
i | P a g e
45
i | Page COLLEGE OF NATURAL AND APPLIED SCIENCE Department of zoology and wildlife science conservation FINAL YEAR RESEARCH PROJECT TOPIC: COMPARISON OF TERMITE ASSEMBLAGES IN TWO HABITAT TYPES AT UNIVERSITY OF DAR ES SALAAM MWALIMU NYERERE CAMPUS STUDENT NAME: WARYOBA EMMANUEL M STUDENT NUMBER: 2010-04-03984 COURSE NAME: Bachelor of Science in wildlife science & conservation DEPARTMENT: Department of zoology and wildlife conservation FACULTY: Science COURSE CODE: BL 390: RESEARCH PROJECT SUPERVISOR: Dr. B.N. Nyundo DATE OF SUBMISSION: 17.MAY .2013, submitted as dissertation report in fulfillment of the requirement for the degree of Bachelor of Science in wildlife science and conservation of university of dare s salaam
Transcript of i | P a g e

i | P a g e
COLLEGE OF NATURAL AND APPLIED SCIENCE
Department of zoology and wildlife science conservation
FINAL YEAR RESEARCH PROJECT
TOPIC:
COMPARISON OF TERMITE ASSEMBLAGES IN TWO HABITAT TYPES AT
UNIVERSITY OF DAR ES SALAAM MWALIMU NYERERE CAMPUS
STUDENT NAME: WARYOBA EMMANUEL M STUDENT NUMBER: 2010-04-03984 COURSE NAME: Bachelor of Science in wildlife science & conservation DEPARTMENT: Department of zoology and wildlife conservation FACULTY: Science COURSE CODE: BL 390: RESEARCH PROJECT
SUPERVISOR: Dr. B.N. Nyundo
DATE OF SUBMISSION: 17.MAY .2013, submitted as dissertation report in
fulfillment of the requirement for the degree of Bachelor of Science in wildlife
science and conservation of university of dare s salaam

i | P a g e
ACKNOWLEGEMENT
I would like to express my acknowledge to the Higher Education student Loan Board (
HESLB) for financial support provided to conduct this work under university of Dar es salaam,
zoology and wildlife conservation department for their permission to allow this study to be
conducted and their moral support and necessary materials required to accomplish this study.
The necessary useful information and positive instruction about my research work would not be
met if not successful provided by Dr. F. Magige (research coordinator). My best regards should
go to Dr. B.A. Nyundo (My supervisor) for his relentless encouragement, moral support, skillful
and useful information for a comprehensive work and produce coherent report.
Many people assisted me in one way or another though their names are not mentioned here. But
mention must be made of Professor KIM Howell, Dr. H. Kiwia for advice and useful discussion
during project proposal presentation. Their contributions are highly appreciated. I wish to thank
Mr. Mwangoka for his materials support.
I am also grateful to Mr. Mfaume from department of Zoology and Wildlife Conservation,
University of Dar es Salaam (UDSM) for assisting termite identification. The head of
Department for his authorization in different aspects help this study to be conducted in terms of
laboratory equipments and other staff of the UDSM for their assistance at various levels of my
study. My gratitude is also extended to my friend and colleagues in the Zoology building who
encouraged me during difficult time. Their challenges made me work even harder; I thank them
for that too.

ii | P a g e
DEDICATION
Here dedication is made to My Sister for her patience, love and support. This work is also
dedicated to my late parents; Mr. and Mrs. Waryoba who with the God love could not remain
alive until I become the first member of the family to attain a stage of being awarded a Bachelor
degree. Furthermore to my young brother (Godfrey) who devotes much his time for
encouragement

iii | P a g e
ABSTRACT
Termite species richness, evenness, and abundance in university of Dar es Salaam are poorly
known at a time when biodiversity is being threatened by human activities. Termites family
richness of two selected habitat at University of dare s salaam were determined between 1st of
January and 12nd
of January 2013 in bush thicket forest and wooded grassland habitats. The
objective of study was to compare the assemblages of termite in two selected habitat at
university of dare s salaam Mwalimu Nyerere campus
Termites were sampled along a transect line 100m and 2m wide in two habitat types. A total of
12 subfamilies were found. From which 1988 individuals were recorded during early wet
season, 690 were from forest and 1298 from wooded grassland. Termites abundance was
significantly higher in the wooded grassland (U = 277.500, P= 0.011). Termites subfamilies
richness was higher in bush thicket forest habitat than wooded grassland but the difference was
not statistically significant (U= 59.5, P = 0.47). Comparison between termite functional groups
(i, ii and iii) show no significant difference (K=0.671, P = 0.715). Taxonomic composition
between two selected habitats was not statistically significant ( 2= 3.000, 0.99<p<0.995) and
diversity between two sites was not significant different (t = 0.74) at level of 5%. This study offers
a baseline data on termite communities at University of Dar es salaam. This results can be used
for further research, monitoring and develop sustainable conservation measures.

iv | P a g e
Table of Contents
ACKNOWLEGEMENT .................................................................................................................. i
DEDICATION ................................................................................................................................ ii
ABSTRACT ................................................................................................................................... iii
Table of Contents ........................................................................................................................... iv
LIST OF FIGURES ....................................................................................................................... vi
LIST OF TABLES ........................................................................................................................ vii
LIST OF APPENDICES .............................................................................................................. viii
Chapter One .................................................................................................................................... 1
1.0 INTRODUCTION .................................................................................................................... 1
1.1 General introduction ............................................................................................................. 1
1.1.1 Termite functional group between habitats........................................................................ 1
1.1.2 Termite species richness and diversity between habitat types ........................................... 2
1.1.3 Termite abundance between habitats ................................................................................. 3
1.2 Statement of Research Problem ............................................................................................ 4
1.3 General Objectives of Research ............................................................................................ 4
1.3.1 The Specific objectives ...................................................................................................... 4
1.4. Significance of the Study ..................................................................................................... 5
1.5 Literature Review.................................................................................................................. 5
1.7 Study Area Description ......................................................................................................... 9
1.6 research hypothesis ............................................................................................................. 10
Chapter Two.................................................................................................................................. 12
2.0 RESEARCH DESIGN AND METHODS .............................................................................. 12
2.1 Sampling of Termites .......................................................................................................... 12

v | P a g e
2.2 Population and Study Sample ............................................................................................. 12
2.3 Materials and Equipment .................................................................................................... 13
2.4 Data Analysis .......................................................................................................................... 13
Chapter Three................................................................................................................................ 14
3.0 RESULTS ........................................................................................................................... 14
3.1 The Termite Abundance in Study Habitats ......................................................................... 14
3.2 Comparison of Termite Abundance between Habitat types ............................................... 15
3.2 The diversity of termite ....................................................................................................... 16
3.3 Diversity and Evenness ....................................................................................................... 16
3.4 Species Accumulation Curves ............................................................................................ 19
Chapter Four ................................................................................................................................. 23
4.0 DISCUSSION ......................................................................................................................... 23
4.1 Introductory Remarks ......................................................................................................... 23
4.1.1Termite Abundance in the selected Habitat ...................................................................... 23
4.1.2 Termites Species Diversity between Habitat types .......................................................... 24
4.1.3 Termites Species Richness ............................................................................................... 25
4.1.4 Termite Functional Group in Different Habitat Types .................................................... 25
4.1.5 Termite Taxonomic Composition .................................................................................... 26
5.0 CONCLUSION ....................................................................................................................... 27 5.1Finding from this Study ....................................................................................................... 27 5.2 Recommendation ................................................................................................................ 27
6.0 REFERENCES ....................................................................................................................... 29
7.0 APPENDICES ........................................................................................................................ 33 Appendix 1: .............................................................................................................................. 33 Appendix 2: .............................................................................................................................. 34 Appendix 3: .............................................................................................................................. 35

vi | P a g e
LIST OF FIGURES
Figure 1: shows the University of Dare S Salaam Map showing parts of studying site (Bush
thicket Forest). The site is behind auxiliary police office and wooded grassland along
university Road to Ubungo and adjacent to university sports area ........................................... 11 Fig 2 above Shows Abundance of Termite in Two Selected Habitat of Wooded Garland and
Bush Thicket Forest .................................................................................................................. 15 Figure 4: Species accumulation curves for wooded grassland the habitat ................................ 20
Figure 5: Species accumulation curves for the habitat ............................................................. 20

vii | P a g e
LIST OF TABLES
Table 1: Termites Abundance (Summary Table) In the Two Habitats Types during the Wet
Season ....................................................................................................................................... 14 Table 2: Statistical Comparison of Significance Difference in Abundance of Termite between
Two Habitats at University of Dar Es Salaam Main Campus .................................................. 16 Table 3 comparison of (H`) and Evenness (J) of two selected habitats .................................... 17 Table 4: termite subfamilies checklist absent/present in their respective habitats at university
of Dar es salaam main campus.................................................................................................. 18 Table 5: termite family richness and abundance in two habitats at university of Dar es salaam
main campus ............................................................................................................................. 19 Table 6: termite functional groups from the two habitats at university of Dar es Salaam main
campus ...................................................................................................................................... 21 Table 7 termite Functional groups and their respective subfamilies categorized at University of
dare s salaam main campus ....................................................................................................... 22

viii | P a g e
LIST OF APPENDICES
Appendix 1: comparison of termite functional groups ............................................................. 33
Appendix 2: comparison of termite microhabitats.................................................................... 34 Appendix 3: shows 30 samples collected on each habitat and their respective subfamily
richness between two selected habitats for three days to obtain cumulative curve .................. 35

1 | P a g e
Chapter One
1.0 INTRODUCTION
1.1 General introduction
Termite is common name for social insects of about 2800 species in seven(7) families so far
known to occur in tropical forest and grassland savanna area worldwide include Mastotermitidae,
Kalotermitidae, Kodotermitidae, Kermopsidae, Rhinotermitidae, Seritermitidae, and Termitidae.
Taxonomically, termite placed into Phylum Arthropoda, Class Insecta, and Order Isoptera
(Krishna et al 1970)
Within a community there are often sets of species that tend to congregate, due to using
particular resources in similar manners these are known ‘guild’. Community structure is the
species abundance, composition, and interaction of various abiotic (environmental) and biotic
(relations with other species or individuals and abiotic factors that may affect this structure).
Declining of forest cover, loss of biodiversity and increased human population contribute
directly to termite abundance and species richness (Kayan, et al., 1979; Fragoso et al., 1994;
Nash et al., 1996)
1.1.1 Termite functional group between habitats
Functional groups of termite’s assemblages are based on feeding habits and nest group (
Eggleton et al. 1996), the functional groups classified as; Soil-feeding: termites that feed on
humus and mineral soil (their food sources include plant-decaying materials, dead foliage,
woody materials, roots, seeds and the faces of higher animals. Wood-feeding: termites that feed
on dead wood. Soil/wood interface-feeding: termites that feed on extremely decayed wood that
has lost its structure and become soil-like. This is synonymous with the `intermediate feeders'
(Brouwer at el 1993) Litter-feeding: termites that feed predominantly on leaf litter and small

2 | P a g e
items of woody trash. Epiphyte-feeding: Hospitalitermes is known to feed on lichens and other
free-living non-vascular plants, which they graze from the surface of tree trunks (Collins et al
1979). Hypogeal or subterranean nesters: termites whose colony centers are below the ground
without any indication of their presence (wood and Johnson, 1986). They use their faeces or
mixture of feaces and mineral soil in nest construction. The colony centers are often poorly
defined and characteristics, especially in the soldier less Apicotermitinae. But in some
Macrotermitinae, Apicotermes, and Aomallotermes, a little internal structure or surface holes are
present together with complex underground nests to enable foraging on above vegetation. This
group also includes many species that are facultative secondary inhabitant of epigeal mounds
such as Microcerotermes, Pericarpritermies and soldier less Apicotermitinae (Eggleton et al.,
1996)
1.1.2 Termite species richness and diversity between habitat types
Differences in food availability (diverse food resources), vegetation structure, (sparse and dense),
and microclimate differs from one habitat to another and consequently affects the species
richness and diversity of termites between habitats. Species diversity in the forest is high
compare to other habitats such as grassland, woodland, and cropland due to availability of
various foods resources in the forest. Retention of canopy cover in the forest was stated as the
factors contribute to high species richness than in grassland, and cropland (Materu, 2012)
More heterogeneous and complex habitat has greater species diversity than homogeneous
habitats perhaps, due to more opportunities for niche differentiation and resource partitioning
hence a significant decrease in competition between similar or different species (Wood, 1978)

3 | P a g e
Dense vegetation in the forest provides more diverse food resources which attract many more
diverse termite species in the forest microclimate than less dense vegetation plant cover in other
habitats, such as wooded grassland, grassland and farmland which all characterized by sparse
vegetation. Tree canopy cover reduces solar radiation falling on the forest floor compared to that
of other habitat such as wooded grassland, grassland, grassland, and farmland at which when
radiation falls down the ground it reduce soil moisture holding capacity and consequently, makes
the habitat not favorable for termites assemblages.
1.1.3 Termite abundance between habitats
In tropics, habitat types influences abundance, seasonal distribution and diversity of termites
(Eggleton 2000). Other environmental biotic factor such as vegetation types and ground cover
affects termite abundance and species richness. The distribution of termite at different habitats
probably might be influenced by food availability and plants biomass (Watson and Gay 1991).
Termite abundance from different habitats is known to vary from one habitat to another in
accordance to species type and human activities (Magurran, 2004). Probably altering the forest
habitat to grassland or farmland influences food availability, nesting site and reduce natural
predators (Holloway et al, 1992) and this will contribute to high abundance of termite’s
assemblages in these habitat and lower species richness and diversity. For example, slashing
stems and leaves produce large amount of biomass in the field as termite food.
Termites form component of the ecosystem and hence their species richness abundance should
be understood to all habitats not for the sake of only comparison among habitat but enabling to

4 | P a g e
monitor distribution, abundance, species richness, and take appropriate conservation measures.
However, no comprehensive study on the abundance and species richness of termite has been
undertaken in wooded grassland and thicket forest at university of Dar es salaam Mwalimu
Nyerere campus. Therefore this study will try to fill the gap.
1.2 Statement of Research Problem
Lack of baseline information from different habitats hampers the design of remedial measures
and management strategies; loss of biodiversity in the earth is now a major global concern before
we know what is missing, there is progressive change of habitats types now at which natural
forest, woodland and grassland has been changed into farmland, rangeland, settlement, and other
land use forms and consequently affects species richness, and diversity of termites assemblages.
However, no comprehensive studies done in wooded grassland and only few studies has been
done on forest, grassland, and farmland in East Africa and Tanzania as well (Nyeko and
Olubayo, 2007) while no any studies has been done at my study area (university of dar es
salaam). The current studies assess and compare the termite assemblages in terms of species
richness, abundance and diversity in wooded grassland and Thicket forest.
1.3 General Objectives of Research
To assess the influence of habitat types, namely; wooded grassland and thicket forest on termite
assemblages (community)
1.3.1 The Specific objectives
1.0 To compare species richness and diversity between the selected habitat
2.0 To compare termites functional groups between selected habitat
3.0 To compare termites taxonomic groups between two selected habitats

5 | P a g e
1.4. Significance of the Study
Following incredible niche termites fulfills; soil formation, nutrient cycling, and foods to human,
birds, reptiles such as gecko, skink, lizard, and echidnas.(Martin, 2000), waste recycling ,Seed
germination emergence residence room for beetles, mice, python and as good and efficient
bioreactors. This research will have significance future usages to entomologist and conservation
biologist. The study will provide base line information and supplements the required information
necessary for conservation action using termites as key taxa and also, this study will have
considerable implication for the management and protection of termite species in forest, and
wooded grassland. These finding will provide important information crucial for the formulation
of management policies and plans as part of the effort to conserve the overall biodiversity values
(Wood, 1978)
The study will also contribute on the information that helps in biotechnology, biochemistry, and
Ecological future researches about termites more ever, research in Tanzania will incorporate the
company generated geochemical data, regolith and landform characteristics and local geology
have provided much information concerning Africa mound-building termites, and the use of
Termitaria in mineral exploration, both artisanal and modern.
1.5 Literature Review
A biodiversity base line study on species Richness, Functional Diversity and Relative
Abundance of Termites under Different Land Use Regimes in Jambi Province, Central Sumatra,
Indonesia by, 2000 showed termites are more abundant in the primary forest and least abundant
in the cassava garden. The collected termites followed into four feeding group. Wood-feeding

6 | P a g e
and soil-feeding species are relatively abundant in most transects, while epiphyte-feeders are rare
and interface-feeders (those species that feed on extremely decayed soil-like wood) vary
considerably in abundance among transects
A biodiversity study done in the southern Cameroon on The diversity, abundance, and biomass
of termites under differing levels of disturbance in the Mbalmayo Forest Reserve, mentioned
termites to achieve very high population densities where by termites were one of the numerous
of all arthropods group and this, was supported by (Eggleton et al. 1996)
Detailed knowledge of local species diversity as well as the diversity of functional groups of
termites (different groups of species that have different ecological strategies, e.g. different
feeding habits) is critical (Dawes-Gromadzki, 2005). It is a prerequisite for evaluating and
quantifying the type and extent of their functional roles in ecosystem processes, i.e. the
contributions that different termite species and functional groups make to the regulation of
different ecosystem processes. It is also important for the development of sound management
practices, for example agricultural, grazing and fire management (Jones, 2002) also noted that
accurate assessments of structures of termite assemblages are necessary to explain the ecological
influence of termites on various sites. Further observed that sustainable management of soil
macro fauna in Uganda is constrained by a shortage of information on the activity, behavior and
environmental tolerances of many species of this fauna and by the limited understanding of the
structural and functional stability of soil fauna communities in general
At a given site the composition of a local termite assemblage will govern the overall impacts of
termites on ecosystem processes (Lavelle et al., 1997; Jones & Prasetyo, 2002). This is because

7 | P a g e
most termite assemblages are made up of species that represent a diverse range of feeding for
example; wood, soil, grass) and nesting ( mounds, hypogeal and arboreal nests, nests in wood)
strategies ( different ‘functional groups’). Consequently different species and groups of species
are likely to have different ecological effects on ecosystem components (Dawes-Gromadzki,
2005).
Termites assemblages study conducted at an altitude of 1000 m by David T. Jones (2000) at two
primary forest habitats: Termite lower Montane forest and stunted fancies of upper Montane
forest. Soil, wood, litter, termite nests, and arboreal microhabitats up to a height of 2 m above
ground were sampled using a belt transect protocol. Two transects were run in each forest type.
The upper montane forest and the lower montane forest transect showed similar species richness
and relative abundance of wood-feeding termites was found in both forest types. However, the
lower montane forest had greater richness and relative abundance of species that feed on soil and
extremely decayed soil-like wood. This difference is attributed mainly to (i) the well developed,
organic-rich soils of the lower montane forest being more suitable for soil-dwelling and mound-
building termites compared with the upper montane forest which has sandy soils with a thick
covering of peat, and (ii) the low-stature and open canopy of the upper montane forest creates
microclimatic conditions which are adverse for soil-feeding termites compared with the high,
closed canopy in the lower montane forest. As expected, comparisons with similar studies in
lowland forests in Southeast Asia confirmed that the upland forests are relatively depauperate.
(Jones, 2000)

8 | P a g e
At a given site the composition of a local termite assemblage will govern the overall impacts of
termites on ecosystem processes (Lavelle et al., 1997). This is because most termite assemblages
are made up of species that represent a diverse range of feeding (e.g. wood, soil, grass) and
nesting (e.g. mounds, hypogeal and arboreal nests, nests in wood) strategies (i.e. different
‘functional groups’). Consequently different species and groups of species are likely to have
different ecological effects on ecosystem components (Dawes-Gromadzki, 2005)
The composition of termite assemblages was analyzed in study done by Alves, between 2007
and 2009 at three sites in the state of Rio Grande does Norte, Brazil. Where by four feeding
groups were sampled: wood-feeders, soil-feeders, wood-soil interface feeders and leaf-feeders.
The wood-feeders were dominant in number of species and number of encounters at all sites. In
general, the sites were not significantly different in relation to the environmental variables
measured. The same pattern was observed for termite assemblages, where no significant
differences in species richness, relative abundance and taxonomic and functional composition
were observed between the three sites. The agreement between composition of assemblages and
environmental variables reinforces the potential of termites as biological indicators of habitat
quality.
Given the ecological importance of termites, there is a need to characterize termite assemblage
structure within and between sites. As a consequence of their highly patchy spatial distribution,
combined with the many and varied field sampling regimes adopted by previous researchers, it
has not been possible to use the existing data to make reliable direct comparisons of termite
diversity and abundance between sites this was according to Eggleton at el, 1995. Also it is

9 | P a g e
necessary to develop and test standardized sampling methods that can be applied easily
throughout the tropics. To this end, a standardized transects sampling method designed to
measure termite species richness and functional diversity in tropical forests has been developed.
The protocol has been used in Cameroon (Eggleton et al, 1995), Thailand (Davies, 1997) in his
study titled Termite species richness in fire-prone and fire-protected dry deciduous dipterocarp
forest in DoiSuthep-PuiNational Park
Study carried out in Malaysia observed that termite is more abundant in the forest and grassland
during the wet season than during dry season. Diversity of termite has been stated as the
indicator of disturbance in natural ecosystem (Bandeira, 1979). Low level of species richness and
abundance is an indicator of level of environment disturbance (Avenant, 2000). A change in
termite habitat characteristics such as plants species composition often affects termite species
richness. Destruction of forest and grassland habitats influences termite abundance and species
richness.
1.7 Study Area Description
The present study was conducted at the University of Dar es Salaam main campus which is
situated close to the equator on the coast of Indian Ocean and located at 6 o 48’S to 39
o 17’E
.(Encarta encyclopedia 2007. Dar es Salaam city experiences tropical climatic conditions,
characterized by hot and humid weather throughout the year. Average January High Temperature
is 34 °C/92 °F. Average January Low Temperature is 21 °C/70 °F. Average July High
Temperature is 30 °C/87 °F. Average July Low Temperature is16 °C/60 °F Average Annual
rainfall is approximately 1,105 mm and in a normal year there are two distinct rainy seasons:

10 | P a g e
"the long rains", which fall during April and May, and "the short rains", which fall during
October and November (Encarta encyclopedia, 2007)
1.7.1Study site
This research was conducted at University of Dar es salaam main campus area, in which two
sites were used. The bush thickets forest behind Hill Park and wooded grassland at gate maji
adjacent to university play ground.
1.6 research hypothesis
o Termite richness, and diversity between the selected habitats are significantly different
from each other
o Termite functional group between two selected habitat types are significantly different
o Taxonomic composition between two selected habitats are significantly different

11 | P a g e
Figure 1: shows the University of Dare S Salaam Map showing parts of studying site (Bush
thicket Forest). The site is behind auxiliary police office and wooded grassland along
university Road to Ubungo and adjacent to university sports area
Key= location of bushthicket and grassland patches (study sites).

12 | P a g e
Chapter Two
2.0 RESEARCH DESIGN AND METHODS
2.1 Sampling of Termites
Standardized transect method of Jones and Eggleton (2000) was used for sampling termites.
Belts transect (2 x 100 m) laid in each habitat types. Each transect was divided into 10 (2 x 5 m)
sections and in each section two man- half an hour will be spent searching for termites. Within
each section the following microhabitats were searched; surface soil, leaf litter and humus on the
forest floor (at the base of trees, between buttress roots etc.), inside dead logs, tree stumps,
branches, twigs, mounds, carton sheeting and runways on trees up to a height of 2 m above
ground and arboreal nests up to 2 m height. Mainly soldier and worker castes were collected
from the different microhabitats. Termite family diversity was compared using Shannon
Diversity Index, In addition to sampling. Collected termite specimens were stored in 70%
isopropyl alcohol for identification purposes. Encountered termite nests were examined and
recorded both during sampling and random collections. Identification of termite species collected
from the study area was made by using taxonomic key and literature.
2.2 Population and Study Sample
The study sample of the population was mainly base on the one species of insect (termites) in
two habitat types. The data of the topics under study was delimited obtained through collection
in the field or sampled plot (wooded grassland and bush thicket forest). Collection of Data was
on the basis only one season, wet season. Sorting and identification of collected specimens was
conducted at the laboratory of the University of Dar es salaam, Department of zoology and
wildlife conservation

13 | P a g e
2.3 Materials and Equipment
Data of my topics under study was be delimited obtained through collection in the field or plot,
to meet the demands of quality data, the different materials will be in use to accomplish the
process of data collection and management ( for the case, preservation in 70% alcohol). The
following materials used; note books, pencil, pen, identification books (key), and 70% alcohol.
Equipments used were; microscope, tape measures, machete, plastic bags transect belt, hummer.
2.4 Data Analysis
Descriptive data analysis was used to summarize the mass of information; total number of
termites collected, percentages, and mean.
Diagrammatic presentation of data in this report used the following:
Tabular and Bar charts and graphs.
The main component of species diversity; richness and equitability computed using the Shannon-
Weiner index (H) was and jaccard (J) index of similarity respectively with aid of Diversity
program (2007). The Significance established at P ≤0.05 (significant) and the treatment of
continuous data followed in all cases a similar procedure: Initially, for all variables, descriptive
statistics (mean, median, mode, range, standard deviation, variance, skewness, kurtosis and box
plots) were produced in to assess for normality. Not normally distributed variables were obtained
and analyzed with Mann-Whitney U; Chi-squared tests of independence was used to detect
possible significant differences of the proportion of species in each feeding group to compare
taxonomic composition of the termite between two selected habitats types while non parametric
test, kruskal Wallis employed to detect the significant differences in three functional group of
termite (group, i, ii, and iii)

14 | P a g e
Chapter Three
3.0 RESULTS
3.1 The Termite Abundance in Study Habitats
60 samples were sampled from both habitats (wooded grassland and bush thicket forest). Total
of 1988 Individuals were collected from which 1298 obtained at wooded grassland and 690 from
bush thicket forest. The average maximum number of termite was 343 ranging in 343
individuals with mean of 82.83. The variance of termite abundance is 8576.93 with standard
deviation of 93 and the number of termite skewed by 1.49. Their respective means, standard
deviation, maximum and minimum number are summarized in Table1.
Table 1: Termites Abundance (Summary Table) In the Two Habitats Types during the Wet
Season
wooded grassland Bush Thicket Forest
Minimum 0.0 Minimum 0.0
Maximum 343.0 Maximum 180.0
Range 343.0 Range 180.0
Sum 1298.0 Sum 690.0
Mean 108.17 Mean 57.50
Standard Dev 117.52 Standard Dev 52.18
Variance 13810.15 Variance 2723.13
Skewness(G1) 0.94 Skewness(G1) 1.26
Kurtosis(G2) -0.29 Kurtosis(G2) 1.54
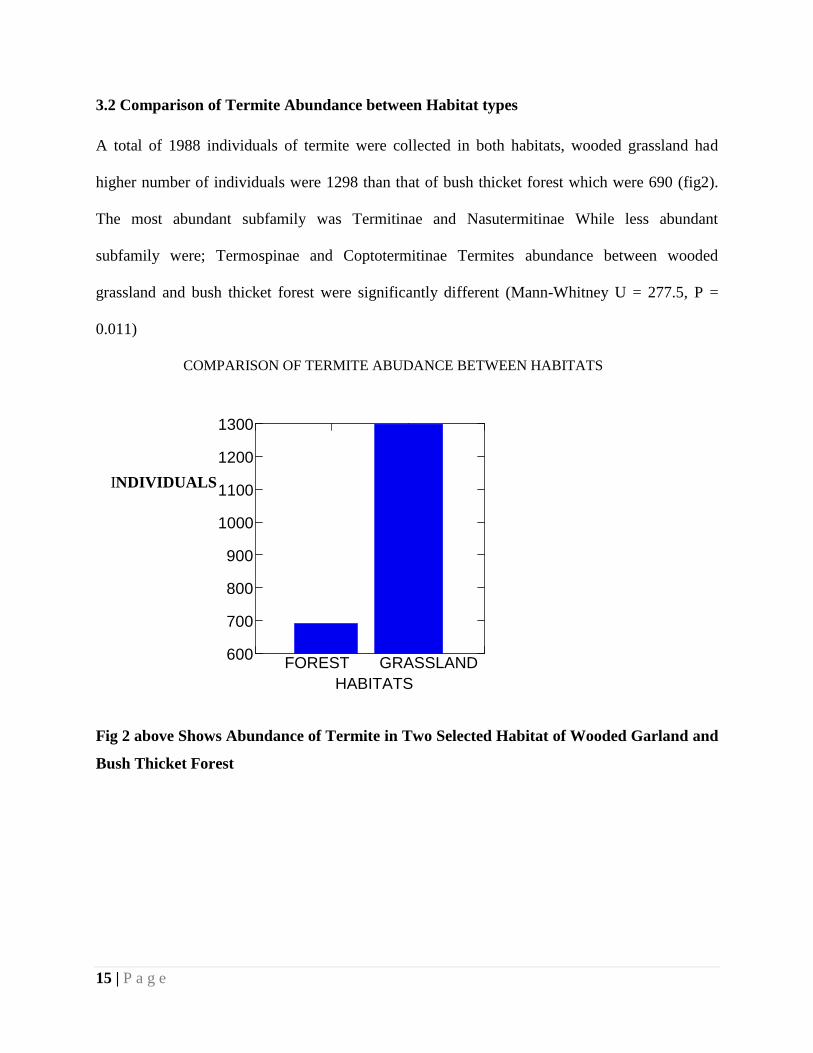
15 | P a g e
3.2 Comparison of Termite Abundance between Habitat types
A total of 1988 individuals of termite were collected in both habitats, wooded grassland had
higher number of individuals were 1298 than that of bush thicket forest which were 690 (fig2).
The most abundant subfamily was Termitinae and Nasutermitinae While less abundant
subfamily were; Termospinae and Coptotermitinae Termites abundance between wooded
grassland and bush thicket forest were significantly different (Mann-Whitney U = 277.5, P =
0.011)
Fig 2 above Shows Abundance of Termite in Two Selected Habitat of Wooded Garland and
Bush Thicket Forest
COMPARISON OF TERMITE ABUDANCE BETWEEN HABITATS
FOREST GRASSLAND
HABITATS
600
700
800
900
1000
1100
1200
1300
INDIVIDUALS

16 | P a g e
Table 2: Statistical Comparison of Significance Difference in Abundance of Termite
between Two Habitats at University of Dar Es Salaam Main Campus
Habitat U Statistic P Significant
forest vs. grassland 277.5 0.011 *
3.2 The diversity of termite
11subfamily of termite species were found in the bush Thicket Forest habitat and only 9 sub
families were found in wooded grassland habitat. The differences in numbers were not
statistically significant.(). Members of subfamily Termitinae and Nasutitermitinae (21%) were
more abundant in wooded grassland, followed by Amitermitinae (16%) and Heterotermitinae
(14%) while other subfamilies such as Hodotermitinae, Macrotermitinae, Psammotermitinae,
kalotermitidae were less abundant (1-4%) had little number of termites. In the bush Thicket
Forest dominant group of termite were recorded to be; Macrotermitinae (26%), genus
Postelectotermes.
3.3 Diversity and Evenness
Subfamily diversity using Shannon- Wiener index (H`) Observed to be (H) = 2.12 for bush
thicket forest habitat and that of wooded grassland (H) = 1.91. And therefore bush thicket forest
was more diverse than wooded grassland. But both had equal diversity at 5% Level (P = 0.7441)
Using an Equitability J index with 10000 random partitions Observed Diversity, bush thicket
forest- FR, J = 0.85 while that of 2nd sample wooded grassland (WG), J = 0.77. And Delta was
0.0838583. Hence species in forest habitat is more evenly distributed than wooded grassland

17 | P a g e
Table 3 comparison of (H`) and Evenness (J) of two selected habitats
Habitats Number of subfamily Diversity(H) Similarity(J)
WOODED GRASSLAND 9 1.91 0.77
BUSH THICKET FOREST 11 2.11 0.85
Overall 11 4.03 1.62
delta 0.20 0.08
Probability(two sided test) 0.7441 0.7405
A total of 12 subfamily of termite were recorded after identification of samples collected in the
field from both habitats. Termospinae, Kaloterimitidae, and Coptotermitinae subfamilies were
only recorded in bush thicket but in few relative numbers. Distinctiveness in termite assemblages
also observed in wooded grassland where by subfamily Psammotemitinae recorded but not in
bush thicket forest. Table 4, summarized subfamilies and their absence or presence in their
respective habitat.
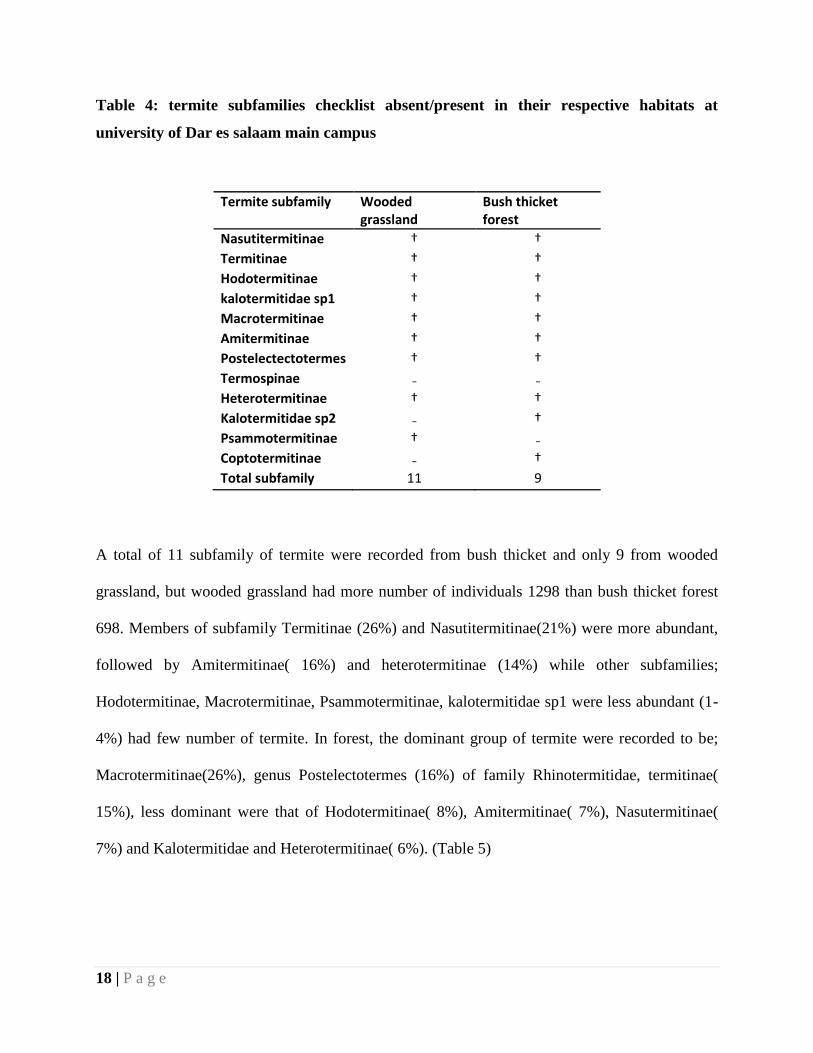
18 | P a g e
Table 4: termite subfamilies checklist absent/present in their respective habitats at
university of Dar es salaam main campus
Termite subfamily Wooded grassland
Bush thicket forest
Nasutitermitinae † †
Termitinae † †
Hodotermitinae † †
kalotermitidae sp1 † †
Macrotermitinae † †
Amitermitinae † †
Postelectectotermes † †
Termospinae ₋ ₋
Heterotermitinae † †
Kalotermitidae sp2 ₋ †
Psammotermitinae † ₋
Coptotermitinae ₋ †
Total subfamily 11 9
A total of 11 subfamily of termite were recorded from bush thicket and only 9 from wooded
grassland, but wooded grassland had more number of individuals 1298 than bush thicket forest
698. Members of subfamily Termitinae (26%) and Nasutitermitinae(21%) were more abundant,
followed by Amitermitinae( 16%) and heterotermitinae (14%) while other subfamilies;
Hodotermitinae, Macrotermitinae, Psammotermitinae, kalotermitidae sp1 were less abundant (1-
4%) had few number of termite. In forest, the dominant group of termite were recorded to be;
Macrotermitinae(26%), genus Postelectotermes (16%) of family Rhinotermitidae, termitinae(
15%), less dominant were that of Hodotermitinae( 8%), Amitermitinae( 7%), Nasutermitinae(
7%) and Kalotermitidae and Heterotermitinae( 6%). (Table 5)

19 | P a g e
Table 5: termite family richness and abundance in two habitats at university of Dar es
salaam main campus
Habitat types
Termite subfamily Wooded grassland Bush Thicket forest Total
Nasutitermitinae 279 51 330
Termitinae 343 104 447
Hodotermitinae 62 60 122
Kalotermitidae sp1 15 48 63
Macrotermitinae 58 180 238
Amitermitinae 203 52 255
Postelectectotermes 116 113 229
Termospinae 0 12 12
Heterotermitinae 182 44 226
Kalotermitidae sp2 0 16 16
Psammotermitinae 40 0 40
Coptotermitinae 0 10 10
Total abundance 1298 690 1988
Family richness 9 11 12
3.4 Species Accumulation Curves
Termite’s species accumulation curves were drawn after pooling the data from different habitats
and plotting the cumulative number of termite species against the area. The number of species
from each habitat increased with the area sampled until the asymptote level was reached. Termite

20 | P a g e
species richness from the wooded grassland and bush thicket forest was recorded from
10quadrats (fig 3 & 5)
Figure 3: Species accumulation curves for wooded grassland the habitat
Figure 4: Species accumulation curves for the habitat

21 | P a g e
3.5 Termite Functional Group between Habitat Types
Termite functional group provides useful information on the roles termite in ecosystem. Four
different groups were identified as wood feeders, fungus growing (wood and leaf litter feeders)
and soil feeders with different roles in the decomposition process. Termite in functional group I
feed on Wood and nest in wood material. This group consisted of 334 Individuals from bush
thicket forest and 65 Individuals from wooded grassland
On the other hand termite in functional group (ii) feeds on the wood, leaf litter, and nest on soil
or dry logs and brunches. A total of 696 individuals were recorded, the highest number of
individuals (458) were recorded from wooded grassland and238 from bush thicket forest
The functional group (iii) consisted of soil feeding and nesting termite, these were collected from
soil upper horizon and in damp wood or wood losing its structure. Out of 1988 individuals
collected, 775 Collected from wooded grassland and 118 from bush thicket forest. Table 6
displays termite functional group and their respective abundance. Comparison between termite
functional groups (i, ii and iii) show no significant difference (K=0.671, P = 0.715)
Table 6: termite functional groups from the two habitats at university of Dar es Salaam
main campus
FUNCTIONAL
GROUP
WOODED
GRASLAND
BUSH THICKET
FOREST
TOTAL PERCENTAGES (%)
I 65 334 399 20.07
II 458 238 696 35.01
III 775 118 893 44.92
TOTAL 1298 690 1988 100

22 | P a g e
Table 7 termite Functional groups and their respective subfamilies categorized at
University of dare s salaam main campus
Termite Subfamily Woded grassland Bush thicket forest Fh Functional group
Nasutitermitinae † † L/W II
Termitinae † † S/W III
Hodotermitinae † † S III
kalotermitidae sp1 † † L/W II
Macrotermitinae † † W I
Amitermitinae † † W I
Postelectectotermes † † S III
Termospinae ₋ † L/W II
Heterotermitinae † † L/W II
Kalotermitidae sp2 ₋ † W I
Psammotermitinae † ₋ W I
Coptotermitinae ₋ †
Total family 9 11
Note: Fh= functional habit, W= dry wood material, S= soil, L= leaf litter, += present, -
=absent

23 | P a g e
Chapter Four
4.0 DISCUSSION
4.1 Introductory Remarks
In this chapter results obtained from selected habitat in my study in university Main campus
including effect of habitat types, disturbances on termite’s species richness, abundance, and
diversity are discussed. The discussion merges certain aspects of termite ecology in the different
habitats and provides an overview of local termite communities.
4.1.1Termite Abundance in the selected Habitat
Termite was more abundant in wooded grassland than in bush thicket forest. Differences in
abundance are considered to have resulted due to ecological impact. The high abundance in
wooded grassland may be explained by availability and reliability of food such condition leading
to rapid individuals and population growth. Some researches reports have reported that land
clearing affects soil fauna population in the subtropical Possoh rain forest in Malaysia. Studies
on termite abundance and distribution at Mokwa in Nigeria have shown that Microtermes species
were more abundant in savannah woodland and agricultural ecosystem than in the forest in West
Africa
By slashing stems and leaves produces large amount of biomass in the field as termite food, this
activities is common to the studied site (wooded grassland) as the university administration
maintain its regular slashing activities, Studies on the effect of cultivation on termites carried out
at Mokwa in Nigeria showed that Macrotermes spp were able to exploit available food resources
after deforestation (Black and Wood, 1989). The presence of tree canopy and vegetation ground

24 | P a g e
cover improves soil moisture holding capacity which is favorable to termite but its removal
increases solar radiation reaching the soil surface.
4.1.2 Termites Species Diversity between Habitat types
Species diversity was higher in the forest habitat than in the wooded grassland habitats probably
due to availability of different types of food resources. The forest habitat has high dense of
vegetation providing more diverse food resources than wooded grassland characterized by sparse
vegetation. The canopy cover reduces solar radiation falling on the forest floor compared to the
grassland. Diverse food resources attract many more termites were to the forest micro climate
conditions.
Changes in micro climate to leads to reduction of termite species, similar studies carried out in
West Africa suggested that dense forest cover is one of the key factors favoring termite species
richness (Eggleton et al., 1996). Studies reported Amazon forest showed that the forest habitat
had microclimate that favors termite’s species. Furthermore, similar studies carried Central
Amazon showed that human activities like planting secondary forests and grazing in primary
forest areas reduced termite species richness. Other studies carried out in Mbalmayo forest
showed that species richness and the functional groups differ under different levels of forest
disturbance (Eggleton et al., 1995) and that reducing the canopy due to land use system lead into
reduction of termite assemblage. Furthermore, studies carried out in Amazon and Lowland
Sumatra, Indonesia showed that termite assemblages collapse along with land use these
contrasting findings suggest that different termite taxa may respond differently.

25 | P a g e
4.1.3 Termites Species Richness
There were 11 number of termite subfamily in the bush thicket forest, 9 in the grassland (table5
& fig2). Probably the number of termite species sampled was under estimated in contrast to
studies carried out in Malysia ( Tho,1992) The number are lower during the Early rainfall season
possibly because during that season the soil surface is a beat heats up quickly during the day
there by influencing termite foraging activity of subterranean termite due to forest landscape.
Though both selected studying sites experience the same condition of heat, but situation in forest
was little better compared to grassland at which canopy cover and denser vegetation provide
humid condition for which termite prefer unlike wooded grassland where vegetation cover is
sparse and allow light to heat up the soil and many termite not prefer such condition. Regular
mowing activities observed in the wooded grassland could be a leading factor at which reduce its
species richness as many functional habits of termite are reduced by pruning trees and destroy
mound around the area. Cutting down of some trees in bush thicket for fire wood observed in a
site at which this leads to increases number of termite prefer dry wood and leaf litter
microhabitats and hence increases number of termite subfamilies.
4.1.4 Termite Functional Group in Different Habitat Types
Knowledge of functional types in different habitat types is critical in predicting responses of
different species. Several important assumptions should be considered such as diversity of
functional groups before disturbances and their linkages in relation to key stone species (table 6
and 7). results from this work have shown that termite have a wide range of dietary, foraging,
and nesting habits, with many species showing a high degree of food specialization in agreement
with previous studies( Wood 1978; sleaford et al., 1996). The termite species recorded from
UDSM were categorized according to their food preference as follows; wood feeding termite,

26 | P a g e
dry wood and leaf litter, and soil. Many number of termite (898) found prefer soil habit and feeds
on soil and makes III categorized group of termite functional group. The followed category was
group II (696) which feeds on dry wood and leaf litter and few termite numbers were in category
I (399) which prefer feeding on wood. Assumed reason for many individuals of termite feeding
on soil to be found could be these; first, large part of sampled area featured by presence of
mounds especially in wooded grassland to which contribute for high abundance of termite in
wooded grassland than bush thicket forest and sampling activities done in morning leads to
accounting with many foraging termite which come to forage above the mound soil surface in
early morning. Secondary, there were few number of dead wood and few tree runways of fresh
plant trees and hence makes few number of termite in group I and group II
4.1.5 Termite Taxonomic Composition
A total of 12 subfamilies of termite were recorded after identification of collected samples of
termite from both habitats. Termospinae, Kaloterimitidae species 2, and Coptotermitinae
subfamilies were only recorded in bush thicket but in few relative numbers. Distinctiveness in
termite assemblages also observed in wooded grassland where by subfamily Psammotemitinae
recorded but not in bush thicket forest. Table 4, summarized subfamilies and their absence or
presence in their respective habitat.
Studies carried out near Darwin; northern Australia have shown that the termite fauna of the
monsoonal forest were under the same sub families Macrotermitinae, Termitinae, and
Nasutermitinae ( Tracy, 2003). They include the most advanced and diverse group of termite,
exhibit a wide variety of social specialization. Termite species in this group have elaborate
external and internal anatomy compared to the lower termite species. (Breznak and Brune, 1994)

27 | P a g e
5.0 CONCLUSION
The overall termite species abundance in grassland and forest habitats varies significantly due to
availability and reliability of food. The species richness and evenness were higher in the forest
due to higher productivity. The alternation of natural forest for various human activities has
reduced species richness in grassland habitat. Diversification of termite subfamilies members in
bush thicket forest than wooded grassland could be due to clearance and other related
disturbances in wooded grassland that reduces the variation in food quality and increases
competition among termite species and their natural enemies. Comparison of termite functional
group was significantly different. This study provides a baseline data on termite communities in
UDSM. Habitat changes have lead to moderate modification of termite communities in terms of
species richness, diversity and evenness.
5.1Finding from this Study
Habitat clearance for various purposes in the university campus impacts negatively on food
availability, nesting sites and termite species richness and Forest disturbance does not
immediately affect species evenness and distribution in UDSM probably because tree stumps,
dead root, and bunches provided adequate food for different species
5.2 Recommendation
1. All forest habitats should be protected and conserved to sustain termite species in
the campus as some termite are unable to withstand harsh environment resulting
from habitat alternation

28 | P a g e
2. Termite species richness in different habitat during dry season is not done.
Therefore there is need to carry out a campus wide survey aiming at identifying
different termite species in relation to factors influencing their richness and
diversity and help to contribute to additional information in Tanzania.
3. Termite molecular biology should be used in combination with morphological
characters to improve identification of termite species in Tanzania
4. Here I recommend for future research of termites covering under one areas of
termites specie richness during dry season,

29 | P a g e
6.0 REFERENCES
Alves, W., Mota A.S, de Lima. A., Bellezoni. R (2009), Termites as Bioindicators of Habitat Quality
in the Caatinga, Brazil
Avenant, N. L (200), “Small mammals community characteristics as indicator of ecological disturbances
in the Willem Pretorius Nature Reserve, Free State, South Africa’’, South Africa Journal of
Wildlife Research, 30: 26-30
Bandeira, A. G (1979), “Ecological de cupins (insect: Isoptera) da Amazonian Central: efeitos
doesmatamento sobre as populaces’’, Acta Amazoan, 9(3):481-99
Baroni, U. C., Josens, G. & Peakin, G.J. (1978). Empirical data and demographic parameters. In:
Production ecology of ants and termites (Brian, M.V. Ed.). Cambridge University Press,
Cambridge, UK,
Black, H. I. J. and Wood, T.G (1989), “The effect of cultivation on the vertical distribution of
Microtermes species (Isoptera: Termitidae; Macrotermitinae) in soil at Mokwa, Nigeria’’,
Sociobiology 15: 133-138
Breznack, J.A and Brune, A (1994), “Role of Micro organism in the Digestion of Lignocelluloses by
Termite” Annual Review of Entomology, Vol, 39: 453-487
Brouwer, J., Fussel, L. K. and Herman, (1993), “Soil and crop growth micro variability in the West
Africa semi arid topics a possible risk reducing factor for disturbance for subsistence farmers’’,
Agriculture Ecosystem and Environment, 45(3-4)
Collins, N.M. (1979), “Observations on the foraging activity of Hospitalitermes umbrinus (Havilland),
(Isoptera: Termitidae) in the Gunung MuluNational Park. Sarawak’’ Ecological Entomology, 4:
231-238

30 | P a g e
Davies, R.G. (1997), “Termite species richness in fire-prone and fire-protected dry deciduous
dipterocarp forest in DoiSuthep-PuiNational Park. Northern Thailand”, Journal of Tropical
Ecology, 13: 153-160.
Dawes-Gromadzki, T. Z (2005), “Bugs beneath the surface: the functional significance of soil macro
invertebrates to landscape health in Australia’s tropical savannas”, Insect Science 12: 307–312.
Eggleton, P. Bignell, D.E. (1995), Monitoring the response of tropical insects to changes in the
environment: Troubles with termites. In: R. Harrington and N.E. Stork, eds. Insects in a changing
environment, pp. 473-497. Academic Press, London.
Encarta encyclopedia 2007
Eggleton, P., Bignell, D.E., Sands, W.A., Mawdsley, N.A., Lawton, J.H., Wood, T.G. & Bignell, N.C.
(1996), “The diversity, abundance, and biomass of termites under differing levels of disturbance
in the Mbalmayo Forest Reserve Southern Cameroon”, Philosophical Transactions of the Royal
Society of London, series B 35:561-68
Eggleton, P (2000), Global diversity pattern of termite in: termite evolution, society symbiosis, Ecology
(Eds T. abe, M. Higashi and D.E. Bignell) Kluwer Academic press Dordrecht, The Netherlands.
Eligne, J. (1966), “Caracteres adaptifs au regime alimetaire dans la mandibule des termites (Insectes
Isopteres)”, Compte Rendu d'Academie des Sciences. Paris, 263: 1323-1325.
Fragoso, B.I., Goanzales, C., Arteaga, C., and Patron, J.C (1994), Relationship between earth worms and
soil organic matter levels in natural and managed ecosystem in the Mexico tropics; In (K.
Mulongoy and Mecchx eds), Soil organic matter dynamics and sustainability of tropical
agriculture. Pp231-240 Wiley and Sons, Chichester, UK

31 | P a g e
Holloway, J. D. A. H. A Kirk Spring, And V.K. Chey (1992), “the Response of Rainforest Insect groups
to logging and Conversion to Plantation. Philosophical Transaction’’, Royal Society, 335: 255-
236
Jones D.T., Susilo, F. X., Bignell, D. E., Suryo, H., Gillison, A. N. and Eggleton, P (2003), “Termite
assemblage collapse along a land use intensification gradient in lowland Central Sumatra,
Indonesia’’, Journal of Applied Ecology 40(2): 380-391
Jones, D.T (2002), “A survey of the termites (Insecta: Isoptera) of Tabalong District, South Kalimantan,
Indonesia”. Raffles Bulletin of Zoology 50, 117–128.
Jones D.T. (2000), “Termite assemblages in two distinct montane forest types at 1000 m elevation in the
Maliau Basin, Sabah”, Journal of Tropical Ecology, 16: 271-286
Kayan, S.A.; Sheikh, K.H. and Ahmad, M (1979), “Altitudinal distribution of termites in relation to the
vegetation and soil condition”, Parkistan journal of zoology, 11: 123-137
Krishna, K. and Weesner, F.M (1970), Biology of termite, vol.2, PP.477-525, New York and London:
academic press
Lavelle, P., Bignell, D. E and Lepage, M (1997), “Soil function in a changing world: the role of
invertebrate ecosystem engineers”, European Journal of Soil Biology 33, 159–193.
Magurran, A.E (2004), Measuring Biological Diversity, Blackwell Science, Oxford.
Nash, M. S., Anderson, J.P and Whitforld, W.G (1996), “Spatial and temporal variability in relative
abundance and foraging behavior of subterranean termite decertified and relatively intact”,
Chihuahua Desert ecosystems, Applied soil Ecology, 12: 149-157

32 | P a g e
Nyeko, P and Olubayo, F. M (2007), Participatory assessment of farmers’ experiences of termite
problems in agro forestry in Tororo district, Uganda. Agricultural Research and Extension
Network Paper No. 143, Overseas Development Institute, London, UK
Sleaford, F., Bignell, D.E. & Eggleton, P. (1996), “A pilot analysis of gut contents in termites from the
Mbalmayo Forest Reserve, Cameroon’’. Ecological Entomology, 21: 279-288
Tho, Y. P. (1978), Termite of peninsular Malaysia, (Kirton, L.G.Ed.), Malayan Forest Records, No.
36:224pp. Forest Research Institute Malaysia, Kepong
Tracy, G (2003), “The termite (isopteran) fauna of tropical rain forest near Darwin, northern Australia”
Australian Journal of Entomology,. Vol 44(2), P152-157
Watson, J.A.L. and Gay, F.J (1991), “Eutermes Exitiosus”, Hill, 1925(insect, isopteran) proposed
conservation by use of the plenary powers- bulleting zoology 38: 142-146
Wood, T.G. & Sands, W.A. (1978), the role of termites in ecosystems. Pp. 245-292 in Brian, M.V. (Ed)
Production ecology of ants and termites, Cambridge University Press, Cambridge
Wood, T.G. (1978), Food and feeding habits of termite. In production ecology of ants and termite
Production ecology of ants and termites (ed. M.V Brian,) Pp.55-80 University Press, Cambridge

33 | P a g e
7.0 APPENDICES
Appendix 1: comparison of termite functional groups
FUNCTIONAL GROUP
ABUNDANCE UNCTIONAL FROUP
ABUNDANCE
ii 18 iii 38
ii 56 iii 26
ii 9 iii 8
ii 5 iii 140
ii 18 iii 22
ii 5 iii 80
ii 21 iii 19
ii 18 iii 115
ii 27 iii 20
ii 12 iii 30
ii 40 iii 37
ii 29 iii 13
ii 113 iii 10
ii 50 iii 61
ii 33 iii 80
ii 80 iii 32
ii 12 iii 3
ii 80 iii 34
ii 10 iii 7
ii 20 iii 12
ii 40 iii 26
i 65 iii 34
i 40 iii 5
i 9 iii 31
i 1 iii 5
i 15 iii 5
i 39 iii 16
i 23 i 80
i 10 i 39
i 23 i 0
i 39

34 | P a g e
Appendix 2: comparison of termite microhabitats
MICRO HABITAT TOTAL MICRO HABITAT
TOTAL
Dry WOOD 18 SOIL 20
Dry WOOD 56 SOIL 30
Dry WOOD 9 SOIL 37
Dry WOOD 5 SOIL 13
Dry WOOD 18 SOIL 10
Dry WOOD 5 SOIL 61
Dry WOOD 21 SOIL 80
Dry WOOD 18 SOIL 32
LEAF LITTER 27 SOIL 3
LEAF LITTER 12 SOIL 34
LEAF LITTER 40 SOIL 7
LEAF LITTER 29 SOIL 12
LEAF LITTER 113 SOIL 26
LEAF LITTER 50 SOIL 34
LEAF LITTER 33 SOIL 5
LEAF LITTER 80 SOIL 31
LEAF LITTER 12 SOIL 5
LEAF LITTER 80 SOIL 5
LEAF LITTER 10 SOIL 16
LEAF LITTER 20 WOOD 65
LEAF LITTER 40 WOOD 40
SOIL 38 WOOD 9
SOIL 26 WOOD 1
SOIL 8 WOOD 15
SOIL 140 WOOD 39
SOIL 22 WOOD 23
SOIL 80 WOOD 10
SOIL 19 WOOD 23
SOIL 115 WOOD 39
WOOD 39 WOOD 80

35 | P a g e
Appendix 3: shows 30 samples collected on each habitat and their respective subfamily
richness between two selected habitats for three days to obtain cumulative curve
WOODED GRASSLAND
BUSH THICKET FOREST
SAMPLES COMMULATIVE SUB FAMILIES
SAMPLES COMMULATIVE SUBFAMILIES
1 1 1 1
2 2 2 2
3 3 3 3
4 6 4 4
5 7 5 5
6 8 6 6
7 10 7 7
8 11 8 7
9 12 9 8
10 13 10 9
11 13 11 10
12 15 12 10
13 15 13 11
14 15 14 11
15 16 15 12
16 17 16 12
17 18 17 12
18 18 18 12
19 18 19 12
20 18 20 12
21 18 21 12
22 18 22 12
23 18 23 12
24 18 24 12
25 18 25 12
26 18 26 12
27 18 27 12
28 18 28 12
29 18 29 12
30 18 30 12

36 | P a g e




















