FDR) <
0.01; intersection size (intersection between gene set vs. a number of
genes in a term) >3; and precision (intersection size divided by gene set)
>0.03. We used Gene Ontology (GO or by branch GO:MF, GO:BP, GO:CC),
Kyoto Encyclopedia of Genes and Genomes (KEGG), Reactome (REAC),
WikiPathways (WP), TRANSFAC (TF), miRTarBase (MIRNA), Human Protein
Atlas (HPA), CORUM (CORUM), Human phenotype ontology (HP) as
sources. For improved visualization, we plotted results only for GO:BP,
GO:CC and KEGG and show only FDR related to terms reaching all criteria
of significance.
Selection of splicing-associated genes
To select splicing-related genes, we searched for terms containing thewords “splicing” or “spliceosome” in gProfiler bank (https://biit.cs.ut.ee/gprofiler/gost). Taking only GO and WP datasets, 25 terms and 441 genesrelated to those terms were selected (Supplementary Table 6).
Selection of AD risk/causal genes
The complete list of AD risk/causal genes used in this study is described inSupplementary Table 10. Briefly, AD risk loci were selected from previouswork using genome-wide association studies and whole exome sequen-cing29,30. AD risk genes within these loci were determined based onregional association plots, assuming that the functional risk variants arelocated in the vicinity of the SNP producing the top signal and taking intoaccount the linkage disequilibrium patterns and the recombination peakswithin the loci of interest31. Early-onset AD causal genes used in this studyare APP, PSEN1, and PSEN2.
Western blotting
Frozen brain samples obtained from the frontal cortex (FCx) andhippocampus (hip) of three non-pathology (age: 80.33 ± 3.78 years;Braak: 2.66 ± 1.15; PMI: 37.33 ± 22.50 h) and six AD patients (age: 79.57 ±6.70 years; Braak: 6; PMI: 26.57 ± 13.40 h) were lysed with RIPA buffer andsonicated at 100% during 10 s before use for the western blottinganalyses. The controls for BIN1 isoforms 1 (Iso1) and 9 (Iso9) wereobtained using HEK cells transiently transfected with 1 µg/ml DNAsolution containing plasmids encoding for BIN1 isoforms mixed with thetransfection reagent FuGENE HD (Promega) at the ratio 1:3. Cells werelysed using RIPA buffer 48 h after transfection and frozen for furtheranalyses.Protein quantification was performed using the BCA protein assay
(Thermo Scientific). In total, 10–20 μg of total protein from extracts wereseparated in SDS–polyacrylamide gels 4–12% (NuPAGE Bis-Tris, ThermoScientific) and transferred to nitrocellulose membranes (Bio-Rad). Next,membranes were incubated in milk (5% in Tris-buffered saline with 0.1%Tween-20, TTBS) to block non-specific binding sites during 1 h at RT,followed by several washes with TTBS. Immunoblotting was carried outwith primary antibodies anti-BIN1 (Abcam, ab182562), anti-β-ACTIN(Sigma-Aldrich, A1978), and anti-GAPDH (Millipore, AB2302) overnight at4 °C on 20 RPM. The membranes were washed three times in TTBS,followed by incubation with HRP-conjugated secondary antibodies(Jackson, anti-mouse 115-035-003 and anti-rabbit 111-035-003; ThermoScientific, anti-chicken A16054) overnight at 4 °C on 20 RPM agitation. Themembranes were washed three times in TTBS, and the immunoreactivitywas revealed using the ECL chemiluminescence system (SuperSignal,Thermo Scientific) and imaged using the Amersham Imager 600 (GE LifeSciences). Optical densities of bands were quantified using “Gel Analyzer”plugin in Fiji56.All western blot experiments were performed in compliance with
relevant guidelines provided by the Neuro-CEB - Biological ResourcesPlatform and the protocols were approved by the National Institute ofHealth and Medical Research (INSERM), at the Institut Pasteur de Lille,University of Lille.
Reporting summary
Further information on research design is available in the Nature ResearchReporting Summary linked to this article.
DATA AVAILABILITY
MSBB RNAseq3: https://doi.org/10.7303/syn3388564; ROSMAP RNAseq4: https://doi.
org/10.7303/syn3157743; MAYO RNAseq2: https://doi.org/10.7303/syn5550404.
CODE AVAILABILITY
Codes used for analyses are available, differentially expressed genes and differential
transcript usage in individual datasets can be assessed using https://diegomscoelho.
github.io/AD-IsoformSwitch/.
Received: 5 May 2020; Accepted: 12 November 2020Published online: 04 January 2021
D. Marques-Coelho et al.
13
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

REFERENCES
1. Masters, C. L. et al. Alzheimer’s disease. Nat. Rev. Dis. Primers 1, 15056 (2015).
2. Allen, M. et al. Human whole genome genotype and transcriptome data for
Alzheimer’s and other neurodegenerative diseases. Sci. Data 3, 160089
(2016).
3. De Jager, P. L. et al. Data descriptor: a multi-omic atlas of the human frontal
cortex for aging and Alzheimer’s disease research. Sci. Data 5, 180142–180142
(2018).
4. Wang, M. et al. The Mount Sinai cohort of large-scale genomic, transcriptomic
and proteomic data in Alzheimer’s disease. Sci. Data 5, 1–16 (2018).
5. Raj, T. et al. Integrative transcriptome analyses of the aging brain implicate
altered splicing in Alzheimer’s disease susceptibility. Nat. Genet. 50, 1584–1592
(2018).
6. Grubman, A. et al. A single-cell atlas of entorhinal cortex from individuals with
Alzheimer’s disease reveals cell-type-specific gene expression regulation. Nat.
Neurosci. 22, 2087–2097 (2019).
7. Mathys, H. et al. Single-cell transcriptomic analysis of Alzheimer’s disease. Nature
570, 332–337 (2019).
8. Braak, H. & Braak, E. Neuropathological stageing of Alzheimer-related changes.
Acta Neuropathologica 82, 239–259 (1991).
9. Xu, J. et al. Regional protein expression in human Alzheimer’s brain correlates
with disease severity. Commun. Biol. 2, 1–15 (2019).
10. Yi, L., Pimentel, H., Bray, N. L. & Pachter, L. Gene-level differential analysis at
transcript-level resolution. Genome Biol. 19, 53 (2018).
11. Anders, S., Reyes, A. & Huber, W. Detecting differential usage of exons from RNA-
seq data. Genome Res. 22, 2008–2017 (2012).
12. Vitting-Seerup, K. & Sandelin, A. IsoformSwitchAnalyzeR: analysis of changes in
genome-wide patterns of alternative splicing and its functional consequences.
Bioinformatics 35, 4469–4471 (2019).
13. Canchi, S. et al. Integrating gene and protein expression reveals perturbed
functional networks in Alzheimer’s disease. Cell Rep. 28, 1103–1116.e4 (2019).
14. Van den Berge, K., Soneson, C., Robinson, M. D. & Clement, L. stageR: a general
stage-wise method for controlling the gene-level false discovery rate in dif-
ferential expression and differential transcript usage. Genome Biol. 18, 151
(2017).
15. Abuznait, A. H. & Kaddoumi, A. Role of ABC transporters in the pathogenesis of
Alzheimers disease. ACS Chem. Neurosci. 3, 820–831 (2012).
16. Benoit, M. E. et al. C1q-induced LRP1B and GPR6 proteins expressed early in
Alzheimer disease mouse models are essential for the C1q-mediated protection
against amyloid- β neurotoxicity. J. Biol. Chem. 288, 654–665 (2013).
17. Zullo, J. M. et al. Regulation of lifespan by neural excitation and REST. Nature 574,
359–364 (2019).
18. Prévot, T. & Sibille, E. Altered GABA-mediated information processing and cog-
nitive dysfunctions in depression and other brain disorders. Mol. Psychiatry 1–17
(2020) https://www.nature.com/articles/s41380-020-0727-3.
19. Beckmann, N. D. et al. Multiscale causal networks identify VGF as a key regulator
of Alzheimer’s disease. Nat. Commun. 11, 18 (2020).
20. Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based
approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci.
USA 102, 15545–15550 (2005).
21. Vitting-Seerup, K. & Sandelin, A. The landscape of isoform switches in human
cancers. Mol. Cancer Res. 15, 1206–1220 (2017).
22. Kamenetz, F. et al. APP processing and synaptic function. Neuron 37, 925–937
(2003).
23. Alberi, S. et al. The endosomal protein NEEP21 regulates AMPA receptor-
mediated synaptic transmission and plasticity in the hippocampus. Mol. Cell.
Neurosci. 29, 313–319 (2005).
24. Norstrom, E. M., Zhang, C., Tanzi, R. & Sisodia, S. S. Identification of NEEP21 as a
β-amyloid precursor protein-interacting protein in vivo that modulates amyloi-
dogenic processing in vitro. J. Neurosci. 30, 15677–15685 (2010).
25. Yu, N. N., Tan, M. S., Yu, J. T., Xie, A. M. & Tan, L. The role of reelin signaling in
Alzheimer’s disease. Mol. Neurobiol. 53, 5692–5700 (2016).
26. Limon, A., Reyes-Ruiz, J. M. & Miledi, R. Loss of functional GABA A receptors in
the Alzheimer diseased brain. Proc. Natl Acad. Sci. USA 109, 10071–10076
(2012).
27. Wan, Y. W. et al. Meta-analysis of the Alzheimer’s disease human brain tran-
scriptome and functional dissection in mouse models. Cell Rep. 32, 107908
(2020).
28. Raj, B. & Blencowe, B. J. Alternative splicing in the mammalian nervous system:
recent insights into mechanisms and functional roles. Neuron 87, 14–27 (2015).
29. Lambert, J. C. et al. Meta-analysis of 74,046 individuals identifies 11 new sus-
ceptibility loci for Alzheimer’s disease. Nat. Genet. 45, 1452–1458 (2013).
30. Kunkle, B. W. et al. Genetic meta-analysis of diagnosed Alzheimer’s disease
identifies new risk loci and implicates Aβ, tau, immunity and lipid processing. Nat.
Genet. 51, 414–430 (2019).
31. Dourlen, P., Kilinc, D., Malmanche, N., Chapuis, J. & Lambert, J. C. The new genetic
landscape of Alzheimer’s disease: from amyloid cascade to genetically driven
synaptic failure hypothesis? Acta Neuropathologica 138, 221–236 (2019).
32. Zhou, Y. et al. Intracellular clusterin interacts with brain isoforms of the bridging
integrator 1 and with the microtubule-associated protein Tau in Alzheimer’s
disease. PLoS ONE 9, e103187 (2014).
33. Kitaguchi, N., Takahashi, Y., Tokushima, Y., Shiojiri, S. & Ito, H. Novel precursor of
Alzheimer’s disease amyloid protein shows protease inhibitory activity. Nature
331, 530–532 (1988).
34. Johnson, S. A., Rogers, J. & Finch, C. E. APP-695 transcript prevalence is selectively
reduced during Alzheimer’s disease in cortex and hippocampus but not in cer-
ebellum. Neurobiol. Aging 10, 755–760 (1989).
35. Tanzi, R. E. et al. Protease inhibitor domain encoded by an amyloid protein
precursor mRNA associated with Alzheimer’s disease. Nature 331, 528–530
(1988).
36. Lee, T. I. & Young, R. A. Transcriptional regulation and its misregulation in disease.
Cell 152, 1237–1251 (2013).
37. Haroutunian, V., Katsel, P. & Schmeidler, J. Transcriptional vulnerability of brain
regions in Alzheimer’s disease and dementia. Neurobiol. Aging 30, 561–573
(2009).
38. Costa-Silva, J., Domingues, D. & Lopes, F. M. RNA-Seq differential expression
analysis: an extended review and a software tool. PLoS ONE 12, e0190152 (2017).
39. Canter, R. G., Penney, J. & Tsai, L. H. The road to restoring neural circuits for the
treatment of Alzheimer’s disease. Nature 539, 187–196 (2016).
40. Milind, N. et al. Transcriptomic stratification of late-onset Alzheimer’s cases
reveals novel genetic modifiers of disease pathology. PLoS Genet. 16, e1008775
(2020).
41. Odero, G. L. et al. Evidence for the involvement of calbindin D28k in the pre-
senilin 1 model of Alzheimer’s disease. Neuroscience 169, 532–543 (2010).
42. Verret, L. et al. Inhibitory interneuron deficit links altered network activity and
cognitive dysfunction in alzheimer model. Cell 149, 708–721 (2012).
43. Mehr, A. et al. Lack of APP and APLP2 in GABAergic forebrain neurons impairs
synaptic plasticity and cognition. Cereb. Cortex 30, 4044–4063 (2020).
44. Dörrbaum, A. R., Alvarez-Castelao, B., Nassim-Assir, B., Langer, J. D. & Schuman, E.
M. Proteome dynamics during homeostatic scaling in cultured neurons. eLife 9,
e52939 (2020).
45. Alam, S., Suzuki, H. & Tsukahara, T. Alternative splicing regulation of APP exon 7
by RBFox proteins. Neurochem. Int. 78, 7–17 (2014).
46. Lee, M. H. et al. Somatic APP gene recombination in Alzheimer’s disease and
normal neurons. Nature 563, 639–645 (2018).
47. Glennon, E. B. C. et al. BIN1 is decreased in sporadic but not familial Alzheimer’s
disease or in aging. PLoS ONE 8, e78806 (2013).
48. Sartori, M. et al. BIN1 recovers tauopathy-induced long-term memory deficits in
mice and interacts with Tau through Thr348 phosphorylation. Acta Neuropatho-
logica 138, 631–652 (2019).
49. Strotzer, M. One century of brain mapping using Brodmann areas. Clin. Neuror-
adiol. 19, 179–186 (2009).
50. Bray, N. L., Pimentel, H., Melsted, P. & Pachter, L. Near-optimal probabilistic RNA-
seq quantification. Nat. Biotechnol. 34, 525–527 (2016).
51. Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and
dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
52. Love, M. I., Soneson, C. & Patro, R. Swimming downstream: statistical analysis of
differential transcript usage following Salmon quantification. F1000Research 7,
952–952 (2018).
53. Soneson, C., Love, M. I. & Robinson, M. D. Differential analyses for RNA-seq:
transcript-level estimates improve gene-level inferences [version 2; referees: 2
approved]. F1000Research 4, 1521 (2016).
54. Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell
transcriptomic data across different conditions, technologies, and species. Nat.
Biotechnol. 36, 411–420 (2018).
55. Kolberg, L., Raudvere, U., Kuzmin, I., Vilo, J. & Peterson, H. gprofiler2—an R
package for gene list functional enrichment analysis and namespace conversion
toolset g:Profiler. F1000Research 9, 709 (2020).
56. Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat.
Methods 9, 676–682 (2012).
ACKNOWLEDGEMENTS
We thank Charles Duyckaerts (Neuroceb, GIE Neuro-CEB BB-0033-00011) for
providing the brain samples. We would also like to thank Dr. Tarciso Velho, Pierre
Dourlen, and Fabien Delahaye for reading and suggestions on the paper. The results
published here are in whole or in part based on the data obtained from the AMP-AD
Knowledge Portal (https://adknowledgeportal.synapse.org/). Study data were pro-
vided by the following sources: The Mayo Clinic Alzheimer’s Disease Genetic Studies,
led by Dr. Nilufer Taner and Dr. Steven G. Younkin, Mayo Clinic, Jacksonville, FL using
D. Marques-Coelho et al.
14
npj Aging and Mechanisms of Disease (2021) 2 Published in partnership with the Japanese Society of Anti-Aging Medicine

samples from the Mayo Clinic Study of Aging, the Mayo Clinic Alzheimer’s Disease
Research Center, and the Mayo Clinic Brain Bank. Data collection was supported
through funding by NIA grants P50 AG016574, R01 AG032990, U01 AG046139, R01
AG018023, U01 AG006576, U01 AG006786, R01 AG025711, R01 AG017216, R01
AG003949, NINDS grant R01 NS080820, CurePSP Foundation, and support from Mayo
Foundation. Study data include samples collected through the Sun Health Research
Institute Brain and Body Donation Program of Sun City, Arizona. The Brain and Body
Donation Program is supported by the National Institute of Neurological Disorders
and Stroke (U24 NS072026 National Brain and Tissue Resource for Parkinson’s Disease
and Related Disorders), the National Institute on Aging (P30 AG19610 Arizona
Alzheimer’s Disease Core Center), the Arizona Department of Health Services
(contract 211002, Arizona Alzheimer’s Research Center), the Arizona Biomedical
Research Commission (contracts 4001, 0011, 05-901, and 1001 to the Arizona
Parkinson’s Disease Consortium) and the Michael J. Fox Foundation for Parkinson’s
Research. Study data were also provided by the Rush Alzheimer’s Disease Center,
Rush University Medical Center, Chicago. Data collection was supported through
funding by NIA grants P30AG10161 (ROS), R01AG15819 (ROSMAP; genomics and
RNAseq), R01AG17917 (MAP), the Illinois Department of Public Health (ROSMAP), and
the Translational Genomics Research Institute (genomic). For ROSMAP, additional
phenotypic data can be requested at www.radc.rush.edu. Part based on the data
generated from postmortem brain tissue collected through the Mount Sinai VA
Medical Center Brain Bank and were provided by Dr. Eric Schadt from Mount Sinai
School of Medicine. This work was co-funded by the European Union under the
European Regional Development Fund (ERDF) and by the Hauts de France Regional
Council (contrat no.18006176), the MEL (contract_2016_ESR_05), and the French
State (contract no. 2018-3-CTRL_IPL_Phase2) to MRC. This work was also funded by
the Lille Métropole Communauté Urbaine and the French government’s LABEX
DISTALZ program (Development of innovative strategies for a transdisciplinary
approach to Alzheimer’s disease) to J.C.L. D.M.C. and L.I.C.C. are supported by
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) scholarships.
AUTHOR CONTRIBUTIONS
M.R.C. conceived of the presented idea and supervised the experiments. D.M-C.
performed all bioinformatic analyses and prepared the figures and tables of the
paper. L.I.C.C. contributed with bioinformatic analyses. A.F. prepared protein lysates
and A.R.M.F. performed western blot analysis in human samples. D.M.-C., J.-C.L., and
M.R.C. wrote the paper. All authors revised and approved the final version of
the paper.
COMPETING INTERESTS
The authors declare no competing interests.
ADDITIONAL INFORMATION
Supplementary information is available for this paper at https://doi.org/10.1038/
s41514-020-00052-5.
Correspondence and requests for materials should be addressed to M.R.C.
Reprints and permission information is available at http://www.nature.com/
reprints
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims
in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons
Attribution 4.0 International License, which permits use, sharing,
adaptation, distribution and reproduction in anymedium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative
Commons license, and indicate if changes were made. The images or other third party
material in this article are included in the article’s Creative Commons license, unless
indicated otherwise in a credit line to the material. If material is not included in the
article’s Creative Commons license and your intended use is not permitted by statutory
regulation or exceeds the permitted use, you will need to obtain permission directly
from the copyright holder. To view a copy of this license, visit http://creativecommons.
org/licenses/by/4.0/.
© The Author(s) 2021
THE BRAINBANK NEURO–CEB NEUROPATHOLOGY NETWORK
Franck Letournel4, Marie-Laure Martin-Négrier5, Françoise Chapon6, Maxime Faisant6, Catherine Godfraind7, Claude-Alain Maurage8,
Vincent Deramecourt8, Mathilde Duchesne9, David Meyronnet10, Nathalie Streichenberger10, André Mauès de Paula11, Valérie Rigau12,
Fanny Vandenbos-Burel13, Charles Duyckaerts14, Danielle Seilhean14, Serge Milin15, Dan Christian Chiforeanu16, Annie Laquerrière17,
Florent Marguet17 and Béatrice Lannes18
4CHU Hôpital St Jean, Angers, France. 5CHRU Pellegrin, Bordeaux, France. 6CHU Côte de Nacre, Caen, France. 7CHU Gabriel Montpied, Clermont-Ferrand, France. 8CHRU Hôpital
Roger Salengro, Lille, France. 9CHRU Hôpital Dupuytren, Limoges, France. 10Groupe Hospitalier Est. Hospices Civils de Lyon, Lyon, France. 11CHU Hôpital de La Timone, APHM,
Marseille, France. 12CHU de Montpellier, Montpellier, France. 13CHU Hôpital Pasteur, Nice, France. 14GH Pitié-Salpêtrière, APHP, Paris, France. 15CHU La Milétrie, Poitiers, France.16CHU Hôpital Pontchaillou, Rennes, France. 17CHU Hôpital Charles Nicolle, Rouen, France. 18CHRU Hôpital de Hautepierre, Strasbourg, France.
D. Marques-Coelho et al.
15
Published in partnership with the Japanese Society of Anti-Aging Medicine npj Aging and Mechanisms of Disease (2021) 2

&DStWXOR�,,
'LIIHUHQWLDO�H[SUHVVLRQ�OHYHOV�RI�6R[��LQ�HDUO\�QHRFRUWLFDO UDGLDO�JOLDO�FHOOV
UHJXODWH�WKH�GHFLVLRQ�EHWZHHQ�VWHP�FHOO�PDLQWHQDQFH DQG�GLIIHUHQWLDWLRQ
$XWRUHV��-DLPH�)DEUD�%HVHU��-HVVLFD�$OYHV�0HGHLURV GH�$UDXMR� 'LHJR
0DUTXHV�&RHOKR��/R\DO�$��*RII��0DUFRV�5��&RVWD��8OULFK 0�OOHU��&ULVWLQD�*LO�6DQ]
$UWLJR�VXEPHWLGR�DR�SHULyGLFR -RXUQDO�RI�1HXURVFLHQFH

)DEUD�%HVHU�HW�DO�
'LIIHUHQWLDO�H[SUHVVLRQ�OHYHOV�RI�6R[��LQ�HDUO\�QHRFRUWLFDO UDGLDO�JOLDO�FHOOV�UHJXODWH�WKH
GHFLVLRQ�EHWZHHQ�VWHP�FHOO�PDLQWHQDQFH�DQG�GLIIHUHQWLDWLRQ�
-DLPH�)DEUD�%HVHU���-HVVLFD�$OYHV�0HGHLURV�GH�$UDXMR ���� 'LHJR�0DUTXHV�&RHOKR�����/R\DO�$
*RII���0DUFRV�5�&RVWD�����8OULFK�0�OOHU���&ULVWLQD *LO�6DQ]��
�%,27(&0('�,QVWLWXWH��8QLYHUVLGDG�GH�9DOHQFLD��%XUMDVVRW� 9DOHQFLD���������6SDLQ
�7KH�6RORPRQ�+��6Q\GHU�'HSDUWPHQW�RI�1HXURVFLHQFH� -RKQV�+RSNLQV�8QLYHUVLW\�6FKRRO�RI
0HGLFLQH��%DOWLPRUH��0'��������86$
�%UDLQ�,QVWLWXWH��)HGHUDO�8QLYHUVLW\�RI�5LR�*UDQGH GR�1RUWH��1DWDO��51������������%UD]LO
�%LRLQIRUPDWLFV�0XOWLGLVFLSOLQDU\�(QYLURQPHQW��,0'� )HGHUDO�8QLYHUVLW\�RI�5LR�*UDQGH�GR
1RUWH��1DWDO��51������������%UD]LO
�8QLY�/LOOH��,QVHUP��&+8�/LOOH��,QVWLWXW�3DVWHXU�GH /LOOH��8�����5,'�$*(�IDFWHXUV�GH�ULVTXH�HW
GpWHUPLQDQWV�PROpFXODLUHV�GHV�PDODGLHV�OLpV�DX�YLHLOOLVVHPHQW� ',67$/=��/LOOH��)UDQFH�
&RUUHVSRQGLQJ�$XWKRU
$GGUHVV�FRPPXQLFDWLRQ�WR
&ULVWLQD�*LO�6DQ]
FULVWLQD�JLO#XY�HV
&RQIOLFW�RI�LQWHUHVW�VWDWHPHQWV� 7KH�DXWKRUV�GHFODUH QR�FRQIOLFWV�RI�LQWHUHVW
$FNQRZOHGJHPHQWV
7KLV�ZRUN�ZDV�VXSSRUWHG�E\�WKH�6SDQLVK�0LQLVWU\�RI 6FLHQFH�DQG�,QQRYDWLRQ��&*6�
6$)������������5��DQG�1,+��80��5)�0+���������&�*�6� KROGV�D�5DPyQ�\�&DMDO�*UDQW
IURP�WKH�6SDQLVK�0LQLVWU\�RI�6FLHQFH�DQG�,QQRYDWLRQ �5\&��������������-�$�0�$��DQG�'�0�&�
ZHUH�ILQDQFHG�LQ�SDUW�E\�WKH�&RRUGHQDomR�GH�$SHUIHLoRDPHQWR GH�3HVVRDO�GH�1tYHO�6XSHULRU
�&$3(6��%(;�������������8�0��LV�D�%ORRPEHUJ�'LVWLQJXLVKHG 3URIHVVRU�IRU�1HXURVFLHQFH�DQG
%LRORJ\��:H�WKDQN�PHPEHUV�RI�WKH�ODERUDWRU\�IRU�FRPPHQWV DQG�FULWLFLVPV��,VDEHO�)DULxDV�DQG
6DFUDPHQWR�5��)HUUyQ�IRU�FULWLFDO�FRPPHQWV��UHDJHQWV DQG�VKDULQJ�WKHLU�HTXLSPHQW�ZLWK�XV��:H
DUH�JUDWHIXO�WR�&ULVWLQD�$QGUpV�&DUERQHOO�IRU�DVVLVWDQFH ZLWK�PRXVH�PDLQWHQDQFH�
�

)DEUD�%HVHU�HW�DO�
$%675$&7
5DGLDO JOLDO SURJHQLWRU FHOOV �5*&V� LQ WKH GRUVDO WHOHQFHSKDORQ GLUHFWO\ RU LQGLUHFWO\ SURGXFH
H[FLWDWRU\ SURMHFWLRQ QHXURQV DQG PDFURJOLD RI WKH QHRFRUWH[� 5HFHQW HYLGHQFH VKRZV WKDW WKH
SRRO RI 5*&V LV PRUH KHWHURJHQHRXV WKDQ RULJLQDOO\ WKRXJKW DQG WKDW SURJHQLWRU VXESRSXODWLRQV
FDQ JHQHUDWH SDUWLFXODU QHXURQDO FHOO W\SHV� 8VLQJ VLQJOH FHOO 51$ VHTXHQFLQJ� ZH KDYH VWXGLHG
JHQH H[SUHVVLRQ SDWWHUQV RI 5*&V ZLWK GLIIHUHQW QHXURJHQLF EHKDYLRU DW HDUO\ DJHV RI FRUWLFDO
GHYHORSPHQW� 6RPH 5*&V UDSLGO\ SURGXFH SRVWPLWRWLF QHXURQV� ZKHUHDV RWKHUV SUHVHQW GHOD\HG
QHXURJHQLF GLYLVLRQV DW WKH DQDO\]HG DJH� :H KDYH LGHQWLILHG FDQGLGDWH JHQHV WKDW DUH
GLIIHUHQWLDOO\ H[SUHVVHG EHWZHHQ WKHVH SRVVLEOH 5*&V VXESRSXODWLRQV� LQFOXGLQJ WKH
WUDQVFULSWLRQ IDFWRU 6R[�� 8VLQJ LQ XWHUR HOHFWURSRUDWLRQ� ZH GHPRQVWUDWH WKDW HOHYDWHG 6R[�
H[SUHVVLRQ LQ SURJHQLWRUV DIIHFWV 5*& FHOO�F\FOH GXUDWLRQ DQG OHDGV WR WKH JHQHUDWLRQ RI
XSSHU�OD\HU FRUWLFDO QHXURQV� 2XU GDWD WKXV UHYHDO PROHFXODU GLIIHUHQFHV EHWZHHQ SURJHQLWRU
FHOOV ZLWK GLIIHUHQW QHXURJHQLF EHKDYLRU DW HDUO\ FRUWLFRJHQHVLV DQG LQGLFDWHV WKDW 6R[� LV FULWLFDO
IRU�WKH�PDLQWHQDQFH�RI�5*&V�WR�UHJXODWH�WKH�JHQHUDWLRQ RI�XSSHU�OD\HU�QHXURQV�
6,*1,),&$1&(�67$7(0(17
7KH H[LVWHQFH RI KHWHURJHQHLW\ LQ WKH SRRO RI 5*&V DQG LWV UHODWLRQVKLS ZLWK WKH JHQHUDWLRQ RI
FHOOXODU GLYHUVLW\ LQ WKH FHUHEUDO FRUWH[ KDV EHHQ DQ LQWHUHVWLQJ WRSLF RI GHEDWH IRU PDQ\ \HDUV�
+HUH ZH GHVFULEH WKH H[LVWHQFH RI 5*&V ZLWK UHGXFHG QHXURJHQLF EHKDYLRU DW HDUO\ HPEU\RQLF
DJHV SUHVHQWLQJ D SDUWLFXODU PROHFXODU VLJQDWXUH� 7KLV PROHFXODU VLJQDWXUH FRQVLVWV RI
GLIIHUHQWLDO H[SUHVVLRQ RI VRPH JHQHV LQFOXGLQJ WKH WUDQVFULSWLRQ IDFWRU 6R[�� IRXQG WR EH D
VSHFLILF PDVWHU UHJXODWRU RI WKLV VXESRSXODWLRQ RI SURJHQLWRU FHOOV� )XQFWLRQDO H[SHULPHQWV
SHUWXUELQJ 6R[� H[SUHVVLRQ¶V OHYHOV UHYHDO LWV LQVWUXFWLYH UROH LQ WKH UHJXODWLRQ RI WKH QHXURJHQLF
EHKDYLRU RI 5*&V DQG LWV UHODWLRQVKLS ZLWK WKH JHQHUDWLRQ RI XSSHU OD\HU SURMHFWLRQ QHXURQV DW
ODWHU�DJHV�
�

)DEUD�%HVHU�HW�DO�
,1752'8&7,21
7KH QHRFRUWH[ LV HYROXWLRQDULO\ WKH PRVW UHFHQW DGGLWLRQ WR WKH EUDLQ DQG H[HFXWHV FRPSOH[ WDVNV
LQFOXGLQJ VHQVRU\ LQWHJUDWLRQ DQG FRJQLWLRQ� 1HRFRUWLFDO GHYHORSPHQW GHSHQGV RQ SUHFLVHO\
RUFKHVWUDWHG PHFKDQLVPV UDQJLQJ IURP WKH JHQHUDWLRQ RI WKH QHXUDO FHOOV LQ VSHFLDOL]HG JHUPLQDO
]RQHV� WR WKHLU PLJUDWLRQ WR GHILQHG ORFDWLRQV DQG WKH HVWDEOLVKPHQW RI VSHFLILF FRQQHFWLRQV�
'XULQJ FRUWLFDO GHYHORSPHQW UDGLDO JOLD SURJHQLWRU FHOOV �5*&V� JHQHUDWH GLIIHUHQW VXEW\SHV RI
H[FLWDWRU\ QHXURQV WKDW ZLOO XOWLPDWHO\ IRUP WKH OD\HUV RI WKH FHUHEUDO FRUWH[ DQG PDNH
FRQQHFWLRQV WR GLVWLQFW VXEFRUWLFDO DQG FRUWLFDO WDUJHWV� 7KH PDWXUH FRUWH[ FRQVLVWV RI VL[
SULQFLSDO OD\HUV WKDW DUH JHQHUDWHG IROORZLQJ D WHPSRUDO VHTXHQFH� *OXWDPDWHUJLF QHXURQV WKDW
SRSXODWH GHHS OD\HUV RI WKH FRUWH[ DUH JHQHUDWHG ILUVW� IROORZHG E\ XSSHU OD\HU QHXURQV DQG
ILQDOO\�JOLDO�FHOOV��0RO\QHDX[�HW�DO���������
0XOWLSOH SURJHQLWRU VXEW\SHV ZLWK GLVWLQFW PRUSKRORJLHV DQG EHKDYLRUV KDYH EHHQ GHVFULEHG
LQ WKH QHRFRUWH[� 5*&V RFFXS\ WKH YHQWULFXODU ]RQH �9=� DQG DUH FRQQHFWHG WR WKH YHQWULFXODU
DQG SLDO VXUIDFHV RI WKH QHRFRUWH[ E\ DSLFDO DQG EDVDO SURFHVVHV� UHVSHFWLYHO\� ,QWHUPHGLDWH
SURJHQLWRU FHOOV �,3&V� DUH EDVDO SURJHQLWRUV JURXSHG LQ WKH VXEYHQWULFXODU ]RQH �69=� DQG DUH
PRVWO\ JHQHUDWHG E\ DV\PPHWULF GLYLVLRQV RI 5*&V �0L\DWD HW DO�� ����� 1RFWRU HW DO�� ������
2WKHU W\SHV RI SURJHQLWRU FHOOV KDYH EHHQ GHVFULEHG PRUH UHFHQWO\ LQFOXGLQJ VKRUW QHXUDO
SUHFXUVRUV �613� �*DO HW DO�� ����� DQG RXWHU UDGLDO JOLDO FHOOV �R5*&V� �)LHW] HW DO�� �����
+DQVHQ HW DO�� ����� 5HLOOR HW DO�� ������ R5*&V DUH H[SDQGHG LQ J\UHQFHSKDOLF DQLPDOV DQG DUH
WKRXJKW WR EH LPSRUWDQW IRU WKH H[SDQVLRQ RI QHRFRUWLFDO QHXURQV LQ SULPDWHV� HVSHFLDOO\ WKRVH
RFFXS\LQJ�WKH�HQODUJHG�XSSHU�FRUWLFDO�OD\HUV��/XL HW�DO���������/D0RQLFD�HW�DO���������
'HVSLWH WKH DGYDQFH RI NQRZOHGJH LQ WKH UHFHQW \HDUV LQ WKH LGHQWLILFDWLRQ RI PRUSKRORJLFDO
GLIIHUHQFHV DPRQJ QHRFRUWLFDO SURJHQLWRUV� WKH PHFKDQLVPV WKDW UHJXODWH QHXURQDO SURJHQLWRU
GLYHUVLILFDWLRQ DUH QRW ZHOO XQGHUVWRRG� ,Q IDFW� WKH H[WHQW RI QHRFRUWLFDO SURJHQLWRU
KHWHURJHQHLW\ LV VWLOO XQFOHDU DQG� LQ VSLWH RI WKH VLJQLILFDQW HIIRUW LQ WKH ODVW �� \HDUV WR
LQYHVWLJDWH FRUWLFDO GHYHORSPHQW� DQG LW LV VWLOO XQNQRZQ KRZ WKH SRSXODWLRQ RI QHRFRUWLFDO
SURJHQLWRUV LV DEOH WR JHQHUDWH WKH GLYHUVLW\ RI QHRFRUWLFDO FHOOV� 7KH WUDGLWLRQDO K\SRWKHVLV VWDWHV
�

)DEUD�%HVHU�HW�DO�
WKDW WKH IDWH RI SULQFLSDO QHXURQV LV GHWHUPLQHG E\ LWV ELUWKGDWH� VXJJHVWLQJ WKH H[LVWHQFH RI D
FRPPRQ 5*& SURJHQLWRU WKDW EHFRPHV SURJUHVVLYHO\ UHVWULFWHG ZLWK GHYHORSPHQWDO WLPH
�0F&RQQHOO DQG .D]QRZVNL� ����� )UDQW] DQG 0F&RQQHOO� ����� 'HVDL DQG 0F&RQQHOO� �����
6KHQ HW DO�� ����� *DR HW DO�� ������ +RZHYHU� UHFHQW VWXGLHV VXJJHVW WKDW WKH SRRO RI 5*&V LV
PRUH GLYHUVH� ZLWK VRPH 5*&V SULPDULO\ JHQHUDWLQJ FHUWDLQ W\SHV RI FHOOV LQFOXGLQJ XSSHU OD\HU
FRUWLFDO JOXWDPDWHUJLF QHXURQV DQG %� FHOOV �)UDQFR HW DO�� ����� )XHQWHDOED HW DO�� �����
*DUFtD�0RUHQR DQG 0ROQiU� ����� *LO�6DQ] HW DO�� ����� /ORUFD HW DO�� ������ 1HYHUWKHOHVV� WKH
PHFKDQLVPV WKDW PLJKW OHDG WR SURJHQLWRU GLYHUVLILFDWLRQ DUH XQFOHDU� DQG PROHFXODU PDUNHUV
WKDW�ZRXOG�GLVWLQJXLVK�GLVSDUDWH�SRSXODWLRQV�RI�5*&V DUH�ODUJHO\�XQNQRZQ�
7R LGHQWLI\ PROHFXODU GLIIHUHQFHV EHWZHHQ 5*& SURJHQLWRUV� ZH XVHG DQ LQ XWHUR
HOHFWURSRUDWLRQ DSSURDFK WR LQWURGXFH SODVPLG FRQVWUXFWV FRQWDLQLQJ GLIIHUHQW SURPRWHUV GULYLQJ
WKH H[SUHVVLRQ RI UHSRUWHU JHQHV� EDVHG RQ WKH SUHGLFWLRQ WKDW GLIIHUHQW SURPRWHUV PD\ EH DFWLYH
LQ GLVWLQFW QHRFRUWLFDO 5*& VXEW\SHV DV GHVFULEHG �*DUFtD�0RUHQR DQG 0ROQiU� ������ 8VLQJ
WKLV VWUDWHJ\� ZH LGHQWLILHG 5*&V ZLWK UHGXFHG QHXURJHQLF EHKDYLRU DW HDUO\ HPEU\RQLF DJHV WKDW
HYHQWXDOO\ SURGXFH SUHGRPLQDQWO\ FRUWLFR�FRUWLFDO SURMHFWLRQ QHXURQV RFFXS\LQJ XSSHU
QHRFRUWLFDO FHOO OD\HUV� 8VLQJ VLQJOH FHOO 51$ VHTXHQFLQJ �VF51$ VHT�� ZH FKDUDFWHUL]HG WKH
PROHFXODU VLJQDWXUH RI WKHVH 5*& SURJHQLWRUV DQG LGHQWLILHG E\ WUDQVFULSWLRQDO UHJXODWRU\
QHWZRUN DQDO\VLV WKDW WKH\ H[SUHVV 6R[� DW VLJQLILFDQWO\ KLJKHU OHYHOV FRPSDUHG WR RWKHU 5*&V�
1RWDEO\� 6R[� LV D WUDQVFULSWLRQ IDFWRU LQYROYHG LQ VWHP FHOO PDLQWHQDQFH �6FRWW HW DO�� ����� /LX
HW DO�� ����� DQG DVVRFLDWHG ZLWK SURJHQLWRU TXLHVFHQFH LQVLGH DQG RXWVLGH WKH QHUYRXV V\VWHP
�.DGDMD HW DO�� ����� /ORUHQV�%REDGLOOD HW DO�� ������ 8VLQJ IXQFWLRQDO SHUWXUEDWLRQV� ZH
GHPRQVWUDWH WKDW 6R[� H[SUHVVLRQ OHYHOV DUH FULWLFDO WR FRQWURO FHOO F\FOH GXUDWLRQ RI D VXEVHW RI
5*&V WR VSDUH�UHVHUYH WKHP VLQFH HDUO\ VWDJHV RI FRUWLFRJHQHVLV IRU WKH SURGXFWLRQ RI XSSHU
OD\HU�QHRFRUWLFDO�QHXURQV�DW�ODWHU�GHYHORSPHQWDO�VWDJHV�
�

)DEUD�%HVHU�HW�DO�
0$7(5,$/6�$1'�0(7+2'6
0LFH
$OO H[SHULPHQWV LQYROYLQJ PLFH ZHUH FRQGXFWHG LQ DFFRUGDQFH ZLWK WKH 6SDQLVK OHJLVODWLRQ DV
ZHOO DV ZLWK WKH 'LUHFWLYH ��������(8 RI WKH (XURSHDQ OHJLVODWLRQ DQG WKH 8QLWHG 6WDWHV RI
$PHULFD DQLPDO ZHOIDUH UHJXODWLRQV DQG ZHUH DSSURYHG E\ WKH HWKLFDO FRPPLWWHH RI WKH
8QLYHUVLW\ RI 9DOHQFLD DQG WKH &RQVHOOHULD GH $JULFXOWXUD� 'HVDUUROOR 5XUDO� (PHUJHQFLD
&OLPiWLFD \ 7UDQVLFLyQ (FROyJLFD RI WKH &RPXQLGDG 9DOHQFLDQD DQG ,QVWLWXWLRQDO $QLPDO &DUH
DQG�8VH�&RPPLWWHH��,$&8&��RI�WKH�86$�
&��%/�� ZLOG�W\SH �:7� DQG $L� &UH UHSRUWHU VWUDLQV �0DGLVHQ HW DO� ����� XVHG IRU
H[SHULPHQWDWLRQ ZHUH RULJLQDOO\ REWDLQHG IURP WKH -DFNVRQ ODERUDWRU\ DQG DUH FXUUHQWO\ EUHG LQ
WKH DQLPDO IDFLOLWLHV RI WKH 8QLYHUVLW\ RI 9DOHQFLD� SODFHG LQ WKH %XUMDVVRW FDPSXV� %RWK IHPDOH
DQG PDOH PLFH ZHUH DQDO\VHG LQ WKLV VWXG\� ,QGXFWLRQ RI UHFRPELQDWLRQ LQ $L� PLFH
HOHFWURSRUDWHG ZLWK &UH LQGXFLEOH SODVPLGV ZDV DFKLHYHG E\ LQWUDSHULWRQHDO LQMHFWLRQ RI
SUHJQDQW GDPV ZLWK WKH IDVWHU�DFWLQJ DQG VKRUWHU�OLYHG WDPR[LIHQ PHWDEROLWH
��K\GUR[\�WDPR[LIHQWDPR[LIHQ ���2+7� �6LJPD� �� PJ��� J RI ERG\ ZHLJKW� GLVVROYHG DV
GHVFULEHG��*XHQWKQHU�HW�DO����������
([SUHVVLRQ�&RQVWUXFWV
+HV�S�GV5HG �$GGJHQH SODVPLG ������� ZDV JHQHUDWHG E\ 1LFKRODV *DLDQR �0L]XWDQL HW DO��
������ +HV�S�*)3 ZDV JHQHUDWHG E\ UHSODFLQJ GV5HG� ZLWK H*)3 LQ +HV�S�'V5HG �)UDQFR HW
DO�� ������ S&$*�P5)3� �$GGJHQH SODVPLG ������� ZDV GHSRVLWHG E\ $QQD�.DWHULQD
+DGMDQWRQDNLV �/RQJ HW DO�� ������ S&$*�P5)3�+HV�S�*)3 ZDV JHQHUDWHG E\ VXEFORQLQJ
S&$*�P5)3 DQG D SRO\�DGHQ\ODWLRQ VLJQDO LQWR +HV�S�*)3� S&$*�&UH(57� �$GGJHQH
SODVPLG ������� ZDV GHSRVLWHG E\ &RQQLH &HSNR �0DWVXGD DQG &HSNR� ������ +HV�S�&UH(57�
ZDV JHQHUDWHG E\ UHSODFLQJ S&$* ZLWK +HV�S� S&,* �$GGJHQH SODVPLG ������� ZDV GHSRVLWHG
E\ &RQQLH &HSNR �0DWVXGD DQG &HSNR� ������ &%)5(�*)3 �$GGJHQH SODVPLG ������� ZDV
GHSRVLWHG E\ 1LFKRODV *DLDQR �0L]XWDQL HW DO�� ������ 723�*)3 �$GGJHQH SODVPLG �������
ZDV GHSRVLWHG E\ 5DPHVK 6KLYGDVDQL �+RUVW HW DO�� ������ (0;�S�*)3 ZDV JHQHUDWHG
�

)DEUD�%HVHU�HW�DO�
DFFRUGLQJ WR WKH SXEOLFDWLRQ RI *DUFtD�0RUHQR DQG =ROWiQ 0ROQiU ������� S&,*�6�)/ ZDV
JHQHUDWHG E\ FORQLQJ 6R[� FRGLQJ VHTXHQFH LQWR S&,*� &5�6K51$ �S/.2���VK6&5� $GGJHQH
SODVPLG ������� ZDV GHSRVLWHG E\ 6KHLOD 6WHZDUW �6DKDULD HW DO�� ������ 6��6K51$
�S/.2���VK�P62;���� $GGJHQH SODVPLG ������� ZDV GHSRVLWHG E\ %RE :HLQEHUJ �*XR HW DO��
������
,Q�8WHUR (OHFWURSRUDWLRQ
,Q XWHUR HOHFWURSRUDWLRQ ZDV SHUIRUPHG DV GHVFULEHG �*LO�6DQ] HW DO�� ������ %ULHIO\ WLPHG
SUHJQDQW PLFH ZHUH DQHVWKHWL]HG ZLWK LVRIOXUDQH� 2QFH WKH SHGDO UHIOH[ ZDV ORVW� DQDOJHVLF
VROXWLRQ ZDV LQMHFWHG VXEFXWDQHRXVO\� 7KH DEGRPLQDO DUHD ZDV VKDYHG� DQG DQ LQFLVLRQ LQ WKH
PLFHV DEGRPHQ ZDV SHUIRUPHG WR H[SRVH WKH XWHULQH KRUQV� ����� �/ RI HQGRWR[LQ�IUHH SODVPLG
'1$ VROXWLRQ �a� �J��/� ZDV LQMHFWHG LQWR RQH RI WKH HPEU\RV¶ ODWHUDO YHQWULFOHV� )RU
HOHFWURSRUDWLRQ RI HPEU\RQLF GD\ �(� ���� HPEU\RV� ILYH SXOVHV RI �� 9 ZHUH GHOLYHUHG� VSDFHG
E\ ���PV� $IWHU HOHFWURSRUDWLRQ� WKH XWHUXV ZDV SODFHG LQ WKH DEGRPLQDO FDYLW\� WKH DEGRPLQDO
ZDOO DQG VNLQ ZHUH VXWXUHG� (PEU\RV ZHUH OHW WR GHYHORS LQ XWHUR IRU WKH LQGLFDWHG WLPH RU
IHPDOHV�ZHUH�DOORZHG�WR�JLYH�ELUWK�DQG�SXSV�ZHUH�VDFULILFHG DW�WKH�LQGLFDWHG�DJH�
,Q�8WHUR )ODVKWDJ�,QMHFWLRQ
&DUER[\IOXRUHVFHLQ VXFFLQLPLG\O HVWHU RU )ODVK7DJ �)7� LQMHFWLRQV ZHUH SHUIRUPHG DW (���� DV
GHVFULEHG �*RYLQGDQ HW DO�� ������ RQ SUHYLRXVO\ HOHFWURSRUDWHG HPEU\RV DW (����� $V GHVFULEHG
IRU LQ XWHUR HOHFWURSRUDWLRQ� (���� SUHJQDQW IHPDOHV ZHUH DQHVWKHWL]HG ZLWK LVRIOXUDQH� DQDOJHVLF
VROXWLRQ ZDV LQMHFWHG� DQG DEGRPLQDO LQFLVLRQ ZDV SHUIRUPHG WR H[SRVH WKH XWHULQH KRUQV� ���
�/ RI ��P0 )7 �&HOO7UDFH70 &)6(� /LIH 7HFKQRORJLHV� �&������ ZDV LQMHFWHG LQWR RQH RI
WKH HPEU\RV¶ ODWHUDO YHQWULFOHV� 7KH XWHUR ZDV WKHQ SODFHG LQ WKH DEGRPLQDO FDYLW\� WKH
DEGRPLQDO ZDOO VXWXUHG� DQG HPEU\RV ZHUH DOORZHG WR GHYHORS LQ XWHUR XQWLO FROOHFWLRQ WLPH DW
(�����
�

)DEUD�%HVHU�HW�DO�
,PPXQRKLVWRFKHPLVWU\
7LVVXH SURFHVVLQJ� (PEU\RQLF EUDLQV ZHUH GLVVHFWHG LQ D SKRVSKDWH�EXIIHUHG VDOLQH VROXWLRQ
�3%6�� IL[HG LQ �� SDUDIRUPDOGHK\GH �3)$� LQ 3%6 RYHUQLJKW DW ��&� DQG ODWHU VWRUHG LQ
3%6������ VRGLXP D]LGH DW ��&� $IWHU HPEHGGLQJ LQ �� ORZ PHOWLQJ SRLQW DJDURVH LQ 3%6�
EUDLQV ZHUH VHFWLRQHG FRURQDOO\ DW ��� �P XVLQJ D YLEUDWRPH �/HLFD 97����6�� 3RVWQDWDO PLFH
ZHUH WUDQVFDUGLDOO\ SHUIXVHG ZLWK �� 3)$ EHIRUH GLVVHFWLRQ DQG SRVW�IL[DWLRQ� 9LEUDWRPH
FRURQDO�VHFWLRQV�ZHUH�SHUIRUPHG�REWDLQLQJ�����P�VHFWLRQV�
,PPXQRIOXRUHVFHQFH RQ EUDLQ VHFWLRQV� %UDLQ VHFWLRQV ZHUH LQFXEDWHG IRU � KRXU LQ EORFNLQJ
VROXWLRQ ���� KRUVH VHUXP� ���� 7ULWRQ ;���� LQ 3%6� DW URRP WHPSHUDWXUH� 6HFWLRQV ZHUH
LQFXEDWHG RYHUQLJKW ZLWK WKH SULPDU\ DQWLERG\ DW WKH DSSURSULDWHG GLOXWLRQ LQ EORFNLQJ VROXWLRQ
DW ��&� 6HFWLRQV ZHUH ZDVKHG WKUHH WLPHV ZLWK 3%6 DQG LQFXEDWHG IRU � KRXU ZLWK WKH UHVSHFWLYH
VHFRQGDU\ DQWLERGLHV �������� LQ EORFNLQJ VROXWLRQ DW URRP WHPSHUDWXUH� $IWHU WKUHH ZDVKHV LQ
3%6��VHFWLRQV�ZHUH�PRXQWHG�RQ�VOLGHV�ZLWK�)OXRURPRXQW *��(OHFWURQ�0LFURVFRS\�6FLHQFHV��
$QWLERGLHV� UDEELW DQWL�%URPRGHR[\XULGLQH �������� 0HJDEDVH 5HVHDUFK 3URGXFWV� �������� UDW
DQWL�&WLS� ������� $EFDP� �DE������� UDEELW DQWL�&X[� ������� 6DQWD &UX] %LRWHFKQRORJ\�
VF�������� UDEELW DQWL�&X[� �������� �3URWHLQWHFK� ���������$3U�� UDEELW DQWL�'RXEOHFRUWLQ
�'&;� �������� $EFDP� �DE������� JRDW DQWL�*)3 ������� 5RFNODQG DQWLERGLHV DVVD\V�
������������0�� UDEELW DQWL�.L�� ������� (0'� �$%������ JRDW DQWL�0FP� ������� 6DQWD
&UX] %LRWHFKQRORJ\� VF������� UDEELW DQWL�3D[� ������� *HQH7H[� �*7;�������� PRXVH
DQWL�3KRVSKR 9LPHQWLQ ������� 0%/� '������� PRXVH DQWL�5)3 ������� /6%LR� /6�&�������
UDEELW DQWL�6DWE� �������� $EFDP� �DE������� UDEELW DQWL�6R[� �������� $EFDP� �$%�������
UDEELW DQWL�7OH� ������� $EFDP� �DE�������� UDEELW DQWL�7EU� �(RPHV� ������� $EFDP�
�$%��������)OXRUHVFHQW�VHFRQGDU\�DQWLERGLHV�ZHUH�XVHG DW���������-DFNVRQ�OPPXQRUHVHDUFK��
,PDJLQJ� ,PDJHV ZHUH DFTXLUHG XVLQJ D 1LNRQ�&� RU D )OXR9LHZ )9��L �2O\PSXV� FRQIRFDO
ODVHU�VFDQQLQJ PLFURVFRSH� ,PDJH DQDO\VLV DQG TXDQWLILFDWLRQ ZHUH SHUIRUPHG XVLQJ 3KRWRVKRS
�$GREH���),-,��,PDJH-��DQG�5�VRIWZDUH�
�

)DEUD�%HVHU�HW�DO�
4XDQWLILFDWLRQ�RI�7LVVXH�6HFWLRQV
)RXU GLIIHUHQW DQLPDOV ZHUH XVHG IRU PRVW RI WKH FRQGLWLRQV ZLWK D FRXSOH RI H[FHSWLRQV ZKHUH
WKUHH DQLPDOV ZKHUH DQDO\VHG� 6HYHUDO URVWUR�FDXGDO VHFWLRQV ZHUH VHOHFWHG EDVHG RQ WKH
HOHFWURSRUDWLRQ ORFDWLRQ� OLPLWLQJ WKH XVH RI HOHFWURSRUDWLRQV WR WKH ODWHUDO QHRFRUWH[ LQFOXGLQJ
WKH�VRPDWRVHQVRU\�QHRFRUWH[�
$QDO\VLV RI WKH GHYHORSLQJ QHRFRUWH[ DW (����� 4XDQWLILFDWLRQV LQ WKH GHYHORSLQJ QHRFRUWH[
ZHUH SHUIRUPHG E\ GLYLGLQJ LWV VXUIDFH LQ WZR UHJLRQV� 9=� DQG RXWVLGH WKH 9=� �LQFOXGLQJ WKH
VXEYHQWULFXODU ]RQH �69=�� LQWHUPHGLDWH ]RQH �,=� DQG FRUWLFDO SODWH �&3��� 'HQVLW\ DQG
PRUSKRORJLFDO�IHDWXUHV�ZHUH�XVHG�WR�GLIIHUHQWLDWH WKHVH�UHJLRQV�
$QDO\VLV RI WKH YHQWULFXODU ]RQH DW (����� 4XDQWLILFDWLRQV ZHUH SHUIRUPHG E\ GHOLPLWLQJ WKH 9=
XVLQJ FHOOXODU GHQVLW\ DQG PRUSKRORJLFDO IHDWXUHV� $GGLWLRQDOO\� IRU FHOO GLVWULEXWLRQ DQDO\VLV� WKH
9= ZDV GLYLGHG LQ WZR HTXDOO\�VL]HG UHJLRQV� WKH LQQHU YHQWULFXODU ]RQH �,9=�� KDOI LQ FRQWDFW
ZLWK�WKH�YHQWULFOH¶V�VXUIDFH��DQG�WKH�RXWHU�YHQWULFXODU ]RQH��29=���KDOI�LQ�FRQWDFW�ZLWK�WKH�69=�
$QDO\VLV RI WKH GHYHORSLQJ QHRFRUWH[ DW (����� 4XDQWLILFDWLRQV LQ WKH GHYHORSLQJ QHRFRUWH[
ZHUH SHUIRUPHG E\ GLYLGLQJ LWV VXUIDFH LQ IRXU UHJLRQV� 9=� 69=� ,= DQG &3� &HOOXODU GHQVLW\
DQG PRUSKRORJLFDO IHDWXUHV ZHUH XVHG WR GLIIHUHQWLDWH WKHVH UHJLRQV� )XUWKHUPRUH�
TXDQWLILFDWLRQV XVLQJ FHOO W\SH PDUNHUV 3D[�� 7EU� DQG 'F[ ZHUH FRQGXFWHG� 4XDQWLILFDWLRQV
UHJDUGLQJ�FHOO�F\FOH�OHQJWK�ZHUH�SHUIRUPHG�LQ�WKH YHQWULFXODU�]RQH�
$QDO\VLV RI WKH FRUWLFDO SODWH DW (���� DQG 3�� 4XDQWLILFDWLRQV ZHUH SHUIRUPHG E\ GLYLGLQJ WKH
FRUWLFDO SODWH LQ WHQ HTXDOO\�VL]HG UHJLRQV� UHIHUUHG DV ELQV� 7KH ELQ GLYLVLRQ ZDV XVHG VLQFH WKH
FRUWLFDO�SODWH�LV�VWLOO�XQPDWXUHG�DQG�VRPH�QHXURQV DUH�VWLOO�PLJUDWLQJ�DW�WKLV�VWDJH�
$QDO\VLV RI WKH FRUWLFDO SODWH DW 3�� DQG 3��� 4XDQWLILFDWLRQV RI WKH FRUWLFDO SODWH ZHUH
SHUIRUPHG E\ GLYLGLQJ WKH QHRFRUWH[ VXUIDFH LQ WZR UHJLRQV� XSSHU OD\HUV �8/� DQG ORZHU OD\HUV
�//�� ,Q RUGHU WR GLIIHUHQWLDWH WKHVH WZR UHJLRQV� LPPXQRFKHPLVWU\ ZLWK &X[� DQWLERG\ ZDV
XVHG�WR�GHILQH�WKH�8/��,,�,9��
�

)DEUD�%HVHU�HW�DO�
7LVVXH�3UHSDUDWLRQ�IRU�6LQJOH�&HOO�6RUWLQJ�DQG�6HTXHQFLQJ
%UDLQV HOHFWURSRUDWHG ZLWK WKH &$*�P5)3�+HV��*)3 SODVPLG ZHUH GLVVHFWHG �� K DIWHU
HOHFWURSRUDWLRQ DQG HPEHGGHG LQ �� $JDURVH LQ FRPSOHWH +%66 PHGLXP �3ROOHX[ DQG *KRVK�
����� DQG VHFWLRQHG XVLQJ D YLEUDWRPH �/HLFD 97����6�� (OHFWURSRUDWHG DUHDV RI WKH ODWHUDO
FRUWH[ ZHUH PLFURGLVVHFWHG XQGHU WKH IOXRUHVFHQW GLVVHFWLQJ VFRSH �=HLVV $[LR =RRP�� 7LVVXH
ZDV GLVVRFLDWHG LQWR VLQJOH FHOOV XVLQJ WKH 3DSDLQ 'LVVRFLDWLRQ 6\VWHP �:RUWKLQJWRQ�
�/.��������� 5HVXVSHQGHG FHOOV ZHUH SURFHVVHG XVLQJ D FHOO VRUWHU �0R)OR ;'3� DQG YLDEOH
VLQJOH FHOOV H[SUHVVLQJ P5)3 �&$*�� RU P5)3 DQG *)3 �+HV��� ZHUH LQGLYLGXDOO\ FROOHFWHG
LQ �� ZHOO SODWHV ZLWK O\VLV EXIIHU� ,QGLYLGXDO O\VHG FROOHFWHG FHOOV ZHUH SURFHVVHG IRU VF51$VHT
XVLQJ D PRGLILHG 6PDUW�VHT� SURWRFRO �&KHYpH HW DO�� ������ 'DWD ZHUH DOLJQHG WR PRXVH
UHIHUHQFH JHQRPH �PP��� XVLQJ +LVDW� �.LP HW DO�� ����� DQG TXDQWLILHG DJDLQVW D PRGLILHG
UHIHUHQFH WUDQVFULSWRPH �*(1&2'( Y0��� �)UDQNLVK HW DO�� ����� ZLWK FXIITXDQW �7UDSQHOO HW
DO�� ������ 'HIDXOW SDUDPHWHUV ZHUH VHW IRU ERWK VRIWZDUH H[FHSW SDUDOOHOL]DWLRQ YDOXH ��S �� WR
LPSURYH�SURFHVVLQJ�WLPH�
VF51$VHT�$QDO\VLV
'DWD ZHUH DQDO\]HG XVLQJ 6HXUDW 5 SDFNDJH Y� �%XWOHU HW DO�� ������ 7R LPSRUW GDWD LQWR D 6HXUDW
REMHFW DQG HOLPLQDWH ORZ TXDOLW\ FHOOV�JHQHV �TXDOLW\�FKHFN�� &UHDWH6HXUDW2EMHFW IXQFWLRQ
VHOHFWHG FHOOV ZLWK DW OHDVW ��� JHQHV� DQG JHQHV WKDW ZHUH H[SUHVVHG LQ DW OHDVW � FHOOV� $
JOREDO�VFDOLQJ QRUPDOL]DWLRQ PHWKRG �/RJ1RUPDOL]H E\ ��� IDFWRU� ZDV LPSOHPHQWHG XVLQJ
1RUPDOL]H'DWD� +LJKO\ YDULDEOH JHQHV ZHUH GHWHFWHG �)LQG9DULDEOH*HQHV� DFURVV WKH VLQJOH FHOOV
DQG GDWD ZHUH VFDOHG �6FDOH'DWD� WR OLQHDU WUDQVIRUP RXU GDWD DQG UHJUHVV RXW YDULDEOHV WKDW
ZRXOG QRW JLYH PHDQLQJIXO LQIRUPDWLRQ WR RXU DQDO\VLV VXFK DV D KLJK SHUFHQWDJH RI
PLWRFKRQGULDO JHQHV H[SUHVVLRQ DQG QXPEHU RI PROHFXOHV GHWHFWHG� 5XQ3&$ ZHUH XVHG WR
SHUIRUP OLQHDU GLPHQVLRQDO UHGXFWLRQ DQG DPRQJ WKRVH 3&$V� �� �EDVHG RQ 3&(OERZ3ORW� ZDV
VHOHFWHG WR IXUWKHU DQDO\VLV� &OXVWHUV DQG W61( ��GLPHQVLRQDO YLVXDOL]DWLRQ ZHUH EXLOW XVLQJ
3&$V ZLWK )LQG&OXVWHUV DQG 5XQ761( IXQFWLRQV� UHVSHFWLYHO\� )LQG$OO0DUNHUV ZHUH XVHG WR
LGHQWLI\ GLIIHUHQWLDOO\ H[SUHVVHG JHQHV �'(*V� LQ HDFK JURXS DQG FOXVWHUV ZDV UHQDPHG EDVHG RQ
�

)DEUD�%HVHU�HW�DO�
H[SUHVVLRQ RI LGHQWLILHG NQRZQ FHOO PDUNHUV� 6WDWLVWLFDOO\ VLJQLILFDQW '(*V ZHUH LGHQWLILHG XVLQJ
WKH QRQ�SDUDPHWULF :LOFR[RQ UDQN VXP WHVW �VLJQLILFDQFH WKUHVKROGV� ORJ� IROG FKDQJH ! ��� DQG
S�DGMXVWHG�YDOXH���������
7UDQVFULSWLRQDO�5HJXODWRU\�1HWZRUNV
:H XVHG WKH 5HFRQVWUXFWLRQ RI 7UDQVFULSWLRQDO 1HWZRUNV �571� SDFNDJH �&DVWUR HW DO�� ����� WR
JHQHUDWH D WUDQVFULSWLRQDO UHJXODWRU\ QHWZRUN EDVHG RQ VF51$VHT GDWD JHQHUDWHG IURP WKH
PRXVH GRUVDO WHOHQFHSKDORQ DW (�� �<X]ZD HW DO�� ������ 7KLV QHWZRUN GHILQHV UHJXORQV
�SRVVLEOH WDUJHW JHQHV� IRU D VHW RI FXUDWHG WUDQVFULSWLRQ IDFWRUV� 7KH UHJXORQ IRU HDFK
WUDQVFULSWLRQ IDFWRU �7)� LV FRPSRVHG RI DOO WKH JHQHV ZKRVH H[SUHVVLRQ GDWD GLVSOD\
VLJQLILFDQW PXWXDO LQIRUPDWLRQ ZLWK WKDW RI D JLYHQ WUDQVFULSWLRQ IDFWRU DQG DUH WKHUHIRUH
OLNHO\ WR EH UHJXODWHG E\ WKDW WUDQVFULSWLRQ IDFWRU �&DVWUR HW DO�� ����� <X]ZD HW DO�� ������
7R VHOHFW PRXVH WUDQVFULSWLRQ IDFWRUV �7)V�� D WUDQVFULSWLRQ IDFWRU GDWDEDVH �7FR)�'% Y��
�6FKPHLHU HW DO�� ����� ZDV XVHG WR LGHQWLI\ 7)V LQ RXU GDWD� 7KHQ� 71,�FODVV REMHFW ZDV FUHDWHG
XVLQJ SUHYLRXV GHVFULEHG LQSXWV �QRUPDOL]HG WDEOH DQG VHOHFWHG 7)V� �WQL�FRQVWUXFWRU�� IROORZHG
E\ SHUPXWDWLRQ DQDO\VLV �WQL�SHUPXWDWLRQ� Q3HUPXWDWLRQV ����� DQG ERRWVWUDS �WQL�ERRWVWUDS��
)LQDOO\� GLIIHUHQWLDOO\ H[SUHVVHG JHQHV IURP HDFK FOXVWHU DQG WKHLU UHVSHFWLYH DYHUDJH ORJ�
IROG�FKDQJH DYHUDJH H[SUHVVLRQ ZHUH XVHG WR ILQG PDVWHU UHJXORQV �WQL�PUD�� 5HVXOWLQJ UHJXORQ
QHWZRUN�ZHUH�PDQLSXODWHG�XVLQJ�5HGH5�SDFNDJH���&DVWUR HW�DO��������
0HDQ�RI�)OXRUHVFHQFH�,QWHQVLW\�RI�62;�
,PPXQRVWDLQLQJ ZLWK 6R[� DQWLERG\ �������� $EFDP� �$%������� ZDV SHUIRUPHG RQ VOLFHV
IURP (���� &$*�P5)3�+HV��*)3 HOHFWURSRUDWHG HPEU\RV� FROOHFWHG ��K SRVW�HOHFWURSRUDWLRQ
�(������� ,PDJHV ZHUH REWDLQHG XVLQJ D )OXR9LHZ )9��L �2O\PSXV� FRQIRFDO ODVHU�VFDQQLQJ
PLFURVFRSH DQG DQDO\]HG XVLQJ ),-, �)LML ,V -XVW ,PDJH-�� 0HDQ RI )OXRUHVFHQFH ,QWHQVLW\ �0),�
ZDV PHDVXUHG IRU P5)3�*)3� �+HV��� DQG P5)3�*)3� �&$*�� FHOOV LQ WKH 9= ZLWK 5*&V
PRUSKRORJ\ E\ VHOHFWLQJ HDFK QXFOHXV XVLQJ WKH IUHHKDQG VHOHFWLRQ WRRO� &HOOV IURP DW OHDVW WZR
��

)DEUD�%HVHU�HW�DO�
VHFWLRQV IURP IRXU GLIIHUHQW DQLPDOV ZHUH XVHG� 0), ZDV PHDVXUHG LQ D VFDOH RI ����� DUELWUDU\
XQLWV �D�X�� 0), IRU HYHU\ FRXQWHG FHOO ZDV FRUUHFWHG E\ LWV SDUWLFXODU EDFNJURXQG YDOXH�
PHDVXUHG IRU HYHU\ FRXQWHG VHFWLRQ� >� � �������LNUU�RW]NW\R]bWa
�KJLTP[X^WMa
KJLTP[X^WMa
�@ ����
%DU JUDSK UHSUHVHQWV WKH PHDQ RI 0), FDOFXODWHG IRU HDFK DQLPDO� 'LVWULEXWLRQ RI 6R[� ,QWHQVLW\
RI )OXRUHVFHQFH JUDSK UHSUHVHQWV WKH IUHTXHQF\ LQ SHUFHQWDJH RI FHOOV UHJDUGLQJ WKH LQWHQVLW\ RI
IOXRUHVFHQFH�LQ�DUELWUDU\�XQLWV��D�X���
&HOO�&\FOH�/HQJWK�$QDO\VLV
���PLQ (G8 SXOVH� ,Q XWHUR HOHFWURSRUDWLRQ ZDV SHUIRUPHG DW (���� XVLQJ 3&,* DQG 3&,*6�)/
FRQVWUXFWV� �� K SRVW�HOHFWURSRUDWLRQ �(������� SUHJQDQW GDPV ZHUH LQMHFWHG ZLWK ��PJ�NJ RI
��HWLQLO���GHVR[LXULGLQD �(G8�� (PEU\RV ZHUH FROOHFWHG �� PLQ SRVW�(G8 LQMHFWLRQ� %UDLQ
VHFWLRQV�ZHUH�VWDLQHG�IRU�(G8��&OLFN�LW�3OXV��(G8� 7KHUPR�)LVKHU�6FLHQWLILF��
���KRXU %UG8 SXOVH� ,Q XWHUR HOHFWURSRUDWLRQ ZDV SHUIRUPHG DW (���� XVLQJ 3&,* DQG
3&,*6�)/ FRQVWUXFWV� ��K SRVW�HOHFWURSRUDWLRQ� SUHJQDQW GDPV ZHUH LQMHFWHG ZLWK ��PJ�NJ RI
%UG8��(PEU\RV�ZHUH�FROOHFWHG���K�SRVW�%UG8�LQMHFWLRQ� %UDLQ�VHFWLRQV�ZHUH�VWDLQHG�IRU�%UG8�
'XDO�SXOVH ODEHOLQJ� 7KH PHDQ RI WRWDO FHOO F\FOH �7F� DQG 6�SKDVH GXUDWLRQ �7V� ZDV FDOFXODWHG
IRU (���� FHOOV LQ WKH 9= XVLQJ D GXDO�SXOVH ODEHOLQJ VWUDWHJ\ PRGLILHG IURP �+DUULV HW DO��
������ ,Q EULHI� LQ XWHUR HOHFWURSRUDWLRQ ZDV SHUIRUPHG DW (���� XVLQJ 3&,* DQG 3&,*6�)/
FRQVWUXFWV� �� K SRVW�HOHFWURSRUDWLRQ� SUHJQDQW GDPV ZHUH LQMHFWHG ZLWK ��PJ�NJ RI %UG8� DQG �
K ODWHU ZLWK ��PJ�NJ RI (G8� ��� K DIWHU (G8 LQMHFWLRQ� HPEU\RV ZHUH FROOHFWHG DQG
LPPHUVLRQ�IL[HG LQ �� 3)$� %UDLQ VHFWLRQV ZHUH VWDLQHG IRU %UG8 DQG (G8� )RU TXDQWLILFDWLRQ
ZH LGHQWLILHG *)3� FHOOV ORFDWHG LQ WKH 9= ZLWK 5*& PRUSKRORJ\� 7KH 7V ZDV FDOFXODWHG DV
WKH IUDFWLRQ RI FHOOV WKDW UHPDLQ LQ 6�SKDVH �%UG8�(G8�� WR WKH FHOOV WKDW OHDYH 6�SKDVH EHIRUH
(G8 LQMHFWLRQ �%UG8�(G8��� PXOWLSOLHG E\ WKH LQMHFWLRQ LQWHUYDO �� K�� 7KH 7F ZDV FDOFXODWHG DV
WKH 7V GLYLGHG E\ WKH IUDFWLRQ RI FHOOV LQ WKH 9= WKDW DUH LQ 6�SKDVH �(G8���*)3��� FRUUHFWHG E\
SURSRUWLRQ�RI�(3�FHOO�LQ�WKH�9=�WKDW�DUH�LQ�FHOO�F\FOH� UHIHUUHG�DV�WKH�JURZWK�IUDFWLRQ��*)��
��

)DEUD�%HVHU�HW�DO�
C\ g1[MD
`4MD
`
1[MD`4MD
a] ����CL g �
C\
4MD`
65?`c65
] ���65 g �
65?`0W]RKXMb
`
65?`
([SHULPHQWDO�'HVLJQ�DQG�6WDWLVWLFDO�$QDO\VLV
0HDQ� VWDQGDUG GHYLDWLRQ �6'�� VWDQGDUG HUURU RI WKH PHDQ �6(0� DQG VWDWLVWLFDO DQDO\VLV ZHUH
SHUIRUPHG XVLQJ ([FHO IURP 0LFURVRIW� 6WDWLVWLFDOO\ VLJQLILFDQW GLIIHUHQFHV RI PHDQV ZHUH
DVVHVVHG E\ 6WXGHQW¶V XQSDLUHG W�WHVW FRPSDULQJ WZR JURXSV� 3�YDOXH � ���� ZDV FRQVLGHUHG D
VLJQLILFDQW GLIIHUHQFH � �� 7KH H[DFWV S YDOXH� PHDQ � 6(0 DQG Q DUH UHSRUWHG LQ WKH UHVXOWV
VHFWLRQ�
��

)DEUD�%HVHU�HW�DO�
5(68/76
,Q 8WHUR (OHFWURSRUDWLRQ 8VLQJ 'LIIHUHQW 3URPRWHUV DV D 6WUDWHJ\ WR 'LVWLQJXLVK
3URJHQLWRUV�ZLWK�'LIIHUHQW�1HXURJHQLF�%HKDYLRU
,W KDV EHHQ UHSRUWHG WKDW 5*&V WDUJHWHG ZLWK FRQVWUXFWV XVLQJ WKH (P[� HQKDQFHU�SURPRWHU DW
(���� VKRZHG GHOD\HG QHXURJHQLF GLYLVLRQV FRPSDUHG WR 5*&V WDUJHWHG ZLWK XELTXLWRXV
SURPRWHUV �*DUFtD�0RUHQR DQG 0ROQiU� ������ :H XVHG WKLV VWUDWHJ\ WR LGHQWLI\ SURJHQLWRUV
ZLWK GLIIHUHQW QHXURJHQLF EHKDYLRU DQG ZH FRQGXFWHG WKH H[SHULPHQWV DW WKH VDPH HPEU\RQLF
DJH �(����� UHSRUWHG LQ WKH HDUOLHU VWXG\� :H VHOHFWHG H[SUHVVLRQ YHFWRUV FRQWDLQLQJ SURPRWHU
IUDJPHQWV GULYLQJ IOXRUHVFHQW SURWHLQV WKDW UHSRUW WKH DFWLYLW\ RI LPSRUWDQW PROHFXODU SDWKZD\V
LQYROYHG LQ GHYHORSPHQW RI WKH FHQWUDO QHUYRXV V\VWHP LQFOXGLQJ :QW VLJQDOLQJ �+LUDED\DVKL HW
DO�� ����� &KHQQ� ����� DQG 1RWFK VLJQDOLQJ �<RRQ DQG *DLDQR� ����� /RXYL DQG
$UWDYDQLV�7VDNRQDV� ������ :H FRPSDUHG WKH ODEHOLQJ REVHUYHG XVLQJ WKHVH SURPRWHUV WR WKDW RI
WKH JHQHUDO SURPRWHU &$* GULYLQJ %)3 �)LJ� �$�� 7R UHSRUW :QW VLJQDOLQJ ZH XVHG WKH
7RS�*)3 SODVPLG FRQWDLQLQJ D WDQGHP RI � 7&)�/() SURPRWHU HOHPHQWV��+RUVW HW DO�� ������ 7R
UHSRUW 1RWFK DFWLYLW\� ZH VHOHFWHG D SODVPLG FRQWDLQLQJ WKH +HV� SURPRWHU IUDJPHQW GULYLQJ
GV5HG SURWHLQ H[SUHVVLRQ �0L]XWDQL HW DO�� ������ :H SHUIRUPHG LQ XWHUR HOHFWURSRUDWLRQ XVLQJ
WKH WKUHH SODVPLGV WR YLVXDOL]H SRVVLEOH GLIIHUHQFHV LQ WKH W\SH RI FHOOV DFWLYDWLQJ WKH GLIIHUHQW
SURPRWHUV LQ WKH VDPH WLVVXH� 7ZHQW\�IRXU KRXUV DIWHU HOHFWURSRUDWLRQ XVLQJ WKH &$* SURPRWHU�
ZH IRXQG D FRQVLGHUDEOH QXPEHU RI ODEHOHG FHOOV ORFDWHG LQ WKH 69=� ,QWHUPHGLDWH ]RQH �,=�� $
VLPLODU UHVXOW ZDV REVHUYHG XVLQJ WKH :QW UHSRUWHU SODVPLG� VKRZLQJ QR VLJQLILFDQW GLIIHUHQFHV
LQ WKH SHUFHQWDJH RI HOHFWURSRUDWHG FHOOV LQVLGH WKH 9= �S&$*�%)3� ����� � ������ 723�*)3�
����� � ������ W ��� ������ S ������ XQSDLUHG W WHVW� Q �� )LJ� �%� (�� +RZHYHU� ZLWK WKH
+HV� UHSRUWHU PRVW RI WKH FHOOV ZHUH IRXQG LQ WKH 9= �S&$*�%)3� ����� � ������
+HV�S�GV5HG� ����� � ������ W ��� ����� S ������ XQSDLUHG W WHVW� Q �� )LJ� �%� (�� $
VLPLODU SDWWHUQ ZDV REVHUYHG XVLQJ DQRWKHU 1RWFK UHSRUWHU SODVPLG FRQWDLQLQJ WKH SURPRWHU
&%)�5( �0L]XWDQL HW DO�� ������ D WDQGHP RI � &%)��ELQGLQJ HOHPHQWV GULYLQJ WKH H[SUHVVLRQ
RI D GLIIHUHQW IOXRUHVFHQW SURWHLQ� *)3 �S&$*�%)3� ����� � ������ &%)5(�*)3� ����� �
��

)DEUD�%HVHU�HW�DO�
������ W ��� ������ S ���� [ ����� XQSDLUHG W WHVW� Q �� )LJ� �&� (�� &R�HOHFWURSRUDWLRQ
H[SHULPHQWV RI +HV�S�GV5HG DQG (P[�S�*)3 SODVPLGV DW (���� VKRZHG FR�ORFDOL]DWLRQ RI WKH
WZR UHSRUWHU JHQHV LQ DOO RI WKH WDUJHWHG FHOOV WZHQW\�IRXU KRXUV DIWHU HOHFWURSRUDWLRQ �)LJ�
�'�(�� &RPSDULVRQ RI WKH GLVWULEXWLRQ RI ODEHOHG FHOOV XVLQJ WKH (P[� DQG &$* SURPRWHUV
FRQVLVWHQWO\ UHYHDOHG WKH H[LVWHQFH RI VWDWLVWLFDOO\ VLJQLILFDQW GLIIHUHQFHV �S&$*�%)3� ����� �
������ (P[�S�*)3� ����� � ������ W ��� ���� S �������� XQSDLUHG W WHVW� Q �� )LJ� �'�(��
)RU VLPSOLFLW\� ZH UHIHU WR FHOOV DFWLYDWLQJ WKH +HV� SURPRWHU DV +HV�� FHOOV� DQG WKRVH
DFWLYDWLQJ�WKH�&$*�SURPRWHU�DV�&$*��FHOOV�
,Q RUGHU WR FRQILUP WKDW WKH +HV�� FHOOV LQ WKH 9= ZHUH 5*&V� ZH SHUIRUPHG
LPPXQRKLVWRFKHPLVWU\� :H IRXQG WKDW IRUW\�HLJKW KRXUV DIWHU HOHFWURSRUDWLRQ WKH PDMRULW\ RI WKH
+HV�� FHOOV H[SUHVVHG WKH 5*& PDUNHU 3D[� �)LJ� �)� DUURZV� EXW QRW WKH LQWHUPHGLDWH
SURJHQLWRU��,3��PDUNHU�7EU���)LJ���*��DUURZ�KHDGV��
+HV���5*&V�0RVWO\�*HQHUDWH�8SSHU�/D\HU�&RUWLFDO�1HXURQV
+HV�� 5*&V DW HDUO\ VWDJHV RI FRUWLFRJHQHVLV DSSHDU WR SUHVHQW GHOD\HG QHXURJHQHVLV FRPSDUHG
WR RWKHU QHXURJHQLF SURJHQLWRUV DW WKH VDPH GHYHORSPHQWDO VWDJH� :H QH[W DQDO\]HG GLVWULEXWLRQ
RI HOHFWURSRUDWHG FHOOV DW ODWHU VWDJHV DIWHU LQ XWHUR HOHFWURSRUDWLRQ WR GHWHUPLQH ODPLQDU
ORFDOL]DWLRQ RI WKH QHXURQV SURGXFHG E\ +HV�� FHOOV �+HV�S�GV5HG� LQ FRPSDULVRQ ZLWK &$*�
FHOOV �S&$*�%)3� ZH DQDO\]HG WKH ODPLQDU SRVLWLRQ RI ODEHOHG FHOOV VHYHQ GD\V DIWHU (����
FR�HOHFWURSRUDWLRQ RI WKH WZR SODVPLGV �)LJ� �$�� &$*� QHXURQV ZHUH ORFDWHG ZLWKLQ ERWK WKH
XSSHU DQG ORZHU SDUWV RI WKH GHYHORSLQJ FRUWLFDO SODWH �&3�� ZLWK JUHDWHU DFFXPXODWLRQ LQ WKH
ORZHU SDUW� 2Q WKH RWKHU KDQG� +HV�� QHXURQV PRVWO\ DFFXPXODWHG LQ WKH XSSHU SDUW RI WKH &3
�S&$*�%)3 YV +HV�S�GV5HG� ELQ �� S ���� [ ����� ELQ �� S ���� [ ���� � ELQ �� S ���� [ ����
� ELQ �� S ������� ELQ �� S ������� ELQ �� S ���� [ ���� � ELQ �� S ���� [ ����� ELQ �� S
������� ELQ ��� S ������� XQSDLUHG W WHVW� Q �� )LJ� �%�&�� ,Q RUGHU WR GHWHUPLQH WKH VXEW\SH
RI QHXURQV WKDW ZHUH +HV��� ZH FRQGXFWHG LPPXQRKLVWRFKHPLVWU\ XVLQJ NQRZQ PDUNHUV IRU
VXEW\SHV RI QHRFRUWLFDO QHXURQV� QDPHO\ &X[�� 6DWE�� 7OH� DQG &WLS�� &X[� LV D VSHFLILF PDUNHU
IRU XSSHU OD\HU QHXURQV �1LHWR HW DO�� ����� DQG WKH PDMRULW\ RI +HV�� FHOOV ORFDWHG LQ WKH XSSHU
��

)DEUD�%HVHU�HW�DO�
SDUW RI WKH &3 DOVR H[SUHVVHG &X[� ������ � ������ )LJ� �'� +�� 6DWE� LV D PDUNHU RI
FRUWLFR�FRUWLFDO QHXURQV� PRVWO\ ORFDWHG LQ XSSHU OD\HUV EXW DOVR SUHVHQW LQ VRPH ORZHU OD\HU
QHXURQV �$OFDPR HW DO�� ����� %ULWDQRYD HW DO�� ������ 0RVW RI WKH +HV�� FHOOV ZHUH 6DWE��
������ � ������ )LJ� �(� +�� 9HU\ IHZ +HV�� FHOOV H[SUHVV 7OH�� D PDUNHU IRU FRUWLFR�WKDODPLF
QHXURQV LQ OD\HU 9, �.RRS HW DO�� ����� RU &WLS�� D PDUNHU IRU QHXURQV LQ OD\HUV 9 DQG 9,
�$UORWWD HW DO�� ����� �+HV��7OH���+HV��� ���� � ������ +HV��&WLS���+HV��� ���� � ������
)LJ� �)�+�� 7KHVH UHVXOWV VXJJHVW WKDW +HV�� SURJHQLWRUV DUH LQYROYHG LQ WKH JHQHUDWLRQ RI
FRUWLFR�FRUWLFDO�SURMHFWLRQ�QHXURQV�PRVWO\�ORFDWHG LQ�XSSHU�OD\HUV�
+HV� LV QRW H[SUHVVHG LQ GLIIHUHQWLDWHG QHXURQV �%DVDN DQG 7D\ORU� ������ VXJJHVWLQJ WKDW
WKH ODEHOHG QHXURQV ORFDWHG LQ XSSHU OD\HUV FDPH IURP HDUO\ +HV�� 5*&V� 7R IRUPDOO\ WHVW WKLV
SRVVLELOLW\� ZH FRQGXFWHG OLQHDJH WUDFLQJ H[SHULPHQWV� :H JHQHUDWHG D WDPR[LIHQ LQGXFLEOH
&UH�UHFRPELQDVH FRQVWUXFW GULYHQ E\ WKH +HV� SURPRWHU �+HV�S�&UH(57�� WR FRQGXFW LQ XWHUR
HOHFWURSRUDWLRQ DW (���� LQ $L� UHSRUWHU PLFH �0DGLVHQ HW DO�� ������ $V D FRQWURO� ZH XVHG D
JHQHUDO SURPRWHU �S&$*�&UH(57��� 6LQFH ��2+7 KDV D VKRUW KDOI�OLIH �*XHQWKQHU HW DO�� ������
ZH LQMHFWHG ��2+7 LPPHGLDWHO\ IROORZLQJ LQ XWHUR HOHFWURSRUDWLRQ WR PD[LPL]H UHFRPELQDWLRQ
LQ 5*&V WDNLQJ XS WKH SODVPLG� EHIRUH WKH\ JHQHUDWHG PRUH GLIIHUHQWLDWHG FHOO W\SHV �)LJ� �$��
:H WKHQ DQDO\]HG WKH GLVWULEXWLRQ RI WKH QHXURQDO RIIVSULQJ RI HOHFWURSRUDWHG 5*&V DW 3���
)ROORZLQJ +HV�S�&UH(57� HOHFWURSRUDWLRQV DQG ��2+7 LQMHFWLRQ� WKHUH ZDV D VWURQJ ELDV LQ WKH
ORFDOL]DWLRQ RI WG�7RPDWR� S\UDPLGDO QHXURQV WRZDUGV XSSHU OD\HUV ZKHQ FRPSDUHG WR
S&$*�&UH(57� HOHFWURSRUDWLRQV �S&$*�&UH(57� YV +HV�S�&UH(57�� ELQ �� S ������� ELQ
�� S �������� ELQ �� S ��� ����� ELQ �� S �������� ELQ �� S ������ ELQ �� S ��� ����� ELQ
�� S ��� ����� ELQ �� S ������� ELQ ��� S ������� XQSDLUHG W WHVW� Q �� )LJ� �%�� 0RVW RI
WKH WG�7RPDWR� QHXURQV LQ EUDLQV HOHFWURSRUDWHG ZLWK +HV�S�&UH(57� H[SUHVVHG WKH XSSHU OD\HU
PDUNHU &X[� ������ � ������ )LJ� �&� (� DQG WKH FRUWLFR�FRUWLFDO PDUNHU 6DWE� ������ � ������
)LJ� �'�(�� HYHQ ZKHQ ORFDWHG LQ ORZHU OD\HUV �)LJ� �'�� 9HU\ IHZ RI WKHP H[SUHVVHG &WLS�
����� � ������� ZKLFK LV PRVWO\ H[SUHVVHG E\ FRUWLFR�IXJDO QHXURQV DQG VRPH LQWHUQHXURQV
�$UORWWD HW DO�� ����� �)LJ� �&�(�� ,Q FRQWUDVW� WG�7RPDWR� QHXURQV LQ EUDLQV HOHFWURSRUDWHG ZLWK
S&$*�&UH(57� FRPSULVHG KLJKHU SHUFHQWDJHV RI &WLS�� QHXURQV ������ � ������ W ��� �����
��

)DEUD�%HVHU�HW�DO�
S ������� XQSDLUHG W WHVW� Q �� DQG UHGXFHG QXPEHUV RI &X[�� DQG 6DWE�� UHFRPELQHG
QHXURQV �&X[�������� ������ � W ��� ������ S ������� 6DWE��� ����� � ������ W ���
������� S �������� XQSDLUHG W WHVW� Q �� )LJ� �(�� ,Q DGGLWLRQ WR WKH QHXURQDO RIIVSULQJ� ZH
REVHUYHG WKH SUHVHQFH RI WUDFHG JOLDO FHOOV ZLWK DVWURF\WH PRUSKRORJ\ �)LJ� �%�&�'� XVLQJ ERWK
SURPRWHU IUDJPHQWV� $OWKRXJK WRWDO QXPEHU RI WUDFHG DVWURF\WHV LV UHGXFHG XVLQJ WKH +HV�
SURPRWHU� WKH SURSRUWLRQ RI DVWURF\WHV YHUVXV S\UDPLGDO QHXURQV LV PDLQWDLQHG LQ ERWK
FRQGLWLRQV �� RI WG�7RPDWR DVWURF\WHV� S&$*�&UH(57�� ����� � ������ +HV�S�&UH(57��
����� � ������ W ��� ������ � S ����� XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ����$��
6LPLODUO\� QR PDVVLYH ODPLQDU GLVWULEXWLRQ RI DVWURF\WH ZDV IRXQG EHWZHHQ WKH WZR FRQGLWLRQV
�S&$*�&UH(57� YV +HV�S�&UH(57�� ELQ �� S ������ XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD
)LJ�����%��
6LQJOH &HOO 51$ 6HTXHQFLQJ RI (DUO\ +HV�� 5*&V 5HYHDOV D 0ROHFXODU 6LJQDWXUH RI
&RUWLFDO�3URJHQLWRUV�ZLWK�'HOD\HG�1HXURJHQLF�3RWHQWLDO
7KH SUHYLRXV UHVXOWV VXJJHVW WKH SRVVLEOH H[LVWHQFH RI WZR VXEW\SHV RI 5*&V DW WKHVH SDUWLFXODU
DJHV� RQH RI WKHP EHLQJ HDUO\ QHXURJHQLF DQG RWKHU UHPDLQLQJ ORQJHU DV 5*&V EHIRUH JHQHUDWLQJ
QHXURQV� 7R VHDUFK IRU PROHFXODU GLIIHUHQFHV EHWZHHQ WKHVH 5*&V� ZH GHVLJQHG DQ H[SHULPHQW
WR LVRODWH QHRFRUWLFDO SURJHQLWRU FHOOV DQG FKDUDFWHUL]H WKHP E\ VLQJOH FHOO 51$ VHTXHQFLQJ
�VF51$�VHT�� )RU WKLV SXUSRVH� ZH JHQHUDWHG D GXDO SURPRWHU SODVPLG FRQWDLQLQJ WZR
IOXRUHVFHQW SURWHLQV XQGHU WKH FRQWURO RI WZR GLIIHUHQW SURPRWHUV� WKH &$* SURPRWHU GULYLQJ
P5)3 DQG WKH +HV� SURPRWHU GULYLQJ *)3� 7KLV VWUDWHJ\ HQVXUHV WKDW ERWK UHSRUWHU FRQVWUXFWV
DUH SUHVHQW ZLWKLQ HDFK HOHFWURSRUDWHG FHOO� 9DOLGDWLRQ RI WKH FRQVWUXFW E\ LQ XWHUR
HOHFWURSRUDWLRQ DW (���� FRQILUPHG VLPLODU GLVWULEXWLRQ RI ODEHOHG FHOOV DW GLIIHUHQW PRPHQWV
DIWHU VXUJHU\ DV ZH GHVFULEHG ZLWK WKH LQGLYLGXDO SODVPLGV �)LJ� �� )LJ� �� ([WHQGHG 'DWD )LJ�
���$�&�� 7ZR GD\V DIWHU HOHFWURSRUDWLRQ D UHGXFHG QXPEHU RI +HV�� FHOOV ZHUH ORFDWHG LQ WKH
&3 ZKHUHDV D VLJQLILFDQWO\ KLJKHU QXPEHU RI &$*� FHOOV ZHUH IRXQG LQ VXFK FRPSDUWPHQW
�&$*�� ����������� +HV��� ���������� W ��� ������ S ����[����� XQSDLUHG W WHVW� Q ��
([WHQGHG 'DWD )LJ� ���$�%�� &RQYHUVHO\� WKH SURSRUWLRQ RI +HV�� FHOOV SUHVHQW LQ WKH 9= ZHUH
��

)DEUD�%HVHU�HW�DO�
VLJQLILFDQWO\ LQFUHDVHG FRPSDUHG ZLWK WKH SURSRUWLRQ RI &$*� FHOOV �&$*�� �����������
+HV��� ����������� W ��� ������ S ������� XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ� ���%��
6PDOOHU GLIIHUHQFHV LQ WKH GLVWULEXWLRQ RI ODEHOHG FHOOV LQ WKH 69= �&$*�� ����������� +HV���
����������� W ��� ������ S ����� XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ����%� DQG WKH ,=
�&$*�� ����������� +HV��� ����������� W��� ���� � S ����� XQSDLUHG W WHVW� Q �� ([WHQGHG
'DWD )LJ� ���%� KDYH EHHQ DOVR IRXQG� $QDO\VLV DW ODWHU DJHV �3�� FRQILUPHG WKH SUHIHUHQW
RFFXSDWLRQ RI +HV�� FHOOV LQ XSSHU OD\HUV DIWHU (���� HOHFWURSRUDWLRQ ZLWK WKH GXDO YHFWRU
�([WHQGHG�'DWD�)LJ�����&��
,Q RUGHU WR FRQGXFW WKH VF51$�VHT VHTXHQFLQJ H[SHULPHQWV ZH DGMXVWHG WKH WLPLQJ EHWZHHQ
WKH VXUJHU\ IRU LQ XWHUR HOHFWURSRUDWLRQ DW (���� DQG FHOO LVRODWLRQ WR RSWLPL]H IOXRUHVFHQFH
LQWHQVLW\ RI FHOOV ODUJHO\ FRQILQHG WR WKH SUROLIHUDWLYH ]RQH RI WKH QHRFRUWH[ �)LJ� �$�� 7KH
RSWLPDO WLPH SRLQW IRU FHOO VRUWLQJ ZDV �� KRXUV DIWHU LQ XWHUR HOHFWURSRUDWLRQ� GLVSOD\LQJ
VXEVWDQWLDO QXPEHUV RI 3D[�� 5*&V �)LJ� �$� H[SUHVVLQJ HLWKHU RQO\ P5)3 �)LJ� �%� DUURZV� RU
ERWK IOXRUHVFHQW SURWHLQV �)LJ� �%� DUURZ KHDGV�� /DEHOHG EUDLQV ZHUH GLVVHFWHG DQG WKH
HOHFWURSRUDWHG DUHDV RI WKH QHRFRUWH[ ZHUH LVRODWHG DQG VRUWHG XVLQJ IOXRUHVFHQFH�DFWLYDWHG FHOO
VRUWLQJ �)$&6� WR VHSDUDWH &$*� FHOOV �RQO\ &$* SURPRWHU DFWLYH� P5)3�*)3� FHOOV� DQG
+HV�� FHOOV �ERWK SURPRWHUV DFWLYH� P5)3�*)3� FHOOV� �)LJ� �&�� 9LDEOH VRUWHG FHOOV RI ERWK
W\SHV FROOHFWHG LQGLYLGXDOO\ ZHUH O\VHG DQG OLEUDULHV ZHUH JHQHUDWHG XVLQJ D PRGLILHG 6PDUW�VHT
��SURWRFRO�IRU�VXEVHTXHQW�VF51$�VHT�VHTXHQFLQJ�DV GHVFULEHG��&KHYpH�HW�DO���������
'DWD ZHUH DQDO\]HG XVLQJ WKH 6HXUDW 5 SDFNDJH �%XWOHU HW DO�� ������ $ WRWDO RI ���� FHOOV
SDVVHG RXU TXDOLW\ FRQWURO WHVW� )URP WKRVH FHOOV� ZH REWDLQHG ��� &$*��+HV�� �P5)3� DQG
*)3�� DQG RI ��� &$*� FHOOV �P5)3� EXW *)3��� 8QVXSHUYLVHG WUDQVFULSWLRQDO DQDO\VLV
DPRQJ VLQJOH FHOOV LGHQWLILHG � FHOO FOXVWHUV FKDUDFWHUL]HG E\ VLPLODU JHQH H[SUHVVLRQ SDWWHUQV
�([WHQGHG 'DWD )LJ� ���(�� %\ XVLQJ FHOO�W\SH VSHFLILF PDUNHUV� WKHVH FOXVWHUV FRXOG EH DVVLJQHG
WR WKH WKUHH PDLQ FHOO W\SHV RI WKH GHYHORSLQJ QHRFRUWH[� 5*&V �3D[�� 9LP� +HV�� 6R[��
�6FKQLW]HU HW DO�� ����� *|W] HW DO�� ����� +DWDNH\DPD HW DO�� ����� 6FRWW HW DO�� ������ ,3V
�(RPHV� +HV�� *DGG��J� �(QJOXQG HW DO�� ����� .DZDJXFKL HW DO�� ������ DQG HDUO\ QHXURQV
�(1V� �1HXURG�� 7XEE�� 6QDS��� 5EIR[�� �&DFFDPR HW DO�� ����� 2\OHU HW DO�� ����� 0XOOHQ HW
��

)DEUD�%HVHU�HW�DO�
DO�� ����� *RHEEHOV HW DO�� ������ 7KH H[SUHVVLRQ RI WKRVH PDUNHUV DOORZHG XV WR DVVLJQ WKH
+HV�� DQG &$*� FHOO W\SHV WR WKH WKUHH GLIIHUHQW ODUJH JURXSV RI FHOOV SUHYLRXVO\ VKRZQ �)LJ�
�'�(��([WHQGHG�'DWD�)LJ�����)��
:H QH[W DQDO\]HG WKH SURSRUWLRQ RI WKHVH PDLQ FHOO W\SHV EHWZHHQ +HV�� �P5)3�*)3��
DQG &$*� �P5)3�*)3�� FHOOV ZHUH GLVWULEXWHG EHWZHHQ WKH GLIIHUHQW FHOO�W\SH FOXVWHUV �)LJ�
�)�� :KLOH WKH SURSRUWLRQ RI ,3V ZDV VLPLODU IRU ERWK JURXSV ������ � RI +HV�� FHOOV DQG �����
� RI &$*� FHOOV�� PRVW RI WKH &$*� FHOOV ZHUH FDWHJRUL]HG DV (1V ��������� EXW RQO\ VPDOO
SHUFHQWDJH RI WKH +HV�� FHOOV ZHUH QHXURQV ����� �� �)LJ� �*�� VXJJHVWLQJ WKDW &$*� 5*&V
DUH IDVW QHXURJHQLF DW WKH WLPH RI HOHFWURSRUDWLRQ �(���� &RQYHUVHO\� +HV�� FHOOV ODUJHO\ KDG WKH
WUDQVFULSWLRQDO VLJQDWXUH RI 5*&V �������� ZKLOH RQO\ D UHGXFHG IUDFWLRQ RI &$*� FHOOV
GLVSOD\HG�5*&�FKDUDFWHULVWLFV������������)LJ���*��
1H[W� ZH VHW RXW WR LGHQWLI\ JHQH H[SUHVVLRQ VLJQDWXUHV WKDW FRXOG GLVWLQJXLVK +HV�� IURP
&$*� 5*&V �)LJXUH ��� 7R WKDW� ZH ILUVW LQYHVWLJDWHG GLIIHUHQWLDOO\ H[SUHVVHG JHQHV �'(*V� LQ
WKHVH WZR SRSXODWLRQV XVLQJ :LOFR[RQ UDQN VXP WHVW �)LJ� �$�� :H LGHQWLILHG �� JHQHV ZKRVH
H[SUHVVLRQ ZDV VLJQLILFDQWO\ XSUHJXODWHG LQ WKH VXESRSXODWLRQ RI +HV�� FRPSDUHG WR &$*�
5*&V �DGMXVWHG S�YDOXH � ���� DQG DYHUDJH ORJ�)& ! ���� )LJ� �$�� $PRQJ WKHVH '(*V� ZH
LGHQWLILHG WKH WUDQVFULSWLRQ IDFWRU 6R[� WKDW FRXOG SOD\ D NH\ UROH LQ WKH UHJXODWLRQ RI 5*& IDWH
�6WROW HW DO�� ����� 9RQJ HW DO�� ����� .DSODQ HW DO�� ������ 7R LQYHVWLJDWH ZKHWKHU 6R[� FRXOG EH
D PDVWHU UHJXODWRU RI JHQH H[SUHVVLRQ LQ +HV�� 5*&V� ZH JHQHUDWHG D WUDQVFULSWLRQDO UHJXODWRU\
QHWZRUN DQG VHDUFKHG IRU PDVWHU UHJXODWRUV �0DUJROLQ HW DO�� ����� )OHWFKHU HW DO�� ����� &DVWUR
HW DO�� ����� XVLQJ SUHYLRXVO\ SXEOLVKHG VF51$�VHT GDWD DW WKH VDPH DJH DV RXU DQDO\VLV �(���
�<X]ZD HW DO�� ����� �WRWDO RI ���� FHOOV DQG ����� JHQHV�� $ WUDQVFULSWLRQDO UHJXODWRU\ QHWZRUN
FRQVLVWV RI D FROOHFWLRQ RI UHJXODWHG WDUJHW JHQHV DQG WUDQVFULSWLRQ IDFWRUV �7)V�� LQ ZKLFK WKH VHW
RI JHQHV FRQWUROOHG E\ D JLYHQ 7) IRUPV D UHJXORQ �0DUJROLQ HW DO�� ����� )OHWFKHU HW DO�� �����
&DVWUR HW DO�� ������ :H IRXQG D UHJXODWRU\ QHWZRUN FRQWDLQLQJ �� UHJXORQV GLIIHUHQWO\
H[SUHVVHG LQ 5*&V� ,3V DQG (1V �)LJ� �%� ([WHQGHG 'DWD )LJ� ��� $�%�� :H DOVR REVHUYHG WKDW
WKH UHJXODWRU\ QHWZRUN RI 5*&V FRXOG EH GLVWLQJXLVKHG IURP WKRVH RI ,3V DQG (1V E\ D KLJKHU
H[SUHVVLRQ RI +HV�� 6R[�� 3D[�� ,G� DQG =IS��O� �)LJ� �%� ([WHQGHG 'DWD )LJ� ��� $�%��
��

)DEUD�%HVHU�HW�DO�
0RUHRYHU� ZH IRXQG WKDW 6R[� DQG +HV� UHJXORQV ZHUH QH[W WR HDFK RWKHU LQ WKH QHWZRUN�
LQGLFDWLQJ WKDW WKHVH WZR WUDQVFULSWLRQ IDFWRUV VKDUH FRPPRQ WDUJHW JHQHV� EXW RQO\ LQ +HV��
5*&V 6R[� ZDV LGHQWLILHG DV D PDVWHU UHJXODWRU �)LJ� �%�� IXUWKHU VXJJHVWLQJ WKDW 6R[� FRXOG EH
DQ�LPSRUWDQW�IDWH�GHWHUPLQDQW�IRU�WKH�+HV���5*&V�
7R YHULI\ GLIIHUHQFHV LQ 6R[� SURWHLQ H[SUHVVLRQ EHWZHHQ WKH WZR 5*&V VXEW\SHV ZH
SHUIRUPHG LQ XWHUR HOHFWURSRUDWLRQ XVLQJ WKH GXDO SURPRWHU SODVPLG DQG FRQGXFWHG 6R[�
LPPXQRKLVWRFKHPLVWU\ �� KRXUV DIWHU WKH VXUJHU\ �)LJ� �&�� 4XDQWLWDWLYH DQDO\VLV RI 6R[�
VWDLQLQJ LQ +HV�� �)LJ� �'� DUURZV� DQG &$*� 5*&V �)LJ� �'� DUURZ KHDGV� FRQILUPHG
VWDWLVWLFDOO\ VLJQLILFDQW GLIIHUHQFHV LQ 62;� H[SUHVVLRQ OHYHOV EHWZHHQ WKRVH 5*&V� DV VKRZQ E\
WKH DQDO\VLV RI WKH PHDQ LQWHQVLW\ RI IOXRUHVFHQFH �&$*�� ������������ +HV��� ������������� W
��� ������ S ������ XQSDLUHG W WHVW� Q �� )LJ� �(� DQG WKH HYLGHQW GLIIHUHQWLDO GLVWULEXWLRQ RI
6R[��IOXRUHVFHQFH�LQWHQVLW\�EHWZHHQ�&$*��DQG�+HV�� 5*&V��([WHQGHG�'DWD�)LJ����&��
6R[��2YHUH[SUHVVLRQ�LQ�5*&V�$IIHFWV�(DUO\�1HXURQDO 'LIIHUHQWLDWLRQ
6R[� LV UHTXLUHG IRU VWHP FHOO PDLQWHQDQFH LQ GLIIHUHQW WLVVXHV �6FRWW HW DO�� ����� 5RFKH HW DO��
����� DV ZHOO DV IRU VXSSUHVVLRQ RI FHOO GLIIHUHQWLDWLRQ �.DGDMD HW DO�� ������ ,Q WKH &16�
RYHUH[SUHVVLRQ RI 6R[� LQ WKH DGXOW VXEYHQWULFXODU ]RQH �D69=� DEROLVKHV WKH SURGXFWLRQ RI
QHXURQV� ZKHUHDV 6R[� NQRFNGRZQ LQFUHDVHG QHXURJHQHVLV DQG UHGXFHG JOLRJHQHVLV �&KHQJ HW
DO�� ������ %HFDXVH 6R[� KDV EHHQ IRXQG WR EH D SXWDWLYH PDVWHU UHJXODWRU VSHFLILF RI WKH +HV��
FHOOV� ZH SUHGLFWHG WKDW LQFUHDVHG OHYHOV RI 6R[� REVHUYHG LQ WKRVH SURJHQLWRUV PD\ DIIHFW WKH
VZLWFK EHWZHHQ QRQ�QHXURJHQLF DQG QHXURJHQLF FHOO GLYLVLRQV� 7R WHVW WKLV K\SRWKHVLV� ZH
RYHUH[SUHVVHG 6R[� LQ 5*&V DW (���� XVLQJ D JHQHUDO SURPRWHU GULYLQJ 6R[� DQG *)3
�S&,*�)/ 6R[�� DQG DQDO\]HG SRVVLEOH FKDQJHV LQ GLIIHUHQWLDWLRQ DW (���� �)LJ� �$�� 6R[�
RYHUH[SUHVVLRQ DIIHFWHG WKH GLVWULEXWLRQ RI WDUJHWHG FHOOV ZLWK UHGXFHG LQYDVLRQ RI WKH &3 DQG D
JUHDWHU DFFXPXODWLRQ RI ODEHOHG FHOOV LQ WKH 9= �S&,* YV S&,*�)/6�� 9=� S �������� ,=� S
������ &3� S ���� ����� XQSDLUHG W WHVW� Q �� )LJ� �%�'�� 2YHUH[SUHVVLRQ RI 6R[� GLG QRW
QRWLFHDEO\ DIIHFW WKH 5*&V VFDIIROG DV UHYHDOHG E\ WKH YLVXDOL]DWLRQ RI WKHLU HQG IHHW �)LJ� �&��
'HWDLOHG DQDO\VLV RI WKH FHOOV DFFXPXODWHG LQ WKH 9= XVLQJ PDUNHUV IRU 5*&V �3D[��� ,3V �7EU��
��

)DEUD�%HVHU�HW�DO�
DQG SRVWPLWRWLF QHXURQV �'F[� UHYHDOHG WKDW WKH 6R[� RYHUH[SUHVVLQJ FHOOV ZHUH 5*&V �)LJ�
�(�*�� 0RUHRYHU� TXDQWLILFDWLRQV RI WKHVH FHOO PDUNHUV VKRZHG D VWDWLVWLFDOO\ VLJQLILFDQW
LQFUHDVH LQ WKH QXPEHU RI 3D[�� FHOOV XSRQ 6R[� RYHUH[SUHVVLRQ �S&,*� ������������
S&,*�)/6�� ������������ W ��� ����� � S ������� XQSDLUHG W WHVW� Q �� )LJ� �(�+� DQG D
FRQFRPLWDQW GHFUHDVH LQ WKH QXPEHU RI 'F[� HDUO\ QHXURQV �S&,*� ������������ S&,*�)/6��
��������������W����� ������ S ����������XQSDLUHG W�WHVW��Q� ����)LJ���*�+��
%HFDXVH 6R[� RYHUH[SUHVVLRQ VHHPV WR GHOD\ QHXURQ SURGXFWLRQ GXULQJ HDUO\ VWDJHV RI
FRUWLFRJHQHVLV� ZH GHFLGHG WR FKHFN LI 6R[� GRZQUHJXODWLRQ LQ WKH HDUO\ FRUWLFDO 9= LQFUHDVHV
WKH QHXURJHQLF EHKDYLRU RI 5*&V DV GHVFULEHG LQ WKH D69= �&KHQJ HW DW� ������ 7R WKLV HQG� ZH
SHUIRUPHG VKRUW�WLPH LQ XWHUR HOHFWURSRUDWLRQ IURP (���� WR (���� XVLQJ SUHYLRXVO\ YDOLGDWHG
6R[� RU VFUDPEOHG VK51$ WRJHWKHU ZLWK WKH 3&,* YHFWRU� 7KH DQDO\VLV RI WKH H[SUHVVLRQ RI
6R[� LQ HOHFWURSRUDWHG VDPSOHV FRQILUPHG WKH UHGXFWLRQ LQ 6R[� OHYHOV LQ 5*&V WDUJHWHG ZLWK
WKH 6R[��VK51$ LQ FRPSDULVRQ ZLWK WKH FRQWURO VLWXDWLRQ �([WHQGHG 'DWD )LJ� ���$��7KHVH
6R[� NQRFN�GRZQ �.'� H[SHULPHQWV VKRZHG DQ LQFUHDVH LQ WKH QXPEHU RI WDUJHWHG QHXURQV
RFFXS\LQJ WKH FRUWLFDO SODWH �&5�6K51$� ������������ 6��6K51$� ������������ W ���
������ S ������ XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ� ��� $�%�� VXJJHVWLQJ WKDW UHGXFWLRQV
LQ�6R[��H[SUHVVLRQ�IDYRUV�QHXURJHQLF�GLYLVLRQV�
6R[��2YHUH[SUHVVLRQ�$IIHFWV�5*&V�&HOO�&\FOH�'XUDWLRQ�
7KH LPEDODQFH LQ WKH QHXURQDO SURGXFWLRQ VHHQ XSRQ 6R[� SHUWXUEDWLRQ FRXOG EH UHODWHG ZLWK
DOWHUDWLRQV LQ WKH FHOO�F\FOH RI 5*&V� 7R YDOLGDWH WKLV K\SRWKHVLV� ZH SHUIRUPHG D FRPSOHWH
FHOO�F\FOH DQDO\VLV FRPSDULQJ WKH FRUWLFDO 9= RI HPEU\RV HOHFWURSRUDWHG DW (���� ZLWK WKH
FRQWURO SODVPLG �S&,*� RU WKH 6R[� RYHUH[SUHVVLRQ SODVPLG �S&,*�)/ 6R[��� �� K DIWHU WKH
VXUJHU\ �(������ :H ILUVW HYDOXDWHG WKH SURSRUWLRQ RI *)3� FHOOV LQ WKH 9= LQ GLIIHUHQW
FHOO�F\FOH SKDVHV XVLQJ LPPXQRKLVWRFKHPLVWU\ IRU NQRZQ FHOO�F\FOH PDUNHUV LQFOXGLQJ
SKRVSKRU\ODWHG YLPHQWLQ �S9LP� IRU 0�SKDVH �.DPHL HW DO�� ����� 1RFWRU HW DO�� ����� DQG WKH
JHQHUDO FHOO�F\FOH PDUNHU .L�� �*HUGHV HW DO�� ������ 0RUHRYHU� ZH FRQGXFWHG VKRUW WLPH SXOVHV
��

)DEUD�%HVHU�HW�DO�
RI WKH WK\PLGLQH DQDORJ (G8 ��� PLQ� WR HYDOXDWH WKH SURSRUWLRQ RI FHOOV LQ 6�SKDVH �%XFN HW DO��
������ ,Q WKH WKUHH FDVHV ZH IRXQG VLJQLILFDQW UHGXFWLRQV LQ WKH QXPEHU RI GRXEOH ODEHOHG FHOOV
XSRQ 6R[� RYHUH[SUHVVLRQ� S9LP �S&,*� ������������ S&,*)/�6�� ����������� W ��� ������
S ���� ����� XQSDLUHG W WHVW� Q �� )LJ� �&�� (G8 �S&,*� ������������ S&,*)/�6��
������������ W ��� ����� S ������� XQSDLUHG W WHVW� Q �� )LJ� �(� DQG .L�� �S&,*�
������������ S&,*)/�6�� ������������ W ��� ����� S �������� XQSDLUHG W WHVW� Q �� )LJ�
�*���7KHVH�GDWD�WRJHWKHU�VXJJHVW�WKH�H[LVWHQFH�RI UHGXFHG�FHOO�GLYLVLRQV�DW�WKH�DQDO\]HG�DJH�
7R LQYHVWLJDWH SRVVLEOH GLIIHUHQFHV LQ FHOO�F\FOH H[LW ZH LQMHFWHG (���� HOHFWURSRUDWHG
IHPDOHV ZLWK %UG8� �� K DIWHU VXUJHU\ �(����� WR ODEHO FHOOV LQ 6�SKDVH DQG DQDO\]H
HOHFWURSRUDWHG EUDLQV ��K DIWHU WKH LQMHFWLRQ �(����� �([WHQGHG 'DWD )LJ� ���$�� 'RXEOH
LPPXQRKLVWRFKHPLVWU\ WR GHWHFW %UG8 DQG .L�� VWDLQLQJ LQ *)3� FHOOV LQ WKH 9= �([WHQGHG
'DWD )LJ� ��� %�&� UHYHDOHG DQ LPSRUWDQW UHGXFWLRQ LQ WKH WRWDO QXPEHU RI .L���%UG8� F\FOLQJ
FHOOV LQ WKH RYHUH[SUHVVLRQ FRQGLWLRQ WRJHWKHU ZLWK DQ LQFUHDVH LQ WKH SURSRUWLRQ RI .L���%UG8�
�� RI *)3�.L���%UG8� FHOOV LQ 9=� S&,*� ������������� S&,*)/�6�� ������������ W ���
����� S ������ � RI *)3�.L���%UG8� FHOOV LQ 9=� S&,*� ����������� S&,*)/�6��
������������ W ��� ������ S ������ XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ� ���'�� 7KHVH
GLIIHUHQFHV ZHUH PDLQO\ GHWHFWHG LQ WKH LQQHU SDUW RI WKH 9= �� RI *)3�.L���%UG8�� S&,* YV
S&,*�)/6�� ,9=� S ������ 29=� S ����� � RI *)3�.L���%UG8�� S&,* YV S&,*�)/6�,�
,9=� S ������� 29=� S ����� Q �� ([WHQGHG 'DWD )LJ� ��� (�)� ZKHUH PDQ\ RI WKH WDUJHWHG
FHOOV DFFXPXODWH XSRQ 6R[� RYHUH[SUHVVLRQ �S&,*� ������������ S&,*�)/6�� ������������ W
��� ������ S ������� XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ� �*�+�� 7KHVH UHVXOWV VXJJHVW
WKDW SDUW RI WKH F\FOLQJ FHOOV LQFRUSRUDWLQJ %UG8 DW (���� FRXOG KDYH H[LWHG FHOO F\FOH�
UHPDLQLQJ LQ WKH 9=� +RZHYHU� WKHVH *)3� GR QRW H[SUHVV PDUNHUV RI GLIIHUHQWLDWHG QHXURQV
OLNH 'F[ �)LJ� �)� ([WHQGHG 'DWD )LJ� ���,� RU ,3V PDUNHUV OLNH 7EU� �)LJ� �(�� DQG H[SUHVV WKH
5*& PDUNHU 3D[� �)LJ �'� ([WHQGHG 'DWD )LJ� ���-�� VXJJHVWLQJ D ODFN RI GLIIHUHQWLDWLRQ LQ
WKRVH�FHOOV�
$OWKRXJK .L�� LV FRQVLGHUHG D EURDG FHOO F\FOH PDUNHU� LWV H[SUHVVLRQ LV ODUJHO\ DEVHQW LQ
HDUO\ *� SKDVH DQG LQ TXLHVFHQW FHOOV WKDW FRXOG EH WUDQVLWLRQLQJ IURP *��*� �0LOOHU HW DO��
��

)DEUD�%HVHU�HW�DO�
������ 0RUHRYHU� LW KDV EHHQ UHSRUWHG WKDW 6R[� RYHUH[SUHVVLRQ LQ D FKRQGURF\WH FHOO OLQH
SURGXFHV DFFXPXODWLRQV RI FHOOV LQ *��*� FRPSDUHG ZLWK WKH FRQWURO VLWXDWLRQ �3DQGD HW DO��
������ 7KHUHIRUH� ZH VHOHFWHG D EURDGHU FHOO F\FOH PDUNHU� WKH PLQLFKURPRVRPDO PDLQWHQDQFH
SURWHLQ � �0FP��� ZLGHO\ H[SUHVVHG LQ UHSOLFDWLRQ�FRPSHWHQW FHOOV� LQFOXGLQJ FHOOV LQ HDUO\ *�
SKDVH RU TXLHVFHQW VWHP FHOOV� VLPLODU WR WKH DGXOW QHXUDO VWHP FHOOV �D16&V� RI WKH D69=
�0DVORY HW DO�� ����� 3RUODQ HW DO�� ������ ,Q WKH QHRFRUWH[� WKH H[SUHVVLRQ RI 0FP� LV ODUJHO\
FRQILQHG LQ WKH SUROLIHUDWLYH UHJLRQV �([WHQGHG 'DWD )LJ� ���.� DQG WKH SURSRUWLRQ RI
0FP���*)3� LQ WKH 9= IRU HDFK FRQGLWLRQ� 6R[� RYHUH[SUHVVLRQ DQG LWV FRQWURO� UHSUHVHQWV
RYHU ��� RI WKH WRWDO WDUJHWHG 5*&V �S&,*� ������������ S&,*)/�6�� ������������ W ���
����� S ������� XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ� ���/�� 'RXEOH LPPXQRVWDLQLQJV ZLWK
.L�� DQG 0FP� LQ HOHFWURSRUDWHG HPEU\RV DW (���� DQDO\]HG DW (���� �)LJ� �+�,� GLVSOD\HG
VWDWLVWLFDOO\ VLJQLILFDQW GLIIHUHQFHV LQ WKH QXPEHU RI 0FP��.L��� HOHFWURSRUDWHG FHOOV LQ WKH 9=
LQ WKH 6R[� RYHUH[SUHVVLRQ FRQGLWLRQ �S&,*� ����������� S&,*)/�6�� ������������ W ���
������� S ���� ����� XQSDLUHG W WHVW� Q �� )LJ� �-�� 7KHVH GDWD VXJJHVW WKDW 6R[�
RYHUH[SUHVVLRQ LQ 5*&V FRXOG DFW E\ VORZLQJ FHOO F\FOH SURJUHVVLRQ� PDLQWDLQLQJ SDUW RI WKH
FHOOV�LQ�HDUO\�*��RU�HYHQ�NHHSLQJ�WKRVH�FHOOV�LQ�D WHPSRUDU\�TXLHVFHQW�VWDWH�
7R FRQILUP SRVVLEOH DOWHUDWLRQV LQ FHOO�F\FOH OHQJWK DW WKLV DJH XSRQ 6R[� RYHUH[SUHVVLRQ
DQG HVWLPDWH WKH FHOO F\FOH GXUDWLRQ ZH XVHG D GXDO�SXOVH WK\PLGLQH DQDORJ ODEHOLQJ VWUDWHJ\
DFFRUGLQJ WR SXEOLVKHG PHWKRGV �+DUULV HW DO�� ������ :H LQMHFWHG %UG8 LQ (���� SUHJQDQW
IHPDOHV ZKRVH HPEU\RV ZHUH HOHFWURSRUDWHG DW (���� ZLWK 3&,* RU 3&,*�)/ 6R[� �KRXU DQG D
KDOI EHIRUH WKH DQDO\VLV� $ VHFRQG LQMHFWLRQ XVLQJ (G8 ZDV DGPLQLVWHUHG KDOI KRXU EHIRUH WLVVXH
FROOHFWLRQ �)LJ� �$¶¶�� *)3� FHOOV SUHVHQW LQ WKH 9= ZHUH TXDQWLILHG EDVHG RQ %UG8 DQG (G8
H[SUHVVLRQ WR HVWLPDWH WKH GXUDWLRQ RI 6�SKDVH �7V� DQG WRWDO FHOO F\FOH GXUDWLRQ �7F� �)LJ�
�.�/�� $V H[SHFWHG� VXEVWDQWLDO GHFUHDVHV LQ WKH QXPEHU RI GRXEOH ODEHOHG %UG8�(G8 *)3�
5*&V DQG %UG8 VLQJOH ODEHOHG *)3� FHOOV ZHUH IRXQG XSRQ 6R[� RYHUH[SUHVVLRQ� 7KH 7V
FDOFXODWHG DV D IUDFWLRQ RI GRXEOH ODEHOHG YHUVXV %UG8 VLQJOH ODEHOHG 5*&V DV GHVFULEHG
�0DUW\QRJD HW DO�� ����� +DUULV HW DO�� ������ ZDV VLJQLILFDQWO\ LQFUHDVHG LQ WKH RYHUH[SUHVVLRQ
FRQGLWLRQ �S&,*� ���������K� S&,*)/�6�� ���������K� W ��� ������ S ������� XQSDLUHG W WHVW�
��

)DEUD�%HVHU�HW�DO�
Q �� )LJ� �0�� 7KH 7F ZDV FDOFXODWHG XVLQJ WKH 7V GXUDWLRQ DQG WKH WRWDO QXPEHU RI 5*&V LQ
6�SKDVH� %HFDXVH FDOFXODWLRQ RI WKH 7F DVVXPHV WKDW DOO RI WKH FHOOV DUH F\FOLQJ� ZH FDQ DSSO\ D
FRUUHFWLRQ IDFWRU �JURZWK IDFWLRQ� DFFRUGLQJ WR WKH NQRZQ SURSRUWLRQ RI F\FOLQJ FHOOV� 8VLQJ
0FP� H[SUHVVLRQ LQ *)3� 5*&V DV D PDUNHU RI F\FOLQJ FHOOV ZH ILQG WKDW WKH GXUDWLRQ RI WKH
FHOO F\FOH LV WKUHH WLPHV ORQJHU LQ 5*&V RYHUH[SUHVVLQJ 6R[� ZLWK UHVSHFW WR WKH FRQWURO �S&,*�
����������K� S&,*)/�6�� ����������K� W ��� ������ S �������� XQSDLUHG W WHVW� Q �� )LJ�
�0�� ,I ZH XVH WKH SHUFHQWDJH RI .L��� WDUJHWHG 5*&V� EHFDXVH 0FP��.L��� FHOOV FRXOG
LQFOXGH FHOOV LQ *� LQ DGGLWLRQ WR HDUO\ *�� WKH GXUDWLRQ RI WKH FHOO�F\FOH LV DOPRVW GRXEOHG LQ
WLPH LQ WKRVH 5*&V LQ FRPSDULVRQ ZLWK WKH FRQWURO VLWXDWLRQ �S&,*� ����������K� S&,*)/�6��
����������K� W ��� ������ S ������� XQSDLUHG W WHVW� Q �� ([WHQGHG 'DWD )LJ� ���0��
$OWRJHWKHU WKH GDWD VXJJHVW WKH H[LVWHQFH RI LPSRUWDQW DOWHUDWLRQV LQ WKH FHOO F\FOH G\QDPLFV
PHGLDWHG E\ WKH HOHYDWHG OHYHOV RI 6R[� WKDW VHHPV WR H[WHQG FHOO F\FOH GXUDWLRQ DW WKH DQDO\]HG
WLPHV� ([WHQGHG FHOO F\FOH LV FRPSDWLEOH ZLWK ORQJHU PDLQWHQDQFH RI WKRVH SURJHQLWRU FHOOV LQ
WKH�9=�DQG�OHVV�QHXURJHQLF�GLYLVLRQV�
6R[� 2YHUH[SUHVVLQJ 5*&V (QJDJH LQ 1HXURJHQLF 'LYLVLRQV DW /DWHU 6WDJHV RI &RUWLFDO
'HYHORSPHQW
2XU GDWD VXJJHVW WKDW 6R[� OHYHOV DIIHFW 5*& SUROLIHUDWLRQ DQG WKHLU QHXURJHQLF SRWHQWLDO� VXFK
WKDW 5*&V H[SUHVVLQJ KLJK OHYHOV RI 6R[� DUH PDLQWDLQHG DV 5*&V GXULQJ HDUO\ VWDJHV RI
QHRFRUWLFDO QHXURJHQHVLV� SUHVHQWLQJ H[WHQGHG FHOO F\FOH GXUDWLRQ� :H K\SRWKHVL]HG WKDW WKHVH
FHOOV ZRXOG JHQHUDWH QHXURQV DW ODWHU VWDJHV RI QHXURJHQHVLV� 7R WHVW WKLV K\SRWKHVLV� ZH
GHVLJQHG DQ H[SHULPHQW WR WDUJHW WKH 5*&V LQ YLYR DW WZR GLIIHUHQW HPEU\RQLF VWDJHV� :H ILUVW
FDUULHG RXW LQ XWHUR HOHFWURSRUDWLRQ DW (���� WR FR�H[SUHVV 6R[� DQG *)3 RU RQO\ *)3 LQ 5*&V�
�� KRXUV DIWHU WKH ILUVW VXUJHU\ ZH SHUIRUPHG DQ LQWUDYHQWULFXODU LQMHFWLRQ RI D
FDUER[\IOXRUHVFHLQ HVWHU DOVR NQRZQ DV )ODVK7DJ �7HOOH\ HW DO�� ����� *RYLQGDQ HW DO�� �����
�)LJ� �$�� )ODVK7DJ LQMHFWLRQ ZLOO ODEHOV 5*&V OLQLQJ WKH ODWHUDO YHQWULFOH DW (����� LQFOXGLQJ
WKRVH 5*&V WDUJHWHG DW (���� E\ LQ XWHUR HOHFWURSRUDWLRQ WKDW DUH VWLOO XQGHUJRLQJ FHOO GLYLVLRQV�
%HFDXVH 6R[� RYHUH[SUHVVLRQ DW (���� FDXVHV WKH DFFXPXODWLRQ RI PDQ\ 5*&V LQ FORVH FRQWDFW
��

)DEUD�%HVHU�HW�DO�
ZLWK WKH YHQWULFOH� YLVXDOL]HG �� KRXU ODWHU� WKRVH FHOOV ZRXOG EH SRWHQWLDOO\ ODEHOHG E\ )ODVK7DJ
DW WKH WLPH RI WKH LQMHFWLRQ� &HOOV ODEHOHG ZLWK ERWK *)3 DQG )ODVK7DJ DUH SURJHQLWRUV WKDW ZHUH
LQ FRQWDFW ZLWK WKH YHQWULFOH DW ERWK (���� DQG (����� ZKHUHDV *)3� FHOOV WKDW DUH QRW ODEHOHG
ZLWK )ODVK7DJ KDG OHIW WKH 9= E\ (����� :H WKHQ DQDO\]HG FHOO GLVWULEXWLRQ RI WKH RIIVSULQJ DW
(����� ,Q FRQWUROV� PRVW *)3� FHOOV ZHUH )ODVK7DJ�QHJDWLYH DQG DFFXPXODWHG LQ WKH ORZHU SDUW
RI WKH &3� ZKLOH )ODVK7DJ��*)3� FHOOV �EOXH� ZHUH ODUJHO\ ORFDWHG LQ WKH XSSHU &3 �)LJ� �%�&��
)ROORZLQJ 6R[� RYHUH[SUHVVLRQ� D VXEVWDQWLDO QXPEHU RI *)3� FHOOV RFFXSLHG WKH XSSHU &3� WKH
VDPH DUHD DOVR RFFXSLHG E\ )ODVK7DJ� FHOOV �)LJ� �&�'�� $QDO\VLV RI VLQJOH RSWLFDO VHFWLRQV
FRQILUPHG FR�H[SUHVVLRQ RI *)3 DQG )ODVK7DJ LQ WKHVH FHOOV �S&,* )7� LQ 8/� ���� � ���� ��
S&,*�)/6� )7� LQ 8/� ����� � ���� �� W ��� ������ S ������� XQSDLUHG W WHVW� Q �� )LJ�
�%� '�(�� VXEVWDQWLDWLQJ WKDW WKRVH QHXURQV ZHUH JHQHUDWHG IURP 5*&V WDUJHWHG DW (���� DQG
VWLOO SUHVHQW LQ WKH 9= DW (����� :H WKXV FRQFOXGH WKDW 5*&V H[SUHVVLQJ KLJK OHYHOV RI 6R[� DUH
PDLQWDLQHG�ORQJHU�DW�WKH�9=�EHIRUH�XQGHUJRLQJ�QHXURJHQLF GLYLVLRQV�
6R[��2YHUH[SUHVVLQJ�5*&V�3URGXFH�1HXURQV�'HVWLQHG IRU�8SSHU�&RUWLFDO�/D\HUV
7R IXUWKHU LQYHVWLJDWH WKH IDWH RI QHXURQV GHULYHG IURP 5*&V ZLWK KLJK 6R[� H[SUHVVLRQ OHYHOV�
ZH TXDQWLILHG WKHLU ODPLQDU SRVLWLRQV� :H FDUULHG RXW LQ XWHUR HOHFWURSRUDWLRQ DW (���� WR
RYHUH[SUHVV 6R[� ZLWK *)3 RU *)3 DORQH DQG GHWHUPLQHG WKH ODPLQDU SRVLWLRQ RI WKH QHXURQDO
RIIVSULQJ RI HOHFWURSRUDWHG 5*&V DW (���� DQG 3��� )ROORZLQJ 6R[� RYHUH[SUHVVLRQ� PRUH
QHXURQV RFFXSLHG WKH XSSHU SDUW RI WKH &3 FRPSDUHG WR FRQWUROV DW (����� H[SUHVVLQJ WKH XSSHU
OD\HU PDUNHU &X[� �S&,* YV S&,*�)/6�� ELQ �� S ������� ELQ �� S ������� ELQ �� S
������� ELQ �� S ������� ELQ �� S ������� ELQ �� S ������� ELQ ��� S ������� XQSDLUHG W
WHVW� Q �� )LJ� �%�&�� $W 3��� ZKHQ FHOO PLJUDWLRQ LV FRPSOHWH� WKHVH QHXURQV H[SUHVVLQJ &X[�
RFFXSLHG XSSHU QHRFRUWLFDO FHOO OD\HUV �S&,*� ����� � ������ S&,*�)/6�� ���� � ������ W ���
����� S ���������XQSDLUHG�W�WHVW��Q� ����)LJ���'�(��
6LQFH 6R[� RYHUH[SUHVVLRQ VHHPV WR LQFUHDVH WKH SURGXFWLRQ QHXURQV RFFXS\LQJ WKH XSSHU
OD\HUV� ZH GHFLGHG WR FKHFN WKH LQIOXHQFH RI 6R[� GRZQUHJXODWLRQ LQ WKH ODPLQDU ORFDOL]DWLRQ RI
+HV��5*&V RIIVSULQJ� 7R WKLV HQG� ZH SHUIRUPHG (���� LQ XWHUR HOHFWURSRUDWLRQ ZLWK WKH VDPH
��

)DEUD�%HVHU�HW�DO�
6R[� RU VFUDPEOHG VK51$ SUHYLRXVO\ VKRZQ �([WHQGHG 'DWD )LJ� ���� WRJHWKHU ZLWK D
+HV��*)3 YHFWRU� 7KH DQDO\VLV RI WKH ODPLQDU SRVLWLRQ RI WDUJHWHG FHOOV DW (���� VKRZHG
LPSRUWDQW FKDQJHV LQ WKH GLVWULEXWLRQ RI WKH SURGXFHG QHXURQV� 6LJQLILFDQW UHGXFWLRQV LQ WKH
QXPEHU RI *)3� QHXURQV ZHUH IRXQG LQ WKH XSSHU SDUW RI WKH &3 LQ FRPSDULVRQ ZLWK WKH FRQWURO
VLWXDWLRQ �&56K51$ YV 6�6K51$� ELQ �� S ������ ELQ �� S ������� ELQ �� S ������ ELQ ��
S �������� ELQ �� S �������� ELQ �� S �������� ELQ �� S ������� XQSDLUHG W WHVW� Q ��
([WHQGHG 'DWD )LJ� ���%�� 7KHVH UHVXOWV DUH FRQVLVWHQW ZLWK WKH PRGHO WKDW WKH FRUWLFDO 9=
FRQWDLQV D VXEVHW RI 5*&V H[SUHVVLQJ KLJK OHYHOV RI 6R[�� ZKLFK DUH PDLQWDLQHG DV SURJHQLWRUV
GXULQJ�HDUO\�VWDJHV�RI�QHXURJHQHVLV�DQG�JHQHUDWH�XSSHU OD\HU�QHXURQV�ODWHU�RQ�GHYHORSPHQW�
',6&866,21
:H SURYLGH KHUH HYLGHQFH RI WKH H[LVWHQFH RI PROHFXODU GLIIHUHQFHV LQ WKH SRRO RI QHRFRUWLFDO
5*&V DW HDUO\ HPEU\RQLF DJHV� 6SHFLILFDOO\� ZH LGHQWLILHG WKH PROHFXODU VLJQDWXUH RI 5*&V
SUHVHQWLQJ D PRUH GHOD\HG QHXURJHQLF EHKDYLRU DW WKHVH DJHV WKDQ RWKHU SURJHQLWRU FHOOV� )URP
WKH �� GLIIHUHQWLDOO\ H[SUHVVHG JHQHV EHWZHHQ WKH WZR VXESRSXODWLRQV ZH LGHQWLILHG 6R[� DV D
PDVWHU UHJXODWRU RI WKHVH SURJHQLWRUV� 2XU IXQFWLRQDO H[SHULPHQWV PRGLI\LQJ 6R[� H[SUHVVLRQ
UHYHDOHG WKDW 6R[� OHYHOV SOD\ D FULWLFDO UROH LQ UHJXODWLQJ WKH GHFLVLRQ RI QHRFRUWLFDO SURJHQLWRUV
WR EH PDLQWDLQHG DV 5*&V RU WR XQGHUJR QHXURJHQLF GLYLVLRQV� 2XU GDWD VXJJHVW WKDW KLJK OHYHOV
RI 6R[� FRXOG DFW H[WHQGLQJ FHOO F\FOH GXUDWLRQ RI SDUW RI 5*&V� GHOD\LQJ QHXURJHQHVLV GXULQJ
HDUO\ VWDJHV RI FRUWLFDO GHYHORSPHQW� WR WKHQ JHQHUDWH QHXURQV GXULQJ ODWHU VWDJHV RI QHRFRUWLFDO
GHYHORSPHQW�
5ROHV IRU 6R[� LQ WKH FRQWURO RI QHXUDO VWHP FHOOV PDLQWHQDQFH DQG GLIIHUHQWLDWLRQ KDYH
DOUHDG\ EHHQ GHVFULEHG LQ VHYHUDO SDUWV RI WKH EUDLQ� LQFOXGLQJ WKH FHUHEHOOXP �9RQJ HW DO��
������ VSLQDO FRUG �6WROW HW DO�� ����� DQG WKH SRVWQDWDO VXEYHQWULFXODU ]RQH �&KHQJ HW DO�� ������
2XU GDWD LQGLFDWH WKDW 6R[� LV LPSRUWDQW LQ WKH UHJXODWLRQ RI QHXUDO VWHP FHOO VHOI�UHQHZDO LQ WKH
HDUO\ HPEU\RQLF QHRFRUWH[� ,QWHUHVWLQJO\� 6R[� LV H[SUHVVHG ZLGHO\ LQ DOO 5*&V� DQG OLQHDJH�
WUDFLQJ H[SHULPHQWV XVLQJ 6R[��&UH(57� PLFH VKRZHG WKDW QHXURQV RI DOO QHRFRUWLFDO FHOO
OD\HUV ZHUH ODEHOHG �.DSODQ HW DO�� ������ 7KHVH GDWD VXJJHVW WKDW WKH OHYHOV RI 6R[� LQ 5*&V DUH
��

)DEUD�%HVHU�HW�DO�
OLNHO\ PRUH LPSRUWDQW WKDQ SUHVHQFH RU DEVHQFH� ,QGHHG� D VLPLODU GRVH�GHSHQGHQW UHJXODWLRQ RI
5*& QHXURJHQLF EHKDYLRU KDV EHHQ GHVFULEHG IRU 6R[�� DQRWKHU WUDQVFULSWLRQ IDFWRU RI WKH VDPH
IDPLO\ DV 6R[� �+DJH\ DQG 0XKU� ������ +LJKHU OHYHOV RI 6R[� KDYH EHHQ IRXQG LQ
VORZ�F\FOLQJ 5*&V� DQG 6R[� RYHUH[SUHVVLRQ E\ LQ XWHUR HOHFWURSRUDWLRQ UHGXFHV WKHLU
SUROLIHUDWLRQ�UDWH�
,Q WKH DGXOW VXEYHQWULFXODU ]RQH� LQFUHDVHG 6R[� OHYHOV DUH D PDUNHU RI TXLHVFHQW 16&V
�/ORUHQV�%REDGLOOD HW DO�� ������ +RZHYHU� WKHVH SURJHQLWRUV QHHG WR EH DFWLYDWHG WR JHQHUDWH
QHXURQV DW DSSURSULDWH WLPHV� VLPLODU WR WKH DFWLYDWLRQ RI 9= SURJHQLWRUV WR VZLWFK IURP D
VHOI�UHQHZDO VWDWH GXULQJ HDUO\ SKDVHV RI FRUWLFRJHQHVLV WR QHXURJHQLF GLYLVLRQV GXULQJ ODWH
VWDJHV RI FRUWLFDO GHYHORSPHQW� 1RWHZRUWKLO\� RQH RI WKH SDUDPHWHUV GHVFULEHG WR FKDQJH LQ WKH
WUDQVLWLRQ EHWZHHQ VHOI�UHQHZLQJ WR QHXURJHQLF SURJHQLWRUV LV WKH GXUDWLRQ RI 6�SKDVH �$UDL HW
DO�� ����� 7DYHUQD HW DO�� ������ 2XU IXQFWLRQDO DQDO\VLV FRQILUPHG WKDW 5*&V RYHUH[SUHVVLQJ
6R[� SUHVHQW HQODUJHG 6�SKDVH GXUDWLRQ� LQ FRPSDULVRQ ZLWK WKH FRQWURO VLWXDWLRQ� DQG SURJUHVV
PRUH VORZO\ WKURXJK WKH FHOO F\FOH� +RZHYHU� HYHQWXDOO\ WKHVH FHOOV HQJDJH LQ QHXURJHQLF
GLYLVLRQV SURGXFLQJ LQFUHDVHG QXPEHU RI 8/ QHXURQV� WKDW DUH DEOH WR SURSHUO\ PLJUDWH�
VXJJHVWLQJ WKDW HOHYDWHG 6R[� OHYHOV DUH QRW DIIHFWLQJ WKH PLJUDWLRQ SURFHVV� +RZ WKH
QHXURJHQLF EUDNH LV RYHUFRPH� DQG ZKLFK PROHFXODU SDWKZD\V DUH LQYROYHG LV XQFOHDU� 2QH
LQWHUHVWLQJ PROHFXODU SDWKZD\ LV WKH <DS�+LSSR SDWKZD\� +LSSR VLJQDOLQJ KDV EHHQ UHODWHG ZLWK
WLVVXH KRPHRVWDVLV� RUJDQ VL]H DQG VWHP FHOO VHOI�UHQHZDO �3DQ� ����� 6FKOHJHOPLOFK HW DO�� �����
=DQFRQDWR HW DO�� ������ $ UHFHQW UHSRUW KDV GHVFULEHG WKDW <DS��7D] SURPRWHV 16&
PDLQWHQDQFH LQ WKH FRUWH[ DQG WKH SURGXFWLRQ RI FRUWLFR�FRUWLFDO QHXURQV DW WKH H[SHQVH RI
FRUWLFRIXJDO QHXURQV �0XNKWDU HW DO�� ������ ,QWHUHVWLQJO\� 6R[� KDV EHHQ GHVFULEHG DV D
GRZQVWUHDP WDUJHW RI <DS� VLJQDOLQJ DQG <DS��WDUJHWLQJ PLFUR51$V� LQGXFHG E\ 6R[��
SRVW�WUDQVFULSWLRQDOO\ UHSUHVV <DS� H[SUHVVLRQ� FRQWULEXWLQJ WR D QHJDWLYH IHHGEDFN ORRS �:DQJ
HW DO�� ������ <DS� H[SUHVVLRQ LV KLJK DW HDUO\ FRUWLFRJHQHVLV VWDJHV DQG LWV H[SUHVVLRQ GHFUHDVHV
DW PLG FRUWLFRJHQHVLV �(����(����� WR LQFUHDVH DJDLQ DW HDUO\ SRVWQDWDO DJHV �0XNKWDU HW DO��
������ ,W ZLOO EH LQWHUHVWLQJ WR WHVW LQ WKH IXWXUH ZKHWKHU WKH GHFUHDVH LQ <DS� H[SUHVVLRQ GXULQJ
PLG�QHXURJHQHVLV�LV�FULWLFDO�IRU�GHWHUPLQLQJ�6R[� OHYHOV�DQG�WKH�QHXURJHQLF�SRWHQWLDO�RI�5*&V�
��

)DEUD�%HVHU�HW�DO�
7KH H[WHQW LQ ZKLFK GLIIHUHQW VXESRSXODWLRQV RI 5*&V FRQWULEXWH GLUHFWO\ WR WKH JHQHUDWLRQ
RI FHOOXODU GLYHUVLW\ LQ WKH QHRFRUWH[ LV D FRQWURYHUVLDO WRSLF �)UDQFR HW DO�� ����� )XHQWHDOED HW
DO�� ����� *DUFtD�0RUHQR DQG 0ROQiU� ����� *LO�6DQ] HW DO�� ����� � 7KH UHVXOWV VKRZQ KHUH
UHYHDO WKH H[LVWHQFH RI PROHFXODU KHWHURJHQHLW\ LQ 5*&V DVVRFLDWHG ZLWK WKH SURGXFWLRQ RI
SDUWLFXODU FHOO W\SHV� $ SUHYLRXVO\ SXEOLVKHG VF51$ VHTXHQFLQJ VWXG\ GLG QRW GHWHFW VXFK
PROHFXODU GLYHUVLW\ �7HOOH\ HW DO�� ������ +RZHYHU� WKLV VWXG\ XVHG D GLIIHUHQW PHWKRG WR ODEHO
DQG LVRODWH 5*&V� LQVWHDG RI LQ XWHUR HOHFWURSRUDWLRQ� WKH\ XVHG )ODVK7DJ WHFKQRORJ\� 1RWDEO\�
)ODVK7DJ KDV EHHQ GHVFULEHG WR RQO\ ODEHO DFWLYHO\ GLYLGLQJ 5*&V LQ 0�SKDVH� EHFDXVH WKRVH
FHOOV PDLQWDLQ ODUJH FRQWDFW VXUIDFH ZLWK WKH YHQWULFOH �*RYLQGDQ HW DO�� ������ $W HDUO\
HPEU\RQLF DJHV �(������ WKH SURJHQ\ RI WKHVH ODEHOHG FHOOV UDSLGO\ H[LWV WKH 9= WR SURGXFH ,3V
RU (1V �7HOOH\ HW DO�� ����� ������ 7KXV� �� KRXUV DIWHU )ODVK7DJ ODEHOLQJ DV FRQGXFWHG LQ WKLV
VWXG\� PRVW RI WKH ODEHOHG FHOOV ZHUH IRXQG LQ WKH 69=�,=� ,Q FRQWUDVW� RXU LQ XWHUR
HOHFWURSRUDWLRQ DSSURDFK DW WKH VDPH DJHV XVLQJ GLIIHUHQW SURPRWHU IUDJPHQWV DOORZHG WKH
LGHQWLILFDWLRQ RI ODWH QHXURJHQLF 5*&V WKDW FDQ EH YLVXDOL]HG LQ WKH 9= HYHQ �� KRXUV DIWHU
ODEHOLQJ E\ HOHFWURSRUDWLRQ� ZKLFK SURGXFH QHXURQV DW ODWHU DJHV� 7KHVH FHOOV ZRXOG KDYH EHHQ
PLVVHG LQ WKH SUHYLRXV VWXG\� JLYHQ WKDW )ODVK7DJ DSSDUHQWO\ GRHV QRW ODEHO VORZO\ GLYLGLQJ
5*&V� WKDW ZRXOG QRW EH LQ 0�SKDVH� 1RWDEO\� RXU GDWD DOVR VKRZ WKDW WKHVH SURJHQLWRUV KDYH D
VSHFLILF PROHFXODU VLJQDWXUH ZLWK �� JHQHV EHLQJ PRUH KLJKO\ H[SUHVVHG� LQFOXGLQJ 6R[�� :KLOH
ZH GHVFULEH KHUH D IXQFWLRQ IRU 6R[� IRU WKH UHJXODWLRQ RI GLIIHUHQWLDO SURJHQLWRU EHKDYLRU� WKH
IXQFWLRQ�RI�WKH�UHPDLQLQJ����JHQHV�VWLOO�QHHGV�WR EH�LQYHVWLJDWHG�
,Q VXPPDU\� ZH GHPRQVWUDWHG KHUH WKH H[LVWHQFH RI PROHFXODU KHWHURJHQHLW\ DPRQJ
QHRFRUWLFDO SURJHQLWRUV� ZKLFK GLVWLQJXLVKHV 5*&V ZLWK GLIIHUHQW QHXURJHQLF DFWLYLW\ DW HDUO\
DJHV RI FRUWLFDO GHYHORSPHQW� $PRQJ WKH GLIIHUHQWLDOO\ H[SUHVVHG JHQHV WKDW ZH IRXQG� 6R[� KDV
DQ LQVWUXFWLYH UROH LQ GHILQLQJ QHXURJHQLF SRWHQWLDO RI 5*&V� 7KH OHYHOV RI H[SUHVVLRQ RI WKLV
JHQH DUH IXUWKHU DVVRFLDWHG ZLWK WKH SURGXFWLRQ RI GLVWLQFW FHOOV W\SHV� ZLWK KLJKHU OHYHOV RI 6R[�
LQVWUXFWLQJ WKH SURGXFWLRQ RI XSSHU OD\HU QHXURQV� 7KHVH UHVXOWV VXJJHVW WKDW WKH GHOD\HG
QHXURJHQLF EHKDYLRU LQVWUXFWHG E\ 6R[� LV LPSRUWDQW IRU SUHYHQWLQJ VRPH 5*&V IURP HDUO\
H[KDXVWLRQ� DOORZLQJ WKHP WR JHQHUDWH XSSHU OD\HU QHXURQV DW ODWHU HPEU\RQLF DJHV� %HFDXVH WKH
��

)DEUD�%HVHU�HW�DO�
GLIIHUHQWLDO QHXURJHQLF EHKDYLRU RI WKHVH 5*&V DQG WKHLU SDUWLFXODU PROHFXODU VLJQDWXUH KDYH
EHHQ RQO\ DQDO\]HG DW RQH SDUWLFXODU DJH� DGGLWLRQDO ZRUN ZLOO EH UHTXLUHG LQ RUGHU WR IXOO\
FODULI\ ZKHWKHU WKLV PROHFXODU VLJQDWXUH LV PDLQWDLQHG RU QRW WKURXJKRXW DOO HPEU\RQLF
GHYHORSPHQW�
��

)DEUD�%HVHU�HW�DO�
),*85(�/(*(1'6
)LJXUH �� ,Q XWHUR HOHFWURSRUDWLRQ VWUDWHJ\ WR LGHQWLI\ 5*& VXESRSXODWLRQV� �$� 6FKHPDWLFV
RI WKH HOHFWURSRUDWLRQ VWUDWHJ\ IRU ODEHOLQJ GLIIHUHQW VXESRSXODWLRQV RI 5*&V� 3520� SURPRWHU�
)3� IOXRUHVFHQFH SURWHLQ� �%� 7ULSOH HOHFWURSRUDWLRQ DW (���� RI S&$*�%)3 �EOXH�� :QW UHSRUWHU
SODVPLG 723�*)3 �JUHHQ� DQG 1RWFK UHSRUWHU SODVPLG +HV�S�GV5HG �UHG�� DQDO\VLV DW (�����
1RWH WKH VLPLODU GLVWULEXWLRQ RI ODEHOHG FHOOV XVLQJ WKH JHQHUDO SURPRWHU �S&$*�� DQG WKH :QW
UHSRUWHU SODVPLG VKRZLQJ D QRWRULRXV QXPEHU RI FHOOV LQ WKH 69=� 7KH 1RWFK UHSRUWHU SODVPLG
DOORZHG WKH ODEHOLQJ RI FHOOV PRVWO\ SUHVHQW LQ WKH 9=� �&� 7ULSOH HOHFWURSRUDWLRQ DW (���� RI
S&$*�%)3 �EOXH�� 1RWFK UHSRUWHU SODVPLGV +HV�S�GV5HG �UHG�� DQG &%)5(�*)3 �JUHHQ��
DQDO\VLV DW ����� 1RWH &%))�5( DQG +HV� SURPRWHU IUDJPHQWV GULYH D VLPLODU ODEHOLQJ SDWWHUQ�
�'� &R�HOHFWURSRUDWLRQ RI +HV�S�GV5HG �UHG� DQG (P[�S�*)3 �JUHHQ� DW (����� DQDO\VLV DW
����� 1RWH WKH YLUWXDO FR�ORFDOL]DWLRQ RI ERWK UHSRUWHU SURWHLQV LQ FHOOV DFWLYDWLQJ WKHVH SURPRWHU
IUDJPHQWV� �(� 4XDQWLILFDWLRQ �PHDQ 6(0� RI HOHFWURSRUDWHG �(3� FHOOV LQ WKH 9= DW ���� ZLWKb
GLIIHUHQW H[SUHVVLRQ YHFWRUV H[KLELWLQJ GLIIHUHQFHV LQ WKHLU QHXURJHQLF EHKDYLRU� � 3 �������
�)�*� ,PPXQRVWDLQLQJ �JUHHQ� DW (���� IRU 3D[� �)� DQG 7EU� �*� RI (���� +HV��GV5HG �UHG�
HOHFWURSRUDWHG HPEU\RV� �)� $UURZV SRLQW H[DPSOHV RI GV5HG��3D[�� FHOOV� �*� ([DPSOHV RI
GV5HG��7EU�� FHOOV �DUURZV� DQG GV5HG��7EU�� FHOOV �DUURZKHDGV�� &3� FRUWLFDO SODWH� 69=�
VXEYHQWULFXODU�]RQH��9=��YHQWULFXODU�]RQH��6FDOH�EDUV� ����P�
)LJXUH �� /DPLQDU ORFDOL]DWLRQ RI QHXURQV JHQHUDWHG IURP HDUO\ 5*&V ZLWK GHOD\HG
QHXURJHQLF SRWHQWLDO� �$� 6FKHPDWLFV RI WKH HOHFWURSRUDWLRQ VWUDWHJ\ IRU ODPLQDU ORFDOL]DWLRQ
GHWHUPLQDWLRQ� �%� &R�HOHFWURSRUDWLRQ RI WKH S&$*�%)3 �EOXH� DQG +HV�S�GVUHG �UHG�
FRQVWUXFWV DW (����� DQDO\VLV DW 3�� �&� 4XDQWLILFDWLRQ �PHDQ 6(0� RI WKH GLVWULEXWLRQ RIb
HOHFWURSRUDWHG FHOOV LQ WKH &3 DW 3� IURP HDFK VXESRSXODWLRQ RI SURJHQLWRUV� )RU TXDQWLILFDWLRQ�
WKH &3 ZDV GLYLGHG LQ WHQ HTXDO�VL]H ELQV �HQXPHUDWHG ���� IURP EDVDO WR DSLFDO�� � 3 � ����
� 3 � ������ �'�*� ,PPXQRVWDLQLQJ �JUHHQ� DW 3� IRU &X[� �'�� 6DWE� �(�� &WLS� �)� DQG 7OH�
�*�� RI (���� +HV�S�GV5HG �UHG� HOHFWURSRUDWHG HPEU\RV� �+� 4XDQWLILFDWLRQ �PHDQ 6(0� DWb
��

)DEUD�%HVHU�HW�DO�
3� RI +HV�S�GV5HG HOHFWURSRUDWHG FHOOV H[SUHVVLQJ &X[�� 6DWE�� &WLS� RU 7OH�� ,,�9,� FRUWLFDO
OD\HUV������:0��ZKLWH�PDWWHU��//��ORZHU�OD\HUV��8/� XSSHU�OD\HUV��6FDOH�EDUV�������P�
)LJXUH �� /LQHDJH WUDFLQJ RI HDUO\ 5*&V DFWLYDWLQJ +HV� SURPRWHU� �$� 6FKHPDWLFV RI WKH
OLQDJH WUDFLQJ VWUDWHJ\� $L� UHSRUWHU HPEU\RV ZHUH HOHFWURSRUDWHG DW (���� ZLWK &$*�&UH(57�
DQG +HV��&UH(57� FRQVWUXFWV� 3UHJQDQW GDPV ZHUH LQMHFWHG ZLWK ��2+7 ULJKW DIWHU
HOHFWURSRUDWLRQ� 3XSV ZHUH GHOLYHUHG DW (���� E\ &DHVDUHDQ VHFWLRQ� %UDLQV ZHUH DQDO\]HG DW
3��� �%� &RQIRFDO LPDJHV DQG TXDQWLILFDWLRQ �PHDQ 6(0� RI WKH GLVWULEXWLRQ RI WG7RPDWR�b
FHOOV DW 3��� 1RWH PRVW WG7RPDWR� FHOOV LQ WKH +HV�S�&UH(57� HOHFWURSRUDWHG EUDLQV ZHUH
SUHVHQW LQ WKH XSSHU OD\HUV� ZKHUHDV &$*�&UH(57� VKRZ D PRUH KRPRJHQHRXV GLVWULEXWLRQ�
)RU TXDQWLILFDWLRQ� WKH &3 ZDV GLYLGHG LQ WHQ HTXDO�VL]H ELQV �HQXPHUDWHG ���� IURP EDVDO WR
DSLFDO�� � 3 � ���� � 3 � ������ �&� 'RXEOH LPPXQRIOXRUHVFHQFH IRU &X[� �JUHHQ� DQG &WLS�
�EOXH� RQ +HV�S�&UH(57� �UHG� HOHFWURSRUDWHG EUDLQV VKRZLQJ PDJQLILFDWLRQ LQ WKH XSSHU
OD\HUV� $UURZV SRLQW H[DPSOHV RI WG7RPDWR��&X[�� FHOOV� �'� 'RXEOH LPPXQRIOXRUHVFHQFH IRU
6DWE� �JUHHQ� DQG &WLS� �EOXH� RQ +HV��&UH(57� �UHG� HOHFWURSRUDWHG EUDLQV VKRZLQJ
PDJQLILFDWLRQ LQ WKH ORZHU OD\HUV� $UURZV SRLQW H[DPSOHV RI WG7RPDWR��6DWE�� FHOOV� �(�
4XDQWLILFDWLRQ �PHDQ 6(0� RQ S&$*�&UH(57� DQG +HV�S�&UH(57� HOHFWURSRUDWHG EUDLQVb
RI UHFRPELQHG WG7RPDWR� FHOOV H[SUHVVLQJ &X[�� 6DWE� RU &WLS�� � 3 � ���� � 3 � �������
$EEUHYLDWLRQV DV LQ )LJXUH �� 6FDOH EDUV� % ��� �P� &�' ��� �P ORZ PDJQLILFDWLRQ DQG �� �P
ER[HG�DUHDV�
)LJXUH �� 6LQJOH�FHOO VRUWLQJ� VHTXHQFLQJ DQG VF51$VHT LGHQWLILFDWLRQ DQDO\VLV� �$�
,PPXQRVWDLQLQJ IRU 3D[� �EOXH� RQ (3 (���� HOHFWURSRUDWHG HPEU\RV ZLWK WKH GXDO SURPRWRU
SODVPLG S&$*�P5)3� +HV�S�*)3 �UHG DQG JUHHQ UHVSHFWLYHO\�� DQDO\]HG �� KRXUV DIWHU
HOHFWURSRUDWLRQ� �%� +LJKHU�PDJQLILFDWLRQ YLHZ RI ER[HG DUHD LQ �$�� $UURZKHDGV SRLQW
P5)3��*)3� FHOOV DQG DUURZV SRLQW P5)3��*)3� FHOOV� �&� 6FKHPDWLFV RI VF51$VHT VWUDWHJ\�
7KH HOHFWURSRUDWHG DUHD ZDV LVRODWHG DQG FHOOV ZHUH LQGLYLGXDOO\ VHSDUDWHG E\ )$&6 �� KRXUV
DIWHU HOHFWURSRUDWLRQ� ,VRODWHG FHOOV ZHUH VHTXHQFHG DQG DQDO\]HG XVLQJ ELRLQIRUPDWLFV WRROV� �'�
��

)DEUD�%HVHU�HW�DO�
W�61( YLVXDOL]DWLRQV RI WKH VRUWHG FHOOV GLVSOD\LQJ JHQH H[SUHVVLRQ �JUD\ WR GDUN EOXH FRORU
JUDGLHQW� RI FHOO�W\SH VSHFLILF PDUNHUV IRU 5*&V �+HV�� 3D[�� 1HV�� ,3V �(RPHV DQG +HV��� DQG
HDUO\ QHXURQV �1HXURG�� 7XEE�� 'F[ DQG 6QDS��� �(� W�61( YLVXDOL]DWLRQV RI WKH VF51$�VHT
GDWDVHW DFFRUGLQJ WR H[SUHVVLRQ RI WKH FHOO�W\SH VSHFLILF PDUNHUV �'� DQG SDUWLFXODU FHOO
SRSXODWLRQV LGHQWLILHG �%OXH� 5*&V� *UHHQ� ,3V� 5HG� (DUO\ 1HXURQV� �)� W�61( YLVXDOL]DWLRQV
RI WKH VF51$�VHT GDWDVHW H[KLELWLQJ &$*� FHOO JURXS �P5)3�� UHG� YV� +HV�� FHOO JURXS
�P5)3�� *)3�� JUHHQ�� �*� 3URSRUWLRQ DQDO\VLV RI &$*� DQG +HV�� FHOOV LQ WKH WKUHH FHOO
W\SHV JURXSV� +HV�� FHOOV DFFRXQWHG IRU WKH PDMRULW\ RI FHOOV ZLWK DQ 5*& WUDQVFULSWLRQDO
VLJQDWXUH ZKHUHDV &$*� FHOOV FRQVWLWXWHG WKH YDVW PDMRULW\ RI (1V� (1� HDUO\ QHXURQ� )$&6�
IOXRUHVFHQFH�DFWLYDWHG FHOO VRUWLQJ� ,3� LQWHUPHGLDWH SURJHQLWRU� 5*&� UDGLDO JOLDO FHOO� 69=�
VXEYHQWULFXODU�]RQH��9=��YHQWULFXODU�]RQH��6FDOH�EDUV� $������P�DQG�%������P�
)LJXUH �� 7UDQVFULSWLRQDO DQDO\VLV UHYHDOV GLIIHUHQWLDOO\ H[SUHVVHG JHQHV DQG PDVWHU
UHJXORQV� �$� +HDWPDS RI GLIIHUHQWLDOO\ H[SUHVVHG JHQHV �URZV� LQ 5*&V EHORQJLQJ WR RQH RI
WKH VXEJURXSV FRPSDULVRQV �+HV�� YV� &$*�� �FROXPQV�� *HQH H[SUHVVLRQ OHYHOV DUH FRORUHG
UDQJLQJ IURP WZR�IROG GRZQUHJXODWLRQ �SLQN� WR WZR�IROG XSUHJXODWLRQ �\HOORZ�� DFFRUGLQJ WR
WKH FRORU NH\� 2QO\ JHQHV ZLWK S DGMXVWHG YDOXH � ���� DQG ORJ IROG FKDQJH ! ��� DUH VKRZQ� �%�
7UDQVFULSWLRQDO UHJXODWRU\ QHWZRUN �UHJXORQV� RI VF51$�VHT GDWD H[KLELWV 6R[� DV D PDVWHU
UHJXODWRU JHQH VSHFLILF RI +HV�� 5*&V� &LUFOHV DUH UHJXORQV DQG GLDPRQG PDVWHU UHJXORQV�
&LUFOH VL]H LV UHODWHG WR QXPEHU RI JHQHV RUFKHVWUDWHG E\ LWV WUDQVFULSWLRQ IDFWRU� *HQH
H[SUHVVLRQ OHYHOV DUH FRORUHG DFFRUGLQJ WR DYHUDJH H[SUHVVLRQ RI 5*&V� 7UHH QHWZRUN UHSUHVHQW
FRUUHODWLRQ DPRQJ UHJXORQV� �&� 6FKHPDWLF LOOXVWUDWLRQ RI WKH HOHFWURSRUDWLRQ VWUDWHJ\ WR YHULI\
6R[� H[SUHVVLRQ GLIIHUHQFHV LQGLFDWHG E\ VF51$�6HT DQDO\VLV� (���� HPEU\RV ZHUH
HOHFWURSRUDWHG ZLWK WKH GXDO SURPRWRU SODVPLG &$*�P5)3�+HV��*)3 �UHG DQG JUHHQ�
UHVSHFWLYHO\�� DQDO\]HG ��K DIWHU HOHFWURSRUDWLRQ� �'� ,PPXQRVWDLQLQJ IRU 6R[� �EOXH� DIWHU
&$*�P5)3�+HV��*)3 HOHFWURSRUDWLRQ �UHG�JUHHQ�� �'¶�'¶¶� +LJKHU�PDJQLILFDWLRQ RI ' ER[HG
DUHDV� ([DPSOHV RI +HV�� 5*&V �DUURZKHDGV� DQG +HV�� 5*&V �DUURZV�� 1RWH WKDW +HV��
5*&V VKRZ KLJKHU OHYHOV RI 6R[� LPPXQRVWDLQLQJ� 6PDOO DUURZKHDGV SRLQW WR WKH EDVDO SURFHVV
��

)DEUD�%HVHU�HW�DO�
RI DQ 5*&� �(� 4XDQWLILFDWLRQ �PHDQ 6(0� RI WKH 0HDQ RI )OXRUHVFHQW ,QWHQVLW\ �0),� RIb
6R[� LQ &$*� YV +HV�� 5*&V� 0), LQ D VFDOH RI ����� DUELWUDU\ XQLWV �D�X��� � 3 � ������� &3�
FRUWLFDO SODWH� (1� HDUO\ QHXURQ� ,3� LQWHUPHGLDWH SURJHQLWRU� ,=� LQWHUPHGLDWH ]RQH� 5*&� UDGLDO
JOLDO FHOO� 69=� VXEYHQWULFXODU ]RQH� 9=� YHQWULFXODU ]RQH� 6FDOH EDUV� '� �� �P DQG '¶�'¶¶ ��
�P�
)LJXUH �� 6R[� RYHUH[SUHVVLRQ DIIHFWV QHXURJHQLF EHKDYLRU RI 5*&V� �$� 6FKHPDWLFV RI WKH
HOHFWURSRUDWLRQ VWUDWHJ\� (���� HPEU\RV ZHUH HOHFWURSRUDWHG ZLWK WKH S&,*�)/ 6R[� SODVPLG� WR
RYHUH[SUHVV 6R[�� DQG LWV HPSW\ YHUVLRQ� WKH S&,* SODVPLG �*)3 RQO\�� ZDV XVHG DV FRQWURO�
%UDLQV ZHUH DQDO\]HG �� KRXUV DIWHU HOHFWURSRUDWLRQ� �%� &RQIRFDO LPDJHV RI (���� HPEU\R¶V
QHRFRUWH[ DIWHU S&,* RU S&,*�)/ 6R[� HOHFWURSRUDWLRQ �JUHHQ�� 1RWH WKDW 6R[� RYHUH[SUHVVLRQ
FDXVHG D ODFN RI QHXURQV LQ WKH &3 DQG FHOOXODU DFFXPXODWLRQ LQ WKH 9= FRPSDUHG WR WKH FRQWURO
FRQGLWLRQ� �&� +LJKHU�PDJQLILFDWLRQ LPDJH RI WKH &3 VKRZLQJ 5*&V¶ HQG IHHW� 1RWH WKDW 6R[�
RYHUH[SUHVVLRQ GLG QRW DIIHFW WKH 5*& VFDIIROG� �'� 4XDQWLILFDWLRQ �PHDQ 6(0� RI WKHb
GLVWULEXWLRQ RI WDUJHWHG FHOOV ZLWKLQ WKH GHYHORSLQJ QHRFRUWH[ �(������ � 3 � ����� � 3 �
������� �(�*� ,PPXQRFKHPLVWU\ DW (���� EUDLQV IRU 3D[� �(�� 7EU� �)� DQG 'F[ �*� �EOXH� RQ
(���� S&,* RU S&,*�)/ 6R[� �JUHHQ� HOHFWURSRUDWHG HPEU\RV� �+� 4XDQWLILFDWLRQ �PHDQ �
6(0� DW (���� RI S&,* DQG S&,*�)/ 6R[� HOHFWURSRUDWHG FHOOV H[SUHVVLQJ 3D[�� 7EU� RU 'F[�
� 3 � ������� &3� FRUWLFDO SODWH� ,=� LQWHUPHGLDWH ]RQH� 69=� VXEYHQWULFXODU ]RQH� 9=�
YHQWULFXODU�]RQH��6FDOH�EDUV��%�������P��&������P� (�*�����P�
)LJXUH����6R[��RYHUH[SUHVVLRQ�DOWHUV�FHOO�F\FOH�NLQHWLFV LQ�5*&V�
�$� 6FKHPDWLFV RI WKH HOHFWURSRUDWLRQ VWUDWHJ\� (���� HPEU\RV ZHUH HOHFWURSRUDWHG ZLWK WKH
S&,* RU S&,*�)/ 6R[� SODVPLGV DQG ZHUH DQDO\]HG �� K SRVW�HOHFWURSRUDWLRQ �(������ �$¶�$¶�
(OHFWURSRUDWLRQ ZDV FRPELQHG ZLWK WKH LQMHFWLRQ RI QXFOHRVLGH DQDORJXHV� �%�
,PPXQRFKHPLVWU\ IRU 0�SKDVH PDUNHU S9LP �UHG� �� K SRVW�HOHFWURSRUDWLRQ �JUHHQ�� 1RWH WKDW
PRVW FHOOV SUHVHQW LQ WKH YHQWULFOH¶V VXUIDFH LQ WKH 6R[� RYHUH[SUHVVLRQ FRQGLWLRQ DUH QRW LQ
0�SKDVH� �&� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI HOHFWURSRUDWHG FHOOV H[SUHVVLQJ S9LP DW (�����
��

)DEUD�%HVHU�HW�DO�
� 3 �������� �'� (G8 VWDLQLQJ �UHG� LQ HOHFWURSRUDWHG EUDLQV �JUHHQ� DIWHU D VLQJOH SXOVH RI
(G8� ��� K EHIRUH GLVVHFWLRQ DV LOOXVWUDWHG LQ �$¶�� $UURZV SRLQW H[DPSOHV RI *)3�(G8� FHOOV�
�(� 4XDQWLILFDWLRQ �PHDQ � 6(0� DW (����� RI HOHFWURSRUDWHG FHOOV ODEHOHG ZLWK (G8� � 3
������� �)� ,PPXQRFKHPLVWU\ IRU SUROLIHUDWLRQ PDUNHU .L�� �UHG� �� K SRVW�HOHFWURSRUDWLRQ
�JUHHQ�� �*� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI HOHFWURSRUDWHG FHOOV H[SUHVVLQJ .L�� DW (����� �
3 �������� �+�,� &RVWDLQLQJ IRU SUROLIHUDWLRQ PDUNHUV 0FP� �UHG� DQG .L�� �EOXH� �� K DIWHU
HOHFWURSRUDWLRQ �JUHHQ� ZLWK S&,* �+� RU S&,*�)/ 6R[� �,� SODVPLGV� �+¶�+¶¶�
+LJKHU�PDJQLILFDWLRQ YLHZV RI �+� ER[HG DUHDV� �,¶�,¶¶� +LJKHU�PDJQLILFDWLRQ YLHZV RI �,� ER[HG
DUHDV� $UURZV SRLQW *)3� 0FP��.L��� FHOOV DQG DUURZKHDGV SRLQW *)3� 0FP��.L��� FHOOV�
�-� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI HOHFWURSRUDWHG FHOOV H[SUHVVLQJ 0FP� DQG�RU .L�� DW (�����
� 3 ������ � 3 �������� �.�/� &RVWDLQLQJ RI %UG8 �EOXH� DQG (G8 �UHG� DW (���� DIWHU
GXDO�SXOVH ODEHOLQJ VWUDWHJ\ RQ S&,* �.� DQG S&,*�)/ 6R[� �/� HOHFWURSRUDWHG EUDLQV �JUHHQ��
$V LOOXVWUDWHG LQ �$¶¶�� SUHJQDQW GDPV UHFHLYHG D %UG8 SXOVH ��� K EHIRUH GLVVHFWLRQ� DQG D (G8
SXOVH ��� K EHIRUH GLVVHFWLRQ� �.¶�.¶¶� +LJKHU�PDJQLILFDWLRQ YLHZV RI �.� ER[HG DUHDV� �/¶�/¶¶�
+LJKHU�PDJQLILFDWLRQ YLHZV RI �/� ER[HG DUHDV� $UURZV SRLQW *)3�%UG8�(G8� FHOOV�
DUURZKHDGV SRLQW *)3�%UG8�(G8� FHOOV DQG DVWHULVNV SRLQW *)3�%UG8�(G8� FHOOV� �0�
4XDQWLILFDWLRQ �PHDQ � 6(0� RI 6�SKDVH GXUDWLRQ �7V� DQG WRWDO FHOO�F\FOH GXUDWLRQ �7F� EDVHG
RQ GXDO�SXOVH ODEHOLQJ VWUDWHJ\� )RU 7F FDOFXODWLRQ� JURZWK IUDFWLRQ �*)� RI ODEHOOHG FHOOV ZDV
PHDVXUHG XVLQJ 0FP� PDUNHU� � 3 ������ � 3 �������� 9=� YHQWULFXODU ]RQH� 6FDOH EDUV� �%�
'� )� �� �P ORZ PDJQLILFDWLRQ DQG �� �P ER[HG DUHDV� �+� ,� .� /� �� �P� �+¶� ,¶� .¶� /¶� ��
�P�
)LJXUH �� 6R[� RYHUH[SUHVVLQJ 5*&V HQJDJH LQ QHXURJHQLF GLYLVLRQ DW ODWHU DJHV� �$�
6FKHPDWLFV RI WKH H[SHULPHQWDO VWUDWHJ\ FRPELQLQJ HOHFWURSRUDWLRQ DQG )ODVK7DJ WHFKQRORJLHV�
(���� HPEU\RV ZHUH HOHFWURSRUDWHG ZLWK WKH S&,* RU S&,*�)/ 6R[� SODVPLGV �JUHHQ�� �� KRXUV
DIWHU HOHFWURSRUDWLRQ HPEU\RV ZHUH LQMHFWHG ZLWK )ODVK7DJ �EOXH�� %UDLQV ZHUH DQDO\]HG DW
(����� �%� 4XDQWLILFDWLRQ �PHDQ 6(0� RI *)3��)7� GRXEOH ODEHOHG FHOOV LQ WKH &3 DW (����b
LQ ERWK FRQGLWLRQV� � 3 � ������ �&� &RQIRFDO LPDJH RI (���� EUDLQV DIWHU HOHFWURSRUDWLRQ DQG
��

)DEUD�%HVHU�HW�DO�
)7 LQMHFWLRQ DW GHVFULEHG WLPHSRLQWV� �'� +LJKHU�PDJQLILFDWLRQ YLHZ RI �&� VKRZLQJ WKH )7
ODEHOHG FHOOV DUHD� �(� +LJKHU�PDJQLILFDWLRQ YLHZ RI �'� ER[HG DUHDV� 'RWWHG RXWOLQH SRLQWV
H[DPSOHV RI *)3��)7� GRXEOH ODEHOHG FHOOV� //� ORZHU OD\HUV� 8/� XSSHU OD\HUV� 6FDOH EDUV� &�
�����P��'������P��(������P�
)LJXUH �� ,QFUHDVHG XSSHU OD\HU SURGXFWLRQ XSRQ 6R[� RYHUH[SUHVVLRQ DW HDUO\ DJHV� �$�
,OOXVWUDWLRQ RI HOHFWURSRUDWLRQ VWUDWHJ\� (���� HPEU\RV ZHUH HOHFWURSRUDWHG ZLWK S&,* RU
S&,*�)/ 6R[� FRQVWUXFWV �JUHHQ�� %UDLQV ZHUH DQDO\]HG DW (���� RU 3��� �%� ,PPXQRVWDLQLQJ
IRU &X[� �UHG� RQ (���� EUDLQV DIWHU (���� HOHFWURSRUDWLRQ RI ERWK FRQGLWLRQV �JUHHQ�� �%¶�
+LJKHU�PDJQLILFDWLRQ YLHZ RI �%� ER[HG DUHD� $UURZV SRLQW H[DPSOHV RI S&,*�)/ 6R[�
HOHFWURSRUDWHG FHOOV H[SUHVVLQJ &X[� DW (����� �&� 4XDQWLILFDWLRQ �PHDQ 6(0� RI WKHb
GLVWULEXWLRQ RI HOHFWURSRUDWHG FHOOV ZLWKLQ WKH &3 DW (����� )RU TXDQWLILFDWLRQ� WKH &3 ZDV
GLYLGHG LQ WHQ HTXDO�VL]H ELQV �HQXPHUDWHG ���� IURP EDVDO WR DSLFDO�� � 3 � ����� � 3 � ������
�'� ,PPXQRVWDLQLQJ IRU &X[� �UHG� RQ 3�� SRVWQDWDO EUDLQV DIWHU (���� HOHFWURSRUDWLRQ RI ERWK
FRQGLWLRQV �JUHHQ�� �'¶� +LJKHU�PDJQLILFDWLRQ YLHZ RI �'� ER[HG DUHD� $UURZV SRLQW H[DPSOHV
RI S&,*�)/ 6R[� HOHFWURSRUDWHG FHOOV H[SUHVVLQJ &X[� DW 3��� �(� 4XDQWLILFDWLRQ �PHDQ b
6(0� RI WKH GLVWULEXWLRQ RI HOHFWURSRUDWHG FHOOV ZLWKLQ WKH &3 DW 3��� )RU TXDQWLILFDWLRQ� WKH &3
ZDV GLYLGHG LQ WZR DUHDV� WKH ORZHU DQG XSSHU OD\HUV RI WKH QHRFRUWH[� � 3 � ����� ,=�
LQWHUPHGLDWH ]RQH� //� ORZHU OD\HUV� 8/� XSSHU OD\HUV� 6FDOH EDUV� �%� '� ��� �P� �%¶� '¶� ��
�P�
(;7(1'('�'$7$�),*85(�/(*(1'6
([WHQGHG 'DWD )LJXUH ���� *OLDO YHUVXV QHXUDO GLIIHUHQWLDWLRQ RI 5*&V DFWLYDWLQJ +HV�
SURPRWHU� �$� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI FHOO W\SH RI 7G�7RPDWR� FHOOV DW 3��� $L�
UHSRUWHU HPEU\RV ZHUH HOHFWURSRUDWHG ZLWK +HV�S�&UH(57� RU &$*�&UH(57� SODVPLGV�
0RUSKRORJLFDO IHDWXUHV ZHUH XVHG WR GLVWLQJXLVK S\UDPLGDO FHOOV DQG DVWURF\WHV� �%�
4XDQWLILFDWLRQ �PHDQ � 6(0� RI WKH GLVWULEXWLRQ RI 7G�7RPDWR� DVWURF\WHV ZLWKLQ WKH FRUWLFDO
SODWH �&3� DW 3��� $L� UHSRUWHU HPEU\RV ZHUH HOHFWURSRUDWHG ZLWK +HV�S�&UH(57� RU
��

)DEUD�%HVHU�HW�DO�
&$*�&UH(57� SODVPLGV� )RU TXDQWLILFDWLRQ� WKH &3 ZDV GLYLGHG LQ WHQ HTXDO�VL]H ELQV
�HQXPHUDWHG������IURP�EDVDO�WR�DSLFDO��� � 3 ������
([WHQGHG 'DWD )LJXUH ���� 'XDO UHSRUWHU YHFWRU FKDUDFWHUL]DWLRQ DQG H[WHQGHG
VF�51$VHT DQDO\VLV� �$� ,Q XWHUR HOHFWURSRUDWLRQ H[SHULPHQW DW (���� XVLQJ WKH GXDO UHSRUWHU
SODVPLG &$*�P5)3�+HV��*)3 DQDO\]HG DW (����� 1RWH WKH GLIIHUHQWLDO GLVWULEXWLRQ RI WKH FHOOV
H[SUHVVLQJ P5)3 �5HG� DQG WKH FHOOV H[SUHVVLQJ *)3 �JUHHQ�� �%� 4XDQWLILFDWLRQ RI WKH GDWD LQ
�$� �PHDQ 6(0� VKRZLQJ WKH GLIIHUHQW SURSRUWLRQ RI &$*� DQG +HV�� FHOOV LQ WKH 9=� 69=�b
,= DQG &3 DW ����� � 3 ������ � 3 �������� �&� ,Q XWHUR HOHFWURSRUDWLRQ H[SHULPHQW DW (����
XVLQJ WKH GXDO UHSRUWHU SODVPLG &$*�P5)3�+HV��*)3 DQDO\]HG DW 3�� 1RWH WKH GLIIHUHQWLDO
ODPLQDU ORFDOL]DWLRQ RI WKH SURMHFWLRQ QHXURQV DFWLYDWLQJ WKH GLIIHUHQW SURPRWHU� �'� W�61(
YLVXDOL]DWLRQV RI WKH VRUWHG FHOOV DIWHU WKH XQVXSHUYLVHG WUDQVFULSWLRQDO DQDO\VLV GLVSOD\LQJ WKH
LGHQWLILFDWLRQ RI � FOXVWHUV RI FHOOV� �(� W�61( YLVXDOL]DWLRQV RI WKH VRUWHG FHOOV GLVSOD\LQJ JHQH
H[SUHVVLRQ �JUD\ WR GDUN EOXH FRORU JUDGLHQW� RI FHOO�W\SH VSHFLILF PDUNHUV IRU 5*&V �6R[��� ,3V
�*DGG��J� DQG HDUO\ QHXURQV �5E)R[��� &3� FRUWLFDO SODWH� ,=� LQWHUPHGLDWH ]RQH� //� ORZHU
OD\HUV��69=��VXEYHQWULFXODU�]RQH��8/��XSSHU�OD\HUV� 9=��YHQWULFXODU�]RQH��6FDOH�EDUV������P�
([WHQGHG 'DWD )LJXUH ���� 0DVWHU UHJXODWRU DQDO\VLV LQ PRUH GLIIHUHQWLDWHG FHOOV DQG
GLVWULEXWLRQ RI 6R[� LQWHQVLW\ RI IOXRUHVFHQFH� �$�%� 7UDQVFULSWLRQDO UHJXODWRU\ QHWZRUN RI
VF51$�VHT GLVSOD\LQJ H[SUHVVLRQ OHYHOV RI HDFK UHJXORQ LQ ,3V �$� DQG (1V �%� LQ &$*� DQG
+HV�� FHOOV� �&� 'LVWULEXWLRQ RI WKH 6R[� LQWHQVLW\ RI IOXRUHVFHQFH �D�X�� IRU &$*� DQG +HV��
5*&V� 1RWH WKH GLIIHUHQFHV LQ WKH SHUFHQWDJHV RI HDFK VXEW\SH RI FHOOV IRU WKH GLIIHUHQW
LQWHQVLWLHV�RI�IOXRUHVFHQFH�
([WHQGHG 'DWD )LJXUH ���� 6R[� NQRFN�GRZQ LQFUHDVHV 5*& QHXURJHQLF EHKDYLRU� �$�
,PPXQRVWDLQLQJ DW WKH (���� 9= IRU 6R[� �EOXH� DIWHU (���� HOHFWURSRUDWLRQ �JUHHQ� ZLWK
6F�6K51$�S&,* DQG 6R[��6K51$�S&,* SODVPLGV� 1RWH WKDW LQ WKH NQRFN�GRZQ FRQGLWLRQ
6R[� LQWHQVLW\ ZDV VXFFHVVIXOO\ UHGXFHG LQ HOHFWURSRUDWHG FHOOV LQ WKH 9=� �%� 4XDQWLILFDWLRQ
��

)DEUD�%HVHU�HW�DO�
�PHDQ � 6(0� RI WKH SHUFHQWDJH RI HOHFWURSRUDWHG FHOOV ORFDWHG LQ WKH &3 DW (����� � 3 ������
�&� &RQIRFDO LPDJH RI (���� EUDLQV DIWHU (���� HOHFWURSRUDWLRQ ZLWK 6F�6K51$�S&,* SODVPLG
�FRQWURO� DQG 6R[��6K51$�S&,* SODVPLG �NQRFN�GRZQ�� &3� FRUWLFDO SODWH� ,=� LQWHUPHGLDWH
]RQH��69=��VXEYHQWULFXODU�]RQH��9=��YHQWULFXODU�]RQH� 6FDOH�EDUV���$������P���&������P�
([WHQGHG 'DWD )LJXUH ���� $GGLWLRQDO FHOO F\FOH DQDO\VLV LQ FRUWLFDO 9= XSRQ 6R[�
RYHUH[SUHVVLRQ� �$� 6FKHPDWLFV RI WKH H[SHULPHQWDO VWUDWHJ\� (PEU\RV ZHUH HOHFWURSRUDWHG DW
(���� ZLWK WKH S&,* DQG S&,*�)/ 6R[� SODVPLGV� �� K SRVW�HOHFWURSRUDWLRQ (���� SUHJQDQW
GDPV UHFHLYHG D VLQJOH SXOVH RI %UG8� (PEU\RV ZHUH GLVVHFWHG �� K SRVW�%UG8 LQMHFWLRQ� �%�&�
&RVWDLQLQJ IRU %UG8 �EOXH� DQG SUROLIHUDWLRQ PDUNHU .L�� �UHG� �� K DIWHU HOHFWURSRUDWLRQ
�JUHHQ� ZLWK S&,* �%� RU S&,*�)/ 6R[� SODVPLG �&�� �%¶�%¶¶� +LJKHU�PDJQLILFDWLRQ YLHZ RI
�%� ER[HG DUHDV� �&¶�&¶¶� +LJKHU�PDJQLILFDWLRQ YLHZ RI �&� ER[HG DUHDV� $UURZV SRLQW H[DPSOHV
RI (3 FHOOV VWDLQHG IRU .L�� DQG %UG8� DUURZKHDGV SRLQW (3 FHOOV VWDLQHG RQO\ IRU %UG8 DQG
DVWHULVNV SRLQW (3 FHOOV RQO\ VWDLQHG IRU *)3� �'� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI (3 FHOOV WKDW
DUH .L���%UG8� RU .L���%UG8� LQ WKH 9= DW (����� � 3 ������ �(�)� 4XDQWLILFDWLRQ �PHDQ �
6(0� RI WKH GLVWULEXWLRQ ZLWKLQ WKH 9= RI (3 FHOOV WKDW DUH .L���%UG8� RU .L���%UG8� DW
(����� � 3 ������ )RU WKHVH TXDQWLILFDWLRQV� WKH 9= ZDV GLYLGHG LQ WZR HTXDO�VL]HG UHJLRQV� WKH
RXWHU �(� DQG WKH LQQHU �)� 9=� � 3 ������ 1RWH WKDW WKH GLIIHUHQFHV LQ WKH � RI (3 FHOOV
.L���%UG8� EHWZHHQ WKH FRQWURO DQG RYHUH[SUHVVLRQ FRQGLWLRQ VKRZQ LQ �'� DUH PDLQO\ FDXVHG
E\ WKH GLIIHUHQFHV LQ WKH LQQHU UHJLRQ RI WKH 9= VKRZQ LQ �)�� �*� &RQIRFDO LPDJH RI WKH 9= DW
(����� DIWHU (���� S&,* RU S&,*�)/ 6R[� HOHFWURSRUDWLRQ� �+� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI
WKH GLVWULEXWLRQ ZLWKLQ WKH 9= RI (3 FHOOV ZLWK S&,* DQG S&,*�)/ 6R[�� � 3 ������ �,�
&RVWDLQLQJ IRU %UG8 �EOXH� DQG 'F[ �UHG� �� K SRVW�HOHFWURSRUDWLRQ �JUHHQ�� 1RWH WKDW (3 FHOOV
%UG8� LQ WKH LQQHU UHJLRQ RI WKH 9= DUH QRW 'F[�� �-� ,PPXQRFKHPLVWU\ IRU 3D[� �UHG� �� K
SRVW�HOHFWURSRUDWLRQ �JUHHQ�� $UURZV SRLQW H[DPSOHV RI HOHFWURSRUDWHG FHOOV WKDW H[SUHVV 3D[��
�.� )XOO�FRUWH[ YLHZ RI WKH VWDLQLQJ IRU WKH SUROLIHUDWLRQ PDUNHU 0FP� DW (����� �/�
4XDQWLILFDWLRQ �PHDQ � 6(0� RI (3 FHOOV WKDW H[SUHVV 0FP� LQ WKH 9= DW (����� � 3 ������
��

)DEUD�%HVHU�HW�DO�
�0� 4XDQWLILFDWLRQ �PHDQ � 6(0� RI WKH WRWDO FHOO�F\FOH GXUDWLRQ �7V� EDVHG RQ GXDO�SXOVH
ODEHOLQJ VWUDWHJ\ GHVFULEHG LQ )LJ��� � 3 �������� )RU WKLV 7F FDOFXODWLRQ� JURZWK IUDFWLRQ �*)�
RI ODEHOOHG FHOOV ZDV PHDVXUHG XVLQJ .L�� PDUNHU� &3� FRUWLFDO SODWH� ,=� LQWHUPHGLDWH ]RQH�
69=� VXEYHQWULFXODU ]RQH� 9=� YHQWULFXODU ]RQH� 6FDOH EDUV� �-� ��� �P� �%� &� *�,� �� �P�
�%¶�&¶¶������P�
([WHQGHG 'DWD )LJXUH ���� 6R[� NQRFN�GRZQ DOWHUV ODPLQDU GLVWULEXWLRQ RI +HV�� 5*&
RIIVSULQJ� �$� &RQIRFDO LPDJH RI (���� EUDLQV DIWHU HOHFWURSRUDWLRQ DW (���� ZLWK
6F�6K51$�+HV��*)3 �FRQWURO� DQG 6R[��6K51$�+HV��*)3 SODVPLG �NQRFN�GRZQ�� �(�
4XDQWLILFDWLRQ �PHDQ � 6(0� RI WKH GLVWULEXWLRQ RI HOHFWURSRUDWHG FHOOV ZLWKLQ WKH &3 DW (�����
)RU TXDQWLILFDWLRQ� WKH &3 ZDV GLYLGHG LQ WHQ HTXDO�VL]H ELQV �HQXPHUDWHG ���� IURP EDVDO WR
DSLFDO�� � 3 ������ � 3 ������� � 3 ������� &3� FRUWLFDO SODWH� ,=� LQWHUPHGLDWH ]RQH� 6FDOH
EDU��$�������P�
%,%/,2*5$3+<
$OFDPR�($��&KLULYHOOD�/��'DXW]HQEHUJ�0��'REUHYD�*� )DULxDV�,��*URVVFKHGO�5��0F&RQQHOO�6.�������6DWE��5HJXODWHV�&DOORVDO�3URMHFWLRQ�1HXURQ ,GHQWLW\�LQ�WKH�'HYHORSLQJ�&HUHEUDO&RUWH[��1HXURQ�������±����
$UDL�<��3XOYHUV�-1��+DIIQHU�&��6FKLOOLQJ�%��1�VVOHLQ ,��&DOHJDUL�)��+XWWQHU�:%��������1HXUDOVWHP�DQG�SURJHQLWRU�FHOOV�VKRUWHQ�6�SKDVH�RQ�FRPPLWPHQW WR�QHXURQ�SURGXFWLRQ��1DW&RPPXQ�������
$UORWWD�3��0RO\QHDX[�%-��&KHQ�-��,QRXH�-��.RPLQDPL 5��0DFNOLV�-'��������1HXURQDO6XEW\SH�6SHFLILF�*HQHV�WKDW�&RQWURO�&RUWLFRVSLQDO 0RWRU�1HXURQ�'HYHORSPHQW�,Q�9LYR�1HXURQ�������±����
%DVDN�2��7D\ORU�9��������,GHQWLILFDWLRQ�RI�VHOIòUHSOLFDWLQJ PXOWLSRWHQW�SURJHQLWRUV�LQ�WKHHPEU\RQLF�QHUYRXV�V\VWHP�E\�KLJK�1RWFK�DFWLYLW\�DQG +HV��H[SUHVVLRQ��(XU�-�1HXURVFL�������±�����
%ULWDQRYD�2��5RPHUR�&�GH�-��&KHXQJ�$��.ZDQ�.<��6FKZDUN 0��*\RUJ\�$��9RJHO�7��$NRSRY�6�0LWNRYVNL�0��$JRVWRQ�'��âHVWDQ�1��0ROQiU�=��7DUDE\NLQ 9��������6DWE��,V�D�3RVWPLWRWLF'HWHUPLQDQW�IRU�8SSHU�/D\HU�1HXURQ�6SHFLILFDWLRQ�LQ WKH�1HRFRUWH[��1HXURQ�������±����
%XFN�6%��%UDGIRUG�-��*HH�.5��$JQHZ�%-��&ODUNH�67� 6DOLF�$��������'HWHFWLRQ�RI�6�SKDVH�FHOOF\FOH�SURJUHVVLRQ�XVLQJ���HWK\Q\O���GHR[\XULGLQH�LQFRUSRUDWLRQ ZLWK�FOLFN�FKHPLVWU\��DQDOWHUQDWLYH�WR�XVLQJ���EURPR���GHR[\XULGLQH�DQWLERGLHV� %LRWHFKQLTXHV�������±����
%XWOHU�$��+RIIPDQ�3��6PLEHUW�3��3DSDOH[L�(��6DWLMD 5��������,QWHJUDWLQJ�VLQJOH�FHOOWUDQVFULSWRPLF�GDWD�DFURVV�GLIIHUHQW�FRQGLWLRQV��WHFKQRORJLHV� DQG�VSHFLHV��1DW�%LRWHFKQRO������±����
��

)DEUD�%HVHU�HW�DO�
&DFFDPR�'��.DWVHWRV�&'��+HUPDQ�00��)UDQNIXUWHU�$��&ROOLQV�93��5XELQVWHLQ�/-�������,PPXQRKLVWRFKHPLVWU\�RI�D�VSRQWDQHRXV�PXULQH�RYDULDQ WHUDWRPD�ZLWK�QHXURHSLWKHOLDOGLIIHUHQWLDWLRQ��1HXURQ�DVVRFLDWHG�EHWD�WXEXOLQ�DV D�PDUNHU�IRU�SULPLWLYH�QHXURHSLWKHOLXP�/DERUDWRU\�LQYHVWLJDWLRQ��D�MRXUQDO�RI�WHFKQLFDO�PHWKRGV DQG�SDWKRORJ\�������±����
&DVWUR�0$$��6DQWLDJR�,�GH��&DPSEHOO�70��9DXJKQ�&� +LFNH\�7(��5RVV�(��7LOOH\�:'�0DUNRZHW]�)��3RQGHU�%$-��0H\HU�.%��������5HJXODWRUV RI�JHQHWLF�ULVN�RI�EUHDVW�FDQFHULGHQWLILHG�E\�LQWHJUDWLYH�QHWZRUN�DQDO\VLV��1DW�*HQHW �����±���
&KHQJ�/�&��3DVWUDQD�(��7DYD]RLH�0��'RHWVFK�)������� PL5�����UHJXODWHV�DGXOW�QHXURJHQHVLV�LQWKH�VXEYHQWULFXODU�]RQH�VWHP�FHOO�QLFKH��1DW�1HXURVFL ������±����
&KHQQ�$��������:QW���FDWHQLQ�VLJQDOLQJ�LQ�FHUHEUDO FRUWLFDO�GHYHORSPHQW��2UJDQRJHQHVLV����±���
&KHYpH�0��5REHUWVRQ�-'-��&DQQRQ�*+��%URZQ�63��*RII /$��������9DULDWLRQ�LQ�$FWLYLW\�6WDWH�$[RQDO�3URMHFWLRQ��DQG�3RVLWLRQ�'HILQH�WKH�7UDQVFULSWLRQDO ,GHQWLW\�RI�,QGLYLGXDO�1HRFRUWLFDO3URMHFWLRQ�1HXURQV��&HOO�5HSRUWV�������±����
'HVDL�$5��0F&RQQHOO�6.��������3URJUHVVLYH�UHVWULFWLRQ LQ�IDWH�SRWHQWLDO�E\�QHXUDO�SURJHQLWRUVGXULQJ�FHUHEUDO�FRUWLFDO�GHYHORSPHQW��'HYHORSPHQW �&DPEULGJH��(QJODQG����������±�����
(QJOXQG�&��)LQN�$��/DX�&��3KDP�'��'D]D�5$0��%XOIRQH $��.RZDOF]\N�7��+HYQHU�5)�������3D[���7EU���DQG�7EU��$UH�([SUHVVHG�6HTXHQWLDOO\�E\ 5DGLDO�*OLD��,QWHUPHGLDWH�3URJHQLWRU&HOOV��DQG�3RVWPLWRWLF�1HXURQV�LQ�'HYHORSLQJ�1HRFRUWH[� -�1HXURVFL�������±����
)LHW]�6$��.HODYD�,��9RJW�-��:LOVFK�%UlXQLQJHU�0��6WHQ]HO '��)LVK�-/��&RUEHLO�'��5LHKQ�$�'LVWOHU�:��1LWVFK�5��+XWWQHU�:%��������269=�SURJHQLWRUV RI�KXPDQ�DQG�IHUUHW�QHRFRUWH[DUH�HSLWKHOLDO�OLNH�DQG�H[SDQG�E\�LQWHJULQ�VLJQDOLQJ� 1DW�1HXURVFL�������±����
)OHWFKHU�01&��&DVWUR�0$$��:DQJ�;��6DQWLDJR�,�GH��2¶5HLOO\ 0��&KLQ�6�)��5XHGD�20��&DOGDV&��3RQGHU�%$-��0DUNRZHW]�)��0H\HU�.%��������0DVWHU UHJXODWRUV�RI�)*)5��VLJQDOOLQJ�DQGEUHDVW�FDQFHU�ULVN��1DW�&RPPXQ��������
)UDQFR�6-��*LO�6DQ]�&��0DUWLQH]�*DUD\�,��(VSLQRVD $��+DUNLQV�3HUU\�65��5DPRV�&��0�OOHU�8�������)DWH�5HVWULFWHG�1HXUDO�3URJHQLWRUV�LQ�WKH�0DPPDOLDQ &HUHEUDO�&RUWH[��6FLHQFH�������±����
)UDQNLVK�$�HW�DO���������*(1&2'(�UHIHUHQFH�DQQRWDWLRQ IRU�WKH�KXPDQ�DQG�PRXVH�JHQRPHV�1XFOHLF�$FLGV�5HV����JN\�����
)UDQW]�*'��0F&RQQHOO�6.��������5HVWULFWLRQ�RI�ODWH FHUHEUDO�FRUWLFDO�SURJHQLWRUV�WR�DQXSSHU�OD\HU�IDWH��1HXURQ������±���
)XHQWHDOED�/&��5RPSDQL�6%��3DUUDJXH]�-,��2EHUQLHU .��5RPHUR�5��&HSNR�&/��$OYDUH]�%X\OOD$��������(PEU\RQLF�2ULJLQ�RI�3RVWQDWDO�1HXUDO�6WHP &HOOV��&HOO���������±�����
*DO�-6��0RUR]RY�<0��$\RXE�$(��&KDWWHUMHH�0��5DNLF 3��+D\GDU�7)��������0ROHFXODU�DQG0RUSKRORJLFDO�+HWHURJHQHLW\�RI�1HXUDO�3UHFXUVRUV�LQ WKH�0RXVH�1HRFRUWLFDO�3UROLIHUDWLYH=RQHV��-�1HXURVFL��������±�����
*DR�3��3RVWLJOLRQH�03��.ULHJHU�7*��+HUQDQGH]�/��:DQJ &��+DQ�=��6WUHLFKHU�&��3DSXVKHYD�(�,QVROHUD�5��&KXJK�.��.RGLVK�2��+XDQJ�.��6LPRQV�%'� /XR�/��+LSSHQPH\HU�6��6KL�6�+�������'HWHUPLQLVWLF�3URJHQLWRU�%HKDYLRU�DQG�8QLWDU\ 3URGXFWLRQ�RI�1HXURQV�LQ�WKH1HRFRUWH[��&HOO��������±����
*DUFtD�0RUHQR�)��0ROQiU�=��������6XEVHW�RI�HDUO\�UDGLDO JOLDO�SURJHQLWRUV�WKDW�FRQWULEXWH�WR�WKHGHYHORSPHQW�RI�FDOORVDO�QHXURQV�LV�DEVHQW�IURP�DYLDQ EUDLQ��3URF�1DWLRQDO�$FDG�6FL����(����±(�����
*HUGHV�-��6FKZDE�8��/HPNH�+��6WHLQ�+��������3URGXFWLRQ RI�D�PRXVH�PRQRFORQDO�DQWLERG\UHDFWLYH�ZLWK�D�KXPDQ�QXFOHDU�DQWLJHQ�DVVRFLDWHG�ZLWK FHOO�SUROLIHUDWLRQ��,QW�-�&DQFHU�����±���
*LO�6DQ]�&��(VSLQRVD�$��)UHJRVR�63��%OXVNH�..��&XQQLQJKDP &/��0DUWLQH]�*DUD\�,��=HQJ�+�)UDQFR�6-��0�OOHU�8��������/LQHDJH�7UDFLQJ�8VLQJ�&X[��&UH DQG�&X[��&UH(57��0LFH�1HXURQ��������±�����
*LO�6DQ]�&��)UDQFR�6-��0DUWLQH]�*DUD\�,��(VSLQRVD $��+DUNLQV�3HUU\�6��0�OOHU�8�������&DMDO�5HW]LXV�&HOOV�,QVWUXFW�1HXURQDO�0LJUDWLRQ�E\ &RLQFLGHQFH�6LJQDOLQJ�EHWZHHQ�6HFUHWHGDQG�&RQWDFW�'HSHQGHQW�*XLGDQFH�&XHV��1HXURQ�������±����
*RHEEHOV�6��%RUPXWK�,��%RGH�8��+HUPDQVRQ�2��6FKZDE 0+��1DYH�.��������*HQHWLF�WDUJHWLQJRI�SULQFLSDO�QHXURQV�LQ�QHRFRUWH[�DQG�KLSSRFDPSXV RI�1(;ò&UH�PLFH��*HQHVLV�������±����
��

)DEUD�%HVHU�HW�DO�
*|W]�0��6WR\NRYD�$��*UXVV�3��������3D[��FRQWUROV�UDGLDO�JOLD�GLIIHUHQWLDWLRQ�LQ�WKH�FHUHEUDOFRUWH[��1HXURQ��������±�����
*RYLQGDQ�6��2EHUVW�3��-DEDXGRQ�'��������,Q�YLYR�SXOVH ODEHOLQJ�RI�LVRFKURQLF�FRKRUWV�RI�FHOOV�LQWKH�FHQWUDO�QHUYRXV�V\VWHP�XVLQJ�)ODVK7DJ��1DW�3URWRF �������±�����
*XHQWKQHU�&-��0L\DPLFKL�.��<DQJ�++��+HOOHU�+&��/XR /��������3HUPDQHQW�*HQHWLF�$FFHVV�WR7UDQVLHQWO\�$FWLYH�1HXURQV�YLD�75$3��7DUJHWHG�5HFRPELQDWLRQ LQ�$FWLYH�3RSXODWLRQV�1HXURQ�������±����
*XR�:��.HFNHVRYD�=��'RQDKHU�-/��6KLEXH�7��7LVFKOHU 9��5HLQKDUGW�)��,W]NRYLW]�6��1RVNH�$�=�UUHU�+lUGL�8��%HOO�*��7DP�:/��0DQL�6$��YDQ�2XGHQDDUGHQ $��:HLQEHUJ�5$��������6OXJDQG�6R[��&RRSHUDWLYHO\�'HWHUPLQH�WKH�0DPPDU\�6WHP &HOO�6WDWH��&HOO���������±�����
+DJH\�':��0XKU�-��������6R[��$FWV�LQ�D�'RVH�'HSHQGHQW )DVKLRQ�WR�5HJXODWH�3UROLIHUDWLRQ�RI&RUWLFDO�3URJHQLWRUV��&HOO�5HSRUWV�������±�����
+DQVHQ�'9��/XL�-+��3DUNHU�35/��.ULHJVWHLQ�$5������� 1HXURJHQLF�UDGLDO�JOLD�LQ�WKH�RXWHUVXEYHQWULFXODU�]RQH�RI�KXPDQ�QHRFRUWH[��1DWXUH��������±����
+DUULV�/��=DOXFNL�2��3LSHU�0��������%UG8�(G8�GXDO ODEHOLQJ�WR�GHWHUPLQH�WKH�FHOO�F\FOHG\QDPLFV�RI�GHILQHG�FHOOXODU�VXESRSXODWLRQV��-�0RO +LVWRO�������±����
+DWDNH\DPD�-��%HVVKR�<��.DWRK�.��2RNDZDUD�6��)XMLRND 0��*XLOOHPRW�)��.DJH\DPD�5�������+HV�JHQHV�UHJXODWH�VL]H��VKDSH�DQG�KLVWRJHQHVLV�RI WKH�QHUYRXV�V\VWHP�E\�FRQWURO�RI�WKHWLPLQJ�RI�QHXUDO�VWHP�FHOO�GLIIHUHQWLDWLRQ��'HYHORSPHQW ��������±�����
+LUDED\DVKL�<��,WRK�<��7DEDWD�+��1DNDMLPD�.��$NL\DPD 7��0DVX\DPD�1��*RWRK�<��������7KH:QW���FDWHQLQ�SDWKZD\�GLUHFWV�QHXURQDO�GLIIHUHQWLDWLRQ RI�FRUWLFDO�QHXUDO�SUHFXUVRU�FHOOV�'HYHORSPHQW���������±�����
+RUVW�'��&KHQ�-��0RULNDZD�7��2JLQR�6��.LUFKQHU�7� 6KLYGDVDQL�5$��������'LIIHUHQWLDO�:17$FWLYLW\�LQ�&RORUHFWDO�&DQFHU�&RQIHUV�/LPLWHG�7XPRULJHQLF 3RWHQWLDO�DQG�,V�5HJXODWHG�E\0$3.�6LJQDOLQJ��&DQFHU�5HV��������±�����
.DGDMD�0��.H\HV�%(��/LQ�0��3DVROOL�+$��*HQDQGHU�0� 3RODN�/��6WRNHV�1��=KHQJ�'��)XFKV�(�������62;���D�VWHP�FHOO�WUDQVFULSWLRQDO�UHJXODWRU RI�VHFUHWHG�QLFKH�VLJQDOLQJ�IDFWRUV��*HQH'HY�������±����
.DPHL�<��,QDJDNL�1��1LVKL]DZD�0��7VXWVXPL�2��7DNHWDQL <��,QDJDNL�0��������9LVXDOL]DWLRQ�RIPLWRWLF�UDGLDO�JOLDO�OLQHDJH�FHOOV�LQ�WKH�GHYHORSLQJ UDW�EUDLQ�E\�&GF��NLQDVHòSKRVSKRU\ODWHGYLPHQWLQ��*OLD�������±����
.DSODQ�(6��5DPRV�/DJXQD�.$��0LKDODV�$%��'D]D�5$0� +HYQHU�5)��������1HRFRUWLFDO�6R[��UDGLDO�JOLD�JHQHUDWH�JOXWDPDWHUJLF�QHXURQV�IRU�DOO OD\HUV��EXW�ODFN�GLVFHUQLEOH�HYLGHQFH�RIHDUO\�ODPLQDU�IDWH�UHVWULFWLRQ��1HXUDO�'HY�������
.DZDJXFKL�$��,NDZD�7��.DVXNDZD�7��8HGD�+5��.XULPRWR .��6DLWRX�0��0DWVX]DNL�)�������6LQJOH�FHOO�JHQH�SURILOLQJ�GHILQHV�GLIIHUHQWLDO�SURJHQLWRU VXEFODVVHV�LQ�PDPPDOLDQQHXURJHQHVLV��'HYHORSPHQW���������±�����
.LP�'��/DQJPHDG�%��6DO]EHUJ�6/��������+,6$7��D�IDVW VSOLFHG�DOLJQHU�ZLWK�ORZ�PHPRU\UHTXLUHPHQWV��1DW�0HWKRGV�������±����
.RRS�.(��0DF'RQDOG�/0��/REH�&*��������7UDQVFULSWV RI�*UJ���D�PXULQH�JURXFKR�UHODWHGJHQH��DUH�GHWHFWHG�LQ�DGMDFHQW�WLVVXHV�WR�RWKHU�PXULQH QHXURJHQLF�JHQH�KRPRORJXHV�GXULQJHPEU\RQLF�GHYHORSPHQW��0HFKDQLVPV�RI�GHYHORSPHQW������±���
/D0RQLFD�%(��/XL�-+��:DQJ�;��.ULHJVWHLQ�$5������� 269=�SURJHQLWRUV�LQ�WKH�KXPDQ�FRUWH[�DQ�XSGDWHG�SHUVSHFWLYH�RQ�QHXURGHYHORSPHQWDO�GLVHDVH� &XUU�2SLQ�1HXURELRO�������±����
/LX�&��/LX�/��&KHQ�;��&KHQJ�-��=KDQJ�+��6KHQ�-��6KDQ -��;X�<��<DQJ�=��/DL�0��4LDQ�&�������6R[��UHJXODWHV�VHOIòUHQHZDO�DQG�WXPRULJHQLFLW\�E\ SURPRWLQJ�V\PPHWULFDO�FHOO�GLYLVLRQ�RIFDQFHU�VWHP�FHOOV�LQ�KHSDWRFHOOXODU�FDUFLQRPD��+HSDWRORJ\ ������±����
/ORUFD�$��&LFHUL�*��%HDWWLH�5��:RQJ�).��'LDQD�*��6HUDIHLPLGRX�3RXOLRX (��)HUQiQGH]�2WHUR0��6WUHLFKHU�&��$UQROG�6-��0H\HU�0��+LSSHQPH\HU�6� 0DUDYDOO�0��0DULQ�2��������$VWRFKDVWLF�IUDPHZRUN�RI�QHXURJHQHVLV�XQGHUOLHV�WKH DVVHPEO\�RI�QHRFRUWLFDO�F\WRDUFKLWHFWXUH�(OLIH���H������
/ORUHQV�%REDGLOOD�(��=KDR�6��%DVHU�$��6DL]�&DVWUR *��=ZDGOR�.��0DUWLQ�9LOODOED�$�������6LQJOH�&HOO�7UDQVFULSWRPLFV�5HYHDOV�D�3RSXODWLRQ�RI 'RUPDQW�1HXUDO�6WHP�&HOOV�WKDW%HFRPH�$FWLYDWHG�XSRQ�%UDLQ�,QMXU\��&HOO�6WHP�&HOO ������±����
��

)DEUD�%HVHU�HW�DO�
/RQJ�-=��/DFNDQ�&6��+DGMDQWRQDNLV�$�.��������*HQHWLF�DQG�VSHFWUDOO\�GLVWLQFW�LQ�YLYRLPDJLQJ�HPEU\RQLF�VWHP�FHOOV�DQG�PLFH�ZLWK�ZLGHVSUHDG�H[SUHVVLRQ�RI�D�PRQRPHULF�UHG�IOXRUHVFHQWSURWHLQ��%PF�%LRWHFKQRO������
/RXYL�$��$UWDYDQLV�7VDNRQDV�6��������1RWFK�VLJQDOOLQJ LQ�YHUWHEUDWH�QHXUDO�GHYHORSPHQW��1DW5HY�1HXURVFL�����±����
/XL�-+��+DQVHQ�'9��.ULHJVWHLQ�$5��������'HYHORSPHQW DQG�(YROXWLRQ�RI�WKH�+XPDQ1HRFRUWH[��&HOO�������±���
0DGLVHQ�/��=ZLQJPDQ�7$��6XQNLQ�60��2K�6:��=DULZDOD +$��*X�+��1J�//��3DOPLWHU�5'�+DZU\O\F]�0-��-RQHV�$5��/HLQ�(6��=HQJ�+��������$�UREXVW DQG�KLJK�WKURXJKSXW�&UHUHSRUWLQJ�DQG�FKDUDFWHUL]DWLRQ�V\VWHP�IRU�WKH�ZKROH PRXVH�EUDLQ��1DW�1HXURVFL�������±���$YDLODEOH�DW��KWWS���ZZZ�QDWXUH�FRP�QHXUR�MRXUQDO�Y���Q��IXOO�QQ������KWPO�
0DUJROLQ�$$��:DQJ�.��/LP�:.��.XVWDJL�0��1HPHQPDQ�,� &DOLIDQR�$��������5HYHUVHHQJLQHHULQJ�FHOOXODU�QHWZRUNV��1DW�3URWRF������±����
0DUW\QRJD�%��0RUULVRQ�+��3ULFH�'-��0DVRQ�-2������� )R[J��LV�UHTXLUHG�IRU�VSHFLILFDWLRQ�RIYHQWUDO�WHOHQFHSKDORQ�DQG�UHJLRQ�VSHFLILF�UHJXODWLRQ RI�GRUVDO�WHOHQFHSKDOLF�SUHFXUVRUSUROLIHUDWLRQ�DQG�DSRSWRVLV��'HY�%LRO��������±����
0DVORY�$<��%DURQH�7$��3OXQNHWW�5-��3UXLWW�6&������� 1HXUDO�6WHP�&HOO�'HWHFWLRQ�&KDUDFWHUL]DWLRQ��DQG�$JH�5HODWHG�&KDQJHV�LQ�WKH�6XEYHQWULFXODU =RQH�RI�0LFH��-�1HXURVFL�������±�����
0DWVXGD�7��&HSNR�&/��������(OHFWURSRUDWLRQ�DQG�51$ LQWHUIHUHQFH�LQ�WKH�URGHQW�UHWLQD�LQ�YLYRDQG�LQ�YLWUR��3URF�1DWLRQDO�$FDG�6FL�������±���
0DWVXGD�7��&HSNR�&/��������&RQWUROOHG�H[SUHVVLRQ�RI WUDQVJHQHV�LQWURGXFHG�E\�LQ�YLYRHOHFWURSRUDWLRQ��3URF�1DWLRQDO�$FDG�6FL���������±�����
0F&RQQHOO�6.��.D]QRZVNL�&(��������&HOO�F\FOH�GHSHQGHQFH RI�ODPLQDU�GHWHUPLQDWLRQ�LQGHYHORSLQJ�QHRFRUWH[��6FLHQFH��������±����
0LOOHU�,��0LQ�0��<DQJ�&��7LDQ�&��*RRNLQ�6��&DUWHU '��6SHQFHU�6/��������.L���LV�D�*UDGHG5DWKHU�WKDQ�D�%LQDU\�0DUNHU�RI�3UROLIHUDWLRQ�YHUVXV 4XLHVFHQFH��&HOO�5HSRUWV�������������H��
0L\DWD�7��.DZDJXFKL�$��6DLWR�.��.DZDQR�0��0XWR�7� 2JDZD�0��������$V\PPHWULFSURGXFWLRQ�RI�VXUIDFH�GLYLGLQJ�DQG�QRQ�VXUIDFH�GLYLGLQJ FRUWLFDO�SURJHQLWRU�FHOOV�'HYHORSPHQW���������±�����
0L]XWDQL�.��<RRQ�.��'DQJ�/��7RNXQDJD�$��*DLDQR�1������� 'LIIHUHQWLDO�1RWFK�VLJQDOOLQJGLVWLQJXLVKHV�QHXUDO�VWHP�FHOOV�IURP�LQWHUPHGLDWH SURJHQLWRUV��1DWXUH��������±����
0RO\QHDX[�%-��$UORWWD�3��0HQH]HV�-5/��0DFNOLV�-'������� 1HXURQDO�VXEW\SH�VSHFLILFDWLRQ�LQWKH�FHUHEUDO�FRUWH[��1DW�5HY�1HXURVFL������±����
0XNKWDU�7��%UHGD�-��*ULVRQ�$��.DULPDGGLQL�=��*UREHFNHU 3��,EHU�'��%HLVHO�&��1LPZHJHQ�(�YDQ�7D\ORU�9��������7HDG�WUDQVFULSWLRQ�IDFWRUV�GLIIHUHQWLDOO\ UHJXODWH�FRUWLFDO�GHYHORSPHQW��6FL5HS�XN���������
0XOOHQ�5-��%XFN�&5��6PLWK�$0��������1HX1��D�QHXURQDO VSHFLILF�QXFOHDU�SURWHLQ�LQYHUWHEUDWHV��'HY�&DPE�(QJO��������±����
1LHWR�0��0RQXNL�(6��7DQJ�+��,PLWROD�-��+DXEVW�1��.KRXU\ 6-��&XQQLQJKDP�-��*RW]�0��:DOVK&$��������([SUHVVLRQ�RI�&X[ò��DQG�&X[ò��LQ�WKH�VXEYHQWULFXODU ]RQH�DQG�XSSHU�OD\HUV,,±,9�RI�WKH�FHUHEUDO�FRUWH[��-�&RPS�1HXURO��������±����
1RFWRU�6&��)OLQW�$&��:HLVVPDQ�7$��:RQJ�:6��&OLQWRQ %.��.ULHJVWHLQ�$5��������'LYLGLQJSUHFXUVRU�FHOOV�RI�WKH�HPEU\RQLF�FRUWLFDO�YHQWULFXODU ]RQH�KDYH�PRUSKRORJLFDO�DQG�PROHFXODUFKDUDFWHULVWLFV�RI�UDGLDO�JOLD��7KH�-RXUQDO�RI�1HXURVFLHQFH �������±�����$YDLODEOH�DW�KWWS���HXWLOV�QFEL�QOP�QLK�JRY�HQWUH]�HXWLOV�HOLQN�IFJL"GEIURP SXEPHGLG ��������UHWPRGH UHIFPG SUOLQNV�
1RFWRU�6&��0DUWtQH]�&HUGHxR�9��,YLF�/��.ULHJVWHLQ $5��������&RUWLFDO�QHXURQV�DULVH�LQV\PPHWULF�DQG�DV\PPHWULF�GLYLVLRQ�]RQHV�DQG�PLJUDWH WKURXJK�VSHFLILF�SKDVHV��1DW�1HXURVFL�����±����
2\OHU�*$��+LJJLQV�*$��+DUW�5$��%DWWHQEHUJ�(��%LOOLQJVOH\ 0��%ORRP�)(��:LOVRQ�0&�������7KH�LGHQWLILFDWLRQ�RI�D�QRYHO�V\QDSWRVRPDO�DVVRFLDWHG SURWHLQ��61$3�����GLIIHUHQWLDOO\H[SUHVVHG�E\�QHXURQDO�VXESRSXODWLRQV��-�&HOO�%LRORJ\ ��������±�����
3DQ�'��������7KH�+LSSR�6LJQDOLQJ�3DWKZD\�LQ�'HYHORSPHQW DQG�&DQFHU��'HY�&HOO�������±����
��

)DEUD�%HVHU�HW�DO�
3DQGD�'.��0LDR�'��/HIHEYUH�9��+HQG\�*1��*ROW]PDQ�'��������7KH�7UDQVFULSWLRQ�)DFWRU62;��5HJXODWHV�&HOO�&\FOH�DQG�'LIIHUHQWLDWLRQ�*HQHV LQ�&KRQGURF\WLF�&).��&HOOV ��-�%LRO&KHP����������±������
3ROOHX[�)��*KRVK�$��������7KH�6OLFH�2YHUOD\�$VVD\� $�9HUVDWLOH�7RRO�WR�6WXG\�WKH�,QIOXHQFH�RI([WUDFHOOXODU�6LJQDOV�RQ�1HXURQDO�'HYHORSPHQW��6FL 6WNH������SO�±SO��
3RUODQ�(��0DUWt�3UDGR�%��0RUDQWH�5HGRODW�-0��&RQVLJOLR $��'HOJDGR�$&��.\SWD�5��/ySH]�2WtQ&��.LUVWHLQ�0��)DULxDV�,��������07��003�UHJXODWHV DGXOW�QHXUDO�VWHP�FHOO�IXQFWLRQDOTXLHVFHQFH�WKURXJK�WKH�FOHDYDJH�RI�1�FDGKHULQ��1DW &HOO�%LRO�������±����
5HLOOR�,��5RPHUR�&�GH�-��*DUFtD�&DEH]DV�0È��%RUUHOO 9��������$�5ROH�IRU�,QWHUPHGLDWH�5DGLDO*OLD�LQ�WKH�7DQJHQWLDO�([SDQVLRQ�RI�WKH�0DPPDOLDQ &HUHEUDO�&RUWH[��&HUHE�&RUWH[�������±�����
5RFKH�.&��*UDF]�$'��/LX�;)��1HZWRQ�9��$NL\DPD�+��0DJQHVV 67��������62;��0DLQWDLQV5HVHUYH�6WHP�&HOOV�DQG�3UHVHUYHV�5DGLRUHVLVWDQFH�LQ 0RXVH�6PDOO�,QWHVWLQH�*DVWURHQWHURORJ\���������������H���
6DKDULD�$��*XLWWDW�/��&URFNHU�6��/LP�$��6WHIIHQ�0� .XONDUQL�6��6WHZDUW�6$��������)ODS(QGRQXFOHDVH���&RQWULEXWHV�WR�7HORPHUH�6WDELOLW\� &XUU�%LRO�������±����
6FKOHJHOPLOFK�.��0RKVHQL�0��.LUDN�2��3UXV]DN�-��5RGULJXH] -5��=KRX�'��.UHJHU�%7�9DVLRXNKLQ�9��$YUXFK�-��%UXPPHONDPS�75��&DPDUJR�)' �������<DS��$FWV�'RZQVWUHDP�RI.�&DWHQLQ�WR�&RQWURO�(SLGHUPDO�3UROLIHUDWLRQ��&HOO �������±����
6FKPHLHU�6��$ODP�7��(VVDFN�0��%DMLF�9%��������7FR)�'% Y���XSGDWH�RI�WKH�GDWDEDVH�RI�KXPDQDQG�PRXVH�WUDQVFULSWLRQ�FR�IDFWRUV�DQG�WUDQVFULSWLRQ IDFWRU�LQWHUDFWLRQV��1XFOHLF�$FLGV�5HV���'���±'����
6FKQLW]HU�-��)UDQNH�:��6FKDFKQHU�0��������,PPXQRF\WRFKHPLFDO GHPRQVWUDWLRQ�RI�YLPHQWLQ�LQDVWURF\WHV�DQG�HSHQG\PDO�FHOOV�RI�GHYHORSLQJ�DQG�DGXOW PRXVH�QHUYRXV�V\VWHP��-�&HOO%LRORJ\�������±����
6FRWW�&(��:\QQ�6/��6HVD\�$��&UX]�&��&KHXQJ�0��*DYLUR 0�9*��%RRWK�6��*DR�%��&KHDK�.6(�/RYHOO�%DGJH�5��%ULVFRH�-��������62;��LQGXFHV�DQG PDLQWDLQV�QHXUDO�VWHP�FHOOV��1DW1HXURVFL��������±�����
6KHQ�4��:DQJ�<��'LPRV�-7��)DVDQR�&$��3KRHQL[�71��/HPLVFKND ,5��,YDQRYD�1%��6WLIDQL�6�0RUULVH\�((��7HPSOH�6��������7KH�WLPLQJ�RI�FRUWLFDO QHXURJHQHVLV�LV�HQFRGHG�ZLWKLQOLQHDJHV�RI�LQGLYLGXDO�SURJHQLWRU�FHOOV��1DW�1HXURVFL �����±����
6WROW�&&��/RPPHV�3��6RFN�(��&KDERLVVLHU�0�&��6FKHGO $��:HJQHU�0��������7KH�6R[�WUDQVFULSWLRQ�IDFWRU�GHWHUPLQHV�JOLDO�IDWH�FKRLFH LQ�WKH�GHYHORSLQJ�VSLQDO�FRUG��*HQH�'HY�������±�����
7DYHUQD�(��*|W]�0��+XWWQHU�:%��������7KH�&HOO�%LRORJ\ RI�1HXURJHQHVLV��7RZDUG�DQ8QGHUVWDQGLQJ�RI�WKH�'HYHORSPHQW�DQG�(YROXWLRQ�RI WKH�1HRFRUWH[��$QQX�5HY�&HOO�'HY�%L����±���
7HOOH\�/��$JLUPDQ�*��3UDGRV�-��$PEHUJ�1��)LqYUH�6� 2EHUVW�3��%DUWROLQL�*��9LWDOL�,��&DGLOKDF�&�+LSSHQPH\HU�6��1JX\HQ�/��'D\HU�$��-DEDXGRQ�'������� 7HPSRUDO�SDWWHUQLQJ�RI�DSLFDOSURJHQLWRUV�DQG�WKHLU�GDXJKWHU�QHXURQV�LQ�WKH�GHYHORSLQJ QHRFRUWH[��6FLHQFH�����HDDY�����
7HOOH\�/��*RYLQGDQ�6��3UDGRV�-��6WHYDQW�,��1HI�6� 'HUPLW]DNLV�(��'D\HU�$��-DEDXGRQ�'�������6HTXHQWLDO�WUDQVFULSWLRQDO�ZDYHV�GLUHFW�WKH�GLIIHUHQWLDWLRQ RI�QHZERUQ�QHXURQV�LQ�WKH�PRXVHQHRFRUWH[��6FLHQFH���������±�����
7UDSQHOO�&��&DFFKLDUHOOL�'��*ULPVE\�-��3RNKDUHO�3� /L�6��0RUVH�0��/HQQRQ�1-��/LYDN�.-�0LNNHOVHQ�76��5LQQ�-/��������7KH�G\QDPLFV�DQG�UHJXODWRUV RI�FHOO�IDWH�GHFLVLRQV�DUHUHYHDOHG�E\�SVHXGRWHPSRUDO�RUGHULQJ�RI�VLQJOH�FHOOV� 1DW�%LRWHFKQRO�������±����
9RQJ�.,��/HXQJ�&.<��%HKULQJHU�55��.ZDQ�.0��������6R[� LV�FULWLFDO�IRU�VXSSUHVVLRQ�RIQHXURJHQHVLV�EXW�QRW�LQLWLDWLRQ�RI�JOLRJHQHVLV�LQ WKH�FHUHEHOOXP��0RO�%UDLQ������
:DQJ�/��=KDQJ�=��<X�;��+XDQJ�;��/LX�=��&KDL�<��<DQJ /��:DQJ�4��/L�0��=KDR�-��+RX�-��/L�)�������8QEDODQFHG�<$3±62;��FLUFXLW�GULYHV�VWHPQHVV DQG�PDOLJQDQW�SURJUHVVLRQ�LQHVRSKDJHDO�VTXDPRXV�FHOO�FDUFLQRPD��2QFRJHQH��������±�����
<RRQ�.��*DLDQR�1��������1RWFK�VLJQDOLQJ�LQ�WKH�PDPPDOLDQ FHQWUDO�QHUYRXV�V\VWHP��LQVLJKWVIURP�PRXVH�PXWDQWV��1DW�1HXURVFL������±����
��

)DEUD�%HVHU�HW�DO�
<X]ZD�6$��%RUUHWW�0-��,QQHV�%7��9RURQRYD�$��.HWHOD�7��.DSODQ�'5��%DGHU�*'��0LOOHU�)'�������'HYHORSPHQWDO�(PHUJHQFH�RI�$GXOW�1HXUDO�6WHP &HOOV�DV�5HYHDOHG�E\�6LQJOH�&HOO7UDQVFULSWLRQDO�3URILOLQJ��&HOO�5HSRUWV��������±�����
=DQFRQDWR�)��&RUGHQRQVL�0��3LFFROR�6��������<$3�7$= DW�WKH�5RRWV�RI�&DQFHU��&DQFHU�&HOO������±����
�
��

texto
Figure 1

Figure 2
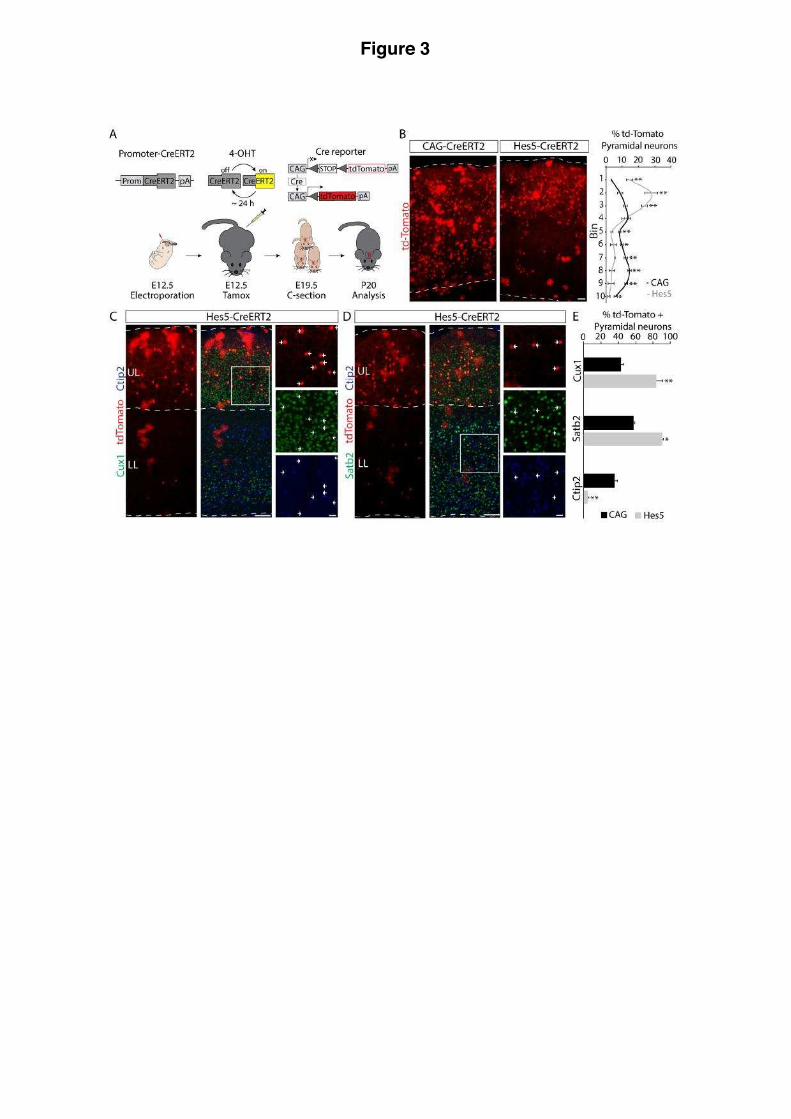
Figure 3

Figure 4

Figure 5
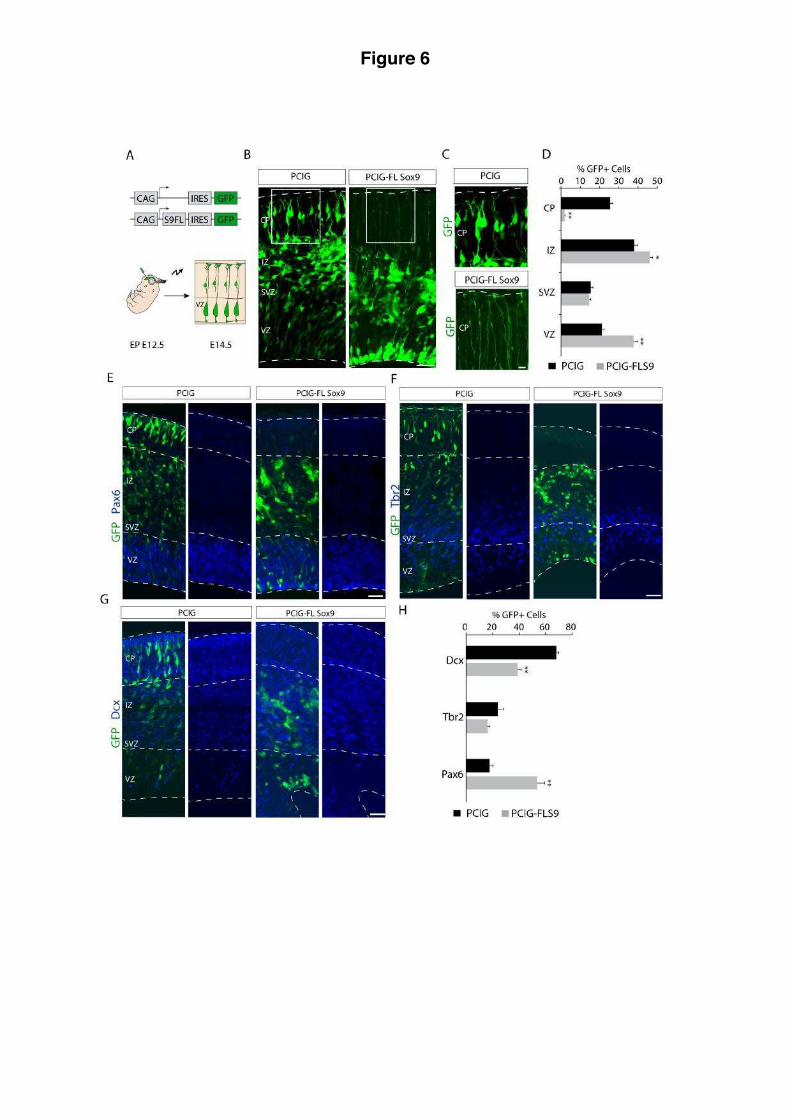
Figure 6

Figure 7

Figure 8

Figure 9

Figure 3-1

Figure 4-1

Figure 5-1

Figure 6-1
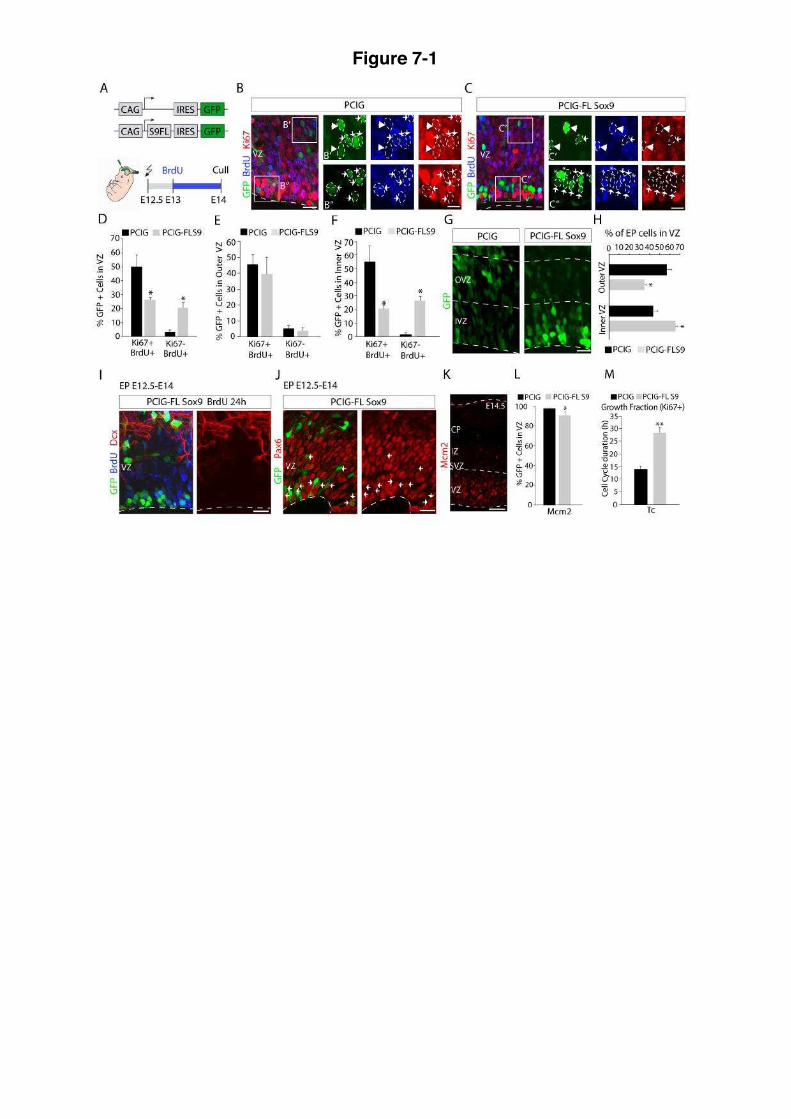
Figure 7-1

Figure 9-1

'LVFXVVmR
(YHQWRV ELROyJLFRV� VHMDP ILVLROyJLFRV RX SDWROyJLFRV� VmR RUTXHVWUDGRV SHOD
FRQVWDQWH LQWHUDomR HQWUH GLYHUVDV SURWHtQDV� TXH SRU VXD YH] VmR R SURGXWR GD
WUDQVFULomR JrQLFD H WUDGXomR GR P51$� 'HVWD IRUPD� R HVWXGR GR WUDQVFULSWRPD p
XPD IRUPD LQGLUHWD GH DYDOLDUPRV SRVVtYHLV PHFDQLVPRV UHJXODGRUHV GRV HYHQWRV
ELROyJLFRV GH LQWHUHVVH� (QWHQGHU R SDSHO GH XP WUDQVFULWR� VXD FRPSRVLomR H VXD
H[SUHVVmR QXP GHWHUPLQDGR PRPHQWR�VLVWHPD ELROyJLFR� SRGH DFDUUHWDU QXPD
GHVFREHUWD UHOHYDQWH SDUD R HYHQWR HVWXGDGR� 1HVWD WHVH� QyV DSUHVHQWDPRV GRLV
H[HPSORV GH FRPR R HVWXGR GD H[SUHVVmR JrQLFD SRGH FRQWULEXLU SDUD R HVWXGR GH
IHQ{PHQRV ILVLROyJLFRV �GHVHQYROYLPHQWR GR FyUWH[ FHUHEUDO� H SDWROyJLFRV �GRHQoD
GH $O]KHLPHU�� OHYDQGR HP FRQVLGHUDomR R XVR GH JHQHV�WUDQVFULWRV
GLIHUHQFLDOPHQWH�H[SUHVVRV�XVDGRV�
$XPHQWDQGR D UHVROXomR GR HVWXGR GH DOWHUDo}HV GD H[SUHVVmR JrQLFD QD GRHQoD
GH�$O]KHLPHU�XWLOL]DQGR�R�XVR�GLIHUHQFLDO�GH�WUDQVFULWRV �'78�
1R DUWLJR 0DUTXHV�&RHOKR H FRODERUDGRUHV ������� YLVWR QHVWD WHVH� QyV
XWLOL]DPRV WUrV GLIHUHQWHV GDWDVHWV FRP GDGRV GH 51$�VHT GR FpUHEUR GH LQGLYtGXRV
GLDJQRVWLFDGRV FRP RX VHP GRHQoD GH $O]KHLPHU� 1yV PRVWUDPRV TXH DV
DOWHUDo}HV GD H[SUHVVmR JrQLFD HP GLIHUHQWHV iUHDV GR FpUHEUR VHJXHP R PHVPR
SDGUmR GHVFULWR QD SDWRORJLD SDUD R GHSyVLWR GH SURWHtQD 7$8 KLSHUIRVIRULODGD�
6XUSUHHQGHQWHPHQWH� QRVVRV GDGRV PRVWUDP TXH XP Q~PHUR VLJQLILFDWLYR GH JHQHV
DSUHVHQWD XVR GLIHUHQFLDO GH WUDQVFULWRV �'78�� VHP DSUHVHQWDU DOWHUDo}HV QD VXD
���

H[SUHVVmR JOREDO� (P RXWUDV SDODYUDV� HVWHV JHQHV QmR VmR GHWHFWDGRV HP DQiOLVHV
FOiVVLFDV GH H[SUHVVmR GLIHUHQFLDO� PDV DSUHVHQWDP PXGDQoDV QR VHX SDGUmR GH
VSOLFLQJ DOWHUQDWLYR TXH SRGH WHU LPSRUWDQWHV FRQVHTXrQFLDV IXQFLRQDLV� 'H IDWR�
YiULRV JHQHV FRP '78 HVWmR UHODFLRQDGRV j DWLYLGDGH VLQiSWLFD� TXH p VHYHUDPHQWH
DIHWDGD�QD�GRHQoD�GH�$O]KHLPHU�
1mR REVWDQWH� DOJXQV DVSHFWRV WpFQLFRV GHYHP VHU REVHUYDGRV QD DQiOLVH
GHVWHV UHVXOWDGRV� 3ULPHLUDPHQWH� DV LVRIRUPDV GHVFULWDV HP QRVVR DUWLJR IRUDP
LGHQWLILFDGDV D SDUWLU GR EDQFR GH GDGRV XWLOL]DGR *(1&2'( �)UDQNLVK HW DO� ������
6HQGR DVVLP� VHULD UHOHYDQWH GHVFUHYHU R XVR GLIHUHQFLDO GH WUDQVFULWRV �'78� FRP R
XVR GH XPD HVWUDWpJLD GH SUHGLomR GH QRYRV WUDQVFULWRV� WDO FRPR R XVR GH
DOLQKDGRUHV GLUHWRV FRPR 67$5 �'RELQ HW DO� ����� RX +LVDW� �.LP HW DO� �����
DOLDGR D DOJXP PRQWDGRU GH WUDQVFULWRV FRPR &XIIOLQNV �7UDSQHOO HW DO� ����� RX
6WULQJ7LH� �3HUWHD HW DO� ������ $ ~QLFD OLPLWDomR TXH SRGHULD VHU HQFRQWUDGD p QR
GDWDVHW 06%% TXH DSUHVHQWD UHDGV VLQJOH�HQG GH WDPDQKR UHGX]LGR VH FRPSDUDGR
DRV RXWURV GDWDVHWV� &RPR GHVFULWR SRU 5RVVHOO H FRODERUDGRUHV UHDGV VLQJOH�HQG VH
OLPLWDP D XP ODGR GR IUDJPHQWR GH P51$� HQTXDQWR UHDGV SDLUHG�HQG SRGHP WHU
DPEDV DV SRVLo}HV GR IUDJPHQWR GH P51$� VHQGR DVVLP� WRUQD�VH PDLV IiFLO
LGHQWLILFDU TXDLV H[RQV HVWmR SUHVHQWHV QXP UHDG H FRQVHTXHQWHPHQWH TXDO
WUDQVFULWR�RX�HYHQWR�GH�VSOLFLQJ�HVWi�VHQGR�LGHQWLILFDGR �5RVVHOO�HW�DO��������
2XWUD HVWUDWpJLD SDUD DYDOLDU R VLJQLILFDGR GDV DOWHUDo}HV GD H[SUHVVmR
JrQLFD QD GRHQoD GH $O]KHLPHU p EDVHDGD QR HVWXGR GH UHGHV GH FRH[SUHVVmR H
VHXV PyGXORV �0LOLQG HW DO� :DQ HW DO� ������ :DQ H FRODERUDGRUHV XWLOL]DUDP DV
DVVLQDWXUDV HQFRQWUDGDV HP PRGHORV GH $O]KHLPHU FRP FDPXQGRQJR H RV PyGXORV
���

GH FRH[SUHVVmR UHWLUDGRV GH GDGRV GH 51$VHT HP KXPDQRV SDUD GHPRQVWUDU FRPR
DOJXQV PRGHORV GH $' HP FDPXQGRQJR VmR FDSD]HV GH PLPHWL]DU R TXH DFRQWHFH
HP KXPDQR� 'H PDQHLUD VHPHOKDQWH� 0LOLQG H FRODERUDGRUHV IDEULFDUDP PyGXORV GH
FRH[SUHVVmR FRP D IHUUDPHQWD :*&1$ XWLOL]DQGR GDGRV GH 51$VHT KXPDQR H
REWLYHUDP WDQWR PyGXORV FRPR VXEPyGXORV FDSD]HV GH FRUUHODFLRQDU FRP
GLIHUHQWHV HYHQWRV GH VLQDOL]DomR FpOXODV�HVSHFtILFR� (P DPERV RV DUWLJRV� RV GDGRV
XWLOL]DGRV IRUDP RV PHVPRV GR DUWLJR 0DUTXHV�&RHOKR HQFRQWUDGR QHVWD WHVH� $LQGD
VHULD UHOHYDQWH FRPSDUDU DV DVVLQDWXUDV XWLOL]DGDV QRV PRGHORV GH FDPXQGRQJR
XVDGRV SRU :DQ FRP DV DVVLQDWXUDV HQFRQWUDGDV QR QRVVR WUDEDOKR� SULQFLSDOPHQWH
FRP RV QRVVRV UHVXOWDGRV GH '78� 2XWUD DERUGDJHP SRVVtYHO p WDPEpP JHUDU R
UHVXOWDGR GH '78 HP PRGHORV GH FDPXQGRQJRV Mi SXEOLFDGRV� VHQGR HVVD
DERUGDJHP OLPLWDGD D H[SHULPHQWRV FRP 51$VHT� $OpP GLVVR� SRGHUtDPRV XWLOL]DU
RV PyGXORV GH FRH[SUHVVmR IDEULFDGRV SRU 0LOLQG SDUD YHULILFDU TXDLV HYHQWRV
FHOXODUHV�SRGHULDP�VHU�DIHWDGRV�SHOR�XVR�GLIHUHQFLDO GH�WUDQVFULWRV�
3RU RXWUR ODGR� 5DM H FRODERUDGRUHV GHVFUHYHUDP D SUHVHQoD TXDQWLWDWLYD GH
ORFXV UHODFLRQDGR D VSOLFLQJ �V47/� GR LQJOrV� VSOLFLQJ 4XDQWLWDWLYH 7UDLW /RFL�� QR
HQWDQWR� R WUDEDOKR VH OLPLWRX DR GDWDVHW 5260$3 TXH SRVVXL VRPHQWH GDGRV
REWLGRV D SDUWLU GR FyUWH[ SUHIURQWDO GRUVRODWHUDO �'/3)&� GR LQJOrV� GRUVRODWHUDO
SUHIURQWDO FRUWH[� H XPD UHSOLFDomR SDUFLDO QR GDWDVHW 06%% FRP � iUHDV GLVWLQWDV
�5DM HW DO� ������ &RPR GHVFULWR SHOR DUWLJR GR FDStWXOR � GD WHVH� H[LVWH XPD
GLIHUHQoD H[SUHVVLYD HQWUH DOWHUDo}HV QDV UHJL}HV GR FyUWH[ IURQWDO YV� FyUWH[
WHPSRUDO� R TXH WRUQD D IDOWD GH VHSDUDomR HQWUH DV iUHDV GR GDWDVHW 06%% QR
DUWLJR XP SRQWR D VHU GLVFXWLGR� $OpP GLVVR� RV DXWRUHV LQFOXtUDP QR JUXSR
���

�$O]KHLPHU� GR GDGR 5260$3� XP VXEJUXSR GH LQGLYtGXRV FODVVLILFDGRV FRP
$O]KHLPHU H WDPEpP XPD RXWUD FRQGLomR �QmR LGHQWLILFDGD� TXH FRQWULEXL SDUD D
SHUGD FRJQLWLYD �V\QDSVH LG� V\Q��������� R TXH SRGH WHU OHYDGR D LGHQWLILFDomR GH
DOWHUDo}HV GD H[SUHVVmR JrQLFD LQHVSHFtILFDV �5DM HW DO� ������ 7DO VXEJUXSR TXDQGR
DQDOLVDGR VHSDUDGDPHQWH DSUHVHQWRX GLIHUHQoDV VLJQLILFDWLYDV �YLGH� '(*� GDGRV
QmR SXEOLFDGRV� WDQWR FRP R JUXSR FRQWUROH TXDQWR FRP R JUXSR $O]KHLPHU VHP
RXWUDV�FDXVDV�GH�SHUGD�FRJQLWLYD�
2XWUD DERUGDJHP FRPSOHPHQWDU DR QRVVR WUDEDOKR� H TXH SRGHULD FRQWULEXLU
SDUD FRQILUPDU DV DVVLQDWXUDV JrQLFDV FpOXODV�HVSHFtILFDV TXH GHVFUHYHPRV
XWLOL]DQGR XPD DERUGDJHP LQGLUHWD� VHULD R XVR GH DOJXP DOJRULWPR GH GHFRQYROXomR
XVDQGR GDGRV GH VF51$VHT H GDWDVHWV GH 51$VHT GH WHFLGR SUHVHQWHV QHVVH
WUDEDOKR� $ WpFQLFD GH GHFRQYROXomR QRV SHUPLWH LQIHULU DV SURSRUo}HV GH WLSRV
FHOXODUHV SUHVHQWHV QRV GDGRV GH EXON 51$VHT SRU PHLR GR XVR GH GDGRV GH
VF51$VHT� VHQGR DVVLP� SRGHUtDPRV LQIHULU RV WLSRV FHOXODUHV DIHWDGRV QDV
DOWHUDo}HV�SUHVHQWHV�QR�FpUHEUR�FRP�$O]KHLPHU��'X HW�DO��������:DQJ�HW�DO��������
'DGR LVVR� RV UHVXOWDGRV HQFRQWUDGRV QR DUWLJR GH 0DUTXHV�&RHOKR
FRUURERUDP HYLGrQFLDV Mi GHVFULWDV QD OLWHUDWXUD FRPR� DXPHQWR PDLV DFHQWXDGR H
FHGR GH DOWHUDo}HV QD UHJLmR WHPSRUDO VH FRPSDUDGR FRP D UHJLmR SUHIURQWDO �%UDDN
HW DO� ����� 0RUDELWR HW DO� ������ DVVLP FRPR DSUHVHQWD XP QRYR FDPLQKR QD
GLYHUVLGDGH GH VSOLFLQJ DOWHUQDWLYR� SUHGLomR GH GRPtQLRV IXQFLRQDLV UHOHYDQWHV H
SRVVtYHLV DOWHUDo}HV HQFRQWUDGRV QD UHJLmR IURQWR�WHPSRUDO GR FpUHEUR FRP
$O]KHLPHU �5DM HW DO� ������ $OpP GLVVR� QRVVR WUDEDOKR WUD] QRYDV LQIRUPDo}HV GH
JUDQGH VLJQLILFkQFLD SDUD R HQWHQGLPHQWR GD SDWRORJLD GD GRHQoD GH $O]KHLPHU�
���

FRPR R JUDQGH Q~PHUR GH '78V H VXD SURJUHVVmR FRP RV HVWiJLRV %UDDN GD
GRHQoD�
3RU ILP� QyV XWLOL]DPRV XPD WpFQLFD LQGLUHWD SDUD PDSHDU DV DOWHUDo}HV GD
H[SUHVVmR JrQLFD LGHQWLILFDGDV HP EXON 51$�VHT SDUD WLSRV FHOXODUHV HVSHFtILFRV GR
FpUHEUR DGXOWR� (VVD HVWUDWpJLD RULJLQDO QRV SHUPLWLX LGHQWLILFDU DVVLQDWXUDV JrQLFDV
WLSR�FHOXODU HVSHFtILFDV FRPSDUiYHLV FRP DTXHODV LGHQWLILFDGDV DWUDYpV GH
VF51$VHT �*UXEPDQ� 0DWK\V�� $OpP GD YDQWDJHP GH SHUPLWLU LQWHUSUHWDU DOWHUDo}HV
GD H[SUHVVmR JrQLFD WHFLGXDO DR QtYHO GDV FpOXODV� QRVVD HVWUDWpJLD SHUPLWH DYDOLDU
LQGLUHWDPHQWH D H[SUHVVmR GH WUDQVFULWRV HP WLSRV FHOXODUHV ~QLFRV� R TXH QmR p
SRVVtYHO�QD�PDLRU�SDUWH�GRV�GDGRV�GH�VF51$VHT�GHYLGR DR��¶�ELDV�
8VDQGR VF51$VHT H UHGHV SDUD LGHQWLILFDU UHJXODGRUHV GD HVSHFLILFDomR FHOXODU
GXUDQWH�R�GHVHQYROYLPHQWR�GR�FyUWH[�FHUHEUDO
-i QR DUWLJR GH )DEUD�%HVHU H FRODERUDGRUHV ������� WDPEpP SUHVHQWH
QHVWD WHVH� QyV GHPRQVWUDPRV R SRWHQFLDO GR VHTXHQFLDPHQWR GH 51$ HP FpOXODV
~QLFDV SDUD LGHQWLILFDU VXESRSXODo}HV FHOXODUHV� 3ULPHLUR� QyV PRVWUDPRV TXH XPD
VXESRSXODomR GH SURJHQLWRUHV QHXUDLV DSUHVHQWD XPD DVVLQDWXUD JrQLFD
FDUDFWHUL]DGD SRU JHQHV UHJXODGRV SHOR IDWRU GH WUDQVFULomR 6R[�� (P VHJXLGD�
WDPEpP GHPRQVWUDPRV TXH HVWH IDWRU GH WUDQVFULomR QmR Vy ID] SDUWH GH XPD UHGH
GH UHJXORQV� DVVLP FRPR p XP UHJXODGRU PHVWUH HVSHFtILFR GD VXESRSXODomR GH
SURJHQLWRUHV� (VWH p SURYDYHOPHQWH R WUDEDOKR SLRQHLUR D XVDU GDGRV GH VF51$VHT
H D PHWRGRORJLD FULDGD SRU &DVWUR H FRODERUDGRUHV �&DVWUR HW DO� ������ 'H DFRUGR
FRP HVWD LQWHUSUHWDomR� D PDQLSXODomR GD H[SUHVVmR GH 6R[� HP SURJHQLWRUHV IRL
���

VXILFLHQWH SDUD DOWHUDU R SRWHQFLDO GH SURJHQLWRUHV QHXUDLV� FRQILUPDQGR D UHOHYkQFLD
GRV�DFKDGRV�GH�ELRLQIRUPiWLFD�
8PD HVWUDWpJLD WDPEpP DERUGDGD FRP RV GDGRV GH )DEUD�%HVHU IRL D
FDUDFWHUL]DomR SVHXGRWHPSRUDO GRV JUXSRV FHOXODUHV HQFRQWUDGRV �GDGRV QmR
SXEOLFDGRV�� &RP R XVR GD ELEOLRWHFD 0RQRFOH� �4LX HW DO� ������ IRL YLVWR XP
JUDGLHQWH GH H[SUHVVmR FRPHoDQGR QRV 5*&V� SDVVDQGR SHOR ,3V H WHUPLQDQGR QRV
QHXU{QLRV� (VVH PDSD SVHXGRWHPSRUDO HVWi GH DFRUGR FRP R TXH Mi p FRQKHFLGR QD
IRUPDomR GR FyUWH[ FHUHEUDO� 1R HQWDQWR� QmR IRL SRVVtYHO YLVXDOL]DU� SRU PHLR GD
DQiOLVH SVHXGRWHPSRUDO� FDPLQKRV PROHFXODUHV LQGHSHQGHQWHV QD IRUPDomR GDV
FDPDGDV LQIUD H VXSUDJUDQXODUHV Mi QD VXESRSXODomR 5*&V� 'HYLGR DR EDL[R
Q~PHUR GH FpOXODV QR VXEJUXSR 5*& +HV�� QmR p SRVVtYHO GHILQLU VH D DXVrQFLD GD
VHSDUDomR IRL FDXVDGD SRU XPD EDL[D FREHUWXUD RX SUR[LPLGDGH QDWXUDO QRV HYHQWRV
PROHFXODUHV�HQFRQWUDGRV�HP�DPERV�RV�VXEJUXSRV�GH�5*&V�
'HYLGR D RVFLODomR GD VLQDOL]DomR GH 1RWFK Mi WHU VLGR GHVFULWD HP
SURJHQLWRUHV QHXUDLV �6KLPRMR HW DO� ������ XPD RXWUD SURSRVWD VHULD D DQiOLVH GD
RVFLODomR GH JHQHV FRP R XVR GRV GDGRV GH VF51$VHT� /HQJ H FRODERUDGRUHV Mi
WHVWDUDP D KLSyWHVH GD RVFLODomR� FRP R XVR GR DOJRULWPR 2VFRSH� REWHQGR
IDFLOPHQWH D RVFLODomR GH PDUFDGRUHV FRPR RV GR FLFOR�FHOXODU �/HQJ HW DO� ������
(VVD KLSyWHVH OHYDULD HP FRQVLGHUDomR TXH RV VXEJUXSRV GH 5*&V ³QmR H[LVWHP´ H
VmR QD YHUGDGH JUDGLHQWHV GH H[SUHVVmR GH XP PHVPR WLSR FHOXODU� 8P SRQWR
UHOHYDQWH p TXH R DOJRULWPR 2VFRSH OLGD EHP FRP XP YROXPH GH FpOXODV DEDL[R GH
�����R�TXH�SRGH�VHU�YDQWDMRVR�GHYLGR�DR�Q~PHUR�OLPLWDGR GH�FpOXODV�SRU�VXEJUXSR�
���

&RPR DV FpOXODV VHTXHQFLDGDV QR WUDEDOKR GH )DEUD�%HVHU XVDUDP D
WpFQLFD VPDUW�6HT� �3LFHOOL HW DO� ������ DLQGD VHULD SRVVtYHO ODQoDU PmR GH
HVWUDWpJLDV TXH XWLOL]HP R VHTXHQFLDPHQWR GH WRGR R WUDQVFULWR� $ PDLRULD GRV
WUDEDOKRV FRP VF51$VHT XWLOL]DP D WpFQLFD ��; JHQRPLFV TXH SHUPLWH R
VHTXHQFLDPHQWR GH PLOKDUHV D PLOK}HV GH FpOXODV HP XP ~QLFR H[SHULPHQWR� QR
HQWDQWR� R �¶�ELDV GHVVH WLSR VHTXHQFLDPHQWR SHUPLWH VRPHQWH DQiOLVHV GD
H[SUHVVmR GDV FpOXODV� &RPR VPDUW�6HT� YLDELOL]D R VHTXHQFLDPHQWR GH WRGR R
WUDQVFULWR� VHULD SRVVtYHO REWHU QRYRV WUDQVFULWRV QXQFD DQWHV GHVFULWRV QRV
GLIHUHQWHV�WLSRV�FHOXODUHV�HQFRQWUDGRV�
/RJR� RV DFKDGRV HQFRQWUDGRV QR WUDEDOKR GH )DEUD�%HVHU UHIRUoDP D LGHLD
GD H[LVWrQFLD GH VXESRSXODo}HV SXWDWLYDV HP HVWiJLRV LQLFLDLV GR GHVHQYROYLPHQWR
GR FyUWH[ �)UDQFR HW DO� ������ VHQGR TXH HVVH HYHQWR VHULD UHJXODGR SULQFLSDOPHQWH
SHOR�IDWRU�GH�WUDQVFULomR�6R[��
���

&RQFOXVmR
2V UHVXOWDGRV HQFRQWUDGRV QHVWD WHVH QRV SHUPLWHP FKHJDU HP DOJXPDV
FRQFOXV}HV�
� $ DQiOLVH GH LVRIRUPDV SHUPLWH LGHQWLILFDU QRYDV DOWHUDo}HV GD H[SUHVVmR
JrQLFDV�QR�FpUHEUR�GH�SDFLHQWHV�FRP�D�GRHQoD�GH�$O]KHLPHU�
� $OWHUDo}HV QR XVR GH LVRIRUPDV GHWHFWDGDV QD GRHQoD GH $O]KHLPHU
SRGHP WHU LPSRUWDQWHV FRQVHTXrQFLDV IXQFLRQDLV� FRQWULEXLQGR SDUD D
SURJUHVVmR�GD�SDWRORJLD�
� 2 XVR GH VF51$�VHT SHUPLWH PDSHDU JHQHV�WUDQVFULWRV GLIHUHQFLDOPHQWH
H[SUHVVRV�HP�WLSRV�FHOXODUHV�HVSHFtILFRV�GR�FpUHEUR KXPDQR�
� (VWXGRV IXWXURV GHYHUmR DYDOLDU GH TXH IRUPD DV DOWHUDo}HV GH VSOLFLQJ
DOWHUQDWLYR LGHQWLILFDGDV QD GRHQoD GH $O]KHLPHU WDPEpP RFRUUHP HP
RXWUDV�GRHQoDV�QHXURGHJHQHUDWLYDV�
� $QiOLVHV GRV GDGRV GH VF51$�VHT XWLOL]DQGR UHGHV UHJXODWyULDV
SHUPLWLUDP LGHQWLILFDU R IDWRU GH WUDQVFULomR 6R[� FRPR XP LPSRUWDQWH
UHJXODGRU GD SUROLIHUDomR�GLIHUHQFLDomR GH SURJHQLWRUHV QHXUDLV QR
GHVHQYROYLPHQWR�GR�FyUWH[�FHUHEUDO�HP�FDPXQGRQJRV�
� 1R IXWXUR� VHULD LQWHUHVVDQWH XWLOL]DU GDGRV GH VF51$VHT REWLGRV GR
FyUWH[ FHUHEUDO KXPDQR HP GHVHQYROYLPHQWR SDUD DYDOLDU VH D H[SUHVVmR
GH 6R[� HVWi DXPHQWDGD HP SRSXODo}HV GH SURJHQLWRUHV DVVRFLDGDV FRP
D JHUDomR GH QHXU{QLRV VXSUDJUDQXODUHV� FRPR RV GD ]RQD VXEYHQWULFXODU
H[WHUQD�
���

� $ H[SUHVVmR GLIHUHQFLDO GH JHQHV� DOLDGD D XP ERP GHVHQKR
H[SHULPHQWDO� p XP H[FHOHQWH PpWRGR SDUD WUDoDU D LPSRUWkQFLD GH JHQHV
H DWp PHVPR VtWLRV GH OLJDomR LPSRUWDQWHV� $OpP GLVVR� UHGHV GH
LQWHUDomR SURWHtQD�SURWHtQD DVVLP FRPR RQWRORJLDV VmR LPSRUWDQWHV
PHWRGRORJLDV�DX[LOLDUHV�SDUD�VDOLHQWDU�R�HYHQWR�ELROyJLFR HVWXGDGR�
� (VWD WHVH H VHXV UHVXOWDGRV QRV SHUPLWHP HQWHQGHU D LPSRUWkQFLD GR
WUDQVFULSWRPD QD HOXFLGDomR GH HYHQWRV ELROyJLFRV HQYROYLGRV HP
SURFHVVRV�ILVLROyJLFRV�H�SDWROyJLFRV�
���

5HIHUrQFLDV�RUGHP�DOIDEpWLFD�
$LEDU� 6�� *RQ]iOH]�%ODV� &� %�� 0RHUPDQ� 7�� +X\QK�7KX� 9� $�� ,PULFKRYD� +��
+XOVHOPDQV� *�� 5DPERZ� )�� 0DULQH� -� &�� *HXUWV� 3�� $HUWV� -�� 9DQ 'HQ 2RUG� -��
$WDN� =� .�� :RXWHUV� -�� $HUWV� 6� ������� 6&(1,&� 6LQJOH�FHOO UHJXODWRU\ QHWZRUN
LQIHUHQFH DQG FOXVWHULQJ� 1DWXUH 0HWKRGV� ������� ����±�����
KWWSV���GRL�RUJ���������QPHWK�����
$OOHQ� 0�� &DUUDVTXLOOR� 0� 0�� )XQN� &�� +HDYQHU� %� '�� =RX� )�� <RXQNLQ� &� 6��
%XUJHVV� -� '�� &KDL� +� 6�� &URRN� -�� (GG\� -� $�� /L� +�� /RJVGRQ� %�� 3HWHUV� 0� $��
'DQJ� .� .�� :DQJ� ;�� 6HULH� '�� :DQJ� &�� 1JX\HQ� 7�� /LQFROQ� 6�� « (UWHNLQ�7DQHU�
1� ������� +XPDQ ZKROH JHQRPH JHQRW\SH DQG WUDQVFULSWRPH GDWD IRU $O]KHLPHU¶V
DQG RWKHU QHXURGHJHQHUDWLYH GLVHDVHV� 6FLHQWLILF 'DWD� ��
KWWSV���GRL�RUJ���������VGDWD��������
$QGHUV� 6�� 3\O� 3� 7�� +XEHU� :� ������� +76HT��D 3\WKRQ IUDPHZRUN WR ZRUN ZLWK
KLJK�WKURXJKSXW VHTXHQFLQJ GDWD� %LRLQIRUPDWLFV� ������ ���±����
KWWSV���GRL�RUJ���������ELRLQIRUPDWLFV�EWX���
%HFN� $� +�� :HQJ� =�� :LWWHQ� '� 0�� =KX� 6�� )ROH\� -� :�� /DFURXWH� 3�� 6PLWK� &� /��
7LEVKLUDQL� 5�� YDQ GH 5LMQ� 0�� 6LGRZ� $�� :HVW� 5� %� ������� ���(QG 6HTXHQFLQJ
IRU ([SUHVVLRQ 4XDQWLILFDWLRQ ��6(4� IURP $UFKLYDO 7XPRU 6DPSOHV� 3/R6 21(�
������H������KWWSV���GRL�RUJ���������MRXUQDO�SRQH��������
%HOOHQJXH] & HW DO� ������ 1HZ LQVLJKWV RQ WKH JHQHWLF HWLRORJ\ RI $O]KHLPHU¶V DQG
UHODWHG GHPHQWLD� 0HG5[LY�
KWWS���PHGU[LY�RUJ�FRQWHQW�HDUO\�������������������������������
%HUWRQH� 3�� 6WROF� 9�� 5R\FH� 7� (�� 5R]RZVN\� -� 6�� 8UEDQ� $� (�� =KX� ;�� 5LQQ� -�
/�� 7RQJSUDVLW� :�� 6DPDQWD� 0�� :HLVVPDQ� 6�� *HUVWHLQ� 0�� 6Q\GHU� 0� �������
*OREDO LGHQWLILFDWLRQ RI KXPDQ WUDQVFULEHG VHTXHQFHV ZLWK JHQRPH WLOLQJ DUUD\V�
6FLHQFH� ���������������±������KWWSV���GRL�RUJ���������VFLHQFH��������
%UDDN� +�� %UDDN� (� ������� 1HXURSDWKRORJLFDO VWDJHLQJ RI $O]KHLPHU�UHODWHG
FKDQJHV� ,Q $FWD 1HXURSDWKRORJLFD �9RO� ��� ,VVXH �� SS� ���±����� 6SULQJHU�9HUODJ�
KWWSV���GRL�RUJ���������%)��������
%UDDN� +�� $ODIX]RII� ,�� $U]EHUJHU� 7�� .UHW]VFKPDU� +�� 7UHGLFL� .� ������� 6WDJLQJ
RI $O]KHLPHU GLVHDVH�DVVRFLDWHG QHXURILEULOODU\ SDWKRORJ\ XVLQJ SDUDIILQ VHFWLRQV DQG
���

LPPXQRF\WRFKHPLVWU\� $FWD 1HXURSDWKRORJLFD� ������� ���±����
KWWSV���GRL�RUJ���������V���������������]
%UD\� 1� /�� 3LPHQWHO� +�� 0HOVWHG� 3�� 3DFKWHU� /� ������� 1HDU�RSWLPDO SUREDELOLVWLF
51$�VHT TXDQWLILFDWLRQ� 1DWXUH %LRWHFKQRORJ\� ������ ���±����
KWWSV���GRL�RUJ���������QEW�����
&DVWUR� 0� $� $�� 'H 6DQWLDJR� ,�� &DPSEHOO� 7� 0�� 9DXJKQ� &�� +LFNH\� 7� (�� 5RVV�
(�� 7LOOH\� :� '�� 0DUNRZHW]� )�� 3RQGHU� %� $� -�� 0H\HU� .� %� ������� 5HJXODWRUV RI
JHQHWLF ULVN RI EUHDVW FDQFHU LGHQWLILHG E\ LQWHJUDWLYH QHWZRUN DQDO\VLV� 1DWXUH
*HQHWLFV� ���������±����KWWSV���GRL�RUJ���������QJ�����
&KHQ� (� <�� 7DQ� &� 0�� .RX� <�� 'XDQ� 4�� :DQJ� =�� 0HLUHOOHV� *� 9�� &ODUN� 1� 5��
0D¶D\DQ� $� ������� (QULFKU� ,QWHUDFWLYH DQG FROODERUDWLYH +70/� JHQH OLVW
HQULFKPHQW DQDO\VLV WRRO� %0& %LRLQIRUPDWLFV� ���
KWWSV���GRL�RUJ�������������������������
'H -DJHU� 3� /�� 0D� <�� 0F&DEH� &�� ;X� -�� 9DUGDUDMDQ� %� 1�� )HOVN\� '�� .OHLQ� +�
8�� :KLWH� &� &�� 3HWHUV� 0� $�� /RGJVRQ� %�� 1HMDG� 3�� 7DQJ� $�� 0DQJUDYLWH� /� 0��
<X� /�� *DLWHUL� &�� 0RVWDIDYL� 6�� 6FKQHLGHU� -� $�� %HQQHWW� '� $� ������� 'DWD
GHVFULSWRU� $ PXOWL�RPLF DWODV RI WKH KXPDQ IURQWDO FRUWH[ IRU DJLQJ DQG $O]KHLPHU¶V
GLVHDVH UHVHDUFK� 6FLHQWLILF 'DWD� �� ������±�������
KWWSV���GRL�RUJ���������VGDWD���������
'RXUOHQ� 3�� .LOLQF� '�� 0DOPDQFKH� 1�� &KDSXLV� -�� /DPEHUW� -��&� ������� 7KH
QHZ JHQHWLF ODQGVFDSH RI $O]KHLPHU¶V GLVHDVH� IURP DP\ORLG FDVFDGH WR JHQHWLFDOO\
GULYHQ V\QDSWLF IDLOXUH K\SRWKHVLV" $FWD 1HXURSDWKRORJLFD� �±���
KWWSV���GRL�RUJ���������V�����������������
'X� 5�� &DUH\� 9�� :HLVV� 6� 7� ������� 'HFRQY6HT� 'HFRQYROXWLRQ RI FHOO PL[WXUH
GLVWULEXWLRQ LQ VHTXHQFLQJ GDWD� %LRLQIRUPDWLFV� ������� ����±�����
KWWSV���GRL�RUJ���������ELRLQIRUPDWLFV�EW]���
)LOOHQEDXP� *� *�� YDQ %HOOH� *�� 0RUULV� -� &�� 0RKV� 5� &�� 0LUUD� 6� 6�� 'DYLV� 3� &��
7DULRW� 3� 1�� 6LOYHUPDQ� -� 0�� &ODUN� &� 0�� :HOVK�%RKPHU� .� $�� +H\PDQ� $�
������� &RQVRUWLXP WR (VWDEOLVK D 5HJLVWU\ IRU $O]KHLPHU¶V 'LVHDVH �&(5$'�� 7KH
ILUVW WZHQW\ \HDUV� ,Q $O]KHLPHU¶V DQG 'HPHQWLD �9RO� �� ,VVXH �� SS� ��±����� 1,+
3XEOLF�$FFHVV��KWWSV���GRL�RUJ���������M�MDO]������������
*RRODP� 0�� 6FLDOGRQH� $�� *UDKDP� 6� -� /�� 0DF$XOD\� ,� &�� -HGUXVLN� $��
+XSDORZVND� $�� 9RHW� 7�� 0DULRQL� -� &�� =HUQLFND�*RHW]� 0� ������� +HWHURJHQHLW\
���

LQ 2FW� DQG 6R[� 7DUJHWV %LDVHV &HOO )DWH LQ ��&HOO 0RXVH (PEU\RV� &HOO� �������
��±����KWWSV���GRL�RUJ���������M�FHOO������������
*UHLJ� /� &�� :RRGZRUWK� 0� %�� *DOD]R� 0� -�� 3DGPDQDEKDQ� +�� 0DFNOLV� -� '�
������� 0ROHFXODU ORJLF RI QHRFRUWLFDO SURMHFWLRQ QHXURQ VSHFLILFDWLRQ� GHYHORSPHQW
DQG GLYHUVLW\� ,Q 1DWXUH 5HYLHZV 1HXURVFLHQFH �9RO� ��� ,VVXH ��� SS� ���±�����
1DWXUH�3XEOLVKLQJ�*URXS��KWWSV���GRL�RUJ���������QUQ����
+DVKLPVKRQ\� 7�� :DJQHU� )�� 6KHU� 1�� <DQDL� ,� ������� &(/�6HT� 6LQJOH�&HOO
51$�6HT E\ 0XOWLSOH[HG /LQHDU $PSOLILFDWLRQ� &HOO 5HSRUWV� ����� ���±����
KWWSV���GRL�RUJ���������M�FHOUHS������������
+HU]RJ� 9� $�� 5HLFKKROI� %�� 1HXPDQQ� 7�� 5HVFKHQHGHU� 3�� %KDW� 3�� %XUNDUG� 7� 5��
:ORW]ND� :�� 9RQ +DHVHOHU� $�� =XEHU� -�� $PHUHV� 6� /� ������� 7KLRO�OLQNHG
DON\ODWLRQ RI 51$ WR DVVHVV H[SUHVVLRQ G\QDPLFV� 1DWXUH 0HWKRGV� �������
����±������KWWSV���GRL�RUJ���������QPHWK�����
+RZDUG� 5�� /LX� .� <� �Q�G��� 4XHVWLRQV (0(5*( DV %LRJHQ FODLPV DGXFDQXPDE
WXUQDURXQG��KWWSV���GRL�RUJ���������V����������������
+XEHUW� )��;�� .LQNHO� 6� $�� &UHZWKHU� 3� (�� &DQQRQ� 3� =� )�� :HEVWHU� .� (�� /LQN�
0�� 8LER� 5�� 2¶%U\DQ� 0� .�� 0HDJHU� $�� )RUHKDQ� 6� 3�� 6P\WK� *� .�� 0LWWD]� /��
$QWRQDUDNLV� 6� (�� 3HWHUVRQ� 3�� +HDWK� :� 5�� 6FRWW� +� 6� ������� $LUH�'HILFLHQW
&��%/�� 0LFH 0LPLFNLQJ WKH &RPPRQ +XPDQ ���%DVH 3DLU 'HOHWLRQ 0XWDWLRQ
3UHVHQW ZLWK 2QO\ D 0LOG $XWRLPPXQH 3KHQRW\SH� 7KH -RXUQDO RI ,PPXQRORJ\�
������������±������KWWSV���GRL�RUJ���������MLPPXQRO��������
+XJKHV� &� 3�� %HUJ� /�� 'DQ]LJHU� :� /�� &REHQ� /� $�� 0DUWLQ� 5� /� ������� $ QHZ
FOLQLFDO VFDOH IRU WKH VWDJLQJ RI GHPHQWLD� %ULWLVK -RXUQDO RI 3V\FKLDWU\� �������
���±�����KWWSV���GRL�RUJ���������EMS����������
-DUPRORZLF]� $� ,�� &KHQ� +� <�� 3DQHJ\UHV� 3� .� ������� 7KH SDWWHUQV RI
LQKHULWDQFH LQ HDUO\�RQVHW GHPHQWLD� $O]KHLPHU¶V GLVHDVH DQG IURQWRWHPSRUDO
GHPHQWLD� $PHULFDQ -RXUQDO RI $O]KHLPHU¶V 'LVHDVH DQG 2WKHU 'HPHQWLDV� ������
���±�����KWWSV���GRL�RUJ�������������������������
.LP� '�� 3DJJL� -� 0�� 3DUN� &�� %HQQHWW� &�� 6DO]EHUJ� 6� /� ������� *UDSK�EDVHG
JHQRPH DOLJQPHQW DQG JHQRW\SLQJ ZLWK +,6$7� DQG +,6$7�JHQRW\SH� 1DWXUH
%LRWHFKQRORJ\� ����������±�����KWWSV���GRL�RUJ���������V����������������
���

.LYLRMD� 7�� 9lKlUDXWLR� $�� .DUOVVRQ� .�� %RQNH� 0�� (QJH� 0�� /LQQDUVVRQ� 6��
7DLSDOH� -� ������� &RXQWLQJ DEVROXWH QXPEHUV RI PROHFXOHV XVLQJ XQLTXH PROHFXODU
LGHQWLILHUV� 1DWXUH�0HWKRGV� ��������±����KWWSV���GRL�RUJ���������QPHWK�����
.XPDU� $�� 6LQJK� $�� (NDYDOL� ������� $ UHYLHZ RQ $O]KHLPHU¶V GLVHDVH
SDWKRSK\VLRORJ\ DQG LWV PDQDJHPHQW� $Q XSGDWH� 3KDUPDFRORJLFDO 5HSRUWV� ������
���±�����KWWSV���GRL�RUJ���������M�SKDUHS������������
.XPDU� 3�� 7DQ� <�� &DKDQ� 3� ������� 8QGHUVWDQGLQJ GHYHORSPHQW DQG VWHP FHOOV
XVLQJ VLQJOH FHOO�EDVHG DQDO\VHV RI JHQH H[SUHVVLRQ� ,Q 'HYHORSPHQW �&DPEULGJH�
�9RO� ���� ,VVXH �� SS� ��±���� &RPSDQ\ RI %LRORJLVWV /WG�
KWWSV���GRL�RUJ���������GHY�������
/DPEHUW� -��&�� ,EUDKLP�9HUEDDV� &� $�� +DUROG� '�� 1DM� $� &�� 6LPV� 5�� %HOOHQJXH]�
&�� -XQ� *�� 'H6WHIDQR� $� /�� %LV� -� &�� %HHFKDP� *� :�� *UHQLHU�%ROH\� %�� 5XVVR�
*�� 7KRUQWRQ�:HOOV� 7� $�� -RQHV� 1�� 6PLWK� $� 9� &KRXUDNL� 9�� 7KRPDV� &�� ,NUDP� 0�
$�� =HOHQLND� '�� « $PRX\HO� 3� ������� 0HWD�DQDO\VLV RI ������ LQGLYLGXDOV LGHQWLILHV
�� QHZ VXVFHSWLELOLW\ ORFL IRU $O]KHLPHU¶V GLVHDVH� 1DWXUH *HQHWLFV� �������
����±������KWWSV���GRL�RUJ���������QJ�����
/RYH� 0� ,�� 6RQHVRQ� &�� 3DWUR� 5� ������� 2SHQ 3HHU 5HYLHZ 6ZLPPLQJ
GRZQVWUHDP� VWDWLVWLFDO DQDO\VLV RI GLIIHUHQWLDO WUDQVFULSW XVDJH IROORZLQJ 6DOPRQ
TXDQWLILFDWLRQ >YHUVLRQ �� SHHU UHYLHZ� � DSSURYHG@�
KWWSV���GRL�RUJ����������I����UHVHDUFK��������
/RYH� 0� ,�� +XEHU� :�� $QGHUV� 6� ������� 0RGHUDWHG HVWLPDWLRQ RI IROG FKDQJH
DQG GLVSHUVLRQ IRU 51$�VHT GDWD ZLWK '(6HT�� *HQRPH %LRORJ\� ������� ����
KWWSV���GRL�RUJ���������V����������������
0DF/LQ� -� 0� $�� :DQJ� 7�� ;LDR� 6� ������� %LRPDUNHUV IRU WKH GLDJQRVLV RI
$O]KHLPHU¶V GLVHDVH� GHPHQWLD /HZ\ ERG\� IURQWRWHPSRUDO GHPHQWLD DQG YDVFXODU
GHPHQWLD� ,Q *HQHUDO 3V\FKLDWU\ �9RO� ��� ,VVXH �� S� ���� %0- 3XEOLVKLQJ *URXS�
KWWSV���GRL�RUJ���������JSV\FK������������
0DQQVSHUJHU� +� $�� *DGH� 6�� +HQMHV� )�� %HLVVEDUWK� 7�� .RUI� 8� �������
533DQDO\]HU� $QDO\VLV RI UHYHUVH�SKDVH SURWHLQ DUUD\ GDWD� %LRLQIRUPDWLFV� �������
����±������KWWSV���GRL�RUJ���������ELRLQIRUPDWLFV�EWT���
0LUUD� 6� 6� ������� 1HXURSDWKRORJLFDO DVVHVVPHQW RI $O]KHLPHU¶V GLVHDVH� 7KH
H[SHULHQFH RI WKH &RQVRUWLXP WR (VWDEOLVK D 5HJLVWU\ IRU $O]KHLPHU¶V 'LVHDVH�
,QWHUQDWLRQDO 3V\FKRJHULDWULFV� ��6833/� ��� ���±����
KWWSV���GRL�RUJ���������6����������������
���

0RFDQX� 0� 0�� 1LVVHQ� $�� (FNHUPDQQ� .�� .KOLVWXQRYD� ,�� %LHUQDW� -�� 'UH[OHU� '��
3HWURYD� 2�� 6FK|QLJ� .�� %XMDUG� +�� 0DQGHONRZ� (�� =KRX� /�� 5XQH� *��
0DQGHONRZ� (� 0� ������� 7KH SRWHQWLDO IRU ��VWUXFWXUH LQ WKH UHSHDW GRPDLQ RI WDX
SURWHLQ GHWHUPLQHV DJJUHJDWLRQ� V\QDSWLF GHFD\� QHXURQDO ORVV� DQG FRDVVHPEO\ ZLWK
HQGRJHQRXV 7DX LQ LQGXFLEOH PRXVH PRGHOV RI WDXRSDWK\� -RXUQDO RI 1HXURVFLHQFH�
����������±�����KWWSV���GRL�RUJ���������-1(8526&,�������������
0ROO� 3�� $QWH� 0�� 6HLW]� $�� 5HGD� 7� ������� 4XDQW6HT �� P51$ VHTXHQFLQJ IRU
51$�TXDQWLILFDWLRQ� 1DWXUH�0HWKRGV� ��������L±LLL� KWWSV���GRL�RUJ���������QPHWK�I����
0RO\QHDX[� %� -�� $UORWWD� 3�� 0HQH]HV� -� 5� /�� 0DFNOLV� -� '� ������� 1HXURQDO
VXEW\SH VSHFLILFDWLRQ LQ WKH FHUHEUDO FRUWH[� ,Q 1DWXUH 5HYLHZV 1HXURVFLHQFH �9RO� ��
,VVXH����SS�����±������1DW�5HY�1HXURVFL��KWWSV���GRL�RUJ���������QUQ����
0RUDELWR� 6�� 0L\RVKL� (�� 0LFKDHO� 1�� 6ZDUXS� 9� ������� ,QWHJUDWLYH JHQRPLFV
DSSURDFK LGHQWLILHV FRQVHUYHG WUDQVFULSWRPLF QHWZRUNV LQ $O]KHLPHU¶V GLVHDVH�
+XPDQ 0ROHFXODU *HQHWLFV� ������� ����±�����
KWWSV���GRL�RUJ���������KPJ�GGDD���
0RUODQ� -� '�� 4X� .�� 6LQLFURSL� '� 9� ������� 6HOHFWLYH GHSOHWLRQ RI U51$ HQDEOHV
ZKROH WUDQVFULSWRPH SURILOLQJ RI DUFKLYDO IL[HG WLVVXH� 3/R6 21(� ����� �±��
KWWSV���GRL�RUJ���������MRXUQDO�SRQH��������
0RUULV� -� &� ������� 7KH FOLQLFDO GHPHQWLD UDWLQJ �FGU�� &XUUHQW YHUVLRQ DQG VFRULQJ
UXOHV� 1HXURORJ\� ������������±������KWWSV���GRL�RUJ���������ZQO������������D
0RUWD]DYL� $�� :LOOLDPV� %� $�� 0F&XH� .�� 6FKDHIIHU� /�� :ROG� %� ������� 0DSSLQJ
DQG TXDQWLI\LQJ PDPPDOLDQ WUDQVFULSWRPHV E\ 51$�6HT� 1DWXUH 0HWKRGV� �����
���±�����KWWSV���GRL�RUJ���������QPHWK�����
1DJDODNVKPL� 8�� :DQJ� =�� :DHUQ� .�� 6KRX� &�� 5DKD� '�� *HUVWHLQ� 0�� 6Q\GHU�
0� ������� 7KH WUDQVFULSWLRQDO ODQGVFDSH RI WKH \HDVW JHQRPH GHILQHG E\ 51$
VHTXHQFLQJ� 6FLHQFH� ���������� ����±�����
KWWSV���GRL�RUJ���������VFLHQFH��������
2NRQLHZVNL� 0� -�� 0LOOHU� &� -� ������� +\EULGL]DWLRQ LQWHUDFWLRQV EHWZHHQ
SUREHVHWV LQ VKRUW ROLJR PLFURDUUD\V OHDG WR VSXULRXV FRUUHODWLRQV� %0&
%LRLQIRUPDWLFV� ��0P����±����KWWSV���GRL�RUJ������������������������
3HDUW� 0� -�� 6P\WK� *� .�� 9DQ /DDU� 5� .�� %RZWHOO� '� '�� 5LFKRQ� 9� 0�� 0DUNV� 3�
$�� +ROORZD\� $� -�� -RKQVWRQH� 5� :� ������� ,GHQWLILFDWLRQ DQG IXQFWLRQDO
VLJQLILFDQFH RI JHQHV UHJXODWHG E\ VWUXFWXUDOO\ GLIIHUHQW KLVWRQH GHDFHW\ODVH
���

LQKLELWRUV� 3URFHHGLQJV RI WKH 1DWLRQDO $FDGHP\ RI 6FLHQFHV RI WKH 8QLWHG 6WDWHV RI
$PHULFD� �������������±������KWWSV���GRL�RUJ���������SQDV�����������
3HUWHD� 0�� 3HUWHD� *� 0�� $QWRQHVFX� &� 0�� &KDQJ� 7� &�� 0HQGHOO� -� 7��
6DO]EHUJ� 6� /� ������� 6WULQJ7LH HQDEOHV LPSURYHG UHFRQVWUXFWLRQ RI D WUDQVFULSWRPH
IURP 51$�VHT UHDGV� 1DWXUH %LRWHFKQRORJ\� ������ ���±����
KWWSV���GRL�RUJ���������QEW�����
3LFHOOL� 6�� )DULGDQL� 2� 5�� %M|UNOXQG� c� .�� :LQEHUJ� *�� 6DJDVVHU� 6�� 6DQGEHUJ�
5� ������� )XOO�OHQJWK 51$�VHT IURP VLQJOH FHOOV XVLQJ 6PDUW�VHT�� 1DWXUH 3URWRFROV�
���������±�����KWWSV���GRL�RUJ���������QSURW���������
3RVSLFK� 6�� 5DXQVHU� 6� ������� 7KH PROHFXODU EDVLV RI $O]KHLPHU¶V SODTXHV� ,Q
6FLHQFH �9RO� ���� ,VVXH ����� SS� ��±���� $PHULFDQ $VVRFLDWLRQ IRU WKH
$GYDQFHPHQW�RI�6FLHQFH��KWWSV���GRL�RUJ���������VFLHQFH�DDS����
3ULQFH� 0DUWLQ� :LPR� $QGHUV� *XHUFKHW� 0DsOHQQ� $OL� *HPPD�&ODLUH� :X� <X�7]X�
3ULQD� 0� ������� :RUOG $O]KHLPHU 5HSRUW�
KWWSV���ZZZ�DO]�FR�XN�UHVHDUFK�:RUOG$O]KHLPHU5HSRUW�����SGI
4LX� ;�� 0DR� 4�� 7DQJ� <�� :DQJ� /�� &KDZOD� 5�� 3OLQHU� +� $�� 7UDSQHOO� &� �������
5HYHUVHG JUDSK HPEHGGLQJ UHVROYHV FRPSOH[ VLQJOH�FHOO WUDMHFWRULHV� 1DWXUH
0HWKRGV��KWWSV���GRL�RUJ���������QPHWK�����
5DPVN|OG� '�� /XR� 6�� :DQJ� <� &�� /L� 5�� 'HQJ� 4�� )DULGDQL� 2� 5�� 'DQLHOV� *� $��
.KUHEWXNRYD� ,�� /RULQJ� -� )�� /DXUHQW� /� &�� 6FKURWK� *� 3�� 6DQGEHUJ� 5� �������
)XOO�OHQJWK P51$�6HT IURP VLQJOH�FHOO OHYHOV RI 51$ DQG LQGLYLGXDO FLUFXODWLQJ WXPRU
FHOOV� 1DWXUH�%LRWHFKQRORJ\� ����������±�����KWWSV���GRL�RUJ���������QEW�����
5HJDQ� 3�� :KLWFRPE� '� -�� &KR� .� ������� 3K\VLRORJLFDO DQG SDWKRSK\VLRORJLFDO
LPSOLFDWLRQV RI V\QDSWLF WDX� ,Q 1HXURVFLHQWLVW �9RO� ��� ,VVXH �� SS� ���±����� 6$*(
3XEOLFDWLRQV�,QF��KWWSV���GRL�RUJ�������������������������
5LWFKLH� 0� (�� 3KLSVRQ� %�� :X� '�� +X� <�� /DZ� &� :�� 6KL� :�� 6P\WK� *� .�
������� /LPPD SRZHUV GLIIHUHQWLDO H[SUHVVLRQ DQDO\VHV IRU 51$�VHTXHQFLQJ DQG
PLFURDUUD\ VWXGLHV� 1XFOHLF $FLGV 5HVHDUFK� ������ H���
KWWSV���GRL�RUJ���������QDU�JNY���
5\PDQ� '� &�� $FRVWD�%DHQD� 1�� $LVHQ� 3� 6�� %LUG� 7�� 'DQHN� $�� )R[� 1� &�� *RDWH�
$�� )URPPHOW� 3�� *KHWWL� %�� /DQJEDXP� -� %� 6�� /RSHUD� )�� 0DUWLQV� 5�� 0DVWHUV� &�
/�� 0D\HX[� 5� 3�� 0F'DGH� (�� 0RUHQR� 6�� 5HLPDQ� (� 0�� 5LQJPDQ� -� 0��
6DOORZD\� 6�� « %DWHPDQ� 5� -� ������� 6\PSWRP RQVHW LQ DXWRVRPDO GRPLQDQW
���

$O]KHLPHU GLVHDVH� $ V\VWHPDWLF UHYLHZ DQG PHWD�DQDO\VLV� 1HXURORJ\� ������
���±�����KWWSV���GRL�RUJ���������:1/�����������������
6KLPRMR� +�� 2KWVXND� 7�� .DJH\DPD� 5� ������� 2VFLOODWLRQV LQ 1RWFK 6LJQDOLQJ
5HJXODWH 0DLQWHQDQFH RI 1HXUDO 3URJHQLWRUV� 1HXURQ� ������ ��±���
KWWSV���GRL�RUJ���������M�QHXURQ������������
6PLWK� 7�� +HJHU� $�� 6XGEHU\� ,� ������� 80,�WRROV� 0RGHOLQJ VHTXHQFLQJ HUURUV LQ
8QLTXH 0ROHFXODU ,GHQWLILHUV WR LPSURYH TXDQWLILFDWLRQ DFFXUDF\� *HQRPH 5HVHDUFK�
����������±�����KWWSV���GRL�RUJ���������JU�����������
6WDUN� 5�� *U]HODN� 0�� +DGILHOG� -� ������� 51$ VHTXHQFLQJ� WKH WHHQDJH \HDUV�
1DWXUH 5HYLHZV *HQHWLFV� ������� ���±����
KWWSV���GRL�RUJ���������V����������������
6XOWDQ� 0�� 6FKXO]� 0� +�� 5LFKDUG� +�� 0DJHQ� $�� .OLQJHQKRII� $�� 6FKHUI� 0�� 6HLIHUW�
0�� %RURGLQD� 7�� 6ROGDWRY� $�� 3DUNKRPFKXN� '�� 6FKPLGW� '�� 2¶.HHIIH� 6�� +DDV� 6��
9LQJURQ� 0�� /HKUDFK� +�� <DVSR� 0� /� ������� $ JOREDO YLHZ RI JHQH DFWLYLW\ DQG
DOWHUQDWLYH VSOLFLQJ E\ GHHS VHTXHQFLQJ RI WKH KXPDQ WUDQVFULSWRPH� 6FLHQFH�
��������������±�����KWWSV���GRL�RUJ���������VFLHQFH��������
6YHQVVRQ� 9�� 9HQWR�7RUPR� 5�� 7HLFKPDQQ� 6� $� ������� ([SRQHQWLDO VFDOLQJ RI
VLQJOH�FHOO 51$�VHT LQ WKH ODVW GHFDGH� 1DWXUH 3URWRFROV� ������ ���±����
KWWS���DU[LY�RUJ�DEV�����������
7DQJ� )�� %DUEDFLRUX� &�� :DQJ� <�� 1RUGPDQ� (�� /HH� &�� ;X� 1�� :DQJ� ;�� %RGHDX�
-�� 7XFK� %� %�� 6LGGLTXL� $�� /DR� .�� 6XUDQL� 0� $� ������� P51$�6HT
ZKROH�WUDQVFULSWRPH DQDO\VLV RI D VLQJOH FHOO� 1DWXUH 0HWKRGV� ����� ���±����
KWWSV���GRL�RUJ���������QPHWK�����
7LDQ� %�� 0DQOH\� -� /� ������� $OWHUQDWLYH SRO\DGHQ\ODWLRQ RI P51$ SUHFXUVRUV� ,Q
1DWXUH 5HYLHZV 0ROHFXODU &HOO %LRORJ\ �9RO� ��� ,VVXH �� SS� ��±���� 1DWXUH
3XEOLVKLQJ�*URXS��KWWSV���GRL�RUJ���������QUP���������
7UDSQHOO� &�� :LOOLDPV� %� $�� 3HUWHD� *�� 0RUWD]DYL� $�� .ZDQ� *�� 9DQ %DUHQ� 0� -��
6DO]EHUJ� 6� /�� :ROG� %� -�� 3DFKWHU� /� ������� 7UDQVFULSW DVVHPEO\ DQG
TXDQWLILFDWLRQ E\ 51$�6HT UHYHDOV XQDQQRWDWHG WUDQVFULSWV DQG LVRIRUP VZLWFKLQJ
GXULQJ FHOO GLIIHUHQWLDWLRQ� 1DWXUH %LRWHFKQRORJ\� ������ ���±����
KWWSV���GRL�RUJ���������QEW�����
9HUJKHVH� 3� %�� &DVWHOODQR� -� 0�� +ROW]PDQ� '� 0� ������� $SROLSRSURWHLQ ( LQ
$O]KHLPHU¶V GLVHDVH DQG RWKHU QHXURORJLFDO GLVRUGHUV� ,Q 7KH /DQFHW 1HXURORJ\ �9RO�
���

��� ,VVXH �� SS� ���±����� /DQFHW 1HXURO�
KWWSV���GRL�RUJ���������6��������������������
9LWWLQJ�6HHUXS� .�� 6DQGHOLQ� $� ������� ,VRIRUP6ZLWFK$QDO\]H5� DQDO\VLV RI
FKDQJHV LQ JHQRPH�ZLGH SDWWHUQV RI DOWHUQDWLYH VSOLFLQJ DQG LWV IXQFWLRQDO
FRQVHTXHQFHV� %LRLQIRUPDWLFV �2[IRUG� (QJODQG�� ������� ����±�����
KWWSV���GRL�RUJ���������ELRLQIRUPDWLFV�EW]���
9LWWLQJ�6HHUXS� .�� 6DQGHOLQ� $� ������� 7KH ODQGVFDSH RI LVRIRUP VZLWFKHV LQ
KXPDQ FDQFHUV� 0ROHFXODU &DQFHU 5HVHDUFK� ������ ����±�����
KWWSV���GRL�RUJ�������������������0&5��������
:DQJ� 0�� %HFNPDQQ� 1� '�� 5RXVVRV� 3�� :DQJ� (�� =KRX� ;�� :DQJ� 4�� 0LQJ� &��
1HII� 5�� 0D� :�� )XOODUG� -� )�� +DXEHUJ� 0� (�� %HQGO� -�� 3HWHUV� 0� $�� /RJVGRQ� %��
:DQJ� 3�� 0DKDMDQ� 0�� 0DQJUDYLWH� /� 0�� 'DPPHU� (� %�� 'XRQJ� '� 0�� « =KDQJ�
%� ������� 7KH 0RXQW 6LQDL FRKRUW RI ODUJH�VFDOH JHQRPLF� WUDQVFULSWRPLF DQG
SURWHRPLF GDWD LQ $O]KHLPHU¶V GLVHDVH� 6FLHQWLILF 'DWD� ��
KWWSV���GRL�RUJ���������VGDWD���������
:DQJ� ;�� 3DUN� -�� 6XV]WDN� .�� =KDQJ� 1� 5�� /L� 0� ������� %XON WLVVXH FHOO W\SH
GHFRQYROXWLRQ ZLWK PXOWL�VXEMHFW VLQJOH�FHOO H[SUHVVLRQ UHIHUHQFH� 1DWXUH
&RPPXQLFDWLRQV� ��������±���KWWSV���GRL�RUJ���������V����������������[
:DQJ� =�� *HUVWHLQ� 0�� 6Q\GHU� 0� ������� 51$�6HT� $ UHYROXWLRQDU\ WRRO IRU
WUDQVFULSWRPLFV� ,Q 1DWXUH 5HYLHZV *HQHWLFV �9RO� ��� ,VVXH �� SS� ��±����
KWWSV���GRL�RUJ���������QUJ����
<DPDGD� .�� /LP� -�� 'DOH� -� +�� &KHQ� +�� 6KLQQ� 3�� 3DOP� &� -�� 6RXWKZLFN� $� 0��
:X� +� &�� .LP� &�� 1JX\HQ� 0�� 3KDP� 3�� &KHXN� 5�� .DUOLQ�1HZPDQQ� *�� /LX� 6�
;�� /DP� %�� 6DNDQR� +�� :X� 7�� <X� *�� 0LUDQGD� 0�� « (FNHU� -� 5� �������
(PSLULFDO $QDO\VLV RI 7UDQVFULSWLRQDO $FWLYLW\ LQ WKH $UDELGRSVLV *HQRPH� 6FLHQFH�
��������������±�����KWWSV���GRL�RUJ���������VFLHQFH��������
<L� /�� 3LPHQWHO� +�� %UD\� 1� /�� 3DFKWHU� /� ������� *HQH�OHYHO GLIIHUHQWLDO DQDO\VLV
DW WUDQVFULSW�OHYHO UHVROXWLRQ� *HQRPH %LRORJ\� ������ ���
KWWSV���GRL�RUJ���������V���������������]
=KDQJ� <�� &KHQ� .�� 6ORDQ� 6� $�� %HQQHWW� 0� /�� 6FKRO]H� $� 5�� 2¶.HHIIH� 6��
3KDWQDQL� +� 3�� *XDUQLHUL� 3�� &DQHGD� &�� 5XGHULVFK� 1�� 'HQJ� 6�� /LGGHORZ� 6� $��
=KDQJ� &�� 'DQHPDQ� 5�� 0DQLDWLV� 7�� %DUUHV� %� $�� :X� -� 4� ������� $Q
51$�VHTXHQFLQJ WUDQVFULSWRPH DQG VSOLFLQJ GDWDEDVH RI JOLD� QHXURQV� DQG YDVFXODU
���

FHOOV RI WKH FHUHEUDO FRUWH[� -RXUQDO RI 1HXURVFLHQFH� ������� �����±������
KWWSV���GRL�RUJ���������-1(8526&,�������������
=KDQJ� <�� 6ORDQ� 6� $�� &ODUNH� /� (�� &DQHGD� &�� 3OD]D� &� $�� %OXPHQWKDO� 3� '��
9RJHO� +�� 6WHLQEHUJ� *� .�� (GZDUGV� 0� 6� %�� /L� *�� 'XQFDQ� -� $�� &KHVKLHU� 6� +��
6KXHU� /� 0�� &KDQJ� (� )�� *UDQW� *� $�� *HSKDUW� 0� *� +�� %DUUHV� %� $� �������
3XULILFDWLRQ DQG &KDUDFWHUL]DWLRQ RI 3URJHQLWRU DQG 0DWXUH +XPDQ $VWURF\WHV
5HYHDOV 7UDQVFULSWLRQDO DQG )XQFWLRQDO 'LIIHUHQFHV ZLWK 0RXVH� 1HXURQ� ������
��±����KWWSV���GRL�RUJ���������M�QHXURQ������������
=KDR� 6�� =KDQJ� <�� *DPLQL� 5�� =KDQJ� %�� 9RQ 6FKDFN� '� ������� (YDOXDWLRQ RI
WZR PDLQ 51$�VHT DSSURDFKHV IRU JHQH TXDQWLILFDWLRQ LQ FOLQLFDO 51$ VHTXHQFLQJ�
3RO\$� VHOHFWLRQ YHUVXV U51$ GHSOHWLRQ� 6FLHQWLILF 5HSRUWV� ����� �����
KWWSV���GRL�RUJ���������V�����������������
���

$3Ç1',&(
���

$OYHV HW DO� ����� &RPR FRODERUDGRU� DMXGHL QDV DQiOLVHV GH VLQJOH�FHOO 57�T3&5SDUD LGHQWLILFDU DV GLIHUHQoDV QD H[SUHVVmR JrQLFD GDV FpOXODV UHSURJUDPDGDV� YLGH
ILJXUD���
*XLPDUmHV HW DO� ����� &RPR FRODERUDGRU� IL] D PLQHUDomR GH GDGRV Mi SXEOLFDGRVGH VF51$VHT GH UHWLQD GH FDPXQGRQJR� IRUQHFHQGR GDGRV GH H[SUHVVmR JrQLFD
XVDGRV�QR�WUDEDOKR��YLGH�ILJXUD���
0DUTXHV�&RHOKR HW DO� ����� &RPR DXWRU� IRUDP XVDGRV GDGRV GH VF51$VHT GH
FpOXODV UHSURJUDPDGDV PDLV GDGRV GH EXON 51$VHT SDUD DYDOLDU RV PHFDQLVPRV
PROHFXODUHV DVVRFLDGRV DR SURFHVVR ILVLROyJLFR GH GLIHUHQFLDomR GH FpOXODV�WURQFR
QHXUDLV HP QHXU{QLRV FRP DTXHOHV SUHVHQWHV GXUDQWH D UHSURJUDPDomR GH OLQKDJHP
GH�ILEUREODVWRV�HP�QHXU{QLRV�
.LOLQF HW DO� ����� &RPR FRODERUDGRU� IRUQHFL RV GDGRV GH H[SUHVVmR GR JHQH37.�% HP GLIHUHQWHV GDWDVHWV GH $O]KHLPHU �0$<2� 5260$3 H 06%%�� YLGH
WDEHOD�VXSOHPHQWDU�
���

ORIGINAL RESEARCHpublished: 08 June 2018
doi: 10.3389/fncel.2018.00155
Frontiers in Cellular Neuroscience | www.frontiersin.org 1 June 2018 | Volume 12 | Article 155
Edited by:
Christophe Heinrich,
INSERM U1208 Institut Cellule
Souche et Cerveau, France
Reviewed by:
Zhiping P. Pang,
Rutgers University, The State
University of New Jersey,
United States
Carol Schuurmans,
Sunnybrook Health Science Centre,
Canada
*Correspondence:
Marcos R. Costa
†Present Address:
Markus M. Hilscher,
Institute for Analysis and Scientific
Computing, Vienna University of
Technology, Vienna, Austria
Received: 23 April 2018
Accepted: 17 May 2018
Published: 08 June 2018
Citation:
Araújo JAM, Hilscher MM,
Marques-Coelho D, Golbert DCF,
Cornelio DA, Batistuzzo de Medeiros
SR, Leão RN and Costa MR (2018)
Direct Reprogramming of Adult
Human Somatic Stem Cells Into
Functional Neurons Using Sox2,
Ascl1, and Neurog2.
Front. Cell. Neurosci. 12:155.
doi: 10.3389/fncel.2018.00155
Direct Reprogramming of AdultHuman Somatic Stem Cells IntoFunctional Neurons Using Sox2,Ascl1, and Neurog2
Jessica Alves de Medeiros Araújo 1, Markus M. Hilscher 1†, Diego Marques-Coelho 1,2,
Daiane C. F. Golbert 1, Deborah A. Cornelio 3, Silvia R. Batistuzzo de Medeiros 3,
Richardson N. Leão 1 and Marcos R. Costa 1*
1 Brain Institute, Federal University of Rio Grande do Norte, Natal, Brazil, 2 Bioinformatics Multidisciplinary Environment, IMD,
Federal University of Rio Grande do Norte, Natal, Brazil, 3 Laboratório de Biologia Molecular e Genômica, Centro de
Biociências, Federal University of Rio Grande do Norte, Natal, Brazil
Reprogramming of somatic cells into induced pluripotent stem cells (iPS) or directly
into cells from a different lineage, including neurons, has revolutionized research in
regenerative medicine in recent years. Mesenchymal stem cells are good candidates for
lineage reprogramming and autologous transplantation, since they can be easily isolated
from accessible sources in adult humans, such as bone marrow and dental tissues. Here,
we demonstrate that expression of the transcription factors (TFs) SRY (sex determining
region Y)-box 2 (Sox2), Mammalian achaete-scute homolog 1 (Ascl1), or Neurogenin
2 (Neurog2) is sufficient for reprogramming human umbilical cord mesenchymal stem
cells (hUCMSC) into induced neurons (iNs). Furthermore, the combination of Sox2/Ascl1
or Sox2/Neurog2 is sufficient to reprogram up to 50% of transfected hUCMSCs into
iNs showing electrical properties of mature neurons and establishing synaptic contacts
with co-culture primary neurons. Finally, we show evidence supporting the notion that
different combinations of TFs (Sox2/Ascl1 and Sox2/Neurog2) may induce multiple and
overlapping neuronal phenotypes in lineage-reprogrammed iNs, suggesting that neuronal
fate is determined by a combination of signals involving the TFs used for reprogramming
but also the internal state of the converted cell. Altogether, the data presented here
contribute to the advancement of techniques aiming at obtaining specific neuronal
phenotypes from lineage-converted human somatic cells to treat neurological disorders.
Keywords: induced neurons, lineage reprogramming, human mesenchymal stem cells, umbilical cord, proneural
genes
INTRODUCTION
Reprogramming of somatic cells into induced pluripotent stem cells (iPS) that can generateall three major embryonic lineages, stem cells and even a new animal has revolutionizedresearch in regenerative medicine in recent years (Takahashi and Yamanaka, 2006; Okitaet al., 2007). Somatic cells isolated from different sources can be converted into iPS(Meissner et al., 2007; Aoi et al., 2008; Hanna et al., 2008; Espejel et al., 2010; Imamuraet al., 2010), which in turn can be converted into specific cell types including neurons(Wernig et al., 2008; Kuzmenkin et al., 2009; Mizuno et al., 2010; Zhang et al., 2013).

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
However, generation of iPS and further differentiation intoneuronal cells is time consuming and the cells retain tumorigenicpotential (Takahashi and Yamanaka, 2006; Okita et al., 2007).
In contrast, direct lineage reprogramming of somatic cells is afast process and bypasses the pluripotent stage associated withtumor transformation. Astrocytes isolated from the postnatalcerebral cortex of mice were the first cells to be directlyreprogrammed into induced neurons (iNs) following expressionof the transcription factor (TF) Neurogenin 2 (Neurog2) orMammalian achaete-scute homolog 1 (Mash1/Ascl1) (Berningeret al., 2007; Heinrich et al., 2011). Subsequently, the list ofmurine cell types liable to lineage reprogramming into iNsgrew substantially, including non-neural cells, such as mousefibroblasts and hepatocytes (Vierbuchen et al., 2010; Marro et al.,2011). However, non-neural cells typically require more than oneTF to achieve a full neuronal conversion.
Direct reprogramming of human somatic cells into neuronscan also be achieved through expression of Ascl1 in combinationwith other TFs (Ambasudhan et al., 2011; Son et al., 2011;Karow et al., 2012). It has also been reported that expressionof Ascl1 or Neurog2 alone is sufficient to induce conversion ofhuman fibroblasts into induced neurons (Chanda et al., 2014;Gascón et al., 2016), but the efficiency of this process is low(<10%). Moreover, the phenotypes of iNs obtained throughdirect cell lineage reprogramming using human cells remainslargely elusive. Pinpointing strategies capable of producing iNsexhibiting defined neurochemical phenotypes is a critical steptowards translation of the lineage reprogramming techniquesinto clinics.
Here, we show that the expression of the transcriptionfactor SRY (sex determining region Y)-box 2 (Sox2), Ascl1 orNeurog2 is sufficient to lineage-convert a small fraction of humanumbilical cord mesenchymal stem cells (hUCMSCs) into iNs. Incontrast, the co-expression of either Sox2/Ascl1 or Sox2/Neurog2is sufficient to convert a large fraction of hUCMSCs (upto 50%) into iNs displaying electrophysiological hallmarks ofmature neurons and establishing synaptic contacts with othercells. Furthermore, we show that iNs may express transcriptsassociated with the acquisition of different neurochemicalphenotypes, independently of the combination of transcriptionfactors used. Also, Sox2/Ascl1 and Sox2/Neurog2 may inducethe expression of genes involved in the acquisition of thesame neurochemical phenotypes, suggesting that iNs fateduring lineage-conversion is influenced by other aspectsthan the transcription factors used. Collectively, our dataindicate that hUCMSCs are good candidates for lineagereprogramming into iNs, but more studies are required to furtheradvance protocols capable of producing iNs with a particularphenotype.
MATERIALS AND METHODS
Cell CultureHuman multipotent mesenchymal stem cells (hMSC) wereisolated from umbilical cords donated with informed consentof the pregnant mothers at maternity Januário Cicco, FederalUniversity of Rio Grande do Norte, Natal, Brazil. The study
was approved by the Research Ethics Committee of the FederalUniversity of Rio Grande do Norte (Project Number 508.459),and in strict agreement with Brazilian law (Resolution 196/96).All subjects gave written informed consent in accordance withthe Declaration of Helsinki.
In this study, Wharton’s jelly mesenchymal stem cells wereisolated from umbilical cord. Following isolation from thesubendothelium vein, according to the method previouslypublished (Duarte et al., 2012), the remaining umbilical cordtissue was cut in small pieces and washed with phosphate-buffered saline (PBS; 137mM NaCl, 2.7mM KCl, 4.3mMNa2HPO4, and 1.47mM KH2PO4; Merck), supplemented with3% antibiotic–antimycotic solution (prepared with 10,000units/ml penicillin G sodium, 10,000µg/ml streptomycin sulfateand 25µg/ml amphotericin B; HyClone). Then, the tissue wascentrifuged at 200 g for 10min, and the pellet resuspended in10mL of 0.1% collagenase type IV (Worthington) diluted inPBS. After that, the explants were incubated for 16 h at 37◦Cin a water bath. The tissue was centrifuged again at 200 gfor 10min, the pellet washed twice with PBS and then gentlydissociated in a digestion solution containing 0.25% trypsin and0.02% EDTA (Invitrogen) for 15min at room temperature. Tointerrupt trypsin activity, we added fetal bovine serum (FBS;HyClone). Once again, the cell suspension was centrifuged,and the cell pellet resuspended in minimum essential mediuma (α MEM; Gibco Invitrogen) supplemented with 10% FBSand 1% antibiotic solution. Cells were plated onto T25 tissueculture flasks (TPP) and these cultures maintained at 37◦C ina humidified atmosphere containing 5% CO2. After 2 or 4days, the medium was changed and non-adherent cells wereremoved. Cultures consisting of small, adherent and spindleshaped fibroblastoid cells reaching 60–70% of confluence weredetached and subcultured at 4,000 cells/cm2.
Characterization of hMSCsThe cells isolated from Wharton’s jelly human umbilical cordwere characterized as MSCs, according to the criteria proposedby the International Society for Cellular Therapy (Horwitz et al.,2005; Dominici et al., 2006). The hMSCs were labeled witha panel of monoclonal antibodies against several cell markers,including CD105-FITC, CD90PE-Cy5 (Bioscience), CD73PE,CD34PE, HLA-DR-FITC, CD45-FITC, and CD14PE (BectonDickinson’s). Briefly, the cells were detached of the tissue cultureplates using 0.25% trypsin/EDTA, washed, and homogenizedwith PBS. They were then incubated with monoclonal antibodyfor 30min in darkness at room temperature. At the end ofthis period, the cell suspension was centrifuged, washed in PBS,and re-suspended in cold fixing solution, 0.5% formaldehydein PBS. For each test, isotype-matched monoclonal antibodieswere used as negative controls (IgG1-FITC, PE, and PE-Cy5;Becton Dickinson’s). The fluorescence intensity of labeled cellswas determined with a fluorescence-activated cell analyzer(FACScan) using cell quest software (Cell QuestTM Software,Becton Dickinson Immunocytometry Systems), a total of20,000 events for each sample were recorded. The followingparameters were considered: forward scatter in linear scale(which evaluates cell size), side scatter in linear scale (assessing
Frontiers in Cellular Neuroscience | www.frontiersin.org 2 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
cell complexity), and cell marker expression in fluorescenceanalysis by FL1, FL2, and FL3 in logarithmical scale, representingthe antigen–antibody reaction conjugated to FITC, PE, and PE-Cy5, respectively. Results were expressed as a percentage of cellslabeled with monoclonal antibodies. Osteogenic, adipogenic, andchondrogenic differentiation assays were carried out according tomethodology previously published (Duarte et al., 2012).
PlasmidsThe pro-neural genes Ascl1, Neurog2, or Sox2 were expressedunder control of an internal chicken β-actin promoter withcytomegalovirus enhancer (pCAG) together with DsRed or GFPbehind an internal ribosomal entry site (pCAG-Ascl1-IRES-DsRed, pCAG-Neurog2-IRES-DsRed, and pCAG-Sox2-IRES-GFP). For control experiments, cultures were transfected withplasmids encoding only DsRed or GFP (pCAG-IRES-DsRed orpCAG-IRES-GFP) (Heinrich et al., 2010; Karow et al., 2012).Plasmid stocks were prepared in Escherichia coli and purifiedusing the endotoxin-free Maxiprep plasmid kit (Invitrogen).DNA concentration was adjusted to 1 µg/µL in TE bufferendotoxin free, and plasmids were stored at−20◦C.
TransfectionFor transfections, hMSC were seeded in 24-well plates ontopoly-D-lysine (Sigma-Aldrich) and laminin (L-2020; SigmaAldrich) coated glass coverslips at a density of 3 × 104
cells per well in 0.5mL α MEM (Gibco) supplemented with10% fetal bovine serum (FBS) and 1% antibiotic solution(penicillin/streptomycin). The cells were grown in theseconditions for 1–3 days until 70–80% confluent.
Both DNA plasmids (1 µg/µL) and a lipophilic cationicreagent (Lipofectamine 2000, Invitrogen) were diluted in 50µL Opti-MEM (Reduced Serum Medium, Invitrogen). Mixtureswere incubated for 5min and then combined for a further 20minaccording to the manufacturer’s instructions. Complexes wereadded to the cells in a total volume of 0.5mL Opti-MEM (Gibco)and incubated at 37◦C in a humidified atmosphere containing5% CO2 for 10–12 h. Antibiotics and serum were not used duringtransfection procedures.
Co-culture With Hippocampal NeuronsFor co-culture experiments, mouse hippocampus at postnatalday 0 to 4 (P0-4) were dissected in ice-cold PBS and dissociatedin a the digestion solution for 10min at 37◦C. Trypsinaction was interrupted with fetal bovine serum and the tissuedissociated mechanically with a fire-polished glass Pasteurpipette. The suspension was then centrifuged at 200 g for 5minand washed twice in DMEM/F12 10% FBS in DMEM/F12medium (Gibco). Mouse hippocampal cells were added to thehuman cultures 1–2 days after transfection at a density of50,000 cells per well. The local University Animal Care andUse Committee (CEUA/UFRN) approved experiments involvingmice. All experiments were carried out in accordance withinternational guidelines and regulations for animal use.
ImmunocytochemistryCell cultures were fixed in 4% paraformaldehyde (PFA) inPBS for 15min at room temperature. Primary antibodies werediluted in PBS, 0.5% Triton X-100 and 5% normal goatserum. Specimens were incubated overnight at 4◦C. After threewashes with PBS, cells were incubated with species-specificsecondary antibodies conjugated to fluorophores for 2 h atroom temperature. Once again, samples were washed with PBSthree times. For nuclei staining, cells were incubated for 5minwith 0.1µg/mL DAPI (4′6′-diamino-2-phenylindone) in PBS0.1M. Coverslips were finally mounted onto a glass slide witha mounting medium (Aqua Poly/Mount; Polysciences). Thefollowing primary antibodies and dilutions were used: chickenanti-Green Fluorescent Protein (GFP, Aves Labs, 1:1,000), rabbitanti-Red Fluorescent Protein (RFP, Rockland, 1:1,000), mouseanti-majormicrotubule associated protein (MAP2; Sigma, 1:500),guinea pig polyclonal anti-vesicular GABA transporter (vGAT,Synaptic Systems, 1:200), and polyclonal anti-vesicular glutamatetransporter 1 (vGLUT11, Synaptic Systems, 1:1,000).
ElectrophysiologyCell cultures with induced neurons were transferred to arecording chamber mounted on the stage of a microscopeequipped with a water immersion 40X objective (Zeiss Examiner.A1, 1 NA) and perfused with oxygenated external solution (1–1.25 ml/min) at 37◦C. Data were acquired using a patch-clampamplifier Axopatch 200B (Molecular Devices) in current orvoltage clamp mode, a 16-bit data acquisition card (NationalInstruments), and WinWCP or WinEDR software implementedby Dr. John Dempster (University of Strathclyde). Patch-pipettesof borosilicate glass capillaries (GC150F-10 Harvard Apparatus)were pulled on a vertical puller (Narishige) with resistances from5–7 M�. Pipettes were filled with internal solution (∼290 Osm)containing (in mM) 130 K+-gluconate, 7 NaCl, 0.1 EGTA, 0.3MgCl2, 0.8 CaCl2, 2 Mg-ATP, 0.5 NaGTP, 10 HEPES, and 2 EGTA(pH 7.2 adjusted with KOH 1M). The external solution (∼300Osm) contained (in mM) 120 NaCl, 3 KCl, 1.2 MgCl2, 2.5 CaCl2,23 NaHCO3, 5 HEPES, and 11 Glucose (pH 7.4 adjusted withNaOH 1M).
Patch-clamped cells were measured for input resistance,resting membrane potential, and capacitance. Recordings wereanalyzed with custom routines in MATLAB. Action potentialswere triggered by 400-ms depolarizing current injections from100 pA, 400ms, with 10 pA increments. The first fired actionpotential in response to minimal current injection was analyzedfor amplitude (peak to afterhyperpolarization voltage), half-width (halfway between threshold voltage and peak), andafterhyperpolarization amplitude (threshold to minimum ofvoltage trough between the first and the second actionpotential in a spike train). Instantaneous and steady-statevoltage were analyzed in response to hyperpolarizing currentinjections (−100 pA, 400ms). Excitatory postsynaptic currentswere analyzed for amplitude and rise time in free-run tracesof 150 s. Active and passive electrophysiological membraneproperties, including action potential parameters were analyzedusing a Student’s unpaired, two-tailed t-test.
Frontiers in Cellular Neuroscience | www.frontiersin.org 3 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
Calcium ImagingCalcium imaging was performed on human MSC 3 weekspost-transfection using Oregon green 488 BAPTA-1 (Invitrogen,10µM). Imaging was performed in physiological saline solutioncontaining (in mM) 140 NaCl, 5 KCl, 2 MgCl2, 2 CaCl2, 10HEPES, 10 glucose, and 6 sucrose (pH 7.35). Images wereacquired approximately every 10ms using a scientific CMOScamera (Andor). The microscope was controlled by Micro-Manager software together with the image processor ImageJ.Changes in fluorescence were measured for individual cells andaverage of the first 10 time-lapse images for each region of interest(ROI) was defined as initial fluorescence (F0).
Single Cell RT-qPCRAfter electrophysiological recordings, the cell was suckedinto the recording pipette. Pipettes were quickly removedand broken into 1.5mL tubes containing 20U of RNaseinhibitor and 8.3mM DTT. Samples were frozen immediatelyon dry ice and stored at −80◦C. Immediately after thaw,the samples were treated to eliminate contaminating DNAmolecules. Complementary DNA (cDNA) synthesis and pre-amplified reactions were performed with the RT2 PreAMPcDNA Synthesis Kit following the manufacturer’s procedure(QIAGEN). Amplification was performed on the AppliedBiosystems ViiA 7 Real-Time PCR (Applied Biosystems).RT2 Profiler PCR Array were customized in 96-well plates,designed for analyzing the expression of the following genes:Choline O-acetyltransferase (CHAT), Tyrosine hydroxylase(TH), Tryptophan hydroxylase 2 (TPH2), Vesicular glutamatetransporter 1 (VGLUT1 or SLC17A7), GABA Vesiculartransporter (VGAT or SLC32A1), FEZ family zinc finger 2(FEZF2), T-box brain 1 (TBR1), SATB homeobox 2 (SATB2),COUP-TF-interacting protein 2 (CTIP2 or BCL11B), Platelet-derived growth factor receptor, beta polypeptide (PDGFRB),Thy-1 cell surface antigen (THY-1), Atonal homolog 8 (ATOH8),Neurogenic differentiation 1 (NEUROD1), Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and Hypoxanthinephosphoribosyltransferase 1 (HPRT1). The RT-qPCR wasperformed using the RT2 profiler PCR customized array(QIAGEN). Each array included genomic DNA control primerset, reverse transcription control, positive PCR control to reportthe efficiency of the polymerase chain reaction itself, and theendogenous reference genes GAPDH and HPRT1.
Analysis of Single-Cell RT-qPCRA single-cell RT-qPCR pre-processing was performed based onmethod described by Ståhlberg et al. (2013). Melting curveanalysis performed elimination of false positives. Next, relativequantities were calculated using a cycle of quantification cutoff(Cq-cutoff) and relative-quantities of cDNA molecule equation.Missing data were imputed with absolute value 0.5, followedby conversion to log2-scale. Mean center and auto scale, foreach gene mean center and auto scale were calculated separatelyusing log2-values. Heat map and Principal Component Analysis(PCA) were used to visualize expression differences betweengroups. Statistical analysis and plotting were performed using thesoftware R version 3.3.3.
Statistical AnalysisAll statistical data are presented as the mean ± standard errorof the mean (SEM) of at least three independent experiments.Statistically significant differences were assessed by Student’sunpaired t-test or one-way Analysis of variance (ANOVA),comparing two or more groups, respectively. P < 0.05 wasconsidered a significant difference (∗).
RESULTS
Direct Lineage Reprogramming of HumanUmbilical Cord MSCsMesenchymal stem cells (MSCs) can be isolated from differentsources in adult humans, including the bone marrow andumbilical cord (Ding et al., 2011). These cells are highlyplastic, retaining the potential to generate chondroblasts,adipocytes, and osteoblasts (Caplan, 1991; Dominici et al.,2006; Afanasyev et al., 2010; Keating, 2012). In order tocharacterize the cells isolated from Wharton’s jelly umbilicalcord, we first evaluated the expression of MSC-specificantigens using flow cytometry. Virtually all cells exhibitedexpression of CD105, CD73, and CD90 markers, and lackedthe expression of hematopoietic lineage markers, such asCD14, CD34, and CD45 (Supplementary Figures 1A–I). TheMSCs also demonstrated capacity for osteogenic, adipogenic,and chondrogenic differentiation (Supplementary Figures 1J–L). Given this versatility, we hypothesized that expressionof neurogenic transcription factors in MSCs could directlyreprogram these cells into neurons. To test this possibility,we transfected plasmids carrying the genes encoding for Sox2,Neurog2, or Ascl1 into human umbilical cord mesenchymal stemcells (hUCMSCs) using lipophilic cationic reagent. To monitortransduced cells, all vectors carried a fluorescent protein (GFPor DsRed) under control of an internal chicken β-actin promoterwith cytomegalovirus enhancer (pCAG). Vectors expressing GFPor DsRed alone were used as control (Figures 1A,B). One dayafter transfection, cultured medium of hUCMSCs was replacedwith neuronal differentiation medium containing B27. In thismedium, most transfected hUCMSCs underwent cell deathprecluding analysis of lineage reprogramming (SupplementaryFigure 2). To overcome this limitation, we co-cultured neonatalmouse hippocampal cells with hUCMSCs. We found an averageof about 50% GFP+/DsRed+ hUCMSC per field 15 daysafter transfection and in the presence of co-cultured neonatalmouse hippocampal cells. In contrast, the average number ofGFP+/DsRed+ hUCMSCs in the absence of co-cultured cellswas<4% (Supplementary Figure 2). Low number of cells was alsoobserved in Hucmsc cultures transfected with control plasmids,indicating that the cell death under these culture conditionswas independent of lineage-reprogramming. It is likely thatthe withdrawal of serum performed after transfection (aimingat the differentiation of induced neurons) affects the survivalof hUCMSCs, whereas addition of co-cultured cells, somehow,counteracts this cell-death effect. We, therefore, concluded thatco-cultures are necessary to support hUCMSC in the cultureconditions used.
Frontiers in Cellular Neuroscience | www.frontiersin.org 4 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 1 | Direct lineage reprogramming of hUCMSC into iN by forced expression of Sox2, Ascl1, or Neurog2 alone, Sox2/Ascl1 or Sox2/Neurog2. (A–U)
Immunostaining for DSRED (red), GFP (green), MAP2 (white), and DAPI (blue), 15 days post transfection (dpt). Scale bar represents 20µm. (A–D) Example of
hUCMSC transfected with control plasmids encoding only reporter proteins GFP and DSRED. Note that cell displayed classical mesenchymal cell morphologies and
did not express MAP2. (E–H) Example of hUCMSC transfected with Sox2 and Ascl1 (white arrows indicate hippocampal neurons expressing MAP2). (I–L) hUCMSC
transfected with Sox2 and Neurog2. (M–O) hUCMSC transfected with only Sox2. (P–R) hMSC transfected with only Neurog2. (S–U) hUCMSC transfected with only
Ascl1. (V) Histograms show the percentage of induced neurons, measured by the number of expressing MAP2 cells over the total number of reporter positive cells.
Data are presented as mean ± s.e.m. from three independent experiments. ANOVA followed by Dunn’s post-hoc test, *p < 0.05; no statistically significant difference
(n.s.). hUCMSC transfected with control plasmids did not express MAP2, thereby the bar is not being shown.
A few days after transfection, we observed that somehUCMSCs transfected with proneural genes acquired neuronal-like morphology. To confirm this possible lineage conversionof hUCMSCs into iNs, we further analyzed the expression of
the neuronal-specific microtubule-associate protein 2 (MAP2)15 days after transfection with proneural genes (Figure 1). Afraction of cells transfected with proneural genes expressedMAP2 and acquired small-round cell bodies and thin and
Frontiers in Cellular Neuroscience | www.frontiersin.org 5 June 2018 | Volume 12 | Article 155
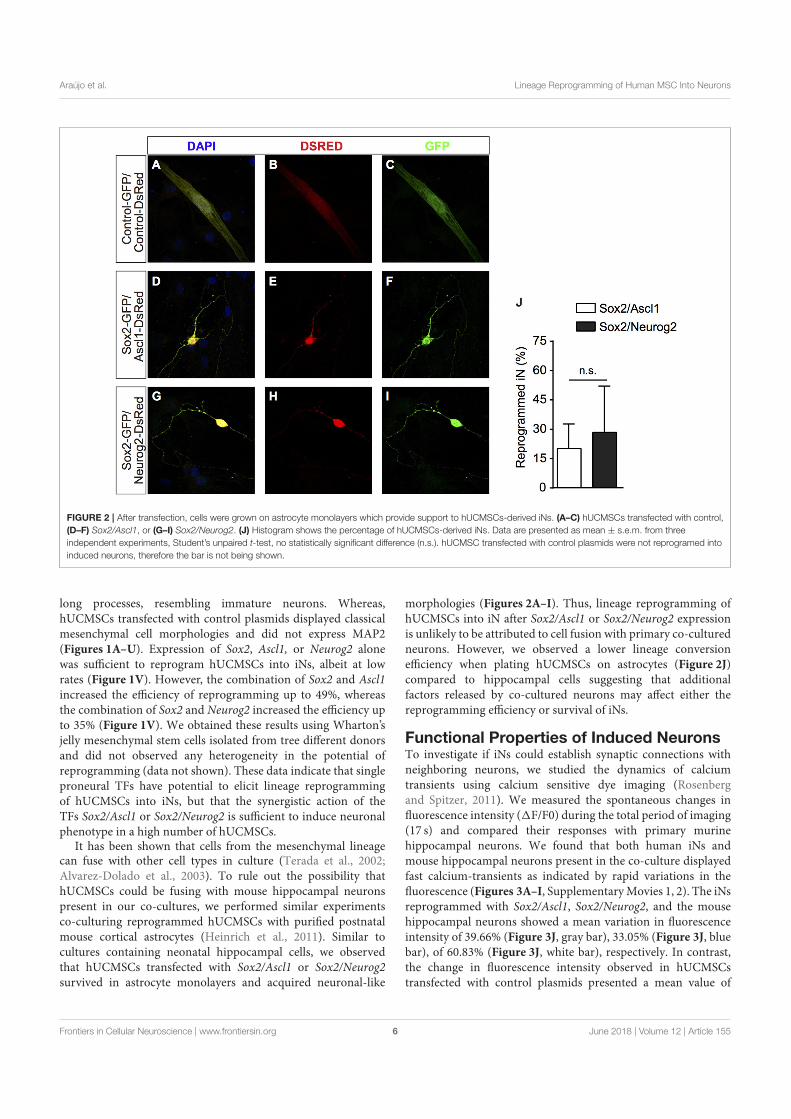
Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 2 | After transfection, cells were grown on astrocyte monolayers which provide support to hUCMSCs-derived iNs. (A–C) hUCMSCs transfected with control,
(D–F) Sox2/Ascl1, or (G–I) Sox2/Neurog2. (J) Histogram shows the percentage of hUCMSCs-derived iNs. Data are presented as mean ± s.e.m. from three
independent experiments, Student’s unpaired t-test, no statistically significant difference (n.s.). hUCMSC transfected with control plasmids were not reprogramed into
induced neurons, therefore the bar is not being shown.
long processes, resembling immature neurons. Whereas,hUCMSCs transfected with control plasmids displayed classicalmesenchymal cell morphologies and did not express MAP2(Figures 1A–U). Expression of Sox2, Ascl1, or Neurog2 alonewas sufficient to reprogram hUCMSCs into iNs, albeit at lowrates (Figure 1V). However, the combination of Sox2 and Ascl1increased the efficiency of reprogramming up to 49%, whereasthe combination of Sox2 and Neurog2 increased the efficiency upto 35% (Figure 1V). We obtained these results using Wharton’sjelly mesenchymal stem cells isolated from tree different donorsand did not observed any heterogeneity in the potential ofreprogramming (data not shown). These data indicate that singleproneural TFs have potential to elicit lineage reprogrammingof hUCMSCs into iNs, but that the synergistic action of theTFs Sox2/Ascl1 or Sox2/Neurog2 is sufficient to induce neuronalphenotype in a high number of hUCMSCs.
It has been shown that cells from the mesenchymal lineagecan fuse with other cell types in culture (Terada et al., 2002;Alvarez-Dolado et al., 2003). To rule out the possibility thathUCMSCs could be fusing with mouse hippocampal neuronspresent in our co-cultures, we performed similar experimentsco-culturing reprogrammed hUCMSCs with purified postnatalmouse cortical astrocytes (Heinrich et al., 2011). Similar tocultures containing neonatal hippocampal cells, we observedthat hUCMSCs transfected with Sox2/Ascl1 or Sox2/Neurog2survived in astrocyte monolayers and acquired neuronal-like
morphologies (Figures 2A–I). Thus, lineage reprogramming ofhUCMSCs into iN after Sox2/Ascl1 or Sox2/Neurog2 expressionis unlikely to be attributed to cell fusion with primary co-culturedneurons. However, we observed a lower lineage conversionefficiency when plating hUCMSCs on astrocytes (Figure 2J)compared to hippocampal cells suggesting that additionalfactors released by co-cultured neurons may affect either thereprogramming efficiency or survival of iNs.
Functional Properties of Induced NeuronsTo investigate if iNs could establish synaptic connections withneighboring neurons, we studied the dynamics of calciumtransients using calcium sensitive dye imaging (Rosenbergand Spitzer, 2011). We measured the spontaneous changes influorescence intensity (1F/F0) during the total period of imaging(17 s) and compared their responses with primary murinehippocampal neurons. We found that both human iNs andmouse hippocampal neurons present in the co-culture displayedfast calcium-transients as indicated by rapid variations in thefluorescence (Figures 3A–I, SupplementaryMovies 1, 2). The iNsreprogrammed with Sox2/Ascl1, Sox2/Neurog2, and the mousehippocampal neurons showed a mean variation in fluorescenceintensity of 39.66% (Figure 3J, gray bar), 33.05% (Figure 3J, bluebar), of 60.83% (Figure 3J, white bar), respectively. In contrast,the change in fluorescence intensity observed in hUCMSCstransfected with control plasmids presented a mean value of
Frontiers in Cellular Neuroscience | www.frontiersin.org 6 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 3 | Human MSC lineage-converted iNs show fast calcium transients. (A,B) hUCMSCs transfected with control plasmids encoding only reporter proteins GFP
and DSRED 23 dpt (yellow arrow). (D,E) hUCMSCs transfected with Sox2 and Neurog2 with neuronal morphology 23 dpt (yellow arrow). (G,H) Example of hUCMSCs
transfected with Sox2 and Ascl1 with neuronal morphology 23 dpt (yellow arrow). Note the presence of mouse hippocampal neurons (green arrowheads) in the same
fields. (C,F,I) Representative traces of time course calcium-transients of transfected hUCMSCs are shown by spontaneous variations in the fluorescence intensity (FI).
(J) Histograms show the mean change in fluorescence of mouse hippocampal neurons and iNs. Responses were calculated as the change in fluorescence (1F) over
the initial fluorescence (F0). Number of cells analyzed is indicated in the bars for each group. (ANOVA followed by Tukey’s post-hoc test, *p < 0.05; **p < 0.01;
***p < 0.001).
1.1% (Figure 3J, black bar), significantly lower than the responsesobserved in iNs and hippocampal neurons (ANOVA followedby Tukey’s post-test, p < 0.0001). These observations indicatethat hUCMSCs-derived iNs present fast calcium transientsqualitatively similar of those observed in primary neurons.
Notably, this sudden increase in fluorescence intensity inhUCMSC-derived iNs was temporally synchronized mousehippocampal neurons in the same field of observation(Figures 4A–I). To quantify this phenomenon, we measuredthe percentage of hippocampal neurons showing elevationin the fluorescence intensity within a time-range (ms) of the
fluorescence fluctuation observed in a single iN within the samefield of observation. The time of the iN calcium transient wasconsidered as t = 0. We found that the majority of mousehippocampal neurons showed changes in fluorescence intensitywithin 15ms of fluctuations observed in iNs (Figure 4J),indicating a strong synchronization of calcium transientsamong primary neurons and iNs. Such a strong synchronizationwithin the frame of milliseconds may suggest that cells aresynaptically connected (Dawitz et al., 2011). To further confirmthat hUCMSCs-derived iNs could receive synaptic inputs, weperformed patch clamp recordings on these cells.
Frontiers in Cellular Neuroscience | www.frontiersin.org 7 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 4 | Synchronization of calcium transients between iNs and primary neurons. (A,D,G) Photomicrographs show hUCMSC cultures 23 dpt and labeled with
BAPTA Oregon Green. Colored circles delimit the regions of interest (ROI) where fluorescence intensity (FI) was measured (colored arrowheads). (B,E,H)
Photomicrographs of the same fields show DsRed expression. Yellow arrowheads (A,B, D,E) point to hUCMSC-derived iNs. Blue, red, green and purple arrowheads
point to primary hippocampal neurons. (C,F,I) Graphics show spontaneous calcium transients. The color of ROIs in the left panel (A,D,G) corresponds to the color of
each trace in the right panel (C,F,I). The traces show spontaneous variations in the FI during 17s of recording. (J) Histogram shows percentage of hippocampal
neurons that respond within a range time difference (ms) in the same area, t=0 was considered the time of spontaneous calcium responses of the reprogrammed iN
present in the same field of imaging. Density curve is represented in black.
We performed patch-clamp recordings on iNs reprogrammedwith one transcription factor (1 TF; n = 5) and compared theiractive and passive properties to cells reprogrammed with twotranscription factors (2 TF; n = 10). Cells with 1 TF had a meaninput resistance of 726 ± 119 M�, resting membrane potentialof −61 ± 2mV and capacitance of 22 ± 2 pF. Out of the 5cells, 2 responded with regular spiking pattern (Figure 5A, topleft), 1 responded with startle onset (Figure 5A, top right), and2 with a spikelet in response to depolarizing current injections(0–100 pA, 400ms, with 10 pA increments). Spikes were analyzedfor action potential amplitude, action potential half-width andafterhyperpolarization amplitude. Spikes of cells with 1 TF hada mean action potential amplitude of 44± 8mV, action potentialhalf-width of 13 ± 0.5ms and afterhyperpolarization amplitude
of −6 ± 2mV (Figure 5C). In comparison, cells reprogrammedwith 2 TF had a mean input resistance of 605 ± 110 M�
(p= 0.51), restingmembrane potential of−59± 2mV (p= 0.54)and capacitance of 26 ± 1 pF (p = 0.13). Of the 10 cells with 2TF, 6 responded with regular spiking pattern and 4 respondedwith startle onset (0–100 pA, 400ms, with 10 pA increments).Spikes of cells with 2 TF had a mean action potential amplitudeof 76 ± 3mV (p = 0.0004), action potential half-width of 4 ±
1ms (p = 0.0006) and afterhyperpolarization amplitude of −11± 2mV (p = 0.22; Figure 5B). While hyperpolarizing currentinjections (0 to−100 pA, 400ms, with 10 pA decrements) causedsome cells to rebound (n = 3), prominent membrane sagscould only be detected in cells transfected with Sox2/Neurog2(n= 7; Figure 5B, right) suggesting that these cells have a sizeable
Frontiers in Cellular Neuroscience | www.frontiersin.org 8 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
hyperpolarization-activated current. Comparing instantaneousand steady state voltage in response to negative current injections(−100 pA, 400ms) showed a significant difference betweeninstantaneous (−71 ± 2mV) and steady state (−66 ± 1mV)values (p= 0.0427).
We also observed spontaneous excitatory postsynapticcurrents (EPSCs) in iN during voltage clamp (Figure 5D),suggesting that lineage-reprogrammed iNs could receive synapticcontacts from other neurons.We compared the first 100 events of1 TF cells and 2 TF cells against each other. Postsynaptic currentsof cells with 1 TF had an amplitude of 82 ± 9.7 pA and rise timeof 11 ± 0.4ms. The postsynaptic currents of cells with 2 TF hadan amplitude of 86 ± 15.1 pA (p = 0.8312) and a mean rise timeof 9± 0.3ms (p= 0.0211).
Sox2/Neurog2 and Sox2/Ascl1 InduceDifferent Neuronal Phenotypes inhUCMSCsNext, we set out to evaluate the expression of messengerRNAs (mRNA) of genes commonly expressed in either hMSCsor neurons. To that, we collected single cells using a glass-micropipette, isolated the total mRNA, reverse transcribed,and pre-amplified cDNAs that were used in RT-qPCRreactions. We observed that the average expression level ofcommon MSCs genes THY1 and PDGFB was decreased inthe iNs (Supplementary Figure 3A), whereas the expressionof the neuronal genes ATHO8 or NEUROD1 increased afterexpression of Sox2/Ascl1 or Sox2/Neurog2 in hMSCs, respectively(Supplementary Figure 3B). Combined with our previousobservations, these data indicate that hMSCs were effectivelyconverted into iNs by forced expression of proneural TFs.
To evaluate the possible phenotypes adopted by hMSC-derived iNs, we analysed the expression of known genesexpressed by cholinergic (CHAT), dopaminergic (TH),serotoninergic (TPH2), glutamatergic (SLC17A7), andGABAergic (SLC32A1) neurons, as well as genes encodingfor transcription factors associated with specific classes ofglutamatergic neurons within the cerebral cortex (FEZF2and BCL11B—corticofugal neurons; TBR1—cortico-thalamicneurons; SATB2—callosal neurons). Relative expression ofthese transcripts was calculated using a cycle of quantificationcutoff (Cq-cutoff) and relative-quantities of cDNA moleculeequation (Ståhlberg et al., 2013). Next, we used unsupervisedPCA analysis to classify iNs obtained from hUCMSCs expressingeither Sox2/Neurog2 or Sox2/Ascl1. We observed that theexpression levels of the transcripts for CHAT, TH, TPH2,SLC17A7, SLC32A1, FEZF2, BCL11B, TBR1, and SATB2could not clearly distinguish the two populations of cells(Figure 6A), indicating that similar genes were regulatedby both combinations of TFs in hUCMSCs-derived iNs.Indeed, we observed that both Sox2/Ascl1 and Sox2/Neurog2could induce the expression of genes associated with distinctneurochemical phenotypes in hUCMSCs-derived iNs, althoughsome phenotypes were more commonly observed for a givenTF combination. For instance, Sox2/Ascl1 generated more iNsexpressing high levels of TPH2, whereas Sox2/Neurog2 generated
more CHAT expressing iNs. Nevertheless, the expression ofall transcripts analyzed was regulated by both combinations ofTFs, suggesting that Ascl1 and Neurog2 do not have a uniquerole in the phenotypic specification of lineage reprogrammedhUCMSC-derived iNs (Figure 6B). These data suggest thatthe expression of Sox2/Ascl1 or Sox2/Neurog2 in hUCMSCactivates a transcriptional program associated with loss ofmesenchymal phenotype and acquisition of multiple neuronalphenotypes.
To further evaluate the neurochemical phenotypes ofhUCMSC-derived iNs, we investigated the expression ofSLC17A7 (Vesicular Glutamate Transporter 1 or VGLUT1)and SLC32A1 (GABA Vesicular Transporter or VGAT) usingimmunocytochemistry (Figure 7). We observed that only afew Sox2/Ascl1-derived iNs showed expression of VGAT fifteendays after reprogramming (Figures 7I–L), whereas most of thehUCMSC-derived iNs did not express any of these markers daysafter (Figures 7M–Y). Although iNs expressed MAP2 15 daysafter transfection with proneural genes (Figure 1), expressionof vesicular neurotransmitter transporters is likely to occur atlater stages of neuronal differentiation. Further analyses andimmunostaining for other vesicular transporters isoforms arenecessary to confirm the phenotypes of hUCMSCs-derivediNs.
DISCUSSION
Direct lineage reprogramming of human somatic cells intoneurons is a promising strategy to advance cell-based therapiesto treat neurological disorders, as well as to study the basicmechanisms of neuronal differentiation. In this work, we furtherexpand the list of cells suitable for direct lineage reprogramming
using transcription factors. More importantly, we show that
combined expression of either Neurog2/Sox2 or Ascl1/Sox2
is sufficient to convert human MSCs into iNs displayingelectrophysiological properties typical of neuronal cells. Finally,we show that these two combinations of transcription factorsmay elicit diverse and non-exclusive neuronal phenotypes inreprogrammed cells.
Human MSCs are versatile cells, capable of differentiation
into adipocytes, chondrocytes, and osteoblasts (Horwitz et al.,2005; Dominici et al., 2006). This potential, combined with
the fact that hMSC can be easily isolated from adult donors,has encouraged researchers to further exploit the versatility of
reprogramming MSCs to other lineages, such as muscle andneural cells for therapeutic purposes (Fan et al., 2011; Kwonet al., 2016). However, the capacity to convert MSCs into fullyfunctional neurons using extrinsic signals remains a matter ofintense debate.
Here, we show that forced expression of Ascl1, Neurog2, orSox2 alone is sufficient to convert hUCMSCs into iNs expressingkey neuronal proteins and exhibiting electrophysiologicalproperties of mature neurons. Importantly, combination ofNeurog2 or Ascl1 with Sox2 significantly increases the rate ofhUCMSC conversion into iNs (up to 35%with Sox2/Neurog2 and49% with Sox2/Ascl1). This efficiency is similar to the conversion
Frontiers in Cellular Neuroscience | www.frontiersin.org 9 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 5 | hUCMSCs-derived iNs show electrical properties of mature neurons and establish synaptic contacts with co-culture mouse primary neurons.
Electrophysiological properties of cells reprogrammed with one transcription factor (1 TF) compared to cells reprogrammed with two transcription factors (2 TF).
(A) Current clamp traces from cells with 1 TF (left: Ascl11; right: Neurog2) showing regular spiking pattern (left), and startle onset (right) in response to depolarizing
(Continued)
Frontiers in Cellular Neuroscience | www.frontiersin.org 10 June 2018 | Volume 12 | Article 155
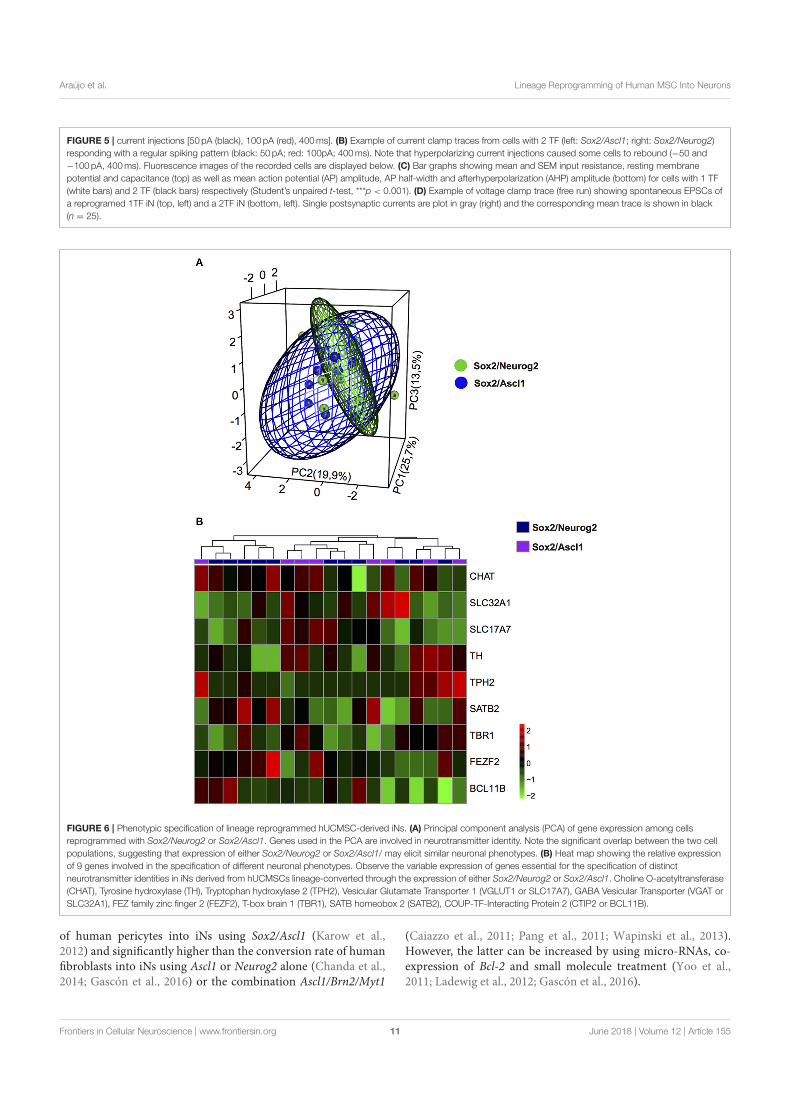
Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 5 | current injections [50 pA (black), 100 pA (red), 400ms]. (B) Example of current clamp traces from cells with 2 TF (left: Sox2/Ascl1; right: Sox2/Neurog2)
responding with a regular spiking pattern (black: 50 pA; red: 100pA; 400ms). Note that hyperpolarizing current injections caused some cells to rebound (−50 and
−100pA, 400ms). Fluorescence images of the recorded cells are displayed below. (C) Bar graphs showing mean and SEM input resistance, resting membrane
potential and capacitance (top) as well as mean action potential (AP) amplitude, AP half-width and afterhyperpolarization (AHP) amplitude (bottom) for cells with 1 TF
(white bars) and 2 TF (black bars) respectively (Student’s unpaired t-test, ***p < 0.001). (D) Example of voltage clamp trace (free run) showing spontaneous EPSCs of
a reprogramed 1TF iN (top, left) and a 2TF iN (bottom, left). Single postsynaptic currents are plot in gray (right) and the corresponding mean trace is shown in black
(n = 25).
FIGURE 6 | Phenotypic specification of lineage reprogrammed hUCMSC-derived iNs. (A) Principal component analysis (PCA) of gene expression among cells
reprogrammed with Sox2/Neurog2 or Sox2/Ascl1. Genes used in the PCA are involved in neurotransmitter identity. Note the significant overlap between the two cell
populations, suggesting that expression of either Sox2/Neurog2 or Sox2/Ascl1/ may elicit similar neuronal phenotypes. (B) Heat map showing the relative expression
of 9 genes involved in the specification of different neuronal phenotypes. Observe the variable expression of genes essential for the specification of distinct
neurotransmitter identities in iNs derived from hUCMSCs lineage-converted through the expression of either Sox2/Neurog2 or Sox2/Ascl1. Choline O-acetyltransferase
(CHAT), Tyrosine hydroxylase (TH), Tryptophan hydroxylase 2 (TPH2), Vesicular Glutamate Transporter 1 (VGLUT1 or SLC17A7), GABA Vesicular Transporter (VGAT or
SLC32A1), FEZ family zinc finger 2 (FEZF2), T-box brain 1 (TBR1), SATB homeobox 2 (SATB2), COUP-TF-Interacting Protein 2 (CTIP2 or BCL11B).
of human pericytes into iNs using Sox2/Ascl1 (Karow et al.,2012) and significantly higher than the conversion rate of humanfibroblasts into iNs using Ascl1 or Neurog2 alone (Chanda et al.,2014; Gascón et al., 2016) or the combination Ascl1/Brn2/Myt1
(Caiazzo et al., 2011; Pang et al., 2011; Wapinski et al., 2013).However, the latter can be increased by using micro-RNAs, co-expression of Bcl-2 and small molecule treatment (Yoo et al.,2011; Ladewig et al., 2012; Gascón et al., 2016).
Frontiers in Cellular Neuroscience | www.frontiersin.org 11 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
FIGURE 7 | Protein expression of vesicular transporters of lineage reprogrammed hUCMSC-derived iNs. Immunostaining for DSRED (red), GFP (green), vGAT (white;
left panel) or vGLUT1 (white; right panel) and DAPI (blue), 15 days post transfection (dpt). Scale bar represents 20µm. (A–H) hUCMSC transfected with control
plasmids encoding only reporter proteins GFP and DSRED. Note that cells displayed classical mesenchymal cell morphologies and did not express neither vGAT or
vGLUT1. (I–P) hUCMSC transfected with Sox2 and Ascl1. (Q–Y) hUCMSC transfected with Sox2 and Neurog2. (I,J) Observe the expression of the vesicular GABA
transporter (VGAT) in Sox2/Ascl1-derived iN (yellow arrows). The inset shows a high magnification view of the boxed area.
Single expression of Ascl1 is sufficient to convert other humansomatic cells into iNs (Chanda et al., 2014). This potential ofAscl1is attributed to its ability to recognize and bind to the regulatoryelements of its target genes even when they are nucleosome-bound (Wapinski et al., 2017). In contrast, the same pioneeringactivity has not been shown to Neurog2, which is believed tobind exclusively to accessible regulatory elements within thegenome. This could help to explain the prominent potential ofNeurog2 to lineage-reprogram astrocytes (Berninger et al., 2007;Heinrich et al., 2010; Chouchane et al., 2017) and pluripotentstem cells (Zhang et al., 2013) in comparison tomouse embryonicfibroblasts into iNs (Gascón et al., 2016).
Our results indicate that a fraction of hUCMSCs (1–2%) hasan epigenetic state compatible with the binding of NEUROG2to regulatory elements of neuronal genes, allowing for theconversion into iNs. However, combination with Sox2, whichhas a well-known role in chromatin modification of Neurog2-target genes (Amador-Arjona et al., 2015), largely increases theefficiency of neuronal conversion mediated by Neurog2 (∼35%).Similarly, combination with Sox2 increases the percentage ofhUCMSCs converted by Ascl1 into iNs by an order of magnitude.These observations suggest that key regulatory elements ofneuronal genes identified by ASCL1 and NEUROG2 are notaccessible in the vast majority of hUCMSCs cultured under theconditions described in this study.
In addition to the low frequency of neuronal conversionelicited in hUCMSC by forced expression of Ascl1 or Neurog2alone, iNs also display electrophysiological properties less robustcompared to iNs generated using Sox2/Ascl1 or Sox2/Neurog2.In fact, the action potential of iNs reprogrammed with asingle TF has a smaller amplitude and a shorter half-widthas compared to iNs reprogrammed with 2 TFs (Sox2/Ascl1or Sox2/Neurog2), indicating that the latter express a morecomplete set of ion channels. It is possible that thesedifferences represent a delay in the maturation of single-TF iNs.Alternatively, the combination of Sox2/Ascl1 or Sox2/Neurog2may be necessary to induce the complete transcriptional cascaderequired for thorough neuronal maturation. Calcium transientsare implicated in distinct aspects of neuronal differentiation byregulating neurotransmitter phenotype, dendritic morphology,and axonal growth and guidance (Rosenberg and Spitzer,2011). While control-transfected hUCMSCs never displayedfast calcium transients, both Sox2/Ascl1- and Sox2/Neurog2-iNs showed spontaneous fast calcium transients, indicative ofsynaptic activity (Bonifazi et al., 2009). Likewise, the calciumtransients of primary neurons and iNs are synchronized,suggesting that these cells are synaptically connected. Together,these findings suggest that hUCMSCs are lineage-converted intoiNs capable of firing action potentials and establishing pre- andpost-synaptic compartments.
Frontiers in Cellular Neuroscience | www.frontiersin.org 12 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
The cellular and molecular mechanisms of direct lineagereprogramming remain largely unknown. It has been reportedthat the metabolic state is particularly important in directneuronal reprogramming of somatic cells into iNs. Accordingly,co-expression of Bcl2/Neurog2 or Bcl2/Ascl1 greatly enhancesthe conversion efficiency of astrocytes into iNs by inhibitinglipid peroxidation, consistent with a caspase-independent role.Similarly, co-expression of Bcl-2 alongside Ascl1 improves therates of lineage conversion of mouse embryonic fibroblastinto iNs, demonstrating that the metabolic shift is necessary
to support survival of lineage-converted iNs (Gascón et al.,2016). Our data suggest that mouse astrocytes and hippocampalneurons may contribute to enhance hUCMSCs survival duringlineage conversion. Future experiments should elucidate whetherBcl2 co-expression or small molecules treatment would allowfor the conversion of Sox2/Neurog2- or Sox2/Ascl1-iNs fromhUCMSCs even in the absence of co-cultured cells.
Despite the large number of studies showing the conversion ofhuman somatic cells into iNs, it remains largely unknown whatis the phenotype of reprogrammed neurons (Ambasudhan et al.,2011; Pang et al., 2011; Son et al., 2011; Karow et al., 2012; Chandaet al., 2014; Hu et al., 2015). Moreover, it is still unclear whetherdifferent TFs could induce particular neuronal fates in lineage-converted cells. Here, we show that lineage-reprogrammedhUCMSCs generate iNs expressing genes associated with theacquisition of diverse neurotransmitter identities, regardlessof the use of Sox2/Ascl1 or Sox2/Neurog2. These differentcombinations of TFs can regulate similar sets of genes,suggesting that Sox2/Ascl1 and Sox2/Neurog2 are not sufficientto drive unambiguous neurotransmitter identities in hUCMSCs-derived iNs. However, the expression of genes associatedwith a specific neuronal phenotype is only an indicationof the possible phenotype of the iNs. Future experimentsusing electrophysiological and pharmacological techniques arenecessary to confirm the phenotypes of hUCMSCs-derived iNs.
According to the notion that Neurog2 and Ascl1 may besufficient to induce a pro-neuronal program during somaticcell lineage reprogramming but not be sufficient to determinea specific phenotype of the iN, studies of the developingcentral nervous system reveal that those TFs may be associatedwith diverse neuronal phenotypes. For instance, while inthe telencephalon, Neurog2 plays important roles for thespecification of glutamatergic neurons (Schuurmans andGuillemot, 2002). Progenitors in the cerebellum and spinal cordexpress Neurog2 generate GABAergic and cholinergic neurons,respectively (Bertrand et al., 2002). Similarly, progenitorsexpressing Ascl1 contribute to different neuronal lineages inthe cerebral cortex, cerebellum, and retina (Chouchane andCosta, 2018). Most protocols aiming at obtaining fibroblast-derived iNs with a particular phenotype through direct lineagereprogramming require the use of several TFs (Victor et al., 2014;Blanchard et al., 2015).
Expression of either Ascl1 and Neurog2 in cortical astrocytesleads to the activation of transcriptional networks with only asmall subset of shared target genes (Masserdotti et al., 2015),which could partly explain the role of those TFs in instructingdifferent iNs phenotypes (Berninger et al., 2007; Heinrich et al.,
2010). However, co-expression of Ascl1, Myt1L, and Brn2induces a glutamatergic neuronal fate in fibroblast-derived iNs(Vierbuchen et al., 2010), whereas Neurog2 drives motor neurondifferentiation associated with forskolin and dorsomorphintreatments in the same cells (Liu et al., 2013), suggesting thatthe fate-specification of iNs is not only dependent on theTF used. Recent work in our laboratory using direct lineagereprogramming of mouse astrocytes isolated from different brainregions further supports the versatile roles of Neurog2 andAscl1 to affect the phenotypes of iNs (Chouchane et al., 2017).While cerebral cortex astrocytes reprogrammed into iNs withNeurog2 adopt mostly a glutamatergic fate, cerebellum astrocyte-derived iNs show GABAergic phenotypes. Taken together, thesedata indicate that the cell of origin with its specific epigeneticlandscape can influence the final fate of iNs.
A comprehensive understanding of the molecularmechanisms involved in the acquisition of particularneurochemical phenotypes will greatly improve the protocols forlineage reprogramming of human somatic cells into iNs, allowingfor the generation of homogeneous neuronal populations thatcould be later used in cell-based therapies.
AUTHOR CONTRIBUTIONS
All authors reviewed the manuscript. JAMA contributed todesign, performed most of the experiments, analyzed the data,discussed the results, and wrote the manuscript. DAC performedisolation and characterization of hUCMSC. SRBM assisted andprovided financial support with isolation and characterizationof hUCMSC. RNL performed electrophysiology experiments.MMH analyzed electrophysiology experiments data, discussedthe results, and helped writing the manuscript. DCFG performedqPCR experiments. DM-C analyzed the single cell qPCR data.MRC provided financial support, directed the project, conceivedthe experiment, analyzed data, discussed the results, and wrotethe manuscript.
FUNDING
This work was supported by Conselho Nacional deDesenvolvimento Científico e Tecnológico (CNPq) andCoordenação de Aperfeiçoamento de Pessoal de Nível Superior(CAPES).
ACKNOWLEDGMENTS
We thank Ana Raquel for her excellent technical help. Theumbilical cord cells were obtained through the Laboratório deBiologia Molecular e Genômica (Natal, Brazil), we thank SusanaMoreira and Tatiana Bressel for their assistance providing thecells.
SUPPLEMENTARY MATERIAL
The Supplementary Material for this article can be foundonline at: https://www.frontiersin.org/articles/10.3389/fncel.2018.00155/full#supplementary-material
Frontiers in Cellular Neuroscience | www.frontiersin.org 13 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
REFERENCES
Afanasyev, B. V., Elstner, E. E., and Zander, A. R. (2010). A. J. Friedenstein,
founder of the mesenchymal stem cell concept. Cell. Ther. Transpl. 1, 35–38.
doi: 10.3205/ctt-2009-en-000029.01
Alvarez-Dolado, M., Pardal, R., Garcia-Verdugo, J. M., Fike, J. R., Lee, H.
O., Pfeffer, K., et al. (2003). Fusion of bone-marrow-derived cells with
Purkinje neurons, cardiomyocytes and hepatocytes. Nature 425, 968–973.
doi: 10.1038/nature02069
Amador-Arjona, A., Cimadamore, F., Huang, C. T., Wright, R., Lewis, S., Gage,
F. H., et al. (2015). SOX2 primes the epigenetic landscape in neural precursors
enabling proper gene activation during hippocampal neurogenesis. Proc. Natl.
Acad. Sci. U.S.A. 112, E1936–E1945. doi: 10.1073/pnas.1421480112
Ambasudhan, R., Talantova, M., Coleman, R., Yuan, X., Zhu, S., Lipton, S.
A., et al. (2011). Direct reprogramming of adult human fibroblasts to
functional neurons under defined conditions. Cell Stem Cell 9, 113–118.
doi: 10.1016/j.stem.2011.07.002
Aoi, T., Yae, K., Nakagawa, M., Ichisaka, T., Okita, K., Takahashi, K., et al. (2008).
Generation of pluripotent stem cells from adult mouse liver and stomach cells.
Science 321, 699–702. doi: 10.1126/science.1154884
Berninger, B., Costa, M. R., Koch, U., Schroeder, T., Sutor, B., Grothe,
B., et al. (2007). Functional properties of neurons derived from in
vitro reprogrammed postnatal astroglia. J. Neurosci. 27, 8654–8664.
doi: 10.1523/JNEUROSCI.1615-07.2007
Bertrand, N., Castro, D. S., and Guillemot, F. (2002). Proneural genes and
the specification of neural cell types. Nat. Rev. Neurosci. 3, 517–530.
doi: 10.1038/nrn874
Blanchard, J. W., Eade, K. T., Szucs, A., Lo Sardo, V., Tsunemoto, R. K., Williams,
D., et al. (2015). Selective conversion of fibroblasts into peripheral sensory
neurons. Nat. Neurosci. 18, 25–35. doi: 10.1038/nn.3887
Bonifazi, P., Goldin, M., Picardo, M. A., Jorquera, I., Cattani, A., Bianconi, G.,
et al. (2009). GABAergic hub neurons orchestrate synchrony in developing
hippocampal networks. Science 326, 1419–1424. doi: 10.1126/science.1175509
Caiazzo, M., Dell’Anno, M. T., Dvoretskova, E., Lazarevic, D., Taverna, S., Leo,
D., et al. (2011). Direct generation of functional dopaminergic neurons from
mouse and human fibroblasts. Nature 476, 224–227. doi: 10.1038/nature10284
Caplan, A. (1991). Mesenchymal stem cells. J. Orthop. Res. 9, 641–650.
doi: 10.1002/jor.1100090504
Chanda, S., Ang, C. E., Davila, J., Pak, C., Mall, M., Lee, Q. Y., et al. (2014).
Generation of induced neuronal cells by the single reprogramming factor
ASCL1. Stem Cell Rep. 3, 282–296. doi: 10.1016/j.stemcr.2014.05.020
Chouchane, M., and Costa, M. R. (2018). Instructing neuronal identity during CNS
development and astroglial-lineage reprogramming: roles of NEUROG2 and
ASCL1. Brain Res. doi: 10.1016/j.brainres.2018.02.045. [Epub ahead of print].
Chouchane, M., Melo de Farias, A. R., Moura, D. M. S., Hilscher, M. M., Schroeder,
T., Leão, R. N., et al. (2017). Lineage reprogramming of astroglial cells from
different origins into distinct neuronal subtypes. Stem Cell Rep. 9, 162–176.
doi: 10.1016/j.stemcr.2017.05.009
Dawitz, J., Kroon, T., Hjorth, J. J., and Meredith, R. M. (2011). Functional calcium
imaging in developing cortical networks. J. Vis. Exp. 56, 1–8. doi: 10.3791/3550
Ding, D.-C., Shyu, W.-C., and Lin, S.-Z. (2011). Mesenchymal stem cells. Cell
Transplant. 20, 5–14. doi: 10.3727/096368910X
Dominici, M., Le Blanc, K., Mueller, I., Slaper-Cortenbach, I., Marini, F., Krause,
D., et al. (2006). Minimal criteria for defining multipotent mesenchymal
stromal cells. Int. Soc. Cell. Ther. Posit. Stat. Cytother. 8, 315–317.
doi: 10.1080/14653240600855905
Duarte, D. M., Cornélio, D. A., Corado, C., Medeiros, V. K., de Araújo L.
A., Cavalvanti, G. B. Jr. et al., (2012). Chromosomal characterization of
cryopreserved mesenchymal stem cells from the human subendothelium
umbilical cord vein. Regen. Med. 7, 147–157. doi: 10.2217/rme.11.113
Espejel, S., Roll, G. R., McLaughlin, K. J., Lee, A. Y., Zhang, J. Y., Laird, D. J., et al.
(2010). Induced pluripotent stem cell-derived hepatocytes have the functional
and proliferative capabilities needed for liver regeneration in mice. J. Clin.
Invest. 120, 3120–3126. doi: 10.1172/JCI43267
Fan, C. G., Zhang, Q. J., and Zhou, J. R. (2011). Therapeutic potentials of
mesenchymal stem cells derived from human umbilical cord. Stem Cell Rev.
7, 195–207. doi: 10.1007/s12015-010-9168-8
Gascón, S., Murenu, E., Masserdotti, G., Ortega, F., Russo, G. L., Petrik,
D., et al. (2016). Identification and successful negotiation of a metabolic
checkpoint in direct neuronal reprogramming. Cell Stem Cell 18, 396–409.
doi: 10.1016/j.stem.2015.12.003
Hanna, J., Markoulaki, S., Schorderet, P., Carey, B. W., Beard, C., Wernig, M.,
et al. (2008). Direct reprogramming of terminally differentiated mature B
lymphocytes to pluripotency. Cell 133, 250–264. doi: 10.1016/j.cell.2008.03.028
Heinrich, C., Blum, R., Gascón, S., Masserdotti, G., Tripathi, P., Sánchez, R.,
et al. (2010). Directing astroglia from the cerebral cortex into subtype specific
functional neurons. PLoS Biol. 8:e373. doi: 10.1371/journal.pbio.1000373
Heinrich, C., Gascón, S., Masserdotti, G., Lepier, A., Sanchez, R., Simon-Ebert, T.,
et al. (2011). Generation of subtype-specific neurons from postnatal astroglia of
the mouse cerebral cortex.Nat. Protoc. 6, 214–228. doi: 10.1038/nprot.2010.188
Horwitz, E. M., Le Blanc, K., Dominici, M., Mueller, I., Slaper-Cortenbach, I.,
Marini, F. C., et al. (2005). Clarification of the nomenclature for MSC: the
International Society for Cellular Therapy position statement. Cytotherapy 7,
393–395. doi: 10.1080/14653240500319234
Hu, W., Qiu, B., Guan, W., Wang, Q., Wang, M., Li, W., et al. (2015).
Direct conversion of normal and Alzheimer’s disease human fibroblasts
into neuronal cells by small molecules. Cell Stem Cell 17, 204–212.
doi: 10.1016/j.stem.2015.07.006
Imamura, M., Aoi, T., Tokumasu, A., Mise, N., Abe, K., Yamanaka, S., et al.
(2010). Induction of primordial germ cells from mouse induced pluripotent
stem cells derived from adult hepatocytes. Mol. Reprod. Dev. 77, 802–811.
doi: 10.1002/mrd.21223
Karow, M., Sánchez, R., Schichor, C., Masserdotti, G., Ortega, F., Heinrich,
C., et al. (2012). Reprogramming of pericyte-derived cells of the adult
human brain into induced neuronal cells. Cell Stem Cell 11, 471–476.
doi: 10.1016/j.stem.2012.07.007
Keating, A. (2012). Mesenchymal stromal cells: new directions. Cell Stem Cell 10,
709–716. doi: 10.1016/j.stem.2012.05.015
Kuzmenkin, A., Liang, H., Xu, G., Pfannkuche, K., Eichhorn, H., Fatima, A.,
et al. (2009). Functional characterization of cardiomyocytes derived from
murine induced pluripotent stem cells in vitro. FASEB J. 23, 4168–4180.
doi: 10.1096/fj.08-128546
Kwon, A., Kim, Y., Kim, M., Kim, J., Choi, H., Jekarl, D. W., et al. (2016). Tissue-
specific differentiation potency of mesenchymal stromal cells from perinatal
tissues. Sci. Rep. 6, 1–11. doi: 10.1038/srep23544
Ladewig, J., Mertens, J., Kesavan, J., Doerr, J., Poppe, D., Glaue, F., et al.
(2012). Small molecules enable highly efficient neuronal conversion of human
fibroblasts. Nat. Methods 9, 575–578. doi: 10.1038/nmeth.1972
Liu, M. L., Zang, T., Zou, Y., Chang, J. C., Gibson, J. R., Huber, K. M., et al. (2013).
Small molecules enable neurogenin 2 to efficiently convert human fibroblasts
into cholinergic neurons. Nat Commun 4:2183. doi: 10.1038/ncomms3183
Marro, S., Pang, Z. P., Yang, N., Tsai, M. C., Qu, K., Chang, H. Y., et al. (2011).
Direct lineage conversion of terminally differentiated hepatocytes to functional
neurons. Cell Stem Cell 9, 374–382. doi: 10.1016/j.stem.2011.09.002
Masserdotti, G., Gillotin, S., Sutor, B., Drechsel, D., Irmler, M., Jørgensen, H. F.,
et al. (2015). Transcriptional mechanisms of proneural factors and REST in
regulating neuronal reprogramming of astrocytes. Cell Stem Cell 17, 74–88.
doi: 10.1016/j.stem.2015.05.014
Meissner, A., Wernig, M., and Jaenisch, R. (2007). Direct reprogramming of
genetically unmodified fibroblasts into pluripotent stem cells. Nat. Biotechnol.
25, 1177–1181. doi: 10.1038/nbt1335
Mizuno, Y., Chang, H., Umeda, K., Niwa, A., Iwasa, T., Awaya, T., et al. (2010).
Generation of skeletal muscle stem/progenitor cells from murine induced
pluripotent stem cells. FASEB J. 24, 2245–2253. doi: 10.1096/fj.09-137174
Okita, K., Ichisaka, T., and Yamanaka, S. (2007). Generation of germline-
competent induced pluripotent stem cells. Nature 448, 313–317.
doi: 10.1038/nature05934
Pang, Z. P., Yang, N., Vierbuchen, T., Ostermeier, A., Fuentes, D. R.,
Yang, T. Q., et al. (2011). Induction of human neuronal cells by
defined transcription factors. Nature 476, 220–223. doi: 10.1038/nature
10202
Rosenberg, S. S., and Spitzer, N. C. (2011). Calcium signaling in
neuronal development. Cold Spring Harb. Perspect. Biol. 3, 1–13.
doi: 10.1101/cshperspect.a004259
Frontiers in Cellular Neuroscience | www.frontiersin.org 14 June 2018 | Volume 12 | Article 155

Araújo et al. Lineage Reprogramming of Human MSC Into Neurons
Schuurmans, C., and Guillemot, F. (2002). Molecular mechanisms underlying cell
fate specification in the developing telencephalon. Curr. Opin. Neurobiol. 12,
26–34. doi: 10.1016/S0959-4388(02)00286-6
Son, E. Y., Ichida, J. K., Wainger, B. J., Toma, J. S., Rafuse, V. F., Woolf, C. J.,
et al. (2011). Conversion of mouse and human fibroblasts into functional spinal
motor neurons. Cell Stem Cell 9, 205–218. doi: 10.1016/j.stem.2011.07.014
Ståhlberg, A., Rusnakova, V., Forootan, A., Anderova, M., and Kubista, M.
(2013). RT-qPCR work-flow for single-cell data analysis. Methods 59, 80–88.
doi: 10.1016/j.ymeth.2012.09.007
Takahashi, K., and Yamanaka, S. (2006). Induction of pluripotent stem cells from
mouse embryonic and adult fibroblast cultures by defined factors. Cell 126,
663–676. doi: 10.1016/j.cell.2006.07.024
Terada, N., Hamazaki, T., Oka, M., Hoki, M., Mastalerz, D. M., Nakano, Y., et al.
(2002). Bone marrow cells adopt the phenotype of other cells by spontaneous
cell fusion. Nature 416, 542–545. doi: 10.1038/nature730
Victor, M. B., Richner, M., Hermanstyne, T. O., Ransdell, J. L., Sobieski, C.,
Deng, P., et al. (2014). Generation of human striatal neurons by MicroRNA-
dependent direct conversion of fibroblasts. Neuroresource 84, 311–323.
doi: 10.1016/j.neuron.2014.10.016
Vierbuchen, T., Ostermeier, A., Pang, Z. P., Kokubu, Y., Südhof, T. C., andWernig,
M. (2010). Direct conversion of fibroblasts to functional neurons by defined
factors. Nature 463, 1035–1041. doi: 10.1038/nature08797
Wapinski, O. L., Lee, Q. Y., Chen, A. C., Li, R., Corces, M. R., Ang, C. E., et al.
(2017). Rapid chromatin switch in the direct reprogramming of fibroblasts to
neurons. Cell Rep. 20, 3236–3247. doi: 10.1016/j.celrep.2017.09.011
Wapinski, O. L., Vierbuchen, T., Qu, K., Lee, Q. Y., Chanda, S., Fuentes, D. R.,
et al. (2013). Hierarchical mechanisms for direct reprogramming of fibroblasts
to neurons. Cell 155, 621–635. doi: 10.1016/j.cell.2013.09.028
Wernig, M., Zhao, J. P., Pruszak, J., Hedlund, E., Fu, D., Soldner, F., et al. (2008).
Neurons derived from reprogrammed fibroblasts functionally integrate into the
fetal brain and improve symptoms of rats with Parkinson’s disease. Proc. Natl.
Acad. Sci. U.S.A. 105, 5856–5861. doi: 10.1073/pnas.0801677105
Yoo, A. S., Sun, A. X., Li, L., Shcheglovitov, A., Portmann, T., Li, Y., et al. (2011).
MicroRNA-mediated conversion of human fibroblasts to neurons. Nature 476,
228–231. doi: 10.1038/nature10323
Zhang, Y., Pak, C., Han, Y., Ahlenius, H., Zhang, Z., Chanda, S., et al. (2013). Rapid
single-step induction of functional neurons from human pluripotent stem cells.
Neuron 78, 785–798. doi: 10.1016/j.neuron.2013.05.029
Conflict of Interest Statement: The authors declare that the research was
conducted in the absence of any commercial or financial relationships that could
be construed as a potential conflict of interest.
Copyright © 2018 Araújo, Hilscher, Marques-Coelho, Golbert, Cornelio, Batistuzzo
de Medeiros, Leão and Costa. This is an open-access article distributed under the
terms of the Creative Commons Attribution License (CC BY). The use, distribution
or reproduction in other forums is permitted, provided the original author(s) and the
copyright owner are credited and that the original publication in this journal is cited,
in accordance with accepted academic practice. No use, distribution or reproduction
is permitted which does not comply with these terms.
Frontiers in Cellular Neuroscience | www.frontiersin.org 15 June 2018 | Volume 12 | Article 155

ORIGINAL RESEARCHpublished: 12 November 2018doi: 10.3389/fncel.2018.00410
Frontiers in Cellular Neuroscience | www.frontiersin.org 1 November 2018 | Volume 12 | Article 410
Edited by:
Sandra Henriques Vaz,
Instituto de Medicina Molecular (IMM),
Portugal
Reviewed by:
Antje Grosche,
Ludwig-Maximilians-Universität
München, Germany
Xiao-Feng Zhao,
University of Michigan, United States
*Correspondence:
Marcos R. Costa
†These authors have contributed
equally to this work
Received: 24 August 2018
Accepted: 22 October 2018
Published: 12 November 2018
Citation:
Guimarães RPdM, Landeira BS,
Marques-Coelho D, Golbert DCF,
Silveira MS, Linden R, de Melo Reis
RA and Costa MR (2018) Evidence of
Müller Glia Conversion Into Retina
Ganglion Cells Using Neurogenin2.
Front. Cell. Neurosci. 12:410.
doi: 10.3389/fncel.2018.00410
Evidence of Müller Glia ConversionInto Retina Ganglion Cells UsingNeurogenin2
Roberta Pereira de Melo Guimarães 1,2,3†, Bruna Soares Landeira 1†,
Diego Marques Coelho 1,4, Daiane Cristina Ferreira Golbert 1, Mariana S. Silveira 2,
Rafael Linden 2, Ricardo A. de Melo Reis 3 and Marcos R. Costa 1*
1 Brain Institute, Federal University of Rio Grande do Norte, Natal, Brazil, 2 Lab Neurogenesis, Institute of Biophysics Carlos
Chagas Filho, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil, 3 Lab Neurochemistry, Institute of Biophysics Carlos
Chagas Filho, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil, 4 Bioinformatics Multidisciplinary Environment, IMD,
Federal University of Rio Grande do Norte, Rio de Janeiro, Brazil
Degenerative retinopathies are the leading causes of irreversible visual impairment in
the elderly, affecting hundreds of millions of patients. Müller glia cells (MGC), the main
type of glia found in the vertebrate retina, can resume proliferation in the rodent adult
injured retina but contribute weakly to tissue repair when compared to zebrafish retina.
However, postnatal and adult mouse MGC can be genetically reprogrammed through
the expression of the transcription factor (TF) Achaete-scute homolog 1 (ASCL1) into
induced neurons (iNs), displaying key hallmarks of photoreceptors, bipolar and amacrine
cells, which may contribute to regenerate the damaged retina. Here, we show that the
TF neurogenin 2 (NEUROG2) is also sufficient to lineage-reprogram postnatal mouse
MGC into iNs. The efficiency of MGC lineage conversion by NEUROG2 is similar to that
observed after expression of ASCL1 and both TFs induce the generation of functionally
active iNs. Treatment of MGC cultures with EGF and FGF2 prior to Neurog2 or Ascl1
expression enhances reprogramming efficiencies, what can be at least partially explained
by an increase in the frequency of MGCs expressing sex determining region Y (SRY)-box
2 (SOX2). Transduction of either Neurog2 or Ascl1 led to the upregulation of key
retina neuronal genes in MGC-derived iNs, but only NEUROG2 induced a consistent
increase in the expression of putative retinal ganglion cell (RGC) genes. Moreover, in
vivo electroporation of Neurog2 in late progenitors from the neonatal rat retina, which
are transcriptionally similar to MGCs, also induced a shift in the generation of retinal
cell subtypes, favoring neuronal differentiation at the expense of MGCs and resuming
the generation of RGCs. Altogether, our data indicate that NEUROG2 induces lineage
conversion of postnatal rodent MGCs into RGC-like iNs in vitro and resumes the
generation of this neuronal type from late progenitors of the retina in vivo.
Keywords: retina, müller glia cells, induced neurons, lineage-reprogramming, neurogenin2, Ascl1, retina ganglion
cells

Guimarães et al. Reprogramming Müller Cells Into RGCs
INTRODUCTION
The retina is a unique tissue with highly organized architecture,known to be one of the most energetically demanding systemsin the nervous system (Wong-Riley, 2010). Due to oxidativestress, trauma, or genetic mutations, gradual and irreversible celldeath affects specific neuronal types in the retina (Athanasiouet al., 2013). For instance, retinal ganglion cells (RGCs) andtheir axons degenerate in glaucoma, a neurodegenerative diseaseassociated with increased intraocular pressure, eventually leadingto blindness (Kimura et al., 2017). In the last 5 years almost 65million people worldwide were diagnosed with glaucoma (Gillet al., 2016; Liang et al., 2017), which is the leading cause ofvisual impairment in developed countries (WHO). Still, despitethe social and economic burden of such disease, therapeuticapproaches are limited. Recent progress in cell-based therapymay, nonetheless, provide novel means to restore vision inglaucoma patients (Abu-Hassan et al., 2015; Chamling et al.,2016).
Cell lineage-reprogramming techniques, which allow thedirect conversion of a non-neuronal cell into neurons, offer apowerful strategy to regenerate neuronal cells in the injuredretina. In fact, expression of the bHLH neurogenic transcriptionfactor (TF) Achaete-scute homolog 1 (ASCL1) in vitro inducedthe reprogramming of mouse Müller glia cells (MGC) intobipolar cells and, to a lesser extent, amacrine cells (Pollaket al., 2013). Following NMDA-mediated injury in postnatalmouse retina, ASCL1 expression reprogrammed MGCs intoneurons expressing markers of bipolar cells, amacrine cells andphotoreceptors (Ueki et al., 2015). Notably, when combined withthe inhibitor of histone deacetylases trichostatin A, expression ofASCL1 elicited the conversion of someMGC into bipolar (∼18%)and amacrine (∼3%) cells in the injured adult retina (Jorstadet al., 2017). These findings demonstrate that regenerative effectsof transgenic expression of ASCL1 in the adult mouse Müllerglia are more limited as compared to the regenerative responseobserved in non-mammalian vertebrates (Wilken and Reh,2016).Moreover, ASCL1 expression is not sufficient to reprogramMGCs into RGCs either in vitro or in vivo (Pollak et al., 2013;Ueki et al., 2015; Jorstad et al., 2017).
During development, expression of ASCL1 defines a subset ofretinal progenitor cells (RPCs) that generate all neuronal typesin the retina, except RGCs (Brzezinski et al., 2011). In contrast,expression of the bHLH TF Neurogenin 2 (NEUROG2) defines aseparate set of RPCs, co-expressing the POUClass 4 Homeobox 1and 2 (Pou4f1/Brn3a and Pou4f2/Brn3b) and contributing to thegeneration of RGCs (Hufnagel et al., 2010; Brzezinski et al., 2011).Interestingly, knocking down the expression of Pou4f2/Brn3b inMGCs cultured in conditions to induce stem cell-like propertieshampers the differentiation into RGCs (Singhal et al., 2012; Songet al., 2013; Wu et al., 2016).
Here we report that forced expression of NEUROG2 issufficient to convert postnatal rodent MGC into a neurogenicstate. Either ASCL1 or NEUROG2 elicited induced neurons (iNs)that express genes of bipolar, horizontal and amacrine cells,as well as photoreceptors. However, only forced expression ofNEUROG2 led to the generation of iNs expressing hallmarks
of RGCs. We also show that treatment with epidermal growthfactor (EGF) and basic fibroblast growth factor (FGF-2) duringthe expansion of MGCs affects lineage-conversion efficienciesand iN-fate specification. Finally, we provide evidence for aninstructive role of NEUROG2 in the specification of RGC fatein late retinal progenitors that are not competent to generatethis cell type in vivo (Young, 1985; Turner et al., 1990; Rapaportet al., 2004; He et al., 2012). Collectively, our results indicate thatNEUROG2 can regulate the specification program of both lateretinal progenitors and MGC to generate RGCs, and, therefore,might be an interesting candidate for gene-based therapies totreat retinal degenerations.
MATERIALS AND METHODS
AnimalsC57BL/6 mice were obtained from the Biotério Setorial doInstituto do Cérebro (BISIC). All experiments were approvedby and carried out in accordance with the guidelines of theInstitutional Animal Care and Use Committee of the FederalUniversity of Rio Grande do Norte (license number #048/2014).
Müller Glial Cell (MGC) CultureMGCs were purified from postnatal day (P)7-9 mice accordingto previously described protocols (de Melo Reis et al., 2008).Briefly, retinas were dissected out and chemically dissociatedwith TrypLE (Life Technologies) for 10min at 37◦C. Isolatedcells were counted using a Neubeuer chamber and plated ontoT75 culture flasks with DMEM F12 (Gibco) plus 10% fetalbovine serum (Gibco), 3.5mM glucose (Sigma), 4.5g/L GlutaMax(Gibco), 100 U/mL penicillin/streptomycin (Gibco), either withor without 10 ng/mL of epidermal growth factor (EGF, Gibco)and 10 ng/mL of fibroblast growth factor 2 (FGF2, Gibco). Halfof the medium was changed once a week during the period ofMGCs expansion.
PlasmidsPlasmids contain the internal chicken β-actin promoter fusedwith a cytomegalovirus enhancer (pCAG), the coding sequencefor either Ascl1 or Neurog2, an internal ribosomal entry site (I)and coding sequences for either DsRed or GFP (pCAG-Ascl1-I-DsRed, pCAG-Neurog2-I-DsRed and pCAG-Neurog2-I-GFP).Control plasmids encode only DsRed or GFP (pCAG-I-DsRed orpCAG-I-GFP).
Plasmid stocks were prepared in Escherichia coli and purifiedusing an endotoxin-freeMaxiprep plasmid kit (Invitrogen). DNAconcentration was adjusted to 1 µg/µL in endotoxin free TEbuffer, and plasmids were stored at−20◦C.
NucleofectionAfter confluence, MGCs were chemically detached from T75culture flasks with TrypLE enzyme at 37◦C, and ∼3 × 105 cellswere mixed with P3 solution (Lonza) and 1 µg of plasmidsencoding for either NEUROGENIN2 (pCAG-Neurog2-IRES-DsRed) or ASCL1 (pCAG-Ascl1-IRES-DsRed) or only reporterprotein DSRED (pCAG-IRES-DsRed). These solutions wereplaced in a special cuvette and electroporated using Nucleofector
Frontiers in Cellular Neuroscience | www.frontiersin.org 2 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
4D (Lonza) with the P3 primary cell program. Next, 8 ×
104 cells were plated onto glass-coverslips (100013-Knittel)previously coated with laminin (L2020–SIGMA) and Poly-Dlysine (Sigma) containing pre-warmed DMEM F12, 10% fetalbovine serum, 3.5mM glucose, 4.5g/L GlutaMax and 100U/mLpenicillin/streptomycin. After 24h, medium was replaced withdifferentiation medium containing DMEM F12, 3.5mM glucose,4.5 g/l GlutaMax, 100 U/ml penicillin/streptomycin and 2% B27(Gibco).
In vivo ElectroporationIn vivo electroporation was performed as previously described(Matsuda and Cepko, 2004, 2007). Briefly, P0 Lister hooded ratswere anesthetized by placing on ice. One microliter of DNA mix(6.5 µg/µL) containing 0.1% Fast Green dye (Sigma) preparedin HBSS saline was injected into the subretinal space with aHamilton syringe equipped with a 33G blunt end needle. Five99V pulses were administered for 50ms at 950ms intervals,using a forceps-type electrode (Nepagene, CUY650P7) withNeurgel (Spes Medica). The electrodes were oriented such thatthe positive pole electrode was placed over the injected eyeand the negative pole electrode was placed over non-injectedeye to ensure that the electrical field is oriented correctly todrive the injected DNA solution from the subretinal spaceinto progenitors. Fast Green in the DNA mix is an injectiontracer, which facilitates observation of the spread of the injectionsolution into the subretinal space (deMelo and Blackshaw, 2018).In addition, the efficiency of electroporation was verified 10 daysafter electroporation when GFP positive cells were observed infreshly dissected retinas. Retinas were then fixed by immersion in4% paraformaldehyde in PBS for 16 h. Serial transversal sectionsof cryoprotected material (10µm) were mounted on either Poly-L-lysine (300µg/mL) or silane (6%, Sigma)-treated microscopeslides.
ImmunocytochemistryCell cultures were fixed in 4% paraformaldehyde (PFA) inPBS for 15min at room temperature. The cells were incubatedovernight at 4◦C, with primary antibodies diluted in PBS,0.5% Triton X-100 and 5% goat serum, washed and incubatedfor 2 h at room temperature, with species-specific secondaryantibodies conjugated to Alexa fluorophores. To stain thenuclei, cells were incubated for 5min with 0.1µg/mL DAPI(4’6’-diamino-2-phenylindone) in PBS 0.1M. Coverslips weremounted onto glass slides with Aqua Poly/Mount mountingmedium (Polysciences, Warrington, PA). Primary antibodiesand respective dilutions were: chicken anti-Green FluorescentProtein (Aves Labs, cat#GFP-1020, 1:1000), rabbit anti-RedFluorescent Protein (Rockland, cat#600-401-379, 1:1000), mouseanti-microtubule associated protein (Sigma, cat#M1406, 1:500),mouse anti-βIII-TUBULIN (TUBB3; Biolegend, cat#MMS-435P,1:1000), rabbit anti-RBPMS (PhosphoSolutions, cat#1830;1:100), mouse anti-SYNAPSIN 1 (Synaptic Systems, cat#106001,1:2000), mouse anti-PARVALBUMIN (SIGMA, cat#p3088,1:1000), mouse anti-CRALBP (ABCAM, cat# ab15051, 1:500),rabbit anti-GFAP (DakoCytomation, cat#z0334, 1:4000) rabbitanti-SOX2 (ABCAM, cat# ab97959, 1:500), rat anti-BrdU
(ABCAM, cat# ab6326 1:500), rabbit anti-phospho-histone 3(Millipore, cat#06-570, 1:1000), mouse anti-PAX6 (Millipore,cat#MAB5552, 1:500) and mouse anti-NESTIN (Millipore,cat#mab353, 1:200 millipore).
Calcium ImagingCalcium imaging was done on MGC cultures at 2–3 weeks post-nucleofection, using Oregon green 488 BAPTA-1 (Invitrogen,10µM). Imaging was performed in a physiological solutioncontaining 140mMNaCl, 5mMKCl, 2mMMgCl2, 2mMCaCl2,10mM HEPES, 10mM glucose, and 6mM sucrose, and pH 7.35.Images were acquired every 10ms with virtually no intervalsusing a scientific CMOS camera (Andor). The microscope wascontrolled by Micro-Manager software together with the imageprocessor ImageJ. Changes in fluorescence were measured forindividual cells using the time series analyzer plugin v3.0 inImageJ v1.37. The average of the first ten time-lapse images foreach region of interest (ROI) was defined as initial fluorescence(F0), and the variation of fluorescence (1F) in each frame (n) wascalculated as Fn-F0/F0.
Quantitative RT-PCRMGC cultures were harvested at 13 days after nucleofection,and mRNA was isolated from all cells, including non-transfected MGCs. RNA was extracted using RNeasy MiniKit (QIAGEN, CA, USA), which includes a genomic DNAelimination step, and the purity and quantity of total RNAwas estimated using a ND8000 spectrophotometer (ThermoScientific NanoDrop Products, DE, EUA). Extractions werecarried out of cells from each group (Control, Neurog2 or Ascl1),detached chemically with TrypLE and washed with nucleasefree PBS, following the manufacturer’s protocol. The first-strandcDNA was synthetized using the High-Capacity cDNA ReverseTranscription Kit (Applied Biosystems, NY, USA) in accordancewith the manufacturer’s instructions, using 900 ng of extractedRNA per sample. Conditions for each cycle of amplificationwere as follows: 10min at 25◦C; 120min at 37◦C, 5min at85◦C. The final cDNA products were amplified using RT2Real-Timer SyBR Green/ROX PCR Mix (QIAGEN, CA, USA)in 25 µL of a reaction mixture pipetted into each well ofa 96-well in a Mouse RT2 Profiler Custom PCR Array. Thearray was designed to simultaneously examine mRNA levelsof 18 genes commonly expressed in retina cell types (RLBP1,GLUL, NRL, RHO, RCVRN, PDE6G, PROX1, LHX1, VSX2,SLC32A1, TH, CHAT, SLC17A6, POU4F1, CALB2, RBFOX3,SYN1, and PVALB) and 2 housekeeping genes (GAPDH andRPL19), following the manufacturer’s protocol. Real-time PCRwas performed using a two-step cycling program, with an initialsingle cycle of 95◦C for 10min, followed by 40 cycles of 95◦
C for 15 s, then 60◦C for 1min, in an ABI ViiA 7 Real-TimePCR System (Applied Biosystems, NY, USA) with SequenceDetector System software v1.2. The ramp rate was adjusted to1◦C/s following manufacturer’s instruction. A first derivativedissociation curve was built (95◦C for 1min, 65◦C for 2min, thenramped from 65◦C to 95◦C at a rate of 2◦C/min). The formationof a single peak at temperatures higher than 80◦C confirmed thepresence of a single PCR product in the reaction mixture.
Frontiers in Cellular Neuroscience | www.frontiersin.org 3 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
For data analysis the 2(−11Ct) method (Livak and Schmittgen,2001) was implemented using normalized threshold cycle(Ct) values provided by two independent experiments ofnucleofection. Furthermore, we applied experiments usingmRNA pool of two independent transfection experiments andused the Ct data to perform normalization and follow the2(−11Ct) method (Livak and Schmittgen, 2001), consideringthe sensitivity, specificity, and reproducibility expected of real-time PCR using RT2 Profiler PCR Array System from QIAGEN.Endogenous gene control used in the normalization was theaverage of the mouse GAPDH and RPL19. A positive valueindicates gene up-regulation and a negative value indicates genedown-regulation.
To confirm that genes upregulated inMGCs could indicate theacquisition of a retinal neuron-like phenotype in MGC-derivediNs, we performed the same analysis using cerebellum astrogliacell cultures 13 days after nucleofection with either Neurog2 orAscl1 (Chouchane et al., 2017).
BioinformaticsDataset raw count table and published metadata were obtainedfrom GSE63472 accession code. A modified Seurat pipeline wasused to re-analyze single-cell RNAseq data. First, we excludedfrom our analysis genes that were not expressed in at least10 cells. Next, we selected cells expressing 500 to 5,000 genes,and <5% of mitochondrial genes (n = 21,494 cells). Metadatavariables as number of genes and percentage of mitochondrialexpression were also used to regress out some unnecessaryclustering bias. Based on PCElbowPlot, we used 30 PC’s inFindClusters (resolution = 2) and RunTSNE Seurat’s functions.After that, using old assigned clusters and markers found byFindAllMarkers function (Macosko et al., 2015), new assignedclusters were labeled. Retinal cell types (n = 21,176 cells) wereclassified according to the levels of expression of genes in Müllerglia cells, astrocytes, amacrine cells, bipolar cells, horizontal cells,cones, rod cells, and ganglion cells (Macosko et al., 2015). Conesand rod cells were merged in a single-group (Photoreceptors).
QuantificationsTo characterize MGC cultures we examined 20 fields at40× magnification for CRALBP and GFAP, and 20 fieldsat 20× magnification for Nestin, Pax6, and PH3 proteins.To quantify the reprogramming process, cells were examinedfor colocalization of DSRED and TUBB3 immunoreactivityat 13 days post nucleofection (dpn), in 20 fields at 20xmagnification, and the same was done for MAP2 protein. For allprotocols of quantification, we counted immunoreactive cells in3 independent experiments.
To estimate a neuronal polarization index, we divided theneurons into 4 quadrants and measured the axial distributionof neuronal processes. Nineteen neurons were analyzed for theconditionMGCwith EGF/FGF+Neurog2, 31 neurons for MGCwithout+Neurog2, 26 neurons forMGCwith EGF/FGF+Ascl1and 21 neurons for MGC without EGF/FGF+ Ascl1.
Distribution of GFP+ cells in the P10 rat retinas following invivo electroporation at P0 in Lister-hooded pups was examined in26 sections from 5 control-electroporated retinas and 27 sections
from 5 Neurog2-electroporated retinas. The outer nuclear layer(ONL), inner nuclear layer (INL), and ganglion cell layer(GCL) were identified using DAPI counter-staining. MGCs wereidentified by their radial morphology and expression of CRALBP.
Statistical AnalysisAll statistical data are presented as the mean ± standard errorof the mean (SEM) of at least three independent experiments.Statistically significant differences were assessed using unpairedt-test, one-way or two-way analysis of variance (ANOVA).Confidence interval is 95%.
RESULTS
Properties of Cells Expanded in thePresence or Absence of EGF/FGF2Enrichment of MGCs was done according to a previouslydescribed method (Hicks and Courtois, 1990). Using this methodwe (de Melo Reis et al., 2008) and others (Das et al., 2006)have shown that virtually all cells after 2–3 weeks in cultureexpress the MGC markers vimentin, glutamine synthetase(GS), cellular retinaldehyde-binding protein (CRALBP). Basedon the previously reported use of selected growth factors toexpand astroglial populations for lineage reprogramming intoiNs (Berninger et al., 2007; Heinrich et al., 2010; Chouchaneet al., 2017), we cultured cells either with or without EGF/FGF2.After expansion, virtually all cells in cultures obtained using bothprotocols expressed the MGC protein CRALBP (Figures 1A,B).A high percentage of cells also expressed glial acid fibrillaryprotein (GFAP), which is upregulated in MGC cultures (Daset al., 2006). No significant difference was observed between thetwo expansion conditions (Figures 1A,B).
MGCs and late-progenitors in the retina share the expressionof several proteins, including NESTIN, sex determining regionY (SRY)-box 2 (SOX2) and Paired box 6 [PAX6; (Karl et al.,2008; Lin et al., 2009; Bhatia et al., 2011)], what has led to thesuggestion that MGCs may have neural stem cell-like properties(Das et al., 2006; Lawrence et al., 2007; Nickerson et al., 2008;Giannelli et al., 2011). Accordingly, we found that more than97% of cells expressed NESTIN and this percentage was slightlyincreased in the presence of EGF/FGF2 (Figures 1C,D; 97.02 ±
0.95% vs. 99.05 ± 0.32%, p = 0.0466, unpaired t-test. N = 3independent experiments). Similarly, expression of SOX2 wasalso higher amongMGCs expanded in the presence of EGF/FGF2(Figure 1D; 78.71 ± 3.21% vs. 98.20 ± 0.48%, p < 0.0001,unpaired t-test.N = 3 independent experiments). In contrast, theproportion of MGCs expressing PAX6 was higher in the absenceof EGF/FGF2 (Figures 1C,D; 34.87± 2.60% vs. 27.20± 1.58%, p= 0.013, unpaired t-test. N = 3 independent experiments).
To examine the proliferative potential of enriched MGCs,we added the thymidine analog BrdU to the cultures at day20. The percentage of MGCs labeled with BrdU (BrdU+) after36 h of incubation was slightly higher with, rather than withoutEGF/FGF2 (Figures 1C,D; 65.54 ± 2.97% vs. 75.41 ± 3.94%,p = 0.047, unpaired t-test. N = 3 independent experiments).However, the percentage of phospho-histone labeled (PH3+)mitotic MGCs was similar in both conditions (Figure 1D; 8.40
Frontiers in Cellular Neuroscience | www.frontiersin.org 4 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 1 | Characterization of MGC cultures expanded with or without EGF/FGF2. (A) Representative photomicrographs of MGC cultures showing the expression of
CRALBP (green) and GFAP (red) 21 days after expansion in the absence (top) or presence (bottom) of EGF/FGF2. (B) Frequencies of CRALBP and GFAP positive cells
in enriched MGC cultures. (C) Immunolabeling for NESTIN, PAX6, and BRDU (red) in MGCs expanded in the absence (top) or presence (bottom) of EGF/FGF2. (D)
Frequencies of MGCs immunolabeled for NESTIN, SOX2, PAX6, BrdU, and PH3 prior to lineage reprogramming (*p < 0.05; ***p < 0.001; Unpaired t-test). Scale bars:
20 µm.
± 0.72% vs. 7.98 ± 0.69%, p = 0.70, unpaired t-test. N =
3 independent experiments), which could be explained by aselective lengthening of the S-phase upon EGF/FGF2 treatment.Collectively, these observations support the interpretation thatcells enriched in our cultures are presumptive MGCs that retainsome properties observed in late-progenitors of the developingretina. However, we never observed spontaneous neurogenesisin these cultures, suggesting that a potential progenitor stateof presumptive MGCs in culture is associated with a glial-faterestriction.
Expression of Either NEUROG2 or ASCL1Is Sufficient to Convert MGCs Into iNsNext, we tested whether the expression of NEUROG2 mayreprogram enriched MGCs into iNs, as compared with ASCL1as previously described (Pollak et al., 2013). Expanded MGCswere harvested and transfected with control-I-GFP, Neurog2-I-GFP (Figure 2B), Neurog2-I-DsRed (Figures 2D,F) or Ascl1-I-DsRed plasmids (Figures 3A,C), and maintained for 13 days (1day in growth medium + 12 days in differentiation medium;Figure 2A). At the end of this period, cells transfected withcontrol plasmids maintained both their typical glial morphologyand the content of the glia-specific protein GFAP (Figure 2C).In contrast, a substantial fraction of NEUROG2-containingcells underwent robust morphological changes, characterized byreduction of the cell body and extension of thin and long primaryprocesses (usually 2 or 3) with small ramifications, similar tothe morphology of neuronal cells in culture (Figures 2B,D,F).Accordingly, these cells also contained the neuron-specificproteins TUBB3 (Figure 2D) and microtubule associated protein
2 (MAP2) (Figure 2F), indicating that NEUROG2 led toconversion of cultured MGCs into iNs. The frequency of iNs wassignificantly higher in cultures containingMGCs expanded in thepresence, as compared with the absence of EGF and FGF2 (71.0± 4.1 vs. 26.4 ± 5.1%; p < 0.001; Tukey’s multiple comparisontest; Figure 2E), suggesting that these growth factors facilitatereprogramming. Additionally, about 90% of iNs also expressedMAP2 (Figures 2F,G) independently of mitogenic factors duringenrichment, further suggesting the acquisition of a neuronalphenotype.
Lentiviral-mediated expression of ASCL1 reportedly inducedthe conversion of about 30% of cultured MGCs into neuronal-like cells (Pollak et al., 2013). In our model, nucleofection ofASCL1 led more than half of cultured MGCs to convert intoiNs (Figures 3A,B). The proportion of iNs was higher amongMGCs previously expanded in the presence, rather than in theabsence of EGF and FGF2 (68.1 ± 4.9 vs. 53.7 ± 5.5%; p < 0.05;Tukey’s multiple comparison test; Figure 3B), and 97% of iNsalso expressed MAP2 (Figure 3D). Collectively, our data indicatethat the expression of either NEUROG2 or ASCL1 is sufficientto lineage-reprogram MGCs into iNs, as well as that previousexposure of MGCs to EGF and FGF2 facilitates reprogramming.
Functional and MorphologicalDifferentiation of MGC-Derived iNsTo evaluate whether iNs derived from lineage reprogrammedcultured MGCs develop features of mature functional neurons,we performed calcium imaging using a genetically encodedcalcium indicator (GCAMP5) and a fluorescent dye (Oregongreen BAPTA-1). MGC cultures nucleofected with either Ascl1
Frontiers in Cellular Neuroscience | www.frontiersin.org 5 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 2 | Expression of NEUROG2 converts MGCs into iNs. (A) Experimental design. Note that EGF/FGF2 are used only during MGC expansion. (B–B’) Overview
of MGC cultures 13 days post nucleofection (dpn) with Neurog2-I-GFP showing the presence of GFP+ cells (green) adopting a neuronal-like morphology (white inset,
magnified in B’). (C) Representative photomicrographs of MGC cultures at 13 dpn with control-I-GFP, showing GFP+ cells (green) that maintain glial morphology and
contain GFAP (white). (D) Immunolabeling of the neuronal protein βIII-TUBULIN (TUBB3, green) in DsRed (red) cells at 13 dpn with Neurog2-I-DsRed in MGCs cultures
expanded either without (top) or with (bottom) EGF/FGF2. (E) Frequencies of DsRed+/TUBB3+ iNs amongst total DsRed+ cells at 13 dpn (***p < 0.001; Tukey’s
multiple comparison test). (F) Immunolableing for the neuronal protein MAP2 (green) in DsRed (red) cells at 13 dpn with Neurog2-I-DsRed in MGCs expanded either
without or with EGF/FGF2. (G) Frequencies of DsRed+MAP2+ iNs amongst all DsRed+ iNs. Photomicrographs in (C,D,F) are single confocal Z stacks. Nuclei are
stained with DAPI (blue). Scale bars: 20µm.
or Neurog2 expression plasmids were maintained for 15 days,and then treated with Oregon green BAPTA (see Materialsand Methods). Fast fluctuations of intracellular calcium levelsleading to sudden fluorescence changes, likely produced by anabrupt aperture of voltage-gated calcium channels mediated bysynaptic activity (Bonifazi et al., 2009; Yang and Yuste, 2017),were detected inmore than half of iNs (Neurog2 with EGF/FGF2:12/20; Neurog2 without EGF/FGF2: 13/18; and Ascl1 withEGF/FGF2: 11/20; Ascl1 without EGF/FGF: 12/19) (Figure 4
and Supplementary Videos). In contrast, non-transfectedMGCsshowed slow calcium fluctuations (Figure 4).
We also examined the morphology of MGC-derived iNsin the various experimental groups (Figure S1). Expression ofeither Neurog2 or Ascl1 leads to the generation of diverseiNs (Figures S1A,B). Notwithstanding, whereas three-quarters ofNeurog2-converted iNs were either unipolar or bipolar, Ascl1-converted iNs had approximately equal proportions of eithermultipolar or uni/bipolar morphologies (Figure S1C). These
Frontiers in Cellular Neuroscience | www.frontiersin.org 6 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 3 | Expression of ASCL1 converts MGCs into iNs. (A) Representative photomicrographs of MGC cultures expanded without or with EGF/FGF2 at 13 dpn
with Ascl1-I-DsRed, showing DsRed+ cells (red) immunolabeled for βIII-TUBULIN (TUBB3, green) (B) Frequencies of DsRed+/TUBB3+ iNs amongst total DsRed+
cells at 13 dpn (*p < 0.05; ***p < 0.001; Tukey’s multiple comparison test). (C) Immunolabeling of the neuronal protein MAP2 (green) in DsRed (red) cells at 13 dpn
with Ascl1-I-DsRed in MGCs expanded without or with EGF/FGF2. (D) Frequencies of DsRed+MAP2+ iNs amongst all DsRed+ iNs. Photomicrographs in (A,C) are
single confocal Z stacks. Nuclei are stained with DAPI (blue). Scale bars: 20µm.
observations suggest that Neurog2 and Ascl1 iNs may acquiredistinct phenotypes.
MGC-Derived iNs Express Key Genes ofRetinal NeuronsIt has been suggested that the origin of reprogrammedastroglial cell lineages affects the phenotype of induced neurons(Chouchane et al., 2017). We tested whether MGC-derivediNs show hallmarks of bona fide retina neurons, after lineageconversion induced by either ASCL1 or NEUROG2 (Figure 5).First, we took advantage of single-cell transcriptomes availablein the literature (Macosko et al., 2015) to identify molecularsignatures of the major retinal cell types (Figure 5A andFigure S2). Based on single-cell RNA sequence of adult mouseretina cells (Macosko et al., 2015), we defined a small set oftranscripts with enriched expression in MGCs (RLBP1 andGLUL), astrocytes (GFAP), photoreceptors (NRL, RHO, RCVRN,and PDE6G), horizontal cells (PROX1 and LHX1), bipolar cells(VSX2), amacrine cells (PAX6, SLC32A1, GAD1, GAD2, TH,and CHAT) and RGCs (RBPMS, SLC17A6, POU4F1, CALB2,RBFOX3, SYN1, TUBB3, and PVALB). Next, we used qRT-PCR to compare the expression of these molecular markersin MGC cultures expanded in the presence or absence ofEGF/FGF2, and nucleofected with either Neurog2 or Ascl1
(Figures 5B,C). Despite the small number of iNs amongst thetotal population of MGCs (∼1–2%), we detected an increasein the expression of several genes commonly expressed byretina neurons in MGC cultures nucleofected with Neurog2or Ascl1 (Figures 5B, C), but not among cerebellum astroglialcells nucleofected with the same plasmids (Figure S3), which areknown to adopt mostly a GABAergic iN phenotype (Chouchaneet al., 2017). Interestingly, differences were observed among cellsconverted through each of the two expansion protocols. WhileMGC-derived iNs, induced by expression of either ASCL1 andNEUROG2 grown in the absence of EGF/FGF2 upregulated keygenes of photoreceptors, genes of amacrine cells were expressedonly in iNs derived from MGCs expanded in the presence ofgrowth factors. In contrast, increased levels of genes commonlyexpressed by horizontal and bipolar cells were detected inall experimental conditions. Surprisingly, we also found thatmany genes commonly expressed by RGCs were upregulatedin MGC-derived iNs, in particular the specific RGC genesSLC17A6 and POU4F1 (Quina et al., 2005; Martersteck et al.,2017), but the same was not observed in cerebellar astroglia-derived iNs (Figure S3). This effect was much more robustafter expression of NEUROG2 (Figure 5B), independently ofthe expansion protocol. Altogether, the transcriptome panelsuggests that either Neurog2 or Ascl1 may induce distinct, but
Frontiers in Cellular Neuroscience | www.frontiersin.org 7 November 2018 | Volume 12 | Article 410
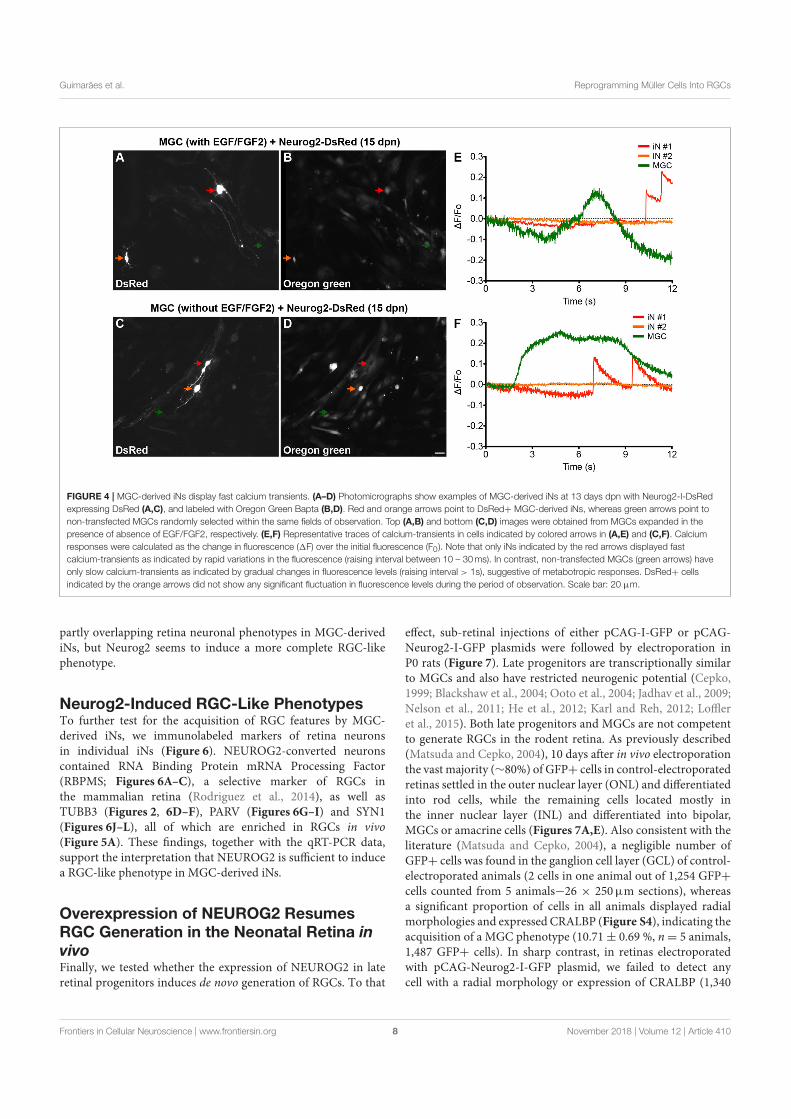
Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 4 | MGC-derived iNs display fast calcium transients. (A–D) Photomicrographs show examples of MGC-derived iNs at 13 days dpn with Neurog2-I-DsRed
expressing DsRed (A,C), and labeled with Oregon Green Bapta (B,D). Red and orange arrows point to DsRed+ MGC-derived iNs, whereas green arrows point to
non-transfected MGCs randomly selected within the same fields of observation. Top (A,B) and bottom (C,D) images were obtained from MGCs expanded in the
presence of absence of EGF/FGF2, respectively. (E,F) Representative traces of calcium-transients in cells indicated by colored arrows in (A,E) and (C,F). Calcium
responses were calculated as the change in fluorescence (1F) over the initial fluorescence (F0). Note that only iNs indicated by the red arrows displayed fast
calcium-transients as indicated by rapid variations in the fluorescence (raising interval between 10 – 30ms). In contrast, non-transfected MGCs (green arrows) have
only slow calcium-transients as indicated by gradual changes in fluorescence levels (raising interval > 1s), suggestive of metabotropic responses. DsRed+ cells
indicated by the orange arrows did not show any significant fluctuation in fluorescence levels during the period of observation. Scale bar: 20 µm.
partly overlapping retina neuronal phenotypes in MGC-derivediNs, but Neurog2 seems to induce a more complete RGC-likephenotype.
Neurog2-Induced RGC-Like PhenotypesTo further test for the acquisition of RGC features by MGC-derived iNs, we immunolabeled markers of retina neuronsin individual iNs (Figure 6). NEUROG2-converted neuronscontained RNA Binding Protein mRNA Processing Factor(RBPMS; Figures 6A–C), a selective marker of RGCs inthe mammalian retina (Rodriguez et al., 2014), as well asTUBB3 (Figures 2, 6D–F), PARV (Figures 6G–I) and SYN1(Figures 6J–L), all of which are enriched in RGCs in vivo(Figure 5A). These findings, together with the qRT-PCR data,support the interpretation that NEUROG2 is sufficient to inducea RGC-like phenotype in MGC-derived iNs.
Overexpression of NEUROG2 ResumesRGC Generation in the Neonatal Retina in
vivoFinally, we tested whether the expression of NEUROG2 in lateretinal progenitors induces de novo generation of RGCs. To that
effect, sub-retinal injections of either pCAG-I-GFP or pCAG-Neurog2-I-GFP plasmids were followed by electroporation inP0 rats (Figure 7). Late progenitors are transcriptionally similarto MGCs and also have restricted neurogenic potential (Cepko,1999; Blackshaw et al., 2004; Ooto et al., 2004; Jadhav et al., 2009;Nelson et al., 2011; He et al., 2012; Karl and Reh, 2012; Loffleret al., 2015). Both late progenitors and MGCs are not competentto generate RGCs in the rodent retina. As previously described(Matsuda and Cepko, 2004), 10 days after in vivo electroporationthe vast majority (∼80%) of GFP+ cells in control-electroporatedretinas settled in the outer nuclear layer (ONL) and differentiatedinto rod cells, while the remaining cells located mostly inthe inner nuclear layer (INL) and differentiated into bipolar,MGCs or amacrine cells (Figures 7A,E). Also consistent with theliterature (Matsuda and Cepko, 2004), a negligible number ofGFP+ cells was found in the ganglion cell layer (GCL) of control-electroporated animals (2 cells in one animal out of 1,254 GFP+cells counted from 5 animals−26 × 250µm sections), whereasa significant proportion of cells in all animals displayed radialmorphologies and expressed CRALBP (Figure S4), indicating theacquisition of a MGC phenotype (10.71± 0.69 %, n= 5 animals,1,487 GFP+ cells). In sharp contrast, in retinas electroporatedwith pCAG-Neurog2-I-GFP plasmid, we failed to detect anycell with a radial morphology or expression of CRALBP (1,340
Frontiers in Cellular Neuroscience | www.frontiersin.org 8 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 5 | Expression of either NEUROG2 or ASCL1 induces upregulation of key retina neuron genes in MGC-derived iNs. (A) Expression levels of selected genes
used to identify different classes of retina neurons using scRNAseq data available in the literature (Macosko et al., 2015) (see also Figure S2). Dot sizes and colors
(green-low to red-high) represent the percentage of cells expressing the gene and its average scaled expression, respectively. Observe that the expression of Slc17a6
and Pou4f1 are specific for a high percentage of cells assigned as RGCs. (B,C) Graphics showing the relative expression levels (log10) of genes enriched in Müller glia
(Rlbp1, Glul), photoreceptors (Rho, Rcvrn, Pde6g), horizontal (Prox1, Lhx1), bipolar (Vsx2), amacrine (Slc32a1, Chat, Th) and retina ganglion cells (Pou4f1, Slc17a6,
Calb2, Rbfox3, Syn1, Pvalb, Rbpms), as shown in (A), in MGC cultures nucleofected with either Neurog2 (B) or Ascl1 (C). White and black bars indicate that MGC
were expanded in the absence or presence of EGF/FGF2, respectively, prior to the expression of Neurog2 or Ascl1.
GFP+ cells analyzed from 5 animals). About 65.5% of GFP+cells settled in the ONL and 33% in the INL, but a significantfraction (17 cells in 5 animals out of 1340 GFP+ cells countedfrom 5 animals−27 × 250µm sections) of cells was found inthe GCL (Figures 7C–I). Quantification of the number of GFP+cells in 250µm of 27 sections of 5 retinas showed that the meannumber of GFP+ cells in the GCL was significantly increasedin Neurog2-electroporated animals as compared to controls,suggesting that NEUROG2 expression resumed the generation ofRGCs (Figure 7F). Accordingly, we also found that GFP+ cellsin the GCL expressed classical markers of RGCs, such as RBPMS(Figures 7 G–I) and TUBB3 (Figure S5) and extended thinaxonal processes that reached the optic nerve (Figure 7J). SomeGFP-expressing cells within the GCL also extended processestoward the inner plexiform layer, consistent with the morphologyof RGCs (Figures 7G, 8). Moreover, GFP+ cells within theGCL also displayed varied patterns of dendrite stratification(Figure 8), consistent with distinct RGC morphologies in therodent retina (Sanes and Masland, 2015). Altogether, these datasupport the hypothesis that the expression of NEUROG2 endows
late retinal progenitors with the potential to generate RGCs, whileit inhibits the differentiation of MGCs in vivo.
DISCUSSION
Here we show that cells isolated in culture and expressingseveral hallmarks of Müller cells, the main type of glia foundin the vertebrate retina, can be reprogrammed into neuronswhen transduced with plasmids encoding either of the basicHelix loop Helix (bHLH) transcription factors Neurog2 orAscl1. Cell–lineage reprogramming is affected by treatmentwith EGF and FGF2 during MGC enrichment, and led to theexpression of typical neuronal markers, MAP-2 and TUBB3.Induced neurons expressed clusters of genes consistent with theprofile of retinal cells, suggesting that distinct retinal neuronphenotypes are elicited in MGC-derived iNs. Notably, only iNsgenerated after expression of NEUROG2 upregulated a set ofgenes compatible with the acquisition of a possible RGC identity.A role for NEUROG2 in the induction of RGC fate was also
Frontiers in Cellular Neuroscience | www.frontiersin.org 9 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 6 | Expression of NEUROG2 induces the acquisition of RGC-like phenotypes in MGC-derived iNs. (A–L) Immunolabeling of the RGC proteins RBPMS (red,
A–C), βIII-TUBULIN (green, D–F), PARV (red, G–I) and SYN1 (green, J–L) in MGC converted into iNs at 15 days post-nucleofection with either Neurog2-I-GFP (green,
A–C and G–I) or Neurog2-I-DsRed (red, D-F and J-L). Scale bars: 20µm.
observed in late progenitors of the neonatal mouse retina invivo, suggesting that Neurog2 is a good candidate for genetherapies aiming at the regeneration of RGCs in the adult injuredretina.
Lineage reprogramming of astroglial cells into neurons hasbeen heralded as a possible treatment for degenerative ortraumatic brain disorders, and recent studies showed evidenceof conversion of glial cells into functional neurons with high
efficiency in the murine brain, induced by virally deliveredtranscription factors (Torper and Gotz, 2017). Astroglial cellsisolated from the postnatal rodent cerebral cortex (Berningeret al., 2007; Heinrich et al., 2010) and cerebellum (Chouchaneet al., 2017), as well as retinal MGCs (Singhal et al., 2012;Pollak et al., 2013; Song et al., 2013; Ueki et al., 2015; Wuet al., 2016; Jorstad et al., 2017) have been reprogrammed intoneurons.
Frontiers in Cellular Neuroscience | www.frontiersin.org 10 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 7 | Forced-expression of NEUROG2 resumes the generation of RGCs in the developing retina in vivo. (A–D) Representative images of P10 rat retinas after
electroporation with control-I-GFP (A,B) and Neurog2-I-GFP (C,D) plasmids at P0. Note the presence of GFP+ radial fibers typical of MGCs (arrows in B) and the
complete absence of GFP+ cells in the GCL in controls. Observe also the decrease of GFP+ cells in the ONL and the presence of GFP+ cells in the GCL of
Neurog2-electroporated retina (arrow in D). (E) Frequencies of GFP+ cells in the ONL, INL e GCL of the retina of control- and Neurog2-electroporated animals. (F)
Mean numbers of GFP+ cells within the ONL, INL or GCL per 250µm longitudinal section (***p < 0.001; Two-way ANOVA, Bonferroni post-hoc test). (G–I) Expression
of RBPMS (red) in GFP+ cells (green) within the GCL of Neurog2-electroporated retinas. White arrow points to the axon-like process of the GFP+ cell observed within
the GCL. Dashed box (G) is amplified in H and I to show the co-localization of GFP and RBPMS in a single confocal Z-stack. (J) Axon-like GFP+ processes (arrows)
reaching the optic nerve in a Neurog2-electroporated retina. Nuclei are stained with DAPI (blue). Scale bars: A-D and J: 50µm; G: 25µm; H-I: 12,5µm.
Lentiviral gene transfer of the proneural factor Ascl1 partiallyreprogrammed P11/12 Müller glial cells in vitro into retinalprogenitors 3 weeks after infection (Pollak et al., 2013), a processfacilitated by microRNA miR-124-9-9∗ (Wohl and Reh, 2016).Combination of Ascl1 and miR-124-9-9∗ led to a peak of 50–60% reprogrammed iNs, whereas Ascl1 alone reached 30–35%(Wohl and Reh, 2016). The latter is lower than what we foundfollowing expression of Ascl1, which may be due to the earlierage of the mice from which cells were isolated in our study(P7-9). Indeed, in our hands, MGCs isolated from P21 retina
failed to reprogram into iNs upon expression of either ASCL1or NEUROG2 (data not shown), similar to what has beendescribed for astroglia isolated from the adult mouse cerebralcortex (Heinrich et al., 2010) or even for astroglia isolated fromthe postnatal cerebral cortex or cerebellum, and maintainedin differentiation conditions for several days before ASCL1 orNEUROG2 expression (Masserdotti et al., 2015; Chouchane et al.,2017). This resistance to lineage conversion in astrocytes isolatedat late developmental stages is likely explained by the epigeneticchanges occurring in these cells during differentiation from an
Frontiers in Cellular Neuroscience | www.frontiersin.org 11 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
FIGURE 8 | Representative examples of GFP+ cells within the GCL of Neurog2-electroporated retinas. (A–H) Images show GFP+ cells (green) within the GCL of P10
rats after electroporation with Neurog2-I-GFP at P0. Nuclei are stained with DAPI (blue, A–D). Note the distinct positions and dendrite stratifications of GFP+ cells
(E-H), resembling bona fide subtypes of rodent RGCs (Sanes and Masland, 2015). Scale bar: 25µm.
immature to mature state (Masserdotti et al., 2015). According tothis interpretation, ASCL1-mediated lineage reprogramming ofMGCs into iNs in adult mouse retina requires co-treatment withhistone deacetylase-inhibitors (Jorstad et al., 2017).
Our data also showed that the presence of mitogenic factorsEGF and FGF2 during MGC expansion substantially increasedthe efficiency of conversion into iNs, suggesting that exposureto those growth factors endows MGCs with higher plasticity.It has been shown that FGF2 induces methylation of Lysine4 and suppresses methylation of Lysine 9 of histone H3 atthe signal transducer and activator of transcription (STAT)binding site (Song and Ghosh, 2004), whereas EGF affectschromatin architecture at the regulatory element of cyclinD1, through a process involving Cre-binding protein (CBP),Histone deacetylase 1 (HDAC1) and Suv39h1 histone/chromatinremodeling complex (Lee et al., 2012). It is conceivable thatsuch effects of EGF/FGF2 upon MGCs may facilitate the bindingof NEUROG2 and ASCL1 to their target genes and, therefore,increases the efficiency of conversion. The effect was morepronounced for NEUROG2 than ASCL1; the latter has a knownrole in the remodeling of the chromatin landscape by itself,which increases its own accessibility to target genes (Raposoet al., 2015). In addition, expression of ASCL1 in NMDA-injuredretinas of adult mice was sufficient to lineage reprogram onlyabout 20% of MGCs into Otx2+ neurons, but this effect wasenhanced by 2-fold in animals receiving concomitant treatmentwith trichostatin-A, a potent inhibitor of histone deacetylase(Jorstad et al., 2017).
Alternative explanations for the effects of EGF/FGF2 uponMGC-lineage conversion into iNs are rooted on the roles
of these growth factors in cell proliferation (Todd et al.,2015) and survival (Rolf et al., 2003; Nickerson et al., 2011),dedifferentiation and possibly reprogramming, reviewed inHamon et al. (2016). Recent work in the adult mouse retinademonstrates that genetic lineage reprogramming of MGCsinto photoreceptors in vivo requires previous activation ofcell proliferation trough activation of the beta-catenin pathway(Yao et al., 2018). Here, we observed a larger fraction ofNESTIN+, SOX2+, and BrdU+ cells in MGC cultures grownin the presence of EGF/FGF2, suggesting an increase in cellproliferation. However, this effect was much smaller than thatobserved in lineage-reprogramming efficiencies, indicating thatthe increased number of iNs observed in cultures treated withEGF/FGF2 cannot be exclusively explained by an increase inMGC proliferation. Thus, the explanation that reprogrammingis facilitated by EGF/FGF2 through effects upon chromatinstructure remains a likely explanation, although further studiesare needed to support this hypothesis.
A critical question in cell reprogramming is whether iNsacquire either single or multiple neuronal phenotypes (Amamotoand Arlotta, 2014; Heinrich et al., 2015). Recent work in ourlaboratory has shown that the origin of astroglial cells, eitherfrom cerebral cortex or cerebellum, affects the phenotype of iNslineage-converted by either ASCL1 or NEUROG2 (Chouchaneet al., 2017). Notably, most iNs generated from astroglia fromeither cerebellum or neocortex showed central hallmarks ofneurons commonly observed in those regions, suggesting thata “molecular memory” in the astroglial cells contributes to theacquisition of specific neuronal phenotypes in iNs (Chouchaneand Costa, 2018).
Frontiers in Cellular Neuroscience | www.frontiersin.org 12 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
This notion is supported by our current finding that MGC-derived iNs upregulate several genes expressed in retinal neurons.Consistent with previous work (Pollak et al., 2013;Wohl and Reh,2016; Jorstad et al., 2017), we found that ASCL1-convertedMGCsgenerated iNs which express genes of horizontal and bipolar cells,but few genes commonly observed in photoreceptors, amacrinecells and RGCs, suggesting that these phenotypes are rare and/orincomplete in the converted cells. In contrast, expression ofNEUROG2 converted MGCs into iNs accompanied by up-regulation of several genes of photoreceptors, amacrine cellsand RGCs. Importantly, the latter phenotype is supported bythe expression of two RGC-specific genes, namely Pou4f1 andScl17a6 (Quina et al., 2005; Martersteck et al., 2017), as well asfour other genes highly enriched in RGCs (Syn1, Parv, Calb2and Tubb3). Immunolabeling confirmed the increased content ofthree of these markers (TUBB3, SYNAPSIN 1, PARVALBUMIN).Moreover, expression of the selective RGC marker RBPMS(Rodriguez et al., 2014) in MGC nucleofected with Neurog2further suggests the acquisition of a RGC-like phenotype in iNs.These observations are particularly important, because previousattempts to lineage reprogram MGCs into iNs, both in vitro andin vivo, have failed to generate iNs with RGC-like phenotypes(Pollak et al., 2013; Wohl and Reh, 2016; Jorstad et al., 2017).Future experiments should address whether the expression ofNEUROG2 in MGCs in the intact or injured adult retina aresufficient to lineage convert these cells into RGCs. Nevertheless,our observations in post-natal retinas indicate that NEUROG2has an important instructive role for the RGC phenotype in vivo.
Altogether, our study corroborates previous evidence of thepotential of MGC to reprogram into iNs following expressionof ASCL1 (Pollak et al., 2013; Wohl and Reh, 2016; Jorstadet al., 2017), and provides compelling evidence that NEUROG2expression is also sufficient to convert MGCs into iNs expressingseveral features of RGCs. Our results may thus contributeto develop new strategies of gene therapy aiming at theregeneration of retinal neurons in patients with glaucoma andother neurodegenerative retinopathies.
AUTHOR CONTRIBUTIONS
All authors reviewed the manuscript. RG and BL contributedto design and perform the experiments, analyzed the data,discussed the results, and prepare figures of the manuscript. DG.performed qPCR experiments. DC analyzed single cell RNAseqdata. MS conceived electroporation experiments, discussed theresults, and contributed to the manuscript. RdMR and RL co-supervised RG’s thesis, and contributed to the manuscript. MCprovided financial support, directed the project, conceived theexperiments, analyzed data, discussed the results, and wrote themanuscript.
ACKNOWLEDGMENTS
We would like to thank Viviane Valença and Pedro LucasFrança for their contribution with in vivo electroporation
experiments. This work was supported by Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq),Coordenação de Aperfeiçoamento de Pessoal de Nível Superior(CAPES), Fundação de Amparo à pesquisa do Rio de Janeiro(FAPERJ) and Instituto Nacional de Neurociência Translacional(INNT).
SUPPLEMENTARY MATERIAL
The Supplementary Material for this article can be foundonline at: https://www.frontiersin.org/articles/10.3389/fncel.2018.00410/full#supplementary-material
Figure S1 | Polarized morphologies are more common in Neurog2- than
Ascl1-iNs. (A) Morphologies of iNs derived from MGCs nucleofected with either
Neurog2 or Ascl1. (B) Cartesian plot representing the orientations of primary
dendrites in iNs generated in different conditions (Neurog2 without GF = 31 iNs;
Neurog2 with GF = 19 iNs; Ascl1 without GF = 21 iNs; Ascl1 with GF = 26 iNs).
(C) Frequencies of iNs showing unipolar, bipolar, or multipolar morphologies
according to the TF used (Neurog2= 50 iNs; Asc1= 47 iNs). Observe the higher
number of uni/bipolar among Neurog2-induced neurons (p < 0.05; Chi-square
test).
Figure S2 | Identification of major retina cell types using single-cell
transcriptomes. (A) Clustering of 21,176 single-cell expression profiles into 39
retinal cell populations using available dataset (Macosko et al., 2015). The plot
shows a two-dimensional representation (tSNE) of global gene expression
relationships among 21,176 cells. (B) Annotation of the same cells into the 7
major retinal cell types, according to the expression of cell-type specific genes. (C)
List of some differently expressed genes used to classify the 7 major retina cell
types and also used to identify the phenotypes of iNs in this study (RT-qPCR and
immunocytochemistry).
Figure S3 | Gene expression in cerebellar astroglia cells nucleofected with
Neurog2 or Asc1. Graphic showing the relative expression levels (log10) of genes
used to identify presumptive retina cell phenotypes (Figure 5) cerebellar astroglia
cell cultures nucleofected with either Neurog2 (white bars) or Ascl1 (black bars).
Observe that genes commonly observed in cerebellar neurons (Prox1, Vsx2,
Slc32a1, Chat, Rbfox3, and Syn1) are upregulated, whereas genes whose
expression is restricted to retinal neurons (Nrl, Rho, Pou4f1, Slc17a6) are not up
regulated in cerebellar astroglia-derived iNs.
Figure S4 | Expression of CRALBP in MGCs generated in the postnatal retina
electroporated with control-I-GFP. (A–C) Coronal section of a P10 rat retina after
electroporation with Control-I-GFP at P0, immunolabeled for GFP (green) and
CRALBP (red). Images are single confocal Z-stacks and show the co-localization
of GFP and CRALB in MGC fibers (arrows). Scale bar: 25µm.
Figure S5 | Expression of βIII-TUBULIN in RGCs generated in the postnatal retina
following Neurog2-electroporation. (A) Coronal section of a P10 rat retina after
electroporation with Neurog2-I-GFP at P0, immunolabeled for GFP (green) and
βIII-TUBULIN (TUBB3, red). Nuclei are stained with DAPI (blue). Image is a
Z-projection of 8 confocal Z-stacks. Dashed box delimits a GFP+ cell within the
ganglion cell layer (GCL). (B,C) Magnification of the dashed box in A showing the
co-localization of GFP and βIII-TUBULIN in a single confocal Z-stack. Scale bars:
A: 50µm; B,C: 25µm.
Supplementary Video 1 | MGC expanded in the presence of EGF/FGF2 and
lineage reprogrammed into iNs by NEUROG2 show fast calcium transients. Movie
shows 600 frames taken with 10ms exposure time and no interval. Observe the
fast fluorescence intensity increase in the MGC-derived iN indicated by a red arrow
in Figures 4A,B. MGCs in the same field show slow oscillations in fluorescence.
Supplementary Video 2 | MGC expanded in the absence of EGF/FGF2 and
lineage reprogrammed into iNs by NEUROG2 show fast calcium transients. Movie
shows 600 frames taken with 10ms exposure time and no interval. Observe the
fast fluorescence intensity increase in the MGC-derived iN indicated by a red arrow
in Figures 4C,D. MGCs in the same field show slow oscillations in fluorescence.
Frontiers in Cellular Neuroscience | www.frontiersin.org 13 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
REFERENCES
Abu-Hassan, D.W., Li, X., Ryan, E. I., Acott, T. S., and Kelley, M. J. (2015). Induced
pluripotent stem cells restore function in a human cell loss model of open-angle
glaucoma. Stem Cells 33, 751–761. doi: 10.1002/stem.1885
Amamoto, R., and Arlotta, P. (2014). Development-inspired reprogramming
of the mammalian central nervous system. Science 343:1239882.
doi: 10.1126/science.1239882
Athanasiou, D., Aguila, M., Bevilacqua, D., Novoselov, S. S., Parfitt, D. A., and
Cheetham, M. E. (2013). The cell stress machinery and retinal degeneration.
FEBS Lett. 587, 2008–2017. doi: 10.1016/j.febslet.2013.05.020
Berninger, B., Costa, M. R., Koch, U., Schroeder, T., Sutor, B., Grothe,
B., et al. (2007). Functional properties of neurons derived from in
vitro reprogrammed postnatal astroglia. J Neurosci 27, 8654–8664.
doi: 10.1523/JNEUROSCI.1615-07.2007
Bhatia, B., Singhal, S., Tadman, D. N., Khaw, P. T., and Limb, G. A.
(2011). SOX2 is required for adult human muller stem cell survival and
maintenance of progenicity in vitro. Invest. Ophthalmol. Vis. Sci. 52, 136–145.
doi: 10.1167/iovs.10-5208
Blackshaw, S., Harpavat, S., Trimarchi, J., Cai, L., Huang, H., Kuo, W. P., et al.
(2004). Genomic analysis of mouse retinal development. PLoS Biol. 2:E247.
doi: 10.1371/journal.pbio.0020247
Bonifazi, P., Goldin, M., Picardo, M. A., Jorquera, I., Cattani, A., Bianconi, G.,
et al. (2009). GABAergic hub neurons orchestrate synchrony in developing
hippocampal networks. Science 326, 1419–1424. doi: 10.1126/science.1175509
Brzezinski, J. A. T., Kim, E. J., Johnson, J. E., and Reh, T. A. (2011). Ascl1
expression defines a subpopulation of lineage-restricted progenitors in the
mammalian retina. Development 138, 3519–3531. doi: 10.1242/dev.064006
Cepko, C. L. (1999). The roles of intrinsic and extrinsic cues and bHLH genes
in the determination of retinal cell fates. Curr. Opin. Neurobiol. 9, 37–46.
doi: 10.1016/S0959-4388(99)80005-1
Chamling, X., Sluch, V. M., and Zack, D. J. (2016). The potential of human stem
cells for the study and treatment of glaucoma. Invest. Ophthalmol. Vis. Sci. 57,
ORSFi1-6. doi: 10.1167/iovs.15-18590
Chouchane, M., and Costa, M. R. (2018). Instructing neuronal identity during CNS
development and astroglial-lineage reprogramming: roles of NEUROG2 and
ASCL1. Brain Res. doi: 10.1016/j.brainres.2018.02.045. [Epub ahead of print].
Chouchane,M., Melo De Farias, A. R., Moura, D.M. S., Hilscher, M.M., Schroeder,
T., Leao, R. N., et al. (2017). Lineage reprogramming of astroglial cells from
different origins into distinct neuronal subtypes. Stem Cell Rep. 9, 162–176.
doi: 10.1016/j.stemcr.2017.05.009
Das, A. V., Mallya, K. B., Zhao, X., Ahmad, F., Bhattacharya, S., Thoreson, W.
B., et al. (2006). Neural stem cell properties of Muller glia in the mammalian
retina: regulation by Notch and Wnt signaling. Dev. Biol. 299, 283–302.
doi: 10.1016/j.ydbio.2006.07.029
de Melo Reis, R. A., Cabral-Da-Silva, M., De Mello, F. G., and Taylor, J. S. (2008).
Muller glia factors induce survival and neuritogenesis of peripheral and central
neurons. Brain Res. 1205, 1–11. doi: 10.1016/j.brainres.2008.02.035
de Melo, J., and Blackshaw, S. (2018). In vivo electroporation of developing mouse
retina.Methods Mol. Biol. 1715, 101–111. doi: 10.1007/978-1-4939-7522-8_8
Giannelli, S. G., Demontis, G. C., Pertile, G., Rama, P., and Broccoli, V.
(2011). Adult human Muller glia cells are a highly efficient source of rod
photoreceptors. Stem Cells 29, 344–356. doi: 10.1002/stem.579
Gill, K. P., Hung, S. S., Sharov, A., Lo, C. Y., Needham, K., Lidgerwood, G. E., et al.
(2016). Enriched retinal ganglion cells derived from human embryonic stem
cells. Sci. Rep. 6:30552. doi: 10.1038/srep30552
Hamon, A., Roger, J. E., Yang, X. J., and Perron, M. (2016). Muller
glial cell-dependent regeneration of the neural retina: an overview across
vertebrate model systems. Dev. Dyn. 245, 727–738. doi: 10.1002/dvdy.
24375
He, J., Zhang, G., Almeida, A. D., Cayouette, M., Simons, B. D., and Harris, W.
A. (2012). How variable clones build an invariant retina. Neuron 75, 786–798.
doi: 10.1016/j.neuron.2012.06.033
Heinrich, C., Blum, R., Gascon, S., Masserdotti, G., Tripathi, P., Sanchez, R.,
et al. (2010). Directing astroglia from the cerebral cortex into subtype specific
functional neurons. PLoS Biol. 8:e1000373. doi: 10.1371/journal.pbio.1000373
Heinrich, C., Spagnoli, F. M., and Berninger, B. (2015). In vivo reprogramming for
tissue repair. Nat. Cell Biol. 17, 204–211. doi: 10.1038/ncb3108
Hicks, D., and Courtois, Y. (1990). The growth and behaviour of rat retinal Muller
cells in vitro. 1. An improved method for isolation and culture. Exp. Eye Res.
51, 119–129. doi: 10.1016/0014-4835(90)90063-Z
Hufnagel, R. B., Le, T. T., Riesenberg, A. L., and Brown, N. L. (2010). Neurog2
controls the leading edge of neurogenesis in the mammalian retina. Dev. Biol.
340, 490–503. doi: 10.1016/j.ydbio.2010.02.002
Jadhav, A. P., Roesch, K., and Cepko, C. L. (2009). Development and neurogenic
potential of Muller glial cells in the vertebrate retina. Prog. Retin. Eye Res. 28,
249–262. doi: 10.1016/j.preteyeres.2009.05.002
Jorstad, N. L., Wilken, M. S., Grimes, W. N., Wohl, S. G., Vandenbosch, L. S.,
Yoshimatsu, T., et al. (2017). Stimulation of functional neuronal regeneration
fromMuller glia in adult mice. Nature 548, 103–107. doi: 10.1038/nature23283
Karl, M. O., Hayes, S., Nelson, B. R., Tan, K., Buckingham, B., and Reh, T. A. (2008).
Stimulation of neural regeneration in the mouse retina. Proc. Natl. Acad. Sci.
U.S.A. 105, 19508–19513. doi: 10.1073/pnas.0807453105
Karl, M. O., and Reh, T. A. (2012). Studying the generation of regenerated retinal
neuron from Muller glia in the mouse eye. Methods Mol. Biol. 884, 213–227.
doi: 10.1007/978-1-61779-848-1_15
Kimura, A., Namekata, K., Guo, X., Noro, T., Harada, C., and Harada, T. (2017).
Targeting oxidative stress for treatment of glaucoma and optic neuritis. Oxid.
Med. Cell Longev. 2017:2817252. doi: 10.1155/2017/2817252
Lawrence, J. M., Singhal, S., Bhatia, B., Keegan, D. J., Reh, T. A., Luthert, P. J.,
et al. (2007). MIO-M1 cells and similar muller glial cell lines derived from adult
human retina exhibit neural stem cell characteristics. Stem Cells 25, 2033–2043.
doi: 10.1634/stemcells.2006-0724
Lee, J. H., Choy, M. L., and Marks, P. A. (2012). Mechanisms of
resistance to histone deacetylase inhibitors. Adv. Cancer Res. 116, 39–86.
doi: 10.1016/B978-0-12-394387-3.00002-1
Liang, Y. B., Zhang, Y., Musch, D. C., and Congdon, N. (2017).
Proposing new indicators for glaucoma healthcare service. Eye Vis. 4:6.
doi: 10.1186/s40662-017-0071-0
Lin, Y. P., Ouchi, Y., Satoh, S., and Watanabe, S. (2009). Sox2 plays a role in the
induction of amacrine and Muller glial cells in mouse retinal progenitor cells.
Invest. Ophthalmol. Vis. Sci. 50, 68–74. doi: 10.1167/iovs.07-1619
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method.
Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Loffler, K., Schafer, P., Volkner, M., Holdt, T., and Karl, M. O. (2015). Age-
dependent Muller glia neurogenic competence in the mouse retina. Glia 63,
1809–1824. doi: 10.1002/glia.22846
Macosko, E. Z., Basu, A., Satija, R., Nemesh, J., Shekhar, K., Goldman, M., et al.
(2015). Highly parallel genome-wide expression profiling of individual cells
using nanoliter droplets. Cell 161, 1202–1214. doi: 10.1016/j.cell.2015.05.002
Martersteck, E. M., Hirokawa, K. E., Evarts, M., Bernard, A., Duan, X., Li, Y., et al.
(2017). Diverse central projection patterns of retinal ganglion cells. Cell Rep. 18,
2058–2072. doi: 10.1016/j.celrep.2017.01.075
Masserdotti, G., Gillotin, S., Sutor, B., Drechsel, D., Irmler, M., Jorgensen, H. F.,
et al. (2015). Transcriptional mechanisms of proneural factors and REST in
regulating neuronal reprogramming of astrocytes. Cell Stem Cell 17, 74–88.
doi: 10.1016/j.stem.2015.05.014
Matsuda, T., and Cepko, C. L. (2004). Electroporation and RNA interference in
the rodent retina in vivo and in vitro. Proc. Natl. Acad. Sci. U.S.A. 101, 16–22.
doi: 10.1073/pnas.2235688100
Matsuda, T., and Cepko, C. L. (2007). Controlled expression of transgenes
introduced by in vivo electroporation. Proc. Natl. Acad. Sci. U.S.A. 104,
1027–1032. doi: 10.1073/pnas.0610155104
Nelson, B. R., Ueki, Y., Reardon, S., Karl, M. O., Georgi, S., Hartman, B. H.,
et al. (2011). Genome-wide analysis of Muller glial differentiation reveals a
requirement for Notch signaling in postmitotic cells to maintain the glial fate.
PLoS ONE 6:e22817. doi: 10.1371/journal.pone.0022817
Nickerson, P. E., Da Silva, N., Myers, T., Stevens, K., and Clarke, D. B.
(2008). Neural progenitor potential in cultured Muller glia: effects of
passaging and exogenous growth factor exposure. Brain Res. 1230, 1–12.
doi: 10.1016/j.brainres.2008.03.095
Nickerson, P. E., Mcleod, M. C., Myers, T., and Clarke, D. B. (2011). Effects of
epidermal growth factor and erythropoietin on Muller glial activation and
phenotypic plasticity in the adult mammalian retina. J. Neurosci. Res. 89,
1018–1030. doi: 10.1002/jnr.22629
Frontiers in Cellular Neuroscience | www.frontiersin.org 14 November 2018 | Volume 12 | Article 410

Guimarães et al. Reprogramming Müller Cells Into RGCs
Ooto, S., Akagi, T., Kageyama, R., Akita, J., Mandai, M., Honda, Y., et al.
(2004). Potential for neural regeneration after neurotoxic injury in the
adult mammalian retina. Proc. Natl. Acad. Sci. U.S.A. 101, 13654–13659.
doi: 10.1073/pnas.0402129101
Pollak, J., Wilken, M. S., Ueki, Y., Cox, K. E., Sullivan, J. M., Taylor, R. J.,
et al. (2013). ASCL1 reprograms mouse Muller glia into neurogenic retinal
progenitors. Development 140, 2619–2631. doi: 10.1242/dev.091355
Quina, L. A., Pak, W., Lanier, J., Banwait, P., Gratwick, K., Liu, Y.,
et al. (2005). Brn3a-expressing retinal ganglion cells project specifically to
thalamocortical and collicular visual pathways. J. Neurosci. 25, 11595–11604.
doi: 10.1523/JNEUROSCI.2837-05.2005
Rapaport, D. H.,Wong, L. L.,Wood, E. D., Yasumura, D., and Lavail, M.M. (2004).
Timing and topography of cell genesis in the rat retina. J. Comp. Neurol. 474,
304–324. doi: 10.1002/cne.20134
Raposo, A. A., Vasconcelos, F. F., Drechsel, D., Marie, C., Johnston, C.,
Dolle, D., et al. (2015). Ascl1 coordinately regulates gene expression and
the chromatin landscape during neurogenesis. Cell Rep. 10, 1544–1556.
doi: 10.1016/j.celrep.2015.02.025
Rodriguez, A. R., De Sevilla Muller, L. P., and Brecha, N. C. (2014). The
RNA binding protein RBPMS is a selective marker of ganglion cells in
the mammalian retina. J. Comp. Neurol. 522, 1411–1443. doi: 10.1002/cne.
23521
Rolf, B., Lang, D., Hillenbrand, R., Richter, M., Schachner, M., and Bartsch, U.
(2003). Altered expression of CHL1 by glial cells in response to optic nerve
injury and intravitreal application of fibroblast growth factor-2. J. Neurosci. Res.
71, 835–843. doi: 10.1002/jnr.10533
Sanes, J. R., and Masland, R. H. (2015). The types of retinal ganglion cells: current
status and implications for neuronal classification. Annu. Rev. Neurosci. 38,
221–246. doi: 10.1146/annurev-neuro-071714-034120
Singhal, S., Bhatia, B., Jayaram, H., Becker, S., Jones, M. F., Cottrill, P. B., et al.
(2012). Human Muller glia with stem cell characteristics differentiate into
retinal ganglion cell (RGC) precursors in vitro and partially restore RGC
function in vivo following transplantation. Stem Cells Transl. Med. 1, 188–199.
doi: 10.5966/sctm.2011-0005
Song, M. R., and Ghosh, A. (2004). FGF2-induced chromatin remodeling regulates
CNTF-mediated gene expression and astrocyte differentiation.Nat. Neurosci. 7,
229–235. doi: 10.1038/nn1192
Song,W. T., Zhang, X. Y., and Xia, X. B. (2013). Atoh7 promotes the differentiation
of retinal stem cells derived from Muller cells into retinal ganglion cells by
inhibiting Notch signaling. Stem Cell Res. Ther. 4:94. doi: 10.1186/scrt305
Todd, L., Volkov, L. I., Zelinka, C., Squires, N., and Fischer, A. J. (2015). Heparin-
binding EGF-like growth factor (HB-EGF) stimulates the proliferation of
Muller glia-derived progenitor cells in avian and murine retinas. Mol. Cell
Neurosci. 69, 54–64. doi: 10.1016/j.mcn.2015.10.004
Torper, O., and Gotz, M. (2017). Brain repair from intrinsic cell sources:
Turning reactive glia into neurons. Prog. Brain. Res. 230, 69–97.
doi: 10.1016/bs.pbr.2016.12.010
Turner, D. L., Snyder, E. Y., and Cepko, C. L. (1990). Lineage-independent
determination of cell type in the embryonic mouse retina. Neuron 4, 833–845.
doi: 10.1016/0896-6273(90)90136-4
Ueki, Y., Wilken, M. S., Cox, K. E., Chipman, L., Jorstad, N., Sternhagen, K., et al.
(2015). Transgenic expression of the proneural transcription factor Ascl1 in
Muller glia stimulates retinal regeneration in young mice. Proc. Natl. Acad. Sci.
U.S.A. 112, 13717–13722. doi: 10.1073/pnas.1510595112
Wilken, M. S., and Reh, T. A. (2016). Retinal regeneration in birds and mice. Curr.
Opin. Genet. Dev. 40, 57–64. doi: 10.1016/j.gde.2016.05.028
Wohl, S. G., and Reh, T. A. (2016). miR-124-9-9∗ potentiates Ascl1-
induced reprogramming of cultured Muller glia. Glia 64, 743–762.
doi: 10.1002/glia.22958
Wong-Riley, M. T. (2010). Energy metabolism of the visual system. Eye Brain 2,
99–116. doi: 10.2147/EB.S9078
Wu, Z. K., Cao, L., Zhang, X. Y., Song, W. T., and Xia, X. B. (2016). Promotion on
the differentiation of retinal Muller cells into retinal ganglion cells by Brn-3b.
Int. J. Ophthalmol. 9, 948–954. doi: 10.18240/ijo.2016.07.03
Yang, W., and Yuste, R. (2017). In vivo imaging of neural activity. Nat. Methods 14,
349–359. doi: 10.1038/nmeth.4230
Yao, K., Qiu, S., Wang, Y. V., Park, S. J. H., Mohns, E. J., Mehta, B., et al.
(2018). Restoration of vision after de novo genesis of rod photoreceptors
in mammalian retinas. Nature 560, 484–488. doi: 10.1038/s41586-018-
0425-3
Young, R.W. (1985). Cell differentiation in the retina of the mouse.Anat. Rec. 212,
199–205. doi: 10.1002/ar.1092120215
Conflict of Interest Statement: The authors declare that the research was
conducted in the absence of any commercial or financial relationships that could
be construed as a potential conflict of interest.
Copyright © 2018 Guimarães, Landeira, Coelho, Golbert, Silveira, Linden, de
Melo Reis and Costa. This is an open-access article distributed under the terms
of the Creative Commons Attribution License (CC BY). The use, distribution or
reproduction in other forums is permitted, provided the original author(s) and the
copyright owner(s) are credited and that the original publication in this journal
is cited, in accordance with accepted academic practice. No use, distribution or
reproduction is permitted which does not comply with these terms.
Frontiers in Cellular Neuroscience | www.frontiersin.org 15 November 2018 | Volume 12 | Article 410



5xgtyixovzoutgr vxulork ul otj{ikj gtj vxosgx� tk{xuty xk|kgry tk} igtjojgzk mktky lux rotkgmk xkvxumxgssotm
%0*� ���������NB{FST������������� .B�FST *44/� //64�5/1- ] �
JO DFMMT UIBU VOEFSHP B DPNQMFUF QSPHSBN PG OFVSPOBM EJVFSFOUJBUJPO BOE JJ HFOFT SFH�
VMBUFE JO DFMMT UIBU GBJMFE UP SFQSPHSBN� "T B DPOTFRVFODF HFOFT XFBLMZ SFHVMBUFE VQPO
OFVSPHFOJD USBOTDSJQUJPO GBDUPS FYQSFTTJPO XIJDI NBZ CF QJWPUBM GPS SFQSPHSBNNJOH
BSF MJLFMZ PWFSMPPLFE� .PSF SFDFOUMZ UIF WSTU DPNQSFIFOTJWF USBOTDSJQUJPOBM BOBMZTJT
PG OFVSPOBM JOEVDUJPO XBT QVCMJTIFE <ûü>� _JT XPSL TIPXFE UIBU NPVTF FNCSZPOJD
WCSPCMBTUT .&' FYQSFTTJOH POMZ "TDMû NPTUMZ GBJMFE UP HP UISPVHI B DPNQMFUF EJVFS�
FOUJBUJPO JOUP J/ HPJOH JOTUFBE UPXBSET B NZPHFOJD QIFOPUZQF� _JT JT BMTP PCTFSWFE
GPS .&'T FYQSFTTJPO B DPNCJOBUJPO PG UISFF 5'T #SOû .ZUûM BOE "TDMû�#". BMUIPVHI
UIF GSFRVFODZ PG DFMMT BEPQUJOH B OFVSPOBM QIFOPUZQF JODSFBTFT JO UIJT MBTU DPOEJUJPO�
)PXFWFS JU SFNBJOT VODMFBS XIFUIFS UIF USBOTDSJQUJPOBM SFHVMBUJPO JOWPMWFE JO UIF MJO�
FBHF SFQSPHSBNNJOH PG .&'T UP JOEVDFE OFVSPOT SFTFNCMFT UIBU PCTFSWFE EVSJOH CPOB
�EF OFVSPO EJVFSFOUJBUJPO�
*O UIJT XPSL XF VTFE BO VOTVQFSWJTFE NBDIJOF�MFBSOJOH BQQSPBDI CBTFE PO QSJODJQBM
DPNQPOFOU BOBMZTJT 1$" UP TFMFDU HFOFT UIBU CFRFS DPSSFTQPOE UP EJVFSFOU DFMM TUBUFT
PG .&'T .&'T ÿ EBZT BOFS USBOTEVDUJPO XJUI "TDMû BTDMûEÿ .&'T üü EBZT BOFS USBOT�
EVDUJPO XJUI "TDMû BTDMûEüü BOE .&'T üü EBZT BOFS USBOTEVDUJPO XJUI #SOü "TDMû BOE
.ZUûM CBNEüü 5SFVUMFJO # FU BM� üúû� QSJNBSZ OFVSPOT GSPN QPTUOBUBM NJDF CSBJOT
1� <ûý> BOE OFVSBM QSPHFOJUPST BOE QSJNBSZ OFVSPOT GSPN FNCSZPOJD NJDF CSBJOT <ûþ>�
/FYU XF DPNQBSFE UIF USBOTDSJQUJPOBM QSPWMFT PG.&'T VOEFSHPJOH OFVSPOBM DPOWFSTJPO
PS .&'T UIBU GBJMFE UP EP TP BOE JEFOUJWFE TPNF QPUFOUJBM DBOEJEBUF HFOFT UP FOIBODF
MJOFBHF SFQSPHSBNNJOH�


5xgtyixovzoutgr vxulork ul otj{ikj gtj vxosgx� tk{xuty xk|kgry tk} igtjojgzk mktky lux rotkgmk xkvxumxgssotm
%0*� ���������NB{FST������������� .B�FST *44/� //64�5/1- ] �
" 1TFVEP�UFNQPSBM DFMM EFDJTJPO NBQ� "O VOTVQFSWJTFE NBDIJOF�MFBSOJOH BQQSPBDI
CBTFE PO 1$" TFMFDUFE þúú HFOFT XIPTF FYQSFTTJPO TJHOBUVSF XBT VTFE UP TFQBSBUF TBN�
QMF DFMMT� " EJNFOTJPO SFEVDUJPO GPMMPXFE CZ DFMM PSEFSJOH XBT UIFO QMPRFE UP HJWF VT
B QTFVEP�UFNQPSBM QPTJUJPOJOH PG DFMMT SFBDIJOH EJWFSHFOU QIFOPUZQFT� $FMM UZQFT BSF�
.&' NPVTF FNCSZPOJD WCSPCMBTUT BTDMûEÿ .&' DFMMT SFQSPHSBNNFE XJUI BTDMû BOFS ÿ
EBZT BTDMûEüü TBNF BT CFGPSF XJUI üü EBZT CBNEüü TBNF XJUI #". USBOTDSJQUJPOBM
GBDUPST OFVSPO1ú� QPTUOBUBM NPVTF DFSFCSBM DPSUFY OFVSPOT OFVSPO&ûþ FNCSZPOJD
OFVSPOT NPVTF EPSTBM UFMFODFQIBMPO /%1 OFVSBM EJVFSFOUJBUFE QSPHFOJUPST NPVTF
EPSTBM UFMFODFQIBMPO BOE /11 OFVSBM QSPMJGFSBUJOH QSPHFOJUPST NPVTF EPSTBM UFMFO�
DFQIBMPO�
# .PVTF (FOF "UMBT DFMM POUPMPHJFT SFMBUFE UP HFOFT TFMFDUFE CZ UIF NBDIJOF�MFBSOJOH
BQQSPBDI� (FOFT TFMFDUFE CZ VOTVQFSWJTFE NBDIJOF�MFBSOJOH XFSF FOSJDIFE GPS WCSPC�
MBTUT DFMM MJOFT .&' BOE /*)�ý5ý NZPCMBTU DFMM MJOFBHF $ü$ûü BOE TFWFSBM OFVSPOBM
DFMM MJOFT )ZQPUIBMBNVT BNZHEBMB DFSFCSBM DPSUFY BOE PUIFST� _F DPNCJOFE TDPSF
JT DPNQVUFE CZ UBLJOH UIF MPH PG UIF Q�WBMVF GSPN UIF 'JTIFS FYBDU UFTU BOE NVMUJQMZJOH
UIBU CZ UIF [�TDPSF PG UIF EFWJBUJPO GSPN UIF FYQFDUFE SBOL�
$ &YQSFTTJPO PG DFMM�UZQF TQFDJWD HFOFT BMPOH UIF QTFVEP�UFNQPSBM NBQ� (FOF FYQSFT�
TJPO MFWFMT BSF QPTJUJWFMZ BTTPDJBUFE UP UIF EJBNFUFS PG UIF DPSSFTQPOEJOH DJSDMFT� /PUF
UIBU HFOFT BTTPDJBUFE XJUI WCSPCMBTU QIFOPUZQF 4ûúúBþ BOE $PMûBü BSF FOSJDIFE JO UIF
CSBODI DPOUBJOJOH .&'T BOE "TDMûEÿ DFMMT XIFSFBT HFOFT BTTPDJBUFE UP NVTDMF QIFOP�
UZQFT .ZPû�C BOE 5OODü BSF FOSJDIFE JO B CSBODI DPOUBJOJOH BTDMûEüü GFX CBNEüü
BOE BTDMûEÿ DFMMT� (FOFT BTTPDJBUFE XJUI OFVSPOBM GBUFT .BQü BOE 5VCCý BSF FOSJDIFE
JO UIF UIJSE CSBODI DPOUBJOJOH NPTU CBNEüü DFMMT GFX BTDMûEÿ BOE QSJNBSZ OFVSPOT
OFVSPO1ú� BOE OFVSPO&ûþ�
% 4BNF BT JO # OPX TIPXJOH UIF SFMBUJWF FYQSFTTJPO PG TFMFDUFE HFOFT UISPVHIPVU
PSEFSFE DFMMT EJTUSJCVUJPO�
& )FBUNBQ PG EJVFSFOUJBMMZ FYQSFTTFE HFOFT BNPOH BTDMûEÿ TVCHSPVQT� 4PNF HFOFT
XFSF IJHIMZ SFMBUFE UP TQFDJWD TVCHSPVQT MJLF GPS FYBNQMF "DUBû UIBU JT NBJOMZ FY�
QSFTTFE JO DFMMT GSPN BTDMûEÿNVTDVMBS CSBODI BTDMûEÿ�NVT� 0UIFS DBTFT JODMVEFT -VN
HFOF NBJOMZ FYQSFTTFE JO UIF VOEJVFSFOUJBUFE BTDMûEÿ CSBODI BTDMûEÿ�VOE BOE $SNQû
IJHIMZ FYQSFTTFE JO TBNQMFT MPDBUFE JO BTDMûEÿ OFVSPOBM CSBODI BTDMûEÿ�OFV�
' (FOFT FOSJDIFE JO BTDMûEÿ DFMMT JO UIF OFVSPOBM CSBODI BMPOH UIF QTFVEP�UFNQPSBM
NBQ� $JSDMFT XJUI EJVFSFOU TJ[FT JMMVTUSBUFT IPX DBOEJEBUF HFOFT BSF FYQSFTTFE� _FTF
QTFVEP�UFNQPSBM NBQT SFBTTVSF IPX UIPTF HFOFT BSF SFMBUFE UP TBNQMFT JO OFVSPOBM
CSBODI�
3FTVMUT � %JTDVTTJPO5P DPNQBSF UIF USBOTDSJQUJPOBM QSPWMFT MJOFBHF�SFQSPHSBNNFE BOE QSJNBSZ DFMMT BO VO�
TVQFSWJTFE NBDIJOF�MFBSOJOH BQQSPBDI CBTFE PO QSJODJQBM DPNQPOFOU BOBMZTJT 1$"
XBT VTFE UP TFMFDU HFOFT UIBU CFRFS DPSSFTQPOE UP EJVFSFOU DFMM TUBUFT <ûÿ>� 8F BOB�
MZ[FE UISFF EBUBTFUT� û 4JOHMF�DFMM 3/"TFR PG NPVTF FNCSZPOJD WCSPCMBTU .&' .&' ÿ
EBZT BOFS USBOTEVDUJPO XJUI "TDMû BTDMûEÿ .&' üü EBZT BOFS USBOTEVDUJPO XJUI "TDMû
BTDMûEüü BOE .&' üü EBZT BOFS USBOTEVDUJPO XJUI #SOü "TDMû BOE .ZUûM CBNEüü
<ûü> (&0� (4&��ýûú� ü 1SJNBSZ OFVSPOT GSPN QPTUOBUBM NJDF CSBJOT 1� <ûý> (&0�
(4&ÿüÿ�þ� BOE ý OFVSBM QSPHFOJUPST BOE QSJNBSZ OFVSPOT GSPN FNCSZPOJD NJDF CSBJOT
<ûþ> (&0� (4&�ÿþ��� "T FBDI HFOF SFQSFTFOUT B EJNFOTJPO B TUSBUFHZ CBTFE PO B
EJNFOTJPOBM SFEVDUJPO XBT BQQMJFE� _VT TBNQMFT XFSF PSEFSFE UP DSFBUF B QTFVEP�
UFNQPSBM NBQ XIJDI QMBDFT DFMMT UPXBSET B DFMM EJVFSFOUJBUJPO TUBUF <ûÿ> 'JHVSF "�
*OUFSFTUJOHMZ XF PCTFSWFE UIBU UIF USBOTDSJQUJPOBM QSPWMFT PG SFQSPHSBNNFE JOEVDFE
OFVSPOT BOE QSJNBSZ OFVSPOT GVMMZ PWFSMBQQFE 'JHVSF "� .PSF JOUFSFTUJOHMZ B DPOUJO�
VVN XBT PCTFSWFE GSPN .&'T UP B TVCQPQVMBUJPO PG "TDMû� PS #".�USBOTEVDFE DFMMT
BOFS ÿ BOE üü EBZT XIFSF QBSU PG UIF DFMMT PWFSMBQ XJUI QSJNBSZ OFVSBM QSPHFOJUPST
BOE JNNBUVSF OFVSPOT XIFSFBT PUIFS DFMMT GPMMPX B EJVFSFOU QBUI 'JHVSF "�_JT WSTU
BOBMZTJT TVHHFTUT UIBU UIF USBOTDSJQUJPOBM QSPHSBNT FOBDUFE EVSJOH UIF DPOWFSTJPO GSPN
.&'T UP JOEVDFE OFVSPOT BOE OFVSBM QSPHFOJUPS DFMMT UP OFVSPOT BSF TJNJMBS�
5P GVSUIFS DPOWSN UIBU UIF PCTFSWFE QTFVEP�UFNQPSBM NBQ DPVME SFQSFTFOU EJVFSFOU
GBUFT PG SFQSPHSBNNFE .&'T XF WSTU BOBMZ[FE DFMM�UZQF POUPMPHJFT PO .PVTF (FOF "U�
MBT VTJOH FOSJDI3 'JHVSF #� 8F DPVME DPOWSN UIBU HFOFT XFSF FOSJDIFE GPS .&' OFVSBM

5xgtyixovzoutgr vxulork ul otj{ikj gtj vxosgx� tk{xuty xk|kgry tk} igtjojgzk mktky lux rotkgmk xkvxumxgssotm
%0*� ���������NB{FST������������� .B�FST *44/� //64�5/1- ] �
DFMMT BOE NVTDMF DFMM POUPMPHJFT 'JHVSF #� /FYU QBO�OFVSPOBM NBSLFST 5VCCý .BQü
NZPDZUF NBSLFST 5OODü .ZPû�C BOE .&' NBSLFST 4ûúúBþ $PMûBü XFSF TFMFDUFE
BNPOH UIF þúú HFOFT UP BOBMZ[F UIF MFWFMT PG FYQSFTTJPO JO EJVFSFOU DFMM TUBUFT 'JHVSF $
BOE%� 4ûúúBþ BOE $PMûBüXFSFNPSF FYQSFTTFE JO UIF CSBODI DPOUBJOJOH.&'T XIFSFBT
5OODü BOE .ZPû�C XFSF NPSF FYQSFTTFE JO UIF CSBODI DPNQSJTJOH NPTU BTDMûEÿ BOE B
TNBMM TVCTFU PG CBNEüü DFMMT� *O DPOUSBTU 5VCCý BOE .BQü FYQSFTTJPO XFSF FOSJDIFE JO
UIF CSBODI DPNQSJTJOH GFX BTDMûEÿ UIF NBKPSJUZ PG CBNEüü BOE QSJNBSZ OFVSBM QSPHFO�
JUPST BOE OFVSPOT� _FTF PCTFSWBUJPOT TVHHFTU UIBU UIF OPEF OVNCFS POF JEFOUJWFE JO
UIF QTFVEP�UFNQPSBM NBQ QJOQPJOUT UIF EJWFSHFODF CFUXFFO DFMMT GPMMPXJOH B NVTDVMBS�
PS OFVSPOBM�DFMM GBUF�
/FYU XF TFU PVU UP JEFOUJGZ HFOFT FOSJDIFE JO UIF BTDMûEÿ DFMMT DMBTTJWFE JO UIF OFV�
SPOBM CSBODI BT DPNQBSFE UP UIF .&' BOE NVTDMF�DFMM CSBODIFT� _VT HFOF FYQSFTTJPO
QBRFSOT PG BTDMEûEÿ DFMM QPQVMBUJPOT DMBTTJWFE JO FBDI PG UIPTF CSBODIFT XFSF DPNQBSFE�
8F JEFOUJWFE ýý HFOFT EJVFSFOUJBMMZ FYQSFTTFE R�WBMVF �ú�úÿ MJLFMJIPPE SBUJP UFTU JO
UIF UISFF EJVFSFOU QPQVMBUJPOT PG BTDMûEÿ DFMMT 'JHVSF &� "NPOH UIFTF HFOFT þ XFSF
IJHIMZ FOSJDIFE JO BTDMûEÿ DFMMT JO UIF OFVSPOBM CSBODI 'JHVSF '� *OUFSFTUJOHMZ UIFTF
HFOFT XFSF BMTP FOSJDIFE JO CBNEüü DFMMT BOE QSJNBSZ OFVSBM DFMMT 'JHVSF '�
*O UIJT XPSL XF TIPX UIBU MJOFBHF�SFQSPHSBNNFE .&'T VOEFSHP USBOTDSJQUJPOBM
DIBOHFT UPXBSET UIF HFOFSBUJPO PG JOEVDFE OFVSPOT UIBU SFTFNCMFT UIPTF PCTFSWFE JO
UIF USBOTJUJPO GSPN QSJNBSZ DFSFCSBM DPSUFY QSPHFOJUPST UP FBSMZ�EJVFSFOUJBUFE OFVSPOT�
ÿ EBZT BOFS FYQSFTTJPO PG "TDMû JO .&'T B TVCTFU PG DFMMT TIPX FOSJDIFE FYQSFTTJPO PG
QBO�OFVSPOBM HFOFT BOE BSF USBOTDSJQUJPOBMMZ TJNJMBS UP OFVSBM QSPHFOJUPST� 4JNJMBSMZ
UIF USBOTDSJQUJPOBM QSPWMF PG CBNEüü DFMMT XIJDI NPTUMZ BEPQU B J/ QIFOPUZQF <ûü> JT
DMPTFMZ SFMBUFE UP 1� DFSFCSBM DPSUFY OFVSPOT� *O DPOUSBTU MJOFBHF�SFQSPHSBNNFE .&'T
UIBU FYQSFTT MPX MFWFMT PG QBO�OFVSPOBM HFOFT TIPXFE FOSJDIFE FYQSFTTJPO PG WCSPC�
MBTU HFOFT PS NVTDMF�DFMM HFOFT JOEJDBUJOH UIBU UIPTF UXP QPQVMBUJPOT SFQSFTFOU .&'T
UIBU GBJMFE UP VOEFSHP MJOFBHF�DPOWFSTJPO PS GPMMPXFE BO BMUFSOBUJWF GBUF <ûü>� 8F BMTP
TIPX UIBU UISFF EJVFSFOU QPQVMBUJPOT PG BTDMûEÿ DFMMT DBO CF EJTUJOHVJTIFE CBTFE PO UIFJS
USBOTDSJQUJPOBM QSPWMFT� (FOF FYQSFTTJPO QBRFSOT PG UIFTF QPQVMBUJPOT BSF DMBTTJWFE JO
UIF VOTVQFSWJTFE NBDIJOF�MFBSOJOH BQQSPBDI JO CSBODIFT DPOUBJOJOH FJUIFS VOEJVFSFO�
UJBUFE .&'T NVTDMF DFMMT PS OFVSPOT� 6TJOH UIJT DMBTTJWDBUJPO XF JEFOUJWFE ýý HFOFT
EJVFSFOUJBMMZ FYQSFTTFE JO BTDMûEÿ DFMM QPQVMBUJPOT BOE GPVS HFOFT TQFDJWDBMMZ FOSJDIFE
JO BTDMûEÿ DFMMT JO UIF OFVSPOBM CSBODI� _FTF HFOFT NBZ CF JOUFSFTUJOH DBOEJEBUFT PS
DPOUSJCVUF UP JEFOUJGZ OFX GBDUPST UP FOIBODF .&' MJOFBHF�SFQSPHSBNNJOH JOUP J/T�
$PMMBQTJO 3FTQPOTF .FEJBUPS 1SPUFJO û $SNQû JT QBSU PG $3.1 GBNJMZ PG QSPUFJOT BOE
JT UZQJDBMMZ BTTPDJBUFE BT NFEJBUPS PG TFNBý" TJHOBMJOH BOE BYPO HVJEBODF <û�> <û�>�
*OUFSFTUJOHMZ TPNF $3.1 QSPUFJOT BSF EJGFSFOUJBMMZ FYQSFTTFE JO BYPO BOE EFOESJUFT PG
EJTUJODU OFVSPOBM UZQFT <û�>� &NCSZPOJD -FUIBM "COPSNBM 7JTJPO %SPTPQIJMB�-JLF þ
&MBWMþ BMTP LOPXO BT )V�"OUJHFO % )V% JT B 3/"�CJOEJOH QSPUFJO JOWPMWFE JO OFV�
SPOBM NBUVSBUJPO <û�> OFVSJUF PVUHSPXUI BOE EFOESJUJD NBJOUFOBODF <û�>� 4UBUINJO ý
4UNOý PS 4$(ûú�-JLF 1SPUFJO 4$-*1 JT BMTP SFMBUFE UP EFOESJUJD GPSNBUJPO <üú> BOE
OFVSJUF PVUHSPXUI <üû>� ;JOD WOHFS $$)$ EPNBJO DPOUBJOJOH ûü ;DDIDûü PS 4NBE�
*OUFSBDUJOH ;JOD 'JOHFS 1SPUFJO û 4[Oû JT B QSPUFJO VTFE JO #.1 <üü> "1�û BOE $3&#
TJHOBMMJOH <üý> BT B DP�BDUJWBUPS� 1PTTJCMZ #SOü BOE .ZUûM NBZ TVTUBJO HFOF FYQSFTTJPO
PG UIPTF DBOEJEBUF HFOFT MPOHFS UIBO "TDMû�POMZ SFQSPHSBNNJOHXIJDI BMMPXT.&' DFMMT
EJVFSFOUJBUF JOUP OFVSPO�MJLF DFMMT� _FSFCZ BMM DBOEJEBUFT BSF SFMBUFE UP OFVSPOBM QIF�
OPUZQF BU TPNF MFWFM XIJDI JOEJDBUF QPTTJCMF QSPUFJOT UP IFMQ "TDMû SFQSPHSBNNJOH
.&'T BDIJFWF B OFVSPO�MJLF TUBUF�
0VS SFTVMUT JOEJDBUF UIBU TPNBUJD DFMMT EVSJOH UIF QSPDFTT PG MJOFBHF SFQSPHSBNNJOH
JOUP JOEVDFE OFVSPOT VOEFSHP USBOTDSJQUJPOBM DIBOHFT SFTFNCMJOH UIPTF FOBDUFE JO UIF
USBOTJUJPO GSPN CPOB �EF OFVSBM QSPHFOJUPST UP OFVSPOBM TUBUFT� $PNQBSJTPO PG USBO�
TDSJQUJPOBM QSPWMFT PG JOUFSNFEJBUF TUBHFT EVSJOH MJOFBHF DPOWFSTJPO NBZ DPOUSJCVUF UP
JEFOUJGZ OFX DBOEJEBUF HFOFT UP JNQSPWF OFVSPOBM SFQSPHSBNNJOH�
0OMZ .&'T XJUI "TDMû BOE #". XFSF BOBMZ[FE XIJDI JMMVTUSBUFT B MJNJUFE TDSFFO PG
SFQSPHSBNNFE DFMMT QPTTJCJMJUJFT� _VT BOBMZTJT PG EJVFSFOU DFMM UZQFT SFQSPHSBNNFE
XJUI "TDMû BOE #". BT XFMM BT .&' MJOFBHF SFQSPHSBNNFE XJUI PUIFS USBOTDSJQUJPO
GBDUPST BSF WJUBM UP GVMMZ VOEFSTUBOE UIPTF QBUIXBZT UBLFO CZ SFQSPHSBNNFE DFMMT� 'VS�
UIFSNPSF B NPSF EJWFSTF TFU PG DPOUSPMT BSF OFFEFE TJODF POMZ DFSFCSBM DPSUFY DFMMT XFSF

VTFE BT OBJWF OFVSPOBM DFMM SFGFSFODF�
*U XPVME CF JOUFSFTUJOH UP FWBMVBUF DBOEJEBUF HFOFTXJUI B TZTUFN CJPMPHZ BQQSPBDI MPPL�
JOH BU GPS FYBNQMF HFOF SFHVMBUPSZ OFUXPSLT� 'VSUIFSNPSF NJ3/"T QMBZT BO JNQPS�
UBOU SPMF PO SFHVMBUJPO PG HFOF OFUXPSLT BOE IBE CFFO BMSFBEZ VTFE BT SFQSPHSBNNJOH
FOIBODFST TP UIFZ TIPVME OPU CF OFHMFDUFE JO UIF GVUVSF BOBMZTJT�
"EEJUJPOBM *OGPSNBUJPO
.FUIPET BOE 4VQQMFNFOUBSZ .BUFSJBM
1MFBTF TFF I{QT���TDJFODFNB{FST�JP�BSUJDMFT��������������
%JFHP .� $PFMIP JT TVQQPSUFE CZ B 1I�%� GFMMPXTIJQ GSPN $"1&4�
'PS UIPVHIUGVM EJTDVTTJPOT XF HMBEMZ UIBOL "OES© 'POTFDB 7BOEFDM©DMJP EB 4JMWB BOE
1SPG� +PSHF &TUFGBOP %F 4PV[B�
&UIJDT 4UBUFNFOU
/PU "QQMJDBCMF�
$JUBUJPOT
<û> #FSOJOHFS #� FU BM� à'VODUJPOBM 1SPQFSUJFT PG /FVSPOT %FSJWFE
GSPN *O 7JUSP 3FQSPHSBNNFE 1PTUOBUBM "TUSPHMJBá� *O� +PVSOBM PG
/FVSPTDJFODF ü��ýü "VH� üúú� QQ� ��ÿþ×���þ� juo�
��������KOFVSPTDJ�������������� {xr� IUUQT���EPJ�PSH���������KOFVSPTDJ��������������
<ü> )FJOSJDI $ISJTUPQIF FU BM� à%JSFDUJOH "TUSPHMJB GSPN UIF $FSFCSBM
$PSUFY JOUP 4VCUZQF 4QFDJWD 'VODUJPOBM /FVSPOTá� *O� 1-P4
#JPMPHZ ��ÿ .BZ üúûú Fûúúúý�ý� juo�
��������KPVSOBM�QCJP��������� {xr� IUUQT���EPJ�PSH���������KPVSOBM�QCJP���������
<ý> 7JFSCVDIFO _PNBT FU BM� à%JSFDU DPOWFSTJPO PG WCSPCMBTUT UP
GVODUJPOBM OFVSPOT CZ EFWOFE GBDUPSTá� *O� /BUVSF þ�ý��ü�þ +BO�
üúûú QQ� ûúýÿ×ûúþû� juo� ��������OBUVSF������ {xr�IUUQT���EPJ�PSH���������OBUVSF������
<þ> .BSSP 4BNVFMF FU BM� à%JSFDU -JOFBHF $POWFSTJPO PG 5FSNJOBMMZ
%JVFSFOUJBUFE )FQBUPDZUFT UP 'VODUJPOBM /FVSPOTá� *O� $FMM 4UFN
$FMM ��þ 0DU� üúûû QQ� ý�þ×ý�ü� juo�
��������K�TUFN������������� {xr� IUUQT���EPJ�PSH���������K�TUFN�������������
<ÿ> $IBOEB 4PIBN FU BM� à(FOFSBUJPO PG *OEVDFE /FVSPOBM $FMMT CZ
UIF 4JOHMF 3FQSPHSBNNJOH 'BDUPS "4$-ûá� *O� 4UFN $FMM 3FQPSUT
ý�ü "VH� üúûþ QQ� ü�ü×ü��� juo�
��������K�TUFNDS������������� {xr� IUUQT���EPJ�PSH���������K�TUFNDS�������������
<�> "NCBTVEIBO 3BKFTI FU BM� à%JSFDU 3FQSPHSBNNJOH PG "EVMU
)VNBO 'JCSPCMBTUT UP 'VODUJPOBM /FVSPOT VOEFS %FWOFE
$POEJUJPOTá� *O� $FMM 4UFN $FMM ��ü "VH� üúûû QQ� ûûý×ûû�� juo�
��������K�TUFN������������� {xr� IUUQT���EPJ�PSH���������K�TUFN�������������
<�> 4PO &TUIFS :� FU BM� à$POWFSTJPO PG .PVTF BOE )VNBO 'JCSPCMBTUT
JOUP 'VODUJPOBM 4QJOBM .PUPS /FVSPOTá� *O� $FMM 4UFN $FMM ��ý
4FQU� üúûû QQ� üúÿ×üû�� juo�
��������K�TUFN������������� {xr� IUUQT���EPJ�PSH���������K�TUFN�������������
<�> ,BSPX .BSJTB FU BM� à-JOFBHF�SFQSPHSBNNJOH PG 1FSJDZUF�EFSJWFE
$FMMT PG UIF "EVMU )VNBO #SBJO JOUP *OEVDFE /FVSPOTá� *O�
+PVSOBM PG 7JTVBMJ[FE &YQFSJNFOUT �� .BZ üúûþ� juo�
�������������� {xr�IUUQT���EPJ�PSH���������������
<�> -JV 9JOKJBO FU BM� à%JSFDU SFQSPHSBNNJOH PG IVNBO WCSPCMBTUT
JOUP EPQBNJOFSHJD OFVSPO�MJLF DFMMTá� *O� $FMM 3FTFBSDI üü�ü /PW�
üúûû QQ� ýüû×ýýü� juo� ��������DS���������� {xr�IUUQT���EPJ�PSH���������DS����������
<ûú> -JV .FOH�-V FU BM� à4NBMM NPMFDVMFT FOBCMF OFVSPHFOJO ü UP
FYDJFOUMZ DPOWFSU IVNBO WCSPCMBTUT JOUP DIPMJOFSHJD OFVSPOTá�
*O� /BUVSF $PNNVOJDBUJPOT þ +VMZ üúûý� juo�
��������ODPNNT����� {xr�IUUQT���EPJ�PSH���������ODPNNT�����
<ûû> 8BQJOTLJ 0SMZ -� FU BM� à)JFSBSDIJDBM .FDIBOJTNT GPS %JSFDU
3FQSPHSBNNJOH PG 'JCSPCMBTUT UP /FVSPOTá� *O� $FMM ûÿÿ�ý 0DU�
üúûý QQ� �üû×�ýÿ� juo� ��������K�DFMM�������������{xr� IUUQT���EPJ�PSH���������K�DFMM�������������
<ûü> 5SFVUMFJO #BSCBSB FU BM� à%JTTFDUJOH EJSFDU SFQSPHSBNNJOH GSPN
WCSPCMBTU UP OFVSPO VTJOH TJOHMF�DFMM 3/"�TFRá� *O� /BUVSF
ÿýþ���ú� +VOF üúû� QQ� ý�û×ý�ÿ� juo�
��������OBUVSF������ {xr�IUUQT���EPJ�PSH���������OBUVSF������
<ûý> ;IBOH :� FU BM� à"O 3/"�4FRVFODJOH 5SBOTDSJQUPNF BOE 4QMJDJOH
%BUBCBTF PG (MJB /FVSPOT BOE 7BTDVMBS $FMMT PG UIF $FSFCSBM
$PSUFYá� *O� +PVSOBM PG /FVSPTDJFODF ýþ�ý� 4FQU� üúûþ
QQ� ûû�ü�×ûû�þ�� juo�
��������KOFVSPTDJ�������������� {xr� IUUQT���EPJ�PSH���������KOFVSPTDJ��������������
<ûþ> "QSFB +VMJFUB FU BM� à*EFOUJWDBUJPO BOE FYQSFTTJPO QBRFSOT PG
OPWFM MPOH OPO�DPEJOH 3/"T JO OFVSBM QSPHFOJUPST PG UIF
EFWFMPQJOH NBNNBMJBO DPSUFYá� *O� /FVSPHFOFTJT ü�û +BO� üúûÿ
F��ÿÿüþ� juo� ����������������������������� {xr�IUUQT���EPJ�PSH������������������������������
<ûÿ> 5SBQOFMM $PMF FU BM� à_F EZOBNJDT BOE SFHVMBUPST PG DFMM GBUF
EFDJTJPOT BSF SFWFBMFE CZ QTFVEPUFNQPSBM PSEFSJOH PG TJOHMF
DFMMTá� *O� /BUVSF #JPUFDIOPMPHZ ýü�þ .BS� üúûþ QQ� ý�û×ý��� juo�
��������OCU������ {xr�IUUQT���EPJ�PSH���������OCU������
<û�> #SFUJO 4ZMWJF FU BM� à%JVFSFOUJBM FYQSFTTJPO PG $3.1û $3.1ü"
$3.1ü# BOE $3.1ÿ JO BYPOT PS EFOESJUFT PG EJTUJODU OFVSPOT JO
UIF NPVTF CSBJOá� *O� +PVSOBM PG $PNQBSBUJWF /FVSPMPHZ þ���û
"QS� üúúÿ QQ� û×û�� juo� ��������DOF������� {xr�IUUQT���EPJ�PSH���������DOF�������
<û�> 5BLBZB 3ZPTVLF FU BM� à$3.1û BOE $3.1þ BSF SFRVJSFE GPS
QSPQFS PSJFOUBUJPO PG EFOESJUFT PG DFSFCSBM QZSBNJEBM OFVSPOT JO
UIF EFWFMPQJOH NPVTF CSBJOá� *O� #SBJO 3FTFBSDI û�ÿÿ +BO� üúû�
QQ� û�û×û��� juo� ��������K�CSBJOSFT�������������{xr� IUUQT���EPJ�PSH���������K�CSBJOSFT�������������

�
<û�> "LBNBUTV 8� FU BM� à_F 3/"�CJOEJOH QSPUFJO )V% SFHVMBUFT
OFVSPOBM DFMM JEFOUJUZ BOE NBUVSBUJPOá� *O� 1SPDFFEJOHT PG UIF
/BUJPOBM "DBEFNZ PG 4DJFODFT ûúü�ûü .BS� üúúÿ QQ� þ�üÿ×þ�ýú�
juo� ��������QOBT������������ {xr�IUUQT���EPJ�PSH���������QOBT������������
<û�> "CEFMNPITFO ,� FU BM� àNJ3�ý�ÿ *OIJCJUT %JVFSFOUJBUJPO PG
/FVSJUFT CZ -PXFSJOH )V% -FWFMTá� *O� .PMFDVMBS BOE $FMMVMBS
#JPMPHZ ýú�û� +VOF üúûú QQ� þû��×þüûú� juo�
��������NDC���������� {xr�IUUQT���EPJ�PSH���������NDC����������
<üú> 1PVMBJO '� &� FU BM� à4$-*1 *T $SVDJBM GPS UIF 'PSNBUJPO BOE
%FWFMPQNFOU PG UIF 1VSLJOKF $FMM %FOESJUJD "SCPSá� *O� +PVSOBM PG
/FVSPTDJFODF ü��ü� +VMZ üúú� QQ� �ý��×�ý��� juo�
��������KOFVSPTDJ�������������� {xr� IUUQT���EPJ�PSH���������KOFVSPTDJ��������������
<üû> .PSJJ )JSPTIJ 4IJSBJTIJ�:BNBHVDIJ :PLP BOE .PSJ /P[PNV�
à4$(ûú B NJDSPUVCVMF EFTUBCJMJ[JOH GBDUPS TUJNVMBUFT UIF OFVSJUF
PVUHSPXUI CZ NPEVMBUJOH NJDSPUVCVMF EZOBNJDT JO SBU
IJQQPDBNQBM QSJNBSZ DVMUVSFE OFVSPOTá� *O� +PVSOBM PG
/FVSPCJPMPHZ ���ûú üúú� QQ� ûûúû×ûûûþ� juo�
��������OFV������� {xr�IUUQT���EPJ�PSH���������OFV�������
<üü> $IP (� FU BM� à4J[Oû *T B /PWFM 1SPUFJO_BU 'VODUJPOT BT B
5SBOTDSJQUJPOBM $PBDUJWBUPS PG #POF .PSQIPHFOJD 1SPUFJO
4JHOBMJOHá� *O� .PMFDVMBS BOE $FMMVMBS #JPMPHZ ü��ÿ %FD� üúú�
QQ� ûÿ�ÿ×ûÿ�ü� juo� ��������NDC���������� {xr�IUUQT���EPJ�PSH���������NDC����������
<üý> -J )� FU BM� à)VNBO ;$$)$ûü BDUJWBUFT "1�û BOE $3&# TJHOBMJOH
BT B USBOTDSJQUJPOBM DP�BDUJWBUPSá� *O� "DUB #JPDIJNJDB FU
#JPQIZTJDB 4JOJDB þû�� .BZ üúú� QQ� ÿýÿ×ÿþþ� juo�
��������BCCT�HNQ���� {xr�IUUQT���EPJ�PSH���������BCCT�HNQ����
<üþ> 5SBQOFMM $PMF FU BM� à%JVFSFOUJBM BOBMZTJT PG HFOF SFHVMBUJPO BU
USBOTDSJQU SFTPMVUJPO XJUI 3/"�TFRá� *O� /BUVSF #JPUFDIOPMPHZ
ýû�û %FD� üúûü QQ� þ�×ÿý� juo� ��������OCU������ {xr�IUUQT���EPJ�PSH���������OCU������
<üÿ> "LFO #SPOXFO -� FU BM� à_F &OTFNCM HFOF BOOPUBUJPO TZTUFNá�
*O� %BUBCBTF üúû� üúû� CBXú�ý� juo�
��������EBUBCBTF�CBX���� {xr�IUUQT���EPJ�PSH���������EBUBCBTF�CBX����
<ü�> 5SBQOFMM $PMF FU BM� à5SBOTDSJQU BTTFNCMZ BOE RVBOUJWDBUJPO CZ
3/"�4FR SFWFBMT VOBOOPUBUFE USBOTDSJQUT BOE JTPGPSN TXJUDIJOH
EVSJOH DFMM EJVFSFOUJBUJPOá� *O� /BUVSF #JPUFDIOPMPHZ ü��ÿ .BZ
üúûú QQ� ÿûû×ÿûÿ� juo� ��������OCU������ {xr�IUUQT���EPJ�PSH���������OCU������
<ü�> 5SBQOFMM $PMF FU BM� à%JVFSFOUJBM HFOF BOE USBOTDSJQU FYQSFTTJPO
BOBMZTJT PG 3/"�TFR FYQFSJNFOUT XJUI 5PQ)BU BOE $VZJOLTá� *O�
/BUVSF 1SPUPDPMT ��ý .BS� üúûü QQ� ÿ�ü×ÿ��� juo�
��������OQSPU���������� {xr�IUUQT���EPJ�PSH���������OQSPU����������
<ü�> $IFO &EXBSE : FU BM� à&OSJDIS� JOUFSBDUJWF BOE DPMMBCPSBUJWF
)5.-ÿ HFOF MJTU FOSJDINFOU BOBMZTJT UPPMá� *O� #.$
#JPJOGPSNBUJDT ûþ�û üúûý Q� ûü�� juo�
������������������������� {xr�IUUQT���EPJ�PSH��������������������������
View publication statsView publication stats

Pyk2 overexpression in postsynaptic neuronsblocks amyloid b1–42-induced synaptotoxicityin microfluidic co-cultures
Devrim Kilinc,1 Anaıs-Camille Vreulx,1 Tiago Mendes,1 Amandine Flaig,1
Diego Marques-Coelho,2,3 Maxime Verschoore,1 Florie Demiautte,1 Philippe Amouyel,1
Neuro-CEB Brain Bank, Fanny Eysert,1 Pierre Dourlen,1 Julien Chapuis,1
Marcos R. Costa,1,2 Nicolas Malmanche,1 Frederic Checler4 and Jean-Charles Lambert1
Recent meta-analyses of genome-wide association studies identified a number of genetic risk factors of Alzheimer’s disease; how-
ever, little is known about the mechanisms by which they contribute to the pathological process. As synapse loss is observed at the
earliest stage of Alzheimer’s disease, deciphering the impact of Alzheimer’s risk genes on synapse formation and maintenance is of
great interest. In this article, we report a microfluidic co-culture device that physically isolates synapses from pre- and postsynaptic
neurons and chronically exposes them to toxic amyloid b peptides secreted by model cell lines overexpressing wild-type or mutated
(V717I) amyloid precursor protein. Co-culture with cells overexpressing mutated amyloid precursor protein exposed the synapses
of primary hippocampal neurons to amyloid b1–42 molecules at nanomolar concentrations and induced a significant decrease in
synaptic connectivity, as evidenced by distance-based assignment of postsynaptic puncta to presynaptic puncta. Treating the cells
with antibodies that target different forms of amyloid b suggested that low molecular weight oligomers are the likely culprit. As
proof of concept, we demonstrate that overexpression of protein tyrosine kinase 2 beta—an Alzheimer’s disease genetic risk factor
involved in synaptic plasticity and shown to decrease in Alzheimer’s disease brains at gene expression and protein levels—selective-
ly in postsynaptic neurons is protective against amyloid b1–42-induced synaptotoxicity. In summary, our lab-on-a-chip device pro-
vides a physiologically relevant model of Alzheimer’s disease-related synaptotoxicity, optimal for assessing the impact of risk genes
in pre- and postsynaptic compartments.
1 Universite de Lille, Institut Pasteur de Lille, CHU Lille, INSERM U1167, LabEx DISTALZ, Lille 59019, France2 Brain Institute, Federal University of Rio Grande do Norte, Natal 59056-450, Brazil3 Bioinformatics Multidisciplinary Environment (BioME), Federal University of Rio Grande do Norte, Natal 59056-450, Brazil4 CNRS UMR7275 Laboratory of Excellence “Distalz”, IPMC, Universite Cote d’Azur, Inserm, Valbonne 06560, France
Correspondence to: Devrim Kilinc, Universite de Lille, Institut Pasteur de Lille, CHU Lille,
INSERM U1167, LabEx DISTALZ, 1 rue du Prof Calmette, BP245, Lille 59019, France
E-mail: [email protected]
Correspondence may also be addressed to: Jean-Charles Lambert
E-mail: [email protected]
Keywords: Alzheimer’s disease; synapses; microfluidics; co-culture; amyloid b
Abbreviations: Ab ¼ amyloid b; APP ¼ amyloid precursor protein; BA ¼ Brodmann area; CHO ¼ Chinese hamster ovary; DIV ¼
days in vitro; HRP ¼ horseradish peroxidase; MW ¼ molecular weight; NBA ¼ neurobasal A; PBS ¼ phosphate-buffered saline;PSD ¼ postsynaptic density; p-Pyk2 ¼ phospho-Pyk2 (Tyr402); Pyk2 ¼ protein tyrosine kinase 2 beta; RNAseq ¼ RNA sequencing;
RT ¼ room temperature
Received April 16, 2020. Revised July 12, 2020. Accepted August 3, 2020. Advance Access publication August 28, 2020VC The Author(s) (2020). Published by Oxford University Press on behalf of the Guarantors of Brain.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/), which
permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited. For commercial re-use, please contact
BRAIN COMMUNICATIONS
doi:10.1093/braincomms/fcaa139 BRAIN COMMUNICATIONS 2020: Page 1 of 18 | 1
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

IntroductionAlzheimer’s disease, the most common neurodegenerative
disorder worldwide, is characterized by two types of
brain lesions: (i) neurofibrillary degeneration due to the
intracellular aggregation of abnormally hyperphosphory-
lated tau protein and (ii) amyloid plaques resulting from
the extracellular accumulation of amyloid b (Ab) peptides
(Nisbet et al., 2015). Ab peptides are generated by the
cleavage of the transmembrane amyloid precursor protein
(APP) and can have different residue lengths (De
Strooper, 2010). The discovery of mutations in the APP,
PS1 and PS2 genes (coding for APP and presenilins 1
and 2) causing early-onset, autosomal-dominant forms of
Alzheimer’s disease has profoundly influenced our under-
standing of the disease and has placed Ab peptides at the
centre of the pathophysiological process. According to
the ‘amyloid cascade hypothesis’, the accumulation of Ab
peptides is the triggering toxic condition that induces the
development of neurofibrillary degeneration and thus
neuronal death (Hardy and Selkoe, 2002).
Ab1–42 species have been the principal focus of research
(Stine et al., 2003) since Ab1–42 is more prone to oligo-
merize (Dahlgren et al., 2002; Resende et al., 2008), and
oligomers of Ab1–42 are more toxic than its monomeric
or fibrillary forms, and other Ab species (Deshpande
et al., 2006; Ferreira et al., 2012; Spires-Jones and
Hyman, 2014). Oligomeric Ab1–42 is in a dynamic equi-
librium with the monomeric forms and fibrils and has
been proposed to be the main promoter of amyloid pla-
ques (Benilova et al., 2012). Although the central role of
the Ab peptide burden as initially enunciated in this hy-
pothesis is strongly debated, several lines of evidence indi-
cate that Ab peptides are still a key actor of the disease
at least via their oligomeric forms. In particular, the Ab
oligomer toxicity has been linked with synapse dysregula-
tion and loss (Brody and Strittmatter, 2018).
Synapse loss is a major pathological correlate of cogni-
tive deficits in Alzheimer’s disease (Lansbury, 1999) and
is observed at the earliest stage of the disease (Scheff
et al., 2007). Several mechanisms have been proposed to
explain Ab-induced synaptotoxicity: (i) membrane-disrupt-
ing activity at high concentrations (Sepulveda et al.,
2010); (ii) deleterious pruning of synapses by microglia
activation (Hong et al., 2016); and (iii) direct interaction
of oligomeric forms with postsynaptic receptors, such
as ionotrophic or metabotropic glutamate receptors
(Wang et al., 2004; Um et al., 2012). In addition, the
new genetic landscape of Alzheimer’s disease, resulting
from the advent of the genome-wide association studies
Graphical Abstract
2 | BRAIN COMMUNICATIONS 2020: Page 2 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

(Lambert et al., 2013; Kunkle et al., 2019), highlights
synaptic (dys)regulation: several among the dozens of
genes/loci identified to be associated with Alzheimer’s dis-
ease risk, e.g. BIN1, CD2AP, FERMT2 and PTK2B,
have been shown to modulate synaptic functions in the
physio- and/or pathophysiological contexts (Giralt et al.,
2017; Eysert et al., 2019; Ojelade et al., 2019; Salazar
et al., 2019; Schurmann et al., 2019). As a result, we re-
cently proposed a genetically driven synaptic failure hy-
pothesis, based on the genetic and post-genome-wide
association study data (Dourlen et al., 2019). In this con-
text, Ab toxicity is one of the elements involved in syn-
apse failure and it may be driven by specific Alzheimer’s
disease genetic risk factors.
However, assessing such an hypothesis requires a number
of considerations to be taken into account: (i) most in vitro
models of Ab toxicity use synthetic Ab oligomers at non-
physiological concentrations, even though synthetic fibrils
are structurally different from Ab fibrils obtained from
Alzheimer’s brains (Kollmer et al., 2019); (ii) only a few of
the genome-wide association study-defined genes have been
analysed in a (physiological and/or pathophysiological) syn-
aptic context; and (iii) genome-wide association study-
defined genes may have different effects when expressed in
the pre- or postsynaptic neurons. For example, protein tyro-
sine kinase 2 beta (Pyk2), product of Alzheimer’s disease
risk gene PTK2B, directly interacts with postsynaptic scaf-
fold proteins (Bartos et al., 2010), regulates dendritic spine
morphology (Giralt et al., 2017) and is involved in synaptic
plasticity through regulating postsynaptic NMDA receptors
via activation of Src (Huang et al., 2001). Recent studies
based on Alzheimer’s disease mouse models linked Pyk2’s
effects to amyloid pathology but reported contradictory
data on whether lack of Pyk2 was detrimental or protective
(Giralt et al., 2018; Salazar et al., 2019).
With this background, we have developed a microfluidic
co-culture device, based on existing tricompartment designs
that physically isolate synapses and provide exclusive access
to pre- and postsynaptic neurons (Taylor et al., 2010;
Virlogeux et al., 2018). This device permits not only the
induction of synaptotoxicity via physiologically relevant
concentrations of Ab molecules secreted by cells stably
overexpressing human APP but also the analysis of synap-
tic density as a function of over- or underexpression of
genetic risk factors in pre- and/or postsynaptic neurons. In
this article, we characterized Ab-induced synaptotoxicity in
primary neurons upon co-culture with cells overexpressing
mutated APP and assessed the impact of Pky2 overexpres-
sion in postsynaptic neurons, as a proof of concept.
Materials and methods
Oligomerization of synthetic Ab
peptides
Ab peptides were oligomerized according to established
protocols (Stine et al., 2003; Chang et al., 2012) with
minor modifications. The inactive control peptide (Ab42–1;
Abcam, Cambridge, UK) has the same composition as
the Ab1–42 peptide (California Peptide Research; Napa,
CA, USA), but with an inverted amino-acid sequence,
and has been widely used as control for oligomeric Ab1–
42 since it is also prone to oligomerization (Walsh et al.,
2002; Xiong et al., 2007; Mairet-Coello et al., 2013).
Ab1–42 and Ab42–1 were treated with hexafluoroisopropa-
nol (Sigma-Aldrich, Saint Louis, MO, USA) to maintain
the oligomeric structure and to reduce fibril formation
(Stine et al., 2003), according to manufacturer’s instruc-
tions. The peptides were resuspended in 1ml hexafluoroi-
sopropanol and incubated for 1 h at room temperature
(RT), with occasional moderate vortexing, followed by
sonication in water bath (Branson; Emerson Electric, St.
Louis, MO, USA) for 10min. The solution was aliquoted
into microcentrifuge tubes, let to evaporate in a chemical
fume hood, dried with SpeedVac system (Thermo Fisher
Scientific, Waltham, MA, USA) for 30min, and stored as
desiccated peptide at �80�C. To produce oligomers,
lyophilized, hexafluoroisopropanol-treated aliquots of
both peptides were resuspended in dimethyl sulphoxide
(DMSO) to reach 5mM, mixed by pipetting, sonicated in
water bath for 10min and diluted to 100 mM in ice-cold
Dulbecco’s phosphate-buffered saline (PBS), followed by
30 s vortexing and 1 h incubation at 25 �C. The solutions
were aliquoted into microcentrifuge tubes and stored at
�20�C for up to 4 weeks. Before adding to neurons, oli-
gomeric peptide stocks were thawed, serial diluted to
100 nM in 2% dimethyl sulphoxide in PBS. Ab1–42 con-
centrations up to 1 mM are considered non-lethal (Kelly
and Ferreira, 2007; Kuperstein et al., 2010).
Microfluidic device design andfabrication
The microfluidic co-culture device was designed based on
a previous tricompartmental neuron culture device (Kilinc
et al., 2014). The device consists of a 300-lm-wide, 7.4-
mm-long central channel flanked by two 750-lm-wide,
3.6-mm-long side channels. All three channels are ca.
100-mm high. The left side channel (termed presynaptic)
and the central channel (termed synaptic) are intercon-
nected via 4-mm-high, 450-mm-long parallel microchannels
that narrow from an entry width of 10 lm to an exit
width of 3 lm. The right side channel (termed postsynap-
tic) and the synaptic chamber are also interconnected via
parallel microchannels with identical dimensions, except
that they were 75-lm long. One end of the synaptic
chamber bifurcates into two branches, one of which ter-
minates in a triangular shape. This terminus is connected
to a diamond-shaped co-culture chamber (based on a pre-
vious design) (Kilinc et al., 2016) via 4-lm-high, 10-lm-
wide and 100-lm-long parallel microchannels.
Master patterns were fabricated at the Institute of
Electronics, Microelectronics and Nanotechnology (Lille,
France) via two-step photolithography (Blasiak et al., 2015).
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 3 of 18 | 3
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

Ca. 4-mm high polydimethysiloxane pads were replica
moulded. Access wells were punched at the termini of
the central channel and the co-culture chamber and of
the side channels using 3-mm and 4-mm biopsy punches
(Harris Unicore), respectively. The devices were perman-
ently bonded to 24mm � 50mm glass coverslips
(Menzel) via O2 plasma (Diener, Ebhausen, Germany).
Prior to cell culture, the devices were sterilized under
ultraviolet light (Light Progress, Anghiari, Italy) for
30min, treated with 0.1mg/ml poly-L-lysine (Sigma) over-
night and rinsed with PBS.
Primary neuron culture
Culture media and supplements were from Thermo
Fisher, unless mentioned otherwise. Primary neurons were
obtained from P0 rats, according to previously described
procedures (Sartori et al., 2019). Briefly, cortices and hip-
pocampi were isolated from new-born rats, washed with
ice-cold dissection medium (Hanks’ balanced salt solution
supplemented with HEPES, sodium pyruvate, and penicil-
lin/streptomycin) and trypsinized (2.5%; at 37 �C for
10min). Trypsin was inactivated with dissociation me-
dium—minimum essential media supplemented with foetal
bovine serum, GlutaMAX, D-glucose (Sigma), minimum
essential media vitamins and Pen/Strep—followed by
DNase (5mg/ml; Sigma) incubation for 1min and wash
with dissection medium. Medium was replaced by dissoci-
ation medium and tissue was triturated with a fire-pol-
ished cotton-plugged Pasteur pipette to obtain a
homogenous cell suspension, followed by centrifugation
(200 � g for 5min). Cells were resuspended in culture
medium [neurobasal A (NBA) supplemented with
GlutaMAX and B27 neural supplement with antioxi-
dants], counted and plated at a density of 100 000 cells/
cm2 in 6- and 24-well plates for immunoblots and in 10-
cm Petri dishes for synaptosome extraction. Plates were
pre-coated with 0.1mg/ml poly-L-lysine in 0.1M borate
buffer (0.31% boric acid, 0.475% sodium tetraborate,
pH ¼ 8.5; Sigma) overnight at 37�C and rinsed thor-
oughly with water. Alternatively, cells were plated in pre-
coated 384-well plates at 50 000 cells/cm2 (ca. 4000
cells/well) and in microfluidic devices at a density of ca.
8� 105 cells/cm2. After 20–24 h, culture medium was
replaced with supplemented NBA medium. The 0.1% eth-
ylenediaminetetraacetic acid (in H2O) was added to the
Petri dishes containing microfluidic devices to minimize
evaporation. Cultures were maintained in a tissue culture
incubator (Panasonic; Osaka, Japan) at 37�C and 5%
CO2.
Immunoblotting
Neurons were harvested in minimum volume of
200 ll/well in 6-well plates, in ice-cold lysis buffer as
described earlier (Chapuis et al., 2017). Lysates were
mixed with 4� lithium dodecyl sulfate (LDS) sample
buffer (Novex; Thermo Fisher) and 10� reducing agent
(Novex), sonicated and boiled at 95�C for 10min.
Samples were loaded at maximum volume into 1.5mm,
10-well, 4–12% Bis–Tris pre-cast NuPage gels (Novex),
along with 5 ll molecular weight (MW) marker (Novex
Sharp pre-stained protein standard, Thermo Fisher). The
gel was run with 2-(N-morpholino)ethanesulphonic acid
running buffer at 200V for 45min and transferred to
0.22-lm nitrocellulose membranes using the Trans-Blot
Turbo transfer system (BioRad, Hercules, CA, USA) using
mixed MW method at 1.3A and 25V for 7min.
Membranes were blocked in 0.05% Tween 20, 20mM
Tris-Base, 150mM NaCl, pH ¼ 8.0 (TNT) containing
5% nonfat dry milk for 1 h at RT and washed 3� in
TNT. Membranes were incubated with the following pri-
mary antibodies in SuperBlock T20 blocking buffer
(Thermo Scientific) at 4�C overnight and washed 3� in
TNT: rabbit anti-phospho-PTK2B (3291; 1/1000; Cell
Signaling Technology, Danvers, MA, USA), rabbit anti-
PTK2B (P3902; 1/1000; Sigma) and mouse anti-b-actin
(A1978; 1/5000; Sigma). Membranes were then incubated
with horseradish peroxidase (HRP)-conjugated secondary
antibodies (HRP-anti-mouse and HRP-anti-rabbit; 1:5000;
Jackson ImmunoResearch, West Grove, PA, USA) in
TNT containing 5% nonfat dry milk for 1 h at RT and
washed 3� in TNT. The membrane was revealed through
chemiluminescence (Luminata Classico, EMD Merck
Millipore) and imaged with Amersham Imager 600 (GE
Healthcare, Mississauga, Canada). The images were
quantified with ImageQuant TL Software (GE
Healthcare).
Culture of Chinese hamster ovarycells and analysis of their media
Chinese hamster ovary (CHO) cell lines (CHO-pcDNA4,
-APPWT and -APPLDN) were maintained according to pre-
viously described procedures (Guillot-Sestier et al., 2012).
Cells were grown in the CHO growth medium:
Dulbecco’s minimum essential medium/Ham’s F-12 1:1
medium, supplemented with 10% heat-inactivated foetal
bovine serum, 0.2% Pen/Strep, 2% HT supplement and
300 mM Proline (Sigma). To stimulate Ab production, the
growth medium was replaced with CHO-NBA medium:
phenol red-free NBA (Gibco) supplemented with 0.2%
Pen/Strep, 2% HT supplement and 300mM Proline.
For media collection, cells were grown in 10-cm Petri
dishes or in 6-well plates until they reached 80% conflu-
ence, at which point the maintenance medium was rinsed
with PBS and replaced with the stimulation medium.
After 48 h of stimulation, the medium was collected into
15-ml Falcon tubes and centrifuged at 4000 � g and 4�C
for 10min to remove the debris. The supernatant was
loaded into a 3-kDa spin column (Amicon Ultra; Merck),
equilibrated with Neurobasal (without serum or Phenol
Red) at 4000 � g and 4�C for 10min and concentrated
at 4000 � g and 4�C for 1 h. Western blotting of
4 | BRAIN COMMUNICATIONS 2020: Page 4 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

conditioned media was performed as described with the
following exceptions: the transferred membrane was
boiled for 5min in PBS and Luminata Crescendo
(Millipore) was used as the HRP substrate. Anti-Ab1–42(clone 6E10; 1:1000; Sigma) was used as primary
antibody.
Exposure of neurons to conditionedmedia
Total protein concentration in the conditioned media col-
lected from different CHO cell lines (CHO-pcDNA4, -
APPWT and -APPLDN) was assessed using the Pierce BCA
Protein Assay Kit (Thermo Fisher) and adjusted to
100 mg/ml with supplemented CHO-NBA medium. A total
of 10 ml of conditioned media (per well) was added to
primary neuronal cultures grown in 384-well plates (con-
taining 40 ml of culture medium per well) at 14 days
in vitro (DIV14), DIV18, DIV19, DIV20 and DIV21 (6 h
prior to media collection). Media from these wells (eight
wells per condition) were collected into a new 384-well
plate prior to the quantification of Ab peptides.
Co-culture of neurons with CHOcells
CHO cells were seeded in the co-culture chamber at a
density of ca. 1.3� 105 cells/cm2 and maintained in a tis-
sue culture incubator (5% CO2; 37�C) in CHO medium.
On the day of primary neuron culture, the medium was
replaced with CHO-NBA medium supplemented with 1%
GlutaMAX and 2% B27 neural supplement with antioxi-
dants. Primary neurons were seeded in the pre- and post-
synaptic chambers. At DIV1, the medium in all wells was
replaced with fresh supplemented CHO-NBA medium.
Every 3–4 days, the medium in the access wells of the co-
culture chamber was replaced with fresh medium, where-
as the medium in other wells was only topped up with
fresh medium.
Lentiviral transductions
At DIV7, neurons cultured in the postsynaptic chamber
were transducted with lentiviruses according to estab-
lished methods.(Sartori et al., 2019) To avoid the trans-
duction of CHO cells and the neurons in the presynaptic
chamber, a hydrostatic pressure gradient was formed
across the microchannels separating synaptic and postsy-
naptic chambers. The following lentiviruses were used for
transduction: Mission shRNA vectors (Sigma) pLenti6/
Ubc/v5-DEST (Invitrogen, Carlsbad, CA, USA) empty
(Mock) or including human PTK2B cDNA sequences,
synthesized via the GeneArt service (Thermo Fisher).
LifeAct-Ruby (pLenti.PGK.LifeAct-Ruby.W; RRID:
Addgene_51009) and LifeAct-GFP (pLenti.PGK.LifeAct-
GFP.W; RRID: Addgene_51010) were kind gifts from
Rusty Lansford. Viral transduction was performed at the
multiplicity of infection of four. Constructs were diluted
in pre-warmed, supplemented CHO-NBA medium con-
taining 2 lg/ml (5�) Polybrene (hexadimethrine bromide;
Sigma). Media from pre- and postsynaptic wells were col-
lected in a common tube. A total of 25, 15 and 20 ml of
the collected media were returned to each presynaptic,
synaptic and postsynaptic wells, respectively. Ten micro-
litres of virus suspension was added to one of the postsy-
naptic wells. Neurons were incubated with viral particles
for 6 h before the wells were topped up with the remain-
der of the collected media. Co-cultures were maintained
as described earlier.
Alpha-LISA measurements
Alpha-LISA is a highly sensitive, quantitative assay based
on biotinylated antibody bound to streptavidin-coated
donor beads and antibody-conjugated acceptor beads. In
the presence of the analyte, the beads come into close
proximity such that the excitation of the donor beads
triggers a cascade of energy transfer in the acceptor
beads, resulting in a sharp peak of light emission at
615 nm. We used Alpha-LISA kits specific to human
Ab1–X and Ab1–42 (AL288C and AL276C, respectively;
PerkinElmer, Waltham, MA, USA) to measure the
amount of Ab1–X and Ab1–42 in culture media. The
human Ab analyte standard was diluted in the CHO-
NBA medium. For the assay, we first added 5ml of cell
culture supernatant or standard solution into an
Optiplate-96 microplate (PerkinElmer). We then added
5 ml of 10� mixture including acceptor beads and bioti-
nylated antibody. Following incubation at RT for 60min,
we added 40 ml of 1.25� donor beads and incubated at
RT for 60min. We measured the fluorescence using an
EnVision-Alpha Reader (PerkinElmer) at 680-nm excita-
tion and 615-nm emission wavelengths. In experiments
where conditioned media were added to primary neurons
in 384-well plates, 2 ml of collected media was transferred
to an Optiplate-384 (PerkinElmer) and Alpha-LISA was
performed using reduced volumes: 2 ml of sample or
standard, 2 ml of acceptor beads and biotinylated anti-
body mix and 16 ml of donor beads.
Enzyme-linked immunosorbentassay measurements
The sandwich enzyme-linked immunosorbent assay was
performed according to the manufacturer’s protocol. We
used microtiter plates pre-coated with anti-human Ab35–
40 antibody (clone 1A10; RE59781, IBL) and anti-human
Ab38–42 (clone 1C3; RE59721, IBL) to detect Ab1–40 and
AbX–42, respectively. Plates were incubated overnight at
4�C with 100 ml of cell culture supernatant or with stand-
ards. The bound antigen was detected by incubating the
wells with 100 ml of 30� mixture containing the HRP-
conjugated anti-human Ab antibodies (clone 82E1 for
Ab1–40 and clone 12B2 for AbX–42) for 60min at 4�C.
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 5 of 18 | 5
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

Signal was revealed by incubating with the 3,30,5,50-
Tetramethylbenzidine (TMB) substrate for 30min at RT
in the dark and stopping the enzymatic reaction with the
TMB stop solution containing 1N H2SO4. Signal inten-
sity was read immediately at 405 nm via a microplate
reader (PowerWave XS2; BioTek Instruments, Winooski,
VT, USA).
Neuronal viability assay usingconventional co-cultures
Cell culture inserts (Falcon 353104; Corning, Corning,
NY, USA) were placed in 24-well plates and pre-incu-
bated with 500 ll of CHO growth medium for 30min.
CHO cells (CHO-pcDNA4, -APPWT and -APPLDN) were
plated at a density of 4000 cells/insert and incubated at
37�C and 5% CO2 for 3 days, during which the cells
reached confluence. CHO growth medium was then
replaced with CHO-NBA medium supplemented with 1%
GlutaMAX and 2% B27 with antioxidants and primary
neurons were plated in the 24-well plate containing the
inserts at a density of 2� 105 cells/well. As control condi-
tions, primary neurons were plated in the wells with
inserts without CHO cells and CHO cells were plated in
inserts without primary neurons, under otherwise identi-
cal conditions. After 24 h, the culture medium was
replaced with fresh, supplemented CHO-NBA medium.
Co-cultures were maintained in a tissue culture incubator
at 37�C and 5% CO2 for 14 days. Two hundred micro-
litres of medium was collected from the wells at DIV1,
DIV7 and DIV14 for lactate dehydrogenase release assay.
Lactate Dehydrogenase Cytotoxicity Detection Kit
(Takara Bio, Saint-Germain-en-Laye, France) was used
according to the manufacturer’s protocol. Briefly, col-
lected culture medium was incubated with reaction mix-
ture at 1:1 ratio for 30min (in dark, at RT) and the
lactate dehydrogenase enzymatic activity was subsequent-
ly measured with a microplate reader at 490 nm
(PowerWave XS2; BioTek Instruments). Cell viability was
calculated as a percentage of the difference between nega-
tive controls (primary neurons without CHO cell co-cul-
ture treated with 0.1% Triton X-100 for 5min) and
positive controls (primary neurons without CHO cell co-
culture). The effect of CHO cell co-culture on neuronal
viability was determined by subtracting the respective
CHO cell monoculture signal from the co-culture signal.
The presence of Ab1–42 in the conventional co-culture
media was measured via Alpha-LISA as described.
Immunocytochemistry andmicroscopy
Co-cultures were fixed at DIV14 in PBS containing 4%
paraformaldehyde for 20min at RT and permeabilized
with 0.3% (v/v) Triton X-100 in PBS for 5min. After
blocking in 5% (w/v) normal donkey serum, samples
were incubated overnight at 4�C with the following
antibodies: mouse anti-MAP2 (188011; Synaptic Systems,
Gottingen, Germany); chicken anti-Homer (160006;
Synaptic Systems); and rabbit anti-PYK2 phospho-Y402
(ab4800; Abcam). Cells were rinsed with PBS and incu-
bated with the following secondary antibodies for 2 h at
RT: Dylight 405 Donkey anti-mouse (715-475-151;
Jackson); AlexaFluor 647 donkey anti-rabbit (711-605-
152; Jackson); and AlexaFluor 594 Donkey anti-chicken
(703-585-155; Jackson). Cells were rinsed with PBS and
incubated with mouse monoclonal anti-Synaptophysin 1
(Syp) pre-labeled with Sulfo-Cyanine 2 (101011C2;
Synaptic Systems) for 2 h at RT. Cells were rinsed with
PBS and microfluidic devices were topped with 90%
glycerol.
Samples were imaged under a LSM 880 confocal
microscope (Zeiss, Oberkochen, Germany) equipped with
a 63� 1.4 numerical aperture objective. Images were
acquired at zoom 2 with a z-stack interval of 0.5 mm.
Typically, four images were acquired from each synapse
chamber, each showing multiple dendrites arriving from
the postsynaptic chamber. Images were deconvoluted
using AutoQuantX3 software (BitPlane, Zurich,
Switzerland) for synaptic connectivity analysis.
In a subset of experiments, neurons were plated only in
the presynaptic chamber, only in the postsynaptic cham-
ber or only in the synaptic chamber. Neurons were fixed
and immunostained at DIV14 as described to reveal nu-
clei, glial fibrillary acidic protein (ABD95; Millipore) and
b3-tubulin (MAB1637; Millipore). All chambers were
imaged using a Zeiss AxioObserver Z1 epifluorescense
microscope equipped with a Prime 95B Scientific CMOS
(Photometrics, Tucson, AZ, USA) camera and 32� ob-
jective. The integrated density (pixel values � area) of
the b3-tubulin signal was quantified in all chambers, at a
distance between 50 and 200lm from the entries and
exits of the microchannels. Neurite penetration rates for
short and long microchannels (in the forward and reverse
directions) were estimated by taking the ratio of the b3-
tubulin-integrated density signals obtained from the emit-
ting and receiving chambers. The ratio between the pene-
tration rates between forward and reverse directions was
defined as the directionality ratio.
Quantification of synapse integrity
We developed an image analysis workflow based on
image segmentation using Imaris software (BitPlane) and
assignment of postsynaptic signals to the nearest pre-
synaptic signal using a custom Matlab (Mathworks,
Natick, MA, USA) code. Briefly, signals obtained for pre-
and postsynaptic structures were first deconvoluted in
Autoquant X3 (Media Cybernetics, Rockville, MD, USA)
and segmented into distinct volumes using the surfaces
function of Imaris in the batch mode that permits the
same parameters to be applied to all images
(Supplementary Fig. 1A–D). Postsynaptic spots were then
assigned to the nearest presynaptic spot (according to the
6 | BRAIN COMMUNICATIONS 2020: Page 6 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

three-dimensional Euclidean distance between intensity
centres) within a pre-defined cut-off distance
(Supplementary Fig. 1E). It is important to note that pre-
and postsynaptic structures are smaller than the axial
resolution of the confocal microscope; the distances meas-
ured are thus not true physical distances between synap-
tic structures but the distances between respective
intensity centres. We empirically determined the cut-off
distance by testing a range of values on a large set of
control cultures (Supplementary Fig. 1F and G). The frac-
tion of Syp puncta with at least one Homer assignments
and the average number of Homer assignments per Syp
were determined to be the most robust read-outs of syn-
apse connectivity. The numbers of assigned pre- and
postsynaptic puncta per image area (in the xy-plane)
were also provided. Note that a small fraction of control
samples for CHO-APPWT and CHO-APPLDN co-cultures
(three and two devices, respectively) do overlap in the
data reported.
Analysis of phospo-Pyk2 signals
relative to synapses
We extended the image analysis workflow to analyse the
positions of phospho-Pyk2 (Tyr402) (p-Pyk2) puncta rela-
tive to the positions of identified synapses. Postsynaptic
spots were first assigned to the nearest presynaptic spot
as described. For each postsynaptic-to-presynaptic assign-
ment, or ‘synapse’, we defined the midpoint as being
equidistant to pre- and postsynaptic puncta. We next
defined pre- and postsynaptic zones, by spanning two
right circular cones with 45� polar angle where the mid-
point is the apex. We then associated p-Pyk2 puncta with
the nearest ‘synapse’ as long as it was within a pre-
defined cut-off distance from the midpoint and catego-
rized these associations as presynaptic, postsynaptic or
others (neither pre- nor postsynaptic). We then calculated
the average number of p-Pyk2 puncta associated with
each ‘synapse’. As each ‘synapse’ inherently consists of a
Syp and Homer pair, a paired statistical test was justified
for this analysis.
Synaptosome extraction
To verify the presence of proteins at the synaptic level,
we conducted subcellular fractionation as previously
described (Frandemiche et al., 2014). Briefly, cortical neu-
rons were cultured in 10-cm Petri dishes as described
(3.5–4.0� 107 neurons per condition). At DIV13, neurons
were exposed to CHO-APPWT or CHO-APPLDN media
for 18 h (final Ab1–X concentration: 40 nM). At the end
of this treatment, neurons were lysed, reconstituted in a
solution (0.32M sucrose and 10mM HEPES, pH ¼ 7.4)
and centrifuged at 1000 � g for 10min to remove nuclei
and debris. The supernatant was centrifuged at 12 000 �
g for 20min to remove the cytosolic fraction. The pellet
was reconstituted in a second solution (4mM HEPES,
1mM ethylenediaminetetraacetic acid, pH ¼ 7.4) and
was centrifuged 2� at 12 000 � g for 20min. The new
pellet was reconstituted in a third solution (20mM
HEPES, 100mM NaCl, 0.5% Triton X-100, pH ¼ 7.2)
for 1 h at 4�C and centrifuged at 12 000 � g for 20min.
The supernatant collected corresponds to the non-postsy-
naptic density (PSD) fraction (Triton-soluble). The
remaining pellet was reconstituted in a fourth solution
(20mM HEPES, 0.15mM NaCl, 1% Triton X-100, 1%
deoxycholic acid, 1% sodium dodecyl sulfate (SDS), pH
¼ 7.5) for 1 h at 4�C and was centrifuged at 10 000 � g
for 15min to obtain a supernatant containing the PSD
fraction (Triton insoluble). The fractions obtained were
then analysed by western blot (WB). There was no differ-
ence in the Syp signal in the non-PSD fraction and the
PSD95 signal in the PSD fraction for neurons treated
with CHO-APPWT and CHO-APPLDN media, suggesting
that pre- and postsynaptic proteins were not affected by
this treatment paradigm (Supplementary Fig. 2).
Analysis of gene expression
alterations in human brain samples
RNA sequencing (RNAseq) data from the Mount Sinai/JJ
Peters VA Medical Center Brain Bank (MSBB) (Wang
et al., 2018), the ROSMAP database (De Jager et al.,
2018) and the Mayo Clinic whole genome and transcrip-
tome data (Allen et al., 2016) were downloaded from
AMP-AD Knowledge Portal (https://adknowledgeportal.syn
apse.org/) according to the terms and conditions concern-
ing the use of the data. Data were aligned using pseudoa-
ligner Kallisto version 0.43.1 (Bray et al., 2016) using a
pre-built index to align fastq files. Differential gene expres-
sion analysis was performed using DESeq2 (Love et al.,
2014). First, a DESeq2 object was created using
DESeqDataSetFromTximport function and rows with sum
of all counts <10 were filtered out. Next, DESeq function
was used with default parameters. Temporal cortex was
analysed in the Mayo Clinic dataset (82 cases and 78
healthy controls). Dorsolateral prefrontal cortex was ana-
lysed in the ROSMAP dataset (222 cases and 201 con-
trols). The following brain areas were analysed in the
MSBB dataset: Brodmann area (BA) 10, which corre-
sponds to the anterior prefrontal cortex (105 cases and 71
controls); BA 22, which is part of the Wernicke’s area in
the superior temporal gyrus (98 cases and 61 controls);
BA 36, which corresponds to the lateral perirhinal cortex
(88 cases and 64 controls); and BA 44, which corresponds
to the inferior frontal gyrus (90 cases and 63 controls).
Analysis of phosphorylated and
total Pyk2 levels in human brain
samples
The brain samples were collected through a brain dona-
tion programme dedicated to neurodegenerative dementias
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 7 of 18 | 7
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

coordinated by the NeuroCEB Brain Bank Network. The
informed consent for post-mortem examination and re-
search studies was signed by the legal representative of
each patient in patient’s name, as allowed by the French
law and approved by the local ethics committee and the
brain bank has been officially authorized to provide sam-
ples to scientists (agreement AC-2013-1887). All proce-
dures performed in this study involving human
participants were in accordance with the ethical standards
of the institutional research committees and with the
1964 Declaration of Helsinki. The brain banks fulfil cri-
teria from the French Law on biological resources includ-
ing informed consent, ethics review committee and data
protection (article L1243-4 du Code de la Sante publique,
August 2007). The Neuro-CEB brain bank (BioResource
Research Impact Factor number BB-0033-00011) has
been declared to the Ministry of Research and Higher
Education, as required by French law.
Assessment of Alzheimer’s disease-related neurofibrillary
pathology (Braak stage) was performed according to pub-
lished procedures (Thierry et al., 2020) by analysing
post-mortem brain tissue samples of 28 individuals
(Supplementary Table 1) via immunohistochemistry
against Ab deposits (Dako M0872; clone 6 F/3D; Agilent,
Santa Clara, CA, USA) and against hyperphosphorylated
Tau at Ser202/Thr205 (clone AT8; Thermo Fisher)
(Braak et al., 2006). Lysis buffer, containing trizma-base
20mM, NaCl 150mM, cOmplete Protease Inhibitor
Cocktail 1� and 1% Triton X-100, was added to single
pieces of whole brain tissue (�100mg) at a ratio of 5 ll
per 1mg of tissue. Brain samples were homogenized by
beads beating using a precellys soft tissue CK14 2ml (3�
30 s at 6500 rpm). The lysate was then centrifuged at
4000 rpm for 15min at 4�C. Fifty microlitres from the
supernatant was used for analysis. Protein quantification
was performed using BCA protein assay. Total proteins
(40 mg/lane) were separated on 4–12% Bis–Tris-polyacryl-
amide gel electrophoresis (NuPAGE; Thermo Scientific)
under reducing conditions and subsequently blotted onto
nitrocellulose membranes using iBlot 2 Dry Blotting
System (BioRad). Primary antibodies against phospho-
Pyk2 Tyr 402 (1:1000, cat. no. 3291; Cell Signalling),
total Pyk2 (1:1000, cat. no. P3902; Sigma) and b-actin
(1:5000, cat. no. ab8226; Abcam) were used for im-
munoblotting. After incubation with the appropriate
HRP-conjugated secondary antibodies, the protein bands
were detected using ImageJ. Samples outside of 3� me-
dian absolute deviations were deemed outliers and were
excluded from the analysis.
Statistical analysis
Synapse connectivity data were analysed using Kruskal–
Wallis ANOVA, followed by Wilcoxon rank-sum test to
compare individual groups. The statistical unit was
microfluidic device. Synapse data were pooled after nor-
malization by the mean of control group for each
primary neuron preparation. Human brain data were
analysed using Wilcoxon rank-sum test between cases
and controls. Other data were analysed using unpaired
or paired t-test as appropriate. A P-value of <0.05 was
considered statistically significant.
Data availability
All data generated or analysed during this study are
included in this published article and its Supplementary
material.
Results
Microfluidic co-culture device to
expose hippocampal synapses to
synthetic or cell-secreted Ab
oligomers
With the goal to better mimic Alzheimer’s disease
in vitro, we developed a microfluidic device that permits
synapse formation between two sets of neurons cultured
in distinct chambers. Based on a previous design, our de-
vice consists of three distinct chambers, interconnected
via parallel microchannels that constrain neuronal cell
bodies but permit axons and dendrites to cross through
(Fig. 1) (Kilinc et al., 2015). Adjusting the lengths of
microchannels made it possible to allow axons and den-
drites from the right (or ‘postsynaptic’) chamber, but
only axons from the left (or ‘presynaptic’) chamber to
reach the central (or ‘synaptic’) chamber (Fig. 1C)
(Taylor et al., 2010). As a novel design feature, one end
of the synaptic chamber bifurcates, where one branch ter-
minates with an access well and the other one connects
to a fourth (co-culture) chamber via a dense series of
short microchannels. In addition, we employed narrowing
microchannels that promote unidirectional neurite cross-
ing (Peyrin et al., 2011). When rat postnatal hippocampal
neurons were cultured in one chamber only
(Supplementary Fig. 3), 2.9� more axons crossed the
long microchannels in the forward direction (from the
wide end towards the narrow end) than in the reverse
direction. This figure decreased to 1.9� in the case of
short microchannels.
At DIV14, no dendrites emanating from neurons cul-
tured in the presynaptic chamber were observed in the
synaptic chamber. Axons from these neurons, however,
invaded the entire synaptic chamber. A total of 11.0 6
4.0% of these axons crossed the short microchannels in
the reverse direction and reached the postsynaptic cham-
ber, as measured by the ratio of the b3-tubulin fluores-
cence between the emitting and receiving chambers. On
the other hand, axons and dendrites from neurons cul-
tured in the postsynaptic chamber fully invaded the syn-
aptic chamber by DIV14. None of these dendrites and
8 | BRAIN COMMUNICATIONS 2020: Page 8 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

only 5.9 6 1.6% of these axons crossed the long micro-
channels in the reverse direction and reached the pre-
synaptic chamber. Synapse formation in the synapse
chamber was confirmed by immunostaining against Syp
and Homer 1, pre- and postsynaptic markers, respectively
(Fig. 1E). In summary, the synaptic chamber receives
axons from both pre- and postsynaptic chambers, receives
dendrites only from the postsynaptic chamber and con-
tains 83.2 6 6.1% of all synaptic connections formed be-
tween pre- and postsynaptic chambers.
Effect of synthetic Ab42 oligomerson synapse connectivity
We oligomerized synthetic Ab1–42 and Ab42–1 (inverted
control peptide) and assessed the presence of oligomeric
species via Coomassie blue staining. We confirmed the
presence of low-MW oligomers in the Ab1–42 sample
(Fig. 2A). We exposed mature synapses (DIV14) to
synthetic Ab peptides by adding the oligomer solution to
the synapse chamber to reach an initial concentration of
100 nM. We kept the media levels in the synaptic reser-
voirs lower than those in the pre- and postsynaptic reser-
voirs. The hydrostatic pressure difference induced a flow
through the microchannels countering the molecular dif-
fusion, which localized the treatment initially to the syn-
aptic chamber. However, as the pressure-driven flow
ceased, the media levels equilibrated and Ab peptides dif-
fused throughout the device. Since the short microchan-
nels offer little fluidic resistance compared to the long
microchannels, it would be safe to assume that the media
levels equilibrated first between the synaptic and postsy-
naptic chambers and then between the synaptic and pre-
synaptic chambers.
At the end of the 16-h treatment period, the neurons
were fixed and immunostained against pre- and postsy-
naptic markers. To quantitatively analyse synaptic con-
nectivity, we developed an image analysis workflow
Figure 1 Design and operating principle of the microfluidic co-culture device. (A) Photograph of the microfluidic device bonded to a
coverslip. (B) The layout of the device showing the presynaptic (pre), synaptic (syn) and postsynaptic (post) chambers, as well as the co-culture
chamber housing the CHO cells. Overlays show immunofluorescence images of primary neurons and CHO cells in their respective chambers,
stained for a-tubulin (red) and MAP2 (green). (C) Subcellular compartmentalization of neurons was shown by immunostaining against b3-tubulin
(red) and MAP2 (green), axonal and somatodendritic markers, respectively. Cell bodies were stained with Hoechst (blue). Microchannel
structure is evident in the brightfield image of the same area. (D) 15� magnification of the square marked in B, showing CHO cells cultured in
the co-culture chamber. CHO cells pass through the microchannels but do not migrate up the synaptic channel. (E) Synapse formation in the
synaptic chamber was evidenced by the localization of Synaptophysin 1 (yellow) and Homer 1 (cyan) puncta, pre- and postsynaptic markers,
respectively, around MAP2-positive dendrites (magenta). Boxed area is 3� magnified.
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 9 of 18 | 9
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

Figure 2 Synthetic and cell-secreted Ab1–42 induce different levels of synaptic toxicity. (A) Coomassie blue stain of in-house
oligomerized synthetic Ab1–42 and Ab42–1 (inverted control) peptides shows the presence of low molecular weight oligomers. (B and C)
Exemplary images and synaptic read-outs 16 h after the introduction of synthetic oligomer solution into the synaptic chamber at DIV14 (final
concentration ¼ 100 nM). Each Homer spot was assigned to the nearest Syp spot within a cut-off distance of 1.0 lm (see Supplementary Fig. 1
for details). Following the assignment of all Homer spots, the fraction of Syp spots assigned by at least one Homer spot and the average number
of Homer assignments per Syp spot were calculated. Densities of all Syp and Homer spots detected (white), Syp spots assigned by a Homer spot
(grey) and Homer spots assigned to a Syp spot (grey) are also shown. (D) Immunoblots of CHO cell media 2 days after stimulation. (E) Relative
Ab1–42 concentration in off-chip CHO cell media quantified via ELISA. c-Secretase inhibitor DAPTwas applied at 18 lg/ml for 5 days. Data
normalized to APPWT control condition. As the sample was concentrated prior to Ab1–42 measurement, Ab1–42 levels cannot be directly
compared to Ab1–40 levels. (F) Alpha-LISA measurement of Ab1–X and Ab1–42 in media collected from the top well of the synaptic chamber at
DIV14. c-Secretase inhibitor DAPTwas applied at 18 lg/ml for 5 days. One-way ANOVA, followed by unpaired t-test. (G and H) Exemplary
images and synaptic read-outs following co-culture with CHO cells at DIV14. In box plots, red circles, red bars, black squares and red plus signs
indicate individual data points, sample median and mean and outliers, respectively. Numbers of microfluidic devices analysed (obtained from at
least three independent cultures) are given in parentheses. Kruskal–Wallis ANOVA, followed by Wilcoxon rank-sum test. *P< 0.05; **P< 0.005.
N/S ¼ not significant; ELISA ¼ enzyme-linked immunosorbent assay.
10 | BRAIN COMMUNICATIONS 2020: Page 10 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

based on scanning confocal microscopy, software-assisted
identification of pre- and postsynaptic puncta and prox-
imity-based assignment of postsynaptic puncta to pre-
synaptic puncta (Supplementary Fig. 1). Briefly, each
Homer spot was assigned to the nearest Syp spot within
a cut-off distance of 1.0lm, which was pre-determined
using a training set. The fraction of Syp puncta with at
least one Homer assignments and the average number of
Homer assignments per Syp were determined to be the
most robust read-outs of synapse connectivity (see
Materials and Methods for details). Exposing synapses to
synthetic Ab1–42 oligomers decreased the fraction of Syp
spots assigned by at least one Homer spot, without
affecting the average number of Homer assignments per
Syp spot (Fig. 2B and C). However, the effect size was
small (15.6%) and the variation within and among
experiments was high.
Co-culture with CHO cellsexpressing human APP with
London mutation (V717I) induces
synaptotoxicity
We cultured CHO cell lines stably overexpressing human
APP, either wild type (CHO-APPWT) or with V717I
(London) mutation (CHO-APPLDN). It has been shown
that CHO-APPWT and CHO-APPLDN continuously secrete
physiologically relevant forms of Ab molecules and
CHO-APPLDN provides toxic Ab species (Guillot-Sestier
et al., 2012). Immunoblot analysis of media collected
from CHO cell cultures confirmed that only the peptides
secreted by CHO-APPLDN formed low-MW oligomers
(Fig. 2D). In contrast, CHO-pcDNA4 cells, which do not
overexpress APP, did not produce any Ab. We conducted
off-chip enzyme-linked immunosorbent assay measure-
ments to determine the relative levels of Ab species in
CHO cell media (Fig. 2E). As expected, treatment with
the c-secretase inhibitor N-[N-(3,5-Difluorophenacetyl)-L-
alanyl]-S-phenylglycine t-butyl ester (DAPT) completely
blocked the secretion of Ab peptides by both CHO-
APPWT and CHO-APPLDN cells. Moreover, Ab1–42 (but
not Ab1–40) levels in CHO-APPLDN media were higher
than in CHO-APPWT media, further supporting the
immunoblot results.
To determine the effect of CHO cell-secreted Ab forms
on synapses, we plated ca. 10 000 CHO cells in the co-
culture chamber 4–6 days prior to the primary neuron
culture. This timing was necessary to overcome the prob-
lems invoked by the differences of growth media compos-
ition between CHO cells and primary neurons. CHO
cells proliferated in their growth medium and fully occu-
pied the co-culture chamber. CHO cells were able to
cross the short microchannels separating the co-culture
chamber from the synaptic chamber (Fig. 1D), but they
did not migrate up the synaptic chamber. When the
growth medium was replaced with the stimulation
medium, the cells stopped proliferating but continued to
secrete Ab peptides. To confirm that CHO cell-secreted
Ab forms diffused into the synaptic chamber, we col-
lected media from different media reservoirs and quanti-
fied their Ab1–X and Ab1–42 peptide contents using
corresponding Alpha-LISA kits. Note that media collected
from the wells of the microfluidic device did not contain
sufficient material for immunoblotting. Measurements
taken at DIV14 revealed that the ratio of Ab1–42 to other
Ab forms in the synaptic chamber was 4.4-fold higher in
CHO-APPLDN co-cultures than in CHO- APPWT co-cul-
tures (Fig. 2F), as expected for the overexpression of
mutated APP (Guillot-Sestier et al., 2012). Ab forms in
the media decreased to undetectable levels when CHO
cells were treated with c-secretase inhibitor DAPT for 5
days prior to media collection, further confirming that
the majority of the measured Ab in the synaptic chamber
was secreted by the CHO cells. Presence of Ab1–X in
media collected from the synaptic chamber, but not in
the co-culture chamber of the CHO-pcDNA4 co-cultures,
is indicative of neuronal APP processing, since CHO-
pcDNA4 cells do not express APP and therefore do not
secrete Ab peptides (Supplementary Fig. 4).
We conducted off-chip experiments to assess the avail-
ability of cell-secreted Ab forms following the addition of
the conditioned media to primary neuron cultures. Alpha-
LISA measurements conducted at different time points
(from 6h to 7 days) suggested that Ab concentrations in
the media decreased logarithmically over time
(Supplementary Fig. 5), suggesting that the peptides
degraded or consumed by the cells. In contrast, the con-
centration of synthetic Ab1–42 peptide did not exhibit
such a decrease when added to the primary neuron cul-
tures (Supplementary Fig. 6). These observations further
support the idea that cell-secreted Ab forms need to be
periodically resupplied to the neuron culture medium un-
less a co-culture model is available. When synapses were
exposed to CHO cell-secreted Ab forms for 14 days in
the co-culture device, a strong decrease in synapse con-
nectivity was observed in CHO-APPLDN but not in CHO-
APPWT co-cultures (Fig. 2G and H). In this case, i.e. fol-
lowing chronic exposure, the effect size was large
(48.9%) and the variation within and among experiments
was low. The density of all Homer puncta detected and
the average number of Homer puncta assigned per Syp
puncta were also significantly lower for CHO-APPLDN
co-cultures (62.5 and 22.0% decrease compared to CHO-
APPWT, respectively). To ensure that the observed de-
crease in synapse connectivity was not a consequence of
neuronal cell death due to CHO cell-secreted Ab forms,
we conducted a classical co-culture experiment using cell
inserts. No aberrant neuronal cell death was observed at
any time point studied, despite the presence of Ab1–42 in
the media of CHO-APPLDN co-cultures (Supplementary
Fig. 7). Separately, microtubule-associated protein 2
(MAP2) protein density in images obtained from the syn-
aptic chamber did not vary with the CHO cell type used
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 11 of 18 | 11
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

in co-cultures (Supplementary Fig. 8), showing that the
effect of Ab forms on synapses was not due to an effect
on the dendritic network.
Ab antibody 3D6 arrests Ab in the
monomeric form and blocks
CHO-APPLDN-induced decrease insynapse connectivity
To further characterize the CHO cell-secreted Ab forms,
we tested two monoclonal Ab antibodies, human
SAR228810 (8810; Sanofi; 3 lg/ml) and the mouse ver-
sion of Bapineuzumab (3D6; Janssen; 3lg/ml). Both anti-
bodies were developed for passive immunotherapy and
tested in clinical trials. 8810 antibody targets soluble pro-
tofibrillar and fibrillar species of Ab and is inactive
against Ab monomers and small oligomeric aggregates
(Santin et al., 2016). In contrast, the 3D6 antibody tar-
gets the N-terminal region of Ab and expected to capture
Ab molecules in the monomeric conformation (Miles
et al., 2013; Vandenberghe et al., 2016).
We conducted off-chip experiments where CHO cells
were treated with these antibodies (or with 18 lg/ml
DAPT as a negative control) for 5 days. Western blots of
the conditioned media showed that DAPT treatment elim-
inated all Ab secretion and induced a slight increase in
the levels of APP (Fig. 3A). However, WBs of condi-
tioned media following DAPT treatment exhibited a 15-
kDa band that overlapped with oligomers of low-MW
Ab1–42. Treatment with neither antibody affected the
presence of low-MW Ab1–42 forms. Interestingly, the 3D6
antibody induced a strong increase in monomeric Ab1–42levels in both CHO-APPWT and CHO-APPLDN media, in
accordance with the idea that 3D6 arrests the peptide in
monomeric form and precludes oligomer formation. We
also performed Alpha-LISA and enzyme-linked immuno-
sorbent assay measurements from media collected from
the co-culture chamber, following the treatment of CHO
cells with the aforementioned antibodies for 5 days.
However, the 3D6 antibody interfered with the measure-
ments and data could not be reported.
We analysed synaptic connectivity in co-cultures treated
with 8810 and 3D6 antibodies for 5 days prior to fix-
ation (Fig. 3B). Similar to Fig. 2H, untreated CHO-
APPLDN co-cultures exhibited a strong (46.6%) decrease
in the fraction of Syp puncta assigned by Homer puncta
and a significant decrease (26.9%) in the number of
Homer assignments per Syp. Treatment with 8810 anti-
body did not induce a significant difference relative to
untreated controls in both cell types. However, treatment
with 3D6 antibody completely blocked the effect of
CHO-APPLDN on the fraction of Syp assigned and par-
tially blocked the effect of CHO-APPLDN on the number
of assignment per Syp. In summary, treatment with an
antibody that prevents Ab monomers from forming
oligomers interfered with CHO-APPLDN-secreted Ab
species and protected synapses from toxicity likely
induced by low-MW oligomers. These findings highlight
the potential use of our disease-on-a-chip model and syn-
aptic connectivity analysis for assessing the synaptopro-
tective effects of therapeutic compounds for Alzheimer’s
disease, such as Ab-targeting antibodies.
Pyk2 overexpression in‘postsynaptic’ neurons blocks
CHO-APPLDN-induced decrease in
synapse connectivity
We next assessed the relevance of our microfluidic tool
to establish whether genetic risk factors of Alzheimer’s
disease may be involved in Ab-dependent synaptotoxicity.
Several lines of evidence indicate that genetically driven
synaptic failure may occur in Alzheimer’s disease
(Dourlen et al., 2019) and, among the different genetic
risk factors susceptible to be studied in our model, we
focused on PTK2B, which has been already described to
be involved in synaptic functions (Giralt et al., 2017).
We evaluated potential changes in the expression of
PTK2B in the brains of Alzheimer’s disease patients com-
pared to healthy individuals by taking advantage of three
publicly available RNAseq datasets: Mayo Clinic data
that probed the temporal cortex (Allen et al., 2016);
ROSMAP data that probed the dorsolateral prefrontal
cortex (De Jager et al., 2018); and the MSBB data that
probed four different brain areas: BA 10, BA 22, BA 36
and BA 44 (Wang et al., 2018). This allowed us to inves-
tigate potential gene expression changes in brain regions
that are affected at different pathological stages of
Alzheimer’s disease (Braak and Braak, 1991). We
observed a decrease in PTK2B expression in Alzheimer’s
cases compared to healthy controls in all brain regions
analysed (Supplementary Table 2); however, after mul-
tiple testing correction, this decrease was significant only
in the BA 22 (16.69% decrease; Padj ¼ 1.99� 10�02), BA
36 (28.89% decrease; Padj ¼ 8.49� 10�05) and BA 44
(15.18% decrease; Padj ¼ 3.87� 10�02) regions of the
MSBB dataset, as well as in the ROSMAP dataset
(28.89% decrease; Padj ¼ 4.57� 10�03). Consistent with
the RNAseq data, we observed a decreasing trend in
Pyk2 total protein levels and an increasing trend in the
p-Pyk2 protein levels in the brains of Alzheimer’s patients
compared to healthy controls (Supplementary Fig. 9).
These changes resulted in a significant increase in the
relative phosphorylation of Pyk2 in the hippocampus
(4.1-fold increase in the p-Pyk2:Pyk2 ratio; P¼ 0.0274;
Wilcoxon rank-sum test; 17 cases versus 6 controls) and
in the cortex (3.6-fold increase in the p-Pyk2:Pyk2 ratio;
P¼ 0.0274; Wilcoxon rank-sum test; 17 cases versus 7
controls), suggesting a compensatory mechanism.
We then evaluated the levels of Pyk2 and phospho-
Pyk2 in primary neurons as a function of Ab exposure.
We conducted off-chip synaptosome extraction using
12 | BRAIN COMMUNICATIONS 2020: Page 12 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

primary cortical neurons treated with CHO cell media
and observed total Pyk2 and p-Pyk2 in both non-PSD
and PSD fractions. Relative to CHO-APPWT medium,
CHO-APPLDN medium caused significant decreases in the
relative amounts of total Pyk2 and p-Pyk2 in the postsy-
naptic fraction (Fig. 4A). In separate experiments
based on hippocampal neurons cultured in microfluidic
devices, we immunostained p-Pyk2 (Tyr402) alongside
synaptic markers and observed that signals localized to
both pre- and postsynaptic puncta, with an increased
tendency towards the latter (Fig. 4B). We extended
our distance-based synaptic connectivity analysis to quan-
titatively analyse the distribution of p-Pyk2 signals rela-
tive to identified synapses, i.e. Syp–Homer pairs. Our
data showed that 1.5-fold more p-Pyk2 puncta were
localized near postsynaptic puncta than near presynaptic
Figure 3 Ab antibody 3D6 modulates Ab secretion and blocks the synaptotoxicity due to CHO-APPLDN co-culture. (A) An
exemplary immunoblot of CHO cell media collected after 5-day-long treatment with the indicated compounds (DAPT, 18lg/ml; antibodies, 3 lg/
ml) showing APP cleavage products of different molecular weights. (B) Synaptic read-outs in antibody-treated co-cultures at DIV14. Each Homer
spot was assigned to the nearest Syp spot within a cut-off distance of 1.0 lm (see Supplementary Fig. 1 for details). Following the assignment of
all Homer spots, the fraction of Syp spots assigned by at least one Homer spot and the average number of Homer assignments per Syp spot were
calculated. Densities of all Syp and Homer spots detected (white), Syp spots assigned by a Homer spot (grey) and Homer spots assigned to a Syp
spot (grey) are also shown. In box plots, red circles, red bars, black squares and red plus signs indicate individual data points, sample median and
mean and outliers, respectively. Numbers of microfluidic devices analysed (obtained from at least 3 independent cultures) are given in
parentheses. Kruskal–Wallis ANOVA, followed by Wilcoxon rank-sum test. *P< 0.05; **P< 0.01. N/S ¼ not significant.
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 13 of 18 | 13
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

puncta regardless of co-culture with CHO-APPWT or
CHO-APPLDN cells (Supplementary Fig. 10).
Since we observed (i) a decrease in PTK2B expression
and a decreasing trend in Pyk2 protein levels in
Alzheimer’s brains, (ii) a decrease in Pyk2 levels in the
PSD fraction of cortical neurons upon treatment with
CHO-APPLDN media and (iii) a decrease in synaptic con-
nectivity in hippocampal neurons upon co-culture with
CHO-APPLDN cells, we hypothesized that Pyk2 was pro-
tective and its overexpression could rescue the detrimental
effect of CHO-APPLDN co-culture on synapses. Since the
active form of Pyk2 was strongly associated with postsy-
napses and CHO-APPLDN medium affected Pyk2 levels
specifically in the PSD fraction, we decided to take ad-
vantage of microfluidic compartmentalization and modu-
late Pyk2 expression in postsynaptic neurons. To this
end, we first verified the overexpression of Pyk2 off-chip
via lentiviral transduction of the relevant cDNA
(Supplementary Fig. 11). Next, using lentiviruses
expressing fluorescent proteins, we confirmed that the
viral transduction in the microfluidic device was restricted
to the target chamber (selectively to pre- or postsynaptic
chambers; Supplementary Fig. 12). Overexpressing Pyk2
in the postsynaptic chamber blocked the detrimental ef-
fect of CHO-APPLDN co-culture on synaptic connectivity,
as evidenced by 12.9% decrease in the fraction of Syp
puncta assigned by Homer puncta (as compared to
33.9% decrease when overexpressing the control vector;
Fig. 4C). As expected, synaptic connectivity in CHO-
APPWT co-cultures was not affected by Pyk2 overexpres-
sion (8.7% increase in the fraction of Syp assigned by
Homer), confirming that the synaptoprotective effect of
postsynaptic Pyk2 overexpression was specific to Ab tox-
icity due to CHO-APPLDN co-culture. Similar to data
shown in Fig. 2H, in cultures expressing the control vec-
tor, CHO-APPLDN co-culture decreased the density of all
Homer puncta detected and the average number of
Homer puncta assigned per Syp puncta (by 46.7% and
Figure 4 Pyk2 overexpression in postsynaptic neurons blocks the synaptotoxicity due to CHO-APPLDN co-culture.
(A) Exemplary immunoblots and quantification of the PSD fraction following synaptosome extraction at DIV14 following 16-h-long treatment
with the indicated CHO cell medium. N¼ 3 independent experiments. Unpaired t-test. (B) An exemplary immunofluorescence image of the
synaptic chamber at DIV14 showing p-Pyk2, Syp, Homer 1 and MAP2. Boxed areas are 2.5� magnified. (C) Synaptic read-outs following Pyk2
overexpression in postsynaptic neurons from DIV7 to DIV14. Each Homer spot was assigned to the nearest Syp spot within a cut-off distance of
1.0lm (see Supplementary Fig. 1 for details). Following the assignment of all Homer spots, the fraction of Syp spots assigned by at least one
Homer spot and the average number of Homer assignments per Syp spot were calculated. Densities of all Syp and Homer spots detected
(white), Syp spots assigned by a Homer spot (grey) and Homer spots assigned to a Syp spot (grey) are also shown. In box plots, red circles, red
bars and black squares indicate individual data points, sample median and sample mean, respectively. Numbers of microfluidic devices analysed
(obtained from at least three independent cultures) are given in parentheses. Kruskal–Wallis ANOVA, followed by Wilcoxon rank-sum test.
Error bars ¼ SEM. *P< 0.05; **P< 0.01; ***P< 0.005. N/S ¼ not significant.
14 | BRAIN COMMUNICATIONS 2020: Page 14 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

17.7%, respectively) relative to CHO-APPWT co-culture.
The former, but not the latter, was blocked when
Pyk2 was overexpressed in the postsynaptic chamber
(Fig. 4C).
DiscussionThe use of microfluidic culture devices for isolating syn-
apses from neuronal cell bodies has been previously dem-
onstrated (Taylor et al., 2010; Virlogeux et al., 2018).
Our co-culture device combines two design concepts:
first, by employing different microchannel lengths, it
guarantees that dendrites from one but not the other neu-
ron chamber can arrive to the synapse chamber. Second,
it minimizes the penetration of axons from the synapse
chamber to the neuron chambers and thereby facilitates
the passage of axons in the intended direction: 2.9- and
1.9-fold more axons crossed the microchannels in the
intended direction relative to the reverse direction, re-
spectively, for long and short microchannels. These ratios
are indicative of a limited effect of the narrowing channel
design on hippocampal neurons and are consistent with
earlier reports (Peyrin et al., 2011). Our co-culture device
also facilitated the concentration of synapses in the syn-
apse chamber: >83% of all synapses formed between
neurons cultured in the pre- and postsynaptic chambers
were found in the synapse chamber. It is important to
note that while all dendrites in the synapse chamber ar-
rive from the postsynaptic chamber, the opposite state-
ment is not true, i.e. not all axons in the synapse
chamber arrive from the presynaptic chamber. Thus, for
all synapses analysed, overexpressing Pyk2 in the postsy-
naptic chamber guarantees Pyk2 overexpression in the
postsynaptic neuron (provided that it is infected by the
lentivirus); however, the possibility of Pyk2 overexpres-
sion also in the presynaptic neuron cannot be ruled out.
However, this uncertainty is not relevant for studies
focusing solely on postsynaptic mechanisms. The micro-
fluidic approach provides a simple yet robust method to
image and analyse synapses independently of densely
plated cell bodies. Although not exploited in the context
of this study, our co-culture device would also permit
acute treatment of synapses independently of their cell
bodies and independently of the co-cultured cells, thanks
to the additional access well connected to the synaptic
chamber (Fig. 1B). However, long-term treatments, as in
the case for Ab molecules secreted by the CHO cells,
cannot be exclusively directed to synapses due to molecu-
lar diffusion.
Several parameters can be considered when inducing
Ab-dependent synaptotoxicity: (i) use of synthetic versus
organic oligomers and (ii) acute versus chronic treatment.
Our microfludic co-culture model combines organic
oligomers at physiological concentrations with chronic
treatments and consistently induces Ab-dependent synap-
totoxicity. This is in agreement with a recent report that
synthetic oligomers do not assume the same molecular
structure as organic oligomers (Kollmer et al., 2019), sug-
gesting potential differences in their biological effects.
Interestingly, as opposed to synthetic Ab oligomers, or-
ganic Ab appears to be degraded in our primary neuron-
al cultures, further highlighting the difference in
bioavailability between the two. This observation also
indicates that the co-culture model is better adapted for
synaptotoxicity studies since it maintains physiological
concentrations thanks to the continuous secretion of Ab
peptides from CHO cells. This allows for the analysis of
synapse connectivity in response to chronic exposure of
synapses to organic Ab oligomers, as opposed to acute
exposure to conditioned media, for instance.
It is important to note that the Ab peptides secreted by
the CHO-APPLDN cells do diffuse into the various cham-
bers of the microfluidic device, considering the long dur-
ation of co-culture experiments. Thus, the synaptic
toxicity observed could be due to a local effect on synap-
ses or due to an effect on neuronal cell bodies. Through
conventional co-culture experiments conducted off-chip
we showed that CHO cell-secreted Ab forms do not in-
duce neuronal death; however, this does not allow us to
rule out any potential mechanisms originating from neur-
onal cell bodies and resulting in the decrease in synaptic
connectivity observed. If true, such a mechanism would
be more likely to occur in neurons in the postsynaptic
chamber, as the fluidic barrier between the synaptic and
postsynaptic chambers is much weaker than that between
the synaptic and presynaptic chambers, allowing the cell-
secreted Ab to easily access the postsynaptic chamber.
The role of Pyk2 in synapses appears to be complex,
considering the seemingly opposite results of recent stud-
ies: Pyk2 has been shown to be required for long-term
potentiation (Huang et al., 2001). However, others have
shown that Pyk2 is not required for long-term potenti-
ation, but for long-term depression (Hsin et al., 2010;
Salazar et al., 2019), and that Pyk2 overexpression inhib-
its long-term potentiation (Hsin et al., 2010) and induces
dendritic spine loss (Lee et al., 2019). Recent analysis of
protein synthesis and degradation during synaptic scaling
showed that Pyk2 protein level was significantly increased
in response to drug-induced decrease in network activity
(via increased synthesis and decreased degradation)
(Dorrbaum et al., 2020). The role of Pyk2 in Ab toxicity
has also been debated, where it has been shown to be
deleterious (Salazar et al., 2019) or protective (Giralt
et al., 2018) in vivo. Our findings are in agreement with
the notion that Pyk2 localizes to the postsynaptic com-
partment (Giralt et al., 2017; Lee et al., 2019; Salazar
et al., 2019). To shed further light into the role of Pyk2
in Ab-induced synapse toxicity, we overexpressed Pyk2
specifically in postsynaptic neurons since we observed
that the active form of Pyk2 was strongly associated with
postsynapses and that Ab treatment affected Pyk2 levels
specifically in the PSD fraction. At the pathophysiological
level, several of our observations fit well with the results
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 15 of 18 | 15
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

of recent in vivo studies using Alzheimer’s disease-like
mouse models: (i) decreased Pyk2 activity has been
reported in 5xFAD mice (Giralt et al., 2018) and (ii) res-
cue of Pyk2 expression improved the behavioural and
synaptic molecular phenotypes of the double transgenic
5xFAD�Pyk�/� mouse model (Giralt et al., 2018).
Interestingly, this rescue seems to have no impact on Ab
loads, suggesting that Pyk2 overexpression impacts patho-
physiological processes downstream of Ab production
and amyloid deposition. This observation is in agreement
with our results, suggesting that Pyk2 overexpression in
postsynaptic neurons may restrict Ab-induced synaptotox-
icity. In stark contrast to these reports, increased Pyk2
activation has been shown in response to acute Ab oligo-
mer treatment in brain slices (Haas et al., 2016; Haas
and Strittmatter, 2016) and in the APP/PS1 mouse model
(Kaufman et al., 2015). Moreover, Pyk2 has been shown
to be detrimental in the double transgenic APP/
PS1�Pyk�/� mouse model, where lack of Pyk2 protected
from synapse loss and memory impairment (Salazar
et al., 2019). These contradictory reports may be due to
differences in the disease model and in the Pyk2�/�
model used (Giralt et al., 2017; Salazar et al., 2019). In
summary, the sparse literature on Pyk2’s role in synapses
is highly controversial and calls for further in vitro and
in vivo work.
In conclusion, our microfluidic co-culture device pro-
vides an in vitro model of Ab synaptotoxicity based on
exposing synapses of primary hippocampal neurons to
cell-secreted Ab1–42 peptides. This disease-on-a-chip model
is highly relevant to Alzheimer’s disease in several
aspects: (i) long-term, low-dose exposure to organic Ab
forms is preferable over acute treatments with synthetic
oligomers at high concentrations; (ii) isolating synapses in
a separate microfluidic chamber facilitates the analysis of
synaptic connectivity via immunostaining pre- and postsy-
naptic markers without the interference of cell bodies;
(iii) providing exclusive access to neurons cultured in the
pre- and postsynaptic chambers to selectively under- or
overexpress Alzheimer’s disease genetic risk factors, there-
in may potentially help dissect their pre- and postsynaptic
roles. Deciphering the mechanisms by which the genetic
risk factors contribute to Alzheimer’s pathology may lead
to novel therapeutic approaches.
Supplementary materialSupplementary material is available at Brain
Communications online.
AcknowledgementsThe authors thank the BICeL platform of the Institut
Biologie de Lille. The authors thank Laurent Pradier and
Philippe Bertrand at Sanofi for fruitful discussions. The
authors thank Karine Blary at the IEMN Lille for the micro-
fabrication work. The authors thank the vectorology plat-
form Transbiomed for lentivirus production. The authors
thank Charles Duyckaerts and the ‘NeuroCEB’ Brain Bank
(GIE Neuro-CEB BB-0033-00011) for providing the brain
tissue samples. The results published here are in whole or in
part based on data obtained from the AMP-AD Knowledge
Portal (https://adknowledgeportal.synapse.org/). These data
were generated from post-mortem brain tissue collected
through the Mount Sinai VA Medical Center Brain Bank
and were provided by Dr. Eric Schadt from Mount Sinai
School of Medicine. Study data were also provided by the
following sources: The Mayo Clinic Alzheimers Disease
Genetic Studies, led by Dr. Nilufer Taner and Dr. Steven G.
Younkin, Mayo Clinic, Jacksonville, FL, using samples from
the Mayo Clinic Study of Aging, the Mayo Clinic Alzheimers
Disease Research Center, and the Mayo Clinic Brain Bank.
Data collection was supported through funding by NIA
(grants P50 AG016574, R01 AG032990, U01 AG046139,
R01 AG018023, U01 AG006576, U01 AG006786, R01
AG025711, R01 AG017216 and R01 AG003949), NINDS
(grant R01 NS080820), CurePSP Foundation and Mayo
Foundation. Study data include samples collected through
the Sun Health Research Institute’s Brain and Body
Donation Program of Sun City, AZ. The Brain and Body
Donation Program is supported by the National Institute of
Neurological Disorders and Stroke (U24 NS072026,
National Brain and Tissue Resource for Parkinson’s Disease
and Related Disorders), the National Institute on Aging (P30
AG19610, Arizona Alzheimer’s Disease Core Center), the
Arizona Department of Health Services (contract 211002,
Arizona Alzheimer’s Research Center), the Arizona
Biomedical Research Commission (contracts 4001, 0011,
05-901 and 1001, Arizona Parkinson’s Disease Consortium)
and the Michael J. Fox Foundation for Parkinson’s
Research. Study data were also provided by the Rush
Alzheimer’s Disease Center, Rush University Medical Center,
Chicago. Data collection was supported through funding by
NIA [grants P30AG10161 (ROS), R01AG15819 (ROSMAP;
genomics and RNAseq), R01AG17917 (MAP), R01AG
30146, R01AG36042 (5hC methylation, ATACseq),
RC2AG036547 (H3K9Ac), R01AG36836 (RNAseq),
R01AG48015 (monocyte RNAseq), RF1AG57473 (single
nucleus RNAseq), U01AG32984 (genomic and whole exome
sequencing), U01AG46152 (ROSMAP AMP-AD, targeted
proteomics), U01AG46161 (TMT proteomics) and
U01AG61356 (whole genome sequencing, targeted proteo-
mics, ROSMAP AMP-AD)], the Illinois Department of
Public Health (ROSMAP) and the Translational Genomics
Research Institute (genomic). Additional phenotypic data can
be requested at https://www.radc.rush.edu/.
FundingThis study was partly supported by the French RENATECH
network (P-16-01891). This study was funded by INSERM,
16 | BRAIN COMMUNICATIONS 2020: Page 16 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

Institut Pasteur de Lille, the EU Joint Programme—
Neurodegenerative Diseases Research (JPND; 3DMiniBrain),
Agence Nationale de la Recherche (ANR-19-CE16-0020)
and Fondation Vaincre Alzheimer (FR-17006p). This study
was also funded by the Lille Metropole Communaute
Urbaine and the French government’s LABEX DISTALZ
program (Development of innovative strategies for a trans-
disciplinary approach to Alzheimer’s disease). D.M.-C. was
supported by a PhD scholarship from Coordenac~ao de
Aperfeicoamento de Pessoal de Nıvel Superior (CAPES). This
study was also co-funded by the European Union under the
European Regional Development Fund (ERDF) and by the
Hauts de France Regional Council (contract no. 18006176),
the Metropole Europeenne de Lille (contract no.
2016_ESR_05) and the French State (contract no. 2018-3-
CTRL_IPL_Phase2). The aforementioned funding bodies did
not play any roles in the design of the study and collection,
analysis and interpretation of data and in writing the article.
Competing interestsThe authors report no competing interests.
ReferencesAllen M, Carrasquillo MM, Funk C, Heavner BD, Zou F, Younkin
CS, et al. Human whole genome genotype and transcriptome data
for Alzheimer’s and other neurodegenerative diseases. Sci Data
2016; 3: 160089.
Bartos JA, Ulrich JD, Li H, Beazely MA, Chen Y, Macdonald JF, et al.
Postsynaptic clustering and activation of Pyk2 by PSD-95.
J Neurosci 2010; 30: 449–63.
Benilova I, Karran E, De Strooper B. The toxic Abeta oligomer and
Alzheimer’s disease: an emperor in need of clothes. Nat Neurosci
2012; 15: 349–57.
Blasiak A, Lee GU, Kilinc D. Neuron subpopulations with different
elongation rates and DCC dynamics exhibit distinct responses to iso-
lated netrin-1 treatment. ACS Chem Neurosci 2015; 6: 1578–90.
Braak H, Alafuzoff I, Arzberger T, Kretzschmar H, Del Tredici K.
Staging of Alzheimer disease-associated neurofibrillary pathology
using paraffin sections and immunocytochemistry. Acta
Neuropathol 2006; 112: 389–404.
Braak H, Braak E. Demonstration of amyloid deposits and neurofibril-
lary changes in whole brain sections. Brain Pathol 1991; 1: 213–6.
Bray NL, Pimentel H, Melsted P, Pachter L. Near-optimal probabilistic
RNA-seq quantification. Nat Biotechnol 2016; 34: 525–7.
Brody AH, Strittmatter SM. Synaptotoxic sgnaling by amyloid beta
oligomers in Alzheimer’s disease through prion protein and
mGluR5. Adv Pharmacol 2018; 82: 293–323.
Chang KH, Vincent F, Shah K. Deregulated Cdk5 triggers aberrant ac-
tivation of cell cycle kinases and phosphatases inducing neuronal
death. J Cell Sci 2012; 125: 5124–37.
Chapuis J, Flaig A, Grenier-Boley B, Eysert F, Pottiez V, Deloison G,
et al.; ADGC, Alzheimer’s Disease Neuroimaging Initiative.
Genome-wide, high-content siRNA screening identifies the
Alzheimer’s genetic risk factor FERMT2 as a major modulator of
APP metabolism. Acta Neuropathol 2017; 133: 955–66.
Dahlgren KN, Manelli AM, Stine WB, Jr., Baker LK, Krafft GA, LaDu
MJ. Oligomeric and fibrillar species of amyloid-beta peptides differ-
entially affect neuronal viability. J Biol Chem 2002; 277: 32046–53.
De Jager PL, Ma Y, McCabe C, Xu J, Vardarajan BN, Felsky D, et al.
A multi-omic atlas of the human frontal cortex for aging and
Alzheimer’s disease research. Sci Data 2018; 5: 180142.
De Strooper B. Proteases and proteolysis in Alzheimer disease: a multi-
factorial view on the disease process. Physiol Rev 2010; 90: 465–94.
Deshpande A, Mina E, Glabe C, Busciglio J. Different conformations
of amyloid beta induce neurotoxicity by distinct mechanisms in
human cortical neurons. J Neurosci 2006; 26: 6011–8.
Dorrbaum AR, Alvarez-Castelao B, Nassim-Assir B, Langer JD,
Schuman EM. Proteome dynamics during homeostatic scaling in cul-
tured neurons. eLife 2020; 9: e52939.
Dourlen P, Kilinc D, Malmanche N, Chapuis J, Lambert JC. The new
genetic landscape of Alzheimer’s disease: from amyloid cascade to
genetically driven synaptic failure hypothesis? Acta Neuropathol
2019; 138: 221–36.
Eysert F. Study of mechanisms involving the genetic risk factor
FERMT2 in the APP metabolism and its consequences in the physio-
pathological process of Alzheimer’s disease [dissertation]. Lille,
France: University of Lille; 2019.
Ferreira IL, Bajouco LM, Mota SI, Auberson YP, Oliveira CR, Rego
AC. Amyloid beta peptide 1-42 disturbs intracellular calcium
homeostasis through activation of GluN2B-containing N-methyl-D-
aspartate receptors in cortical cultures. Cell Calcium 2012; 51:
95–106.
Frandemiche ML, De Seranno S, Rush T, Borel E, Elie A, Arnal I,
et al. Activity-dependent tau protein translocation to excitatory syn-
apse is disrupted by exposure to amyloid-beta oligomers. J Neurosci
2014; 34: 6084–97.
Giralt A, Brito V, Chevy Q, Simonnet C, Otsu Y, Cifuentes DC, et al.
Pyk2 modulates hippocampal excitatory synapses and contributes to
cognitive deficits in a Huntington’s disease model. Nat Commun
2017; 8: 15592.
Giralt A, de Pins B, Cifuentes-Diaz C, Lopez-Molina L, Farah AT,
Tible M, et al. PTK2B/Pyk2 overexpression improves a mouse model
of Alzheimer’s disease. Exp Neurol 2018; 307: 62–73.
Guillot-Sestier MV, Sunyach C, Ferreira ST, Marzolo MP, Bauer C,
Thevenet A, et al. alpha-Secretase-derived fragment of cellular prion,
N1, protects against monomeric and oligomeric amyloid beta
(Abeta)-associated cell death. J Biol Chem 2012; 287: 5021–32.
Haas LT, Salazar SV, Kostylev MA, Um JW, Kaufman AC,
Strittmatter SM. Metabotropic glutamate receptor 5 couples cellular
prion protein to intracellular signalling in Alzheimer’s disease. Brain
2016; 139: 526–46.
Haas LT, Strittmatter SM. Oligomers of amyloid beta prevent physio-
logical activation of the cellular prion protein-metabotropic glutam-
ate receptor 5 complex by glutamate in Alzheimer disease. J Biol
Chem 2016; 291: 17112–21.
Hardy J, Selkoe DJ. The amyloid hypothesis of Alzheimer’s disease:
progress and problems on the road to therapeutics. Science 2002;
297: 353–6.
Hong S, Beja-Glasser VF, Nfonoyim BM, Frouin A, Li S,
Ramakrishnan S, et al. Complement and microglia mediate early
synapse loss in Alzheimer mouse models. Science 2016; 352: 712–6.
Hsin H, Kim MJ, Wang CF, Sheng M. Proline-rich tyrosine kinase 2
regulates hippocampal long-term depression. J Neurosci 2010; 30:
11983–93.
Huang Y, Lu W, Ali DW, Pelkey KA, Pitcher GM, Lu YM, et al.
CAKbeta/Pyk2 kinase is a signaling link for induction of long-term
potentiation in CA1 hippocampus. Neuron 2001; 29: 485–96.
Kaufman AC, Salazar SV, Haas LT, Yang J, Kostylev MA, Jeng AT,
et al. Fyn inhibition rescues established memory and synapse loss in
Alzheimer mice. Ann Neurol 2015; 77: 953–71.
Kelly BL, Ferreira A. Beta-amyloid disrupted synaptic vesicle endocyto-
sis in cultured hippocampal neurons. Neuroscience 2007; 147:
60–70.
Kilinc D, Blasiak A, O’Mahony JJ, Lee GU. Low piconewton towing
of CNS axons against diffusing and surface-bound repellents
Pyk2 blocks Ab1–42
-induced synaptotoxicity BRAIN COMMUNICATIONS 2020: Page 17 of 18 | 17
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

requires the inhibition of motor protein-associated pathways. Sci
Rep 2015; 4: 7128.
Kilinc D, Schwab J, Rampini S, Ikpekha OW, Thampi A, Blasiak A,
et al. A microfluidic dual gradient generator for conducting cell-
based drug combination assays. Integr Biol 2016; 8: 39–49.
Kollmer M, Close W, Funk L, Rasmussen J, Bsoul A, Schierhorn A,
et al. Cryo-EM structure and polymorphism of Abeta amyloid fibrils
purified from Alzheimer’s brain tissue. Nat Commun 2019; 10:
4760.
Kunkle BW, Grenier-Boley B, Sims R, Bis JC, Damotte V, Naj AC,
et al.; Alzheimer Disease Genetics Consortium (ADGC). Genetic
meta-analysis of diagnosed Alzheimer’s disease identifies new risk
loci and implicates Abeta, tau, immunity and lipid processing. Nat
Genet 2019; 51: 414–30.
Kuperstein I, Broersen K, Benilova I, Rozenski J, Jonckheere W,
Debulpaep M, et al. Neurotoxicity of Alzheimer’s disease Abeta pep-
tides is induced by small changes in the Abeta42 to Abeta40 ratio.
EMBO J 2010; 29: 3408–20.
Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R,
Bellenguez C, et al.; European Alzheimer’s Disease Initiative (EADI).
Meta-analysis of 74,046 individuals identifies 11 new susceptibility
loci for Alzheimer’s disease. Nat Genet 2013; 45: 1452–8.
Lansbury PT. Jr., Evolution of amyloid: what normal protein folding
may tell us about fibrillogenesis and disease. Proc Natl Acad Sci
USA 1999; 96: 3342–4.
Lee S, Salazar SV, Cox TO, Strittmatter SM. Pyk2 signaling through
Graf1 and RhoA GTPase is required for amyloid-b oligomer-trig-
gered synapse loss. J Neurosci 2019; 39: 1910–29.
Love MI, Huber W, Anders S. Moderated estimation of fold change
and dispersion for RNA-seq data with DESeq2. Genome Biol 2014;
15: 550.
Mairet-Coello G, Courchet J, Pieraut S, Courchet V, Maximov A,
Polleux F. The CAMKK2-AMPK kinase pathway mediates the syn-
aptotoxic effects of Abeta oligomers through Tau phosphorylation.
Neuron 2013; 78: 94–108.
Miles LA, Crespi GA, Doughty L, Parker MW. Bapineuzumab cap-
tures the N-terminus of the Alzheimer’s disease amyloid-beta peptide
in a helical conformation. Sci Rep 2013; 3: 1302.
Nisbet RM, Polanco JC, Ittner LM, Gotz J. Tau aggregation and its
interplay with amyloid-beta. Acta Neuropathol 2015; 129: 207–20.
Ojelade SA, Lee TV, Giagtzoglou N, Yu L, Ugur B, Li Y, et al. cindr,
the drosophila homolog of the CD2AP Alzheimer’s disease risk
gene, is required for synaptic transmission and proteostasis. Cell
Rep 2019; 28: 1799–813.e1795.
Peyrin JM, Deleglise B, Saias L, Vignes M, Gougis P, Magnifico S,
et al. Axon diodes for the reconstruction of oriented neuronal net-
works in microfluidic chambers. Lab Chip 2011; 11: 3663–73.
Resende R, Ferreiro E, Pereira C, Resende de Oliveira C. Neurotoxic
effect of oligomeric and fibrillar species of amyloid-beta peptide 1–
42: involvement of endoplasmic reticulum calcium release in oligo-
mer-induced cell death. Neuroscience 2008; 155: 725–37.
Salazar SV, Cox TO, Lee S, Brody AH, Chyung AS, Haas LT, et al.
Alzheimer’s disease risk factor Pyk2 mediates amyloid-beta-induced
synaptic dysfunction and loss. J Neurosci 2019; 39: 758–72.
Santin MD, Vandenberghe ME, Herard AS, Pradier L, Cohen C,
Debeir T, et al. In vivo detection of amyloid plaques by gadolinium-
stained MRI can be used to demonstrate the efficacy of an anti-
amyloid immunotherapy. Front Aging Neurosci 2016; 8: 55.
Sartori M, Mendes T, Desai S, Lasorsa A, Herledan A, Malmanche N,
et al. BIN1 recovers tauopathy-induced long-term memory deficits in
mice and interacts with Tau through Thr(348) phosphorylation.
Acta Neuropathol 2019; 138: 631–52.
Scheff SW, Price DA, Schmitt FA, DeKosky ST, Mufson EJ. Synaptic
alterations in CA1 in mild Alzheimer disease and mild cognitive im-
pairment. Neurology 2007; 68: 1501–8.
Schurmann B, Bermingham DP, Kopeikina KJ, Myczek K, Yoon S,
Horan KE, et al. A novel role for the late-onset Alzheimer’s disease
(LOAD)-associated protein Bin1 in regulating postsynaptic traffick-
ing and glutamatergic signaling. Mol Psychiatry 2019. doi:
10.1038/s41380-019-0407-3.
Sepulveda FJ, Parodi J, Peoples RW, Opazo C, Aguayo LG.
Synaptotoxicity of Alzheimer beta amyloid can be explained by its
membrane perforating property. PLoS One 2010; 5: e11820.
Spires-Jones TL, Hyman BT. The intersection of amyloid beta and tau
at synapses in Alzheimer’s disease. Neuron 2014; 82: 756–71.
Stine WB, Jr., Dahlgren KN, Krafft GA, LaDu MJ. In vitro character-
ization of conditions for amyloid-beta peptide oligomerization and
fibrillogenesis. J Biol Chem 2003; 278: 11612–22.
Taylor AM, Dieterich DC, Ito HT, Kim SA, Schuman EM.
Microfluidic local perfusion chambers for the visualization and ma-
nipulation of synapses. Neuron 2010; 66: 57–68.
Thierry M, Boluda S, Delatour B, Marty S, Seilhean D, Potier M-C,
et al.; Brainbank Neuro-CEB Neuropathology Network. Human
subiculo-fornico-mamillary system in Alzheimer’s disease: Tau seed-
ing by the pillar of the fornix. Acta Neuropathol 2020; 139:
443–61.
Um JW, Nygaard HB, Heiss JK, Kostylev MA, Stagi M, Vortmeyer A,
et al. Alzheimer amyloid-beta oligomer bound to postsynaptic prion
protein activates Fyn to impair neurons. Nat Neurosci 2012; 15:
1227–35.
Vandenberghe R, Rinne JO, Boada M, Katayama S, Scheltens P,
Vellas B, et al.; Bapineuzumab 3000 and 3001 Clinical Study
Investigators. Bapineuzumab for mild to moderate Alzheimer’s dis-
ease in two global, randomized, phase 3 trials. Alzheimers Res Ther
2016; 8: 18.
Virlogeux A, Moutaux E, Christaller W, Genoux A, Bruyere J, Fino E,
et al. Reconstituting corticostriatal network on-a-chip reveals the
contribution of the presynaptic compartment to Huntington’s dis-
ease. Cell Rep 2018; 22: 110–22.
Walsh DT, Montero RM, Bresciani LG, Jen AY, Leclercq PD,
Saunders D, et al. Amyloid-beta peptide is toxic to neurons in vivo
via indirect mechanisms. Neurobiol Dis 2002; 10: 20–7.
Wang M, Beckmann ND, Roussos P, Wang E, Zhou X, Wang Q,
et al. The Mount Sinai cohort of large-scale genomic, transcriptomic
and proteomic data in Alzheimer’s disease. Sci Data 2018; 5:
180185.
Wang Q, Walsh DM, Rowan MJ, Selkoe DJ, Anwyl R. Block of long-
term potentiation by naturally secreted and synthetic amyloid beta-
peptide in hippocampal slices is mediated via activation of the kin-
ases c-Jun N-terminal kinase, cyclin-dependent kinase 5, and p38
mitogen-activated protein kinase as well as metabotropic glutamate
receptor type 5. J Neurosci 2004; 24: 3370–8.
Xiong K, Cai H, Luo XG, Struble RG, Clough RW, Yan XX.
Mitochondrial respiratory inhibition and oxidative stress elevate
beta-secretase (BACE1) proteins and activity in vivo in the rat retina.
Exp Brain Res 2007; 181: 435–46.
18 | BRAIN COMMUNICATIONS 2020: Page 18 of 18 D. Kilinc et al.
Do
wn
loa
de
d fro
m h
ttps://a
ca
de
mic
.ou
p.c
om
/bra
inco
mm
s/a
rticle
/2/2
/fca
a1
39
/58
98
62
5 b
y g
ue
st o
n 1
7 M
ay 2
02
1

1
Pyk2 Overexpression in Postsynaptic Neurons Blocks Amyloid �1-42-
induced Synaptotoxicity in Microfluidic Co-Cultures
Devrim Kilinca ��$QD�V-Camille Vreulx
a, Tiago Mendes
a, Amandine Flaig
a, Diego Marques-
Coelhob, c
, Maxime Verschoorea, Florie Demiautte
a, Philippe Amouyel
a, NeuroCEB Brain
Bank, Fanny Eyserta, Pierre Dourlen
a, Julien Chapuis
a, Marcos R. Costa
a, b, Nicolas
Malmanchea, Frédéric Checler
d and Jean-Charles Lambert
a*
a Université de Lille, Institut Pasteur de Lille. CHU Lille, INSERM U1167, LabEx DISTALZ, 1 rue du Prof
Calmette, BP245, Lille 59019 France
b Brain Institute, Federal University of Rio Grande do Norte, Av. Nascimento de Castro 2155 Natal, Brazil
c Bioinformatics Multidisciplinary Environment (BioME), Federal University of Rio Grande do Norte, Av.
Nascimento de Castro 2155 Natal, Brazil
d CNRS UMR7275 /DERUDWRU\�RI�([FHOOHQFH�³'LVWDO]´��,30&��8QLYHUVLWp�&{WH�G$]XU��,QVHUP, 660 Route des
Lucioles, Sophia-Antipolis, Valbonne 06560 France
* Corresponding authors: Devrim Kilinc [email protected]; Jean-Charles Lambert jean-
Supplementary Material

2
Table S1. 'HPRJUDSKLF�GHWDLOV�RI�WKH�QHXURSDWKRORJLFDO�FRKRUW��1'��1RW�GHWHUPLQHG��$'��$O]KHLPHU¶V�GLVHDVH�
Individual Braak stage Gender Age at death Post-mortem
delay (h)
Neuropathological
diagnosis
1 III F 95 ND Non-AD
2 Å F 52 29 Non-AD
3 I F 92 21 Non-AD
4 II M 82 63 Non-AD
5 II F 83 21 Non-AD
6 III F 93 24 Non-AD
7 Å M 69 6 Non-AD
8 IV F 76 28 Non-AD
9 VI F 48 ND AD
10 VI M 57 19 AD
11 VI F 60 ND AD
12 VI M 72 44 AD
13 VI F 72 5 AD
14 VI F 78 24 AD
15 VI M 81 17 AD
16 VI F 85 31 AD
17 VI F 100 67 AD
18 VI M 67 30 AD
19 VI M 67 ND AD
20 VI M 53 ND AD
21 VI F 56 26 AD
22 VI F 70 ND AD
23 VI F 72 27 AD
24 VI M 76 27 AD
25 VI F 77 48 AD
26 VI F 81 41 AD
27 VI F 85 28 AD
28 VI F 90 24 AD
Table S2. PTK2B gene expression analysis in different publicly-available datasets. BA: Brodmann area.
Dataset Brain region Sample size Gene expression
(ENSG00000120899.17)
Cases Controls
log2 fold
change padj
Mayo Clinic Temporal cortex 82 78 í0.2036 0.1308
Mount Sinai
Brain Bank
BA 10 105 71 í0.0932 0.2802
BA 22 98 61 í0.2635 0.0199
BA 36 88 64 í0.4919 8.49E-05
BA 44 90 63 í0.2375 0.0387
ROSMAP Dorsolateral
prefrontal cortex 222 201 í0.1577 0.0046

3
Figure S1. Synapse analysis workflow and the determination of the distance cut-off. A. An exemplary dendrite in the synaptic
chamber. Image shows the maximum intensity projection of Homer 1 (yellow), Synaptophysin 1 (cyan), and MAP2 (magenta)
stains. B. Segmentation of the same image in Imaris. C-D. Marked areas in A and B, respectively, magnified 3.5×. E. Principle
of assigning postsynaptic (Homer) to presynaptic (Syp) puncta based on proximity. Numbers indicate the number of
assignments for each presynaptic puncta. F. Synaptic assignment density as a function of distance cut-off. G. Fractions of
assigned pre- and postsynaptic puncta as a function of distance cut-off, showing that Syp assignments saturate at around 1.0
µm for the dataset analyzed. Data in F and G are based on N = 9 microfluidic devices (small circles) from 4 independent
cultures; 5-8 images per device. Rectangles and error bars show show sample mean and SEM, respectively.

4
Figure S2. Quantification of synaptic markers in the synaptosomal fraction. Synaptophysin (Syp) in the presynaptic fraction
and PSD95 in the postsynaptic fraction for primary neuronal cultures exposed to media collected from CHO-APPWT
and CHO-
APPLDN
cultures. Each data point represents one independent experiment. Unpaired t-test. N/S: not significant.

5
Figure S3. Exemplary images of microfluidic devices where primary neurons were plated only in the presynaptic (A), synaptic
(B), or the postsynaptic (C) FKDPEHU����-tubulin (magenta), Hoechst (cyan), and GFAP (yellow) staining shown at DIV14.
:KLWH�UHFWDQJOHV�LQGLFDWH�UHJLRQV�ZKHUH���-tubulin staining intensity was measured to calculate the penetration ratios.

6
Figure S4. &RQFHQWUDWLRQ�RI�$�1-X DQG�$�1-42 in the media collected from the reservoir of the co-culture chamber at DIV14
measured via Alpha-LISA. �-VHFUHWDVH� LQKLELWRU� '$37�ZDV� DSSOLHG� DW� ����J�P/� IRU� �� G�� 2QH-way ANOVA, followed by
unpaired t-test. * p < 0.05; ** p < 0.005.
Figure S5. &RQFHQWUDWLRQ�RI�$�1-X DQG�$�1-42 in the media collected from primary neurons cultured in 384-well plates and
exposed to CHO cell-conditioned media for 6 h to 7 d. Concentration was determined via Alpha-LISA using 6-8 wells per
condition. Error bars indicate standard deviation of the mean. Individual data points are not shown for time points where all
values were equal to zero.
Figure S6. &RQFHQWUDWLRQ�RI�$�1-42 in the media collected from primary neurons cultured in 384-well plates and treated with
V\QWKHWLF�$�1-42 oligomers for 6 h to 7 d. Concentration was determined via Alpha-LISA using 6-8 wells per condition. Error
bars indicate standard deviation of the mean. Individual data points are not shown for time points where all values were equal
to zero.
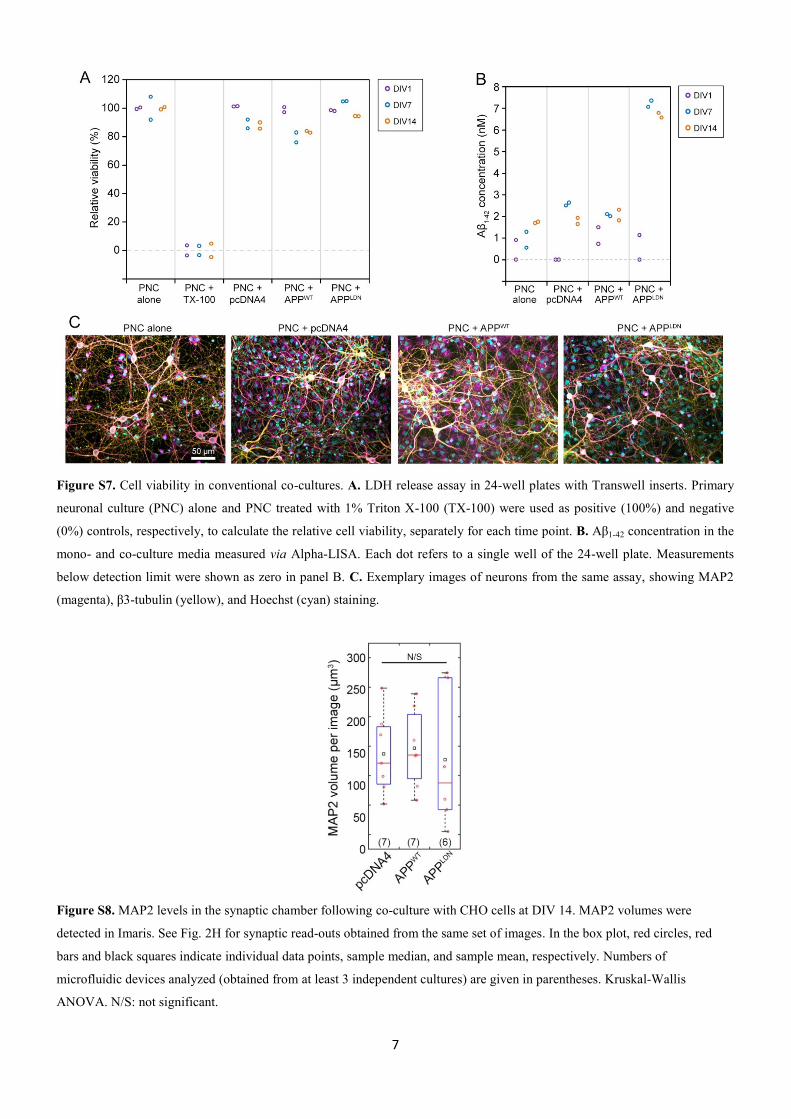
7
Figure S7. Cell viability in conventional co-cultures. A. LDH release assay in 24-well plates with Transwell inserts. Primary
neuronal culture (PNC) alone and PNC treated with 1% Triton X-100 (TX-100) were used as positive (100%) and negative
(0%) controls, respectively, to calculate the relative cell viability, separately for each time point. B. $�1-42 concentration in the
mono- and co-culture media measured via Alpha-LISA. Each dot refers to a single well of the 24-well plate. Measurements
below detection limit were shown as zero in panel B. C. Exemplary images of neurons from the same assay, showing MAP2
�PDJHQWD�����-tubulin (yellow), and Hoechst (cyan) staining.
Figure S8. MAP2 levels in the synaptic chamber following co-culture with CHO cells at DIV 14. MAP2 volumes were
detected in Imaris. See Fig. 2H for synaptic read-outs obtained from the same set of images. In the box plot, red circles, red
bars and black squares indicate individual data points, sample median, and sample mean, respectively. Numbers of
microfluidic devices analyzed (obtained from at least 3 independent cultures) are given in parentheses. Kruskal-Wallis
ANOVA. N/S: not significant.

8
Figure S9. Total and phospho-Pyk2 levels in the post-mortem brains of $O]KHLPHU¶V� GLVHDVH cases and healthy controls.
Demographic details of the cohort are given in Table S1. In box plots, red circles, red bars, and black squares indicate
individual data points, sample median and mean, respectively. Numbers of individual data points analyzed for each condition
are given in parentheses. Wilcoxon rank-sum test.

9
Figure S10. Localization of phospho-Pyk2 Tyr402 (p-Pyk2) puncta relative to synapses. Each Homer spot was assigned to the
nearest Synaptophysin (Syp) spot within cut-off distance of 1.0 µm. Each p-3\N��VSRW�ZDV�DVVLJQHG�WR�WKH�QHDUHVW�³V\QDSVH´��
i.e., to the midpoint between Syp and Homer spots, within cut-off distance of 1.5 µm. Such assignments were then categorized
as presynaptic or postsynaptic, if they were inside of 45° cones emanating from the midpoint towards Syp and Homer spots,
respectively. N = 5 devices per condition from two independent experiments. Paired t-test; * p < 5×10-4
; ** p < 5×10-5
.
Figure S11. An exemplary immunoblot demonstrating the overexpression of Pyk2 in primary neuronal cultures at DIV14
following lentiviral transduction at DIV7.
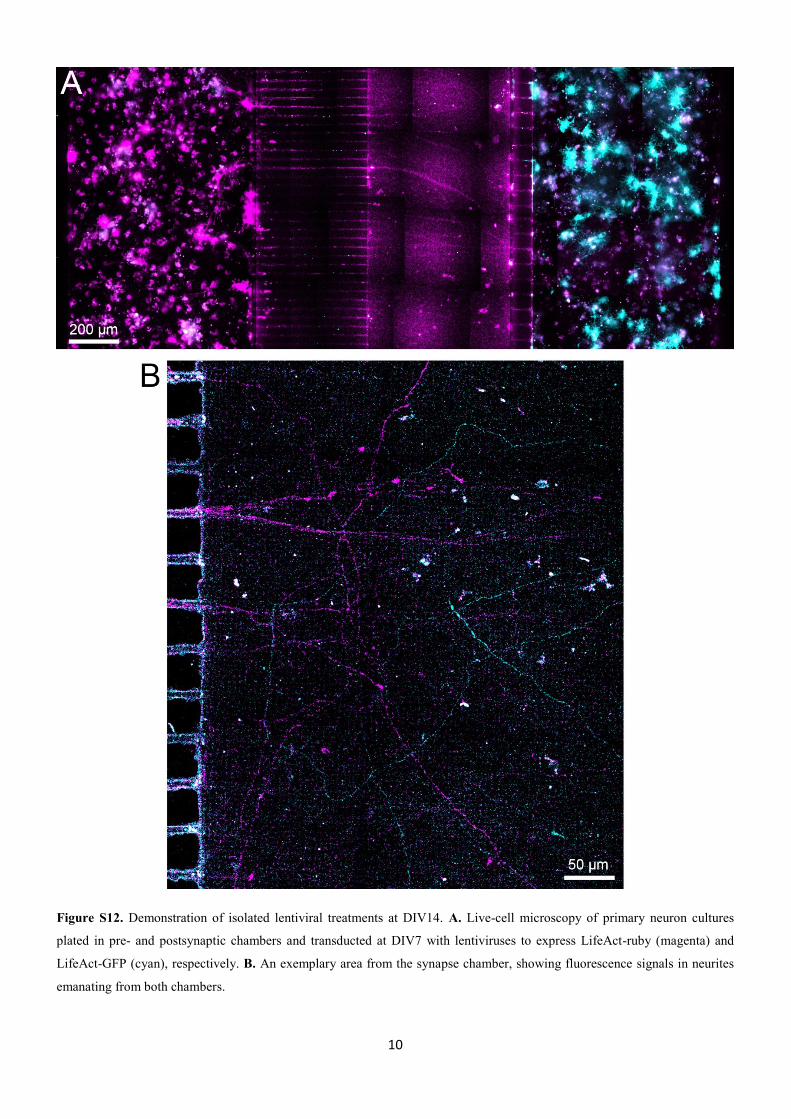
10
Figure S12. Demonstration of isolated lentiviral treatments at DIV14. A. Live-cell microscopy of primary neuron cultures
plated in pre- and postsynaptic chambers and transducted at DIV7 with lentiviruses to express LifeAct-ruby (magenta) and
LifeAct-GFP (cyan), respectively. B. An exemplary area from the synapse chamber, showing fluorescence signals in neurites
emanating from both chambers.
Copyright © 2022 FDOKUMEN




















