
Identification, Characterization, and Variable Expression of a Naturally Occurring Inhibitor Protein...
12
INFECTION AND IMMUNITY, 0019-9567/01/$04.000 DOI: 10.1128/IAI.69.12.7425–7436.2001 Dec. 2001, p. 7425–7436 Vol. 69, No. 12 Copyright © 2001, American Society for Microbiology. All Rights Reserved. Identification, Characterization, and Variable Expression of a Naturally Occurring Inhibitor Protein of IS1106 Transposase in Clinical Isolates of Neisseria meningitidis PAOLA SALVATORE, 1,2 CATERINA PAGLIARULO, 1 ROBERTA COLICCHIO, 1 PATRIZIA ZECCA, 3 GIUSEPPINA CANTALUPO, 1 MAURIZIO TREDICI, 3 ALFREDO LAVITOLA, 1 CECILIA BUCCI, 1,3 CARMELO B. BRUNI, 1 * AND PIETRO ALIFANO 3 * Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano,” Universita ` di Napoli “Federico II,” and Centro di Endocrinologia ed Oncologia Sperimentale “G. Salvatore” of the Consiglio Nazionale delle Ricerche, 80131 Naples, 1 Dipartimento di Scienze Ambientali, Seconda Universita ` di Napoli, 81100 Caserta, 2 and Dipartimento di Scienze e Tecnologie Biologiche e Ambientali, Universita ` degli Studi di Lecce, 73100 Lecce, 3 Italy Received 17 July 2001/Returned for modification 6 September 2001/Accepted 19 September 2001 Transposition plays a role in the epidemiology and pathogenesis of Neisseria meningitidis. Insertion se- quences are involved in reversible capsulation and insertional inactivation of virulence genes encoding outer membrane proteins. In this study, we have investigated and identified one way in which transposon IS1106 controls its own activity. We have characterized a naturally occurring protein (Tip) that inhibits the trans- posase. The inhibitor protein is a truncated version of the IS1106 transposase lacking the NH 2 -terminal DNA binding sequence, and it regulates transposition by competing with the transposase for binding to the outside ends of IS1106, as shown by gel shift and in vitro transposition assays. IS1106Tip mRNA is variably expressed among serogroup B meningococcal clinical isolates, and it is absent in most collection strains belonging to hypervirulent lineages. Many studies have pointed out the importance of mobile genetic elements in microbial pathogenesis and adaptation to changing environmental conditions. Virulence genes of patho- genic bacteria, which code for toxins, adhesins, invasins, cap- sules, pili, resistance determinants, or other virulence factors, may be located on transmissible genetic elements such as transposons, plasmids, or bacteriophages (16, 50). Insertion sequence (IS) elements are known to be involved in microevo- lution of bacterial genomes by several mechanisms (1, 32). (i) In the chromosomes of both gram-negative and gram-positive bacteria, virulence genes are often clustered in “virulence blocks” or “pathogenicity islands,” surrounded by IS elements that promote their transposition and lead to changes in viru- lence in the course of evolution. Pathogenicity islands are invariably found in pathogenic strains of a given species but are either absent or rarely present in nonpathogenic variants of the same species (12, 20, 21, 23, 30, 33, 34). (ii) In addition, two copies of certain IS elements flanking a DNA segment are able to act in concert, mobilizing the intervening region. (iii) Pro- grammed insertion and excision of IS elements and of invert- ible DNA sequences may control the expression of several virulence factors by a mechanism of phase variation (24, 58). (iv) “Jumping” of DNA sequences (transposition) and subse- quent recombination events may also cause gene activation (5) and antigenic variation of virulence factors, leading to the emergence of novel pathogenic variants (48). (v) IS-related DNA rearrangements do occur in resting bacterial cultures and confer plasticity on the genome under conditions of nu- tritional deprivation, thereby playing an adaptive role (1, 3, 18, 35, 36). With the development of studies of the mechanisms of bac- terial pathogenesis and the advent of whole-genome sequenc- ing technologies in recent years, the finding of association between IS elements and pathogenic and virulence functions has become increasingly evident. Such associations have been observed in a variety of animal pathogens (6, 9, 14, 17, 31, 52). Whole-genome sequence analysis and subtractive hybridiza- tion procedures have led to the identification of putative is- lands of horizontally transferred DNA into the genomes of serogroup B and serogroup A meningococci (42, 55). Several of these regions encode proteins that are specific to the patho- genic Neisseria species and may have a role in virulence. These regions do not have the classical characteristics of pathogenic- ity islands. Several, however, have a particularly low GC content and are associated with transposase and integrase genes, suggesting that at some time in the genetic history of these species, the regions were the results of recombination events with DNAs from other species. One of these regions in a serogroup A strain, characterized by a significantly low GC content and containing open reading frames (ORFs) with no homology to genes in databases, is flanked by several copies of IS1106 and a copy of IS50 (42). Transposition plays a role in the epidemiology and patho- genesis of Neisseria meningitidis. IS1301 is involved in revers- ible capsulation by insertion into and excision from the siaA gene locus in serogroup B meningococci (24). This transpos- * Corresponding author. Mailing address for C. B. Bruni: Diparti- mento di Biologia e Patologia Cellulare e Molecolare “L. Califano,” Universita ` di Napoli “Federico II,” and Centro di Endocrinologia ed Oncologia Sperimentale “G. Salvatore” of the Consiglio Nazionale delle Ricerche, Via S. Pansini 5, 80131 Naples, Italy. Phone: (39) 081 7462047. Fax: (39) 081 7703285. E-mail: [email protected]. Mailing address for P. Alifano: Dipartimento di Scienze e Tecnologie Bio- logiche e Ambientali, Universita ` degli Studi di Lecce, Via Monteroni, 73100 Lecce, Italy. Phone: (39) 0832 320856. Fax: (39) 0832 320626. E-mail: [email protected]. 7425 on May 30, 2016 by guest http://iai.asm.org/ Downloaded from
-
Upload
unisalento -
Category
Documents
-
view
1 -
download
0
Transcript of Identification, Characterization, and Variable Expression of a Naturally Occurring Inhibitor Protein...

INFECTION AND IMMUNITY,0019-9567/01/$04.00�0 DOI: 10.1128/IAI.69.12.7425–7436.2001
Dec. 2001, p. 7425–7436 Vol. 69, No. 12
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Identification, Characterization, and Variable Expression of aNaturally Occurring Inhibitor Protein of IS1106 Transposase in
Clinical Isolates of Neisseria meningitidisPAOLA SALVATORE,1,2 CATERINA PAGLIARULO,1 ROBERTA COLICCHIO,1 PATRIZIA ZECCA,3
GIUSEPPINA CANTALUPO,1 MAURIZIO TREDICI,3 ALFREDO LAVITOLA,1 CECILIA BUCCI,1,3
CARMELO B. BRUNI,1* AND PIETRO ALIFANO3*
Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano,” Universita di Napoli “Federico II,” andCentro di Endocrinologia ed Oncologia Sperimentale “G. Salvatore” of the Consiglio Nazionale delle Ricerche, 80131
Naples,1 Dipartimento di Scienze Ambientali, Seconda Universita di Napoli, 81100 Caserta,2 and Dipartimento diScienze e Tecnologie Biologiche e Ambientali, Universita degli Studi di Lecce, 73100 Lecce,3 Italy
Received 17 July 2001/Returned for modification 6 September 2001/Accepted 19 September 2001
Transposition plays a role in the epidemiology and pathogenesis of Neisseria meningitidis. Insertion se-quences are involved in reversible capsulation and insertional inactivation of virulence genes encoding outermembrane proteins. In this study, we have investigated and identified one way in which transposon IS1106controls its own activity. We have characterized a naturally occurring protein (Tip) that inhibits the trans-posase. The inhibitor protein is a truncated version of the IS1106 transposase lacking the NH2-terminal DNAbinding sequence, and it regulates transposition by competing with the transposase for binding to the outsideends of IS1106, as shown by gel shift and in vitro transposition assays. IS1106Tip mRNA is variably expressedamong serogroup B meningococcal clinical isolates, and it is absent in most collection strains belonging tohypervirulent lineages.
Many studies have pointed out the importance of mobilegenetic elements in microbial pathogenesis and adaptation tochanging environmental conditions. Virulence genes of patho-genic bacteria, which code for toxins, adhesins, invasins, cap-sules, pili, resistance determinants, or other virulence factors,may be located on transmissible genetic elements such astransposons, plasmids, or bacteriophages (16, 50). Insertionsequence (IS) elements are known to be involved in microevo-lution of bacterial genomes by several mechanisms (1, 32). (i)In the chromosomes of both gram-negative and gram-positivebacteria, virulence genes are often clustered in “virulenceblocks” or “pathogenicity islands,” surrounded by IS elementsthat promote their transposition and lead to changes in viru-lence in the course of evolution. Pathogenicity islands areinvariably found in pathogenic strains of a given species but areeither absent or rarely present in nonpathogenic variants of thesame species (12, 20, 21, 23, 30, 33, 34). (ii) In addition, twocopies of certain IS elements flanking a DNA segment are ableto act in concert, mobilizing the intervening region. (iii) Pro-grammed insertion and excision of IS elements and of invert-ible DNA sequences may control the expression of severalvirulence factors by a mechanism of phase variation (24, 58).(iv) “Jumping” of DNA sequences (transposition) and subse-
quent recombination events may also cause gene activation (5)and antigenic variation of virulence factors, leading to theemergence of novel pathogenic variants (48). (v) IS-relatedDNA rearrangements do occur in resting bacterial culturesand confer plasticity on the genome under conditions of nu-tritional deprivation, thereby playing an adaptive role (1, 3, 18,35, 36).
With the development of studies of the mechanisms of bac-terial pathogenesis and the advent of whole-genome sequenc-ing technologies in recent years, the finding of associationbetween IS elements and pathogenic and virulence functionshas become increasingly evident. Such associations have beenobserved in a variety of animal pathogens (6, 9, 14, 17, 31, 52).
Whole-genome sequence analysis and subtractive hybridiza-tion procedures have led to the identification of putative is-lands of horizontally transferred DNA into the genomes ofserogroup B and serogroup A meningococci (42, 55). Severalof these regions encode proteins that are specific to the patho-genic Neisseria species and may have a role in virulence. Theseregions do not have the classical characteristics of pathogenic-ity islands. Several, however, have a particularly low G�Ccontent and are associated with transposase and integrasegenes, suggesting that at some time in the genetic history ofthese species, the regions were the results of recombinationevents with DNAs from other species. One of these regions ina serogroup A strain, characterized by a significantly low G�Ccontent and containing open reading frames (ORFs) with nohomology to genes in databases, is flanked by several copies ofIS1106 and a copy of IS50 (42).
Transposition plays a role in the epidemiology and patho-genesis of Neisseria meningitidis. IS1301 is involved in revers-ible capsulation by insertion into and excision from the siaAgene locus in serogroup B meningococci (24). This transpos-
* Corresponding author. Mailing address for C. B. Bruni: Diparti-mento di Biologia e Patologia Cellulare e Molecolare “L. Califano,”Universita di Napoli “Federico II,” and Centro di Endocrinologia edOncologia Sperimentale “G. Salvatore” of the Consiglio Nazionaledelle Ricerche, Via S. Pansini 5, 80131 Naples, Italy. Phone: (39) 0817462047. Fax: (39) 081 7703285. E-mail: [email protected]. Mailingaddress for P. Alifano: Dipartimento di Scienze e Tecnologie Bio-logiche e Ambientali, Universita degli Studi di Lecce, Via Monteroni,73100 Lecce, Italy. Phone: (39) 0832 320856. Fax: (39) 0832 320626.E-mail: [email protected].
7425
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

able element is also responsible for insertional inactivation ofthe porA gene encoding the class 1 outer membrane protein,which is considered to function as a porin and invasin (57) inseveral serogroup B and C meningococcal isolates (38). Inaddition, analysis of the nucleotide sequence of the chromo-somal region downstream of the porA gene has revealed thepresence of a rearranged copy of the IS1106 element in carrierstrains/isolates but not in invasive meningococcal strains/iso-lates of serogroup B, type 15, subtype 16 (B15:P1.16) (26).
IS1106 is an IS present in multiple copies in all of themeningococcal strains so far examined (39, 40) belonging tothe IS5 group of the IS4 family of transposable elements (46,26, 32). It was the first IS to be characterized in N. meningitidisand has been used as a DNA probe in phylogenetic and epi-demiological analyses (25, 39) and to develop rapid, specific,and sensitive PCR-based tests for the diagnosis of meningo-coccal disease (40).
In this study, we have investigated the regulation of trans-posase activity of the element IS1106 in clinical isolates of N.meningitidis and characterized a naturally occurring inhibitorprotein (IS1106Tip) of the transposase of IS1106 (IS1106T) byusing biochemical and genetic approaches.
MATERIALS AND METHODS
Bacterial strains and growth conditions. The meningococcal strains used inthis study are listed in Table 1. Invasive strains/isolates were derived from acollection of strains isolated during outbreaks of epidemic disease that haveoccurred in different places in Italy and France during the last 20 years. Theserotypes and subserotypes of these strains are shown in Table 1. A total of 24carrier strains/isolates were enrolled in this study. These strains were sampledfrom the nasopharynges of different healthy subjects at the time of their militaryenlistment in the course of a routine screening program for the surveillance ofmeningococcal disease. The carrier strains/isolates used in this study were sam-pled in different geographical areas in Italy and France. Two strains, BL9513 andBF9513, were isolated, respectively, from the cerebrospinal fluid and the naso-pharynx of a single sick subject in France. All meningococcal strains were cul-tured on chocolate agar (Becton-Dickinson) or on GC agar or broth (Difco)supplemented with 1% (vol/vol) Polyvitox (Bio-Merieux) at 37°C in 5% CO2.Antibiotics were purchased from Sigma and used at the following concentrations:erythromycin, 7 �g ml�1; rifampin, 36 �g ml�1.
Escherichia coli strain DH5� [F� �80d lacZ�M15 endA1 recA1 hsdR17supE44 thi-1 �� gyrA96 �(lacZYA-argF) U169] was used in cloning procedures.Strain BL21 �DE3 (F� ompT rB
� mB�) was used to overexpress recombinant
proteins (53). E. coli strains were grown in Luria-Bertani broth/agar. Whenneeded, ampicillin was added to a final concentration of 50 �g ml�1.
Transformation of meningococci. Transformations were performed as previ-ously described (19) by using 500 ng of chromosomal DNA extracted fromrifampin-resistant derivatives of strains BL847 and BF52 (Table 1). A recipientstrain was BF52. Transformants were selected on GC agar base supplementedwith rifampin (36 �g ml�1) or erythromycin (7 �g ml�1).
Plasmids and cloning procedures. To obtain plasmid pUCIS1106::ermC�, a1,245-bp-long PCR-derived EcoRV fragment spanning an entire IS1106 elementand flanking direct repeats (GGTC) was cloned into the HincII site of pUC19.The oligonucleotides used to amplify the IS1106 DNA sequence were 5�-TAAGATATCGTCGACGGTCGAGACCTTTGCAAAATTCCCCAAAATC-3� and5�-TTAGATATCGTCGACGACCGAGACCTTTGCAAAATTCCTTTCCCTC-3�(the EcoRV sites are underlined). The template DNA was derived fromstrain BL859. The resulting plasmid, pUC1106, was linearized at the unique ClaIsite within the IS1106T gene, and a 1,573-bp ClaI-AccI fragment containing theermC� gene was inserted.
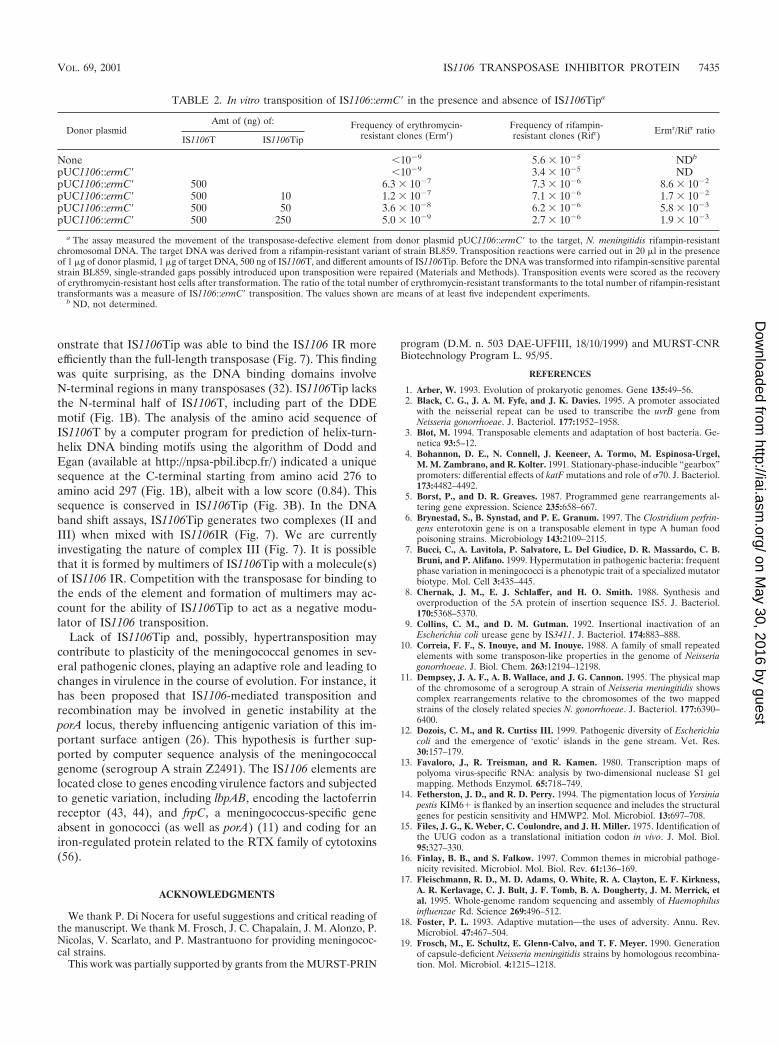
To construct plasmids pET1106T and pET1106Tip, genomic regions contain-ing the entire or 5�-end-truncated transposase were amplified, respectively, byusing oligonucleotides 5�-TTAGGGGATTTCATATGAGCACCTTCTTCCGGCAAACCGC-3� and 5�-CTTTCCCGGATCCCAGCCGAAACCCAAACACAGG-3� (for IS1106T) or 5�-CCTTGTCCTGACATATGTTAATCCACTATACCTCCGCCAATG-3� and 5�-CTTTCCCGGATCCCAGCCGAAACCCAAACACAGG-3� (for IS1106Tip) (the underlined sequences are CATATG for NdeI
sites and GGATCC for BamHI sites). The template DNAs were derived fromstrain BL859 (for IS1106T) and BF18 (for IS1106Tip). The PCR products of1,128 and 518 bp, respectively, were restricted with NdeI and BamHI and clonedinto the NdeI-BamHI sites of pET15b (provided by Novagen). pT71106Tip wasobtained by cloning the 518-bp PCR product into the NdeI-BamHI sites of pT7-7(54).
Plasmid pUH1106Tip was constructed by cloning a 540-bp-long, PCR-derivedBamHI-XbaI fragment spanning the IS1106Tip gene into the polylinker of plas-mid pUH1I (47). The oligonucleotides used to amplify the IS1106 DNA se-quence were 5�-TAAGATGGATCCCTGCGGCTTCGTCGCCTTGTC-3� and5�-CTTTCCCTCTAGACAGCCGAAACCCAAACACAGG-3� (the underlinedsequences are GGATCC for BamHI sites and TCTAGA for XbaI sites). Thetemplate DNA was derived from strain BF18.
The genomic region encompassing the truncated IS1106 element shown in Fig.3 was amplified from meningococcal strains by using oligonucleotides 5�-ATGGACGAAATCGAGGCAGCCG-3� and 5�-TTCCCGCGAACGCGGGAATC-3�as primers.
DNA procedures. High-molecular-weight genomic DNAs from the different N.meningitidis strains were prepared as previously described (7). DNA fragmentswere isolated through acrylamide slab gels and recovered by electroelution aspreviously described (49).
The IS1106T-specific probe used in the Northern blot and S1 nuclease map-ping experiments shown in Fig. 2 was obtained by PCR using the genomic DNAderived from strain BS849 as the template. The oligonucleotides used as primers(5�-ATGAGCACCTTCTTCCGGCAAACCGC-3� and 5�-AGACAGCCGAAACCCAAACACAGG-3�) were designed to amplify a region of 1,126 bp on thebasis of the nucleotide sequence of IS1106 shown in Fig. 1 (from nucleotide 68to nucleotide 1194). The IS1106-specific probe used in the S1 mapping experi-ment shown in Fig. 5 was obtained with oligonucleotides 5�-ACAATGATGATTTCTTTGAACTGATGCGCG-3� and 5�-ACATCGCCTTCAGGTGGCTTTGCGCACTCAC-3� (from nucleotide 456 to nucleotide 482 of Fig. 4B) andgenomic DNA derived from strain BF18 used as a template. The amplificationreactions consisted of 30 cycles including 1 min of denaturation at 94°C, 1 min ofannealing at 55°C, and 1 to 2 min of extension at 72°C. They were carried out ina Perkin-Elmer Cetus DNA Thermal Cycler 480. 5�-end labeling was performedwith the T4 polynucleotide kinase and [-32P]ATP (3,000 Ci mmol�1). In North-ern blot and S1 mapping experiments, the probes were labeled only at the strandcomplementary to the expected transcripts.
The nucleotide sequence of the wild-type IS1106 element (Fig. 1A) was de-termined by sequencing of a 1,227-bp PCR product obtained by amplifying agenomic region(s) of serogroup B strain BL859 (Table 1) using two oligonucle-otides corresponding to the IS1106 arms as primers. The putative arms of theelement were determined by analysis of the available genomic sequence of N.meningitidis serogroup A strain Z2491 (Sanger Centre database) using a NationalCenter for Biotechnology Information sequence similarity search tool.
The nucleotide sequence of the genomic region containing the IS1106Tip genewas determined by sequencing of PCR products obtained by using appropriateoligonucleotides as primers. The oligonucleotides were designed on the basis ofthe available sequences of N. meningitidis serogroup A strain Z2491, in which aregion containing a 5�-truncated element was identified by using a NationalCenter for Biotechnology Information sequence similarity search tool.
DNA sequencing reactions were carried out by the dideoxy-chain terminationprocedure using the T7 SequencingTM kit (Pharmacia Biotech) or the TAQencecycle sequencing kit from USB (distributed by Amersham Life Science) inaccordance with the instructions of the manufacturers.
Processing of the DNA sequences was performed with the software Gene-Jockey Sequence Processor (published and distributed by Biosoft).
Restriction fragment length polymorphism (RFLP) typing of meningococcalstrains. The genetic distances between the meningococcal isolates were deter-mined by comparing RFLP patterns in eight housekeeping genes and usingcollection strains as a reference. Assignment of the different isolates to hyper-virulent lineages was done by comparing the RFLP patterns to those of thereference strains (BL9513 [lineage 3]; 205900 [IV-1 cluster]; 93/4286 andNGP165 [ET-37 complex]; and BZ169, H44/76, and MC58 [ET-5 complex]).
Southern blotting was used to detect polymorphism generated by the presenceor absence of Sau3AI sites in the coding region of the genes recA, uvrA, uvrB,uvrC, uvrD, rep, leuS, and rho. The gene fragments were amplified from chro-mosomal DNA of strain BL859 by using PCR with the following primers: 5�-CCGAATCCTCCGGCAAAACCACCC-3� and 5�-CCGATCTTCATCCGGATTTGGTTGATG-3� (recA), 5�-GCTCGTGGTGGTAACAGGATTGTCGGG-3�and 5�-CAAAAGGCGCAGATAGTCGTGGATTTC-3� (uvrA), 5�-GAACATATCGAGCAGATGCGCCTTTCC-3� and 5�-GCTCATTAAATCGTCGACTTGGGTGGC-3� (uvrB), 5�-GCAAAGTCTTATACGTCGGCAAAGC-3� and 5�-G
7426 SALVATORE ET AL. INFECT. IMMUN.
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from
GTGTGGCTGATGTCGAAGCATTC-3� (uvrC), 5�-GTGCTGACCACGCGCATCGCATGGC-3� and 5�-GTTGGTGTCTTGGAACTCGTCAACGAG-3�(uvrD), 5�-TGCTCGTCCTTGCCGGTGCAGGCAGCG-3� and 5�-CGCGGTGGAGCGGTAGTTTTGCTCCAG-3� (rep), 5�-GAGCTGACTTTTGACGACAAAGGC-3� and 5�-TTCGTCGACTGCCGGCCAGCCAGC-3� (leuS), and 5�-TGCACGTCTCCGAATTACAAACCCTGC-3� and 5�-CACGCTTCCTTCGATGGTGTCGCC-3� (rho). The lengths of the PCR products were as follows: 400bp (recA), 248 bp (uvrA), 955 bp (uvrB), 1,129 bp (uvrC), 561 bp (uvrD), 755 bp(rep), 611 bp (leuS), and 303 bp (rho). Southern blots were performed in accor-dance with standard procedures (49). For higher resolution, a large apparatuswas used for agarose-gel electrophoresis and the DNAs from reference strainswere always included in each run. Bands of the same size were assumed to beidentical and therefore to correspond to the same allele. The genetic diversity (h)at a locus among isolates was calculated as follows: h (1 � �xi
2)(n/n � 1),where xi is the frequency of the ith allele and n is the number of isolates. The hvalues for the loci were: 0.00 (recA), 0.83 (uvrA), 0.70 (uvrB), 0.82 (uvrC), 0.73(uvrD), 0.64 (rep), 0.39 (leuS), and 0.66 (rho). The mean genetic diversity perlocus was the arithmetic average of h values over all of the loci. The geneticdistance (D) between pairs of isolates was calculated as the proportion of loci atwhich dissimilar alleles occurred, and a dendrogram was constructed from amatrix of allelic mismatches by the pair group cluster method with arithmeticaverages. Data were normalized by a weighted coefficient, with the contributionof each locus to D being weighted by the reciprocal of the mean genetic diversityat the locus in the total sample being analyzed (37, 51).
RNA procedures. Total bacterial RNA was extracted from logarithmicallygrowing cells by the guanidine hydrochloride procedure previously described (7).Electrophoretic analysis was done by fractionating the total RNA on 1% agarosegels containing formaldehyde (49). RNA transfer to Hybond (Amersham) mem-branes and hybridization with 32P-labeled fragments were done in accordancewith the standard procedure (49).
RNA-DNA hybridization, S1 nuclease digestion, and analysis of the hybrids ondenaturing polyacrylamide gels were performed as described by Favaloro et al.(13). Quantitative analysis of the different transcripts was performed by densi-tometry using a Scanmaster 3 (Howtek, Inc., Hudson, N.H.) or a high-perfor-mance desktop flat-bed color scanner equipped with the RFLPrint (Pdi, Hunt-ington Station, N.Y.) software package or by directly counting the radioactivebands with a PhosphorImager SI (Molecular Dynamics, Inc., Sunnyvale, Calif.).
In vitro translation assay. In vitro transcription-translation of recombinantplasmids was obtained in an S30 extract prepared from N. meningitidis in thebuffer system described by Zubay (59) by making use of [35S]methionine toobtain labeled gene products. Proteins were analyzed on sodium dodecyl sulfate(SDS)–15% polyacrylamide gels.
In vitro transposition assay. The in vitro transposition assay measured themovement of the transposase-defective element from donor plasmidpUC1106::ermC� to target chromosomal DNA. The target DNA was derivedfrom a rifampin-resistant variant of N. meningitidis strain BL859. Transpositionreactions were carried out in 10% glycerol–2 mM dithiothreitol–250 �g of bovineserum albumin ml�1–25 mM HEPES (pH 7.9)–100 mM NaCl–10 mM MgCl2.They contained, in a final volume of 20 �l, 1 �g of donor plasmid, 1 �g of targetchromosomal DNA, 500 ng of IS1106T, and different amounts of IS1106Tip.Samples were incubated at 30°C for 3 h and then exposed to 75°C for 10 min toinactivate the enzymes. Before the DNA was transformed into rifampin-sensitiveparental strain BL859, single-stranded gaps possibly introduced upon transposi-tion were repaired. Gaps in the DNA were first filled with the Klenow fragmentof E. coli DNA polymerase I by using the same buffer system and each de-oxynucleoside triphosphate at 1 mM. The enzyme was heat inactivated by 10 minof exposure to a temperature of 75°C. The sample volumes were then raised to40 �l and 5 U of T4 DNA ligase and ATP to a final concentration of 1 mM wereadded. The ligation reactions were performed for 3 h at room temperature. Halfof the repaired transposition products were used to transform BL859. Transpo-sition events were scored as the recovery of erythromycin-resistant host cellsafter transformation. The ratio of the total number of erythromycin-resistanttransformants to the total number of rifampin-resistant transformants was a
TABLE 1. Meningococcal strains used in this study and theircharacteristics
Strain Serogroup and/or serotypea Lineageb IS1106Tip
mRNAcClinical
specimen(s)d Sourcee
BF2 B ET-37 � NP iBF3 B ET-37 � NP iBF9 B Other � NP iBF13 B Other � NP iBL847 B:14:P1.12 Lineage 3 � CSF iiBL851 B:4:P1.13 Lineage 3 � CSF iiBL857 B:21:P1.7,16 Other � CSF iiBL858 B:15,21:P1.15 Other � CSF iiBL859 B:4:P1.13 Lineage 3 � CSF iiBS843 B:4:P1.13 Lineage 3 � BL iiBS845 B:4:P1.13 Lineage 3 � BL iiBS849 B:4:P1.13 Lineage 3 � BL iiBL855 B ET-37 � CSF iiB1940 B:NT:P1.3,6,15 Other � CSF iiiBL9513 B:4:P1.4 Lineage 3 � CSF, BL ivBL892 B:4:P1.4 Lineage 3 � CSF ivBL942 B:1:NST Lineage 3 � CSF ivBL911 B:NT:P1.9 Other � CSF ivBL915 B:NT:P1.5 Other � CSF ivBL951 B:NT:P1.1 Lineage 3 � CSF ivBL899 B:4:P1.2,5 ET-5 � CSF ivBL932 B:4:P1.1 Other � CSF ivBL937 B:4:NST ET-5 � CSF ivBL947 B:1:NST Other � CSF ivBL897 B Other � CSF ivBF10 B Other � NP vBF17 B Other � NP vBF18 B Other � NP vBF21 B Other � NP vBF65 B Other � NP vBF16 B Other � NP vBF23 B ET-5 � NP vBF40 B ET-37 � NP vBF52 B ET-37 � NP vBF8960 B Other � NP viBF8961 B Other � NP viBF8964 B Other � NP viBF8969 B Other � NP viBF9216 B Other � NP viBF5425 B Other � NP viBF32B B Lineage 3 NA NP viiBF37B B Other NA NP viiBF43B B Other NA NP viiBF6L B Other NA NP viiBF8L B Other NA NP viiBF57L B Other NA NP vii205900 A IV-1 � CSF, BL viii93/4286 C ET-37 � CSF, BL viiiNGP165 B ET-37 � CSF, BL viiiBZ169 B ET-5 � CSF, BL viiiH44/76 B ET-5 � CSF, BL viiiMC58 B ET-5 � CSF, BL viiiNGF26 B Other � CSF, BL viii1000 B Other � CSF, BL viiiNGE31 B Other � CSF, BL viiiNGH15 B Other � CSF, BL viiiZF15 Z Other � NP vXL929 X Other � CSF, BL ivXF47A X Other NA NP vYL896 Y Other � CSF ivCF5C C Other � NP v
a NST, not serotypeable.b Assignment of isolates to hypervirulent lineages was done as detailed in
Materials and Methods. Other, not belonging to lineage 3, ET-5, or ET-37complex, IV-1 cluster.
c IS1106Tip mRNAs were detected by S1 nuclease protection experiments. �,presence of specific transcripts; �, absence of detectable transcripts; �, barelydetectable transcripts; NA, not analyzed.
d NP, nasopharynx; CSF, cerebrospinal fluid; BL, blood.e i, II Policlinico, Universita di Napoli, Naples, Italy; ii, Istituto Superiore di
Sanita, Rome, Italy; iii, Bayerische Julius-Maximilians Universitat, Wurzburg,Germany; iv, Institut Pasteur, Paris, France; v, Hopital d’Instruction des Armee,Brest Naval, France; vi, Institut de Medecine Tropicale du Service de Sante desArmees, Marseille Armees, France; vii, Laboratory of Microbiology, Universitadi Lecce, Lecce, Italy; viii, IRIS, Chiron S.p.A, Siena, Italy.
VOL. 69, 2001 IS1106 TRANSPOSASE INHIBITOR PROTEIN 7427
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

measure of IS1106::ermC� transposition. Each value is the mean of at least fiveindependent experiments. The variation within one set of assays was usually lessthan twofold.
Cell extract preparation and protein purification. Crude (S30) extracts wereprepared from E. coli BL21 �DE3 cells transfected with plasmid pET15b,pET1106T, pET1106Tip, pT7-7, or pT71106Tip grown to early logarithmic phaseand induced with isopropyl- -D-thiogalactopyranoside (IPTG) for 2 h at 37°C ornot induced. Cells were mechanically broken with a French press in a buffercontaining 20 mM HEPES (pH 8.2), 100 mM KCl, 0.2 mM EDTA, 0.5 mMphenylmethylsulfonyl fluoride, 0.5 mM dithiothreitol, and 20% glycerol. Induc-tion of E. coli cells transfected with pT71106Tip resulted in the appearance ofpolypeptides with an apparent molecular mass of 16,000 Da as determinedby SDS-polyacrylamide gel electrophoresis. Histidine-tagged IS1106T and
IS1106Tip were partially purified by the rapid affinity purification protocol inaccordance with the Novagen pET system manual.
DNA gel mobility shift assay. The double-stranded probe used in the gelmobility shift experiments was obtained by annealing the complementary 5�-end-labeled oligonucleotides 5�-GGTCGAGACCTTTGCAAAATTCC-3� and 5�-GGAATTTTGCAAAGGTCTCGACC-3� spanning the 19-bp sequence of the leftarm of IS1106. The annealing reaction was carried out at 65°C for 1 h.
The gel mobility shift assay was performed by mixing (at 24°C) purified pro-teins, double-stranded labeled probe, unlabeled competitor DNA, and buffer ina total volume of 20 �l. The final buffer contained 20 mM HEPES, 40 mM KCl,4% Ficoll, 5 mM spermidine, 0.25 �g of poly(dI-dC) �l�1. The protein concen-trations ranged between 2.5 and 25 ng �l�1, and the concentration of the probewas 1 ng �l�1 (20,000 cpm). Unlabeled competitor was added in 5- to 50-fold
FIG. 1. Structure and nucleotide sequence of N. meningitidis IS1106. (A) Nucleotide sequence of IS1106 and deduced amino acid sequence ofthe putative transposase (IS1106T) gene. IS1106 has a length of 1,207 bp and is flanked by 19-bp-long IRs (IRL and IRR, arrows). Four-base-pairdirect repeats (GGTC) border the ends of the IS. Amino acids corresponding to the IS1106T ORF are indicated in uppercase letters below thenucleotide sequence. Several control elements are also shown: (i) a14-bp-long perfect IR located 2 bp downstream from IRL (arrows), (ii) arepeated motif resembling part of the core sequence of the neisserial RS3 repeat (underlined sequences), and (iii) putative �10 and �35 promoterand Shine-Dalgarno (SD) sequences (asterisks). The nucleotide sequence downstream from the black arrowhead is also part of the rearrangedelement previously described (26). The sequence downstream from the open arrowhead is shared by IS1106Tip (Fig. 3B). (B) The deduced aminoacid sequence of the putative IS1106T of N. meningitidis (N.m.) is aligned with the amino acid sequence of the 5A transposase protein of E. coli(E.c.) IS5. Plus signs indicate synonymous substitutions. The sequence downstream from the open arrowhead is shared by IS1106Tip (Fig. 3B). Theoverlined amino acids grouped in the N2, N3, and C1 regions are part of the DDE motif, the amino acid triad intimately involved in catalysis (32).
7428 SALVATORE ET AL. INFECT. IMMUN.
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

excess over the labeled DNA. The mixture was incubated for 15 min at 24°Cbefore loading onto a 5% polyacrylamide gel in 0.25� standard TBE buffer (1�TBE is 0.0089 M Tris-borate, 0.089 M boric acid, and 0.002 M EDTA), andelectrophoresis was carried out at 4°C.
RESULTS
Structure and nucleotide sequence of the N. meningitidisIS1106 element and analysis of transposase-specific tran-scripts in meningococcal strains. The IS1106 element locatedwithin a complex repetitive region downstream of porA is arearranged element in which a transposon-like repetitive ele-ment (also known as a small repetitive element [SRE]) (10)interrupts the region encoding the amino terminus of the pu-tative IS1106T protein (26). By PCR using two oligonucleo-tides complementary to the IS1106 inverted-repeat sequences(IRs), we isolated a wild-type element from serogroup B strainBL859 (Fig. 1A). The element has a length of 1,207 bp, isflanked by 19-bp-long IRs (a left IR [IRL] and a right IR[IRR]), and encodes a putative 38,505-Da peptide showingextensive homology to the 5A transposase protein of IS5 (27,8) (Fig. 1B). Analysis of the DNA sequence revealed the pres-ence of several putative control elements upstream of the startcodon of the transposase: (i) a 14-bp-long perfect IR located 2bp downstream from IRL, (ii) a repeated motif resemblingpart of the core sequence of the neisserial RS3 repeat (22), and(iii) putative �10 and �35 promoter sequences (Fig. 1A).
We next analyzed transposon-specific transcripts in differentserogroup B meningococcal strains by Northern blotting. TotalRNAs extracted from either “carrier strain” or “invasivestrain” isolates were probed with an IS1106T-specific probe. Aspecific transcript of about 1,200 nucleotides was detected onlyin carrier strain isolates (Fig. 2A, lanes 1 to 4) and not ininvasive strain isolates (Fig. 2A, lanes 5 to 8). We performed anS1 mapping experiment to define the ends of this transcript(Fig. 2B). After treatment with S1 nuclease, the amounts offull-length protected hybrids were very low in all of the strainstested and detectable only after overexposure of the autora-diogram. On the contrary, a shorter hybrid was present inconsiderable amounts only in carrier strains/isolates BF10 andBF18 (Fig. 2B, lanes 3 and 4). The 5� end corresponded tonucleotide 700 of the IS1106 sequence (Fig. 1A).
Isolation of a transcriptionally active, 5�-end-truncatedIS1106 element from the genome of a carrier strain/isolate.The transcript mapping data led us to hypothesize the exis-tence of a transcriptionally active, rearranged version of theIS1106 element in the meningococcal genome. The sequenceof the IS1106 element in the genome of serogroup B carrierstrain/isolate BF18 was determined (Fig. 3). Inspection of thenucleotide sequence revealed the presence of a rearrangedIS1106 element located downstream of an IS1016-like element(32) (Fig. 3A). Alignment of the rearranged IS1106 sequencewith that of the wild type (Fig. 1A) revealed that the elementis truncated at the 5� end and located immediately downstreamfrom a neisserial SRE (Fig. 3B).
The putative start site(s) of the 5�-end-truncated IS1106-specific transcript was determined by S1 nuclease mapping(Fig. 4A). The analysis revealed two major transcripts whose 5�ends mapped within the SRE located upstream of the rear-ranged IS1106 element (Fig. 3B). These transcripts could bedetected only in carrier strains/isolates BF10 and BF18 (Fig.
FIG. 2. Analysis of the IS1106-specific transcript. (A) Northernblot analysis. Total RNAs (10 �g) from carrier strains/isolates BF10,BF18, BF21, and BF8961 (lanes 1 to 4) and from invasive strains/isolates BL858, BS849, BL892, and BL947 (lanes 5 to-8) were hybrid-ized to a 5�-end-labeled 1,126-bp DNA fragment spanning the entirecoding region of IS1106T (Fig. 1A and Materials and Methods). Onlythe strand complementary to the RNA was labeled. Arrows indicatethe IS1106-specific transcript and its approximate size deduced on thebasis of the relative migration of the rRNAs. nt, nucleotides. (B) S1nuclease mapping analysis. A 5�-end-labeled DNA fragment spanningthe entire coding region of IS1106T (Fig. 1A) was used as a probe. Theprobe (lane 8) was hybridized to total RNA (10 �g) extracted fromcarrier strains/isolates BF10 and BF18 (lanes 3 and 4), from invasivestrains/isolates BL858 and BS849 (panel B, lanes 5 and 6), or fromyeast (lane 7). After treatment with S1 nuclease, the reaction productswere resolved on a 6% polyacrylamide-urea denaturing gel. The sizesof the protected hybrids (arrows) were determined by running in par-allel a sequencing reaction ladder (G and A, lanes 1 and 2) of the sameDNA fragment used as a probe. The bar indicates the relative migra-tion of the probe.
VOL. 69, 2001 IS1106 TRANSPOSASE INHIBITOR PROTEIN 7429
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

4A, lanes 5 and 6) and not in invasive strains/isolates BL858and BS849 (Fig. 4A, lanes 3 and 4). The absence of the IS1106-specific transcripts in invasive strains/isolates BL858 andBS849 does not depend on a lack of the element because PCRanalysis revealed the presence of the truncated IS1106 element(data not shown). We therefore speculated that transcriptionof the 5�-truncated IS1106 element could be impaired by apoint mutation(s). Analysis of the nucleotide sequence ofstrain BS849 revealed four nucleotide substitutions in the pu-tative promoter region in both strains, mapping 12, 24, 82, and86 nucleotides upstream of the longer transcription start site,respectively (Fig. 3B).
In the rearranged element, an ORF initiates at the rarecodon for leucine TTG (15) (nucleotide 140), possibly encod-ing a 14,870-Da peptide corresponding to the carboxy-terminalhalf of the putative wild-type IS1106T . This ORF is foundboth in the carrier strains/isolates and in the invasive strains/isolates (data not shown). To seek evidence that the 5�-end-truncated IS1106-specific transcripts were translatable, we per-formed an in vitro translation assay. To this end, the 5�-end-truncated transposase gene from strain BF18 was cloneddownstream from the E. coli his promoter to obtain plasmidpUH1106Tip. The recombinant and the vector plasmids weretranscribed in vitro and translated by using an S30 extractderived from meningococcal strain BF18 (Fig. 4B). In additionto vector-encoded peptides, a specific translation product ofabout 16,000 Da was produced by pUH1106Tip (Fig. 4B, lane2). The size of this peptide was close to the expected molecular
mass (14,870 Da) of the 5�-end-truncated transposase(IS1106Tip).
Analysis of ISII06Tip mRNA and distribution of IS1106 inclinical isolates of N. meningitidis. We next investigated thepresence of the IS1106Tip mRNA in clinical isolates of me-ningococci by S1 mapping. The meningococcal strains wereisolated in different regions of Italy and France over the last 10years. The genetic relationships among 50 clinical isolates and10 reference strains (45) were determined by comparing RFLPpatterns in eight housekeeping genes that were mapped on aphysical map (55) to ensure that they were unlinked. Therelatedness between strains is shown as a dendrogram (Fig. 5).The results of the S1 mapping analysis, summarized in Table 1,indicated that the 5�-end-truncated form of the IS1106TmRNA was produced in 14 (67%) of 21 meningococcal isolatessampled from the oropharynges of healthy subjects. In con-trast, the transcript was detected in only 6 (18%) of 33 isolatesderived from the blood or cerebrospinal fluid of patients withmeningococcal disease or in strains belonging to several hy-pervirulent lineages, such as lineage 3, the ET-5 complex, andcluster IV-1. However, it was detected in strains belonging tothe ET-37 complex derived either from patients or fromhealthy subjects.
In most cases, lack of IS1106Tip expression was associatedwith the same point mutations previously mapped (Fig. 3A).However, in strains of the ET-5 complex, the IS1106Tip genecould not be amplified by PCR, suggesting that a rearrange-ment or deletion had occurred (data not shown).
FIG. 3. Nucleotide sequence, deduced amino acid sequence, and structural features of the genomic region containing the transcriptionallyactive IS1106Tip gene. (A) Physical and genetic map of the genomic region containing the IS1106Tip gene. The IS1106Tip gene is locateddownstream of an IS1016-like element, which is preceded by the rho gene. The element is truncated at the 5� end and is located immediatelydownstream from a neisserial SRE. Two copies of neisserial repetitive sequence RS3 are arranged in tandem downstream from the truncatedelement. The bent arrow indicates the transcription start point of the IS1106Tip mRNA. (B) Nucleotide and deduced amino acid sequences of theIS1106Tip gene. Arrows indicate IRs of the SRE (SRE-IRL and -IRR) and of IS1106 (IS1106-IRR). RS3 sequences are underlined. Bent arrowsindicate the 5� ends of the IS1106Tip mRNA. Asterisks mark the positions of a putative gearbox promoter sequence (5�-CACCAAGT-3�). Fournucleotide substitutions (indicated above the nucleotide sequence of BF18) were found in invasive strains/isolates BL859 and BL892 and map 12,24, 82, and 86 nucleotides upstream of the transcription start site. Amino acid residues that are different from the deduced sequence of IS1106T(Fig. 1A) are underlined.
7430 SALVATORE ET AL. INFECT. IMMUN.
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from
In an attempt to correlate IS1106Tip expression with IS1106transposition, both the copy number and distribution of theIS1106 elements in the meningococcal clinical isolates weredetermined by Southern blotting. In the experiment whose
results are shown in Fig. 6B, any band in the autoradiographmight be interpreted as an insertion of a single copy of eitheran intact or a truncated IS1106 element. The results demon-strate that IS1106 is present in multiple copies in the genomesof the meningococcal isolates, ranging from about 5 or 6 tomore than 15 or 16. The copy number did not correlate withthe expression of IS1106 Tip. However, in phylogeneticallyrelated strains that do not express IS1106Tip, a high hetero-geneity of the insertion pattern was observed. For instance,strains BL847 and BL859, which did not express IS1106 Tipalthough both belonging to lineage 3 (Fig. 5), appeared to beunrelated on the basis of the IS1106 transposition pattern. Asimilar heterogeneity was observed in several strains of theET-5 complex, for instance, BL899 and BL937. By contrast, allof the examined strains of the ET-37 complex that expressIS1106Tip exhibited very similar transposition patterns. Thissuggested that IS1106Tip might act as a negative modulator ofIS1106 transposition.
Effects of IS1106Tip expression on in vitro transposition ofan IS1106::ermC� element. To investigate the possibility thatIS1106Tip might function as a repressor of IS1106T, an in vitrotransposition assay was developed. Histidine-tagged IS1106Tor IS1106Tip overexpressed in E. coli BL21 �DE3 cells (Fig.7A, lanes 4 and 6) was partially purified and variously mixedwith tester DNA. A modified version of IS1106, IS1106::ermC�,was engineered by inserting the ermC� gene conferring resis-tance to erythromycin into the gene for IS1106T. The assaymeasured the movement of the transposase-defective elementfrom a donor plasmid to target rifampin-resistant N. meningi-tidis chromosomal DNA in the presence of different amountsof partially purified IS1106T and IS1106Tip. The target DNAwas used to transform a sensitive N. meningitidis strain toerythromycin or rifampin resistance. Transposition eventswere scored as the recovery of erythromycin-resistant host cellsafter natural transformation. As the in vitro treatment wasexpected to affect the transforming ability of the target DNA,values were normalized with transformation efficiencies torifampin resistance. Therefore, the ratio of the total numberof erythromycin-resistant transformants to the number of ri-fampin-resistant transformants was taken as a measure ofIS1106::ermC� transposition. The data demonstrate thatIS1106T was able to activate transposition of the transposase-defective element in trans and that transposition efficienciesstrongly decreased in the presence of IS1106Tip. The extent ofinhibition was dependent on the ratio of IS1106T to IS1106Tip(Table 2). In particular, at ratios of 50:1, 10:1, and 2:1(amounts of IS1106T to amounts of IS1106Tip), the frequen-cies of transposition events decreased about 5-, 15-, and 50-fold, respectively.
Binding of IS1106Tip to the IR of IS1106. To elucidate themechanism of inhibition of IS1106 transposition by the trun-cated transposase, we investigated the ability of IS1106Tip tointerfere with binding of the full-length transposase to IS1106termini (IS1106IR). Incubation of partially purified IS1106Twith 5�-end-labeled double-stranded oligonucleotides corre-sponding to the IS1106 IR led to the appearance of a retardedcomplex I (Fig. 7B, lane 2) specifically titrated out by excesscold probe (Fig. 7B, lanes 3 and 4). When the IS1106 IR wasincubated with IS1106Tip, two specific major DNA-proteincomplexes were detected: a faster-migrating one (complex II)
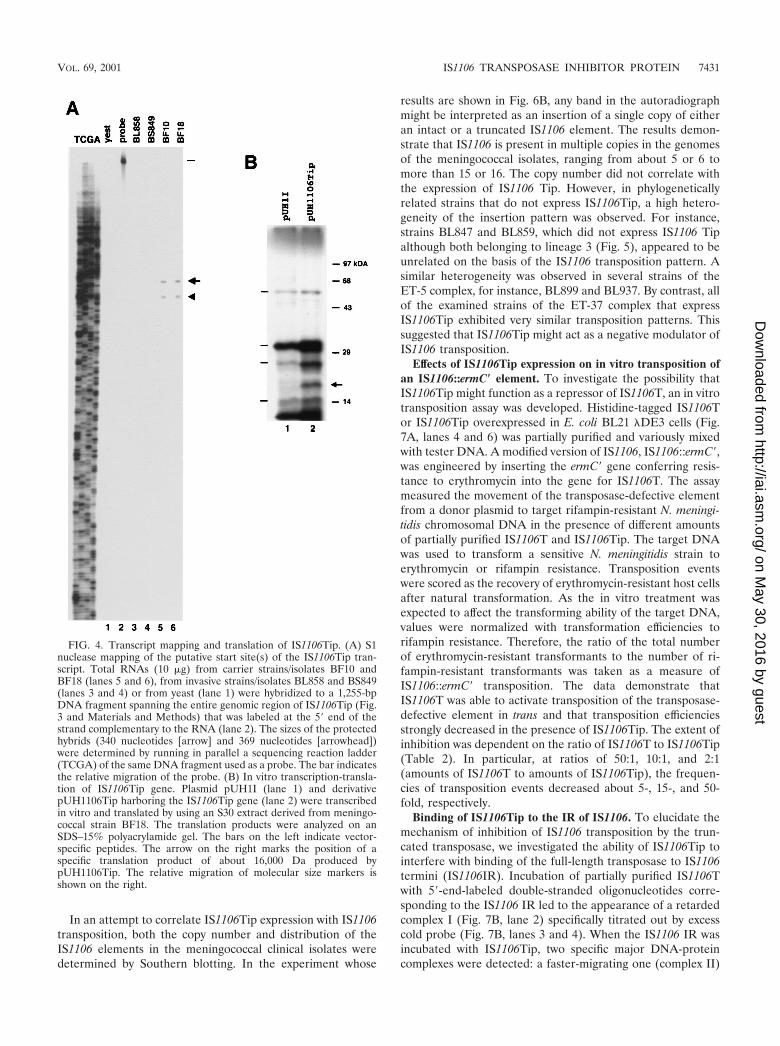
FIG. 4. Transcript mapping and translation of IS1106Tip. (A) S1nuclease mapping of the putative start site(s) of the IS1106Tip tran-script. Total RNAs (10 �g) from carrier strains/isolates BF10 andBF18 (lanes 5 and 6), from invasive strains/isolates BL858 and BS849(lanes 3 and 4) or from yeast (lane 1) were hybridized to a 1,255-bpDNA fragment spanning the entire genomic region of IS1106Tip (Fig.3 and Materials and Methods) that was labeled at the 5� end of thestrand complementary to the RNA (lane 2). The sizes of the protectedhybrids (340 nucleotides [arrow] and 369 nucleotides [arrowhead])were determined by running in parallel a sequencing reaction ladder(TCGA) of the same DNA fragment used as a probe. The bar indicatesthe relative migration of the probe. (B) In vitro transcription-transla-tion of IS1106Tip gene. Plasmid pUH1I (lane 1) and derivativepUH1106Tip harboring the IS1106Tip gene (lane 2) were transcribedin vitro and translated by using an S30 extract derived from meningo-coccal strain BF18. The translation products were analyzed on anSDS–15% polyacrylamide gel. The bars on the left indicate vector-specific peptides. The arrow on the right marks the position of aspecific translation product of about 16,000 Da produced bypUH1106Tip. The relative migration of molecular size markers isshown on the right.
VOL. 69, 2001 IS1106 TRANSPOSASE INHIBITOR PROTEIN 7431
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

and a much more abundant complex that migrated consider-ably more slowly (complex III) (Fig. 7B, lanes 5 to 7). Signif-icantly, the amount of complex III was much greater than thatof the IS1106 IR-IS1106T complex (Fig. 7B, lane 2), althoughIS1106Tip was used at a concentration 10-fold lower than thatof IS1106T. This finding indicated that the truncated trans-posase was able to bind the IS1106 IR more efficiently than wasthe full-length transposase. This result was confirmed when theprobe was incubated in the presence of both IS1106T andIS1106Tip at different relative ratios (Fig. 7B, lanes 8 to 10).Formation of complex I was substantially inhibited when
IS1106Tip and IS1106T were used at a 1:10 ratio (Fig. 7B, lane10).
DISCUSSION
Transposons have evolved various regulatory mechanismsthat limit their movement and the accompanying mutageniceffect within the host cell. Several of these mechanisms aregeneral and involve transcriptional repressors and translationalinhibitors (antisense RNA). Others are more specific and in-clude (i) sequestration of translation initiation signals, (ii) pro-
FIG. 5. Dendrogram showing the genetic relatedness between meningococcal strains. The genetic distance between the meningococcal isolateswas determined by comparing RFLP patterns in eight housekeeping genes and using collection strains as a reference as detailed in Materials andMethods.
7432 SALVATORE ET AL. INFECT. IMMUN.
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

FIG. 6. Southern blot analysis of IS1106 insertions in the genomes of meningococcal strains. (A) Physical and genetic map of an IS1106element. The positions of the ClaI and HindIII sites are indicated. The dashed bar below the map corresponds to the 322-bp-long fragment usedas a probe in the Southern blot experiment shown in panel B. (B) Total DNAs derived from the meningococcal strains indicated above the panelswere digested with EcoRI and HindIII and hybridized to the 332-bp 32P-labeled DNA fragment corresponding to the 5�-proximal one-third of theIS1106 sequence (A). The IS1106 sequence contains a unique HindIII site mapping downstream of the probe but does not contain any EcoRI site(A). Therefore, any band in the autoradiograph might be interpreted as an insertion of a single copy of either an intact or a truncated IS1106element. The probe we used minimized the detection of rearranged elements because truncation of IS1106 occurred mostly at the 5�-proximal end(data not shown). The relative migrations of molecular size markers (sizes are in base pairs) are shown beside each panel. In addition to the namesof the strains, their assignment to hypervirulent lineages is also indicated (lineage 3, ET-5, ET-37, IV-1, and other).
VOL. 69, 2001 IS1106 TRANSPOSASE INHIBITOR PROTEIN 7433
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from
grammed translational frameshifting, (iii) coupling of transla-tion termination, transposase binding and transposon activity,(iv) impinging transcription from an outside promoter and/orfrom within the element, (v) transposase stability, (vi) activityin cis of transposase (32). In this paper, we have characterizedthe wild-type IS1106 element and a novel mechanism by whichthis transposon controls its own activity.
Four copies of the wild-type element and several rearrangedcopies are found in the genome of serogroup A strain Z2491(Sanger Centre database) (41). Computer sequence analysis ofthe regions flanking the IS1106 copies in strain Z2491 did notreveal a consensus target site. However, copies of repetitivesequence RS3 are located downstream of several IS1106 ele-ments in the genome of Z2491, suggesting that the primarysequence and/or the architecture of the RS3 or RS3-like se-quence may be involved in target site selection and possibly inthe orientation of insertion. Interestingly, a repeated motifresembling part of the core sequence of the neisserial RS3repeat is located upstream to the start codon of the trans-posase gene and partially overlaps the IRL (Fig. 1A). Theanalysis of the nucleotide sequence of this region also revealedseveral putative control elements: �10 and �35 promoter el-ements and a 14-bp-long perfect IR located in the space be-tween the IRL and the start codon of the transposase gene.Because formation of this palindromic structure at the level ofRNA is predicted to sequester the ribosomal binding site, onemay speculate that the 14-bp-long IR protects IS1106 fromactivation by impinging transcription following insertion intohighly expressed genes. A similar control mechanism regulatesthe activity of other transposable elements (32).
Transcriptional mapping analysis has demonstrated thepresence, in several meningococcal strains, of an IS1106-spe-cific transcript corresponding to a 5�-end truncated transposasemRNA (Fig. 2). The sequence analysis of the genomic regionencoding the truncated transposase (IS1106Tip) revealed thepresence of a 5�-end truncated IS1106 element downstreamfrom a neisserial SRE (10) (Fig. 3). SREs have been associatedwith transposition in neisseriae. It is therefore reasonable thatthe genomic rearrangement leading to the 5�-end-truncatedtransposase gene has been promoted by insertion of the SREinto an IS1106 element. Transcription of the IS1106Tip genestarts within the SRE (Fig. 3 and 4A). Transcription fromSREs has been reported for other meningococcal genes, in-cluding uvrB (2) and drg (7). No canonical �70-dependent pro-moter consensus sequence is detectable in the SRE upstreamof the truncated transposase gene. However, a putative gear-box promoter sequence (5�-CACCAAGT-3�) is present a fewnucleotides upstream of the transcript start site (Fig. 3). Gear-box promoters appear to be involved in the regulation of sev-eral genes in E. coli that are induced upon entry into thestationary phase (4, 28, 29). In N. gonorrhoeae, putative gear-box sequences have been identified within the SRE upstreamof uvrB and 7 of the 11 opa genes from strain MS11 (2).IS1106Tip mRNA is variably expressed among meningococcalclinical isolates (Table 1). The absence of IS1106Tip-specifictranscripts is associated either with mutations in the putativepromoter region located within the SRE or with rearrange-ment/deletion of the gene (in strains of the ET-5 complex).
The results of the in vitro transposition assay demonstratedthat IS1106Tip might act as a negative modulator of IS1106transposition (Table 2). Incomplete transposase peptides con-tribute to repression of transposition by different mechanisms:(i) binding to an IR, leading to either repression of the trans-posase pIRL promoter or competition with the transposase forbinding to the ends of the element, and (ii) generation ofnonproductive heteromultimers with full-length transposasepeptides (32). The results of the DNA band shift assays dem-
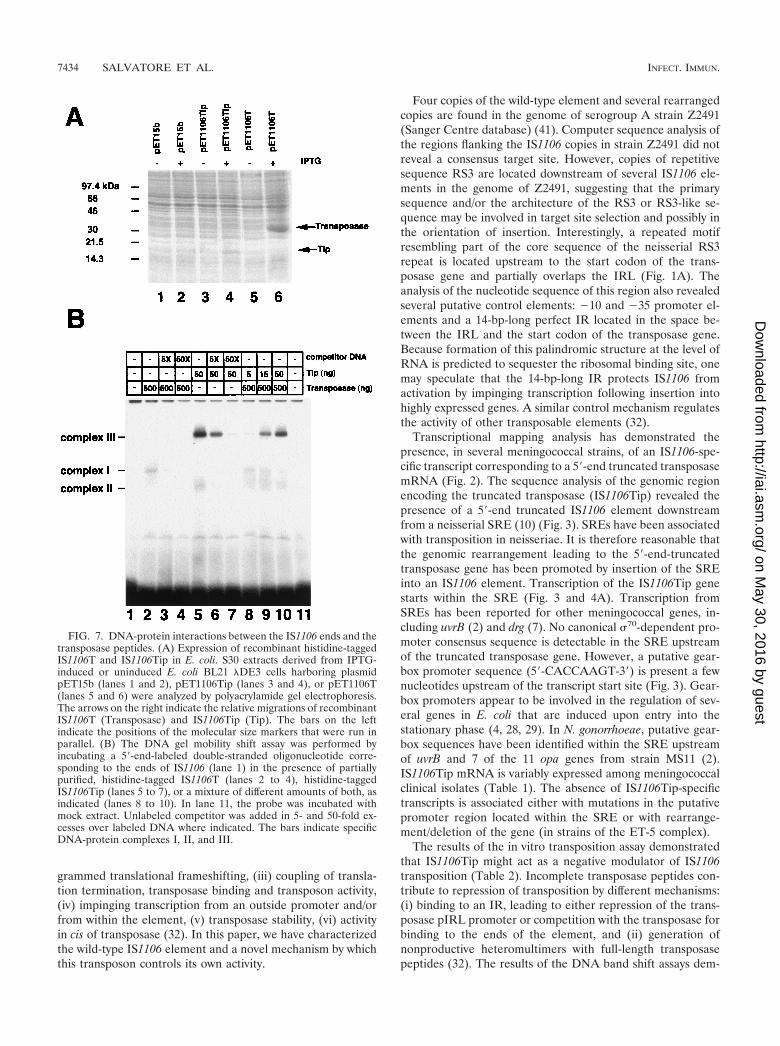
FIG. 7. DNA-protein interactions between the IS1106 ends and thetransposase peptides. (A) Expression of recombinant histidine-taggedIS1106T and IS1106Tip in E. coli. S30 extracts derived from IPTG-induced or uninduced E. coli BL21 �DE3 cells harboring plasmidpET15b (lanes 1 and 2), pET1106Tip (lanes 3 and 4), or pET1106T(lanes 5 and 6) were analyzed by polyacrylamide gel electrophoresis.The arrows on the right indicate the relative migrations of recombinantIS1106T (Transposase) and IS1106Tip (Tip). The bars on the leftindicate the positions of the molecular size markers that were run inparallel. (B) The DNA gel mobility shift assay was performed byincubating a 5�-end-labeled double-stranded oligonucleotide corre-sponding to the ends of IS1106 (lane 1) in the presence of partiallypurified, histidine-tagged IS1106T (lanes 2 to 4), histidine-taggedIS1106Tip (lanes 5 to 7), or a mixture of different amounts of both, asindicated (lanes 8 to 10). In lane 11, the probe was incubated withmock extract. Unlabeled competitor was added in 5- and 50-fold ex-cesses over labeled DNA where indicated. The bars indicate specificDNA-protein complexes I, II, and III.
7434 SALVATORE ET AL. INFECT. IMMUN.
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from
onstrate that IS1106Tip was able to bind the IS1106 IR moreefficiently than the full-length transposase (Fig. 7). This findingwas quite surprising, as the DNA binding domains involveN-terminal regions in many transposases (32). IS1106Tip lacksthe N-terminal half of IS1106T, including part of the DDEmotif (Fig. 1B). The analysis of the amino acid sequence ofIS1106T by a computer program for prediction of helix-turn-helix DNA binding motifs using the algorithm of Dodd andEgan (available at http://npsa-pbil.ibcp.fr/) indicated a uniquesequence at the C-terminal starting from amino acid 276 toamino acid 297 (Fig. 1B), albeit with a low score (0.84). Thissequence is conserved in IS1106Tip (Fig. 3B). In the DNAband shift assays, IS1106Tip generates two complexes (II andIII) when mixed with IS1106IR (Fig. 7). We are currentlyinvestigating the nature of complex III (Fig. 7). It is possiblethat it is formed by multimers of IS1106Tip with a molecule(s)of IS1106 IR. Competition with the transposase for binding tothe ends of the element and formation of multimers may ac-count for the ability of IS1106Tip to act as a negative modu-lator of IS1106 transposition.
Lack of IS1106Tip and, possibly, hypertransposition maycontribute to plasticity of the meningococcal genomes in sev-eral pathogenic clones, playing an adaptive role and leading tochanges in virulence in the course of evolution. For instance, ithas been proposed that IS1106-mediated transposition andrecombination may be involved in genetic instability at theporA locus, thereby influencing antigenic variation of this im-portant surface antigen (26). This hypothesis is further sup-ported by computer sequence analysis of the meningococcalgenome (serogroup A strain Z2491). The IS1106 elements arelocated close to genes encoding virulence factors and subjectedto genetic variation, including lbpAB, encoding the lactoferrinreceptor (43, 44), and frpC, a meningococcus-specific geneabsent in gonococci (as well as porA) (11) and coding for aniron-regulated protein related to the RTX family of cytotoxins(56).
ACKNOWLEDGMENTS
We thank P. Di Nocera for useful suggestions and critical reading ofthe manuscript. We thank M. Frosch, J. C. Chapalain, J. M. Alonzo, P.Nicolas, V. Scarlato, and P. Mastrantuono for providing meningococ-cal strains.
This work was partially supported by grants from the MURST-PRIN
program (D.M. n. 503 DAE-UFFIII, 18/10/1999) and MURST-CNRBiotechnology Program L. 95/95.
REFERENCES
1. Arber, W. 1993. Evolution of prokaryotic genomes. Gene 135:49–56.2. Black, C. G., J. A. M. Fyfe, and J. K. Davies. 1995. A promoter associated
with the neisserial repeat can be used to transcribe the uvrB gene fromNeisseria gonorrhoeae. J. Bacteriol. 177:1952–1958.
3. Blot, M. 1994. Transposable elements and adaptation of host bacteria. Ge-netica 93:5–12.
4. Bohannon, D. E., N. Connell, J. Keeneer, A. Tormo, M. Espinosa-Urgel,M. M. Zambrano, and R. Kolter. 1991. Stationary-phase-inducible “gearbox”promoters: differential effects of katF mutations and role of �70. J. Bacteriol.173:4482–4492.
5. Borst, P., and D. R. Greaves. 1987. Programmed gene rearrangements al-tering gene expression. Science 235:658–667.
6. Brynestad, S., B. Synstad, and P. E. Granum. 1997. The Clostridium perfrin-gens enterotoxin gene is on a transposable element in type A human foodpoisoning strains. Microbiology 143:2109–2115.
7. Bucci, C., A. Lavitola, P. Salvatore, L. Del Giudice, D. R. Massardo, C. B.Bruni, and P. Alifano. 1999. Hypermutation in pathogenic bacteria: frequentphase variation in meningococci is a phenotypic trait of a specialized mutatorbiotype. Mol. Cell 3:435–445.
8. Chernak, J. M., E. J. Schlaffer, and H. O. Smith. 1988. Synthesis andoverproduction of the 5A protein of insertion sequence IS5. J. Bacteriol.170:5368–5370.
9. Collins, C. M., and D. M. Gutman. 1992. Insertional inactivation of anEscherichia coli urease gene by IS3411. J. Bacteriol. 174:883–888.
10. Correia, F. F., S. Inouye, and M. Inouye. 1988. A family of small repeatedelements with some transposon-like properties in the genome of Neisseriagonorrhoeae. J. Biol. Chem. 263:12194–12198.
11. Dempsey, J. A. F., A. B. Wallace, and J. G. Cannon. 1995. The physical mapof the chromosome of a serogroup A strain of Neisseria meningitidis showscomplex rearrangements relative to the chromosomes of the two mappedstrains of the closely related species N. gonorrhoeae. J. Bacteriol. 177:6390–6400.
12. Dozois, C. M., and R. Curtiss III. 1999. Pathogenic diversity of Escherichiacoli and the emergence of ‘exotic’ islands in the gene stream. Vet. Res.30:157–179.
13. Favaloro, J., R. Treisman, and R. Kamen. 1980. Transcription maps ofpolyoma virus-specific RNA: analysis by two-dimensional nuclease S1 gelmapping. Methods Enzymol. 65:718–749.
14. Fetherston, J. D., and R. D. Perry. 1994. The pigmentation locus of Yersiniapestis KIM6� is flanked by an insertion sequence and includes the structuralgenes for pesticin sensitivity and HMWP2. Mol. Microbiol. 13:697–708.
15. Files, J. G., K. Weber, C. Coulondre, and J. H. Miller. 1975. Identification ofthe UUG codon as a translational initiation codon in vivo. J. Mol. Biol.95:327–330.
16. Finlay, B. B., and S. Falkow. 1997. Common themes in microbial pathoge-nicity revisited. Microbiol. Mol. Biol. Rev. 61:136–169.
17. Fleischmann, R. D., M. D. Adams, O. White, R. A. Clayton, E. F. Kirkness,A. R. Kerlavage, C. J. Bult, J. F. Tomb, B. A. Dougherty, J. M. Merrick, etal. 1995. Whole-genome random sequencing and assembly of Haemophilusinfluenzae Rd. Science 269:496–512.
18. Foster, P. L. 1993. Adaptive mutation—the uses of adversity. Annu. Rev.Microbiol. 47:467–504.
19. Frosch, M., E. Schultz, E. Glenn-Calvo, and T. F. Meyer. 1990. Generationof capsule-deficient Neisseria meningitidis strains by homologous recombina-tion. Mol. Microbiol. 4:1215–1218.
TABLE 2. In vitro transposition of IS1106::ermC� in the presence and absence of IS1106Tipa
Donor plasmidAmt of (ng) of: Frequency of erythromycin-
resistant clones (Ermr)Frequency of rifampin-resistant clones (Rifr) Ermr/Rifr ratio
IS1106T IS1106Tip
None �10�9 5.6 � 10�5 NDb
pUC1106::ermC� �10�9 3.4 � 10�5 NDpUC1106::ermC� 500 6.3 � 10�7 7.3 � 10�6 8.6 � 10�2
pUC1106::ermC� 500 10 1.2 � 10�7 7.1 � 10�6 1.7 � 10�2
pUC1106::ermC� 500 50 3.6 � 10�8 6.2 � 10�6 5.8 � 10�3
pUC1106::ermC� 500 250 5.0 � 10�9 2.7 � 10�6 1.9 � 10�3
a The assay measured the movement of the transposase-defective element from donor plasmid pUC1106::ermC� to the target, N. meningitidis rifampin-resistantchromosomal DNA. The target DNA was derived from a rifampin-resistant variant of strain BL859. Transposition reactions were carried out in 20 �l in the presenceof 1 �g of donor plasmid, 1 �g of target DNA, 500 ng of IS1106T, and different amounts of IS1106Tip. Before the DNA was transformed into rifampin-sensitive parentalstrain BL859, single-stranded gaps possibly introduced upon transposition were repaired (Materials and Methods). Transposition events were scored as the recoveryof erythromycin-resistant host cells after transformation. The ratio of the total number of erythromycin-resistant transformants to the total number of rifampin-resistanttransformants was a measure of IS1106::ermC� transposition. The values shown are means of at least five independent experiments.
b ND, not determined.
VOL. 69, 2001 IS1106 TRANSPOSASE INHIBITOR PROTEIN 7435
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from

20. Groisman, E. A., and H. Ochman. 1996. Pathogenicity islands: bacterialevolution in quantum leaps. Cell 87:791–794.
21. Groisman, E. A., and H. Ochman. 1997. How Salmonella became a patho-gen. Trends Microbiol. 5:343–349.
22. Haas, R., and T. F. Meyer. 1986. The repertoire of silent pilus genes inNeisseria gonorrhoeae: evidence for gene conversion. Cell 44:107–115.
23. Hacker, J., G. Blum-Oehler, L. Muhldorfer, and H. Tschape. 1997. Patho-genicity islands of virulent bacteria: structure, function and impact on mi-crobial evolution. Mol. Microbiol. 23:1089–1097.
24. Hammerschmidt, S., R. Hilse, J. P. M. van Putten, R. Gerardy-Schahn, A.Unkenmeir, and M. Frosch. 1996. Modulation of cell surface sialic acidexpression in Neisseria meningitidis via a transposable genetic element.EMBO J. 15:192–196.
25. Knight, A. I., H. Ni, K. A. V. Cartwright, and J. J. McFadden. 1990. Iden-tification of a UK outbreak strain of Neisseria meningitidis with a DNAprobe. Lancet 335:1182–1184.
26. Knight, A. I., H. Ni, K. A. V. Cartwright, and J. J. McFadden. 1992. Iden-tification and characterization of a novel insertion sequence, IS1106, down-stream of the porA gene in B15 Neisseria meningitidis. Mol. Microbiol.6:1565–1573.
27. Kroger, M., and G. Hobom. 1982. Structural analysis of insertion sequenceIS5. Nature 297:159–162.
28. Lange, R., and R. Hengge-Aronis,. 1991. Growth phase-regulated expressionof bolA and morphology of stationary-phase Escherichia coli cells are con-trolled by the novel sigma factor �s. J. Bacteriol. 173:4474–4481.
29. Lange, R., and R. Hengge-Aronis. 1991. Identification of a central regulatorof stationary-phase gene expression in Escherichia coli. Mol. Microbiol. 5:49–59.
30. Lee, C. A. 1996. Pathogenicity islands and the evolution of bacterial patho-gens. Infect. Agents Dis. 5:1–7.
31. Mahillon, J., J. Seurinck, L. van Rompuy, J. Delcour, and M. Zabeau. 1985.Nucleotide sequence and structural organization of an insertion sequence(IS231) from Bacillus thuringiensis strain berliner 1715. EMBO J. 4:3895–3899.
32. Mahillon, J., and M. Chandler. 1998. Insertion sequences. Microbiol. Mol.Biol. Rev. 62:725–774.
33. Mecsas, J. J., and E. J. Strauss. 1996. Molecular mechanisms of bacterialvirulence: type III secretion and pathogenicity islands. Emerg. Infect. Dis.2:270–288.
34. Mobley, H. L. 1997. Helicobacter pylori factors associated with disease devel-opment. Gastroenterology 113(Suppl. 6):S21–S28.
35. Naas, T., M. Blot, W. M. Fitch, and W. Arber. 1994. Insertion sequence-related genetic rearrangements in resting Escherichia coli K-12. Genetics136:721–730.
36. Naas, T., M. Blot, W. M. Fitch, and W. Arber. 1995. Dynamics of IS-relatedgenetic rearrangements in resting Escherichia coli K-12. Mol. Biol. Evol.12:198–207.
37. Nei, M. 1975. Molecular polymorphism and evolution. North-Holland Pub-lishing Co., Amsterdam, The Netherlands.
38. Newcombe, J., K. Cartwright, S. Dyer, and J. McFadden. 1998. Naturallyoccurring insertional inactivation of the porA gene of Neisseria meningitidisby integration of IS1301. Mol. Microbiol. 30:453–457.
39. Ni, H., A. I. Knight, K. A. V. Cartwright, and J. J. McFadden. 1992. Phylo-genetic and epidemiological analysis of Neisseria meningitidis using DNAprobes. Epidemiol. Infect. 109:227–239.
40. Ni, H., A. I. Knight, K. A. V. Cartwright, W. H. Palmer, and J. J. McFadden.1992. Polymerase chain reaction for diagnosis of meningococcal meningiti-dis. Lancet 340:1432–1434.
41. Parkhill, J., M. Achtman, K. D. James, S. D. Bentley, C. Churcher, S. R.Klee, G. Morelli, D. Basham, D. Brown, T. Chillingworth, et al. 2000. Com-
plete DNA sequence of a serogroup A strain of Neisseria meningitidis Z2491.Nature 404:502–506.
42. Perrin, A., X. Nassif, and C. Tinsley. 1999. Identification of regions of thechromosome of Neisseria meningitidis and Neisseria gonorrhoeae which arespecific to the pathogenic Neisseria species. Infect. Immun. 67:6119–6129.
43. Pettersson, A., V. Klarenbeek, J. van Deurzen, J. T. Poolman, and J. Tom-massen. 1994. Molecular characterization of the structural gene for thelactoferrin receptor of the meningococcal strain H44/76. Microb. Pathog.17:395–408.
44. Pettersson, A., T. Prinz, A. Umar, J. van der Biezen, and J. Tommassen.1998. Molecular characterization of LbpB, the second lactoferrin-bindingprotein of Neisseria meningitidis. Mol. Microbiol. 27:599–610.
45. Pizza, M., V. Scarlato, V. Masignani, M. M. Giuliani, B. Arico, M. Coman-ducci, G. T. Jennings, L. Baldi, E. Bartolini, B. Capecchi, et al. 2000.Identification of vaccine candidates against serogroup B meningococcus bywhole-genome sequencing. Science 287:1816–1820.
46. Rezsohazy, R., B. Hallet, J. Delcour, and J. Mahillon, J. 1993. The IS4 familyof insertion sequences: evidence for a conserved transposase motif. Mol.Microbiol. 9:1283–1295.
47. Rivellini, F., P. Alifano, C. Piscitelli, V. Blasi, C. B. Bruni, and M. S. M. S.Carlomagno. 1991. A cytosine over guanosine-rich sequence in RNA acti-vates Rho-dependent transcription termination. Mol. Microbiol. 5:3049–3054.
48. Saint Girons, I., and A. G. Barbour. 1991. Antigenic variation in Borrelia.Res. Microbiol. 142:711–717.
49. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
50. Smith, J. T., and C. S. Lewin. 1993. Mechanisms of antimicrobial resistanceand implications for epidemiology. Vet. Microbiol. 35:233–242.
51. Sneath, P. H. A., and R. R. Sokal. 1973. Numerical taxonomy. W. H. Free-man & Co., San Francisco, Calif.
52. Stroeher, U. H., K. E. Jedani, B. K. Dredge, R. Morona, M. H. Brown, L. E.Karageorgos, M. J. Albert, and P. A. Manning. 1995. Genetic rearrange-ments in the rfb regions of Vibrio cholerae O1 and O139. Proc. Natl. Acad.Sci. USA 92:10374–10378.
53. Studier, F. W., and B. A. Moffatt. 1986. Use of bacteriophage T7 RNApolymerase to direct selective high-level expression of cloned genes. J. Mol.Biol. 189:113–130.
54. Tabor, S., and C. C. Richardson. 1985. A bacteriophage T7 RNA polymer-ase/promoter system for controlled exclusive expression of specific genes.Proc. Natl. Acad. Sci. USA 82:1074–1078.
55. Tettelin, H., N. J. Saunders, J. Heidelberg, A. C. Jeffries, K. E. Nelson, J. A.Eisen, K. A. Ketchum, D. W. Hood, J. F. Peden, R. J. Dodson, et al. 2000.Complete genome sequence of Neisseria meningitidis serogroup B strainMC58. Science 287:1809–1815.
56. Thompson, S. A., L. L. Wang, and P. F. Sparling. 1993. Cloning and nucle-otide sequence of frpC, a second gene from Neisseria meningitidis encoding aprotein similar to RTX cytotoxins. Mol. Microbiol. 9:85–96.
57. White, D. A., A. K. Barlow, I. N. Clarke, and J. E. Heckels. 1990. Stableexpression of meningococcal class 1 protein in an antigenically reactive formin outer membranes of Escherichia coli. Mol. Microbiol. 4:769–776.
58. Ziebuhr, W., V. Krimmer, S. Rachid, L. Lossner, F. Gotz, and J. Hacker.1999. A novel mechanism of phase variation of virulence in Staphylococcusepidermidis: evidence for control of the polysaccharide intercellular adhesinsynthesis by alternating insertion and excision of the insertion sequenceelement IS256. Mol. Microbiol. 32:345–356.
59. Zubay, G. 1973. In vitro synthesis of protein in microbial systems. Annu. Rev.Genet. 7:267–287.
Editor: D. L. Burns
7436 SALVATORE ET AL. INFECT. IMMUN.
on May 30, 2016 by guest
http://iai.asm.org/
Dow
nloaded from




















