
Identification of plant semiochemicals and characterization of new olfactory sensory neuron types in...
15
Chem. Senses 39: 719–733, 2014 doi:10.1093/chemse/bju046 © The Author 2014. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected] Identification of Plant Semiochemicals and Characterization of New Olfactory Sensory Neuron Types in a Polyphagous Pest Moth, Spodoptera littoralis Muhammad Binyameen 1,2 , Peter Anderson 1 , Rickard Ignell 1 , Göran Birgersson 1 , Muhammad Razaq 2 , Sarfraz A. Shad 2 , Bill S. Hansson 1,3 and Fredrik Schlyter 1 1 Unit of Chemical Ecology, Department of Plant Protection Biology, Swedish University of Agricultural Sciences, PO Box 102, SE-23053 Alnarp, Sweden, 2 Insect Chemical Ecology Lab, Department of Entomology, Faculty of Agricultural Sciences and Technology, Bahauddin Zakariya University, Multan 60800, Pakistan and 3 Department of Evolutionary Neuroethology, Max Planck Institute for Chemical Ecology, Hans-Knöll-Strasse 8, D-07745 Jena, Germany Correspondence to be sent to: Muhammad Binyameen, Insect Chemical Ecology Lab, Department of Entomology, Faculty of Agricultural Sciences and Technology, Bahauddin Zakariya University, Multan 60800, Pakistan. e-mail: [email protected] Accepted August 7, 2014 Abstract Phytophagous insects use blends of volatiles released from plants to select hosts for feeding and oviposition. To behaviorally analyze complex blends, we need efficient and selective methods for elucidating neuron types, their ligands, and specific- ity. Gas chromatography-combined single sensillum recordings (GC-SSRs) from antennal olfactory sensilla of female moth, Spodoptera littoralis revealed 38 physiologically active peaks in the headspace volatile blends from both larvae-damaged cot- ton plants and lilac flowers. Using GC-combined mass spectrometry, 9 new physiologically active compounds were identified from damaged cotton and 11 from lilac compared with earlier electrophysiological studies using antennae of female S. litto- ralis. We characterized 14 novel classes of olfactory sensory neurons (OSNs). Among these, we found the first 2 ligands for a frequent type of short trichoid sensillum, for which no ligands were identified earlier. By using GC-SSR, a substantial increase in functional classes of OSNs and active compounds, 40% and 34% more, respectively, compared with recent studies using GC-electroantennogram or SSR using single compounds was detected. Compared with the estimated number of correspond- ing antennal olfactory receptors, the OSN classes now correspond to 83% of a likely maximum. The many specialist OSNs observed may facilitate behavioral confirmation of key plant volatiles in blends. Key words: GC-SSR, Kairomone, Lepidoptera, Noctuidae, olfactory sensory neurons, plant volatiles Introduction Volatile organic compounds emitted by plants mediate a variety of ecological interactions between plants and insects. To understand these interactions, the identification of the volatile compounds that play vital roles in triggering insect behaviors, such as attraction to plants for nectar feeding and oviposition, is essential (de Bruyne and Baker 2008; Riffell et al. 2009; Bruce and Pickett 2011). Although orientation and attraction of moths to plant volatiles have received some attention (Tingle and Mitchell 1992; Tasin et al. 2006; Del Socorro et al. 2010a; Saveer et al. 2012), only a hand- ful of bioactive compounds has been identified (Rojas 1999; Landolt et al. 2006; Del Socorro et al. 2010a, 2010b; Gregg et al. 2010; Saveer et al. 2012). This is probably due to the complexity of the odor blends released by plants, particu- larly in comparison with the more simple blends of, e.g., pheromones. From a practical perspective, unraveling the identity of nonpheromonal semiochemicals from both host and nonhost plants could provide sufficient insight for their future use in insect pest management (Cook et al. 2007; Schlyter 2012). The polyphagous moth Spodoptera littoralis uses a broad range of plants from different families as host plants (Anderson and Alborn 1999), but shows a clear species-specific preference for egg laying (Thöming et al. 2013). Behavioral studies have shown that plant volatiles are important cues driving the host- seeking behavior of S. littoralis, eliciting both attraction and by guest on April 8, 2016 http://chemse.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Identification of plant semiochemicals and characterization of new olfactory sensory neuron types in...

Chem. Senses 39: 719–733, 2014 doi:10.1093/chemse/bju046
© The Author 2014. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected]
Identification of Plant Semiochemicals and Characterization of New Olfactory Sensory Neuron Types in a Polyphagous Pest Moth, Spodoptera littoralis
Muhammad Binyameen1,2, Peter Anderson1, Rickard Ignell1, Göran Birgersson1, Muhammad Razaq2, Sarfraz A. Shad2, Bill S. Hansson1,3 and Fredrik Schlyter1 1Unit of Chemical Ecology, Department of Plant Protection Biology, Swedish University of Agricultural Sciences, PO Box 102, SE-23053 Alnarp, Sweden, 2Insect Chemical Ecology Lab, Department of Entomology, Faculty of Agricultural Sciences and Technology, Bahauddin Zakariya University, Multan 60800, Pakistan and 3Department of Evolutionary Neuroethology, Max Planck Institute for Chemical Ecology, Hans-Knöll-Strasse 8, D-07745 Jena, Germany
Correspondence to be sent to: Muhammad Binyameen, Insect Chemical Ecology Lab, Department of Entomology, Faculty of Agricultural Sciences and Technology, Bahauddin Zakariya University, Multan 60800, Pakistan. e-mail: [email protected]
Accepted August 7, 2014
Abstract
Phytophagous insects use blends of volatiles released from plants to select hosts for feeding and oviposition. To behaviorally analyze complex blends, we need efficient and selective methods for elucidating neuron types, their ligands, and specific-ity. Gas chromatography-combined single sensillum recordings (GC-SSRs) from antennal olfactory sensilla of female moth, Spodoptera littoralis revealed 38 physiologically active peaks in the headspace volatile blends from both larvae-damaged cot-ton plants and lilac flowers. Using GC-combined mass spectrometry, 9 new physiologically active compounds were identified from damaged cotton and 11 from lilac compared with earlier electrophysiological studies using antennae of female S. litto-ralis. We characterized 14 novel classes of olfactory sensory neurons (OSNs). Among these, we found the first 2 ligands for a frequent type of short trichoid sensillum, for which no ligands were identified earlier. By using GC-SSR, a substantial increase in functional classes of OSNs and active compounds, 40% and 34% more, respectively, compared with recent studies using GC-electroantennogram or SSR using single compounds was detected. Compared with the estimated number of correspond-ing antennal olfactory receptors, the OSN classes now correspond to 83% of a likely maximum. The many specialist OSNs observed may facilitate behavioral confirmation of key plant volatiles in blends.
Key words: GC-SSR, Kairomone, Lepidoptera, Noctuidae, olfactory sensory neurons, plant volatiles
Introduction
Volatile organic compounds emitted by plants mediate a variety of ecological interactions between plants and insects. To understand these interactions, the identification of the volatile compounds that play vital roles in triggering insect behaviors, such as attraction to plants for nectar feeding and oviposition, is essential (de Bruyne and Baker 2008; Riffell et al. 2009; Bruce and Pickett 2011). Although orientation and attraction of moths to plant volatiles have received some attention (Tingle and Mitchell 1992; Tasin et al. 2006; Del Socorro et al. 2010a; Saveer et al. 2012), only a hand-ful of bioactive compounds has been identified (Rojas 1999; Landolt et al. 2006; Del Socorro et al. 2010a, 2010b; Gregg et al. 2010; Saveer et al. 2012). This is probably due to the
complexity of the odor blends released by plants, particu-larly in comparison with the more simple blends of, e.g., pheromones. From a practical perspective, unraveling the identity of nonpheromonal semiochemicals from both host and nonhost plants could provide sufficient insight for their future use in insect pest management (Cook et al. 2007; Schlyter 2012).
The polyphagous moth Spodoptera littoralis uses a broad range of plants from different families as host plants (Anderson and Alborn 1999), but shows a clear species-specific preference for egg laying (Thöming et al. 2013). Behavioral studies have shown that plant volatiles are important cues driving the host-seeking behavior of S. littoralis, eliciting both attraction and
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

720 M. Binyameen et al.
repellency to plant odorants (Saveer et al. 2012; Zakir et al. 2013). Using gas chromatography-combined electroantenno-graphic detection (GC-EAD) analysis (Arn et al. 1975), the antennae of S. littoralis females have been shown to respond to compounds present in the volatile blends of undamaged and herbivore larvae-damaged host plants (Zakir et al. 2013) as well as flowers floral odor (Saveer et al. 2012). Moreover, single sensillum recordings (SSRs) have investigated responses of olfactory sensory neurons (OSNs) to plant odors impor-tant for host plant choice by S. littoralis (Anderson et al. 1995; Jönsson and Anderson 1999; Binyameen et al. 2012). These studies have characterized 32 functional classes of OSNs housed in basiconic, auricilic, and coeloconic sensilla. Moreover, 47 odorant receptors (ORs) and 12 ionotropic receptors (IRs) have been identified from male and female S. littoralis expressed sequence tag libraries and antennal transcriptomes (Olivier et al. 2011; Jacquin-Joly et al. 2012; Poivet et al. 2013). Assuming that each OSN class expresses a single OR or IR, this suggests that <55% of the OSN classes have hitherto been characterized in S. littoralis. Hence, there are likely voids in the odor space that could be filled by the identification of novel plant semiochemicals.
Using GC-combined SSR analysis, the aim of this study was to narrow this void by identifying novel biologically active compounds in headspace volatiles of lilac flowers, which strongly attract male and female S. littoralis (Kromann 2012; Saveer et al. 2012), and of cotton plants releasing con-specific larval-induced compounds that act as oviposition deterrents (Zakir et al. 2013). This method provides screen-ing for biologically active compounds in complex odors with high specificity and sensitivity and would be expected to detect substantially more active compounds from sources previously well-analyzed by GC-EAD (Wadhams 1982). In the study, we focus on OSNs housed in the most abundant sensillum types: basiconic and short trichoid sensilla.
GC-SSRs from antennal olfactory sensilla of female moth, S. littoralis revealed 38 physiologically active peaks in the headspace volatile blends from both larvae-damaged cotton plants and lilac flowers. Using GC-combined mass spectrom-etry, 9 new physiologically active compounds were identified from damaged cotton and 11 from lilac compared with ear-lier electrophysiological studies on antennae of female S. lit-toralis. We characterized 14 novel classes of OSNs. Among these, we found the first 2 ligands for a frequent type of short trichoid sensillum, for which no ligands were identified earlier. The many specialist OSNs observed may facilitate behavioral confirmation of key plant volatiles in blends.
Materials and methods
Plant headspace volatile collection
Plants
Cotton (Gossypium hirsutum L.; Malvaceae) plants were grown individually in a growth chamber maintained at
25 ± 2 °C and 65 ± 5% relative humidity. In addition to natu-ral light, artificial light was provided by positioning a light (Philips, SON-T, 400 W) 1 m above the plants. Because lilac (Syringa vulgaris L.; Oleaceae) flowers are seasonal, volatile collections were made from field collected (Alnarpsgården) detached flower bunches during June–July 2009.
Odor collection
Six-week-old cotton plants (6–8 true leaves) were subjected to S. littoralis larval feeding for 48 h before and during odor col-lection. In total, 12–16, 3rd–4th instar larvae were allowed to feed on each plant. Headspace collections were made accord-ing to Saveer et al. (2012), except that polyacetate oven bags (Toppits 3M, Melitta Scandinavia AB) were used instead of glass containers. An air plenum was created inside the bags, to avoid any physical damage to the leaves, by pumping 300 mL min−1 of charcoal-filtered air inside the bag and by taking out 150 mL min−1 through the volatile trapping adsorbent columns, using 12-V diaphragm vacuum pumps (KNF Neuberger). The adsorbent columns were made of Teflon tube (60 × 3 mm) and contained 50–60 mg of Super Q (Supelco). Volatile collections were performed for 12 h and extracts were obtained by eluting the adsorbent columns with 500 μL of redistilled n-hexane and then pooled and condensed, under a gentle stream of nitrogen, to a level so that an amount of 1 µL contained 10 min equiva-lents of aeration. Condensed extracts were used for chemical analyses and in electrophysiological recordings. The volatile samples were stored in air tight glass vials at −18 °C.
Headspace volatiles from lilac flowers were collected in the same way as by Saveer et al. (2012). The flower branches were then enclosed in a 5-L glass container, which was closed from the base with a glass fitting. The adsorption columns were made of glass tubes (60 × 6 mm), containing 150–200 mg of Super Q adsorbent between glass wool plugs.
Electrophysiology
Insects
Spodoptera littoralis were imported as field collected pupae from Egypt to establish a culture as described by Binyameen et al. (2012). Virgin females, 2–3 days of age, were used in the experiments.
Odor stimuli preparation and stimulation; stationary SSR
Synthetic references were used for electrophysiological char-acterization of antennal olfactory sensilla and for confirma-tion of chemical identity of newly identified compounds (chemicals name, CAS, and source details are given in Supplementary Table S1). The odor stimuli used for SSRs included a wide range of compounds, identified in volatile extracts from biological sources (Jönsson and Anderson 1999; Zakir et al. 2013) or compounds that had been used in physiological or behavioral studies on S. littoralis (Anderson
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 721
et al. 1993; Anderson et al. 1995; Galizia et al. 2002; Binyameen et al. 2012; Supplementary Table S1).
To identify the functional types of OSNs characterized in our previous study, stationary SSRs were performed using a set of diagnostic compounds (Binyameen et al. 2012), loaded in pasture pipettes, which were used at most three times for stimulation and were kept at −18 °C when not in use (Andersson et al. 2012). Briefly, a female moth was restrained in a plastic pipette tip with only the head protruding, and a tungsten wire, serving as a reference electrode, was inserted into the abdomen. The head was immobilized with dental wax, and one of the exposed antennae was fixed onto a glass microscope slide using double-sided sticky tape. SSRs were performed by using a tungsten electrode, which was electro-lytically sharpened by repeatedly immersing the tip into a syringe of 10% KNO2 solution. The recording electrode was attached to an olfactory probe (Syntech) and inserted into the base of a sensillum using a motorized micromanipulator equipped with a piezoceramic element (DC3001-MPM10; World Precision Instrument). Recordings were made under a Nikon Eclipse FN1 compound light microscope at ×750 magnification. Extracellular spikes of the OSNs were ampli-fied through an IDAC-4 interface amplifier (Syntech), dis-played on a computer screen, and their frequency recorded using Autospike (Syntech).
GC-combined single sensillum recordings
If a neuron responded to any of the 45 synthetic single stimuli (Supplementary Table S1), or to pipettes loaded with damaged cotton and lilac volatile extracts, we also stimu-lated the cell by combined gas chromatograph and SSR setup to individual constituents of 2 µL of extracts separated by the GC (Wadhams 1982). In the GC, a four-way cross Graphpack (Gerstel) installed at the end of the GC-column and fed by 4 psi N2, led half of the effluent into the charcoal-filtered and humidified air stream (1 L min−1) flushing over the insect antenna through a glass tube (14-cm long, 7-mm inner diameter), and the other half to the flame ionization detector (FID).
Thus, the activity of an OSN and the gas chromatogram of the components separated on the GC-column were recorded simultaneously. For GC-SSR recordings, the GC was equipped with a fused silica capillary column (30 m × 0.25 mm ID, coated with HP-5; df = 0.25 μm), and the GC oven temperature was programmed from 50 °C (2 min hold) at 8 °C min−1 to 250 °C (5 min hold). Hydrogen was used as the mobile phase at an average linear flow of 45 cm s−1. For other details, see Saveer et al. (2012).
Identification and quantification of plant volatiles
The GC-SSR active compounds were identified from the headspace volatile extracts of both plant types by GC-combined mass spectrometry (GC-MS; 6890 GC and
5975 MS; Agilent Technologies) analysis. The HP-5MS coated fused silica capillary column and the temperature program specifications were the same as that used in the GC-SSR experiments (mentioned earlier). In addition, to check for coeluting compounds on the HP-5 column, they were separated on a polar-fused silica capillary column DB-Wax (30 m × 0.25 mm ID, df = 0.25 μm). In this case, the GC oven temperature was programmed from 30 °C (3 min hold) at 8 °C min−1 to 225 °C (5 min hold). Two microliters of each extract sample were injected into the GC-MS by means of an autosampler (7683B Series, Agilent). Helium was used as a mobile phase in all GC-MS analyses, at an average lin-ear flow rate of 35 cm s−1. The identity of active compounds was determined by following a standard protocol (see the editorial in Journal of Chemical Ecology: Anonymous 2008), with Kovat’s retention indices and mass spectral libraries (National Institute of Standards and Technology, Wiley, and our custom Alnarp11). Synthetic standards were then used in GC-SSR and GC-MS to confirm the biological activity and chemical identification of OSN active peaks, respec-tively. For quantification of identified compounds in the bio-logical extracts, 2 µg of heptyl acetate (data not shown) was added to the extract as an internal quantification standard. Furthermore, 2 µL of synthetic standards of known concen-tration (10 ng µL−1) of each compound were injected in the GC-MS employing the same conditions and the response compared for quantification of compounds identified in the plant extracts.
Criteria for classification of functional OSN types
Classification of OSN classes was based on the same criteria used in our previous study (Binyameen et al. 2012), where OSNs were classified based on the morphological sensillum type, spike amplitude of the responding neuron (larger spike amplitude as A-neuron and smaller amplitude as B-neuron), and their receptive range. Confirmation of chemical identity of electrophysiologically active candidate compounds was verified by their synthetic standards run on the GC-SSR. An OSN was considered a novel OSN if its odor profile (response spectra) did not share all odors with any of the previous defined classes.
Results
Chemical analysis
Chromatograms produced from headspace extract of both plant types had the same number of active volatiles, 23 (Figure 1), but lilac had much larger amounts (x = 12 ng min−1 compound−1) than damaged cotton (x = 0.9). Eight compounds were shared between damaged cotton and lilac extracts; however, they differed in quantity, where again their titer for 7 of 8 compounds was much higher (2-fold or more) in lilac than in cotton (Table 1). Among the 23 GC-SSR
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

722 M. Binyameen et al.
Figure 1 Gas chromatograms of headspace volatiles from (A) larval-damaged cotton, Gossypium hirsutum plants and (B) flowers of lilac, Syringa vul-garis produced using HP-5 column. GC peaks with electrophysiological activity are given names (chemical identity). Standard protocol for identification was followed (Anonymous 2008); comparison of mass spectra with National Institute of Standards and Technology, Wiley, and Alnarp11 MS libraries to give candidate compounds and subsequent injection of candidate synthetic standards on GC-MS and GC-SSR. For the (E,E)-cosmene in lilac, a standard was lacking; hence, its identification remains tentative.
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 723
Table 1 List of neuronally active plant odorants identified through GC-SSR and GC-MS analyses listed in elution order on HP-5, and their amounts found in larval-damaged cotton plants and lilac flowers
Compound Present in both species
Odorants released (ng min–1)
Antennal active compounds identified earliera by GC-EAD studies on S. littoralis
Damaged cotton Lilac Damaged cotton Undamaged cotton Lilac
Toluene 0.2
(E)-2-Hexenal 0.4 1, 2
(Z)-3-Hexenol 0.3 1, 2
(E)-2-Hexenol 0.1
Isoamyl acetate 0.2
α-Pinene X 6.2 11
Camphene 0.7
Benzaldehyde X 0.2 15 3 3
β-Pinene X 1.2 5.8 1
Benzyl methyl ether 26 3
4-Methylanisole 2.0 3
Myrcene 2.5 1, 2 3
(Z)-3-Hexenyl acetate 1.7 1, 2 3
β-Phellandrene 0.5
(Z)-β-Ocimene X 0.02 5.5 3
(E)-β-Ocimene X 1.4 120 1, 2 3
Acetophenone 4.5 3
Unidentified MTb 0.4
(S)-(+)-Linalool 4.8 3
(R)-(–)-Linalool 0.2 1, 2 3
DMNTc 0.8 1, 2
Lilac aldehyde isomer 1 15 3
Lilac aldehyde isomer 2 5.6 3
(E)-2-Hexenyl butyrate 0.1
1,4-Dimethoxy benzene 92 3
Decanal 0.04 3
Estragolc 11 3
Lilac alcohol 10 3
Indole 0.01 1, 2
Methyl eugenol 8.3
β-Caryophyllene X 2.7 1.3 1, 2
(E)-β-Farnesene 0.05
α-Humulene X 0.7 4.7 2
α-Cubebene 2.5
(E,E)-α-Farnesene X 0.4 3.0 2
TMTTc 0.3 1, 2
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

724 M. Binyameen et al.
active compounds in cotton headspace, only six were major compounds (>1 ng min−1 from a plant surface); 4 monoter-penes, a sesquiterpene, and 1 green leaf volatile. A minor peak, the mass spectra of which could not be matched to any reference mass spectrum is listed as “unidentified” (Figure 1A, Table 2). Correspondingly, of the 23 GC-SSR active compounds in lilac flower headspace (Figure 1B), most were >1 ng min−1 and eight were >10 ng min−1, of which two were monoterpene hydrocarbons, one of each monoterpenic alcohol and aldehyde, 1 aromatic aldehyde, and 3 aromatic ethers (Table 1). Two of the peaks that were found only in the flower volatiles, the lilac aldehyde A and lilac aldehyde B, did not separate on the HP5 column (Figure 1B), but separated on the DB-WAX column in the GC-MS.
Physiology
SSRs with an extended panel of 47 stimuli and 170 success-ful GC-SSR runs revealed 33 functional classes of OSNs in basiconic sensilla (BC), 13 of which were novel, and 1 novel functional OSN class in short-trichoid (ST) sensilla (Table 2) Twelve novel plant volatiles were identified from the GC-SSR and GC-MS analysis (Table 1). Chemical iden-tity of active compounds was confirmed by their synthetic standards run on the GC-SSR, examples are shown for extracts in Figure 2.
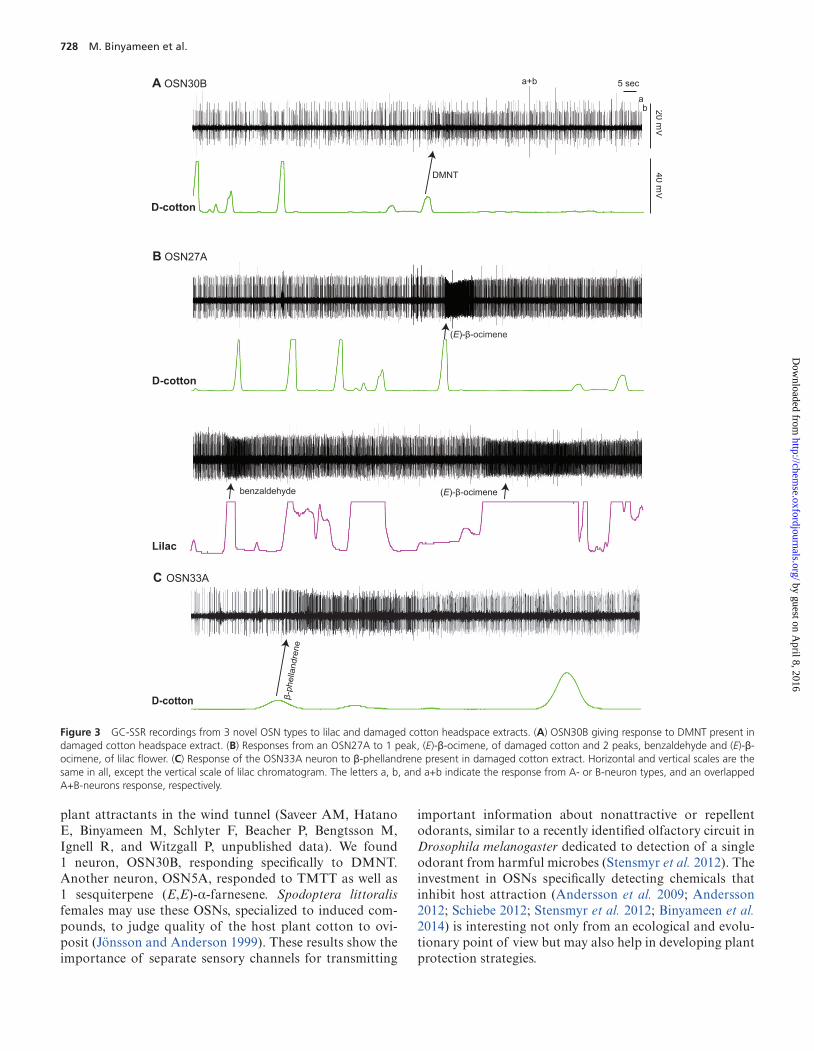
Among novel types, OSN30B responded with high speci-ficity to a herbivore-induced homoterpene, 4,8-dimethyl-1,(E)3,7-nonatriene (DMNT) (Figure 3A). Similarly, a total of seven novel OSNs were specific, responding to 1–3 stim-uli only (Table 2), There were also 3 clear generalists, e.g., OSN27A that strongly responded to 5 compounds (Table 2), including (E)-β-ocimene (Figure 3B). OSN33A responded to nine compounds from low to high sensitivity (Table 2, Figure 3C). Interestingly, the novel responses tended to fall among the volatiles in lowest titer, with the exception of α-pinene (Table 1).
Of the common ST sensilla morphological type, unrespon-sive in our earlier study, we found responses in one case; to DMNT and toluene in cotton extract. This OSN is referred to as ST-OSN1A (Table 2).
For the 20 OSN types previously encountered, 5 new ligands were found, two of which are sesquiterpene hydrocarbons, one of each aliphatic, aromatic, and sesquiterpenic alcohols, and 1 hemiterpenic ester (Table 3). For example, OSN5A, which previously was found specifically tuned to a homoter-pene 4,8,12-trimethyl-1,(E)3,(E)7,11-tridecatetraene (TMTT), now responded also to a sesquiterpene hydrocarbon, (E,E)-α-farnesene (Figure 2). Likewise, OSN3A, which previously responded to 2 sesquiterpene hydrocarbons, also responded to another sesquiterpene, α-cubebene in lilac (Figure 4). The OSN9B also responded to 6 new ligands, 1 sesquiterpene hydrocarbon, 1 short-chained alcohol, one of each aromatic and hemiterpenic esters, and 2 other compounds present either in the synthetic panel or in the biological extracts (Table 3).
Overall, the majority of OSNs were narrowly tuned to one, or at most, a few structurally related compounds, where 25 out of 34 OSN classes responded to only 1–3 compounds. The remaining nine OSN classes responded to 4–10 com-pounds (Tables 2 and 3). When restricting “narrowly tuned” only to those giving stronger responses (50 Hz or higher), we still find such neuron responses in 20 of 34 OSN classes, cor-responding to 59% OSNs.
Discussion
Novel compounds
Using an extended panel of synthetic plant-derived compounds and GC-SSR analysis of behaviorally active headspace extracts of cotton and lilac flowers, we have identified 38 compounds detected by OSNs of female S. littoralis. Twelve of these compounds are novel actives; toluene, camphene, α-pinene, β-phellandrene, (E,E)-cosmene, α-cubebene, (E)-β-farnesene, (E)2-hexenol, methyl eugenol, farnesol, (E)2-hexenyl butyrate, and isoamyl acetate, which have not been shown to elicit an electrophysiological response in S. littoralis previously (Jönsson and Anderson 1999; Saveer et al. 2012; Zakir et al. 2013).
An interesting observation is that earlier studies used the same plant species, but a different technique, i.e., GC-EAD instead of GC-SSR, compared with this study. Thus, the identification of novel compounds could be mainly due to
Compound Present in both species
Odorants released (ng min–1)
Antennal active compounds identified earliera by GC-EAD studies on S. littoralis
Damaged cotton Lilac Damaged cotton Undamaged cotton Lilac
Nerolidol 2.2 2
(E,E)-Farnesol 0.7
Number of compounds 8 23 23 14 5 12
New actives from this study are shown in bold.a1 = Jönsson and Anderson (1999); 2 = Zakir et al. (2013); 3 = Saveer et al. (2012).bUnidentified MT, tentatively identified as (E,E)-cosmene.cDMNT (E)-4,8 dimethyl-1,3,7-nonatriene; TMTT (E,E)-4,8,12-trimethyl-1,3,7,11-tridecatetraene; estragol = 1-methoxy-4-(2-propenyl)-benzene/4-allyl anisole.
Table 1 Continued
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 725
Table 2 Novel functional OSN class response spectra, 13 housed in basiconic sensilla (BC) and one in a short trichoid sensillum (ST)
Sensillum type BC ST
Compound OSN classes 25A 26A 26B 27A 28A 29A 30A 30B 31A 31B 32A 33A 34A 1A
Hydrocarbons
Aromatic
Toluene ● Monoterpenes
Camphene ●
Myrcene ● (Z)-β-Ocimene ● ●
(E)-β-Ocimene ● ● α-Pinene ● ● ● β-Pinene ● ● ● β-Phellandrene ● Unidentified MTa ● Sesquiterpenes
(E,E)-α-Farnesene ● Homoterpenes
DMNT ● ● ● TMTT ●Alcohols
Monoterpenes
Geraniol ● ● Lilac alcohol ● (S)-(+)-Linalool ●
Aldehydes
Aromatics
Benzaldehyde ● 2-Phenyl acetaldehyde ● ● Monoterpenes
Lilac aldehyde isomer 1 ● ●
Lilac aldehyde isomer 2 ● ●
Ethers
Aromatics
Benzyl methyl ether ● 1,4-Dimethoxy benzene ● 1-Methoxy-4-methyl
benzene● ●
Estragol DMNT, TMTT, and Estragol are explained above in Table1
●
Esters
Aliphatics
(E)-2-Hexenyl butyrate ● ●
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

726 M. Binyameen et al.
the employment of GC-SSR. The GC-EAD (Arn et al. 1975) is a standard and common method heavily used to screen for biologically active compounds (Barata et al. 2000; Zhang et al. 2000; Bengtsson et al. 2001; Fraser et al. 2003; Wibe 2004; Zakir et al. 2013). The GC-SSRs in this study allows us to describe a considerably wider chemosphere than previous GC-EAD studies, which clearly demonstrates that GC-EAD by itself may not be sufficient for identifying all compounds detected by the insect antenna (Wibe 2004). This may be due to the fact that the activity of small populations of OSNs results in small sums of receptor potentials, which can eas-ily lie below the threshold of signal detection in GC-EAD recordings. This also could be due to the fact that GC-SSR technique has a higher resolution compared with GC-EAD as in SSR the response is recorded from a single neuron instead of the whole antenna (Wibe 2004; Ghaninia et al. 2008). Therefore, GC-SSR could be a highly sensitive tool in detecting physiologically active components in the vola-tile of host plants, including compounds that do not elicit GC-EAD responses (Blight et al. 1995; Barata et al. 2002; Wibe 2004).
In S. littoralis, EAG appears to be a less suitable tool to detect all of the most relevant signals perceived by this polyphagous species. The importance of using GC-SSR has also been demonstrated for the cabbage seed weevil (Ceutorhynchus assimilis), where 5 volatile compounds of oil seed rape (Brassica napus) elicited responses in single OSNs, but did not elicit EAD response (Blight et al. 1995). Likewise, Barata et al. (2002), using GC-SSR, found six active peaks in Eucalyptus globulus, a host plant of eucalyptus woodborer (Phoracantha semipunctata) and two in a nonhost, Pinus
pinaster that were not found to be active in GC-EAD study (Barata et al. 2000). Several other studies have also reported similar observations (Bichão et al. 2003; Wibe 2004; Bichão et al. 2005), which suggest that GC-SSR is a better tool for identification of physiologically active odorants than the extensively used GC-EAD.
New OSN classes
Altogether 34 classes of OSNs, of which 14 were novel, responded to 51 odorants present in either the synthetic panel or plant headspace extracts. These odorants belonged to several compound classes, including green leaf alcohols and floral aldehydes, which have been shown to be highly attractive for noctuid female moths (Gregg et al. 2010). In addition, quite a few OSNs responded to terpenoid com-pounds, which are induced by herbivore attack, particularly homoterpenes that can inhibit the oviposition behavior of a female moth (Tholl et al. 2011; Zakir et al. 2013). The possible reason for this dedication of OSNs to detect her-bivore-induced plant volatiles (HIPVs) could be the fitness cost, because HIPVs may protect plants either directly by repelling or deterring herbivores, or indirectly by attracting the enemies of herbivores (Turlings and Wäckers 2004). The direct repellence might also be a result of the detection of a signal that indicates that enemies may be present. Detection of herbivore-induced chemicals in plants by S. littoralis has been previously shown both at antennal (Zakir et al. 2013) and neuronal levels (Jönsson and Anderson 1999).
Several of the compounds that we found in damaged cot-ton, e.g. (E,E)-α-farnesene, (E)-β-farnesene, (E)-β-ocimene,
Sensillum type BC ST
Compound OSN classes 25A 26A 26B 27A 28A 29A 30A 30B 31A 31B 32A 33A 34A 1A
(Z)-3-Hexenyl acetate ● Aromatics
Methyl benzoate ● ●
Methyl salicylate ●Others
Indole ●
Sulcatoneb ● ●
UNIDENTIFIEDc ●No. of encountered sensilla 3 2 5 2 3 3 2 2 1 4 2
The spike frequency (Hz) is used as a measure of response strength: (●) indicates 15–50 Hz; (●) 51–80Hz; (●) 81–110 Hz; (●) >110 Hz. The empty cells indicate that the compound did not elicit response in that OSN class. Neurons (A and B) were differentiated on the basis of spike amplitude, A with large and B with small amplitude. The OSNs classes 12, 19, 26, 30, and 31 contain 2 neurons (A & B) responding to different stimuli. No. of encountered sensilla indicates how many times the OSN was found. BC stands for basiconic; ST short trichoid sensilla. For other symbols and abbreviations, see Table 1. New actives from this study are shown in bold.aUnidentified MT, tentatively identified as (E,E)-cosmene.bbsulcatone = 6-methyl-5-hepten-2-one (terpenic).cUnidentified (MS indole-alike).
Table 2 Continued
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 727
linalool, indole, DMNT, and TMTT have been reported earlier in herbivore larvae-damaged cotton plants and are proposed to be produced de novo in response to insect feed-ing (McCall et al. 1994; Paré and Tumlinson 1997; Röse and Tumlinson 2005). Homoterpenes are common constituents
of floral and HIPV bouquets (Tholl et al. 2011). Two homoterpenes, DMNT and TMTT, are produced by angi-osperms with emissions from floral and vegetative tissues upon herbivore feeding. DMNT abolished upwind attrac-tion of S. littoralis females when added to a blend of host
Figure 2 GC-SSR recordings from OSN5A cells using damaged cotton and lilac headspace with synthetic standard candidates for the active peaks. (A) Two peaks, (E,E)-α-farnesene (new active from this study) and TMTT, from the damaged cotton headspace. (B) Response to (E,E)-α-farnesene peak present in lilac headspace. (C, D) OSN5A responses to separate GC-injections of 10 ng synthetic (E,E)-α-farnesene and TMTT, respectively. Horizontal and vertical scales are the same for all A–D.
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from
728 M. Binyameen et al.
plant attractants in the wind tunnel (Saveer AM, Hatano E, Binyameen M, Schlyter F, Beacher P, Bengtsson M, Ignell R, and Witzgall P, unpublished data). We found 1 neuron, OSN30B, responding specifically to DMNT. Another neuron, OSN5A, responded to TMTT as well as 1 sesquiterpene (E,E)-α-farnesene. Spodoptera littoralis females may use these OSNs, specialized to induced com-pounds, to judge quality of the host plant cotton to ovi-posit (Jönsson and Anderson 1999). These results show the importance of separate sensory channels for transmitting
important information about nonattractive or repellent odorants, similar to a recently identified olfactory circuit in Drosophila melanogaster dedicated to detection of a single odorant from harmful microbes (Stensmyr et al. 2012). The investment in OSNs specifically detecting chemicals that inhibit host attraction (Andersson et al. 2009; Andersson 2012; Schiebe 2012; Stensmyr et al. 2012; Binyameen et al. 2014) is interesting not only from an ecological and evolu-tionary point of view but may also help in developing plant protection strategies.
Figure 3 GC-SSR recordings from 3 novel OSN types to lilac and damaged cotton headspace extracts. (A) OSN30B giving response to DMNT present in damaged cotton headspace extract. (B) Responses from an OSN27A to 1 peak, (E)-β-ocimene, of damaged cotton and 2 peaks, benzaldehyde and (E)-β-ocimene, of lilac flower. (C) Response of the OSN33A neuron to β-phellandrene present in damaged cotton extract. Horizontal and vertical scales are the same in all, except the vertical scale of lilac chromatogram. The letters a, b, and a+b indicate the response from A- or B-neuron types, and an overlapped A+B-neurons response, respectively.
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 729
Table 3 Response spectra of 20 earlier known functional OSN classes housed in BC
Sensillum typea Basiconic (BC)
Compound ORN classes 1A 2A 3A 4A 5A 6A 7B 8A 9B 10A 11A 12A 12B 15A 16B 17B 18A 19A 19B 24A
Hydrocarbons
Sesquiterpenes
β-Caryophyllene ● α-Cubebeneb ● (E,E)-α-Farneseneb ● (E)-β-Farneseneb ● α-Humulene ● ● Homoterpenes
TMTTc ●Alcohols
Aliphatic
1-Hexanol ● ● ● (E)-2-Hexenol ● ● ● (Z)-3-Hexenol ● ● ● 1-Heptanol ● ●
Aromatic
Benzyl alcoholb ● ● Methyl eugenolb ●
Monotepenes
Carvacrol ●
(S)-(+)-Linalool ● (R)-(−)-Linalool ● ● Sesquiterpenes
(±)-Nerolidol ●
(E,E)-Farnesol ●Aldehydes
Aliphatic
(E)-2-Hexenal ● ● Decanalb ●
Aromatic
Benzaldehyde ● ● ● ●
2-Phenyl acetaldehyde
● ● ● ● ● ●
Esters
Aliphatic
(Z)-3-Hexenyl acetate ● ● Hemiterpenic
Isoamyl acetateb,c ●
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

730 M. Binyameen et al.
Specificity of OSN responses
We, by GC-SSR, found 7 of the 14 new OSNs highly selec-tive (responding to 1–3 odorants) compared with earlier data. Selectivity of OSNs is not only important for understanding the olfactory coding mechanisms but also for the identifica-tion of odorants, which are the most ecologically relevant to
the moth and should thus be tested in behavioral assays. It has been believed that plant odor-detecting OSNs are broadly tuned, particularly in comparison to pheromone OSNs, but the picture is changing. This may be due to the fact that most previous studies have used high (i.e., not ecologically relevant) stimulus concentrations, especially of highly volatile host
Sensillum typea Basiconic (BC)
Compound ORN classes 1A 2A 3A 4A 5A 6A 7B 8A 9B 10A 11A 12A 12B 15A 16B 17B 18A 19A 19B 24A
Aromatic
Methyl salicylateb ●Others
Methyl jasmonateb ● (Z)-Jasmone ● ●
Acetophenoned,b ● ● Indole ● ●
No. of encountered sensilla
5 4 8 2 8 3 4 7 4 3 2 2 2 3 1 1 3 4
New actives from this study are shown in bold. For other symbols and abbreviations, see Table 1.aBinyameen et al. (2012).bIndicates this compound is a new ligand for that OSN.cIsoamyl acetate = 3-methybutyl acetate (terpenic).dAcetophenone = 1-phenylethanone (aromatic).
Table 3 Continued
Figure 4 Examples of GC-SSR from OSN3A cells showing spontaneous activity and responses to damaged cotton and lilac headspace extracts. (A) OSN3A housed in a basiconic sensillum responding to 2 peaks, β-caryophyllene and α- humulene, of damaged cotton headspace. (B) OSN3A responses to 2 peaks, β-caryophyllene and α-cubebene, of lilac headspace. Horizontal and vertical scales are same for A and B.
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 731
compounds (Andersson et al. 2012). However, when biologi-cally relevant compounds and ecologically relevant stimulus concentrations are used, OSNs narrowly tuned to plant odor-ant have been consistently found in several moth and beetle species (Hansson 1999; Røstelien et al. 2000a, 2000b; Shields and Hildebrand 2000; Wibe 2004; Ignell and Hansson 2005; Røstelien et al. 2005; Andersson et al. 2009; Andersson 2012; Schiebe 2012). In this study, we found that most novel OSNs have selective responses to one, or a few structurally related components in the complex mixtures, released by damaged cot-ton and lilac flowers, suggest that these OSNs are specialized rather than being broadly tuned to many plant compounds. Similarly, in heliothine moths, several OSNs responding selec-tively to monoterpene and sesquiterpene hydrocarbons have been characterized (Røstelien et al. 2000a, 2000b; Stranden et al. 2002, 2003; Røstelien et al. 2005). One interesting exam-ple of a sensitive and selective nonpheromonal neuronal type has been found in heliothine species which responds to the ses-quiterpene germacrene D at picogram doses (Røstelien et al. 2000b). Dose–response studies showed many-fold higher effect of (−)-germacrene D than of the (+)-enantiomer and much lower, if any, effects of a few other sesquiterpenes (Stranden et al. 2002). Similarly, Svensson and Larsson (2008) found kairomonal enantioselective OSNs in 2 species of beetles, Osmoderma eremita and Elater ferrugineus.
Our results show that the antennae of female S. littoralis are equipped with both specialized and more broadly tuned OSNs. The majority of the OSNs responding to >1 odorant, responded to structurally similar odorants, or displayed sec-ondary (weak) responses to structurally dissimilar odorants. We also found some OSNs responding with high sensitivity to several apparently structurally dissimilar compounds, for example, OSN2A that responded equally well to 2-phenyl acetaldehyde (PAA) and (Z)-3-hexenyl acetate. However, the female moth also has a specialized neuron, OSN1A, for detect-ing PAA. It can be hypothesized that the OSN2A responding to both odorants may express 2 ORs, as it has been shown in D. melanogaster, where a single neuron equipped with 2 ORs was found (Goldman et al. 2005). We hope that future studies using heterologous expression system may test this hypoth-esis of double expression in this polyphagous moth.
In contrast, most odorants showed little or no overlap (i.e., elicited response in only 1 neuron), suggesting that informa-tion about many compounds is conveyed to the brain by a single channel. In this study, for example, several odor-ants such as β-caryophyllene, α-cubebene, (E)-β-farnesene, myrcene, camphene, methyl jasmonate, isoamyl acetate, (E,E)-cosmene, β-phellandrene, toluene, estragol, benzyl methyl ether, 1,4-dimethoxy benzene, lilac alcohol, methyl eugenol, (E,E)-farnesol, and decanal elicited responses in only single OSNs. Our results also show that the receptive range of OSNs tuned to plant odorants ranges from specific to broadly tuned, which may correspond to both of the two, the principles by which information about an odorant is transmitted to the brain; either through “labeled lines” and/
or “across-fiber patterns/combinatorial coding” (Malnic et al. 1999; Hallem et al. 2004; Hallem and Carlson 2006; Leal 2013). However, more recent studies postulate that under restrictive experimental conditions, these networks lead to an apparent existence of “labeled line” coding for spe-cial odors (Stensmyr et al. 2012; Galizia 2014). Our data also strengthens the idea of “labeled lines” as we find several new cases of OSN with quite specific responses. The earlier often found less specific responses, corresponding to “across-fiber patterns” suggested for plant compounds, could be a con-sequence of screening doses used in earlier studies (Hallem et al. 2004; Hallem and Carlson 2006) combined with the high volatility of many plant compound causing high com-pound fluxes (concentrations) in stationary SSR (Andersson et al. 2012), compared with the low and known titers pre-sented by GC-SSR in our study.
Overall, many OSNs associated with BC sensilla responded to the biological volatile collections tested. We assume that other sensillum types also described from S. littoralis antennae (coeloconic, auricillic, grooved peg; Binyameen et al. 2012) as well as the predominant ST sensilla (of which, we character-ized only one for the first time in the present study), might be involved in the detection of other volatile blends than those tested here. It is likely that many odorants from other sources, such as microbes or nonhost plants, activating these other morphological sensilla types still remain undiscovered.
In conclusion, GC-SSRs, compared with earlier GC-EAD studies, revealed that the peripheral olfactory system of female S. littoralis can detect many more than earlier known ecologically relevant compounds, which could be used to locate potential food sources and suitable reproduction sites. This study has increased by 40–50% the number of plant odorants known to be encoded by a now larger number of OSNs (from 35 to 49) in female S. littoralis. Furthermore, the current study strengthens the concept of OSN specific-ity in a generalist feeder as many types of OSNs are specifi-cally tuned to one or a few structurally related compounds, while only some overlap in their response spectra. In addi-tion to the discovery of novel classes of OSNs and biological active ligands from ecological sources and OSNs specificity, this study widens the platform for the deorphanization of ORs that will elucidate underlying molecular mechanisms of olfaction in this polyphagous pest species (Jacquin-Joly et al. 2012; Poivet et al. 2013).
Supplementary material
Supplementary material can be found at http://www.chemse.oxfordjournals.org/
Funding
This work was supported by Linnaeus initiative grant [217-2006-1750] IC-E3 (Insect Chemical Ecology, Ethology and
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

732 M. Binyameen et al.
Evolution), from FORMAS (The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning), and SLU (Swedish University of Agricultural Sciences).
AcknowledgementsWe thank Elisabeth Marling for her help in insect rearing. Dr M.S. Ahmed and Dr A.S.H. Kromann are acknowledged for their help and technical advice in headspace odor collection. We also thank Dr M.N. Andersson, Dr W. Walker III, and Dr E. Jacquin-Joly for their valuable comments on an earlier version of the manu-script. Higher Education Commission (HEC) of Pakistan is highly acknowledged for providing scholarship grant for PhD studies to M. Binyameen under the scheme “Overseas Scholarship for MS/MPhil leading to PhD Studies, Phase II batch II, Sweden.” Support for M.B. in analysis of data and revision of manuscript from Department of Entomology, Bahauddin Zakariya University is greatly acknowledged.
ReferencesAnderson P, Alborn H. 1999. Effects on oviposition behaviour and larval
development of Spodoptera littoralis by herbivore induced changes in cotton plants. Entomol Exp Appl. 92:45–51.
Anderson P, Hansson BS, Löfqvist J. 1995. Plant-odour-specific receptor neurones on the antennae of female and male Spodoptera littoralis. Physiol Entomol. 20:189–198.
Anderson P, Hilker M, Hansson BS, Bombosch S, Klein B, Schildknecht H. 1993. Oviposition deterring components in larval frass of Spodoptera littoralis (Boisd) (Lepidoptera, Noctuidae) - a behavioral and electrophys-iological evaluation. J Insect Physiol. 39:129–137.
Andersson MN. 2012. Mechanisms of odor coding in coniferous bark bee-tles: from neuron to behavior and application. Psyche: J Entomol. 2012. Article ID 149572.
Andersson MN, Larsson MC, Schlyter F. 2009. Specificity and redundancy in the olfactory system of the bark beetle Ips typographus: single-cell responses to ecologically relevant odors. J Insect Physiol. 55:556–567.
Andersson MN, Schlyter F, Hill SR, Dekker T. 2012. What reaches the antenna? How to calibrate odor flux and ligand-receptor affinities. Chem Senses. 37:403–420.
Anonymous. 2008. General guidelines for authors for submission of man-uscripts that contain identifications and syntheses of compounds. J Chem Ecol. 34:984–986.
Arn H, Städler E, Rauscher S. 1975. The electroantennographic detector—a selective and sensitive tool in the gas chromatographic analysis of insect pheromones. Zeitschrift für Naturforschung A. 30:722–725.
Barata N, Mustaparta H, Pickett JA, Wadhams LJ, Araujo J. 2002. Encoding of host and non-host plant odours by receptor neurones in the eucalyp-tus woodborer, Phoracantha semipunctata (Coleoptera: Cerambycidae). J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 188:121–133.
Barata EN, Pickett JA, Wadhams LJ, Woodcock CM, Mustaparta H. 2000. Identification of host and nonhost semiochemicals of eucalyptus wood-borer Phoracantha semipunctata by gas chromatography–electroanten-nography. J Chem Ecol. 26:1877–1895.
Bengtsson M, Bäckman AC, Liblikas I, Ramirez MI, Borg-Karlson AK, Ansebo L, Anderson P, Löfqvist J, Witzgall P. 2001. Plant odor
analysis of apple: antennal response of codling moth females to apple volatiles during phenological development. J Agric Food Chem. 49:3736–3741.
Bichão H, Borg-Karlson AK, Araújo J, Mustaparta H. 2003. Identification of plant odours activating receptor neurones in the weevil Pissodes nota-tus F. (Coleoptera, Curculionidae). J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 189:203–212.
Bichão H, Borg-Karlson AK, Araújo J, Mustaparta H. 2005. Five types of olfactory receptor neurons in the strawberry blossom weevil Anthonomus rubi: selective responses to inducible host-plant volatiles. Chem Senses. 30:153–170.
Binyameen M, Anderson P, Ignell R, Seada MA, Hansson BS, Schlyter F. 2012. Spatial organization of antennal olfactory sensory neurons in the female Spodoptera littoralis moth: differences in sensitivity and tempo-ral characteristics. Chem Senses. 37:613–629.
Binyameen M, Jankuvová J, Blaženec M, Jakuš R, Song L, Schlyter F, Andersson MN. 2014. Co-localisation of insect olfactory sensory cells improves the discrimination of closely separated odour sources. Funct Ecol. doi: 10.1111/1365-2435.12252.
Blight MM, Pickett JA, Wadhams LJ, Woodcock CM. 1995. Antennal percep-tion of oilseed rape, Brassica napus (Brassicaceae), volatiles by the cab-bage seed weevil Ceutorhynchus assimilis (Coleoptera, Curculionidae). J Chem Ecol. 21:1649–1664.
Bruce TJ, Pickett JA. 2011. Perception of plant volatile blends by herbivo-rous insects–finding the right mix. Phytochemistry. 72:1605–1611.
Cook SM, Khan ZR, Pickett JA. 2007. The use of push-pull strategies in integrated pest management. Annu Rev Entomol. 52:375–400.
de Bruyne M, Baker TC. 2008. Odor detection in insects: volatile codes. J Chem Ecol. 34:882–897.
Del Socorro AP, Gregg PC, Alter D, Moore CJ. 2010a. Development of a synthetic plant volatile-based attracticide for female noctuid moths. I. Potential sources of volatiles attractive to Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Aust J Entomol. 49:10–20.
Del Socorro AP, Gregg PC, Hawes AJ. 2010b. Development of a synthetic plant volatile-based attracticide for female noctuid moths. III. Insecticides for adult Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Aust J Entomol. 49:31–39.
Fraser AM, Mechaber WL, Hildebrand JG. 2003. Electroantennographic and behavioral responses of the sphinx moth Manduca sexta to host plant headspace volatiles. J Chem Ecol. 29:1813–1833.
Galizia CG. 2014. Olfactory coding in the insect brain: data and conjec-tures. Eur J Neurosci. 39:1784–1795.
Galizia CG, Carlsson MA, Hansson BS. 2002. Spatial representation of odours in the antennal lobe of the moth Spodoptera littoralis (Lepidoptera: Noctuidae). Chem Senses. 27:231–244.
Ghaninia M, Larsson M, Hansson BS, Ignell R. 2008. Natural odor ligands for olfactory receptor neurons of the female mosquito Aedes aegypti: use of gas chromatography-linked single sensillum recordings. J Exp Biol. 211(Pt 18):3020–3027.
Goldman AL, Van der Goes van Naters W, Lessing D, Warr CG, Carlson JR. 2005. Coexpression of two functional odor receptors in one neuron. Neuron. 45:661–666.
Gregg PC, Del Socorro AP, Henderson GS. 2010. Development of a syn-thetic plant volatile-based attracticide for female noctuid moths. II. Bioassays of synthetic plant volatiles as attractants for the adults of the cotton bollworm, Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Aust J Entomol. 49:21–30.
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from

Identification of Plant Semiochemicals and Characterization of OSNs using GC-SSR 733
Hallem EA, Carlson JR. 2006. Coding of odors by a receptor repertoire. Cell. 125:143–160.
Hallem EA, Ho MG, Carlson JR. 2004. The molecular basis of odor coding in the Drosophila antenna. Cell. 117:965–979.
Hansson BS. 1999. Insect olfaction. Heidelberg Berlin: Springer-Verlag.
Ignell R, Hansson BS. 2005. Insect olfactory neuroethology: an electrophysi-ological perspective. In: Christensen TA, editor. Methods in insect sen-sory neuroscience. Boca Raton (FL): CRC Press. p. 319–347.
Jacquin-Joly E, Legeai F, Montagné N, Monsempes C, François MC, Poulain J, Gavory F, Walker WB 3rd, Hansson BS, Larsson MC. 2012. Candidate chemosensory genes in female antennae of the noctuid moth Spodoptera littoralis. Int J Biol Sci. 8:1036–1050.
Jönsson M, Anderson P. 1999. Electrophysiological response to herbivore induced host plant volatiles in the moth Spodoptera littoralis. Physiol Entomol. 24:377–385.
Kromann ASH. 2012. Modulation of olfactory information in the anten-nal lobe of Spodoptera littoralis. Acta Universitatis agriculturae Sueciae. Vol. 2012:60. Alnarp, Sweden: Swedish University of Agricultural Sciences. Available from: http://pub.epsilon.slu.se/9021/1/Kromann_ash_120904.pdf.
Landolt PJ, Adams T, Zack RS. 2006. Field response of alfalfa looper and cabbage looper moths (Lepidoptera: Noctuidae, Plusiinae) to single and binary blends of floral odorants. Environ Entomol. 35:276–281.
Leal WS. 2013. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes. Annu Rev Entomol. 58:373–391.
Malnic B, Hirono J, Sato T, Buck LB. 1999. Combinatorial receptor codes for odors. Cell. 96:713–723.
McCall PJ, Turlings TC, Loughrin J, Proveaux AT, Tumlinson JH. 1994. Herbivore-induced volatile emissions from cotton (Gossypium hirsutum L.) seedlings. J Chem Ecol. 20:3039–3050.
Olivier V, Monsempes C, François MC, Poivet E, Jacquin-Joly E. 2011. Candidate chemosensory ionotropic receptors in a Lepidoptera. Insect Mol Biol. 20:189–199.
Paré PW, Tumlinson JH. 1997. De novo biosynthesis of volatiles induced by insect herbivory in cotton plants. Plant Physiol. 114:1161–1167.
Poivet E, Gallot A, Montagné N, Glaser N, Legeai F, Jacquin-Joly E. 2013. A comparison of the olfactory gene repertoires of adults and larvae in the noctuid moth Spodoptera littoralis. PLoS One. 8:e60263.
Riffell JA, Lei H, Christensen TA, Hildebrand JG. 2009. Characterization and coding of behaviorally significant odor mixtures. Curr Biol. 19:335–340.
Rojas JC. 1999. Electrophysiological and behavioral responses of the cab-bage moth to plant volatiles. J Chem Ecol. 25:1867–1883.
Röse US, Tumlinson JH. 2005. Systemic induction of volatile release in cot-ton: how specific is the signal to herbivory? Planta. 222:327–335.
Røstelien T, Borg-Karlson AK, Fäldt J, Jacobsson U, Mustaparta H. 2000a. The plant sesquiterpene germacrene D specifically activates a major type of antennal receptor neuron of the tobacco budworm moth Heliothis virescens. Chem Senses. 25:141–148.
Røstelien T, Borg-Karlson AK, Mustaparta H. 2000b. Selective receptor neu-rone responses to E-beta-ocimene, beta-myrcene, E,E-alpha-farnesene and homo-farnesene in the moth Heliothis virescens, identified by gas chroma-tography linked to electrophysiology. J Comp Physiol A. 186:833–847.
Røstelien T, Stranden M, Borg-Karlson AK, Mustaparta H. 2005. Olfactory receptor neurons in two Heliothine moth species responding selectively to aliphatic green leaf volatiles, aromatic compounds, monoterpenes and sesquiterpenes of plant origin. Chem Senses. 30:443–461.
Saveer AM, Kromann SH, Birgersson G, Bengtsson M, Lindblom T, Balkenius A, Hansson BS, Witzgall P, Becher PG, Ignell R. 2012. Floral to green: mating switches moth olfactory coding and preference. Proc Biol Sci. 279(1737):2314–2322.
Schiebe C. 2012. Attraction and resistance in the Picea abies-Ips typogra-phus system: host choice in the Eurasian spruce bark beetle [PhD-thesis]. Sweden: Unit of Chemical Ecology, Department of Plant Protection Biology, Swedish University of Agricultural Sciences, SE-23053, Alnarp, Sweden.
Schlyter F. 2012. Semiochemical diversity in practice: antiattractant semio-chemicals reduce bark beetle attacks on standing trees. A first meta-analysis. Psyche: A Journal of Entomology, vol, 2012. Article ID 268621.
Shields VD, Hildebrand JG. 2000. Responses of a population of antennal olfactory receptor cells in the female moth Manduca sexta to plant-asso-ciated volatile organic compounds. J Comp Physiol A. 186:1135–1151.
Stensmyr MC, Dweck HK, Farhan A, Ibba I, Strutz A, Mukunda L, Linz J, Grabe V, Steck K, Lavista-Llanos S, et al. 2012. A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell. 151:1345–1357.
Stranden M, Borg-Karlson AK, Mustaparta H. 2002. Receptor neuron dis-crimination of the germacrene D enantiomers in the moth Helicoverpa armigera. Chem Senses. 27:143–152.
Stranden M, Røstelien T, Liblikas I, Almaas TJ, Borg-Karlson AK, Mustaparta H. 2003. Receptor neurones in three Heliothine moths responding to floral and inducible plant volatiles. Chemoecology. 13:143–154.
Svensson GP, Larsson MC. 2008. Enantiomeric specificity in a pheromone-kairomone system of two threatened saproxylic beetles, Osmoderma eremita and Elater ferrugineus. J Chem Ecol. 34:189–197.
Tasin M, Bäckman AC, Bengtsson M, Ioriatti C, Witzgall P. 2006. Essential host plant cues in the grapevine moth. Naturwissenschaften. 93:141–144.
Tholl D, Sohrabi R, Huh JH, Lee S. 2011. The biochemistry of homoterpe-nes–common constituents of floral and herbivore-induced plant volatile bouquets. Phytochemistry. 72:1635–1646.
Thöming G, Larsson MC, Hansson BS, Anderson P. 2013. Comparison of plant preference hierarchies of male and female moths and the impact of larval rearing hosts. Ecology. 94:1744–1752.
Tingle FC, Mitchell ER. 1992. Attraction of Heliothis virescens (F.) (Lepidoptera: Noctuidae) to volatiles from extracts of cotton flowers. J Chem Ecol. 18:907–914.
Turlings TCJ, Wäckers F. 2004. Recruitment of predators and parasitoids by herbivore-injured plants. In: Cardé RT, Millar JG, editors. Advances in insect chemical ecology. Vol. 2. Cambridge: Cambridge University Press. p. 21–75.
Wadhams L. 1982. Coupled gas chromatography–single cell recording: a new technique for use in the analysis of insect pheromones. Zeitschrift Fur Naturforschung C. 37:947–952.
Wibe A. 2004. How the choice of method influence on the results in elec-trophysiological studies of insect olfaction. J Insect Physiol. 50:497–503.
Zakir A, Bengtsson M, Sadek MM, Hansson BS, Witzgall P, Anderson P. 2013. Specific response to herbivore-induced de novo synthesized plant volatiles provides reliable information for host plant selection in a moth. J Exp Biol. 216(Pt 17):3257–3263.
Zhang Q-H, Schlyter F, Birgersson G. 2000. Bark volatiles from nonhost angi-osperm trees of spruce bark beetle, Ips typographus (L.) (Coleoptera: Scolytidae): Chemical and electrophysiological analysis. Chemoecology. 10:69–80.
by guest on April 8, 2016
http://chemse.oxfordjournals.org/
Dow
nloaded from




















